
Submitted:
12 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
Screening of pollen traits in diploid wild potatoes (Solanum sect. Petota, Solanaceae) is desirable to develop heat-tolerant potato (S. tuberosum) cultivars. To accomplish this goal requires exploring potato genetic resources that are conserved in genebanks. The goal of this study was to assess pollen viability and 2n pollen production of the diploid potato wild relatives under heat stress condition. We assessed pollen viability and size of nine potato accessions conserved at the Embrapa Potato Gene Bank, including eight wild potatoes S. chacoense (BRA 00167447-2, BRA 00167017-3, BRA 00167023-1, BRA 00167028-0), S. commersonii (BRA00167007-4, BRA00167420-9, BRA00183760-8), and S. malmeanum (BRA 00183755-8), along with a control accession from the cultivated species S. tuberosum (BRA 00167251-8). The plant accessions were cultivated in different growth chambers, subjected to both control temperature (ranging from 14 to 27°C) and supraoptimal temperature conditions (ranging from 24 to 34°C). At heat stress, the accessions BRA 00167251-8 did not bloom, and BRA 00167023-1 did not produce pollen. The remaining accessions did not exhibit a significant reduction in pollen viability as the temperature increased. Pollen viability at the control temperature had the lowest value in BRA-00167420-9 (S. commersonii) with 68.5% and the highest in BRA 00183755-8 (S. malmeanum) with 100%. At the supraoptimal temperature the lowest value was in BRA 00167420-9 (S. commersonii) with 54.5% and the highest in BRA 00167017-3 (S. chacoense) with 94%. The average of pollen diameter was 20 μm in all wild potato genotypes, and the increase of temperature did not lead to 2n pollen production. Estimated Genotypic Coefficient of Variation (GCV) was lower than Phenotypic Coefficient of Variation (PCV) for pollen viability. The observed heritability values ranged from 58.82% in BRA00167007-4 to 91.32% in BRA 00183755-8. Our results highlight the genetic variability available in wild potato germplasm concerning pollen viability under heat stress. Furthermore, these first insights offer valuable guidance for ongoing and future endeavors in diploid potato breeding.
Keywords:
Crop wild relatives
; diploid breeding
; pre-breeding
; Solanaceae
; variability
Introduction
As F1-hybrid potato breeding continues to progress, assessing pollen viability becomes an essential trait in germplasm screening (Li et al. 2022). In regions characterized by tropical climates with high temperatures, identify genotypes that exhibit robust blooming and maintain appropriate pollen viability under supraoptimal temperatures can be advantageous for cultivating True Potato Seeds (TPS) propagated cultivars (Malagamba 1988; Buckseth et al. 2022). In the context of global climate change, it is crucial to explore the phenotypic plasticity of crops in response to rising temperatures to anticipate potential heat-related effects (Bloomfield et al. 2014). Given future climate change scenarios projecting a 1 to 3°C increase in global average temperatures during the twenty-first century (IPCC, 2018; Portner et al. 2022), there is a potential for substantial reductions in agricultural production and impacts on natural populations of crop wild relatives in their natural distribution areas, including potatoes (Jarvis et al. 2008; Vincent et al. 2019).
Globally, potato (Solanum tuberosum L., Solanaceae) ranks as the third most important crop, behind rice and wheat (Silva and Lopes 2015; Stokstad 2019). Despite the genetic diversity of the commercially potato crop is relatively limited (Hardigan et al. 2017), potatoes encompass a phylogenetic branch comprising 107 wild species, four domesticated species, and their natural and artificial hybrids (Spooner et al. 2007; Ovchinnikova et al. 2011; Spooner et al. 2014). Natural populations of potato wild species have evolved in extreme environmental conditions, such as drought, high temperatures, intense sunlight, and high altitudes, given their extensive geographical distribution in the Americas continent, ranging from North America in the southwestern United States (latitude 38 °N), through Central and South America to Argentina, and adjacent mainland in Chile (39 °S), including Brazil (Hawkes and Hjerting 1969; Hijmans and Spooner 2001). Genetic variability is essential for maintaining and improving the genetic base of agricultural crops (Magal et al. 2021; Singer et al. 2021; Singh et al. 2023). This variability is abundant within crop wild relatives, which serve as natural repositories of genes possessing agronomic, industrial, and nutritional significance (Jansky 2006; Warschefsky et al. 2014; Bethke et al. 2017; Dempewolf et al. 2017; Bashir et al. 2021).
Domesticated and wild potatoes are members of Solanum sect. Petota, capable of both sexual reproduction, involving botanical seeds, and vegetative propagation through stolons and tubers (Hawkes 1958; Correll 1962). Particularly, the capacity to produce diploid inbred lines and propagate via TPS (Stokstad 2019; Lindhout et al. 2011; Jansky et al. 2016; Bradshaw 2022a) contrasts with the predominant method of propagating tetraploid commercial potato crops (2n=4x=48), which is through vegetative means using tubers (Bradshaw 2022b). Diploid potato cultivars propagated through TPS aid in preventing the transmission of viruses to subsequent generations and facilitate the storage and conservation of propagative material (Jansky et al. 2016; Bradshaw 2022a). This stark contrast has heightened significant interest among breeders, farmers, and the seed industry in the development of F1-hybrid potato cultivars (Bradshaw 2022a; De Vries et al. 2016; Bethke et al. 2022). Breeding potatoes, particularly through F1-hybrid approaches, can facilitate the genetic mapping studies (Endelman and Jansky 2016; Meijer et al. 2018; Song and Endelman 2023). This heightened interest is further fueled by the accessibility of modern plant breeding techniques and genetic tools (Li et al. 2022; Bethke et al. 2019; Watson et al. 2018).
The effect of heat stress can lead to the formation of unreduced (2n) gametes in many crop species, such as roses (Rosa sp.) (Pécrix et al. 2011), tomato (Solanum lycopersicon L.) (Schindfessel et al. 2023), and wheat (Triticum turgidum L.) (Omidi et al. 2014), causing pollen unviability (Alam et al. 2017; Jiang et al. 2015; Jian et al. 2019), and consequently disrupt the stability of the natural populations (Zhou et al. 2022). Pollen viability studies in wild potato species grown under heat stress conditions are scarce in the literature. The identification of heat-tolerant wild potato genotypes regarding pollen viability favors the direction of future studies that relate stress, as plants respond to heat stress, triggering a cascade of physiological, biochemical, and molecular processes and adapt by activating several stress-responsive genes (Hasanuzzaman et al. 2013; Paupière et al. 2014). Conducting a screening for the ability to produce viable pollen under heat stress conditions is crucial for identifying heat tolerant genotypes (Paupière et al. 2017; Mesihovic et al. 2016). This process paves the way for further molecular characterization of genes involved and facilitates the selection of promising genotypes for future stages of a plant breeding program (Paupière et al. 2014; Mesihovic et al. 2016; Ahmed et al. 2022), such as potato.
By investigating the impact of temperature on pollen viability and 2n pollen production in diploid potato wild relatives, our study aims to contribute to the identification of genotypes that can thrive in high-temperature environments. This research is crucial for developing pre-breeding potato lines capable of withstand the challenges posed by global climate change.
Materials and Methods
Plant Material
In Brazil, three potato wild species are naturally distributed: Solanum chacoense Bitter, S. commersonii Dunal, and S. malmeanum Bitter (Spooner et al. 2014; 2016). We evaluated nine wild potato accessions from the species S. chacoense (BRA 00167447-2, BRA 00167017-3, BRA 00167023-1, BRA 00167028-0), S. commersonii (BRA00167007-4, BRA00167420-9, BRA00183760-8), and S. malmeanum (BRA 00183755-8), as well as a control accession from of the commercial cultivar of S. tuberosum named BRSIPR-BEL (BRA 00167251-8) which are conserved at the Embrapa Potato Gene Bank (Table 1).
Tubers of uniform size were planted in a 5L plastic container with TurfaFértil® organomineral substrate and supplemental fertilizer according to the crop instructions. The plants were transported to the growth chambers after 15 days of growth and exposed to two temperature gradients: control temperature, with thermal amplitude ranging from 14 to 27°C (Figure 1, blue line); and supraoptimal temperature, with amplitude ranging from 24 to 34°C (Figure 1, red line). The plant growth chamber was developed by InstalaFrio, a brazilian manufacturer. The photoperiod was programmed with 12 hours (7:00 to 19:00) in a supervisory software LabVIEW®, using ten groups of LED bulb lamps with a light intensity of about 400 mol m-2 s-1. Temperature control was carried out, for heating, by a reverse refrigeration system complemented by electrical resistance, and for cooling, by an air condenser refrigeration unit with hermetic compressor. The plants stayed under these two distinct treatments until harvest.
A double factorial experimental scheme 9 × 2 (genotype vs. temperature), was set up in randomized blocks with two replications each consisting of one plant.
Pollen Viability Assessment
Five flowers in the anthesis stage (when the floral bud opens) from each accession were collected on the same day for anther extraction and pollen processing (Alexander 1969). All the accessions from the two treatments opened the flowers in the same week, and pollen from each plant accession was collected seven days after the anthesis. Pollen from each plant was collected and preserved in 1.5 mL tubes in a freezer (-4°C) until analysis. To determine pollen viability, the percentage of staining pollen (PSP) was assessed in 400-grain sample dyed with red aceto-carmine glycerol (Marks 1954). Colored pollen was considered viable, and non-colored pollen was considered unviable (Figure 2). The examination of samples, as described in the technique provided by the protocol of CIP (International Potato Center) (Ordoñez et al. 2017), was conducted using an optical microscope with 200x magnification.
Pollen viability (percent) was calculated according to the formula:
Based on the viability range data obtained, four distinct categories were defined to classify pollen viability (Ordoñez et al. 2017). The percentage of pollen viability was considered sterile (0%), low (> 0% and ≤ 50%), moderate (>50% and ≤ 80%), or high (> 80% and ≤ 100%).
Pollen Size
For analysis of pollen size, pollen from each accession subjected to both temperature treatments was collected and analyzed following the protocol described by CIP (Ordoñez et al. 2017; Quinn et al. 1974). To distinguish 2n pollen, 50 viable pollen grains were stained with a droplet of 2% (w/v) aceto-carmine solution on a microscope slide. Pollen size was measured using an optical microscope. Enlarged pollen grains were classified as 2n pollen if the average diameter under heat stress condition exceeds 1.2 times the average diameter of the pollen grains in the control condition (Ordoñez et al. 2017; Quinn et al. 1974).
Pollen Viability-Based Heat Susceptibility Index (HSIpv)
The pollen viability-based heat susceptibility index (HSIpv) was calculated in Microsoft Excel 2023 using percent pollen viability values of heat stress and normal conditions, following the formula below (Fischer and Maurer 1978; Khan et al. 2022):
Building upon, HSIpv genotypes were grouped into three classes: tolerant (HSIpv <0.5), moderately tolerant (HSIpv 0.5-0.99), and susceptible (HSIpv >1.0).
Genetic Parameters
The basic genetic parameters—phenotypic coefficient of variation (PCV%), genotypic coefficient of variation (GCV%), broad sense heritability (H²), genetic gain (GG) and genetic advance (GA)—were calculated. The PCV% and GCV% were calculated using the formulas: PCV%=(√VP)/mean×100 and GCV%=(√VG)/mean×100, respectively.
To categorize the GCV and PCV values, the classification proposed by Burton (1952) was employed. GCV and PCV values falling within the range of 0 to ≤10% were classified as low, values ranging from 10 to ≤20% were categorized as moderate, and values exceeding 20% were considered high.
Genotypic variance was calculated using the formula:
where:
MSt = Mean sum of squares for trait of genotype.
MSe = Mean sum of squares for error of genotype.
r = Number of replications.
Phenotypic Variance
VP = Phenotypic variance for each trait of genotype.
VG = Genotypic variance for each trait of genotype.
VE = Environmental variation among the tested traits of genotype.
Heritability
Heritability was estimated as the ratio of total genotypic variance to the phenotypic variance (Allard 1960):
where H2 = % broad sense heritability percentage. The heritability percentage was categorized as low (0–30%), moderate (30–60%), and high ≥60% as given by Robinson et al. (1949):
The genetic gain was calculated by the ratio of genetic advance (GA= (k× √(Vp)× H2), where k represents the constant value 2.06 at 5% selection intensity, and Vp is the phenotypic variance. The overall mean of the population used is denoted by ∑.
Statistical Analysis
The standard error and the significance of differences (p≤0.05) for temperature, genotype, and genotype interaction were determined using analysis of variance (ANOVA) for pollen viability and pollen size. Following that, the Tukey mean comparison test was used, and the analyses were carried out using the function fat2.dbc of the ‘ExpDes.pt’ package (Ferreira et al. 2021) in the RStudio environment (RStudio Team 2023).
Results and Discussion
Analysis of Variance
The results of the ANOVA indicated that the main effects, genotype and temperature, are significant, while block and the genotype by environment (G×E) interaction are not significant (Table 2).
Pollen Viability-Based Heat Susceptibility Index (HSIpv)
Table 3 presents the genetic parameters estimated for pollen viability. GCV (15.76%) was lower than PCV (17.84%). According to Johnston et al. (1955) genetic advance (% average) is categorized as low (<10%), moderate (10–20%) and high (>20%). In our study, the genetic advance for pollen viability was high (23.30%) (Table 3).
Pollen viability of the potato wild genotypes from two experimental conditions is presented in Table 4. As the genotype by environment (G×E) interaction was non-significant, the Tukey test was only applied for each treatment. At the control (CT) conditions, genotypes BRA 00183755-8, BRA 00167017-3, BRA 00167028-0, BRA 00167447-2, and BRA00167007-4 exhibited the highest pollen viability values. Conversely, genotypes BRA00167420-9 and BRA00183760-8 displayed the lowest pollen viability. Under heat stress (HS) conditions, the maximum pollen viability was observed in genotypes BRA 00167017-3, BRA 00183755-8, and BRA 00167028-0. However, the genotypes BRA 00167447-2, BRA00183760-8, BRA00167007-4, and BRA00167420-9 demonstrated lower pollen viability under HS conditions. A non-significant reduction in pollen viability was observed with increasing temperature in genotypes BRA00167007-4, BRA00167420-9, BRA 00167447-2, BRA 00167017-3, BRA 00167028-0, BRA 00183755-8, and BRA00183760-8 (Table 4). Conversely, BRA 00167023-1 failed to produce pollen, and BRA 00167251-8 did not bloom under elevated temperatures; hence, these two genotypes were not included in this analysis.
The heat susceptibility index (HSIpv) is a useful criterion to select heat-tolerant genotypes (Khan et al. 2022). Genotypes with HSIpv < 0.5 were considered as heat-tolerant, those with HSIpv ranging from 0.5–0.99 were considered moderately tolerant, and genotypes with HSIpv > 1.0 were deemed susceptible (Khan et al. 2022). The genotypes categorized as tolerant were BRA00183760-8 (0.06) and BRA 00167017-3 (0.36). BRA 00183755-8 (0.66) was moderately tolerant, while BRA 00167028-0 (1.01), BRA 00167447-2 (1.57), BRA00167420-9 (1.59), and BRA00167007-4 (1.89) were susceptible (Table 4).
Heritability for Pollen Viability
The estimated heritability for pollen viability varied from 58.82% to 90.79%. The lowest H² was observed in BRA00167007-4 (58.82%), while the highest value was observed in BRA 00183755-8 (91.32%), followed by BRA 00167023-1 (90.79%), BRA 00167028-0 (88.56%), BRA 00167017-3 (88.35%), BRA00167420-9 (87.72%), BRA00183760-8 (83.82%), and BRA 00167447-2 (83.16%). Heritability is classified as low (<30%), medium (30–60%), and high (>60%) (Allard 1960; Johnson et al. 1955). High heritability indicates that selection for the trait is effective and is less influenced by environmental effects (Allard 1960; Johnson et al. 1955). Broad-sense heritability becomes particularly intriguing when it applies to situations where the full genetic variability can be harnessed in clones or single-cross hybrids (Bernardo 2010).
A non-significant difference in pollen viability was observed as the temperature increased, indicating that the genotypes maintained their performance under both treatments. Heat stress has been shown to adversely affect or even inhibit pollen viability in various agricultural crops, including corn (Zea mays L.) (Begcy et al. 2019), wheat (Triticum aestivum L.) (Ullah et al. 2022), rice (Oryza sativa L.) (Kumar et al. 2015), tomato (Solanum lycopersicum L.) (Paupière et al. 2017), and wild potatoes (Solanum sect. Petota) (Bamberg 1995). Consistent with our findings, Bamber and colleagues (1995) found that when explodes to elevated temperature stress conditions, certain wild potato genotypes displayed pollen viability rates above 45%. These genotypes included Solanum stoloniferum Schltdl. (cited as S. fendleri A. Gray, 50% for PI 275156, 56% for PI 497998, and 63% for PI 498004), S. stoloniferum Schltdl. (cited as S. polytrichon Rydb., with 49% for PI 255547), S. demissum Lindl. (70% for PI 160208 and 50% for PI 498232) and S. jamesii Torr. (both PI 458425 and PI 195190, each with 45%), S. chacoense (49% for PI 320293), S. commersonii (53% for PI 243503), S. infundibuliforme Phil. (68% for PI 498351), and S. boliviense Dunal (cited as S. megistacrolobum Bitter, with 47% for PI 473133). In our study, except for the genotypes that did not produce pollen and did not bloom under heat stress, S. chacoense (BRA 00167023-1) and S. tuberosum (BRA 00167251-8), respectively, the genotypes of the species S. chacoense (BRA 00167017-3, BRA 00167028-0, BRA 00167447-2), S. commersonii (BRA00167007-4, BRA00183760-8, BRA00167420-9), and S. malmeanum (BRA 00183755-8) exhibited similar pollen viability results to the study by Bamberg and collaborators (1995), with values higher than 54.5%.
Pollen Size
Pollen size was compared using a non-parametric Wilcoxon-Mann-Whitney test. On average, pollen size measured 20 μm across all wild potato genotypes, ranging from 18 to 24 μm in the CT conditions and 16 to 22 μm in the HS conditions. Specifically, pollen size for BRA00167007-4 was 20 μm in the CT conditions and 22 μm in the HS conditions. BRA00167420-9, BRA 00167447-2, BRA 00167017-3, BRA 00167028-0, BRA 00183755-8, BRA00183760-8 exhibited consistent pollen size of 20 μm across both treatments. BRA 00167023-1 produces pollen with a size of 20 μm, while BRA 00167251-8 exhibited pollen with a size of 24 μm in the CT conditions. Notably, these two genotypes failed to produce pollen and did not bloom under the HS treatment (Figure 3).
The temperature elevation did not lead to the production of non-reduced (2n) pollen grains in any of the genotypes assessed in this study. According to Ordoñez et al. (2017), pollen grains are considered diploid if their size increases by 1.2 times compared to normal ones. However, under supraoptimal temperature condition, a 1.1-fold increase in pollen size was observed in BRA 00183755-8 compared to the control temperature condition.
The variations on pollen viability among the tested genotypes present opportunities for selecting desirable genotypes to enhance future potato crop improvement efforts. This variability suggests that promising genotypes can be identified based on their pollen viability trait. However, the phenotypic coefficient of variation (PCV) and genotypic coefficient of variation (GCV) for all genotypes under both conditions were moderate. A moderate level of coefficient of variation indicates an equal influence of additive and nonadditive gene action (Allard and Hansche 1964). In this study, the PCV was higher than the GCV, although the differences between the PCV and GCV ranges were minimal, suggesting the limited impact of the environment on trait expression and implying the heritable nature of the traits (Allard and Hansche 1964). Several studies have also reported higher PCV than GCV (Nagalakshmi et al. 2018; Ganesan et al. 2010), indicating significant potential for precise trait selection based on the phenotypic expression (Schoper et al. 1986; Panthee et al. 2018; Hazra and Ansary 2008).
Heritability serves as a valuable metric for assessing the extent of variation within a population (Visscher et al. 2008). In this study, a high heritability was observed for pollen viability. Previous research consistently indicates that high heritability reflects minimal environmental influence on genotype expression (Bamberg 1995; Allard 1964; Petr and Frey 1966). Consequently, the limited environmental impact on the studied trait allowed for a high accuracy of 94% in selecting genotypes under HS conditions (Khan et al. 2022; Olakojo and Olaoye 2011; Aminu and Izge 2012; Azam et al. 2015). This insight into heritability from the current study aids in selecting optimal traits for crop improvement (Khan et al. 2022; Olakojo and Olaoye 2011; Aminu and Izge 2012; Azam et al. 2015).
Furthermore, this study unveiled a high level of heritability for pollen viability trait in potato wild genotypes. However, relying solely on heritability-based trait selection may not consistently yield success, as broad-sense heritability encompasses total genetic variance, including additive, dominant, and epistatic variances (Hill et al. 2008). Thus, estimating heritability alongside with high genetic advance is more reliable and efficient for selecting desirable traits across a population (Nyquist and Baker 1991). Our study identified high heritability coupled with significant genetic gain for the pollen viability, providing valuable information for selecting superior genotypes (Divakara et al. 2010; Kalloo 2012).
The abortion of pollen development observed in BRA 00167023-1 under HS conditions can be attributed to various factors. These include the degradation of membrane integrity, the accumulation of reactive oxygen species (ROS), alterations in carbohydrate metabolism (Liu et al. 2019), disruptions in protein and lipid metabolism (Narayanan et al. 2018), and changes in phospholipid profiles (Djanaguiraman et al. 2018). These factors collectively leads to degeneration and abnormalities in tapetum cells (Cecchetti et al. 2013).
A comprehensive understanding of the mechanisms underlying the response to supraoptimal temperatures during sexual reproduction, coupled with the identification of reproductive traits associated with thermo-tolerance, will enable breeders with more effective tools to develop heat stress-resilient crops and enhance crop productivity (Aiqing et al. 2018; Chaturvedi et al. 2021; Delph et al. 1997; Djanaguiraman et al. 2013; Djanaguiraman et al. 2014; Endo et al. 2009; Jain et al. 2010; Pécrix et al. 2011).
The identification of heat-tolerant wild potato genotypes exhibiting improved pollen viability is a crucial advancement in pre-breeding initiatives aiming at enhancing the potato crop. Heat stress triggers a series of physiological, biochemical, and molecular changes in plants, stimulating the activation of various stress-responsive genes as an adaptative mechanism (Li et al. 2022; Hasanuzzaman et al. 2013). The capacity to produce viable pollen under elevated temperatures, as demonstrated in this investigation, holds considerable significance in understanding heat tolerance and accelerates the subsequent molecular characterization of genes involved in this intricate process.
Conclusions
Pollen viability in wild potato germplasm exhibits genetic variability when cultivated at optimum temperature (control) and high temperature (stress).
The S. chacoense genotypes (BRA 00167017-3) and S. commersonii (BRA 00183760-8) were identified as heat-tolerant based on their pollen viability, highlighting their potential as promising candidates for future improvement studies.
Elevated temperatures did not result in the formation of 2n pollen in the evaluated potato wild genotypes.
Author Contributions
Performed the experiments: Nicolao, R.; Bashir, I. Conceptualization: Nicolao, R. Data curation: Nicolao, R. Formal analysis: Nicolao, R.; Bashir, I. Supervision: Heiden, G.; Castro, C.M. Writing & editing: Nicolao, R.; Bashir, I.; Heiden, G.; Castro, C.M.
Acknowledgments
This work was supported by CAPES/PROAP and EMBRAPA. RN acknowledges the CAPES (88887.622551/2021-00), The International Association for Plant Taxonomy for the IAPT 2022 Research Grants Program and Botanical Society of America for the BSA 2023 Research Grants Program. IB acknowledges CNPq (429368/2016-0) and its partnership with TWAS (TWAS/CNPq 154585/2017-3). GH acknowledges CNPQ (314590/2020-0) for the productivity research fellowship. CC acknowledge USDA Potato Genebank for sending S. chacoense germplasm.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li D, Lu X, Zhu Y, Pan J, Zhou S, Zhang X, Zhu G, Shang Y, Huang S, Zhang C (2022) The multi-omics basis of potato heterosis. Journal of Integrative Plant Biology 64:3, 671-687. [CrossRef]
- Malagamba, P. (1988) Potato production from true seed in tropical climates. HortScience 23:3, 495-500.
- Buckseth T, Tiwari JK, Singh RK, Kumar V, Sharma AK, Dalamu D, Bhardwaj V, Sood S, Kumar M, Sadawarti M, Challam CL, Naik S, Pandey NK (2022) Advances in innovative seed potato production systems in India. Frontiers in Agronomy 4:956667. [CrossRef]
- Bloomfield JA, Rose TJ, King GJ (2014) Sustainable harvest: managing plasticity for resilient crops. Plant Biotechnology Journal 12:517–533.
- IPCC: Masson-Delmotte, V, Zhai, P, Pörtner, H. O, Roberts, D, Skea, J, Shukla, P.R, Pirani, A, Moufouma-Okia, W, Péan, C, Pidcock, R, Connors, S, Matthews, J. B. R, Chen, Y, Zhou, X, Gomis, M. I, Lonnoy, E, Maycock, T, Tignor, M, Waterfield, T. (eds.)]. Global warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty 2018 In Press.
- Pörtner HO, Roberts DC, Adams H, Adler C, Aldunce P, Ibrahim ZZ (2022) Climate Change 2022: Impacts, Adaptation and Vulnerability. IPCC: Geneva, Switzerland. Available online: https://www.ipcc.ch/report/ar6/wg2/downloads/report/IPCC_AR6_WGII_FrontMatter.pdf (accessed on 1 January 2023).
- Jarvis A, Lane A, Hijmans RJ (2008) The effect of climate change on crop wild relatives. Agriculture, Ecosystem and Environment 126:13–23. [CrossRef]
- Vincent H, Amri A, Castañeda-Álvarez NP, Dempewolf H, Dulloo E, Guarino L, Hole D, Mba C, Toledo A, Maxted N (2019) Modeling of crop wild relative species identifies areas globally for in situ conservation. Communications Biology 2:1, 1-8. [CrossRef]
- Silva GO, Lopes CA (2015) Sistema de produção da Batata. Embrapa, Brasília 2. Available online: http://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/1028425.
- Stokstad E (2019) The new potato. Science 363:6427, 574-577. [CrossRef]
- Hardigan MA, Laimbeer FPE, Newron L, Crisovan E, Hamilton JP, Vaillancourt B, Wiegert-Rininger K, Wood JC, Douches DS, Farré EM, Veilleux RE, Nuell CR (2017) Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proceedings of the National Academy of Sciences of the United States of America 114:46, 9999–10008. [CrossRef]
- Spooner DM, Núñez J, Trujillo G, Herrera MDR, Guzmán F, Ghislain M, Nunez J, del Rosario Herrera M, Guzma F (2007) Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. Proceedings of the National Academy of Sciences 104:19398–19403. [CrossRef]
- Spooner DM, Ghislain M, Simon R, Jansky SH, Gavrilenko T (2014) Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Botanical Review 80:283–383. [CrossRef]
- Ovchinnikova A, Krylova E, Gavrilenko T, Smekalova T, Zhuk M, Knapp S, Spooner DM (2011) Taxonomy of cultivated potatoes (Solanum section Petota: Solanaceae). Botanical Journal of the Linnean Society 165:107–155. [CrossRef]
- Hawkes JG, Hjerting JP (1969) The potatoes of Argentina, Brazil, Paraguay and Uruguay. A biosystematics study. Oxford University Press, London.
- Hijmans RJ, Spooner DM (2001) Geographic distribution of wild potato species. American Journal of Botany 88, 2101–2112. [CrossRef]
- Magar BT, Acharya S, Gyawali B, Timilsena K, Upadhayaya J, Shrestha J (2021) Genetic variability and trait association in maize (Zea mays L.) varieties for growth and yield traits. Heliyon 7(9). [CrossRef]
- Bashir I, Nicolao R, Heiden G (2021) Wild potatoes: A genetic reservoir for potato breeding. In Wild germplasm for genetic improvement in crop plants, Academic Press, pp. 215-240.
- Singh B, Sharma J, Sood S, Bhardwaj V, Siddappa S, Dalamu, Kardile HS, Sharma V, Dipta B, Kumar V, Dua VK, Goutam U, Pandey NK (2023) Genetic Variations in Tuber Dry Matter (%), Yield and Mineral Concentrations in a Diversity Panel of Tetraploid Potatoes. Potato Research 66(1):179-193. [CrossRef]
- Singer SD, Laurie JD, Bilichak A, Kumar S, Singh J (2021) Genetic variation and unintended risk in the context of old and new breeding techniques. Critical Reviews in Plant Sciences 40(1):68-108. [CrossRef]
- Dempewolf H, Baute G, Anderson J, Kilian B, Smith C, Guarino L (2017) Past and future use of wild relatives in crop breeding Crop Science 57:3, 1070-1082. [CrossRef]
- Warschefsky E, Penmetsa RV, Cook DR, Von Wettberg EJ (2014) Back to the wilds: tapping evolutionary adaptations for resilient crops through systematic hybridization with crop wild relatives. American journal of Botany 101:10, 1791-1800. [CrossRef]
- Jansky S (2006) Overcoming hybridization barriers in potato. Plant Breeding 125:1, 1–12. [CrossRef]
- Bethke PC, Halterman DA, Jansky S (2017) Are we getting better at using wild potato species in light of new tools?. Crop Science 57:3, 1241-1258. [CrossRef]
- Hawkes JG (1958) Significance of wild species and primitive forms for potato breeding. Euphytica 7:3, 257–270. [CrossRef]
- Correll D (1962) The potato and its wild relatives. Texas Research Foundation, Renner, USA .
- Lindhout P, Meijer D, Schotte T, Hutten RC, Visser RG, van Eck HJ (2011) Towards F1 hybrid seed potato breeding. Potato Research 54:4, 301-312. [CrossRef]
- Jansky SH, Charkowski AO, Douches DS, Gusmini G, Richael C, Bethke PC, Spooner DM, Novy RG, de Jong H, de Jong WS, Bamberg JB, Thompson AL, Bizimungu B, Holm DG, Brown CR, Haynes KG, Sathuvalli VR, Veilleux RE, Miller JC, Bradeen JM, Jiang J (2016) Reinventing potato as a diploid inbred line–based crop. Crop Science 56:1412–1422. [CrossRef]
- Bradshaw JE (2022a) Breeding diploid F1 hybrid potatoes for propagation from botanical seed (TPS): comparisons with theory and other crops. Plants 11:9, 1121. [CrossRef]
- Bradshaw JE (2022b) A brief history of the impact of potato genetics on the breeding of tetraploid potato cultivars for tuber propagation. Potato Research 65(3): 461-501. [CrossRef]
- De Vries M, ter Maat M, Lindhout P (2016) The potential of hybrid potato for East-Africa. Open Agriculture 1:1:151-156. [CrossRef]
- Bethke PC, Halterman DA, Francis DM, Jiang J, Douches DS, Charkowski AO, Parsons J (2022) Diploid potatoes as a catalyst for change in the potato industry. American Journal of Potato Research 99:5-6, 337-357. [CrossRef]
- Endelman JB, Jansky SH (2016) Genetic mapping with an inbred line-derived F2 population in potato. Theoretical and Applied Genetics 129, 935-943. [CrossRef]
- Meijer D, Viquez-Zamora M, Van Eck HJ, Hutten RCB, Su Y, Rothengatter R, Visser RGF, Lindhout WH, Van Heusden AW (2018) QTL mapping in diploid potato by using selfed progenies of the cross S. tuberosum× S. chacoense. Euphytica 214:1-18. [CrossRef]
- Song L, Endelman JB (2023) Using haplotype and QTL analysis to fix favorable alleles in diploid potato breeding. The Plant Genome e20339. [CrossRef]
- Bethke PC, Halterman DA, Jansky SH (2019) Potato germplasm enhancement enters the genomics era. Agronomy 9:10, 575. [CrossRef]
- Watson A, Ghosh S, Williams MJ, Cuddy WS, Simmonds J, Rey MD, Hatta MAM, Hinchliffe A, Steed A, Reynolds D, Adamsky NM, Breakspear A, Korolev A, Rayuner T, Dixon LE, Riaz A, Martin W, Ryan M, Edward D, Batley J, Raman H, Carter J, Rogers C, Domoney C, Moore G, Harwood W, Nicholson P, Dieters MJ, DeLacy IH, Zhou J, Uauy C, Boden SA, Park RF, Wulff BBH, Hickey LT (2018) Speed breeding is a powerful tool to accelerate crop research and breeding. Nature Plants 4:1, 23-29. [CrossRef]
- Pécrix Y, Rallo G, Folzer H, Cigna M, Gudin S, Le Bris, M (2011) Polyploidization mechanisms: temperature environment can induce diploid gamete formation in Rosa sp. Journal of Experimental Botany 62:3587–3597. [CrossRef]
- Schindfessel C, De Storme N, Trinh HK, Geelen D (2023) Asynapsis and meiotic restitution in tomato male meiosis induced by heat stress. Frontiers in Plant Science 14. [CrossRef]
- Omidi M, Siahpoosh MR, Mamghani R, Modarresi M (2014) The influence of terminal heat stress on meiosis abnormalities in pollen mother cells of wheat. Cytologia 79:1, 49-58. [CrossRef]
- Alam MA, Seetharam K, Zaidi PH, Dinesh A, Vinayan MT, Nath UK (2017) Dissecting heat stress tolerance in tropical maize (Zea mays L.). Field Crops Research 204:110-119. [CrossRef]
- Jiang Y, Lahlali R, Karunakaran C, Kumar S, Davis AR, Bueckert RA (2015) Seed set, pollen morphology and pollen surface composition response to heat stress in field pea. Plant, Cell & Environment (11):2387-2397. [CrossRef]
- Jiang Y, Lahlali R, Karunakaran C, Warkentin TD, Davis AR, Bueckert RA (2019) Pollen, ovules, and pollination in pea: Success, failure, and resilience in heat. Plant, Cell & Environment 42(1):354-372. [CrossRef]
- Zhou Q, Cheng X, Kong B, Zhao Y, Li Z, Sang Y, Wu J, Zhang P (2022) Heat shock-induced failure of meiosis I to meiosis II transition leads to 2n pollen formation in a woody plant. Plant Physiology 189:4:2110-2127. [CrossRef]
- Hasanuzzaman M, Nahar K, Alam M, Roychowdhury R, Fujita M (2013) Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. International Journal of Molecular Sciences 14:5, 9643-9684. [CrossRef]
- Paupière MJ, Van Heusden AW, Bovy AG (2014) The metabolic basis of pollen thermos-tolerance: perspectives for breeding. Metabolites 4:4, 889-920. [CrossRef]
- Paupière MJ, van Haperen P, Rieu I, Visser RG, Tikunov YM, Bovy AG (2017) Screening for pollen tolerance to high temperatures in tomato. Euphytica 213:1-8. [CrossRef]
- Mesihovic A, Iannacone R, Firon N, Fragkostefanakis S (2016) Heat stress regimes for the investigation of pollen thermotolerance in crop plants. Plant Reproduction 29:93-105. [CrossRef]
- Ahmed Z, Khalid M, Ghafoor A, Shah MKN, Raja GK, Rana RM, Mahmood T, Thompson AM (2022) SNP-Based Ge-nome-wide association mapping of pollen viability under heat stress in tropical Zea mays L. inbred lines. Frontiers in Genetics 13:819849. [CrossRef]
- Alexander MP (1969) Differential staining of aborted and non-aborted pollen. Stain Technology 44:117–122. [CrossRef]
- Marks GE (1954) An aceto-carmine glycerol jelly for use in pollen-fertility counts. Stain Technology 29:5, 277-277. [CrossRef]
- Ordoñez B, Orrillo M, Bonierbale MW (2017) Technical manual potato reproductive and cytological biology. International Pota-to Center (CIP) pp.69. Available online: https://cgspace.cgiar.org/server/api/core/bitstreams/5d4bb61f-c49d-40e8-92ee-e41a512f2da1/content.
- Quinn AA, Mok DWS, Peloquin SJ (1974) Distribution and significance of diplandroids among the diploid Solanums. American Potato Journal 51:16-21. [CrossRef]
- Fischer RA, Maurer R (1978) Drought resistance in spring wheat cultivars. I. Grain yield responses. Australian Journal of Agricultural Research 29:5, 897-912. [CrossRef]
- Khan I, Wu J, Sajjad M (2022) Pollen viability-based heat susceptibility index (HSIpv): A useful selection criterion for heat-tolerant genotypes in wheat. Frontiers in Plant Science 13:1064569. [CrossRef]
- Burton GW (1952) Quantitative inheritance in grasses. In Proceedings of the 6th International Grassland Congress, State College, PA, USA 17–23, pp 277–283.
- Allard RW (1960) Principles of plant breeding. John Willey and Sons. Inc. New York.
- Robinson HF, Cornstock RE, Harvey PH (1949) Estimates of heritability and degree of dominance in corn. Agronomy Journal 41:353–359.
- Ferreira EB, Cavalcanti PP, Nogueira DA (2021) ExpDes.pt: Experimental Designs package (Portuguese). R package version 1.1.2. Available online: https://cran.utstat.utoronto.ca/web/packages/ExpDes.pt/ExpDes.pt.pdf (accessed on 1 August 2023).
- RStudio Team (2020) Rstudio: Integrated Development for R. Rstudio, PBC, Boston MA. Available online: http://www.rstudio.com/ (accessed on 1 August 2023).
- Johnson HW, Robinson HF, Comstock RE (1955) Estimation of genetic and environmental variability in soybean. Agronomy Journal 47:314-318.
- Bernardo R (2010) Breeding for quantitative traits in plants. 2nd ed. Woodbury: Stemma Press.
- Begcy K, Nosenko T, Zhou LZ, Fragner L, Weckwerth W, Dresselhaus T (2019) Male sterility in maize after transient heat stress during the tetrad stage of pollen development. Plant Physiology 181:2, 683-700. [CrossRef]
- Ullah A, Nadeem F, Nawaz A, Siddique KH, Farooq M (2022) Heat stress effects on the reproductive physiology and yield of wheat. Journal of Agronomy and Crop Science 208:1, 1-17. [CrossRef]
- Kumar N, Kumar N, Shukla A, Shankhdhar SC, Shankhdhar D (2015) Impact of terminal heat stress on pollen viability and yield attributes of rice (Oryza sativa L.). Cereal Research Communications 43:4, 616-626. [CrossRef]
- Paupière MJ, Müller F, Li H, Rieu I, Tikunov YM, Visser RG, Bovy AG (2017) Untargeted metabolomic analysis of tomato pollen development and heat stress response. Plant Reproduction 30:2, 81-94. [CrossRef]
- Bamberg JB (1995) Screening potato (Solanum) species for male fertility under heat stress. American Potato Journal 72:1, 23-33. [CrossRef]
- Allard RW, Hansche PE (1964) Some parameters of population variability and their implications in plant breeding. Advances in Agronomy 16:281-325. [CrossRef]
- Nagalakshmi RM, Ravikesavan R, Paranidharan V, Manivannan N, Firoz H, Vignesh M, Senthil N (2018) Genetic variability, heritability and genetic advance studies in backcross populations of maize (Zea mays L.). Electronic Journal of Biotechnology 9:1137–1145.
- Ganesan KN, Nallathambi G, Senthil N, Tamilarasi PM (2010) Genetic divergence (Zea mays L.) Analysis in indigenous maize. Electronic Journal of Biotechnology 1:1241–1243.
- Schoper JB, Lambert RJ, Vasilas BL (1986) Maize pollen viability and ear receptivity under water and high temperature stress 1. Crop Science 26:5, 1029-1033. [CrossRef]
- Panthee DR, Kressin JP, Piotrowski A (2018) Heritability of flower number and fruit set under heat stress in tomato. HortScience 53:9, 1294-1299. [CrossRef]
- Hazra P, Ansary SH (2008) Genetics of heat tolerance for floral and fruit set to high temperature stress in tomato (Lycopersicon esculentum Mill.). SABRAO Journal of Breeding & Genetics 40(2), p 117.
- Visscher P, Hill W, Wray N (2008) Heritability in the genomics era — concepts and misconceptions. Nature Review Genetics 9:255–266. [CrossRef]
- Petr FC, Frey KJ (1966) Genotypic Correlations, Dominance, and Heritability of Quantitative Characters in Oats 1. Crop Science 6:3, 259-262. [CrossRef]
- Olakojo SA, Olaoye G (2011) Correlation and heritability estimates of maize agronomic traits for yield improvement and Striga asiatica (L.) Kuntze tolerance. African Journal of Plant Science 5:365–369. Available online: https://academicjournals.org/article/article1380018174_Olakojo%20and%20Olaoye.pdf.
- Aminu D, Izge AU (2012) Heritability and correlation estimates in maize (Zea mays L.) Under drought conditions in northern guinea and sudan savannas of Nigeria. World Journal of Agricultural Sciences 8:598–602. [CrossRef]
- Azam MG, Sarker U, Maniruzzaman M, Banik BR (2015) Genetic variability of yield and its contributing characters on CIMMYT maize inbreds under drought stress. Bangladesh Journal of Agricultural Research 39:419–426.
- Hill WG, Goddard ME, Visscher PM (2008) Data and theory point to mainly additive genetic variance for complex traits. PloS Genetics 4:2, e1000008. [CrossRef]
- Nyquist WE, Baker RJ (1991) Estimation of heritability and prediction of selection response in plant populations. Critical Reviews in Plant Sciences 10(3):235-322. [CrossRef]
- Divakara BN, Upadhyaya HD, Wani SP, Gowda CL (2010) Biology and genetic improvement of Jatropha curcas L.: a review. Applied Energy 87:3, 732-742. [CrossRef]
- Kalloo G (2012) Genetic improvement of tomato. Springer Science & Business Media.
- Liu Y, Li J, Zhu Y, Jones A, Rose RJ, Song Y (2019) Heat stress in legume seed setting: effects, causes, and future prospects. Frontiers in Plant Science 10:938. [CrossRef]
- Narayanan S, Prasad PVV, Welti R (2018) Alterations in wheat pollen lipidome during high day and night temperature stress. Plant, Cell & Environment 41:1749–1761. [CrossRef]
- Djanaguiraman M, Perumal R, Ciampitti IA, Gupta SK, Prasad PVV (2018) Quantifying pearl millet response to high temperature stress: thresholds, sensitive stages, genetic variability and relative sensitivity of pollen and pistil. Plant, Cell & Environment 41:993–1007. [CrossRef]
- Cecchetti V, Celebrin D, Napoli N, Ghelli R, Brunetti P, Costantino P, Cardarelli M (2013) An auxin maximum in the middle layer controls stamen development and pollen maturation in Arabidopsis. New Phytologist 1194–1207. [CrossRef]
- Aiqing S, Somayanda I, Sebastian SV, Singhm K, Gill K, Prasad PV, Jagadish SV (2018) Heat stress during flowering affects time of day of flowering, seed set, and grain quality in spring wheat. Crop Science 58:1, 380–392. [CrossRef]
- Chaturvedi P, Wiese AJ, Ghatak A, Zaveska DL, Weckwerth W, Honys D (2021) Heat stress response mechanisms in pollen development. New Phytologist 231:2, 571-585. [CrossRef]
- Delph LF, Johannsson MH, Stephenson AG (1997) How environmental factors affect pollen performance ecological and evolutionary perspectives. Ecology 78:1632- 1639. [CrossRef]
- Djanaguiraman M, Prasad PV, Boyle D, Schapaugh W (2013) Soybean pollen anatomy, viability and pod set under high temperature stress. Journal of Agronomy and Crop Science 199:171–177. [CrossRef]
- Djanaguiraman M, Prasad PV, Murugan M, Perumal R, Reddy UK (2014) Physiological differences among sorghum (Sorghum bicolor L. Moench) genotypes under high temperature stress. Environmental and Experimental Botany 100:43–54. [CrossRef]
- Endo M, Tsuchiya T, Hamada K, Kawamura S, Yano K, Ohshima M, Higashitani A, Watanabe M, Kawagishi-Kobayashi M (2009) High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development. Plant & Cell Physiology 50:1911–1922. [CrossRef]
- Jain M, Chourey PS, Boote KJ, Allen LHJr (2010) Short-term high temperature growth conditions during vegetative-to-reproductive phase transition irreversibly compromise cell wall invertase-mediated sucrose catalysis and microspore meiosis in grain sorghum (Sorghum bicolor). Journal of Plant Physiology 167:578–582. [CrossRef]
- Pécrix Y, Rallo G, Folzer H, Cigna M, Gudin S, Le Bris M (2011) Polyploidization mechanisms: temperature environment can induce diploid gamete formation in Rosa sp. Journal of Experimental Botany 62:3587–3597. [CrossRef]
Figure 1.
Temperature profiles of the control temperature (represented by the blue line) and supraoptimal temperature (heat stress, represented by the red line).
Figure 1.
Temperature profiles of the control temperature (represented by the blue line) and supraoptimal temperature (heat stress, represented by the red line).
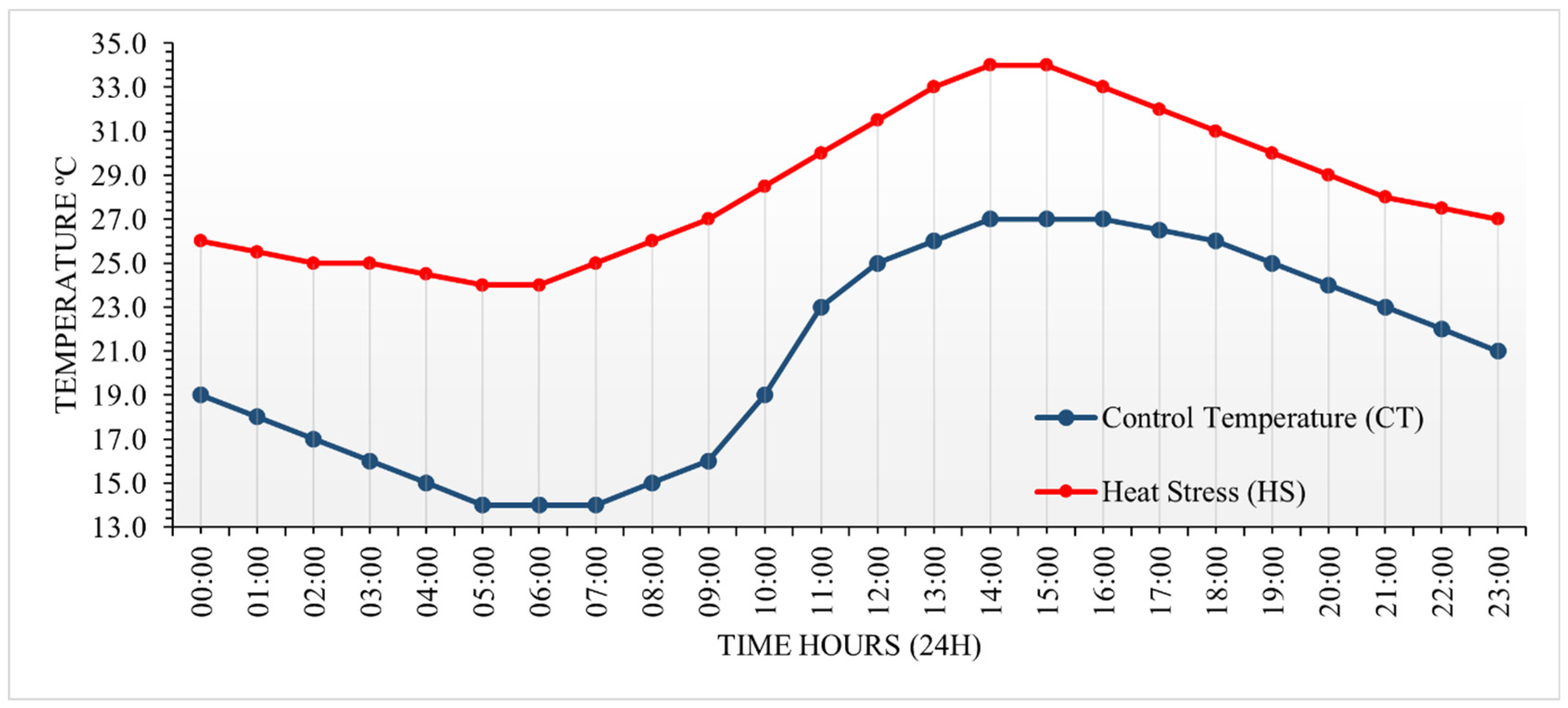
Figure 2.
Pollen viability was assessed under an optical microscope using red acetic carmine staining (scale bar = 20 µm). Arrows indicate unviable (uncolored) and viable (colored) pollen grains.
Figure 2.
Pollen viability was assessed under an optical microscope using red acetic carmine staining (scale bar = 20 µm). Arrows indicate unviable (uncolored) and viable (colored) pollen grains.
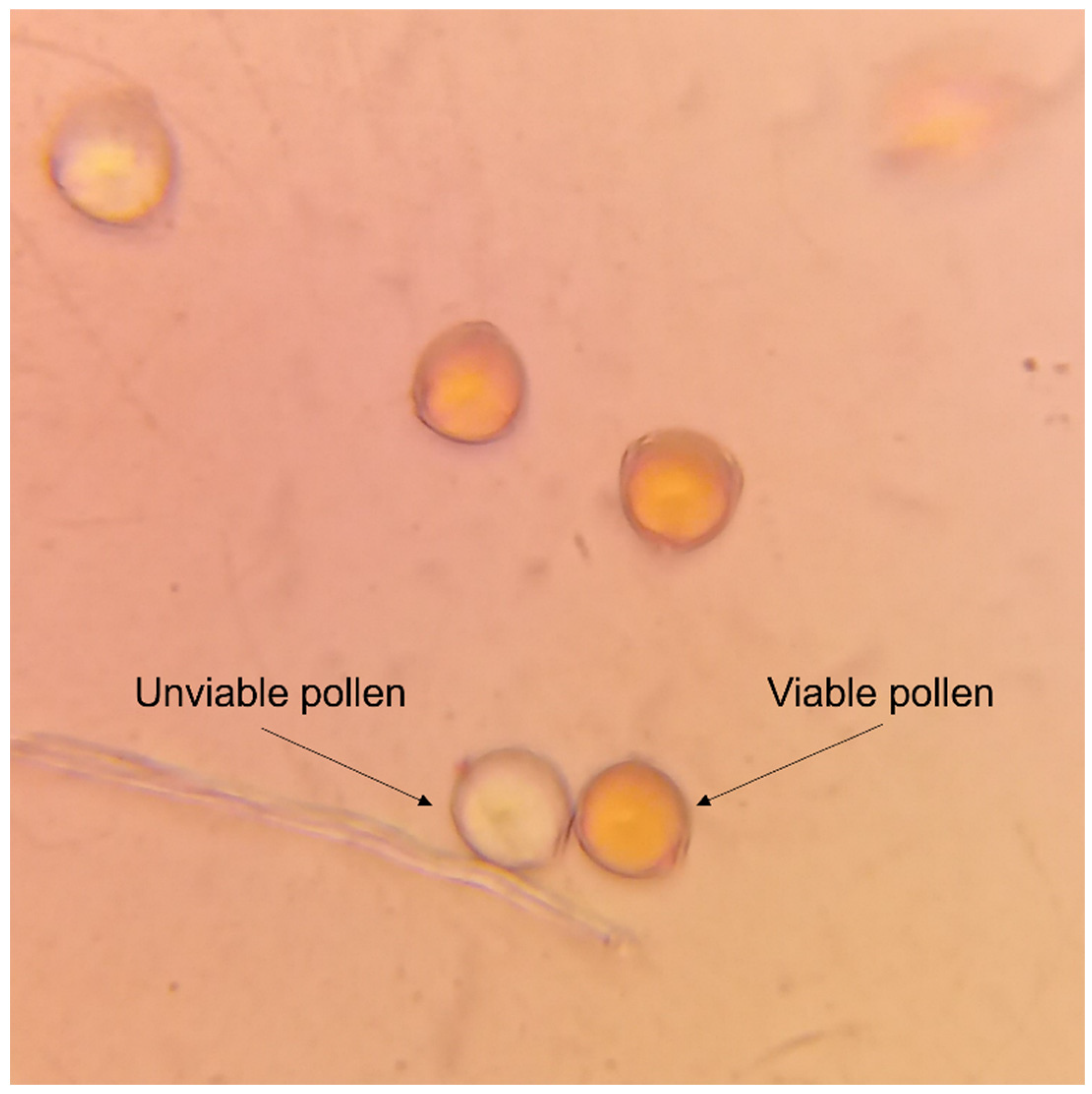
Figure 3.
A non-parametric Wilcoxon-Mann-Whitney test comparing pollen size from each genotype of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank (referred to as local code BGB) under both control (CT; represented by green points) and heat stress (HS; represented by orange points) conditions. Each accession (BGB code) is depicted in the violin plot.
Figure 3.
A non-parametric Wilcoxon-Mann-Whitney test comparing pollen size from each genotype of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank (referred to as local code BGB) under both control (CT; represented by green points) and heat stress (HS; represented by orange points) conditions. Each accession (BGB code) is depicted in the violin plot.
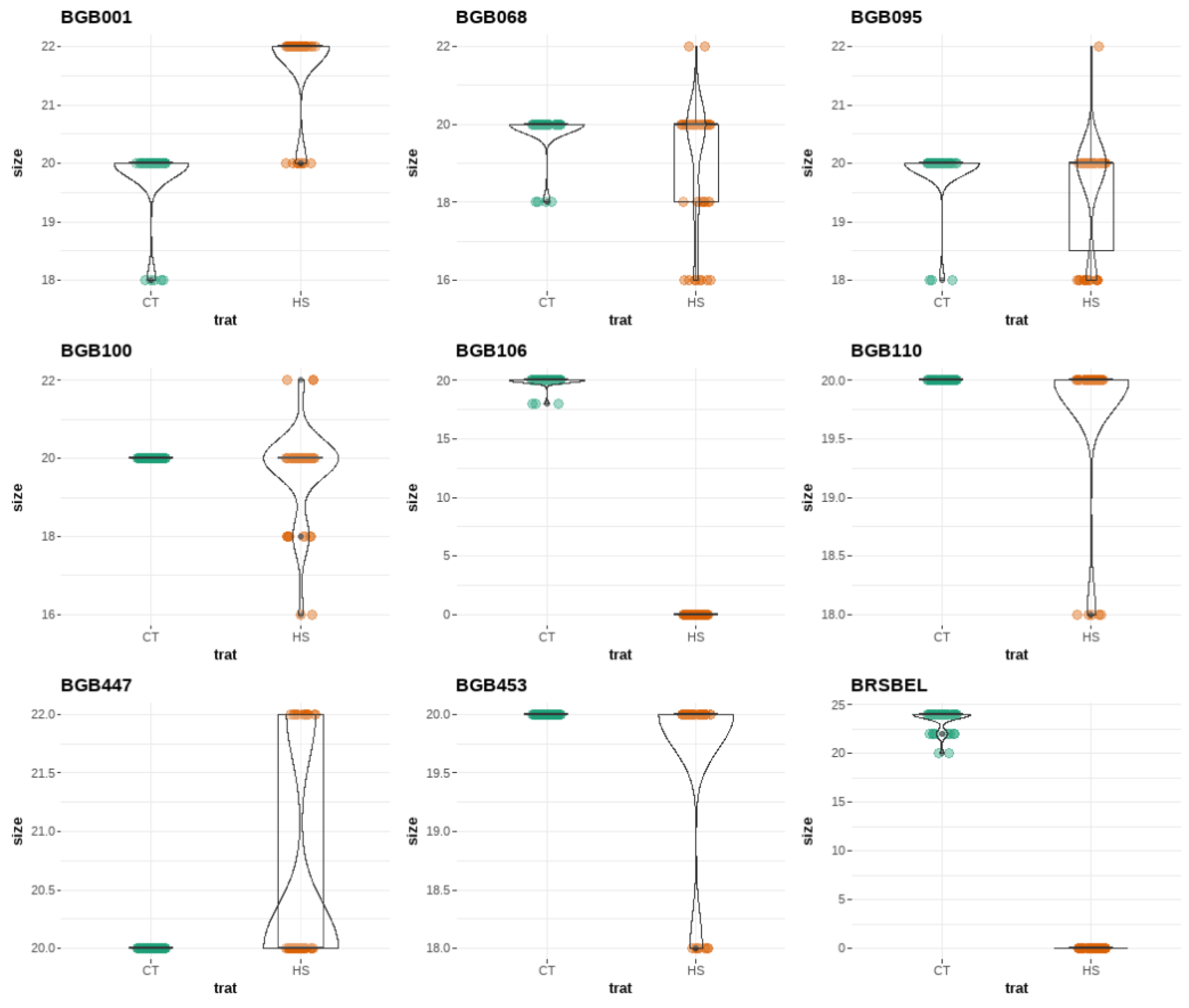

Table 1.
Accessions of potato wild species conserved at Embrapa evaluated in this study. The heading information is followed by local code, genesys code, scientific name (species), origin (city, state, country), latitude (Lat), and longitude (Lon) in UTM.
Table 1.
Accessions of potato wild species conserved at Embrapa evaluated in this study. The heading information is followed by local code, genesys code, scientific name (species), origin (city, state, country), latitude (Lat), and longitude (Lon) in UTM.
| Local code* | Genesys code** | Species | Origin | Lat | Lon |
| BGB447 | BRA 00183755-8 | S. malmeanum | Porto Lucena, RS, Brazil | -27.856100 | -55.016400 |
| BGB100 | BRA 00167017-3 | S. chacoense | Catamarca, Argentina | -28.471588 | -65.787721 |
| BGB110 | BRA 00167028-0 | S. chacoense | Unknown | - | - |
| BGB106 | BRA 00167023-1 | S. chacoense | Unknown | - | - |
| BGB095 | BRA 00167447-2 | S. chacoense | Cordoba, Argentina | -31.13333 | -64.48333 |
| BRSIPR-BEL | BRA 00167251-8 | S. tuberosum | Brazil | - | - |
| BGB001 | BRA 00167007-4 | S. commersonii | Ijuí, RS, Brazil | -28.388000 | -53.915000 |
| BGB453 | BRA 00183760-8 | S. commersonii | Herval, RS, Brazil | -32.023600 | -53.395600 |
| BGB068 | BRA 00167420-9 | S. commersonii | São Gabriel, RS, Brazil | -30.336000 | -54.32000 |
* AleloVegetal, available at < https://av.cenargen.embrapa.br/avconsulta/Passaporte/busca.do >; ** Genesys, available at https://www.genesys-pgr.org/.
Table 2.
Analysis of Variance (ANOVA) for pollen viability of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank cultivated under both control (CT) and heat stress conditions (HS).
Table 2.
Analysis of Variance (ANOVA) for pollen viability of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank cultivated under both control (CT) and heat stress conditions (HS).
| Df | Sum Sq | Mean Sq | F-value | Pr (>F) | |
|---|---|---|---|---|---|
| Block | 1 | 82.3 | 5 | 2.450 | 0.141 |
| Genotype | 6 | 4278.7 | 4 | 21.228 | 0.000* |
| Temperature | 1 | 869.1 | 6 | 25.872 | 0.000* |
| Genotype×Temperature | 6 | 340.9 | 3 | 1.532 | 0.200 |
| Residue | 13 | 436.7 | 2 | ||
| Total | 27 | 6007.7 | 1 | ||
| CV (%) | 7.13 |
The column headers include Block, Genotype, Temperature, Genotype × Temperature interaction, Residue, Total, and Coefficient of Variation (CV %). The row headers include Df: Degree of Freedom, Sum Sq: Sum of Squares, Mean Sq: Mean squares, F distribution, and *Pr(>F): p-value at <0.5%.
Table 3.
Genetic Parameter Estimates for Pollen Viability of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank under Control (CT) and Heat Stress (HS) conditions.
Table 3.
Genetic Parameter Estimates for Pollen Viability of wild potatoes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank under Control (CT) and Heat Stress (HS) conditions.
| Components | Pollen Viability |
|---|---|
| phenotypic variance | 210.29 |
| genotypic variance | 164.10 |
| residual variance | 35.66 |
| genotype × treatment interaction variance | 10.54 |
| broad sense heritability (%) | 78% |
| accuracy in genotypic selection | 0.96 |
| phenotypic coefficient of variation | 17.84 |
| genotypic coefficient of variation | 15.76 |
| genetic gain % | 28.68 |
| Genetic advance | 23.30 |
| General average | 81.29 |
Table 4.
Pollen viability (%) of the wild potato genotypes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank under control (CT) and heat stress (HS) conditions.
Table 4.
Pollen viability (%) of the wild potato genotypes (Solanum sect. Petota, Solanaceae) from Embrapa Potato Genebank under control (CT) and heat stress (HS) conditions.
| Genotype | Genesys code | Species | Temperature treatment | HSIpv | Score | Heritability (h2) % | |
|---|---|---|---|---|---|---|---|
| CT | HS | ||||||
| BGB447 | BRA 00183755-8 | S. malmeanum | 100 aA* | 91.5 abcA | 0.66 | Moderately tolerant | 91.32 |
| BGB100 | BRA 00167017-3 | S. chacoense | 98.5 aA | 94.0 abA | 0.36 | Tolerant | 88.35 |
| BGB110 | BRA 00167028-0 | S. chacoense | 96.0 aA | 83.5 abcdA | 1.01 | Susceptible | 88.56 |
| BGB095 | BRA 00167447-2 | S. chacoense | 89.5 abcdA | 71.5 bcdeA | 1.57 | Susceptible | 83.16 |
| BGB001 | BRA 00167007-4 | S. commersonii | 86.5 abcdA | 65.5 deA | 1.89 | Susceptible | 58.82 |
| BGB453 | BRA 00183760-8 | S. commersonii | 69.0 bA | 69.5 bcA | 0.06 | Tolerant | 83.82 |
| BGB068 | BRA 00167420-9 | S. commersonii | 68.5 cdeA | 54.5 eA | 1.59 | Susceptible | 87.72 |
* Significant differences determined by Tukey’s test (P<0.05) are indicated by average values labeled with different letters. These letters are represented in lowercase within the column when comparing genotypes within the same treatment and in upcase letter within the row when comparing genotypes from the two treatments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



