
Submitted:
01 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
Data regarding the pandemic's impact on tilapia culture remain limited, but it is known that there was a significant decline in production and marketing since 2020. The post-pandemic challenges confronting tilapia farming necessitate prompt solutions, encompassing the management of bacterial infections and the adoption of more advanced technologies by small-scale producers in developing nations. Probiotics, acknowledged as a viable alternative, are presently extensively employed in tilapia aquaculture. Multiple studies have suggested that the application of diverse probiotics in tilapia culture has yielded favorable outcomes. Nonetheless, only a limited number of studies have employed statistical methods to evaluate such findings. To address this gap, a regression analysis was carried out to investigate the existence of a linear relationship between the probiotic dosage added to the feed and two key dependent variables: the specific growth rate (SGR) and the feed conversion ratio (FCR). Additionally, a hierarchical regression analysis was undertaken to ascertain the extent to which the variance observed in these responses could be explained by the variable "probiotic dosage in feed," after accounting for covariates such as initial weight, test duration, water temperature, and number of replicate tanks. Finally, two Pearson correlation matrices were constructed since different studies were included for the SGR and FCR analyses.
Keywords:
tilapia
; probiotics
; linear regression analysis
; hierarchical regression analysis
; Pearson correlation
1. Introduction
Aquaculture involves the controlled breeding of various aquatic animals and is primarily aimed at food production [1,2]. It has emerged as a significant component of today’s food production landscape, representing one of the sectors that recorded the most substantial growth in animal protein production [3]. The expansion of this activity can be ascribed to the increasing global population, which, in recent decades, has heightened the demand for non-traditional protein sources [4]. Aquaculture plays a crucial role in enhancing global food security and reducing poverty [5]. This is due to its widespread and cost-effective nature, extensive distribution, and commercialization, all achieved with minimal adverse environmental impacts. Consequently, it serves as a vital tool in preserving ecosystems [6]. The Asian continent stands out as a primary contributor to this industry, accounting for 89.4% of the world’s aquaculture production in 2016 [2]. According to FAO [3], Asia continued to dominate global aquaculture production in 2020, achieving a record production of 214 million tons (comprising 36 million tons of algae and 178 million tons of aquatic animals). Projections by FAO [3] anticipate further growth in the global production of aquatic animals, with an estimated 2030 target of 202 million tons. There has also been a consistent rise in per capita consumption of aquatic products (excluding algae), with the figure reaching 20.2 kg in 2020. However, it is noteworthy that, due to the COVID-19 pandemic, the trade in aquaculture products experienced a decline in 2020 compared to 2018, with the total value decreasing from a record US$ 165 billion in 2018 to US$ 151 billion [3].
China, as the world’s largest producer, surpassed the 47 million ton mark in aquaculture product output in 2018 [7]. This sector receives substantial investment priority compared to other economic sectors within the country [8,9]. If growth rates continue at their current pace, China will account for 62% of the world’s aquaculture production by 2025 [10]. The significance of Asia in the field of aquaculture can be attributed to the fact that this activity initially originated on the continent. The practice of aquaculture can be traced back to ancient Chinese civilization, dating to around 2000 B.C. During this time, carp (Cyprinus carpio) were first domesticated and utilized both as a food source and for ornamental purposes [11]. The global trade in fish produced through aquaculture started in the 1950s, driven by advancements in farming techniques and the declining marine fish populations available for capture due to intensive fishing [6]. Despite the promising market and favorable outlook for the expansion of tilapia farming [12], there are challenges that hinder productivity and development of this sector [13]. Among the hurdles faced by the industry, the prevalence of various diseases, particularly those of bacterial origin, is notable. These diseases often spread among tilapia populations, resulting in costly losses for producers [14]. Another issue is associated with the use of antibiotics to treat these diseases, as their indiscriminate use can pose environmental and human health risks. Therefore, it is imperative to validate alternative methods for the safe and effective control of diseases caused by antibiotic-resistant microorganisms [15,16].
This review article provides updated and comprehensive insights into the prospects of tilapia aquaculture in the global animal protein market, the challenges posed by infection-related losses, the development of antibiotic resistance, and the beneficial impact of using probiotics and bacteriocins on tilapia health.
2. Largest Global Producers, Their Potential and Challenges
Tilapia farming represents a significant component of the aquaculture sector. According to a report by FAO [8], data on the global production of the key aquaculture species indicates that, in 2020, production of Oreochromis niloticus (inland aquaculture) reached 4,407.2 thousand tonnes, accounting for 9% of the world’s total fish production that year. A substantial increase in production was expected; however, as mentioned earlier, the entire aquaculture sector experienced severe disruptions due to the COVID-19 pandemic [8]. The recent COVID-19 pandemic had a profound impact on global tilapia production, primarily driven by reduced demand for the product across several countries. Mobility restrictions further destabilized numerous production chains, leading to reduced productivity. FAO [8] anticipates that the tilapia farming market will recover in the years ahead, as pandemic-related restrictions are eased. Tilapia culture is a widespread activity, present in over 125 countries, and its global distribution is extensive [17]. The adaptability of tilapia, particularly Nile tilapia (Oreochromis niloticus), which constitutes more than 70% of tilapia culture [17], to various production systems contributes to its high diffusion [18]. FAO [8] reports that 4.4 thousand tons (live weight) of this species were produced in continental aquaculture in 2019, making it the third most produced aquatic species in this segment. One of the consumer-friendly features of tilapia is the absence of troublesome “Y” bones, making the product suitable for industrial filleting [19]. Tilapia culture is economically significant, particularly in countries across Asia, Africa, and the Americas [17]. Leading global producers in this sector, in descending order, include China, Indonesia, Egypt, and Brazil [20].
2.1. China: A Process of Refinement of Production Technique
China ascended to the forefront of this industry during the 1990s [21]. According to FAO [9], China continues to hold a dominant position in the global aquaculture sector, acting as the largest producer, processor, and consumer market. FAO also notes that, particularly since 2018, Chinese tilapia producers have encountered significant challenges in exporting their production, primarily due to tariff barriers imposed by the United States of America. This situation resulted in reduced Chinese exports to the United States, which, in turn, benefited tilapia producers in Latin America. Despite these export challenges, FAO [9] affirms that China remains the world’s largest exporter of tilapia. However, the combination of trade crises and the challenges faced by producers during the COVID-19 pandemic could potentially jeopardize China’s position as the leading tilapia producer in the coming years. Tilapia culture is prevalent in more than 30 Chinese provinces [22], with Guangdong standing out as the major contributor, accounting for 40% of the national production. This is due to its ideal conditions for tilapia cultivation, including a strategic geographical location and a suitable climate [21]. China benefits from an extensive coastal area and abundant water resources, thanks to the presence of lagoons, lakes, streams, and approximately one hundred rivers [23]. Yuan et al. [22] conducted a study that encompassed a range of farming systems in China, varying from rudimentary to highly sophisticated models. They concluded that, irrespective of the adopted model and the differences in expected profit, tilapia cultivation in the southern region proves to be more profitable and compensates for the investments made. The southern region contributes to over 90% of Chinese production due to its abundant water resources and favorable climate.
Southern aquaculture farmers often engage in polyculture, combining tilapia with other aquatic species, primarily Chinese carp and various shrimp species, which enhances the economic viability of this activity [23]. There have also been reports of recent growth in this activity in northern provinces such as Shandong and Beijing, though higher production costs are incurred due to the need for electricity to maintain breeding tanks warm [24]. A study conducted by Phiri and Yuan [25] revealed that most of the country’s facilities operate with high technological efficiency. However, it is estimated that average yields could increase by up to 9% through improvements in fish feeding and training of workers to adopt more effective resource management and strategies [23]. To ensure a balance between the domestic and international markets, the Chinese government encourages the storage and trade of a portion of the production from the southern to the northern provinces, helping to avoid product shortages [24]. The primary breeding system adopted in China is intensive, with the most commonly used facilities being closed tanks and cages, signifying a continuous refinement of production techniques [24]. In 2009, an industrialization project for tilapia culture was launched, supported by funding from central and provincial governments. This initiative saw an initial phase of growth, followed by a period of stagnation attributed to an increased incidence of streptococcosis resulting from high stocking density and stressful breeding conditions [17,24]. Aside from its significance in domestic supply, China also stands out as a significant supplier of tilapia to the international market [8,9]. Chinese exports primarily cater to the strong demand for the product in the United States [8,9]. Until 2014, export rates exhibited year-over-year growth, but in the subsequent three years, they decreased from 69.00% to 63.23% [26]. One of the most prominent reasons for this decline in exports is believed to be competition of Indonesian products, which exhibit significant differences in price and quality. Chinese products have been classified as unsatisfactory by the international market due to the presence of drug residues in tilapia [22].
Between 2005 and 2016, annual tilapia production increased by 45.03%, with an average growth rate of 7.8% per year [23]. According to FAO’s survey in 2019, tilapia production increased from 1.28 million tons in 2010 to 1.56 million tons in 2016, with expectations of reaching 1.93 million tons in 2019 and a positive trajectory for the following year [20]. However, as previously mentioned, the COVID-19 pandemic resulted in production declines in several countries [8,9]. Furthermore, several factors may contribute to potential stagnation in China’s tilapia farming in the future. These include the country’s climatic instability, price fluctuations that impact international demand and reduce the competitiveness of Chinese products [22], and the spread of diseases in the breeding environment, such as bacteriosis caused by Salmonella spp. [27]. The COVID-19 pandemic has also impacted China’s tilapia production system. Although production and exports resumed in 2021, the numbers remain lower compared to 2019. Production levels remained below the 2019 benchmark in 2022 as well, influenced, in part, by the high cost of transportation [28].
2.2. Indonesia: The Association Between Tilapia Culture and Rice Farming
Indonesia has gained international recognition in the field of aquaculture, particularly in the production of tilapia, shrimp, and tuna, as per FAO [8]. The country holds the position of being the third-largest supplier of tilapia to the United States, following China and Colombia. The prospects for tilapia culture in Indonesia are promising, with exports consistently showing growth rates over the past two decades [26]. Tilapia farming began in Indonesia in 1930, initially with the introduction of Mozambique tilapia (Oreochromis mossambicus), although it did not hold significant economic importance. It wasn’t until the 1960s when Nile tilapia was introduced, following the adoption of the GIFT genetic improvement program, that Indonesia emerged as a key player in this sector [29]. Starting in 2004, annual growth reached an impressive 20% [30]. Between 2010 and 2017, tilapia production increased from 458 thousand tons to 1.10 million tons [17,31]. The growth continued between 2017 and 2019, culminating in a total production of 1.35 million tons [20]. According to FAO [8], it’s estimated that in Indonesia 50% of protein sources designated for human consumption come from aquatic animals. Among the species commonly cultivated, Nile tilapia, including the GIFT strain, and Red tilapia (a hybrid of O. niloticus and O. mossambicus) are prominent [17].
The culture of Red tilapia can be carried out in net tanks or in cages set up in brackish waters or lagoons, with the latter being a preferred option for rural farmers in Indonesia [32]. The integration of tilapia farming with rice cultivation, a practice also prevalent in Egypt, is considered a promising approach. It optimizes water resource usage and aids in controlling insects and pests through fish consumption [33]. Additionally, plants used in conjunction with this practice can enhance water quality by absorbing phosphorus and nitrogenous substances, thereby increasing fish production and harvest yields [34]. The range of production modalities includes intensive, extensive, and semi-intensive systems, with the latter being the most common. Semi-intensive practices combine strategies characteristic of intensive models, such as the use of feed additives and fertilizers, while maintaining facilities similar to extensive practices [35]. Despite substantial potential for expansion, certain limitations persist, especially the high cost of feed and significant waste production on small farms [36,37]. This underscores the importance of investments and government support to encourage small producers, particularly in remote areas [30]. Another challenge that has to face production in the country is the spread of bacterial diseases, primarily caused by Aeromonas hydrophila [38].
2.3. Egypt: Good Results from Production Industrialization
The expansion of tilapia farming in the African continent is becoming a reality, with Egypt and Nigeria emerging as the largest producers in the region, as reported by FAO [18]. Egypt has seen a remarkable growth in tilapia production. Between 1995 and 2000, it surged from 21 thousand tons to 157 thousand tons, followed by a substantial increase to 557 thousand tons in 2010 [17,31]. By 2017, the production had reached 967 thousand tons, accounting for 79% of African production. According to El-Sayed and Fitzsimmons [28], this percentage increased to 85% of the total African production in 2019. This rapid growth in productivity can be attributed to the shift from extensive practices to intensive ones, with the government’s support and investment in the introduction of new techniques [33,39]. Since 2015, regional governments have organized workshops and training programs to educate tilapia producers on best management practices. These interventions have resulted in increased tilapia production, improved profitability, and reduction in the environmental impact in the regions surrounding the facilities [40].
The primary producing region is situated along the Nile River, encompassing reservoirs in the lakes of the north coast [18]. Tank cultivation systems are also installed in desert areas [2]. It is noteworthy that some tributaries of the Nile River, such as Lake Manzala in the river delta, are plagued by severe pollution from nearby industrial plants. A histological study on Nile tilapia reared in the region has revealed degenerative changes in the seminiferous tubules, as well as deformations in the ovaries [41]. High levels of heavy metals, including aluminum, iron, nickel, and chromium, have been found in the gills and muscles of tilapia from the Salam canal, which receives water from the Damietta branch of the Nile River [42]. Bacteria associated with human bacteriosis have also been detected, including Escherichia coli, Streptococcus agalactiae, Streptococcus iniae, and A. hydrophila [43]. This underscores the urgency of implementing measures to treat the canal and monitor water quality [44]. Egypt faces a significant challenge in terms of infections, with viral diseases being a major cause of tilapia mortality, particularly during the summer [45]. Small Egyptian producers also encounter difficulties in accessing quality feed with appropriate formulations to promote growth and bolster tilapia immunity [39]. As a result, small producers often engage in the farming of other fish species such as carp or mullet alongside tilapia [2,39]. Nonetheless, the industrialization of farming systems has yielded positive results in Egypt, establishing the country’s global prominence in tilapia production [39].
2.4. Brazil: Favorable Natural Characteristics
Intensive livestock and poultry farming are the primary sources of animal protein in Brazil [46], and the country has made substantial investments in these sectors in recent years [13]. Among the emerging potentials in Brazil, tilapia farming has stood out as one of the fastest-growing activities [47]. Tilapia culture was first introduced in Brazil in 1952 with the Congo tilapia (Tilapia rendalli). However, its slow growth rate resulted in low productivity, rendering the initial attempt at tilapia culture in the country ineffective [48]. The introduction of tilapia gained momentum in the 1970s when the O. niloticus and Oreochromis hornorum species were simultaneously introduced by the National Department of Works Against Drought (DNOCS) [13]. Among the more than 70 known and cataloged tilapia species, only four species, all belonging to the Oreochromis genus, are prominent in the global aquaculture market [49]. In Brazil, O. niloticus predominates [48], representing 45% of fish production on the continent [19]. Brazil experienced rapid growth between 2014 and 2019, with production increasing from 200 thousand tons at the beginning of this period to 450 thousand tons by the end [20,31]. Studies have highlighted Nile tilapia as the species with the greatest productivity potential in Brazil due to its adaptation to the tropical climate, tolerance to intermediate salinity environments [50], resistance to low levels of dissolved oxygen in water [19], and adaptability to various cultivation systems [49]. Tilapia farming in Brazil serves both the national and international high demand markets [46]. In 2018, the export of tilapia alone accounted for 81.35% of all fish from aquaculture, with Mato Grosso do Sul being the leading exporting state [12]. Key destinations for Brazilian tilapia production include Japan, China, and the United States, as per data from the annual survey of the Brazilian Association of Fish Culture [20].
While the domestic consumption of fish in Brazil averaged 11 kg per capita in 2018, indicating a promising market, this figure falls short of the World Health Organization’s recommended 12 kg per capita [51]. Brazil’s favorable conditions for tilapia culture include abundant water resources, particularly river channels [52], a climate suitable for supporting tilapia’s metabolic activities [48], and a growing trend of private investments in management technology [53]. This activity also generates socioeconomic benefits by reducing food deficits [54], providing affordable and nutritious food to local communities [55], and maintaining relatively low production costs [56]. Tilapia farming allows for production models based on cooperatives and associations, contributing to further socioeconomic gains [13]. These models have been successfully implemented in Brazil, particularly in Santa Catarina, the fourth-largest tilapia producer in the country [12], resulting in improved human development indices in producing cities [50]. However, despite the remarkable growth in Brazilian tilapia culture in recent decades, several challenges remain [13], the overcoming of which is crucial. The lack of government policies to incentivize production centers, the inadequate training for producers in proper management, and the preservation of natural resources are among the most pressing challenges [52]. Bureaucracy in granting water use permits and environmental permits for fish farm construction is another significant hindrance to facility expansion [46], which, in turn, hampers new investments in the sector [13]. Feed purchase represents a major expense in the production process, accounting for 70-80% of production costs, depending on the region and production system employed [46]. Additionally, the incidence of diseases is a growing concern, especially in relation to fish stocking density and the chosen culture system [48]. Several investment projects are planned for the coming years, with estimates suggesting that, based on these investments, Brazil could achieve a production of approximately 500,000 million tons (mt) by 2025 [17,39].
3. Main Pathogens for Tilapia
Bacterial infections pose a significant challenge in various sectors of aquaculture [57], and they are particularly problematic for tilapia culture companies. This is because some bacterial pathogens are responsible for substantial production losses worldwide [14]. The prevalence of pathogenic species varies depending on the region [58] and the specific tilapia species in question [59]. The primary bacterial diseases that affect tilapia culture include streptococcosis, francisellosis, aeromonosis, vibriosis, and columnariosis [60].
3.1. Streptococcosis
Streptococcosis is the primary infection responsible for tilapia mortality worldwide and affects various species, including Nile tilapia [61] and Red tilapia [62]. Outbreaks of this disease result in estimated losses of up to $40 million annually in the tilapia industry [63]. Between 2009 and 2019, recurrent streptococcosis outbreaks occurred in China, impacting the profitability of the industry, with fish mortality rates ranging from 30% to 90%, particularly affecting farms in the southern region [64,65]. Streptococcus spp. are Gram-positive, coccus-shaped bacteria [66] that prefer aerobic conditions but can tolerate anaerobic environments [67]. Under experimental conditions, Streptococcus spp. thrive at temperatures between 26°C [68] and 37°C [69]. Consequently, outbreaks of contamination primarily occur during the summer, and higher temperatures lead to increased mortality rates [68]. Hu et al. [70], in a comparison of S. agalactiae cultures incubated at temperatures of 25°C and 35°C, observed that strains grown at higher temperatures exhibited greater pathogenicity. This was evidenced by increased secretion of virulence factors, nucleotides, and compounds associated with stress regulation, such as oxidized glutathione and glyceraldehyde-3-phosphate. Notably, these virulence factors accelerated bacterial growth, enhanced the adhesion capacity of pathogens, created pores in the host cell membrane, and caused tissue damage [68].
The primary etiological agent is the species S. iniae, which is the leading cause of increased tilapia mortality worldwide [71,72,73]. This pathogen also affects other fish species like Crucian carp (Carassius auratus) [74] and can even infect mammals, including humans [68]. Multiple studies [75,76,77] have shown that infections mainly occur through the gastrointestinal tract. The pathogen, after passing through the stomach and intestinal mucosa, adheres to the gastrointestinal epithelium. Following this stage of infection, its cells can spread to other organs, leading to systemic septicemia [75]. Soto et al. [76] found that infections proceeded through intramuscular injection in their tests on Nile tilapia to mimic the natural conditions of disease development, suggesting that lesions in the epidermis may represent an entry route for the pathogen. High stocking density in culture facilities, along with the aggressive behavior of the fish, can increase the likelihood of skin ulcers, which can favor this type of infection. Common symptoms of streptococcosis include erratic swimming, exophthalmos, corneal opacity, and skin lesions [64]. Septicemia and meningoencephalitis are frequently observed, along with complications that affect the liver, kidneys, and spleen [78].
3.2. Francisellosis
Francisellosis is another significant disease in tilapia culture, known for its high mortality rates and the ability to persist in the environment [76]. Although its distribution is more limited, it severely impacts productivity in endemic regions. Francisellosis is frequently found in the United States, Indonesia, the United Kingdom, and Latin America, with southern Brazil being one of the regions most affected by outbreaks [79,80]. The first confirmed cases of francisellosis in Brazil date back to 2014, with a mortality rate of around 60% in the country. The primary etiological agent is Francisella noatunensis subsp. orientalis, which exhibits coccus-rod morphology, is Gram-negative, and can grow strictly aerobically with facultative intracellular growth [48,81]. While the pathogenic mechanisms of this bacterium are not entirely understood, it is known that temperatures below 25°C favor disease outbreaks [82]. Therefore, the incidence of the disease and mortality, particularly among fry and young tilapia, increases in Brazil during the winter months. Intensive farming conditions, such as high stocking density and poor water quality, can increase susceptibility to the development of francisellosis [83,84]. The transmission of the disease can occur horizontally through direct contact between the pathogen present in the water and the fish’s skin, as well as through direct contact between infected animals. Vertical transmission is also possible, where breeding tilapia can transmit the pathogen to fry or eggs. The entry routes for F. noatunensis subsp. orientalis are the skin, peritoneum, and gastrointestinal tract through the consumption of contaminated food. The disease can manifest acutely with symptoms such as anorexia, erratic swimming, anemia, and exophthalmos. Additionally, it can also occur in a chronic or sub-acute form, which involves milder disease symptoms and results in lower mortality rates. The differentiation among clinical conditions depends on the extent of intracellular infiltrates in the central nervous system. Common complications are related to the appearance of multifocal granulomas containing the pathogen, with the most affected regions being the spleen, kidneys, liver, and skeletal muscle tissue [48].
3.3. Aeromonosis
The most common bacterial infections in freshwater aquaculture are linked to the genus Aeromonas spp. [85]. These pathogens primarily impact freshwater fish [66]. Various bacterial species act as opportunistic parasites in tilapia, with A. hydrophila, a rod-shaped, Gram-negative bacterium [19], being the principal agent responsible for the primary transmission of aeromonosis [85] and the development of co-infections with Tilapia lake virus [87,88]. The predisposition to initiate infection depends on specific environmental triggers that activate the secretion of virulence factors [89,90]. Pathogenicity is promoted by the presence of particular pollutants in the culture water, hypoxia, high fish stocking density, pre-existing host infections, and elevated temperatures [89]. It has been reported that the optimal temperature for bacterial growth and disease development is 28 °C [43]. The disease has a significant incidence in Asian countries such as Indonesia and Malaysia [91] but is also present in other tropical regions like Latin America [92,93] and African countries, including Egypt [93]. The mucosal surface is a primary entry route for A. hydrophila [90]. Addo et al. [94] documented that tilapia infected through intraperitoneal injection displayed erratic swimming, multiple areas of hemorrhage, depigmentation, and skin erosion on the fins. Aeromonosis is known to progress into motile Aeromonas septicemia when combined with other secondary infections [94], potentially leading to fatal outcomes due to deep ulcerations in internal organs, necrosis of liver, brain, kidneys, and blood flow congestion [95,96]. In the case of highly virulent strains, infected fish may perish before exhibiting clinical signs, rendering diagnosis and containment of bacterial spread challenging [95].
3.4. Vibriosis
The term vibriosis encompasses a group of diseases that affect a wide range of aquatic species [97]. Pathogens responsible for vibriosis include various species within the genus Vibrio spp., such as Vibrio harveyi, Vibrio anginolyticus, Vibrio cholerae [98], and Vibrio vulnificus [99]. These bacteria belong to the Gram-negative, rod-shaped category [97] and are equipped with flagella to facilitate their movement [100]. While they are more commonly found in saline environments [101], some species can also thrive in freshwater [97]. The geographical distribution of vibriosis primarily spans from coastal regions in European countries [102] to Asian countries [99]. Outbreaks tend to be seasonal, occurring primarily in the summer [102]. However, the expression of virulence can even be observed in cold waters with temperatures exceeding 15 °C [99,103]. While the exact mode of transmission and evasion from the host’s immune system remains unclear, it is known that the virulent bacteria can adhere to the host’s skin and infiltrate the tissues [101]. Clinical symptoms include lethargy, impairment of fish development, tissue necrosis, malformations, scale discoloration, and erythema near the oral cavity [101].
3.5. Columnariosis
Columnariosis is caused by the bacterium Flavobacterium columnare, which is a Gram-negative species characterized by its long, non-flagellated bacillus shape [104]. This disease has a global distribution and is highly contagious [66], with a higher incidence in tropical countries, making it a recurring issue in Brazilian tilapia farming [82]. The susceptibility to infection varies with the developmental stage of tilapia, with fry and young fish being more prone to severe symptoms of the disease [66,105]. The occurrence of columnariosis is more common in the summer, as temperatures exceeding 20 °C promote pathogen growth [104]. However, the optimal temperature for the development of virulence is between 28 and 30 °C [66]. Other factors, such as low dissolved oxygen levels, high stocking density, and high ammonia concentration in the aquatic environment, stimulate the secretion of virulence factors, making infection easier [104]. The disease can be transmitted orally through the gastrointestinal tract and via contact of pathogenic bacteria with pre-existing lesions [66,106]. Initial symptoms, such as lethargy, erratic swimming, and accelerated opercular movements, are nonspecific and can be mistaken for clinical signs seen in other bacterial diseases [66]. As the infection progresses, more characteristic features of columnariosis become apparent, including erosion of the dorsal and caudal fins, the presence of yellowish or gray skin erosion near a reddish hyperemic zone, and tissue necrosis in the cranial and branchial region [104].
4. Use of Antibiotics in Disease Control
Implementing effective measures to control bacterial diseases is vital for sustaining tilapia culture productivity. Antibiotics are commonly employed to treat these diseases. However, their usage is not always accompanied by proper technical oversight, and often, high doses are administered without identifying the specific pathogen responsible for the infection [107]. The lack of accurate diagnoses for bacterial diseases and a surveillance system to determine the necessity of antibiotics application is a prevalent issue in many countries [108,109]. Even in regions where regulations and inspections exist, these measures are primarily applied to systems intended for export [107,109]. The indiscriminate use of antibiotics can lead to severe problems in tilapia culture and pose risks to human health. One significant concern is the development of antibiotic resistance, which renders these drugs ineffective for disease control and promotes the selection of resistant bacterial strains [110]. Moreover, bacteria possess mechanisms for exchanging genetic material [111], potentially facilitating the transfer of resistance genes to previously antibiotic-sensitive strains, rendering existing treatments inefficient [112]. In this context, numerous studies have assessed the incidence of antibiotic-resistant microorganisms in tilapia culture (Table 1). The adverse consequences stemming from the indiscriminate use of antibiotics can also be linked to ecological losses [113]. It is estimated that over 80% of antibiotics used in aquaculture persist in the aquatic environment for decades after use [114]. During this period, they can diminish the populations of phytoplankton and green algae, which are crucial for the primary production of organic matter necessary for the maintenance of biochemical cycles in aquatic ecosystems [115].
To mitigate the impact of antibiotic use, the World Health Organization (WHO), World Organization for Animal Health (OIE), and Food and Agriculture Organization (FAO) have established global guidelines for antibiotic use in animals and monitoring of antibiotic-resistant strains. This information has proven invaluable for governments and regulatory agencies to make informed decisions [116]. As shown in Table 2, many tilapia pathogens have developed resistance to commonly used antibiotics in farming systems.
Notably, some of the key bacteria responsible for tilapia diseases have shown resistance to most of the drugs used to treat bacteriosis (Figure 1). For instance, S. agalactiae exhibits resistance to many of the antibiotics commonly used for infections [117]. Moreover, most studies indicate that the persistence of the use of prohibited drugs or their use without a prescription from a qualified technician is a significant issue, especially in breeding systems, with China being a prominent example. In addition to therapeutic use, many aquaculture farmers resort to these drugs as a preventive measure, believing that prophylactic medication can reduce mortality and accelerate animal growth [129]. While improved growth may indirectly result from pathogen control, the indiscriminate use of antimicrobials carries inherent risks. Therefore, it is essential to explore new, promising alternatives to antibiotics, such as probiotics, as they have demonstrated the ability to combat pathogenic microorganisms while displaying high metabolic efficiency [130].
5. Benefits of Probiotics, Prebiotics and Bacteriocins in Tilapia Culture
5.1. Probiotics in Tilapia Culture
In light of the risks associated with excessive antibiotic exposure, both for the environment and human health [15], the use of probiotics as growth promoters to replace antibiotics has gained momentum [131]. Probiotics are living microorganisms that can have beneficial effects on the host when administered in sufficient quantities (Figure 2) [116,132].
Probiotics can help regulate the intestinal microbiota by competing with enteropathogens for nutrients and space [133], without affecting the bacteria that make up the normal host microbiota [134], and can then be employed in tilapia culture as growth promoters and potential producers of antimicrobial peptides [131]. Furthermore, they can stimulate the immune response and enhance the resistance of tilapia to pathogens, which is particularly important given that the confinement conditions in breeding systems can act as stressors and contribute to immunosuppression [17]. Improved animal growth can occur through various mechanisms [135]. Probiotic microorganisms can stimulate appetite and optimize host nutrition by competing with bacteria responsible for amino acid deamination, thus reducing nitrogen uptake [136]. Additionally, probiotics secrete fatty acids, essential amino acids, biotin, and enzymes capable of breaking down carbohydrates, lipids, and proteins into smaller fragments in the animal’s digestive tract, making them more easily absorbable [137]. The endogenous enzymes produced by tilapia are often insufficient to ensure efficient utilization of feed, and the enzymes from probiotics can enhance nutrient absorption [138].
Table 3 presents a list of recent studies that have reported enzyme secretion by probiotics used in tilapia. Although the number of studies is limited, the most recent ones aimed at assessing the impact of these enzymes on tilapia health have highlighted that probiotics are valuable sources of exogenous enzymes, particularly digestive enzymes. These enzymes play a beneficial role in promoting fish health, with key clinical benefits including an improved immune response, increased resistance to diseases, and faster weight gain. Notably, after the administration of probiotics in the tilapia diet, there was an increase in the nutritional efficiency of the feed. This is because most of the main enzymes secreted by probiotics are involved in the digestion of complex nutrients, indicating an enhancement in development and disease resistance. Well-nourished animals are known to have a lower risk of disease and a higher growth rate.
Table 4 provides a summary of recent investigations on the effects of probiotics on tilapia. Notably, most of the selected studies utilized probiotic concentrations within the range of 107 to 108 CFU/g, administered for periods ranging from 7 to 60 days. Growth promotion was a predominant effect observed, especially in studies with shorter administration periods, suggesting that the benefits can manifest in the initial days of probiotic use. For instance, Dawood et al. [139] administered the probiotic Aspergillus oryzae at concentrations close to the typical range for tilapia studies (~108 CFU) and found significant enzyme activities, including those of superoxide dismutase and catalase, after an extended 60-day dosing period. These enzymes likely led to improvements in the immune response and protection against the tested pathogen (A. hydrophila). A similar pattern was observed by Gobi et al. [143], who also noted these benefits with longer administration periods. In contrast, Selim and Reda [144] administered the probiotic Bacillus amyloliquefaciens for 8 weeks but only observed lysozyme production, with no growth promotion, likely due to the significantly lower probiotic concentration used.
The competition promoted by probiotics in tilapia culture extends beyond receptor sites and includes essential nutrients needed for the metabolism of pathogens [164]. Some probiotics have the capacity to produce siderophores, which are iron-chelating agents that can capture metal ions from the environment, thus reducing the availability of iron to pathogenic bacteria [165]. Probiotics can also enhance tilapia’s resistance to disease by improving water quality [166]. For example, Gram-positive bacteria, primarily belonging to the genus Bacillus spp., are known for their ability to contribute to the aquatic ecosystem by effectively breaking down organic matter [167]. Probiotics can also modulate parameters like salinity, pH, and ammonia concentration in the water [168]. In addition to reducing the susceptibility to diseases, probiotics improve the tilapia immune response, leading to increased survival when exposed to infections [164,165]. Some studies have shown that probiotics influence gene expression related to the inflammatory pathways and regulation of immunological markers [169,170]. This means that probiotic bacteria significantly impact gene expression, and even when the probiotic microorganisms are no longer alive, their secreted metabolites can still produce immunomodulatory effects [171]. Gram-positive probiotics are known to predominantly stimulate the production of pro-inflammatory cytokines such as IL-1, IL-6, IL-12, tumor necrosis factor α (TNF-α), gamma interferon (IFN-γ), and anti-inflammatory cytokines like IL-10. This stimulation enhances the phagocytic activity of leukocytes and increases antibody levels as well as activity of enzymes associated with the innate immune system. On the other hand, Gram-negative probiotics primarily stimulate cell-mediated immunity at the expense of the humoral response, which is associated with serum immunity and mucus production [164].
In tilapia culture, the most commonly used probiotics belong to the genus Bacillus [172]. They are followed, to a lesser extent, by lactic acid bacteria (LABs) [173,174]. The frequent use of Bacillus spp. in tilapia culture can be attributed to the spore-forming capacity of these bacteria. Spore formation makes them easier to handle and apply because spores tend to pass through the stomach relatively unharmed. Moreover, spore formation provides greater resistance to adverse storage conditions, such as drying, exposure to heat, and UV radiation, thereby increasing their viability [175]. In contrast, LABs are Gram-positive bacteria that are catalase and oxidase negative. They typically have a cocci or bacilli shape [176] and do not form spores [177]. LABs synthesize lactic acid, which leads to the secretion of metabolites (acidoline, acidophylline, lactocidine, and lactonin) that contribute to lowering the pH in the surrounding medium, affecting the metabolism of hydrogen peroxide in enteropathogens [178]. LABs also produce molecules with antagonistic effects on other bacteria, such as bacteriocins [179].
The concentration of probiotics used in various studies on tilapia yielded different results, with some important findings. Xia et al. [180] found that a concentration of 108 CFU/mL brought the most benefits to the host, including growth promotion, improved feed conversion ratio, and enhanced immune response. Similar results were reported in studies that maintained a dosage of 108 CFU/g for a two-month period. Lower concentrations of probiotics (105-107 CFU/g) were also effective in promoting growth and improving immunological parameters [142,143,153]. Some studies even demonstrated positive effects with just one week of probiotic administration. Bacillus subtilis used alone was found not to be effective in promoting the growth of Nilotic tilapia in any of the selected studies. However, Adeoye et al. [158] reported growth promotion when B. subtilis was combined with other Bacillus species. This suggests that a combination of probiotic strains may be more efficient than a single strain, but the underlying mechanisms for these synergistic effects are not fully understood. Bacillus licheniformis, administered at various concentrations, improved nutrient absorption, exogenous enzyme production, growth rate, and feed conversion [143,160]. B. amyloliquefaciens did not affect the growth of tilapia after 30 days of administration, but growth performance and weight gain increased even at sub-standard concentrations after 2 months [159].
Lactobacillus plantarum strains administered at concentrations higher than the standard for more than one month enhanced the immune response to infections by stimulating cytokine production and regulating intestinal flora. This reduced the population of potentially pathogenic bacteria, as reported by Foysal et al. [15] and Hamdan et al. [181]. In some studies, like that of Chu et al. [147], it was found that even short-term administration of probiotics at certain concentrations could increase the secretion of digestive enzymes and survival of tilapia when exposed to pathogens. While some studies reported increased survival rates with probiotic administration, others did not reveal significant changes, likely due to the influence of factors such as the probiotic strain used, concentration and treatment duration. It is important to note that the optimal concentration and duration of probiotic administration may vary depending on the strain, fish health condition and environmental factors in the aquaculture system. Further research is needed to determine the most effective probiotic regimens for various conditions and tilapia species.
The use of probiotics in tilapia culture has shown various benefits, particularly in terms of promoting growth, enhancing the immune response, and reducing mortality due to disease. Some key findings from these studies are outlined below. Bacillus spp., especially at concentrations around 107 CFU/g, have been effective in reducing mortality and increasing resistance to diseases like S. agalactiae infection. Similar reduction in mortality have been observed with B. subtilis. B. licheniformis administration at concentrations below the standard dosage resulted in increased lysozyme levels, which is associated with improved innate immune responses. The combination of Bacillus strains, such as B. subtilis and Bacillus cereus, enhanced feed conversion, but did not have any substantial impact on the growth rate. Enterococcus faecium, when administered for a short duration (7 days), had a positive effect on tilapia growth and immune system. L. plantarum, especially at high concentrations (109 to 1010 CFU/g), showed an increased immune response to infections and changes in gut microbiota.
Administration method, animal age, time of use, dosage, and application frequency are important parameters to consider when using probiotics. For tilapia, the recommended dosage typically ranges from 105 to 109 CFU/mL, with a preference for higher concentrations compared to other fish species. Probiotics are typically incorporated into the animal feed as pellets, flours, granules, or flakes, while direct administration in the culture water is less effective. Encapsulation methods, such as using calcium alginate matrices, can increase the viability and adhesion of probiotics in the fish’s digestive tract. It has been shown that mixing different probiotic strains has a better effect than the use of a single strain. The optimal period and frequency of probiotic use are not yet well-defined and should be the focus of future research. In summary, probiotics offer promising advantages in tilapia culture, both in terms of promoting fish health and productivity, thus potentially contribute to disease prevention and control. However, further research is needed to fine-tune the optimal application methods and dosages for various conditions and species of tilapia.
In view of these results, we conducted a linear regression analysis (GraphPad Prism version 9.0, GraphPad Software, San Diego, CA, USA) to explore whether a linear relationship exists between probiotic dose and two response variables of interest, namely specific growth rate (SGR; 13 studies), defined as the ratio of ln final mean weight (g) - ln initial mean weight (g) to the experimental time (days), and feed conversion ratio (FCR; 16 studies), defined as the ratio of feed fed (g) to weight gain (g). Data were the mean values reported in the selected studies at a significance level of 5%. Only the presence or absence of probiotics was considered as a criterion to select experimental group, but not the probiotic format (i.e., single or multi strain probiotic).
Results showed that the independent variable “probiotic dose in feed” does not have any linear relationship with both SGR (p = 0.167) (Figure 3) and FCR (p = 0.66) (Figure 4); therefore, it cannot predict their values.
Further, a hierarchical regression analysis (IBM SPSS Statistics for Mac, version 28., IBM Corp., Armonk, N.Y., USA) was conducted to determine the amount of variance observed in these responses (Table 5 for SGR and Table 6 for FCR) explained by the same independent variable, after accounting for covariables, including initial weight, test duration, water temperature and number of replicate tanks.
The first model, which considered four covariables, explained a significant (p = 0.002) proportion (35.4%) of the variance observed on SGR among studies; however, when adding the variable “probiotic dose in feed” as predictor into the second model, no significant (p > 0.05) changes were detected in the variance of this response, reflected by a R square change of 2.3%. In addition, significant (p < 0.001) negative correlation between initial weight and SGR was detected among studies. For FCR, the first model explained a significant (p = 0.002) proportion (27.9%) of the variance observed among studies; however, in the second model, no significant (p > 0.05) changes were detected in the variance of this response, reflected by no change in R square value. Significant (p < 0.001) negative correlation between temperature and FCR, test duration, and initial weight as well as replicate number and test duration were detected. These results suggest that “probiotic dose in feed” cannot be considered as predictive variable neither for SGR nor FCR. In a previous work, although no significant linear relationship between SGR and probiotic dose in fish feed was found, a significant linear relationship between probiotic dose in fish feed and FCR was detected [16]. The reasons for this inconsistency between reports might be due to the different number of articles taken into consideration, i.e., 32 in that study and only 16 in the present one for FCR regression, thus affecting the statistical inference.
Finally, we built two Pearson correlation matrices since different studies were included for SGR and FCR analyses (Figure 5 and Figure 6), which evidenced significant negative correlations a) between initial weight and SGR (p < 0.01), b) between water temperature and FCR (p < 0.05), and c) between test duration and both initial weight and number of replicates (p < 0.001).
5.2. Probiotics in Combination with Prebiotics (Synbiotics)
The use of synbiotics, which combine probiotics and prebiotics, is gaining attention in aquaculture due to their potential to enhance the growth, health, and immunological parameters of fish. Synbiotics offer a synergistic approach by promoting the growth of beneficial microorganisms in the gut and providing substrates that support their activity.
Here are some key findings and implications related to the use of synbiotics in tilapia culture. Prebiotics are substrates that selectively promote the growth and activity of beneficial gut microorganisms. They can lead to several health benefits, including the modulation of the intestinal microbiota, reduction in intestinal pH (which can enhance mineral absorption), and the production of microbial metabolites like short-chain fatty acids (e.g., butyrate) that are beneficial for gut health. Synbiotics are supplements that combine probiotics (beneficial live microorganisms) with prebiotics (substrates that support their growth). This combination is believed to have more powerful effects on the host’s health than using probiotics or prebiotics alone. Studies have shown that the use of synbiotics can enhance the protection and growth of tilapia. For example, one study found that a synbiotic containing specific bacterial strains and mannan oligosaccharides (MOS) significantly increased the protection of Nile tilapia against infections by A. hydrophila. Synbiotics, like the above-mentioned A. oryzae plus β-glycan synbiotic, have been reported to improve the production of antioxidants and immunomodulation in fish. These effects can help enhance the immune response and overall health of the fish. In some cases, the combination of a probiotic with a prebiotic in the form of a synbiotic has been found to significantly reduce fish mortality [19,43,139,154].
These results suggest that synbiotics can play a crucial role in disease prevention and overall fish survival. The use of synbiotics in tilapia culture offers a promising approach to improving the health and productivity of fish. It demonstrates that a combination of probiotics and prebiotics can have more substantial and beneficial effects on fish growth, health, and disease resistance than using these supplements separately. Further research and experimentation with synbiotics can help optimize their application in tilapia farming and aquaculture more broadly.
5.3. Bacteriocins in Tilapia Culture
The use of bacteriocins and bacteriocin-like inhibitory substances (BLIS) produced by probiotics in tilapia culture is an area of growing interest due to their potential antibacterial activity against pathogenic bacteria (Table 7).
Bacteriocins are ribosomally-synthesized peptides with bactericidal or bacteriostatic effects against a wide range of bacterial strains. Here are some key findings and implications regarding the use of bacteriocins and BLIS in tilapia culture.
5.3.1. Mechanism of Action
Bacteriocins primarily exert their antibacterial effects by disrupting the target bacteria’s cell membrane through the formation of pores. They interact with anionic lipids in the bacterial membrane, affecting amino acid transport and the proton motive force necessary for ATP synthesis. This mode of action makes bacteriocins particularly effective against Gram-positive bacteria, which have a higher content of anionic lipids.
5.3.2. Antimicrobial Potential
LAB bacteriocins have demonstrated significant antibacterial activity against a wide range of pathogenic bacteria and are often used in the food industry as natural preservatives due to their ability to inhibit the growth of spoilage and pathogenic bacteria.
5.3.3. Resistance Mechanisms
Bacteriocins have different mechanisms of action than antibiotics, making them potential candidates for combating bacterial infections caused by antibiotic-resistant strains. This makes them particularly valuable in scenarios where antibiotic resistance is a concern.
5.3.4. Synergistic Effects
While single bacteriocins can be effective, combining different bacteriocins may enhance their effectiveness. The use of bacteriocinogenic probiotics that produce multiple bacteriocins can offer a more robust approach to preventing and controlling diseases in tilapia.
5.3.5. Challenges in Characterization
Characterizing bacteriocins and BLIS can be challenging. In vitro studies have shown that these substances can inhibit pathogenic strains, but the precise details of their properties and characteristics may vary depending on the focus of each study. In On the other hand, determining the antibacterial effects of bacteriocins in in vivo studies can be complex, as the supplementation of probiotics often leads to multiple health improvements in fish, including disease resistance. It can be challenging to isolate the specific contribution of bacteriocins to these effects.
5.3.6. Safety and Efficacy
It’s essential to evaluate the safety and efficacy of probiotics and their bacteriocin production for the specific animal model, in this case, tilapia. Different fish species may have unique metabolic diversity, and the effectiveness of probiotics can vary from one species to another.
5.3.7. Identification Challenges
The literature shows that the identification of bacteriocins used in tilapia culture is limited. Further research is needed to identify and characterize bacteriocins produced by probiotics for targeted applications in tilapia farming. The use of bacteriocins and BLIS produced by probiotics can be an exciting way for improving the health and disease resistance of tilapia in aquaculture. By leveraging the antibacterial potential of these substances, researchers and aquaculturists can explore novel ways to prevent and manage bacterial infections in this important fish species.
6. Conclusions
In conclusion, the tilapia farming industry has experienced steady growth and economic impact in recent years. However, it faces several challenges in the near future, including recovering from production reductions caused by the COVID-19 pandemic, adopting new production technologies, improving disease control, and finding safe and effective alternatives to antimicrobials. Bacterial infections have caused significant losses in tilapia farming in various regions, and the ineffectiveness of some antibiotics due to bacterial resistance has been a growing concern. Probiotics, bacteriocins, and bacteriocin-like inhibitory substances (BLIS) may be promising alternatives to antibiotics for promoting fish health and controlling diseases. To further boost the use of probiotics in tilapia culture, ongoing research is necessary to better understand the microorganisms used as probiotics and identify new candidates. It is essential to investigate their modes of action and determine the optimal application conditions to achieve the best results. The statistical analysis of published studies suggests that the “dose of probiotic in the feed” may not serve as a predictive variable for specific growth rate or feed conversion ratio. The relationship between probiotic dosage and these parameters may be influenced by various other factors, such as initial weight, test duration, water temperature, and number of replicate tanks. Additionally, it is noteworthy that several pathogens causing diseases in tilapia farming have shown high levels of antibiotic resistance. Biotechnological methods, including the use of probiotics and bacteriocins, have shown promise in addressing this issue. While they may not entirely replace antibiotics, they represent valuable alternatives in promoting productivity and disease control in tilapia farming. Continued research, development and responsible application of probiotics, bacteriocins, and synbiotics will be crucial in enhancing the sustainability and productivity of tilapia aquaculture in the years ahead. These biotechnological approaches can contribute to reducing the reliance on antibiotics, mitigating disease outbreaks, and supporting the growth of this important sector.
Author Contributions
Conceptualization, investigation and writing —original draft preparation, W.A.P., I.L.R., A.V., C.R., S.M.F., C.M.N.M., J.R.; data curation, writing—review and editing, D.C.D., L.T., A.C., A.M.E., E.F.V.; supervision and writing—review and editing, R.P.S.O.
Funding
This work was funded by the FAPESP-São Paulo Research Foundation under Grant 2018/25511-1, 2021/01570-1, 2022/07543-9; CNPq-National Council for Scientific and Technological Development of Brazil under Grant 2020-2577; and the Commission of Scientific and Technological Development ANID/FONDECYT/INICIACIÓN under Grant 11230690.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lucas, J.S.; Southgate, P.C.; Tucker, C.S. Aquaculture: farming aquatic animals and plants. John Wiley & Sons, West Sussex, United Kingdom. 2019.
- FAO (Food and Agriculture Organization of the United Nations). The state of World fisheries and aquaculture. Topics fact sheets. Available online: http://www.fao.org/state-of-fisheries-aquaculture/2018/en (accessed on 2023 February 04).
- FAO (Food and Agriculture Organization of the United Nations). Aquaculture News. May 2022, No. 65. Rome. Available online: https://www.fao.org/documents/card/en/c/cc0158en (accessed on 04 February 2023).
- Woods, R.G. Future dimensions of world food and population, 1st ed.; CRC Press: New York, USA, 2019. [Google Scholar]
- Kassam, L.; Dorward, A. A comparative assessment of the poverty impacts of pond and cage aquaculture in Ghana. Aquaculture 2017, 470, 110–122. [Google Scholar] [CrossRef]
- Siqueira, T.V.D. Aquicultura: a nova fronteira para produção de alimentos de forma sustentável. Rev. BNDES 2018, 25, 119–170. Available online: https://web.bndes.gov.br/bib/jspui/bitstream/1408/16085/1/PRArt_Aquicultura%20a%20nova%20fronteira_compl.pdf (accessed on 2023 May 28).
- SOFIA (State of World Fisheries and Aquaculture). The state of world fisheries and aquaculture. Available online: http://www.fao.org/3/ca9229en/CA9229EN.pdf (accessed on 2023 February 04).
- FAO (Food and Agriculture Organization of the United Nations). The state of World fisheries and aquaculture. Topics fact sheets. Available online: http://www.fao.org/3/ca9229en/CA9229EN.pdf (accessed on 2023 February 04).
- FAO (Food and Agriculture Organization of the United Nations). Aquaculture development in China: the role of public sector policies. Available online: http://www.fao.org/3/Y4762E/Y4762E00.htm (accessed on 2023 February 04).
- SOFIA (State of World Fisheries and Aquaculture). The state of World fisheries and aquaculture. Available online: http://www.fao.org/3/i5798e/i5798e.pdf (accessed on 2023 February 04).
- Calado, R.; Olivotto, I.; Oliver, M.P.; Holt, G.J. Marine ornamental species aquaculture; John Wiley & Sons: West Sussex, UK, 2017. [Google Scholar]
- PEIXE BR. Anuário Peixe Br da piscicultura brasileira. São Paulo: Associação Brasileira de Piscicultura. Available online: https://www.peixebr.com.br/anuario-2020/ (accessed on 2023 February 04).
- Schulter, E.P.; Vieira Filho, J.E.R. Desenvolvimento e potencial da tilapicultura no Brasil. Rev. Econo. Agron. 2018, 16, 177–201. [Google Scholar] [CrossRef]
- Hassan, S.; Abdel-Rahman, M.; Mansour, E.S.; Monir, W. Prevalence and antibiotic susceptibility of bacterial pathogens implicating the mortality of cultured Nile tilapia, Oreochromis niloticus. Egyp. Jour. Aquac. 2020, 10, 23–43. [Google Scholar] [CrossRef]
- Foysal, M.J.; Alam, M.; Kawser, A.R.; Hasan, F.; Rahman, M.M.; Tay, C.Y.; Gupta, S.K. Meta-omics technologies reveals beneficiary effects of Lactobacillus plantarum as dietary supplements on gut microbiota, immune response and disease resistance of Nile tilapia (Oreochromis niloticus). Aquaculture 2020, 520, 734–974. [Google Scholar] [CrossRef]
- Pereira, W.A.; Piazentin, A.C.M.; de Oliveira, R.C. Bacteriocinogenic probiotic bacteria isolated from an aquatic environment inhibit the growth of food and fish pathogens. Sci Rep 2022, 12, 5530. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Tilapia culture; Academic Press: London, United Kingdom, 2019. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). National aquaculture overview – Egypt. 2020. Available online: http://www.fao.org/fishery/countrysector/naso_egypt/en (accessed on 2023 February 04).
- De Andrade, L.A.R.; de Azevedo, T.M.P. Manejo experimental de alevinos de tilápia (Oreochromis niloticus), alimentados com ração comercial e pré-probióticos. PUBVET 2018, 12, 133. [Google Scholar] [CrossRef]
- PEIXE BR. Anuário Peixe Br da piscicultura brasileira. São Paulo: Associação Brasileira de Piscicultura. 2019. Available online: https://www.peixebr.com.br/anuario-peixe-br-da-piscicultura-2019/ (accessed on 2023 February 04).
- Gu, D.E.; Yu, F.D.; Yang, Y.X.; Xu, M.; Wei, H.; Luo, D.; Hu, Y.C. Tilapia fisheries in Guangdong Province, China: Socio-economic benefits, and threats on native ecosystems and economics. Fishe. Manag. Ecol. 2019, 26, 97–107. [Google Scholar] [CrossRef]
- Yuan, Y.; Yuan, Y.; Dai, Y.; Gong, Y. Economic profitability of tilapia farming in China. Aquac. Intern. 2017, 25, 1253–1264. [Google Scholar] [CrossRef]
- Yuan, Y.; Yuan, Y.; Dai, Y.; Zhang, Z.; Gong, Y.; Yuan, Y. Technical efficiency of different farm sizes for tilapia farming in China. Aquac. Res. 2020, 51, 307–315. [Google Scholar] [CrossRef]
- Xu, P.; Ming, J. Status and trends of the tilapia farming industry development. Aquac. Indai China: Succ. Stor. Mod. Tren. 2018, 404–420. [Google Scholar] [CrossRef]
- Phiri, F.; Yuan, X. Technical efficiency of Tilapia production in Malawi and China: Application of stochastic frontier production approach. Jour. Aquac. Res. Develop. 2018, 9, 532. [Google Scholar] [CrossRef]
- Dai, Y.Y.; Yuan, Y.M.; Yuan, Y.; Zhou, Z.; Zhang, H.Y. Competitiveness of Chinese and Indonesian tilapia exports in the US market. Aquac. Intern. 2020, 28, 791–804. [Google Scholar] [CrossRef]
- Li, K.; Petersen, G.; Barco, L.; Hvidtfeldt, K.; Liu, L.; Dalsgaard, A. Salmonella Weltevreden in integrated and non-integrated tilapia aquaculture systems in Guangdong, China. Food Microb. 2017, 65, 19–24. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Fitzsimmons, K. From Africa to the world—The journey of Nile tilapia. Revi. Aquac. 2023, 15, 6–21. [Google Scholar] [CrossRef]
- Fathi, M.; Dickson, C.; Dickson, M.; Leschen, W.; Baily, J.; Muir, F.; Weidmann, M. Identification of Tilapia Lake Virus in Egypt in Nile tilapia affected by ‘summer mortality’syndrome. Aquaculture 2017, 473, 430–432. [Google Scholar] [CrossRef]
- Wati, L.A.; Harahab, N.; Riniwati, H.; Utami, T.N. Development of tilapia cultivation strategy based local economy: case study in Wonogiri, Central Java. IOP Conf. Series: Earth and Envir. Sci. 2020, 493, 012042. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Global aquaculture production 2019, 1950-2017. Available online: http://www.fao.org/fishesy/statistics/global-aquaculture-production/query/en (accessed on 2023 February 04).
- Wijayanto, D.; Kurohman, F.; Nugroho, R.A. Bioeconomic of profit maximization of red tilapia (Oreochromis sp.) culture using polynomial growth model. IOP Conf. Series: Earth and Envir. Sci. 2018, 139, 012040. [Google Scholar] [CrossRef]
- Shaalan, M.; El-Mahdy, M.; Saleh, M.; El-Matbouli, M. Aquaculture in Egypt: insights on the current trends and future perspectives for sustainable development. Revi. Fish. Sci. Aquac. 2018, 26, 99–110. [Google Scholar] [CrossRef]
- Goada, A.M.A.; Essa, M.A.; Hassaan, M.S.; Sharawy, Z. Bio economic features for aquaponic systems in Egypt. Turk. Jour. Fish. Aqua. Sci. 2015, 15, 525–532. [Google Scholar] [CrossRef]
- Setiadi, E.; Widyastuti, Y.R.; Prihadi, T.H. Water quality, survival, and growth of red tilapia, Oreochromis niloticus cultured in aquaponics systems. E3S Web of Conf. 2018, 47, 02006. [Google Scholar] [CrossRef]
- Parata, L.; Mazumder, D.; Sammut, J.; Egan, S. Diet type influences the gut microbiome and nutrient assimilation of Genetically Improved Farmed Tilapia (Oreochromis niloticus). Plos one 2020, 15, e0237775. [Google Scholar] [CrossRef]
- Mo, W.Y.; Man, Y.B.; Wong, M.H. Use of food waste, fish waste and food processing waste for China’s aquaculture industry: Needs and challenge. Sci. Total Envir. 2018, 613, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Fadjar, M.; Sanoesi, E.; Mintiya, Y.A.; Hakim, L. Effect of squid powder (Loligo Sp.) on antibody titer and bacteria density in blood of tilapia (Oreochromis niloticus) infected by Aeromonas hydrophila. Aquac. Indo. 2020, 21, 24–31. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. On-farm feed management practices for Nile tilapia (Oreochromis niloticus) in Egypt. On-farm feeding and feed management in aquaculture. FAO Fish. Aquac. Techn. Paper 2013, 583, 101–129. [Google Scholar]
- Dickson, M.; Nasr-Allah, A.; Kenawy, D.; Kruijssen, F. Increasing fish farm profitability through aquaculture best management practice training in Egypt. Aquaculture 2016, 465, 172–178. [Google Scholar] [CrossRef]
- Mansour, A.A.H.; El-kady, A.H.M.; Abu Almaaty, A.H.M.; Ramadan, M.A. Effect of environmental pollution on gonads histology of the Nile tilapia, Oreochromis niloticus from Lake Manzala, Egypt. Egyp. Jour. Aqua. Biol. Fish 2018, 563–572. [Google Scholar] [CrossRef]
- Badawy, R.K.; Omer, A.M.; Othman, D.I. Bio-Fertilization Effect on the Productivity and Biodiesel Quality of Castor Plant Oil under El-Salam Canal Irrigation Condition. Alexa. Sci. Exch. Jour. 2018, 39, 168–182. [Google Scholar] [CrossRef]
- Nicholson, P.; Fathi, M.A.; Fischer, A.; Mohan, C.; Schieck, E.; Mishra, N.; Jores, J. Detection of tilapia lake virus in Egyptian fish farms experiencing high mortalities in 2015. Jour. Fish Dise. 2017, 40, 1925–1928. [Google Scholar] [CrossRef]
- Donia, G.; Hafez, A.; Wassif, I. Studies on some heavy metals and bacterial pollutants in tilapia fish of El Salam Canal, Northern Sinai, Egypt. Egyp. Jour. Aqua. Biol. Fish. 2017, 21, 67–84. [Google Scholar] [CrossRef]
- Ali, S.E.; Jansen, M.D.; Mohan, C.V.; Delamare-Deboutteville, J.; Charo-Karisa, H. Key risk factors, farming practices and economic losses associated with tilapia mortality in Egypt. Aquaculture 2020, 527, 735438. [Google Scholar] [CrossRef]
- Milanez, A.Y.; Guimarães, D.D.; Maia, G.D.S.; Muñoz, A.E.P.; Pedroza Filho, M.X. Potencial e barreiras para a exportação de carne de tilápias pelo Brasil. Embrapa Pesca e Aquic. 2019, 25, 155–213. [Google Scholar]
- Castilho-Barros, L.; Owatari, M.S.; Mouriño, J.L.P.; Silva, B.C.; Seiffert, W.Q. Economic feasibility of tilapia culture in southern Brazil: A small-scale farm model. Aquaculture 2020, 515, 734551. [Google Scholar] [CrossRef]
- Raghiante, F.; de Mattos Ferrasso, M.; Rodrigues, M.V.; Biondi, G.F.; Martins, O.A. Francisella spp. em tilápias no Brasil: Uma revisão. Ver. Bras. Higi. Sanid. Ani. 2017, 11, 119–130. [Google Scholar]
- Mello, S.C.R.P.; de Oliveira, E.D.C.P.; de Seixas Filho, J.T. Aspectos da aquicultura e sua importância na produção de alimentos de alto valor biológico. Semioses 2017, 11, 28–34. [Google Scholar] [CrossRef]
- Barroso, R.; Muñoz, A.; Tahim, E.; Tenório, R.; Muehlmann, L.; Silva, F.; Flores, R. Dimensão socioeconômica da tilapicultura no Brasil, 1st ed.; Embrapa: Brasília, Brasil, 2018. [Google Scholar]
- CNA (Confederação da agricultura e pecuária do Brasil). Tilápia, cenário econômico 2018. Available online: https://www.cnabrasil.org.br/assets/arquivos/artigostecnicos/AntecipaCNACena%CC%81rio-Econo%CC%82mico-Tilapia.pdf (accessed on 2023 February 04).
- Kubitza, D.; Becka, M.; Mueck, W.; Halabi, A.; Maatouk, H.; Klause, N.; Bruck, H. Effects of renal impairment on the pharmacokinetics, pharmacodynamics and safety of rivaroxaban, an oral, direct Factor Xa inhibitor. Brit. Jour. Clin. Pharm. 2010, 70, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Pedroza Filho, M.X. Análise comparativa de resultados econômicos dos polos piscicultores no segundo trimestre de 2015. Ativ. Aqui. CNA 2015. Available online: https://www.embrapa.br/busca-de-publicacoes/-/publicacao/1041290/analise-comparativa-de-resultados-economicos-dos-polos-piscicultores-no-segundo-trimestre-de-2015 (accessed on 2023 February 04).
- Igarashi, M.A. Aspectos tecnológicos e perspectivas de desenvolvimento do cultivo de tilápia no Brasil. Arqu. Ciên. Veter. Zool. da UNIPAR 2018, 21, 123–130. [Google Scholar] [CrossRef]
- Nowland, S.J.; O’Connor, W.A.; Osborne, M.W.; Southgate, P.C. Current status and potential of tropical rock oyster aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 57–70. [Google Scholar] [CrossRef]
- Asche, F.; Cojocaru, A.L.; Roth, B. The development of large scale aquaculture production: A comparison of the supply chains for chicken and salmon. Aquaculture 2018, 493, 446–455. [Google Scholar] [CrossRef]
- Hamom, A.; Alam, M.M.; Iqbal, M.M.; Khalil, S.M.I.; Parven, M.; Sumon, T.A.; Mamun, M.A.A. Identification of pathogenic bacteria from diseased nile tilapia Oreochromis niloticus with their sensitivity to antibiotics. Intern. Jour. Curr. Microb. Appl. Sci. 2020, 9, 716–1738. [Google Scholar] [CrossRef]
- Guerrero-Cabrera, L.; Olivera, B.C.L.; Villavicencio-Pulido, J.G.; Luna, R.J.O. Proximity and density of neighboring farms and water supply, as risk factors for bacteriosis: A case study of spatial risk analysis in tilapia and rainbow trout farms of Oaxaca, Mexico. Aquaculture 2020, 520, 734955. [Google Scholar] [CrossRef]
- Wanja, D.W.; Mbuthia, P.G.; Waruiru, R.M.; Bebora, L.C.; Ngowi, H.A. Natural concurrent infections with Black Spot Disease and multiple bacteriosis in farmed Nile tilapia in Central Kenya. Veter. Med. Intern. 2020, 8821324. [Google Scholar] [CrossRef]
- Soto, E.; Zayas, M.; Tobar, J.; Illanes, O.; Yount, S.; Francis, S.; Dennis, M.M. Laboratory-controlled challenges of Nile Tilapia (Oreochromis niloticus) with Streptococcus agalactiae: comparisons between immersion, oral, intracoelomic and intramuscular routes of infection. Jour. Compar. Pathol. 2016, 155, 339–345. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, J.; Chen, K.; Gao, T.; Yao, H.; Liu, Y.; Lu, C. Development of Streptococcus agalactiae vaccines for tilapia. Disea. Aqua. Organ. 2016, 122, 163–170. [Google Scholar] [CrossRef]
- Ismail, M.S.; Siti-Zahrah, A.; Syafiq, M.R.M.; Amal, M.N.A.; Firdaus-Nawi, M.; Zamri-Saad, M. Feed-based vaccination regime against streptococcosis in red tilapia, Oreochromis niloticus x Oreochromis mossambicus. BMC Veter. Resea. 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Sun, J.; Fang, W.; Ke, B.; He, D.; Liang, Y.; Ning, D.; Ke, C. Inapparent Streptococcus agalactiae infection in adult/commercial tilapia. Scient. Rep. 2016, 6, 26319. [Google Scholar] [CrossRef]
- Ye, X.; Li, J.; Lu, M.; Deng, G.; Jiang, X.; Tian, Y.; Jian, Q. Identification and molecular typing of Streptococcus agalactiae isolated from pond-cultured tilapia in China. Fish. Sci. 2011, 77, 623–632. [Google Scholar] [CrossRef]
- Li, L.; Huang, T.; Liang, W.; Chen, M. Development of an attenuated oral vaccine strain of tilapia Group B Streptococci serotype Ia by gene knockout technology. Fish & Shellf. Immun. 2019, 93, 924–933. [Google Scholar] [CrossRef]
- Bueno, D.; Neto, R.T. As principais bacterioses que acometem a tilápia do Nilo (Oreochromis niloticus): Revisão de literatura. Arq. Bras. Med. Vet. FAG 2019, 2, 103–113. [Google Scholar]
- Veselá, K.; Kumherová, M.; Klojdová, I.; Solichová, K.; Horáčková, Š.; Plocková, M. Selective culture medium for the enumeration of Lactobacillus plantarum in the presence of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. LWT 2019, 114, 108365. [Google Scholar] [CrossRef]
- Palang, I.; Hirono, I.; Senapin, S.; Sirimanapong, W.; Withyachumnarnkul, B.; Vanichviriyakit, R. Cytotoxicity of Streptococcus agalactiae secretory protein on tilapia cultured cells. Jour. Fish Dis. 2020, 43, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Bal, F.A.; Ozkocak, I.; Cadirci, B.H.; Karaarslan, E.S.; Cakdinleyen, M.; Agaccioglu, M. Effects of photodynamic therapy with indocyanine green on Streptococcus mutans biofilm. Photod. Photod. Ther. 2019, 26, 229–234. [Google Scholar] [CrossRef]
- Hu, W.T.; Guo, W.L.; Meng, A.Y.; Sun, Y.; Wang, S.F.; Xie, Z.Y.; He, C. A metabolomic investigation into the effects of temperature on Streptococcus agalactiae from Nile tilapia (Oreochromis niloticus) based on UPLC–MS/MS. Veter. Microb. 2017, 210, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Laith, A.A.; Abdullah, M.A.; Nurhafizah, W.W.I.; Hussein, H.A.; Aya, J.; Effendy, A.W.M.; Najiah, M. Efficacy of live attenuated vaccine derived from the Streptococcus agalactiae on the immune responses of Oreochromis niloticus. Fish & Shellf. Immun 2019, 90, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Saleh, H.; Gabr, A.N.M.; Aboyadak, I.; Saber, N. Subcellular degenerative changes in hepatopancreas and posterior kidney of Streptococcus iniae infected Nile tilapia using transmission electron microscope. Egyp. Jour. Aquat. Biol. Fish. 2019, 23, 305–316. [Google Scholar]
- Suhermanto, A.; Sukenda, S.; Zairin Jr., M.; Lusiastuti, A.M.; Nuryati, S. Characterization of Streptococcus agalactiae bacterium isolated from tilapia (Oreochromis niloticus) culture in Indonesia. Aquac. Aquar. Conserv. Legisl. 2019, 12, 756–766. [Google Scholar]
- Geng, Y.; Wang, K.Y.; Huang, X.L.; Chen, D.F.; Li, C.W.; Ren, S.Y.; Lai, W.M. Streptococcus agalactiae, an emerging pathogen for cultured ya-fish, Schizothorax prenanti, in China. Transb. Emer. Dis. 2012, 59, 369–375. [Google Scholar] [CrossRef]
- Iregui, C.A.; Comas, J.; Vásquez, G.M.; Verjan, N. Experimental early pathogenesis of Streptococcus agalactiae infection in red tilapia Oreochromis spp. Jour. Fish Dis. 2016, 39, 205–215. [Google Scholar] [CrossRef]
- Soto, E.; Wang, R.; Wiles, J.; Baumgartner, W.; Green, C.; Plumb, J.; Hawke, J. Characterization of isolates of Streptococcus agalactiae from diseased farmed and wild marine fish from the US Gulf Coast, Latin America, and Thailand. Jour. Aqua. Ani. Heal. 2015, 27, 123–134. [Google Scholar] [CrossRef]
- Vásquez-Machado, G.; Barato-Gómez, P.; Iregui-Castro, C. Morphological characterization of the adherence and invasion of Streptococcus agalactiae to the intestinal mucosa of tilapia Oreochromis sp.: an in vitro model. Jour. Fish Dis. 2019, 42, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, P.; Mon-on, N.; Jaemwimol, P.; Tattiyapong, P.; Surachetpong, W. Coinfection of tilapia lake virus and Aeromonas hydrophila synergistically increased mortality and worsened the disease severity in tilapia (Oreochromis spp.). Aquaculture 2020, 520, 734746. [Google Scholar] [CrossRef]
- Leal, C.A.G.; Tavares, G.C.; Figueiredo, H.C.P. Outbreaks and genetic diversity of Francisella noatunensis subsp orientalis isolated from farm-raised Nile tilapia (Oreochromis niloticus) in Brazil. Gen. Mol. Res. 2014, 13, 5704–5712. [Google Scholar] [CrossRef] [PubMed]
- Facimoto, C.T.; Chideroli, R.T.; Di Santis, G.W.; Gonçalves, D.D.; Do Carmo, A.O.; Kalapothakis, E.; Pereira, U.P. Complete genome sequence of Francisella noatunensis subsp. orientalis strain F1 and prediction of vaccine candidates against warm and cold-water fish francisellosis. Gen. Mol. Res. 2019, 18, 1–10. [Google Scholar] [CrossRef]
- Soto, E.; Baumgartner, W.; Wiles, J.; Hawke, J.P. Francisella asiatica as the causative agent of piscine francisellosis in cultured tilapia (Oreochromis sp.) in the United States. Jour. Vet. Diagn. Inv. 2011, 23, 821–825. [Google Scholar]
- Sebastião, F.A.; Pilarski, F.; Kearney, M.T.; Soto, E. Molecular detection of Francisella noatunensis subsp. orientalis in cultured Nile tilapia (Oreochromis niloticus L.) in three Brazilian states. Jour. Fish Dis. 2017, 40, 1731–1735. [Google Scholar] [CrossRef] [PubMed]
- Amal, M.N.A.; Saad, M.Z.; Zahrah, A.S.; Zulkafli, A.R. Water quality influences the presence of Streptococcus agalactiae in cage cultured red hybrid tilapia, Oreochromis niloticus × Oreochromis mossambicus. Aquac. Res. 2015, 46, 313–323. [Google Scholar] [CrossRef]
- Assis, G.B.N.; Tavares, G.C.; Pereira, F.L.; Figueiredo, H.C.P.; Leal, C.A.G. Natural coinfection by Streptococcus agalactiae and Francisella noatunensis subsp. orientalis in farmed Nile tilapia (Oreochromis niloticus L.). Jour. Fish Dis. 2017, 40, 51–63. [Google Scholar] [CrossRef]
- Dong, H.T.; Techatanakitarnan, C.; Jindakittikul, P.; Thaiprayoon, A.; Taengphu, S.; Charoensapsri, W.; Senapin, S. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). Jour. Fish Dis. 2017, 40, 1395–1403. [Google Scholar] [CrossRef]
- Fernandes, D.C.; Eto, S.F.; Funnicelli, M.I.; Fernandes, C.C.; Charlie-Silva, I.; Belo, M.A.; Pizauro, J.M. Immunoglobulin Y in the diagnosis of Aeromonas hydrophila infection in Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 500, 576–585. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Koh, C.B.; Nurliyana, M.; Suhaiba, M.; Nor-Amalina, Z.; Santha, S.; Zamri-Saad, M. A case of natural co-infection of Tilapia Lake Virus and Aeromonas veronii in a Malaysian red hybrid tilapia (Oreochromis niloticus × O. mossambicus) farm experiencing high mortality. Aquaculture 2018, 485, 12–16. [Google Scholar] [CrossRef]
- Salem, M.; Zharan, E.; Saad, R.; Zaki, V. Prevalence, molecular characterization, virulotyping, and antibiotic resistance of motile aeromonads isolated from Nile tilapia farms in northern Egypt. Mans. Vet. Med. Jour. 2020, 21, 56–67. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Samir, F.; Abd El-Naby, A.S.; Monier, M.N. Antioxidative and immunostimulatory effect of dietary cinnamon nanoparticles on the performance of Nile tilapia, Oreochromis niloticus (L.) and its susceptibility to hypoxia stress and Aeromonas hydrophila infection. Fish & Shellf. 2018, 74, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Farias, T.H.V.; Arijo, S.; Medina, A.; Pala, G.; da Rosa, P.E.J.; Montassier, H.J.; de Andrade, B.M.A. Immune responses induced by inactivated vaccine against Aeromonas hydrophila in pacu, Piaractus mesopotamicus. Fish & Shellf. Immun. 2020, 101, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Basri, L.; Nor, R.M.; Salleh, A.; Saad, M.Z.; Barkham, T.; Amal, M.N.A. Co-infections of tilapia lake virus, Aeromonas hydrophila and Streptococcus agalactiae in farmed red hybrid tilapia. Animals 2020, 10, 2141. [Google Scholar] [CrossRef]
- Grajales-Hahn, S.; Hahn-von, H.C.M.; Grajales-Quintero, A. Case report of Aeromonas salmonicida in nilotic tilapia (Oreochromis niloticus) in caldas, Colombia. Boletín Científico 2018, 22, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Elsheshtawy, A.; Yehia, N.; Elkemary, M.; Soliman, H. Investigation of Nile tilapia summer mortality in Kafr El-Sheikh governorate, Egypt. Gen. Aqua. Org. 2019, 3, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Addo, S.; Carrias, A.A.; Williams, M.A.; Liles, M.R.; Terhune, J.S.; Davis, D.A. Effects of Bacillus subtilis strains and the prebiotic Previda® on growth, immune parameters and susceptibility to Aeromonas hydrophila infection in Nile tilapia, Oreochromis niloticus. Aquaculture Research 2017, 48, 4798–4810. [Google Scholar] [CrossRef]
- Pauzi, N.A.; Mohamad, N.; Azzam-Sayuti, M.; Yasin, I.S.M.; Saad, M.Z.; Nasruddin, N.S.; Azmai, M.N.A. Antibiotic susceptibility and pathogenicity of Aeromonas hydrophila isolated from red hybrid tilapia (Oreochromis niloticus × Oreochromis mossambicus) in Malaysia. Vet. World 2020, 13, 2166. [Google Scholar]
- Hal, A.M.; Manal, I. Gene expression and histopathological changes of Nile tilapia (Oreochromis niloticus) infected with Aeromonas hydrophila and Pseudomonas fluorescens. Aquaculture 2020, 526, 735392. [Google Scholar] [CrossRef]
- Ceballos-Francisco, D.; Castillo, Y.; De La Rosa, F.; Vásquez, W.; Reyes-Santiago, R.; Cuello, A.; Esteban, M.Á. Bactericidal effect on skin mucosa of dietary guava (Psidium guajava L.) leaves in hybrid tilapia (Oreochromis niloticus × O. mossambicus). Jour. Ethnop. 2020, 259, 112838. [Google Scholar] [CrossRef] [PubMed]
- Aboyadak, I.M.; Ali, N.G.M.; Goda, A.M.A.S.; Saad, W.; Salam, A.M.E. Non-Selectivity of RS media for Aeromonas hydrophila and TCBS media for Vibrio species isolated from diseased Oreochromis niloticus. Jour. Aqua. Res. Devel. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Sumithra, T.G.; Reshma, K.J.; Anusree, V.N.; Sayooj, P.; Sharma, S.R.K.; Suja, G.; Sanil, N.K. Pathological investigations of Vibrio vulnificus infection in Genetically Improved Farmed Tilapia (Oreochromis niloticus L.) cultured at a floating cage farm of India. Aquaculture 2019, 511, 734217. [Google Scholar] [CrossRef]
- Zhu, S.; Kojima, S.; Homma, M. Structure, gene regulation and environmental response of flagella in Vibrio. Front. Microb. 2013, 4, 410. [Google Scholar] [CrossRef] [PubMed]
- Novriadi, R. Vibriosis in aquaculture. Omni-Akuatika 2016, 12, 2–4. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nature Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef]
- Mabrok, M.A.E.; Wahdan, A. The immune modulatory effect of oregano (Origanum vulgare L.) essential oil on Tilapia zillii following intraperitoneal infection with Vibrio anguillarum. Aquac. Intern. 2018, 26, 1147–1160. [Google Scholar] [CrossRef]
- Sebastião, F.A.; Nomura, D.; Sakabe, R.; Pilarski, F. Hematology and productive performance of nile tilapia (Oreochromis niloticus) naturally infected with Flavobacterium columnare. Braz. Jour. Microb. 2011, 42, 282–289. [Google Scholar] [CrossRef]
- Wonmongkol, P.; Sukhavachana, S.; Ampolsak, K.; Srisapoome, P.; Suwanasopee, T.; Poompuang, S. Genetic parameters for resistance against Flavobacterium columnare in Nile tilapia Oreochromis niloticus (Linnaeus, 1758). Jour. Fish Dis. 2018, 41, 321–328. [Google Scholar] [CrossRef]
- Leal, C.A.G.; Carvalho-Castro, G.A.; Sacchetin, P.S.C.; Lopes, C.O.; Moraes, A.M.; Figueiredo, H.C.P. Oral and parenteral vaccines against Flavobacterium columnare: evaluation of humoral immune response by ELISA and in vivo efficiency in Nile tilapia (Oreochromis niloticus). Aquac. Intern. 2010; 18, 657–666. [Google Scholar] [CrossRef]
- Khoi, L.N.D.; Wijngaard, J.; Lutz, C. Farming system practices of seafood production in Vietnam: the case study of Pangasius small-scale farming in the Mekong River Delta. Asean Bus. Case Stud. 2008, 27, 45–69. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). The state of World fisheries and aquaculture. Topics fact sheets. 2016. Available online: http://www.fao.org/3/a-i5798e.pdf (accessed on 2023 February 04).
- Brunton, L.A.; Desbois, A.P.; Garza, M.; Wieland, B.; Mohan, C.V.; Häsler, B.; Nguyen-Viet, H. Identifying hotspots for antibiotic resistance emergence and selection, and elucidating pathways to human exposure: Application of a systems-thinking approach to aquaculture systems. Sci. Total Envir. 2019, 687, 1344–1356. [Google Scholar] [CrossRef]
- Mannan, M.; Islam, S.R.; Osman, M.H.; Rahman, M.K.; Uddin, M.N.; Kamal, M.; Reza, M.S. Antibacterial activity of oxytetracycline on microbial ecology of Nile tilapia (Oreochromis niloticus) gastrointestinal tract under laboratory condition. Aquac. Res. 2020, 51, 2125–2133. [Google Scholar] [CrossRef]
- Singh, S.B.; Young, K.; Silver, L.L. What is an “ideal” antibiotic? Discovery challenges and path forward. Bioch. Pharmac. 2017, 133, 63–73. [Google Scholar] [CrossRef]
- Gastalho, S.; Silva, G.; Ramos, F. Uso de antibióticos em aquacultura e resistência bacteriana: Impacto em saúde pública. Acta Farm. Portug. 2014, 3, 29–45. [Google Scholar]
- Limbu, S.M.; Zhou, L.; Sun, S.X.; Zhang, M.L.; Du, Z.Y. Chronic exposure to low environmental concentrations and legal aquaculture doses of antibiotics cause systemic adverse effects in Nile tilapia and provoke differential human health risk. Envir. Internat. 2018, 115, 205–219. [Google Scholar] [CrossRef]
- Makled, S.O.; Hamdan, A.M.; El-Sayed, A.F.M. Growth promotion and immune stimulation in Nile tilapia, Oreochromis niloticus, fingerlings following dietary administration of a novel marine probiotic, Psychrobacter maritimus S. Prob. Antim. Prot. 2019, 12, 365–374. [Google Scholar] [CrossRef]
- Song, C.; Zhang, C.; Fan, L.; Qiu, L.; Wu, W.; Meng, S.; Chen, J. Occurrence of antibiotics and their impacts to primary productivity in fishponds around Tai Lake, China. Chemosphere 2016, 161, 127–135. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United States) / WHO (World Healt Organization). FAO/WHO expert meeting on foodborne antomicrobial resistance: Role of environment, crops and biocides. 2008. Available online: http://www.fao.org/3/CA0963EN/ca0963en.pdf (accessed on 2023 February 04).
- Lulijwa, R.; Rupia, E.J.; Alfaro, A.C. Antibiotic use in aquaculture, policies and regulation, health and environmental risks: a review of the top 15 major producers. Rev. Aquac. 2020, 12, 640–663. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Antimicrobial usage in aquaculture. 2017. Available online: http://www.fao.org/fi/static-media/MeetingDocuments/WorkshopAMR/presentations/09Lavila Pitogo.pdf (accessed on 2023 February 04).
- FAO (Food and Agriculture Organization of the United Nations). China: Development of nation action plans on AMR: Aquaculture component, project accomplishments and impacts. 2017. Available online: http://www.fao.org/fi/static-media/MeetingDocuments/WorkshopAMR17/presentations/23.pdf (accessed on 2023 February 04).
- Arumugam, U.; Stalin, N.; Rebecca, G.P. Isolation, molecular identification and antibiotic resistance of Enterococcus faecalis from diseased Tilapia. Intern. Jour. Curr. Microb. App. Sci. 2017, 6, 136–146. [Google Scholar] [CrossRef]
- PEIXE BR. Anuário Peixe Br da piscicultura brasileira 2018. São Paulo: Associação Brasileira de Piscicultura. 2018. Available online: https://www.peixebr.com.br/anuario2018/ (accessed on 2023 February 04).
- Monteiro, S.H.; Garcia, F.; Gozi, K.S.; Romera, D.M.; Francisco, J.G.; Moura-Andrade, G.C.; Tornisielo, V.L. Relationship between antibiotic residues and occurrence of resistant bacteria in Nile tilapia (Oreochromis niloticus) cultured in cage-farm. Jour. Envir. Sci. Heal. 2016, 51, 817–823. [Google Scholar] [CrossRef]
- Osman, K.M.; da Silva Pires, Á.; Franco, O.L.; Orabi, A.; Ali, A.H.; Hamada, M.; Elbehiry, A. Nile tilapia (Oreochromis niloticus) as an aquatic vector for Pseudomonas species: Quorum sensing association with antibiotic resistance, biofilm formation and virulence. Aquaculture 2021, 532, 736068. [Google Scholar] [CrossRef]
- Thongkao, K.; Sudjaroen, Y. Beta-lactamase and integron-associated antibiotic resistance genes of Klebsiella pneumoniae isolated from Tilapia fishes (Oreochromis niloticus). Jour.App. Pharm. Sci. 2019, 9, 125–130. [Google Scholar] [CrossRef]
- Dangwetngam, M.; Suanyuk, N.; Kong, F.; Phromkunthong, W. Serotype distribution and antimicrobial susceptibilities of Streptococcus agalactiae isolated from infected cultured tilapia (Oreochromis niloticus) in Thailand: Nine-year perspective. Jour. Med. Microb. 2016, 65, 247–254. [Google Scholar] [CrossRef]
- Hamdan, R.H.; Peng, T.L.; Ong, B.L.; Suhana, M.Y.S.; Hamid, N.H.; Afifah, M.N.F.; Raina, M.S. Antibiotics resistance of Vibrio spp. isolated from diseased seabass and tilapia in cage culture. Procee. Inter. Sem. Lives. Prod. Vet. Tech. 2018, 1, 554–560. [Google Scholar]
- Gao, P.; Mao, D.; Luo, Y.; Wang, L.; Xu, B.; Xu, L. Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res. 2012, 46, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.R.; Santos, F.A.; Ribeiro, A.; Fernandes, C.; Silva, L.H.M.; Gloria, M.B.A. Quinolones and tetracyclines in aquaculture fish by a simple and rapid LC-MS/MS method. Food Chem. 2018, 245, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, P.S.; Gao, D.X.; Wills, R. Preparation of ormetoprim–sulfadimethoxine-medicated discs for disc diffusion assay. North Amer. Jour. Aqua. 2011, 73, 17–20. [Google Scholar] [CrossRef]
- Gaskins, H.R.; Collier, C.T.; Anderson, D.B. Antibiotics as growth promotants: mode of action. Animal Biotec. 2002, 13, 29–42. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.; Abarike, E.D.; Sakyi, M.E.; Lu, Y.; Wang, Z. Modulation of nutrient utilization, growth, and immunity of Nile tilapia, Oreochromis niloticus: the role of probiotics. Aquac. Intern. 2020, 28, 277–291. [Google Scholar] [CrossRef]
- Hasslöf, P.; Stecksén-Blicks, C. Probiotic bacteria and dental caries. In The Impact of Nutrition and Diet on Oral Health; Karger Publishers: Basel, Switzerland, 2020. [Google Scholar]
- Umu, Ö.C.; Rudi, K.; Diep, D.B. Modulation of the gut microbiota by prebiotic fibres and bacteriocins. Mic. Ecol. Heal. Disea. 2017, 28, 1348886. [Google Scholar] [CrossRef]
- Musa, H.H.; Wu, S.L.; Zhu, C.H.; Seri, H.I.; Zhu, G.Q. The potential benefits of probiotics in animal production and health. Jour. Ani. Vet. Adv. 2009, 8, 313–321. [Google Scholar]
- Begum, N.; Islam, M.S.; Haque, A.K.M.F.; Suravi, I.N. Growth and yield of monosex tilapia Oreochromis niloticus in floating cages fed commercial diet supplemented with probiotics in freshwater pond, Sylhet. Bangl. Jour. Zool. 2017, 45, 27–36. [Google Scholar] [CrossRef]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, R.G. Animal Nutrition, 7th edition; Pearson Education Limited: Harlow, England, 2011. [Google Scholar]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.M.; Dequenne, I.; Cani, P.D. How probiotics affect the microbiota. Front. Cell. Infec. Microb. 2020, 9, 454. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, G.; Nandi, A.; Ray, A.K. Assessment of hemolytic activity, enzyme production and bacteriocin characterization of Bacillus subtilis LR1 isolated from the gastrointestinal tract of fish. Arch. Microb. 2017, 199, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Eweedah, N.M.; Moustafa, E.M.; Farahat, E.M. Probiotic effects of Aspergillus oryzae on the oxidative status, heat shock protein, and immune related gene expression of Nile tilapia (Oreochromis niloticus) under hypoxia challenge. Aquaculture 2020, 520, 734669. [Google Scholar] [CrossRef]
- Tan, H.Y.; Chen, S.W.; Hu, S.Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish & Shellf. Immun. 2019, 92, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Srichaiyo, S. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Abarike, E.D.; Cai, J.; Lu, Y.; Yu, H.; Chen, L.; Jian, J.; Kuebutornye, F.K. Effects of a commercial probiotic BS containing Bacillus subtilis and Bacillus licheniformis on growth, immune response and disease resistance in Nile tilapia, Oreochromis niloticus. Fish & Shellf. Immun. 2018, 82, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Gobi, N.; Vaseeharan, B.; Chen, J.C.; Rekha, R.; Vijayakumar, S.; Anjugam, M.; Iswarya, A. Dietary supplementation of probiotic Bacillus licheniformis Dahb1 improves growth performance, mucus and serum immune parameters, antioxidant enzyme activity as well as resistance against Aeromonas hydrophila in tilapia Oreochromis mossambicus. Fish & Shellf. Immun. 2018, 74, 501–508. [Google Scholar] [CrossRef]
- Selim, K.M.; Reda, R.M. Improvement of immunity and disease resistance in the Nile tilapia, Oreochromis niloticus, by dietary supplementation with Bacillus amyloliquefaciens. Fish & Shellf. Immun. 2015, 44, 496–503. [Google Scholar] [CrossRef]
- Efendi, Y. Bacillus subtilis strain VITNJ1 potential probiotic bacteria in the gut of tilapia (Oreochromis niloticus) are cultured in floating net, Maninjau lake, West Sumatra. Pak. Jour. Nut. 2014, 13, 710–715. [Google Scholar] [CrossRef]
- Tachibana, L.; Telli, G.S.; de Carla Dias, D.; Gonçalves, G.S.; Ishikawa, C.M.; Cavalcante, R.B.; Ranzani-Paiva, M.J.T. Effect of feeding strategy of probiotic Enterococcus faecium on growth performance, hematologic, biochemical parameters and non-specific immune response of Nile tilapia. Aquac. Rep. 2020, 16, 100277. [Google Scholar] [CrossRef]
- Chu, T.W.; Chen, C.N.; Pan, C.Y. Antimicrobial status of tilapia (Oreochromis niloticus) fed Enterococcus avium originally isolated from goldfish intestine. Aquac. Rep. 2020, 17, 100397. [Google Scholar] [CrossRef]
- Samson, J.S.; Choresca, C.H.; Quiazon, K.M.A. Selection and screening of bacteria from African nightcrawler, Eudrilus eugeniae (Kinberg, 1867) as potential probiotics in aquaculture. Wor. Jour. Microb. Biotec. 2020, 36, 1–10. [Google Scholar] [CrossRef]
- Poolsawat, L.; Li, X.; He, M.; Ji, D.; Leng, X. Clostridium butyricum as probiotic for promoting growth performance, feed utilization, gut health and microbiota community of tilapia (Oreochromis niloticus × O. aureus). Aquac. Nut. 2020, 26, 657–670. [Google Scholar] [CrossRef]
- Guimarães, M.C.; Dias, D.C.; Araujo, F.V.A.P.; Ishikawa, C.M.; Tachibana, L. Probiotic Bacillus subtilis and Lactobacillus plantarum in diet of nile tilapia. Bol. Inst. Pesca 2019, 45, 1–7. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Ani. Nut. 2020, 6, 69–79. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Tapingkae, W.; Seel-Audom, M.; Jaturasitha, S.; Dawood, M.A.; Esteban, M.Á. Boosted growth performance, mucosal and serum immunity, and disease resistance Nile tilapia (Oreochromis niloticus) fingerlings using corncob-derived xylooligosaccharide and Lactobacillus plantarum CR1T5. Prob. Antim. Prot. 2019, 12, 400–411. [Google Scholar] [CrossRef]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish & Shellf. Immun. 2019, 84, 695–703. [Google Scholar] [CrossRef]
- Sewaka, M.; Trullas, C.; Chotiko, A.; Rodkhum, C.; Chansue, N.; Boonanuntanasarn, S.; Pirarat, N. Efficacy of synbiotic Jerusalem artichoke and Lactobacillus rhamnosus GG-supplemented diets on growth performance, serum biochemical parameters, intestinal morphology, immune parameters and protection against Aeromonas veronii in juvenile red tilapia (Oreochromis spp.). Fish & Shellf. Immun. 2019, 86, 260–268. [Google Scholar] [CrossRef]
- Kaktcham, P.M.; Temgoua, J.B.; Zambou, F.N.; Diaz-Ruiz, G.; Wacher, C.; de Lourdes Pérez-Chabela, M. In vitro evaluation of the probiotic and safety properties of bacteriocinogenic and non-bacteriocinogenic lactic acid bacteria from the intestines of nile tilapia and common carp for their use as probiotics in aquaculture. Prob. Antim. Prot. 2018, 10, 98–109. [Google Scholar] [CrossRef]
- Rahman, M.M.; Aktar, S.; Faruk, M.O.; Uddin, M.S.; Ferdouse, J.; Anwar, M.N. Probiotic potentiality of Lactobacillus coryniformis subsp. Torquens MTi1 and Lactobacillus coryniformis MTi2 isolated from intestine of Nile tilapia: An in vitro evaluation. Jour. Pure App. Microb. 2018, 12, 1037–1045. [Google Scholar] [CrossRef]
- Addo, S.; Carrias, A.A.; Williams, M.A.; Liles, M.R.; Terhune, J.S.; Davis, D.A. Effects of Bacillus subtilis strains on growth, immune parameters, and Streptococcus iniae susceptibility in Nile tilapia, Oreochromis niloticus. Jour. World Aquac. Soc. 2017, 48, 257–267. [Google Scholar] [CrossRef]
- Adeoye, A.A.; Yomla, R.; Jaramillo-Torres, A.; Rodiles, A.; Merrifield, D.L.; Davies, S.J. Combined effects of exogenous enzymes and probiotic on Nile tilapia (Oreochromis niloticus) growth, intestinal morphology and microbiome. Aquaculture 2016, 463, 61–70. [Google Scholar] [CrossRef]
- Reda, R.M; Selim, K.M. Evaluation of Bacillus amyloliquefaciens on the growth performance, intestinal morphology, hematology and body composition of Nile tilapia, Oreochromis niloticus. Aquac. Intern. 2015, 23, 203–217. [Google Scholar] [CrossRef]
- Han, B.; Long, W.Q.; He, J.Y.; Liu, Y.J.; Si, Y.Q.; Tian, L.X. Effects of dietary Bacillus licheniformis on growth performance, immunological parameters, intestinal morphology and resistance of juvenile Nile tilapia (Oreochromis niloticus) to challenge infections. Fish & Shellf. Immun. 2015, 46, 225–231. [Google Scholar] [CrossRef]
- Ayyat, M.S.; Labib, H.M.; Mahmoud, H.K. A probiotic cocktail as a growth promoter in Nile tilapia (Oreochromis niloticus). Jour. App. Aquac. 2014, 26, 208–215. [Google Scholar] [CrossRef]
- Telli, G.S.; Ranzani-Paiva, M.J.T.; de Carla Dias, D.; Sussel, F.R.; Ishikawa, C.M.; Tachibana, L. Dietary administration of Bacillus subtilis on hematology and non-specific immunity of Nile tilapia Oreochromis niloticus raised at different stocking densities. Fish & Shellf. Immun. 2014, 39, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.H.; Refat, N.A.A. Pathological evaluation of probiotic, Bacillus subtilis, against Flavobacterium columnare in Tilapia nilotica (Oreochromis niloticus) fish in Sharkia Governorate, Egypt. The Jour. Amer. Sci. 2011, 7, 244–256. [Google Scholar]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: an update on their multiple modes of action: A review. Vet. Quar. 2016, 36, 228–241. [Google Scholar] [CrossRef]
- Chauhan, A.; Singh, R. Probiotics in aquaculture: a promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish & Shellf. Immun 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Farizky, H.S.; Satyantini, W.H.; Nindarwi, D.D. The efficacy of probiotic with different storage to decrease the total organic matter, ammonia, and total Vibrio on shrimp pond water. E&ES 2020, 441, 012108. [Google Scholar] [CrossRef]
- Elsabagh, M.; Mohamed, R.; Moustafa, E.M.; Hamza, A.; Farrag, F.; Decamp, O.; Eltholth, M. Assessing the impact of Bacillus strains mixture probiotic on water quality, growth performance, blood profile and intestinal morphology of Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2018, 24, 1613–1622. [Google Scholar] [CrossRef]
- Thomas, C.M.; Versalovic, J. Probiotics-host communication: modulation of signaling pathways in the intestine. Gut Mic. 2010, 1, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef] [PubMed]
- Oelschlaeger, T.A. Mechanisms of probiotic actions–a review. Intern. Jour. Med. Micr. 2010, 300, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Opiyo, M.A.; Jumbe, J.; Ngugi, C.C.; Charo-Karisa, H. Different levels of probiotics affect growth, survival and body composition of Nile tilapia (Oreochromis niloticus) cultured in low input ponds. Sci. Afr. 2019, 4, 103. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microb. 2018, 9, 2429. [Google Scholar] [CrossRef]
- Dias, D.D.C.; Furlaneto, F.D.P.B.; Sussel, F.R.; Tachibana, L.; Gonçalves, G.S.; Ishikawa, C.M.; Ranzani-Paiva, M.J.T. Economic feasibility of probiotic use in the diet of Nile tilapia, Oreochromis niloticus, during the reproductive period. Acta Scien., Ani. Sci. 2020, 42, 47960. [Google Scholar] [CrossRef]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Ani. Nut. 2017, 3, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Ismail, Y.S.; Yulvizar, C.; Mazhitov, B. Characterization of lactic acid bacteria from local cow’s milk kefir. IOP Conf. Ser.: Ear. Envir. Sci. 2018, 130, 012019. [Google Scholar] [CrossRef]
- Da Silva, H.R.; do Nascimento, R.C.V.; Talma, S.V.; de Carvalho Furtado, M.; Balieiro, A.L.; Barbosa, J.B. Aplicações tecnológicas de Bactérias do Ácido Lático (BALs) em produtos lácteos. Ver. INGI-Ind. Geog. Inov. 2020, 4, 681–690. [Google Scholar]
- Ewing, W.N. The living gut; Nottingham University Press: Nottingham, United Kingdom, 2008. [Google Scholar]
- Pacheco, K.D.; Del’Duca, A.; Borges, M.L.; Fernandes, R.T.; Cesar, D.E.; Apolônio, A.C.M. Bacteriocin-like inhibitory substance in aquaculture: a classic method of protein precipitation for a new applicability. Acta Sci. Biol. Sci. 2018, 40, e37881. [Google Scholar] [CrossRef]
- Xia, Y.; Cao, J.; Wang, M.; Lu, M.; Chen, G.; Gao, F.; Yi, M. Effects of Lactococcus lactis subsp. lactis JCM5805 on colonization dynamics of gut microbiota and regulation of immunity in early ontogenetic stages of tilapia. Fish & Shellf. Immun. 2019, 86, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, A.M.; El-Sayed, A.F.M.; Mahmoud, M.M. Effects of a novel marine probiotic, Lactobacillus plantarum AH 78, on growth performance and immune response of Nile tilapia (Oreochromis niloticus). Jour. App. Microb. 2016, 120, 1061–1073. [Google Scholar] [CrossRef]
- Mohamed, M.H.; Ammar, M.A.M. Effect of some antimicrobials on quality and shelf life of freshwater tilapia (Oreochromis niloticus). Jour. Food Proc. Pres. 2020, 45, e15026. [Google Scholar] [CrossRef]
- Kaktcham, P.M.; Piame, L.T.; Sileu, G.M.S.; Kouam, E.M.F.; Temgoua, J.B.; Ngoufack, F.Z.; de Lourdes Pérez-Chabela, M. Bacteriocinogenic Lactococcus lactis subsp. lactis 3MT isolated from freshwater Nile Tilapia: isolation, safety traits, bacteriocin characterisation, and application for biopreservation in fish pâté. Arch. Microb. 2019, 201, 1249–1258. [Google Scholar] [CrossRef]
- Kaktcham, P.M.; Kouam, E.M.F.; Tientcheu, M.L.T.; Temgoua, J.B.; Wacher, C.; Ngoufack, F.Z.; de Lourdes Perez-Chabela, M. Nisin-producing Lactococcus lactis subsp. lactis 2MT isolated from freshwater Nile tilapia in Cameroon: Bacteriocin screening, characterization, and optimization in a low-cost medium. LWT 2019, 107, 272–279. [Google Scholar] [CrossRef]
- Abdelfatah, E.N.; Mahboub, H.H.H. Studies on the effect of Lactococcus garvieae of dairy origin on both cheese and Nile tilapia (O. niloticus). Intern. Jour. Vet. Sci. Med. 2018, 6, 201–207. [Google Scholar] [CrossRef]
- Loh, J.Y.; Lim, Y.Y.; Ting, A.S.Y. Bacteriocin-like substances produced by Lactococcus lactis subsp. lactis CF4MRS isolated from fish intestine: Antimicrobial activities and inhibitory properties. Inter. Food Res. Jour. 2017, 24, 394. [Google Scholar]
- Tasaku, S.; Siripornadulsil, S.; Siripornadulsil, W. Inhibitory activity of food-originated Pediococcus pentosaceus NP6 against Salmonella enterica serovar Typhimurium in Nile tilapia by-products. Chi. Mai Jour. Sci. 2017, 44, 383–393. [Google Scholar]
- Etyemez, M.; Balcazar, J.L. Isolation and characterization of bacteria with antibacterial properties from Nile tilapia (Oreochromis niloticus). Res. Vet. Sci. 2016, 105, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Y.; Yao, J.; Li, W. Inhibition ability of probiotic, Lactococcus lactis, against A. hydrophila and study of its immunostimulatory effect in tilapia (Oreochromis niloticus). Inter. Jour. Eng. Sci. Tech. 2010, 2, 73–80. [Google Scholar] [CrossRef]
Figure 1.
Antibiotic susceptibility of a wide diversity of bacterial isolates associated with mortality outbreaks in cultured Nile tilapia. Among the set of genera, Pseudomonas and Vibrio show a high resistance profile to most of the antibiotics tested, such as penicillins, sulfonamides, quinolones, tetracyclines and beta-lactams. Interestingly, Enterococcus faecalis only shows resistant and intermediate susceptibility to the tested antibiotics, highlighting the need for new alternatives to reduce the associated mortalities. A third group of bacterial species, including Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis exhibit susceptibility to all antibiotics tested, excluding penicillins. The remaining bacteria, including two species of Streptococcus and Aeromonas hydrophila, show a pattern similar to the one described above, although describing a high resistance to sulfonamides, and some intermediate resistance to penicillins and tetracyclines.
Figure 1.
Antibiotic susceptibility of a wide diversity of bacterial isolates associated with mortality outbreaks in cultured Nile tilapia. Among the set of genera, Pseudomonas and Vibrio show a high resistance profile to most of the antibiotics tested, such as penicillins, sulfonamides, quinolones, tetracyclines and beta-lactams. Interestingly, Enterococcus faecalis only shows resistant and intermediate susceptibility to the tested antibiotics, highlighting the need for new alternatives to reduce the associated mortalities. A third group of bacterial species, including Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis exhibit susceptibility to all antibiotics tested, excluding penicillins. The remaining bacteria, including two species of Streptococcus and Aeromonas hydrophila, show a pattern similar to the one described above, although describing a high resistance to sulfonamides, and some intermediate resistance to penicillins and tetracyclines.
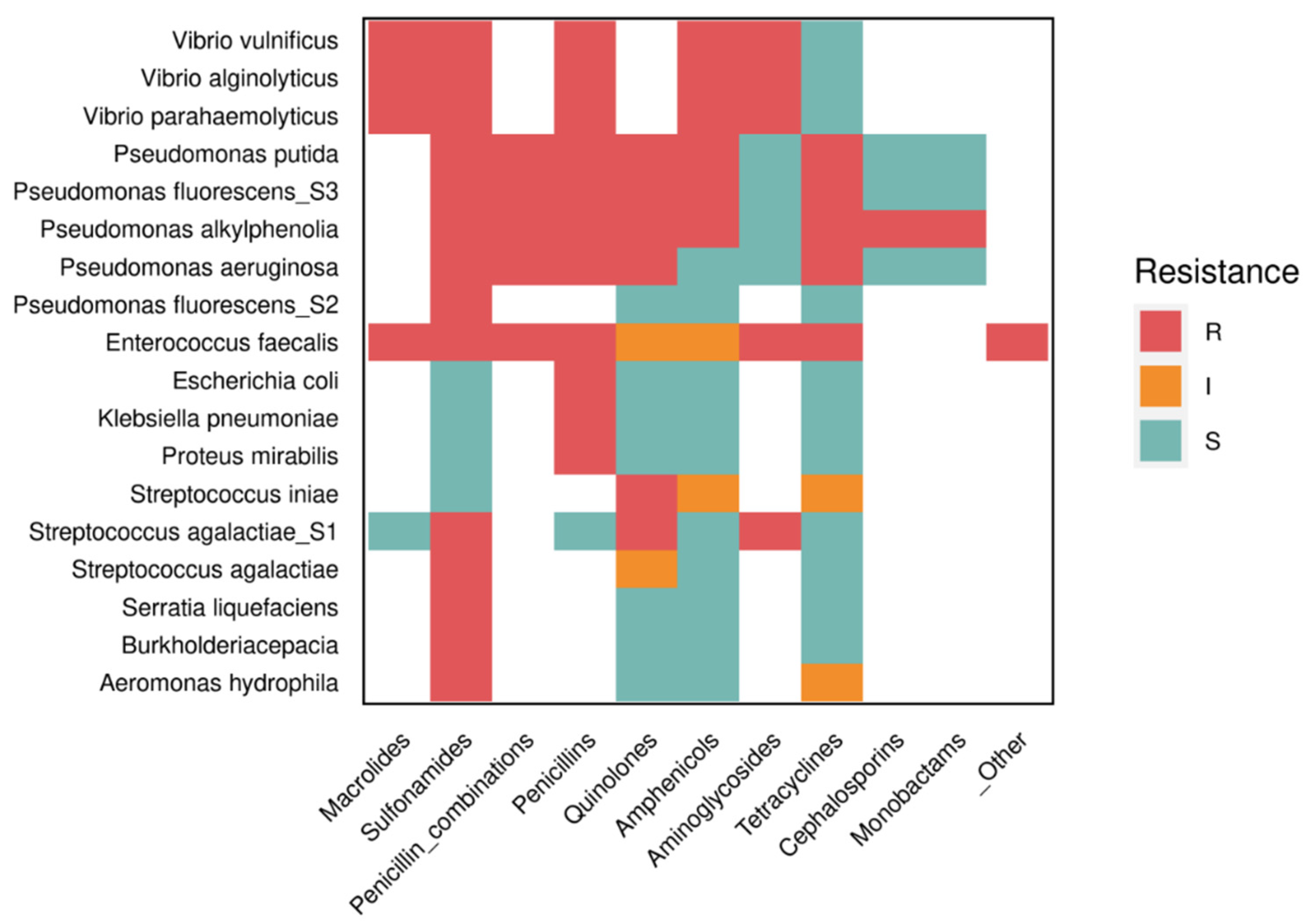
Figure 2.
Effect of probiotic bacteria on the intestinal microbiota and immunological parameters. 1) When probiotics reach the intestine, they start competing for space and nutrients. 2) They produce vitamins and bacteriocins, which inhibit the growth of pathogens and produce 3) digestive enzymes, which improve the host nutrition. 4) Due to their antagonistic effect, they may be associated with Microbe-Associated Molecular Patterns (MAMPs) by Pattern Recognition Receptors (PRRs) and Toll-Like Receptors (TLRs), which lead to the activation of immune system cells. T cells produce cytokines, B cells produce antibodies and they activate phagocytes responsible for neutralizing and destroying pathogens.
Figure 2.
Effect of probiotic bacteria on the intestinal microbiota and immunological parameters. 1) When probiotics reach the intestine, they start competing for space and nutrients. 2) They produce vitamins and bacteriocins, which inhibit the growth of pathogens and produce 3) digestive enzymes, which improve the host nutrition. 4) Due to their antagonistic effect, they may be associated with Microbe-Associated Molecular Patterns (MAMPs) by Pattern Recognition Receptors (PRRs) and Toll-Like Receptors (TLRs), which lead to the activation of immune system cells. T cells produce cytokines, B cells produce antibodies and they activate phagocytes responsible for neutralizing and destroying pathogens.
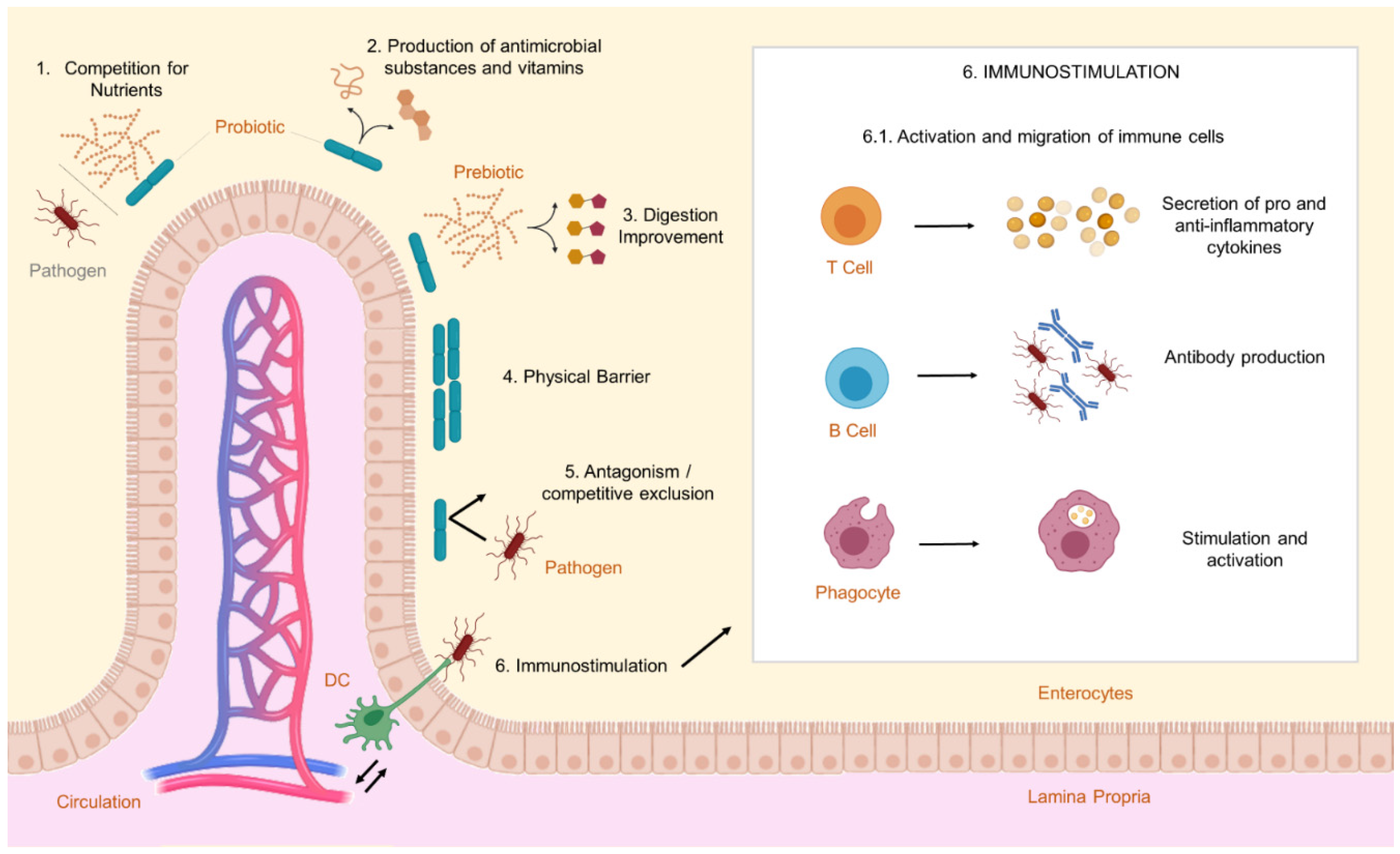
Figure 3.
Tilapia specific growth rate (SGR) versus ln of probiotic dose in the feed. Regression analysis revealed no significant (p > 0.05) relationship. Circles represent the mean of considered experimental groups.
Figure 3.
Tilapia specific growth rate (SGR) versus ln of probiotic dose in the feed. Regression analysis revealed no significant (p > 0.05) relationship. Circles represent the mean of considered experimental groups.
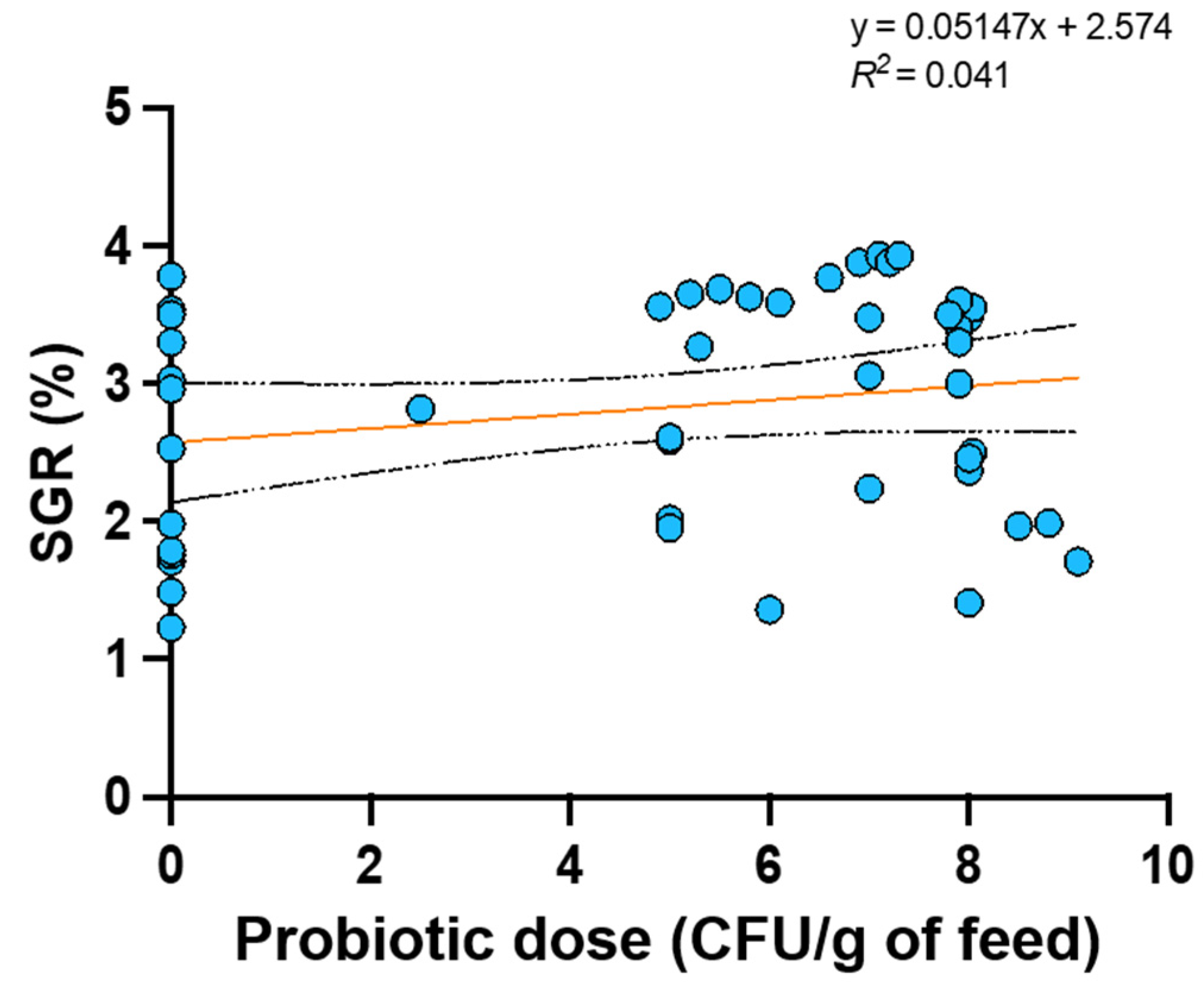
Figure 4.
Feed conversion ratio (FCR) versus ln of probiotic dose in the feed. Regression analysis revealed no significant (p > 0.05) relationship. Circles represent the mean of considered experimental groups.
Figure 4.
Feed conversion ratio (FCR) versus ln of probiotic dose in the feed. Regression analysis revealed no significant (p > 0.05) relationship. Circles represent the mean of considered experimental groups.
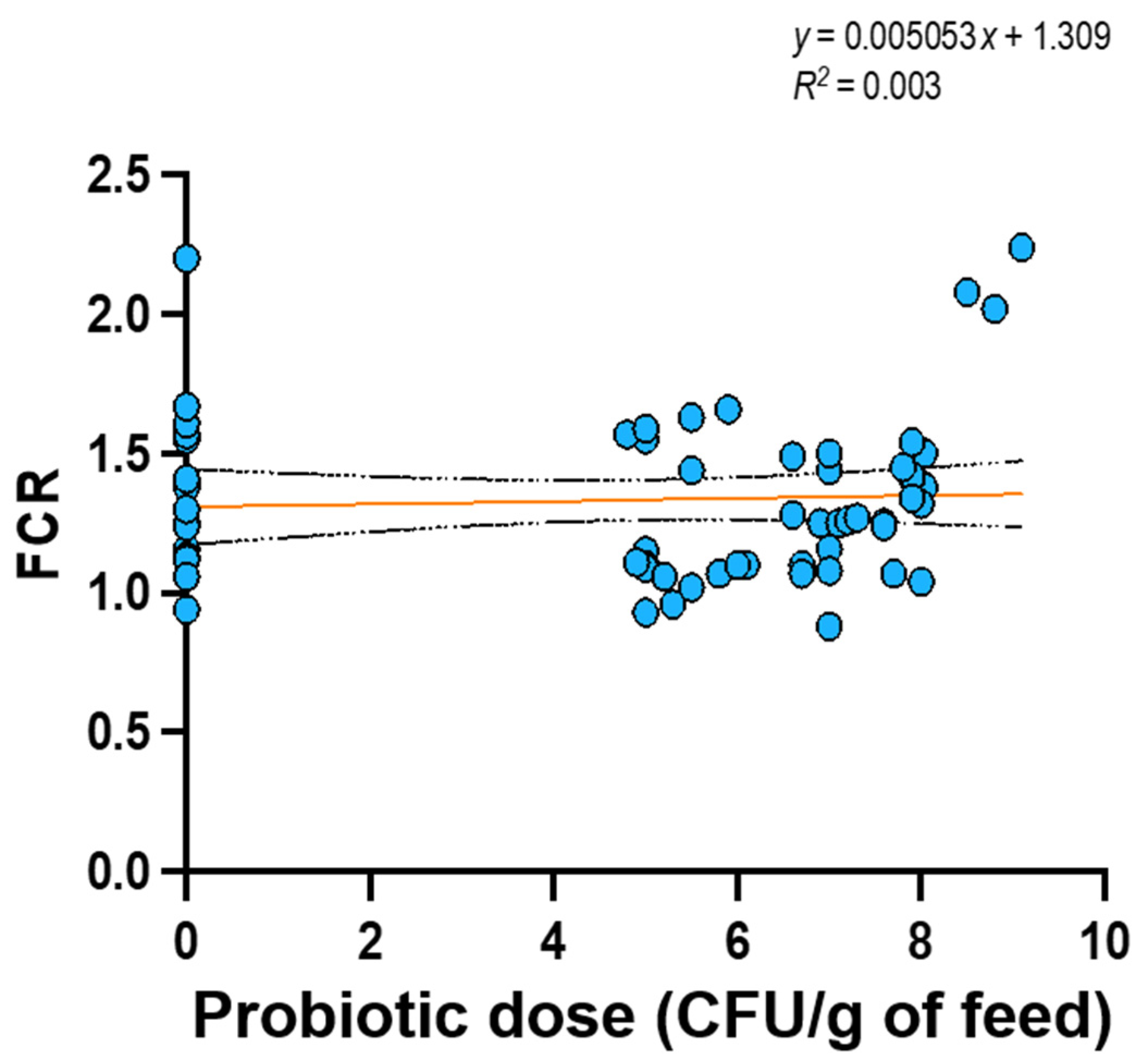
Figure 5.
Pearson correlation matrix between variables of interest and tilapia specific growth rate. ** p < 0.01.
Figure 5.
Pearson correlation matrix between variables of interest and tilapia specific growth rate. ** p < 0.01.
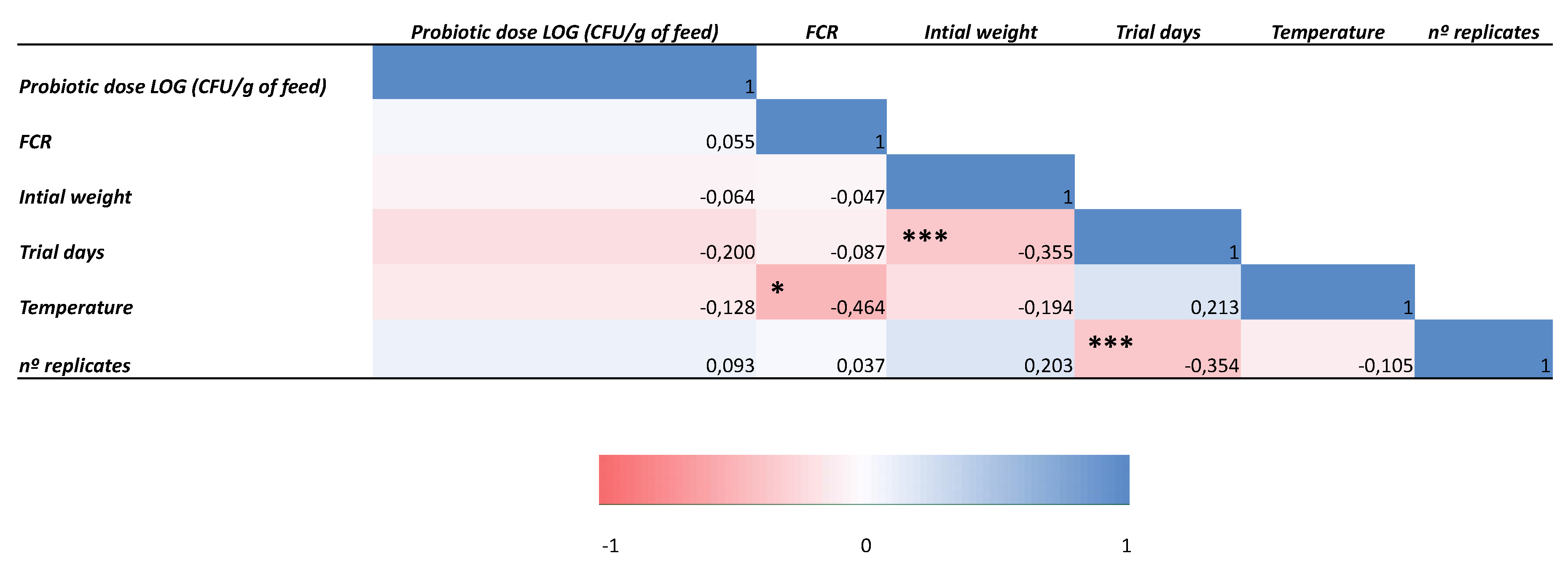
Figure 6.
Pearson correlation matrix between variables of interest and tilapia feed conversion ratio. * p < 0.05, *** p < 0.001.
Figure 6.
Pearson correlation matrix between variables of interest and tilapia feed conversion ratio. * p < 0.05, *** p < 0.001.
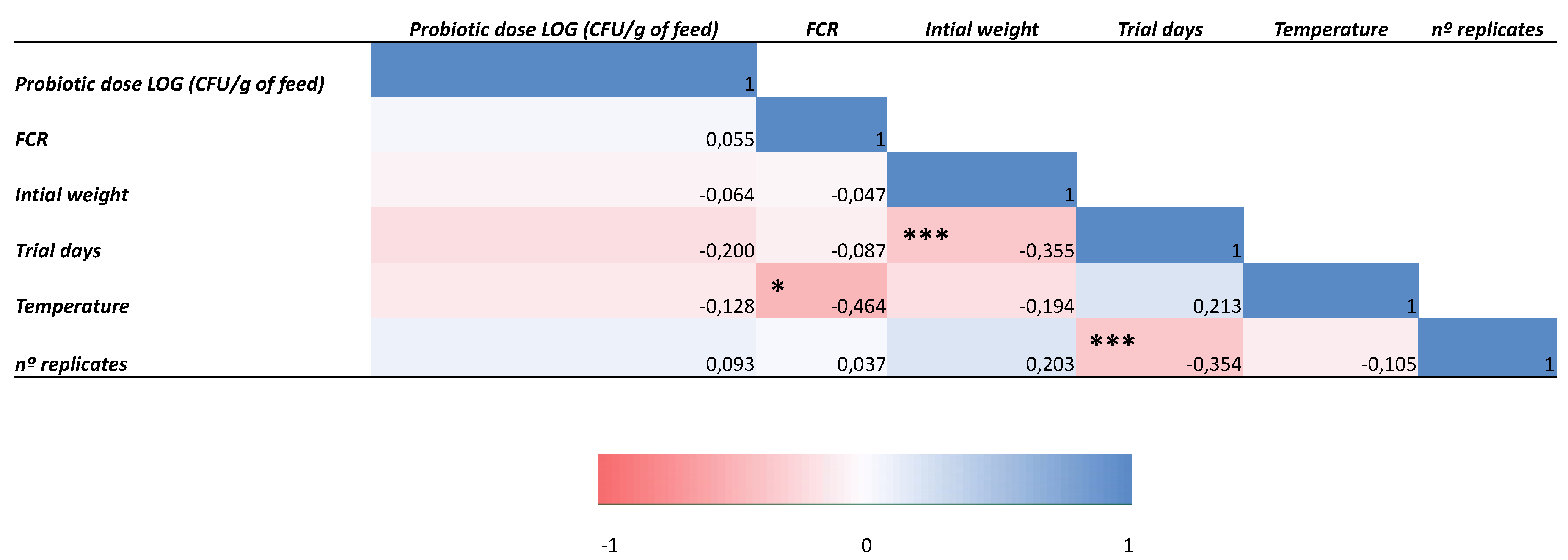

Table 1.
Production systems and technological resources employed by the 12 largest tilapia producers.
Table 1.
Production systems and technological resources employed by the 12 largest tilapia producers.
| Country | Production in 2018 (milliontons) | Installations | Integration with other economic activities | Other technologies | Reference |
| China | 1.86 | Floating cage (high-density), net, ponds (in hydroelectric reservoirs) | Polyculture with carp, mullet or shrimp; rice culture | Hydroponics, GIFTs and ProGIFT*, RAS (Recirculating systems), hatcheries | [17,20,21,24] |
| Indonesia | 1.25 | Floating net cage, two-net cage | Polyculture with carp; rice culture | Biofloc technology, RAS, GIFT and other genetic improved tilapias, nanobubble technology, dual-cage | [17,20] |
| Egypt | 0.86 | Pond-farm, tank, earthen-ponds | Polyculture with mullet; rice culture | RAS, In-Pond Raceway System (IPRS), dual-cage, aquaponics, hatcheries, improved-feeds, GIFT and GIANT*, seed production, Automated Monitoring and Control System (AMCS) | [17,20] |
| Brazil | 0.40 | Earthen-ponds, tank-net, cages (in hydroelectric reservoirs, high-density), periphyton pond | Polyculture with pirapitinga or shrimp | Aerator, automatic feeding, Biofloc technology, GIFTs and GST | [17,20,46,121] |
| Philippines | 0.33 |
Earthen-ponds, floating cages and fixed cages, tank | Most farmers adopt monoculture system; however, there is integration with swine, rabbit and poultry cultures in lesser extent | GIFT and other genetic improved tilapias, monosex tilapia, supermale technology, Biofloc technology | [12,17,20] |
| Thailand | 0.32 | Floating-cages (high-density), Bamboo cages | Integration with poultry culture; rice culture | GIFT and other genetic improved tilapias, supermale technology, improved seaweed | [17,20] |
| Bangladesh | 0.22 | Pond-dike, cages | Polyculture with carp, rice culture (rotational) | GIFT, feed supplements, improved seeds, water-saving technologies | [17,20] |
| Vietnam | 0.20 | Cages, net | Polyculture with silver barb and carp or shrimp, rice culture | RAS, GIFT | [17,20] |
| Colombia | 0.077 | Cages (in hydroelectric reservoirs, high-density), pond, tanks, raceways | Polyculture with carp or bocachico | Improved seeds, Biofloc technology | [2,17] |
| Uganda | 0.070 | Earthen-ponds, Cages (low-density), cage/pens, tank/raceways | Most farmers adopt monoculture system; however there is integration between farming/aquaculture activities | Hatcheries, improved seeds | [2,17] |
| Taiwan | 0.062 | Cages, octogonal tanks/raceway ponds | Polyculture with shrimp | Aerator, automatic feeders, RAS | [2,17] |
| Mexico | 0.052 | Net-pens, cages | Polyculture with Mayan cichlids shrimp or prawn | RAS, Biofloc technology | [2,17] |
| China | 1.86 | Floating cage (high-density), net, ponds (in hydroelectric reservoirs) | Polyculture with carp, mullet or shrimp; rice culture | Hydroponics, GIFTs and ProGIFT*, RAS (Recirculating systems), hatcheries | [17,20,21,24] |
*GIANT and ProGIFTs are types of genetic improved tilapia.
Table 2.
Tilapia pathogens that showed resistance to antibiotics and other antimicrobials used in farming systems in some countries.
Table 2.
Tilapia pathogens that showed resistance to antibiotics and other antimicrobials used in farming systems in some countries.
| Resistant Pathogen | Ineffective antibiotic (resistance) | Tilapia origin | Reference | Most common antibiotics detected in the country | Reference | Antibiotic allowed in the country (active principle) | Reference |
| Pseudomonas aeruginosa | Ampicillin, sulfamethoxazole/trimethoprim, tetracycline and nalidixic acid | Giza (Egypt) | [123] | Ciprofloxacin and florfenicol | [117] | Florfenicol, ciprofloxacin | [117] |
| Klebsiella pneumoniae, Escherichia coli, Proteus mirabilis, and Streptococcus agalactiae | Penicillin, ampicillin, oxytetracyclines, trimethoprim, oxolinic acid, gentamicin, and sulfamethoxazole | Bangkok and Nakhon Si Thammarat (Thailand) | [124,125] | Enrofloxacin, norfloxacin, amoxicillin, oxolinic acid, penicillin, florfenicol, tetracycline, oxytetracycline, sulfadiazine, trimethoprim, ormetoprim, sulfadiazine + trimethoprim, sulfadimethoxine + trimethoprim, sulfaguanidine | [117,118] | Oxytetracycline, tetracycline, sulfadimethozine, trimethoprim, sulfadimethoxine-ormethoprim and amoxicillin | [117] |
| Vibrio spp. | Erythromycin and chloramphenicol | Sri Tujuh (Malaysia) | [126] | Oxolinic acid, virginiamycin, chloramphenicol and sulfonamides, tetracyclines, nitrofurans* | [117,118] | Amoxicillin, oxytetracycline, flumequine and florfenicol, oxolinic acid, virginiamycin and tetracyclines | [117,118] |
| Enterococcus spp. | Tetracycline | Chennai (India) | [120] | Erythromycin, chloramphenicol, sulfadiazine, sulfadimethoxine, sulfamethazine, sulfapyridine, sulfamethoxypyridazine, sulfadoxine, sulfamethoxazole, sulfanilamide, sulfathiazole | [117] | Sulfadimethoxin, sulfabromomethazin, erythromycin, oxytetracycline, althrocin, ampicillin, sparfloxacin, and enrofloxacin and sulfaethoxypyridazine | [117,120] |
| Aeromonas hydrophila | Tetracycline, sulfathiazole | Solteira Island (Brazil) | [122] | Florfenicol, tetracycline, oxytetracycline and enrofloxacin** | [117] | Florfenicol, oxytetracycline and neomycin (ornamental fish) | [20,119,121,122] |
| Acinetobacter spp. | Sulfamethoxazole, tetracycline | Tianjin (China) | [127] | Neomycin sulphate, doxycycline hydrochloride, thiamphenicol, florfenicol, sulfadiazine, sulfamethoxazole + trimethoprim, sodium sulfamonomethoxine, enrofloxacin, flumequine, oxolinic acid, oxytetracycline, ciprofloxacin, norfloxacin, ofloxacin, amoxicillin, cephalexin, cefradine, cefotaxime, erythromycin, gentamicin S, neomycin, tetracycline, lycomicin, sulfamethoxazole*** | [117,119] | Neomycin sulfate, doxycycline hydrochloride, thiamphenicol, florfenicol, sulfadiazine, sulfamethoxazole + trimethropim, sodium sulfamonomethoxin, enrofloxacin, flumequine, oxolinic acid, oxytetracycline |
[119] |
| Pseudomonas aeruginosa | Ampicillin, sulfamethoxazole/trimethoprim, tetracycline and nalidixic acid | Giza (Egypt) | [123] | Ciprofloxacin and florfenicol | [117] | Florfenicol, ciprofloxacin | [117] |
| Klebsiella pneumoniae, Escherichia coli, Proteus mirabilis, and Streptococcus agalactiae | Penicillin, ampicillin, oxytetracyclines, trimethoprim, oxolinic acid, gentamicin, and sulfamethoxazole | Bangkok and Nakhon Si Thammarat (Thailand) | [124,125] | Enrofloxacin, norfloxacin, amoxicillin, oxolinic acid, penicillin, florfenicol, tetracycline, oxytetracycline, sulfadiazine, trimethoprim, ormetoprim, sulfadiazine + trimethoprim, sulfadimethoxine + trimethoprim, sulfaguanidine | [117,119] | Oxytetracycline, tetracycline, sulfadimethozine, trimethoprim, sulfadimethoxine-ormethoprim and amoxicillin | [117] |
| Vibrio spp. | Erythromycin and chloramphenicol | Sri Tujuh (Malaysia) | [126] | Oxolinic acid, virginiamycin, chloramphenicol and sulfonamides, tetracyclines, nitrofurans* | [117,119] | Amoxicillin, oxytetracycline, flumequine and florfenicol, oxolinic acid, virginiamycin and tetracyclines | [117,119] |
*According to Lulijwa et al. [117] report, there were no updated data showing specifically which drugs are used in farming systems in Malaysia. The FAO survey [118] mainly registered most used classes of antibiotics. ** Although enrofloxacin is not allowed in aquaculture in Brazil [128], some studies [117] have reported this antibiotic in fish samples. *** According to FAO [119], only 13 antibiotics are allowed in Chinese aquaculture; however, Lulijwa et al. [117] reported that 33 different drugs were detected in farming systems, some of the most recurrent of which are listed in this table.
Table 3.
Enzymes produced by probiotics and health impacts observed in studies performed on tilapia.
Table 3.
Enzymes produced by probiotics and health impacts observed in studies performed on tilapia.
| Tilapia species | Probiotic | Concentration | Duration | Secreted enzyme | Results | Reference |
| Oreochromis niloticus | Aspergillus oryzae | 106 and 108 CFU/g | 60 days | PR, AP, LY, SD and CA (Results obtained under hypoxia) |
IR, AA, PP, FCR, GR ↑; BC, SR, AN←; CH, PC, ROS↓ | [139] |
| Oreochromis niloticus | Rummeliibacillus stabekisii | 107 CFU/g | 7 days | PR, CE, AM and XY | GR, FE, WG ↑ | [140] |
| Oreochromis spp. | Lactobacillus plantarum, Bacillus velezensis | 108 CFU/g- Lb. plantarum 107 CFU/g- B. velezensis |
15 and 30 days | PE and LY | IR, GR, DR ↑ | [141] |
| Oreochromis niloticus | Bacillus spp. | 107 CFU/g | 14 days | LY, SD, CA, MPO and AP | WG, GR, IR, DR, FCR, PP↑ | [142] |
| Oreochromis niloticus | Lactobacillus plantarum (KC426951) | 105 and 107 CFU/g | 14 and 28 days | ALP, MPO and LY | WG, GR, IR, DR, FCR, PP, ROS↑ | [143] |
| Oreochromis niloticus | Bacillus amyloliquefaciens | 104 and 106 CFU/g | 56 days | LY | IR, DR, PP ↑ | [144] |
| Oreochromis niloticus | Bacillus subtilis V1TNJ1 | Not specified | Not specified | PR | PI ↑ | [145] |
Results: Significant change ←; No significant change →; Increase/Growth ↑; Decrease/Reduction ↓. Parameters: Antioxidants activity (AA), Anti-protease activity (AP), Bacterial community (BC), Cholesterol (CH), Disease resistance (DR), Feed conversion ratio (FCR), Feed efficiency (FE), Growth rate (GR), Immune response (IR), Plasma cortisol (PC), Plasma glucose (PG), Protection against pathogen (PP), Proteolysis index (PI), Reactive oxygen species (ROS), Survival rate (SR), Weight gain (WG). Enzymes: Amylase (AM), Catalase (CA), Cellulase (CE), Lysozyme (LY), Myeloperoxidase (MPO), Peroxidase (PE), Protease (PR), Superoxide dismutase (SD), Xylanase (XY).
Table 4.
Probiotic effects against some pathogens observed in tests performed with tilapia.
| Tilapia species | Probiotic | Concentration | Pathogen | Duration | Results | Reference |
| Oreochromis niloticus | Enterococcus faecium | 1010 CFU/g | Aeromonas hydrophila | 84 days (CON); 7 days (P7); 14 days (P14) | SR, HE, HM, PG, PC, MO (regardless of the period) →; PP (CON)↑; GR, WG (P7) ↑; RB (P14) ← | [146] |
| Oreochromis niloticus | Enterococcus avium | 107, 108, 109 and 1010 CFU/g | Streptococcus agalactiae | 7, 14 and 21 days | SR, AM, PR, LA ↑ (107 CFU/g during 7 days) | [147] |
| Oreochromis niloticus | Lactobacillus plantarum | 1.02 × 109 CFU/mL/kg | Enterococcus faecalis | 56 days | GR →; IR (innate), DR, IG (cytokines), SR↑; MO↓; BC← | [15] |
| Oreochromis niloticus | Bacillus spp. | 107 and 108 CFU/g | Aeromonas hydrophila, Micrococcus luteus, Pseudomonas fluorescens, Enterococcus faecalis, Streptococcus agalactiae | 14 days | SR↑ DE← | [148] |
| Oreochromis niloticus | Aspergillus oryzae | 106 and 108 CFU/g | Aeromonas hydrophila | 60 days | FCR, GR, GLx, IR (immunoglobulin M), SR, LY, PP, TP, PA, CA↑; AN←; PG, CH↓ | [139] |
| Hybrid tilapia (Oreochromis niloticus x Oreochromis aureus) | Clostridium butyricum | 1.50 × 108 CFU/g | Aeromonas hydrophila | 56 days | PRE, LR, ADC, VH, GR, FCR↑; BC←; MO↓ | [149] |
| Oreochromis niloticus | Bacillus subtilis and Lactobacillus plantarum | 1.51 × 106 CFU/g for Lb. plantarum 1.34 × 107 CFU/g for B. subtilis |
Aeromonas hydrophila | 28 days | GR, SR, IG →; BC← | [150] |
| Oreochromis niloticus | Bacillus cereus NY5 and Bacillus subtilis | 108 CFU/g | Streptococcus agalactiae | 42 days | WG, FCR, SR→ (only B. subtilis); FCR↑ (B. cereus alone, and B. cereus + B. subtilis); DR, LY, ML, MD↑; BC← | [151] |
| Oreochromis niloticus | Rummeliibacillus stabekisii | 107 CFU/g | Streptococcus iniae and Aeromonas hydrophila | 56 days | WG, FCR, GR, FE, DE, SR, DR, IR, IG (cytokines)↑; PA, RB, LY← | [140] |
| Oreochromis niloticus | Lactococcus lactis subsp. lactis JCM5805 | 104 CFU/mL (T1) and 108 CFU/mL (T2) | Streptococcus agalactiae | 15 days | DR, GR, IG, SR↑ (only in T2); BC ← | [151] |
| Oreochromis niloticus | Lactobacillus plantarum CR1T5 | 108 CFU/g | Streptococcus agalactiae | 84 days | WG, GR, FCR, PA, PE, RB, LY, IR↑; SM←; SR→ | [152] |
| Oreochromis niloticus | Paenibacillus ehimensis NPUST1 | 106 and 107 CFU/g | Streptococcus iniae and Aeromonas hydrophila | 70 days | WG, FCR, FE, PA, RB, SD, LY, IG (TNF-α and IL-1β), PY, LA, AM, PR↑ | [153] |
| Oreochromis spp. | Lactobacillus rhamnosus | 108 CFU/g | Aeromonas veronii | 30 days | GR, FCR, LY, WG, CH, PG, VH, VW, GC, AB, MP↑; AL, TR, AST→; ALT, BUN, MO↓ | [154] |
| Oreochromis niloticus | Lactococcus lactis | Not specified | Staphylococcus spp., Vibrio spp., Pseudomonas aeruginosa | Not specified | AN ← | [155] |
| Oreochromis niloticus | Lactococcus coryniformis subsp. torquens MTi1 and MTi2 | 1.50 × 108 CFU/mL | Escherichia coli, Salmonella typhi, Pseudomonas aeruginosa, Staphylococcus aureus | Not specified | AN← | [156] |
| Oreochromis niloticus | Bacillus spp. | 107 CFU/g | Streptococcus agalactiae | 30 days | GR, WG, LY, PR, CA, SD, ALP, MPO, ROS, GC, PP↑; IR, IG ← MO↓ | [142] |
| Oreochromis niloticus | Bacillus licheniformis | 105 and 107 CFU/g | Aeromonas hydrophila | 14 or 28 days | GR, WG, FCR, IR, DR, ROS↑; ALP, LY← | [143] |
| Oreochromis niloticus | Bacillus subtilis | 3.9 × 107 CFU per fish | Aeromonas hydrophila | 56 days | WG, GR, FCR, LY, RB→; SR, PP↑; MO↓ (even lower when probiotic was combined with Previda® prebiotic) | [94] |
| Oreochromis niloticus | Bacillus subtilis | 4 x 107 CFU/g | Streptococcus iniae | 21 days | GR→; AN, LY↑; MO↓ | [157] |
| Oreochromis niloticus | Lactobacillus plantarum AH78 | 1010 CFU/mL | Aeromonas hydrophila | 40 days | GR, IR, FCR, AL, GLx↑; IG (cytokines), VH, BC, ABA, ← | [126] |
| Oreochromis niloticus | Bacillus subtilis, Bacillus licheniformis and Bacillus pumilus | Not specified | Cetobacterium spp. and Plesiomonas spp. | 49 days | GR, FCR, LY, GC, WG, ABA, VH↑ | [158] |
| Oreochromis niloticus | Bacillus amyloliquefaciens | 104 and 106 CFU/g | Yersinia ruckeri and Clostridium perfringens | 30 days | LY, NO, PA, IR, DR, IG, PP (at higher concentration) ↑ | [144] |
| Oreochromis niloticus | Bacillus amyloliquefaciens | 104 and 106 CFU/g | Not evaluated | 30 and 60 days | WG, GR (after 60 days), SR, GC; GB, AL, TP (at higher concentration) ↑VH, BC← | [159] |
| Oreochromis niloticus | Bacillus licheniformis | 4.4 × 106 CFU/g | Streptococcus iniae | 70 days | WG, GR, DR↑; LY, ML←; FCR, SD, SR→ | [160] |
| Oreochromis niloticus | Lactobacillus acidophilus, Bifidobacterium bifidum, Streptococcus thermophilus | Not specified | Aeromonas hydrophila | 98 days | GR, DR, AL, GC, FCR↑; MO↓; ALT, AL, GB, SR→; AST← | [161] |
| Oreochromis niloticus | Bacillus subtilis | 5 × 106 CFU/g | Not evaluated | 84 days | GR, PC, PG→; LY, PA, HE↑; HM↓; IR← | [162] |
| Oreochromis niloticus | Bacillus subtilis | 0.1 g/mL (in water), 0.2 g/mL (in diet) | Flavobacterium columnare | 60 days | MO↓; WQ→; DR↑ | [163] |
Results: Significant change ←; No significant change →; Increase/Growth ↑; Decrease/Reduction ↓; Parameters: Absorptive area (ABA), Apparent digestibility coefficient (ADC), Albumin (AL), Alanine aminotransferase (ALT), Amylase (AM), Antibacterial activity (AN), Alkaline phosphatase (ALP), Aspartate aminotransferase (AST), Bacterial community (BC), Blood urea nitrogen (BUN), Catalase (CA), Cholesterol (CH), Digestive enzyme activities (DE), Disease resistance (DR), Feed conversion ratio (FCR), Feed efficiency (FE), Globulin (GB), Goblet cells (GC), Glutathione (GLx), Growth rate (GR), Hematocrit (HM), Hemoglobin (HE), Immune-related genes expression (IG), Immune response (IR), Lipase activity (LA), Lipid retention (LR), Lysozyme (LY), Microvilli density (MD), Microvilli length (ML), Mucin production (MP), Myeloperoxidase (MPO), Mortality (MO), Nitric oxide (NO), Phagocytic activity (PA), Plasma cortisol (PC), Peroxidase (PE), Plasma glucose (PG), Protection against pathogen (PP), Protease (PR), Protein retention (PRE), Phytase activity (PY), Respiratory burst (RB), Reactive oxygen species (ROS), Superoxide dismutase (SD), Skin mucous (SM), Survival rate (SR), Total blood protein (TP), Triglyceride (TR),Villus height (VH), Villus width (VW), Water quality (WQ), Weight gain (WG). Certain strains of Saccharomyces cerevisiae are added to the tilapia diet due to their ability to improve microbiota regulation through the production of polyamines involved in metabolite biosynthesis. This leads to changes in the intestinal tract morphology, such as the distribution of commensal bacteria, which can enhance nutrient absorption capacity and promote the development and maturation of mucins and epithelial cells. When selecting a probiotic, it is crucial to consider safety criteria recommended by FAO and WHO, such as antibiotic sensitivity, the absence of toxin production, and hemolytic activity. Additionally, the ability of a probiotic to colonize the host’s intestinal tract, resist stomach acids, bile salts, and enzymes, adhere to the gastrointestinal tract, and exhibit antagonism against pathogenic species are essential factors to assess. Probiotics can compete for receptor sites on the epithelium, reducing susceptibility to infections, as they can exclude pathogens from attaching to these sites. Some probiotics enhance their adhesion by synthesizing glycol conjugates on the gastrointestinal tract wall, which serve as receptors for bacteria attachment. The adhesion of probiotics is favored over that of pathogens because the host’s immune system recognizes probiotic antigens as harmless and does not trigger an inflammatory response. Moreover, probiotics can regulate mucins and proteins related to tight junctions, such as claudins, occludins, and ZO-1, thus preventing the attachment of harmful bacteria. For instance, probiotics belonging to the genus Lactobacillus can prevent the adhesion of E. coli, Klebsiella spp., and Pseudomonas aeruginosa strains to host intestinal cells.
Table 5.
Results of the Hierarchical Regression Analysis performed on tilapia specific growth rate, probiotic dose in feed, number of replicates, initial weight, water temperature and test duration.
Table 5.
Results of the Hierarchical Regression Analysis performed on tilapia specific growth rate, probiotic dose in feed, number of replicates, initial weight, water temperature and test duration.
| Model | Predictors | R2 | ∆R2 | ∆F | p-value |
| 1 | Number of replicates, initial weight, temperature and test duration. | 0.354 | 0.354 | 5.063 | 0.002 |
| 2 | Number of replicates, initial weight, temperature, test duration and probiotic dose in feed | 0.377 | 0.023 | 1.319 | 0.258 |
R2 = R square (coefficient of determination); ∆R2 = Delta R Square; ∆F = Final variation.
Table 6.
Results of the Hierarchical Regression Analysis performed on feed conversion ratio, probiotic dose in feed, number of replicates, initial weight, water temperature and test duration.
Table 6.
Results of the Hierarchical Regression Analysis performed on feed conversion ratio, probiotic dose in feed, number of replicates, initial weight, water temperature and test duration.
| Model | Predictors | R2 | ∆R2 | ∆F | p-value |
| 1 | Number of replicates, initial weight, temperature and test duration. | 0.279 | 0.279 | 5.036 | 0.002 |
| 2 | Number of replicates, initial weight, temperature, test duration and probiotic dose in feed | 0.279 | 0.000 | 0.014 | 0.905 |
R2 = R square (coefficient of determination); ∆R2 = Delta R Square; ∆F = Final variation.
Table 7.
Bacteriocins or bacteriocin-like inhibitory substances (BLIS) with antimicrobial effects against some pathogens in tests performed on tilapia.
Table 7.
Bacteriocins or bacteriocin-like inhibitory substances (BLIS) with antimicrobial effects against some pathogens in tests performed on tilapia.
|
Tilapia Species |
Bacteriocin or BLIS | Pathogen | Research mode | Results | Reference |
| Oreochromis niloticus | Nisin | Enterobacteriaceae | In vitro study, the capacity of preserving tilapia meat was evaluated | Biopreservation effect, bacteriocin did not affect sensory properties of the product, there was no biogenic amine production. | [182] |
| Oreochromis niloticus | BLIS produced by Paenibacillus ehimensis NPUST1 | Streptococcus iniae and Aeromonas hydrophila | In vivo study, administration of probiotics with BLIS production | Low pH tolerance, high thermal tolerance, broad spectrum, BLIS had antibacterial activity and improved fish immunity | [153] |
| Oreochromis niloticus | BLIS produced by Lactococcus lactis subsp. lactis 3MT | Vibrio spp. | In vitro study, evaluation of the biopreservation capacity of bacteriocin isolated from tilapia in fish pâté | Stable to heat and pH, antibacterial properties, free of virulence, no production of biogenic amines | [183] |
| Oreochromis niloticus | Nisin Z | Staphylococcus aureus ATCC 25923 | In vitro study, screening for bacteriocin production in LAB isolates and identification | High stability to heat, resistance to pH variations, detergents and NaCl, wide range of antibacterial activity | [184] |
| Oreochromis niloticus | BLIS produced by Bacillus spp. | Aeromonas hydrophila, Salmonella typhi | In vitro study, purification and evaluation of antibacterial capacity of BLIS extracted from tilapia gut | BLIS was not resistant to acid treatment and denatured in ammonium sulfate (20% of saturation), antibacterial activity against both tested pathogens. | [179] |
| Oreochromis niloticus | BLIS produced by Lactococcus garvieae | Staphylococcus aureus | In vivo study, administration of probiotics with BLIS production | BLIS showed moderate zones of inhibition against closed related species; fish that received bacteriocinogenic probiotics were protected against pathogens and had improved immune response. | [185] |
| Oreochromis niloticus | BLIS produced by Lactococcus coryniformis subsp. torquens MTi1 and MTi2 | Escherichia coli | In vitro study, purification and evaluation of antibacterial capacity of BLIS extracted from tilapia gut | BLIS exhibited antibacterial activity, but when submitted to enzyme treatment, the inhibitory properties were inactivated, according to the protein nature of these compounds | [156] |
| Oreochromis niloticus | BLIS produced by Lactococcus lactis subsp. lactis CF4MRS | Pseudomonas aeruginosa, Klebsiella pneumoniae, Escherichia coli, Aeromonas hydrophila, Edwardsiella tarda and Serratia marcescens | In vitro study, purification and evaluation of antibacterial activity of BLIS extracted from tilapia intestine | BLIS concentration was too low to significantly inhibit the pathogens | [186] |
| Oreochromis niloticus | Supernatant produced by Pediococcus pentosaceus NP6 | Salmonella enterica serovar typhimurium | In vitro study, capacity of preserving tilapia by-products | Supernatant exhibited antibacterial activity; partial purification indicates that it may be a bacteriocin | [187] |
| Oreochromis niloticus | BLIS produced by Bacillus endophyticus, Bacillus flexus, Bacillus mojavensis, Bacillus sonorensis and Bacillus subtilis | Streptococcus iniae | In vivo study, administration of probiotics with BLIS production | BLIS exhibited antibacterial activity; the enzyme treatment suggested that the inhibitory substance may be a bacteriocin | [188] |
| Oreochromis niloticus | BLIS produced by Lactococcus lactis RQ516 | Aeromonas hydrophila | In vivo study, administration of probiotics with BLIS production | Immunostimulant effect and antibacterial activity against a wide spectrum of bacteria, including A. hydrophila. | [189] |
| Oreochromis niloticus | Nisin | Enterobacteriaceae | In vitro study, capacity of preserving tilapia meat | Biopreservation effect, the bacteriocin did not affect sensory properties of the product, there was no biogenic amine production. | [182] |
| Oreochromis niloticus | BLIS produced by Paenibacillus ehimensis NPUST1 | Streptococcus iniae and Aeromonas hydrophila | In vivo study, administration of probiotics with BLIS production | Low pH tolerance, high thermal tolerance, broad spectrum, BLIS had antibacterial activity and improved fish immunity | [153] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



