
Submitted:
02 November 2023
Posted:
03 November 2023
Read the latest preprint version here
Abstract
Peach (L.) is a kind of fruit tree with considerable economic and nutritional significance. However, it shows a high sensitivity to drought stress, which has a profound impact on its growth. L-methionine (MET) and L-proline (PRO) are two amino acids synthesized by plants, which play an important role in abiotic stress response. However, their exact mechanism of action remains elusive. Through physiological analysis, MET (500ppm) and PRO (500ppm) treatments can effectively reduce the adverse effects of drought stress on the growth of peach seedlings. Studies have shown that amino acid (AA) can effectively inhibit the stomatal opening of plant leaves and increase the photosynthetic rate. In addition, the application of MET or PRO promoted the accumulation of carbohydrates in leaf tissues, increased intracellular water potential (24%, 24%), reduced osmotic potential (21%, 24%), and alleviated membrane damage caused by drought. Using Evans Blue, nitroblue tetrazolium (NBT) and DAB (3, 3-diaminobenzidine) staining experiments, We confirmed that the increase in antioxidant enzyme activity in leaves may be due to the removal of reactive oxygen species (ROS) in vivo, namely the downregulation of malondialdehyde (18%, 19%) ,peroxidase (50%, 5%) and superoxide dismutase (12%, 14%) activities. In addition, the application of exogenous amino acids also improved the root morphology and vitality of peach seedlings, which was conducive to absorbing external buffer substances and alleviating drought stress. In summary, the application of exogenous amino acids can effectively alleviate the effects of drought stress on plants and reduce the harm.
Keywords:
peach
; amino acid
; drought stress
; osmotic
; physiological index
1. Introduction
Peach (scientific name: Prunus persica L.): Rose family, peach plants, originated in the western region of China. Currently, 29 provinces and autonomous regions in China have industrialized peach cultivation. Therefore, peaches have become the third largest deciduous fruit tree after pears and apples in our country [1]. As one of China's most important crops, peaches are widely cultivated in China's arid and semi-arid areas. However, in recent years, due to the increase of drought caused by global climate change, drought has become a major factor limiting the growth and development of peach trees, affecting the development of the global peach industry to a large extent, resulting in greater crop loss [2]. Studies have shown that drought stress adversely affects plant growth by reducing leaf relative water content (RWC), root length, leaf area and leaf water potential [3]. The long-term stress of drought resulted in the accumulation of a large number of ROS in plant leaves, resulting in membrane lipid peroxidation, which damaged cell membrane function and weakened photosynthesis, which seriously affected the growth of plant seedlings [4].
Amino acids (AA) are organic nitrogen compounds that are building blocks of proteins and play a role in protein synthesis and it plays structural, metabolic and transport functions in plants [5,6]. In addition Amino acids are precursors of plant hormones and other growth agents, and have important functions such as improving product quality, increasing resistance to abiotic stresses,facilitating transport-assimilation-nutrient utilization, and improving plant metabolic efficiency. Amino acids are involved in stress-mediated signaling responses, cryoprotectant, and secondary metabolism [7,8]. AA has a high ROS clearance capacity and acts as a precursor to bioregulators involved in carbohydrate and protein metabolic pathways, enhancing photosynthesis and promoting plant growth [9]. Met is a sulfur-containing amino acid (SAA) that is essential for biological growth. Studies have shown that Met induces the production of hydrogen peroxide (H2O2), a key element of plant defense signaling, and up-regulates the expression of a series of defense-related genes [10].Met plays an important role in the biosynthesis of auxin, cytokinin and brassinolide cofactors, polyamines, vitamins, and antioxidants [11], to further synthesize defense compounds and participate in cellular homeostasis[12].In addition, Met, as a metabolic sulfide donor, produces substances such as secondary metabolites, vitamin cofactors, and phytochelates in organisms. Pro plays an important role in the growth process of many different organisms, including tolerance to drought stress in many higher plants [13]. It also changes the concentration of osmotic components in biological cells to balance the osmotic potential of the cytoplasm [14], and as an important substance in osmotic regulation in plants, accumulates in large quantities as ROS scavenger in plants under drought stress [15,16]. Under drought stress, tomato can increase the gene expression of SlProT1 and SlProT2 in tomato, increase the plant Pro content, improve the plant osmoregulation ability, and alleviate the damage caused by drought stress [17]. By regulating Pro metabolism, Glu can activate the expression of calc-dependent protein kinase (CPK5) and ICS1 genes related to salicylic acid synthesis, increase the levels of salicylic acid and cytoplasmic Ca2+, and enhance the drought resistance of rape plants [18]. These studies have shown that amino acids play a key role in alleviating abiotic stress in plants. However, there are few studies on fruit trees, especially peach trees (plum), which are mostly in the process of research and discussion, and there is a lack of evidence that can specifically show that amino acids can alleviate drought stress in plants. Therefore, the purpose of this study was to investigate whether Met and Pro can alleviate the effects of drought stress on peach plants, and to prove this through physiological characteristics. The results showed that the application of Met and Pro could ameliorate drought-related oxidative damage and alleviate the effects of drought. These studies provide strategies for increasing agricultural fruit tree yields by improving drought tolerance.
2. Results
2.1. Methionine and Proline Could Relieve Drought Stress of Peach Seedlings
500ppm Met and Pro treatment mitigated the drought stress of peach seedlings (Figure 1). Relative water content (RWC) in plant leaves is one of the factors reflecting the water status of plants. Under drought conditions, RWC content in plants is greatly reduced, but the application of Met and Pro with exogenous aid can effectively slow down the decrease of RWC content (Figure 2A). The content of EL in plants under natural drought stress was significantly higher than that in peach leaves under normal conditions, and the content of EL in plants under stress was significantly lower after Met and Pro treatment (Figure 2B). The results showed that Met and Pro were able to mitigate the effects of drought, increasing the RWC content and decreasing the EL content in the leaves.
3.2. Effects of Exogenous Amino Acids on Leaf Photosynthetic Parameter
In peach seedlings treated with water (CK), the values of leaf net photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate (Tr) significantly decreased after drought stress, while the intercellular carbon dioxide concentration (Ci) was significantly increased (Figure 3). Exogenous application of Met and Pro significantly slowed down the decline of Pn, Gs, Tr under drought stres. Compared to drought stress under CK treatment, pn, Gs, Tr of MET-D and PRO-D peach saplings increased by 147.06%, 38.19%, 54.59%, 123.53%, 40.58%, 44.64%. In turn, plant leaves treated with Met-D and Pro-D showed reductions in Ci values of 19.02 percent and 13.20 percent, respectively, compared to the CK-D group. Experiments have shown that Met and Pro can alleviate plant stress by improving the photosynthetic performance of plants through external auxiliary application.
3.3. Effects of Exogenous Amino Acids on Stomata in Leaves
In drought conditions, plants reduce leaf transpiration by closing stomata and other means to ensure the water content needed for leaf growth. In group CK-D, the stomata of the plants remained open, which was not conducive to maintaining the water required for normal growth of the leaves. In contrast, leaf stomata were closed in Met-D and Pro-D groups (Figure 4). Experiments have shown that Met and Pro treatments can alleviate drought stress by promoting stomatal closure to reducing water transpiration.
3.4. Effects of Exogenous Amino Acids on Leaf Pigment
Drought stress reduced levels of SPAD , chlorophyll-A, chlorophyll-B and carotenoids in peach seedling leaves by 25.48%,27.77%, 21.87% and 28.91%, respectively, while Met-D treatment increased levels by 12.59%,17.87%, 19.12% and 7.48%, respectively. The blades after Pro-D treatment increased by19.64%, 18.76%, 29.11% and 17.68% (Figure 5). In contrast, the effect of Pro applied by exogenous aid is more pronounced. The results suggest that the application of Met and Pro may facilitate the synthesis of pigments in leaves to alleviate drought stress. Exogenous amino acids can alleviate drought stress by affecting photosynthetic pathways and electron transfer pathways between leaf cells.
3.5. Effects of Exogenous Amino Acids on Chlorophyll Fluorescence of Peach Seedlings
In the absence of Met and Pro treatment, the initial fluorescence value (F0), maximum fluorescence value (Fm), maximum quantum yield (Fv/Fm) of PSII, actual photochemical efficiency (FPSII) and photochemical quenching coefficient (qP) of peach seedlings treated with drought were significantly reduced. After Met-D and Pro-D treatment, the contents of Fo, Fm, Fv/Fm, FPSII and qp increased by 32.98%, 19.29%, 2.23%, 9.01%, 2.74% and 44.45%, 2.48%, 4.25%, 5.01% and 2.94%, respectively (Figure 6).
3.6. Effects of Exogenous Amino Acids on the Function of Leaf Penetration System
Drought stress significantly increased the contents of soluble sugar, free amino acid, free proline and glutathione in the leaves of peach seedlings (Figure 7). In contrast, Met-D and Pro-D treatments significantly increased the antioxidant capacity of leaves under drought conditions. Compared with CK-D, levels of soluble sugars, free amino acids, free proline and glutathione increased by 1.40%, 4.62%, 8.23%, 19.13% and 7.26%, 5.6%, 13.97% and 10.52%, respectively, after Met-D and Pro-D treatment. Studies have shown that the application of Met and Pro can improve the antioxidant capacity of leaves and alleviate drought stress.
3.7. Effects of Exogenous Amino Acids on Active Oxygen Content and Antioxidant Enzyme Activity in Leaves
Under drought stress, the levels of O2- and H2O2 and antioxidant levels in leaves of peach seedlings in different treatment groups were significantly different.Under drought conditions, the accumulation levels of O2- and H2O2 in the peach leaf are higher and the degree of oxidative stress in the peach leaf is higher. The lower levels of O2- and H2O2 in the leaves of the Met-D and Pro-D groups compared to the levels in the leaves of the Met-D and Pro-D groups suggests that Met and Pro may alleviate oxidative damage caused by ROS. CAT, POD and SOD activity were significantly increased in the Met-D and Pro-D treatment groups by 1.75%, 5.47%, 5.26% and 5.94%, 8.56% and 9.69%, respectively, compared to the CK-D treatment group. Compared to the control group, MDA levels in the peach leaf increased significantly under drought stress. The MDA content of peach leaves treated with Met and Pro was significantly lower than that of leaves under drought stress without Met and Pro treatment, but was significantly higher than that of the control (Figure 8).
3.8. Effects of Exogenous Amino Acids on Root Growth of Peach Seedlings
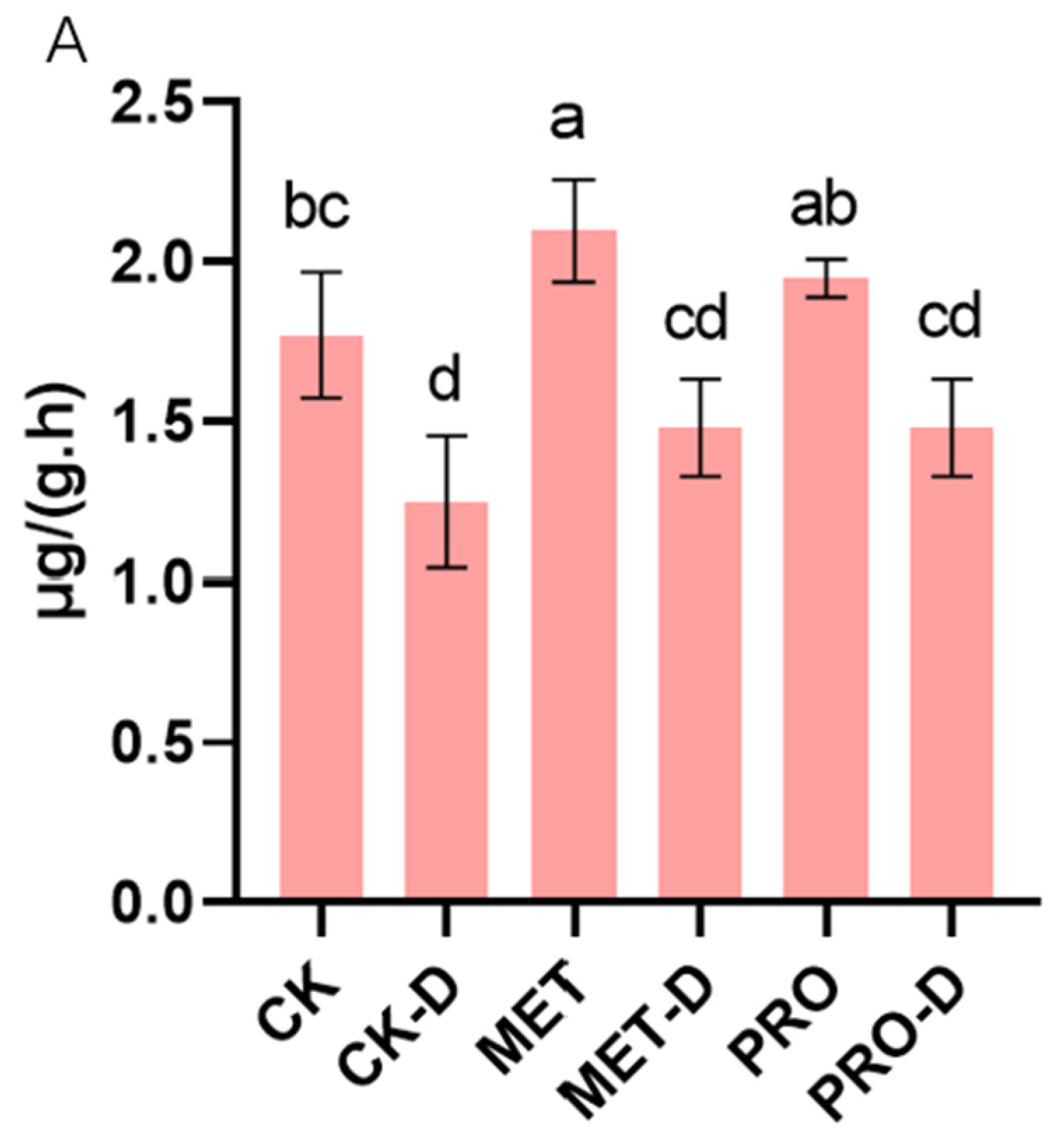
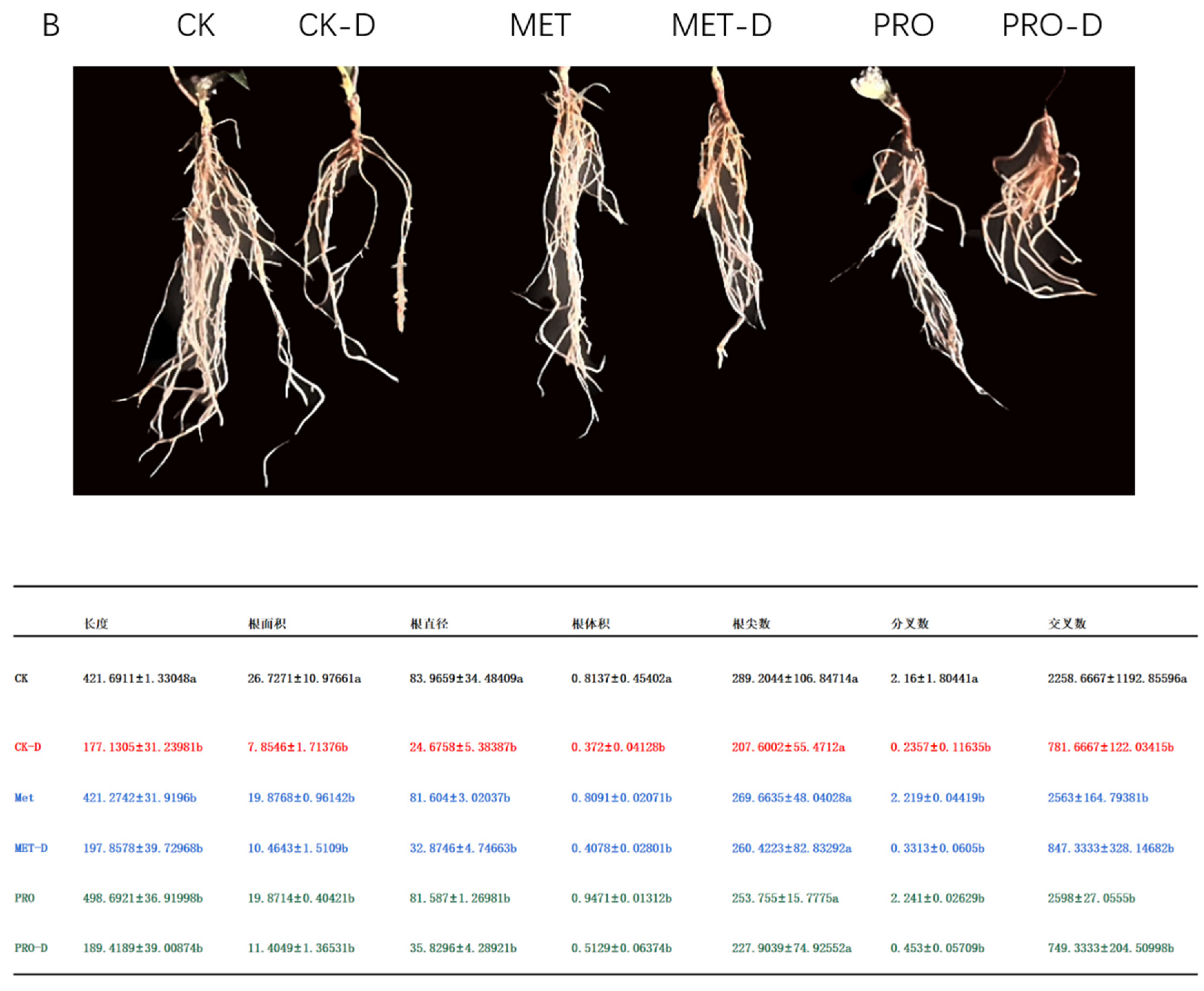
The root system is an important part of the normal growth and development of a plant, and the vitality of the root system reflects the plant's ability to absorb external substances. Under drought stress, the root activity of peach seedlings decreased by 29.59% compared with control group. Compared with CK-D, Met-D and Pro-D treatments, the root activity increased by 18.59% and 18.54%, respectively (Figure 9A). In addition to root activity, drought stress also affected plant root configuration (Figure 9B). Studies have shown that the application of Met and Pro can alleviate drought stress by improving plant root activity and structure.
3. Discussion
As a component of protein, amino acids also participate in excessive cellular reactions, so amino acids can directly or indirectly affect some physiological processes of plants, such as plant growth and development, intracellular pH control, metabolic energy generation and resistance to abiotic and biological stress [19,20,21,22,23,24,25]. Methionine (Met) and Proline (Pro) are two amino acids that play important roles in plants, among which MET plays a central role in cell metabolism, including protein synthesis, transmethylation through adenosine methionine (AdoMet), and polyamine and ethylene biosynthesis [26,27]. In bacteria, methionine in protein acts as a cellular endogenous antioxidant, and methionine residues in protein react with reactive oxygen species (ROS) to form methionine sulfoxide, thereby clearing ROS [28]. Proline, a plant multifunctional amino acid, is involved in many cell signaling pathways, accumulates under various stresses, and triggers or participates in cellular defense, including maintaining osmotic pressure homeostasis, regulating cellular state, and clearing ROS [29,30]. The effects of exogenous Met and Pro on plant photosynthetic performance under drought conditions have been studied and then investigate the mechanism of Met and Pro in photosynthesis by evaluating photosynthetic pigments, gas exchange parameters, and chlorophyll fluorescence parameters. Osmoregulatory and antioxidant systems were examined later to describe our findings, However the use of Met and Pro under abiotic stress is not well-studied and the mechanism of partial inhibition is unclear.
The morphological observation of seedling leaves showed that the deeper the degree of drought, the more serious the water loss of leaves and the greater the degree of leaf curl (Figure 1), the leaves are adjusted to ensure that they contain normal water content, thus enhancing the response ability of plants to drought stress [31]. Drought stress reduced the relative water content (RWC) of peach leaves, and after irrigating seedlings with Met and Pro roots, RWC in one of the treated subgroups was significantly higher than RCW in the CK-D subgroup (Figure 2). Relative electrolyte leakage (REL) is a good predictor of membrane permeability and drought tolerance [32]. Results showed a significant improvement in REL for CK-D treatment, but there was no significant difference between the MET-D and Pro-D treatment subgroups and the Met and Pro treatment subgroups under drought conditions. The results showed that Met and Pro therapy prevented cell damage under drought conditions.
Photosynthesis is an efficient way for plants to obtain carbon and energy, but this process is particularly vulnerable to environmental influences [33]. Experiments have shown that drought stress can destroy the pigment complex in plants, hinder electron transfer, damage the structure of chloroplasts, and then affect the reduction of photosynthetic rate [34,35]. Under the influence of drought stress, the chlorophyll content and stomatal conductance of plants decrease, and the Pn value decreases [36]. Drought has been found to reduce levels of chlorophyll a, chlorophyll b and carotenoids in plant leaves. However, the application of Met and Pro mitigated the adverse effects of drought on the reduction of chlorophyll content. Our experimental results are consistent with other studies that have found that photosynthetic pigments in crops decrease under drought stress [37]. Drought stress reduces the size of the stomatal pore, which causes a decrease in the rate of Pn and a change in the rate of photosynthetic metabolism. In addition, drought stress reduces the concentration and transpiration rate of carbon dioxide through stomata [38]. It has been shown that Met and Pro treatments promote stomatal closure under drought stress to reduce leaf transpiration. The results show that Met and Pro enhance drought tolerance in plants through a variety of mechanisms, including leaf hole restriction, protection from chlorophyll degradation, and increased photosynthetic capacity.
Evaluation of leaf chlorophyll fluorescence values is an essential technique for determining plant water status under drought stress, as these characteristics can quickly, accurately and safely indicate the effects of drought stress on plant photosynthesis [39]. Fo is a key indicator for assessing plant stress damage, and Fm can indicate the electron transport capacity of PS II after dark adaptation [40]. Studies have shown that both Fo and Fm decrease significantly under drought stress, but Met and Pro treatments can significantly delay the downward trend (Figure 7). Fv/Fm is a good indicator to determine the degree of plant leaf damage, and Fv/Fm decreases with the increase of environmental stress [41,42].
Plant resistance to abiotic stress is heavily dependent on the accumulation of antioxidant [43]. Both Met and Pro have substantial effects on the various percolating fluids in our survey. The levels of soluble sugar and glutathione were higher in plants under drought stress than in plants not treated with Met and Pro (Figure 6). The levels of proline and free amino acids in plants are the main factors regulating the osmotic potential of cells. The establishment of these molecules contributes to the integrity of the cell membrane and also plays a role in preventing osmotic and oxidative damage [44]. In our past experimental studies, Met and Pro treatments significantly increased the proline content of hickory seedlings under drought stress. Similar results were obtained for Apple [45]. In addition to its osmoregulatory role, the accumulation of proline under drought conditions also protects the photosynthetic system from damage. [46].
After drought stress, it is very important to maintain intracellular ROS level and osmotic balance [47]. In our study, drought stress leads to excessive ROS accumulation in the leaves, which damages the plant cell membrane system and leads to electrolyte leakage, which in turn leads to chlorophyll degradation. This process has been shown to significantly limit photochemical processes, impair photosynthesis, and accelerate leaf aging [48]. In this study, the levels of O2 -, hydrogen peroxide and MDA in peach leaves significantly increased under drought stress, but decreased significantly after the application of Met and Pro (Figure 8). The antioxidant defense system of plants is composed of both enzymatic and non-enzymatic antioxidants, which strictly controls the balance of reactive oxygen species [49]. Studies have shown that exogenous application of amino acids can control the homeostasis of reactive oxygen species (ROS) by increasing the activity of antioxidant enzymes in apple, thereby protecting fruit from ROS damage under drought stress. According to our findings, in our study, Met and Pro treatment significantly increased SOD, POD and CAT activity levels (Figure 8). It has been found in other studies that excess reactive oxygen species can be eliminated by SOD, POD and CAT enzymes. SOD can convert O2- to hydrogen peroxide, which can then be converted back to water via POD and CAT to reduce damage from reactive oxygen species.
Pearson correlation analysis was used to investigate the interplay of these metrics to assess how Met and Pro improve plant tolerance in drought conditions. Correlation analysis in this study shows that ROS and MDA are positively correlated with enzymatic antioxidants, while RWC is positively correlated with ROS, REL, reactive oxygen species, the osmotic system, and the enzymatic antioxidant system. The results show that drought stress has a significant effect on the photosynthetic system, osmotic regulation system, and antioxidant system of the active oxygen-enzyme-promoting peach seedlings, and Met and Pro can alleviate these effects and thus drought stress.
4. Materials and Methods
4.1. Experimental Materials
This experiment was conducted in 2022 at the Horticultural Experimental Base of Shandong Agricultural University (36°17 '7459 "N, 117°16' 7712" E). The experimental materials were evenly plump peach seeds, soaked in 400ppm of gibberellin solution for one day, and then put into a virus-free and sterile culture pot for germination promotion. Peach buds with good growing conditions were then selected and transplanted into pots of 7 cm diameter and 11 cm height for culture. The culture medium is soil: substrate: vermiculite with a ratio of 2:1:0.5 and a substrate weight of about 200g per pot, mixed culture, awaiting use.
4.2. Drought Treatment
Peach seedlings of the same growth and development were divided into five groups of ten seedlings each. When the seedlings grew to about 7-8 cm, the experimental treatment was carried out on the peach seedlings at six treatment levels: (a) no treatment (CK); (b) Natural Drought(Dro); (c) 500ppm methionine root irrigation (Met); (d) 500ppm methionine root irrigation + drought (Met+ Dro); (e) 500ppm proline root irrigation (Pro); (f) 500ppm proline root irrigation + drought (Pro+Dro). Previous experiments had shown that the two amino acids applied by exogenous aid did not adversely affect the growth of peach seedlings. This experiment could be divided into two groups: one group was treated with regular irrigation (CK, Met, Pro), and the other group was treated with natural drought after 7 days of normal irrigation (CK-D, MET-D, PRO-D). After a total of 14 days of treatment, sampling, all the leaves are cleaned with water, frozen with liquid nitrogen, stored at -80° for use.
4.3. Measurements of Physiological and Biochemical Indicators
4.3.1. Light and Index Measurements
4.3.1.1. Measurements of the Photosynthetic Parameters
On the 14th day after treatment, the photosynthetic parameters of the peach seedlings were measured from 10:00 am to 11:30 am in suitable light conditions. The CIRAS-3 Portable Photosynthesis System (PP System, Massachusetts, USA) was used by us to measure the following parameters: net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) and intercellular carbon dioxide concentration (Ci).
4.3.1.2. Chlorophyll Fluorescence Parameter Determination:
The relative chlorophyll content (SPAD) was measured 5 times using the SPAD502 instrument and its average value was calculated. The leaves was we kept in the dark for 30 minutes and then used a portable pulse-debug fluorometer (Hansatech, UK) to determine the fluorescence parameters: minimum fluorescence (F0), maximum fluorescence (Fm), maximum quantum efficiency (Fv/Fm) and maximum fluorescence (FV/FM).
The formula for calculating some of the parameters is:
4.3.2. Relative water content and relative electrical conductivity of blades
Three peach seedlings were randomly selected for each treatment, and two leaves from the same part of each peach seedling were immediately placed in a petri dish with known weight, and the fresh leaf weight (Wf) was measured. The leaves were then immersed in distilled water for about 1 hour, then removed and weighed to determine the saturated fresh weight (WT) of the sample. Dry weight (Wd) is then obtained by drying the blades to a constant weight. The relative water content (RWC) is determined by the following formula [50] : (Wf-Wd)/(Wt-Wd) ×100%
the magnitude of relative conductivity (REL) by randomly taking 10 small disks from peach leaves and placing them into a 20 mL centrifuge tube containing 10 mL of distilled water. After 4 hours at room temperature, the conductivity of the solution was tested using a Raymag DDS-307 conductivity meter, designated S1. At the same time, the conductivity of the distilled water was measured and assigned to s0. The centrifuge tube was then placed in a 100°C water bath for 20 min. After cooling, shake the mixture well and measure the conductivity, designated s2. To represent the relative permeability of the plasma membrane, the relative electrolyte leakage was calculated using the following formula [51]: REL (%) = (S1-S0)/(S2-S0) ×100%.
4.3.3. Determination of Leaf Stomata
Nikon positive microscope (Nikon Precision Machinery (Shanghai) Co., LTD., China) was used to observe the stomatal opening degree of plants under drought stress. Different treated leaves was selected from the same section, tore the membrane from the back of the leaf with tweezers, placed it on the slide, secured it with water, then covered the slide, secured it on the platform, and adjusted the eyepiece to 400X. The state of the leaf stomata was observed.
4.3.4. Determination of Root Vitality and Configureuration Indexes of Peach Seedlings
4.3.4.1. The root activity is usually measured by TTC method [52]. 0.1g fresh root sample is weighed and placed in a test tube, and 0.1mmolL phosphate buffer (PH=7.8) and 2.5mL 0.4%TTC are added, so that the root sample is fully immersed in the solution and bathed in a 37°C water bath for 1h. The reaction was terminated by adding 1mol/L H2SO41mL, the roots were removed immediately, the water was dried with absorbent paper, then ground and filtered with ethyl acetate, and the absorption value of the solution at 485nm was measured with the Pharma-Spec UC-2450 ultraviolet spectrophotometer [53].
4.3.4.2 Desktop scanner Epson Expression 1200XL was used to scan the roots, and analysis system software (WinHIZO 2003) was used to analyze the root images after scanning. Obtain total root length, total root surface area, average root diameter, root volume, and root cusp number. There are other relevant parameters as well.
4.3.5. Determination of Osmotic Regulatory Substances
4.3.5.1. Determination of Soluble Sugar Content
The soluble sugar content of the plant leaves was determined and 0.2g of the leaves were weighed, chopped and mixed in a test tube. Ten mL of distilled water was then added and the test tube was placed in a boiling water bath for 30 min to generate an extraction solution which was filtered and diluted to a volume of 25 mL bottle. The absorbance at 620 nm was determined by mixing 1 mL of the extract with 4 mL of 0.2% anthrone reagent [54].
4.3.5.2. Determination of Proline Content
The proline content of the plant leaves was determined. Firstly, 0.5g leaf sample was weighed and put into grinding, 5 mL 3% sulfosalicylic acid was added, the sample was ground to prepare homogenate, then all the samples were transferred to a test tube and boiled in a water bath for 10 min. After cooling to room temperature, 2 mL of supernatant was collected in a new test tube, and 3 mL of indanhydrin and 2 mL of glacial acetic acid were added. Then the test tube was heated in boiling water for 40 min, and finally 5 mL of toluene was added to the mixture for extraction, and the absorbance value of the toluene layer was determined by spectrophotometer at 520 nm [55].
4.3.5.3. Determination of Free Amino Acid Content
Total free amino acids were measured. Weigh about 0.5 g of leaves, 10% acetic was added acid to make a homogenized solution, then dilute to 100mL. 2 mL of the sample was extracted from the homogenate and mixed with aqueous solution of 3 mL of niacin reagent and 0.1% ascorbate. The mixture was heated in a water bath of boiling water for 15min, then cooled to room temperature, followed by 5ml of 60% ethanol solution, and determined at 570nm for absorbance [56].
4.3.5.4. Determination of Glutathione Content
Glutathione content was determined using the glutathione GSH/GSSG kit.
4.3.6. Determination of Active Oxygen Species in Leaves
The O2 content in the leaves was measured. 1g sample peach leaves was chopped and took and added 3 mL phosphate buffer (pH = 7.8). It was ground in an ice bath, followed by centrifuge at 4° centrifuge 4000r/min for 15 min. The supernatant was collected and mixed with 0.1 mL 10 mmol/L hydroxylamine hydrochloride solution and incubated at 25°C for 20 min. Then 1 mL 17 mmol/L p-aminobenzenesulfonic acid and 1 mL 7mmol/L naphthylamine solution were added and incubated at 25°C for 20 min. Then, Equal volume of chloroform was added to extract the pigment and centrifuged it at 10000 r/min for 3 min. Pink extracts were collected and absorbance was determined at 450 nm [57].
4.3.7. Determination of MAD Content,CAT,POD and SOD Activity in Leaves
4.3.7.1. Determination of MAD Content
Malondialdehyde (MDA) is a biomarker of lipid peroxidation caused by oxidative stress. 0.2 g of leaf samples with 2 mL of 0.1% trichloroacetic acid (TCA) were mixed in an ice bath and ground them to prepare homogenates. The homogenate was centrifuged at 4000 r/min for 10 min. After completion, 1 mL of supernatant was collected and mixed with 4 mL 20%TCA (containing 0.5% thiobarbituric acid [TBA]), the mixture was placed in a boiling water bath for 60 min, and the mixture was rapidly cooled on ice at 4°C. After centrifugation at 4000 rpm for 10 min, absorbance OD values at 600, 532 and 450 nm were measured [58].
4.3.7.2. Determination of SOD Activity of CAT POD
Chop and take 0.5g peach seedling leaves, add 4 mL phosphate buffer (0.05mmol, ph=7.8), grind them into homogenate under ice bath condition, transfer to centrifuge tube, centrifuge at 4°C and 12000 r/min for 20 min, collect supernatant and store at 4°C for use.
0.2 mL of supernatant was added to 2.8 mL of 30 ml of H2O2 and measured the absorbance at 240 nm after full mixing.The initial absorption was measured value a1 and absorption value a2 after 1 min. The activity of CAT enzyme was expressed by the absorbance change per unit time [59].
The formula is calculated as follows.
Total CAT activity (△OD240˙min-1˙g-1˙FW) =(OD240*V)/(a*W*t)
20ul of supernatant was added into the colorimetric cup, add 20ul of phosphoric acid buffer(0.1mol/L pH=6.0)solution to the contrast, add 3ml of reaction solution, read OD value at 470nm immediately and time, read it every 1min .
The formula for the calculation is as follows.
Total POD activity (△OD470˙min-1, g-1, FW) = (OD470*V)/(a*W*t)
Take a tube of the same type, and add 50 l. of supernatant; a buffer solution of 50ul 0.05mol/L pH=7.8 phosphoric acid was added to the four controls, and 3ml of reaction liquid was added to all. Two controls were placed in the dark, and the other tubes were placed under 4000xl (33% light intensity) sunlight for 20-30min. (All tubes were required to be exposed to the same light, and the time was shortened when the temperature was high. After the reaction was over, the absorbance was measured at 560nm using the unilluminated control as a blank [60].
The formula is calculated as follows.
Total SOD activity (units˙s-1˙Fw) =(ACK-A *V)/(0.5*ACK*W*Vt)
Statistical analysis
SPSS 26.0 (IBM, New York, USA) statistical analysis software was used to conduct one-way ANOVA and Duncan's multiple comparison test. The statistical significance was 5%, assuming 5% (P < 0.05). The data on all tables are expressed as average ± standard deviation (error bar).
5. Conclusions
In this study, both Met and Pro were able to mitigate the growth of peach saplings under drought stress. The results showed that drought stress could be alleviated by various mechanisms, including: (1) Met and Pro could effectively increase the chlorophyll content of peach leaves, increase the stomatal openness of leaves, increase the net photosynthetic rate, and thus enhance photosynthesis, so as to reduce the damage degree of drought stress on the photosystem; (2) Met and Pro can alleviate osmotic stress. The relative conductivity of the control vane, The water content of the leaves is maintained. (3) Met and Pro can increase the content of antioxidants, reduce the content of ROS in plants, and regulate the activity of antioxidant enzymes . Overall, Met and Pro can help reduce drought stress levels. The results of this study shed light on the mechanisms by which Met and Pro alleviate drought stress in peach plants. More research is needed to better understand these pathways.
References
- Yu Ming-liang, WANG Li-Rong, WANG Zhi-qiang, Peng Futian, Zhang Fan, YE Zhengwen. 70 years of fruit science research in New China -- Peach [J]. Journal of Fruit Science, 2019,36, 1283-1291.
- Najla Ksouri, Sergio Jiménez, Christina E. Wells,Bruno Contreras-Moreira and Yolanda Gogorcena1.Transcriptional Responses in Root and Leaf of Prunus persica under Drought Stress Using RNA Sequencing. Frontiers in Plant Science. [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin Improves Drought Stress Tolerance of Tomato by Modulating Plant Growth, Root Architecture, Photosynthesis, and Antioxidant Defense System. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Roles of exogenous glutathione in antioxidant defense system and methylglyoxal detoxifification during salt stress in mung bean. Biologia Plantarum. 2015, 59, 745–756. [Google Scholar] [CrossRef]
- Liu, X.Q., Ko K.Y., Kim S.H., Lee K.S. 2008. Effect of amino acid fertilization on nitrate assimi lation of leafy radish and soil chemical properties in high nitrate soil. Commun Soil Sci Plant Anal 39: 269-281. [CrossRef]
- Sarojnee D.Y., Navindra B., Chandrabose S. 2009. Effect of naturally occurring amino acid stimulants on the growth and yield of hot peppers. J Anim Plant Sci, 5, 414-424.
- Rennenberg H, Herschbach C (2014) A detailed view on sulphur metabolism at the cellular and whole-plant level illustrates challenges in metabolite fux analyses. J Exp Bot 65:5711–5724.
- Hildebrandt TM, Nunes-Nesi A, Araújo WL, Braun HP (2015) Amino acid catabolism in plants. Mol Plant 8:1563–1579.
- Bakhoum GS, Badr EA, Sadak MS, Kabesh MO, Amin GA (2019) Improving growth, some biochemical aspects and yield of three cultivars of soybean plant by methionine treatment under sandy soil condition. Int J Enviro Res 13,35–43.
- Zhengxuan Wang, Liang Cai, Hui Li,Mingcai Liang, Yan Zhang,Qiong Wu,Lin Yang(2020) Rice protein stimulates endogenous antioxidant response attributed to methionine availability in growing rats. Journal Of Food Biochemistry. [CrossRef]
- El-Awadi M, El-Bassiony A, Fawzy Z, El-Nemr M (2011) Response of snap bean (Phaseolus vulgaris L) plants to nitrogen fertilizer and foliar application with methionine and tryptophan. Nat Sci 9,87–94.
- Paungfoo-Lonhienne C, Thierry G, Lonhienne A, Doris R, Nicole R, Michael C, Richard IW, Gamage HK, Bernard JC, Peer MS (2008) Plants can use protein as a N source without assistance from other organisms. Proc Natl Acad Sci 105:4524–4529.
- Yakhin OI, Aleksandr AL, Ildus AY, Patrick HB (2017) Biostimulants in plant science: a global perspective. Front Plant Sci 7:2049.
- Hasegawa, P.M., Bressan, R.A., Zhu, J.K., Bohnert, H.J., 2000. Plant cellular and molecular response to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 51, 463–469.
- Wated, A.F., Reinhard, L., Erner, H.R.L., 1983. Comparison between a stabile NaCl selected Nicotiana cell line and the wide type. Na, K and proline pools as a function of salinity. Plant Physiol. 73, 624–629.
- Hare PD, Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 21:79–102.
- Hare PD, Cress WA, Van Staden J (1998) Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ 21:535–553.
- Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59:206–216.
- Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ 33:453–467.
- Akbudak M.A.and Filiz E..Genome-wide investigation of proline transporter (ProT) gene family in tomato: Bioinformatics and expression analyses in response to droughtstress. Plant Physiology and Biochemistry, 2020, 157, 13-22.
- Moe, L.A. (2013). Amino acids in the rhizosphere: from plants to microbes. Am. J. Bot. 100:1692–1705.
- Watanabe, M., Balazadeh, S., Tohge, T., Erban, A., Giavalisco, P., Kopka, J., Mueller-Roeber, B., Fernie, A.R., and Hoefgen, R. (2013). Comprehensive dissection of spatiotemporal metabolic shifts in primary, secondary, and lipid metabolism during developmental senescence in Arabidopsis. Plant Physiol. 162:1290–1310.
- Zeier, J. (2013). New insights into the regulation of plant immunity by amino acid metabolic pathways. Plant Cell Environ. 36:2085–2103.
- Fagard, M., Launay, A., Cle´ ment, G., Courtial, J., Dellagi, A., Farjad, M., Krapp, A., Soulie´ , M.C., and Masclaux-Daubresse, C. (2014). Nitrogen metabolism meets phytopathology. J. Exp. Bot. 65:5643– 5656.
- Galili, G., Avin-Wittenberg, T., Angelovici, R., and Fernie, A.R. (2014). The role of photosynthesis and amino acid metabolism in the energy status during seed development. Front. Plant Sci. 5:447.
- Hausler, R.E., Ludewig, F., and Krueger, S. (2014). Amino acids - a life between metabolism and signaling. Plant Sci. 229:225–237.
- Pratelli, R., and Pilot, G. (2014). Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 65:5535–5556.
- Ravanel S, Gakière B, Job D, Douce R. The specifific features of methionine biosynthesis and metabolism in plants. Proc Natl Acad Sci USA 1998;95:7805–12.
- Levine RL, Moskovitz J, Stadtman ER. Oxidation of methionine in proteins: roles in antioxidant defense and cellular regulation. IUBMB Life 2000;50:301–7.
- Luo S, Levine RL. Methionine in proteins defends against oxidative stress. FASEB J 2009;23:464–72.
- Rehman, A.U., Bashir, F., Ayaydin, F., Kota, Z., Pali, T., Vass, I., 2021. Proline is a quencher of singlet oxygen and superoxide both in in vitro systems and isolated thylakoids. Physiol. Plant 172, 7–18.
- Szabados, L., Savoure, A., 2010. Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97.
- Han, D., Tu, S., Dai, Z., Huang, W., Jia, W., Xu, Z., et al. (2022). Comparison of selenite and selenate in alleviation of drought stress in nicotiana tabacum l. Chemosphere. 287, 132136. [CrossRef]
- un, X., Wang, P., Jia, X., Huo, L., Che, R., and Ma, F. (2018). Improvement of drought tolerance by overexpressing MdATG18a is mediated by modifified antioxidant system and activated autophagy in transgenic apple. Plant Biotechnol. J. 16, 545–557. [CrossRef]
- Muhammad, I., Shalmani, A., Ali, M., Yang, Q.-H., Ahmad, H., and Li, F. B. (2021). Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 11, 2310. [CrossRef]
- Liu, E., Mei, X., Yan, C., Gong, D., and Zhang, Y. (2016). Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agric. Water Management. 167, 75–85. [CrossRef]
- Li, C., Zhao, Q., Gao, T., Wang, H., Zhang, Z., Liang, B., et al. (2018). The mitigation effects of exogenous melatonin on replant disease in apple. J. pineal Res. 65, e12523. [CrossRef]
- Liang, B., Ma, C., Zhang, Z., Wei, Z., Gao, T., Zhao, Q., et al. (2018). Long-term exogenous application of melatonin improves nutrient uptake flfluxes in apple plants under moderate drought stress. Environ. Exp. botany. 155, 650–661. [CrossRef]
- Moaveni, P. (2011). Effect of water defificit stress on some physiological traits of wheat (Triticum aestivum). Agric. Sci. Res. J. 1, 64–68.
- Shahid, M. A., Balal, R. M., Pervez, M. A., Garcia-Sanchez, F., Gimeno, V., Abbas, T., et al (2014). Treatment with 24-epibrassinolide mitigates NaCl-induced toxicity by enhancing carbohydrate metabolism, osmolyte accumulation, and antioxidant activity in Pisum sativum. Turk J Bot. 38, 511–525. [CrossRef]
- Maxwell, K., and Johnson, G. N. (2000). Chlorophyll flfluorescence–a practical guide. J. Exp. botany. 51, 659–668. [CrossRef]
- Rao, L., Li, S., and Cui, X. (2021). Leaf morphology and chlorophyll flfluorescence characteristics of mulberry seedlings under waterlogging stress. Sci. Rep. 11, 1–11. [CrossRef]
- Pettigrew, W. (2004). Physiological consequences of moisture defificit stress in cotton. Crop Sci. 44, 1265–1272. [CrossRef]
- Raja, V., Qadir, S. U., Alyemeni, M. N., and Ahmad, P. (2020). Impact of drought and heat stress individually and in combination on physio-biochemical parameters, antioxidant responses, and gene expression in solanum lycopersicum. 3 Biotech. 10, 1–18. [CrossRef]
- Jespersen, D., Yu, J., and Huang, B. (2017). Metabolic effects of acibenzolar-smethyl for improving heat or drought stress in creeping bentgrass. Front. Plant sci. 8, 1224. [CrossRef]
- Gao, D., Shi, C., Li, Q., Wei, Z., Liu, L., and Feng, J. (2021). Drought tolerance monitoring of apple rootstock m. 9-T337 based on infrared and flfluorescence imaging. Photosynthetica. 59, 458–467. [CrossRef]
- Khan, M., Khan, N. A., Masood, A., Per, T. S., and Asgher, M. (2016). Hydrogen peroxide alleviates nickel-inhibited photosynthetic responses through increase in use-effificiency of nitrogen and sulfur, and glutathione production in mustard. Front. Plant sci. 7, 44. [CrossRef]
- Wang, P., Sun, X., Li, C., Wei, Z., Liang, D., and Ma, F. (2013). Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. pineal Res. 54, 292–302. [CrossRef]
- Gao, T., Zhang, Z., Liu, X., Wu, Q., Chen, Q., Liu, Q., et al. (2020). Physiological and transcriptome analyses of the effects of exogenous dopamine on drought tolerance in apple. Plant Physiol. Biochem. 148, 260–272. [CrossRef]
- La V. H., Lee B. R., Islam M., Mamun M., Park S. H., Bae D. W. and Kim, T. H.. Characterization of glutamate-mediated hormonal regulatory pathway of the droughtresponses in relation to proline metabolism in Brassica napus L. Plants, 2020, 9 (4),512.
- Bielsa, B., Sanz, M.A, and Rubio-Cabetas, M. J. (2021). ‘Garnem’and myrobalan‘P. 2175’: Two different drought responses and their implications in drought tolerance. Horticulturae. 7, 299. [CrossRef]
- Zhang Xiong. Determination of wheat root and pollen activity and its application [J]. Plant Physiology Bulletin, 1982, 48-50.
- Li,Y., Lv,Y., Lian,M., Peng, F., andXiao,Y.(2021). Effects of combined glycine and urea fertilizer application on the photosynthesis, sucrose metabolism, and fruit development of peach. Scientia Horticulturae. 289, 110504. [CrossRef]
- Mehdizadeh, L., Farsaraei, S., and Moghaddam, M. (2021). Biochar application modifified growth and physiological parameters of ocimum ciliatum l. and reduced human risk assessment under cadmium stress. J. Hazardous Materials. 409, 124954. [CrossRef]
- Vardharajula, S., Zulfifikar Ali, S., Grover, M., Reddy, G., and Bandi, V. (2011). Drought-tolerant plant growth promoting bacillus spp.: effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interactions. 6, 1–14. [CrossRef]
- Han, D., Tu, S., Dai, Z., Huang, W., Jia, W., Xu, Z., et al. (2022). Comparison of selenite and selenate in alleviation of drought stress in nicotiana tabacum l. Chemosphere. 287, 132136. [CrossRef]
- Zhang, X., Wang, X., Zhong, J., Zhou, Q., Wang, X., Cai, J., et al (2016). Drought priming induces thermo-tolerance to post-anthesis high-temperature in offspring of winter wheat. Environ. Exp. botany. 127, 26–36. [CrossRef]
- Li, Y., Zhao, H., Duan, B., Korpelainen, H., and Li, C. (2011). Effect of drought and ABA on growth, photosynthesis and antioxidant system of cotinus coggygria seedlings under two different light conditions. Environ. Exp. Botany. 71, 107–113. [CrossRef]
- Liang, B., Ma, C., Zhang, Z., Wei, Z., Gao, T., Zhao, Q., et al. (2018). Long-term exogenous application of melatonin improves nutrient uptake flfluxes in apple plants under moderate drought stress. Environ. Exp. botany. 155, 650–661. [CrossRef]
- Zhang, X., Wang, X., Zhong, J., Zhou, Q., Wang, X., Cai, J., et al (2016). Drought priming induces thermo-tolerance to post-anthesis high-temperature in offspring of winter wheat. Environ. Exp. botany. 127, 26–36. [CrossRef]
Figure 1.
Comparison of growth of peach seedlings affected by drought stress after Met and Pro applied by exogenous aid.
Figure 1.
Comparison of growth of peach seedlings affected by drought stress after Met and Pro applied by exogenous aid.

Figure 2.
The effects of Met and Pro treatments on leaf relative water content (A) relative electrolyte leakage (B) under controlled and drought conditions are shown in the figure. These values represent the standard deviation of the mean of the three repetitions. Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 2.
The effects of Met and Pro treatments on leaf relative water content (A) relative electrolyte leakage (B) under controlled and drought conditions are shown in the figure. These values represent the standard deviation of the mean of the three repetitions. Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
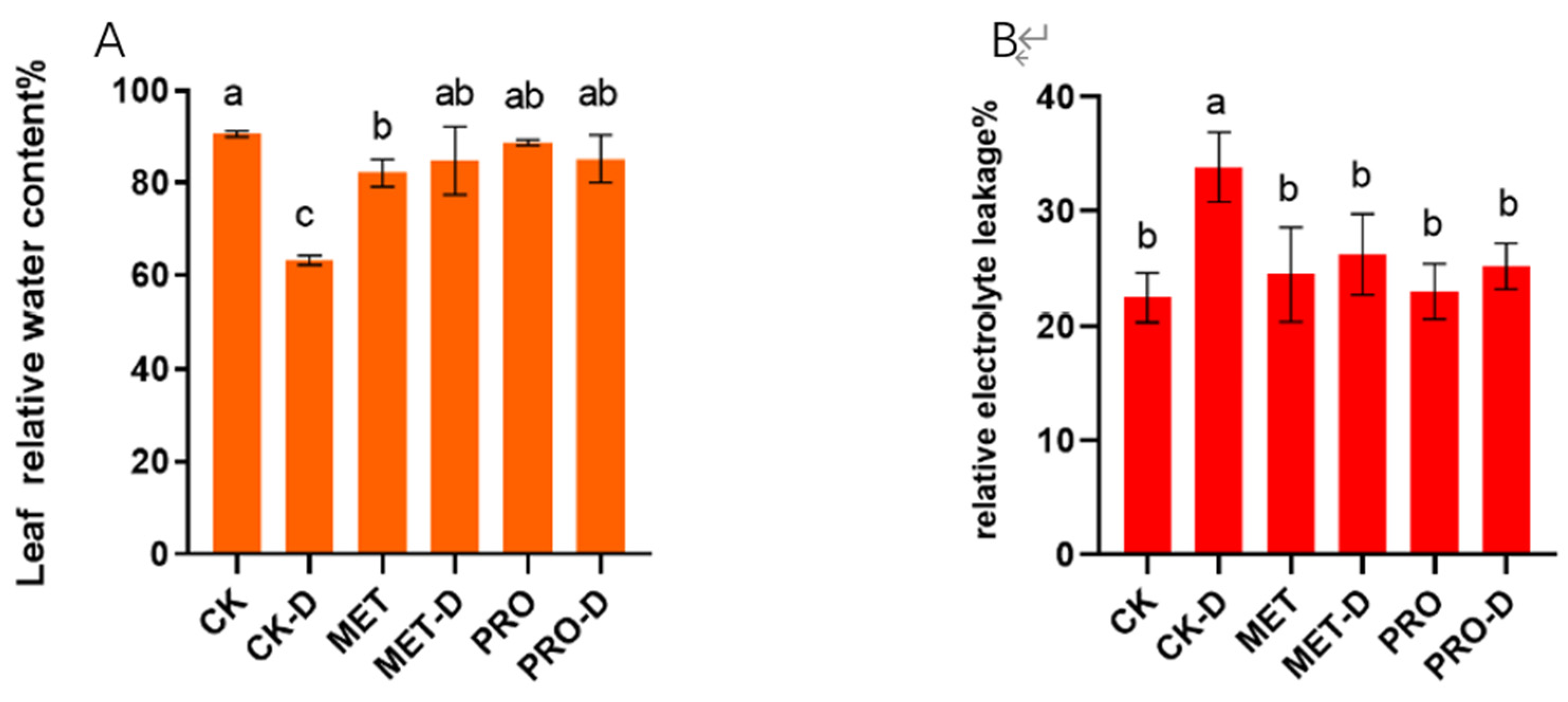
Figure 3.
Effects of Met and Pro in (A) Pn, (B) Gs, (C) Ci, (D) Tr on photosynthetic characteristics of peach grass under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 3.
Effects of Met and Pro in (A) Pn, (B) Gs, (C) Ci, (D) Tr on photosynthetic characteristics of peach grass under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
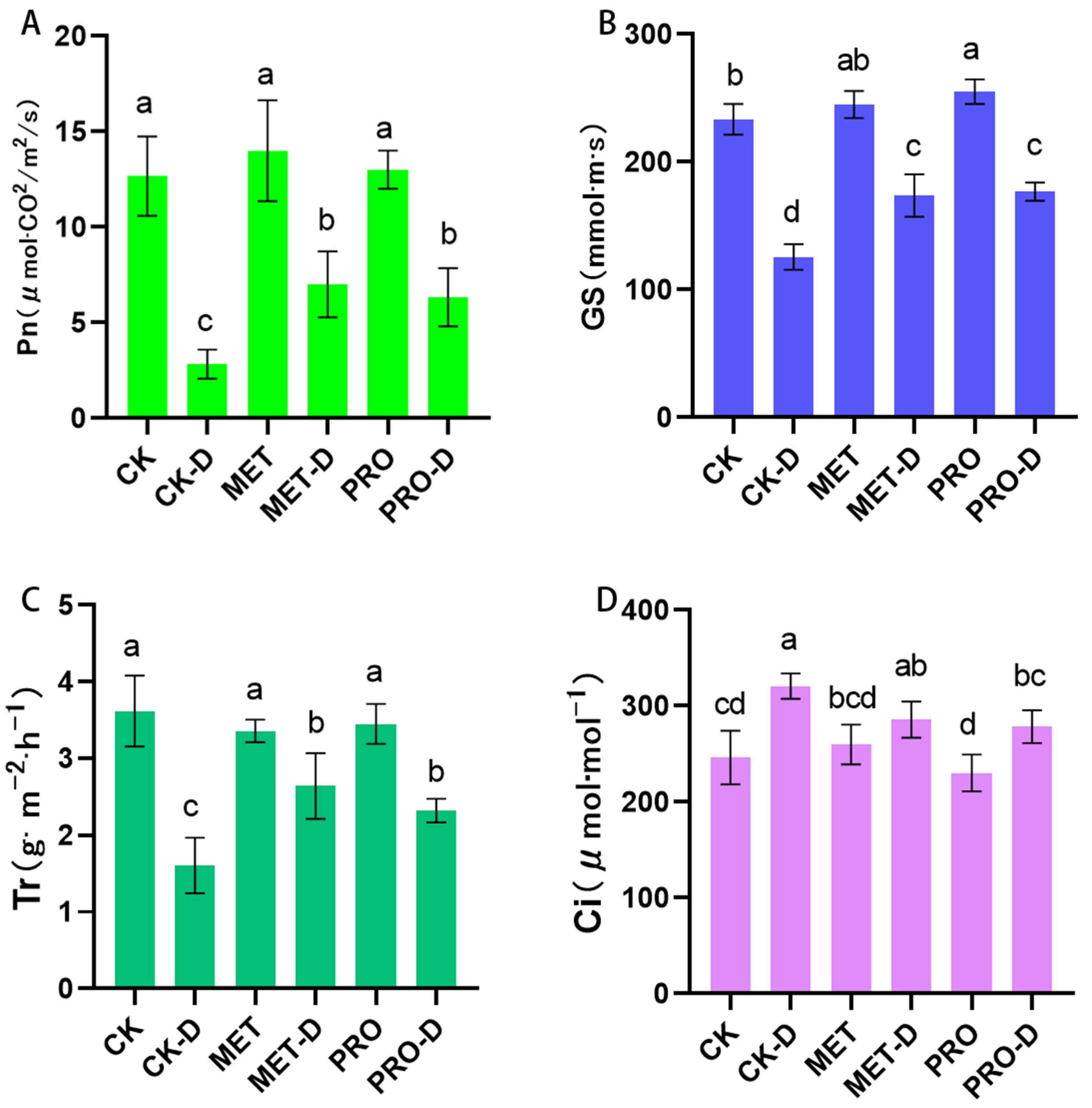
Figure 4.
Stomatal opening degree of leaves under drought stress after amino acid application by exogenous aid.
Figure 4.
Stomatal opening degree of leaves under drought stress after amino acid application by exogenous aid.

Figure 5.
Effects of Met and Pro on (A)SPAD, (B)chlorophyll a content (C) chlorophyll a content and (D) carotenoid content under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 5.
Effects of Met and Pro on (A)SPAD, (B)chlorophyll a content (C) chlorophyll a content and (D) carotenoid content under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
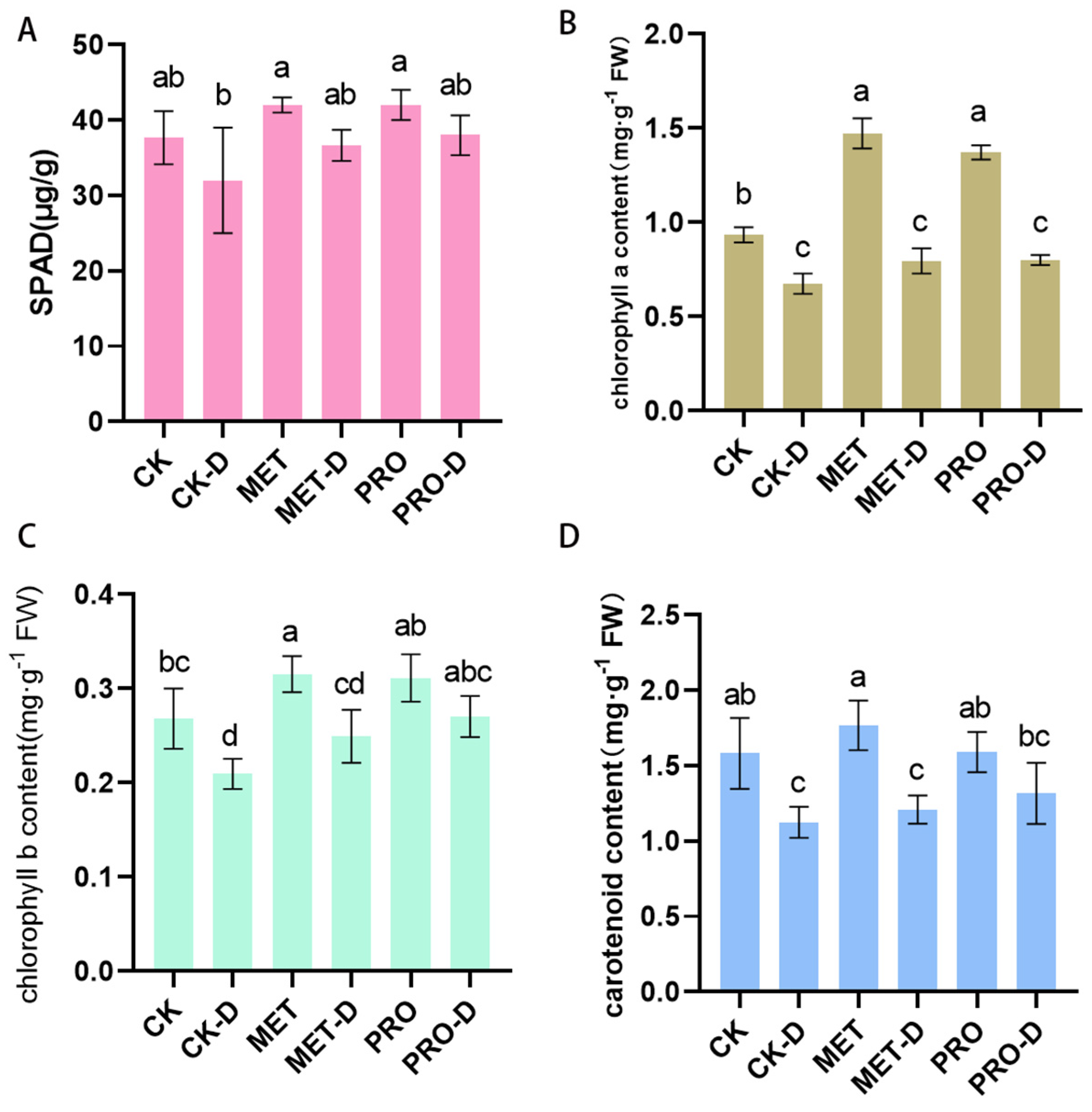
Figure 6.
Changes of chlorophyll fluorescence parameters of peach seeds under drought conditions. (A) Fm; (B) Fo; (C) Fv/Fm; (D) qP; (E) FPSII. Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 6.
Changes of chlorophyll fluorescence parameters of peach seeds under drought conditions. (A) Fm; (B) Fo; (C) Fv/Fm; (D) qP; (E) FPSII. Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
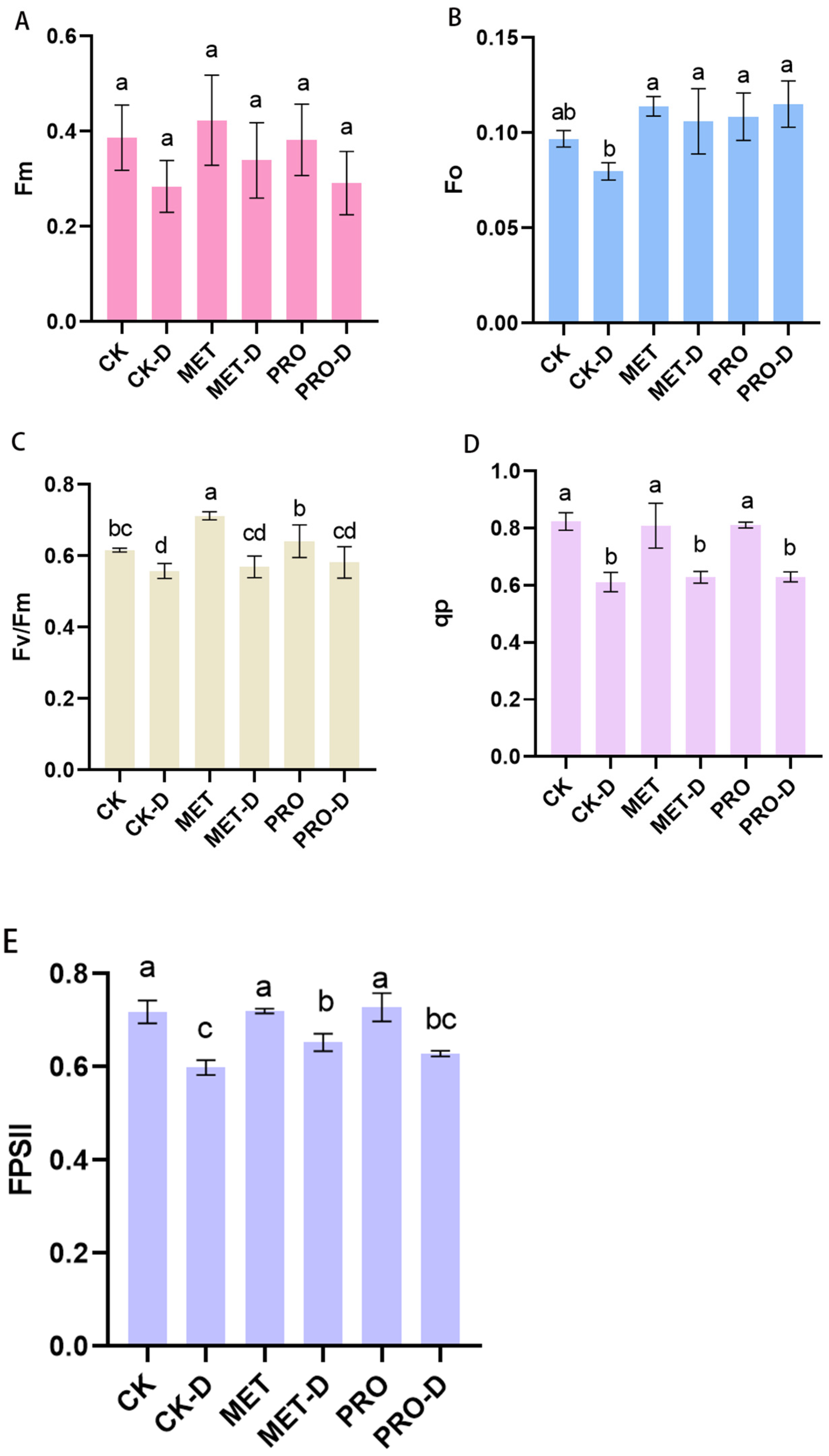
Figure 7.
Effects of Met and Pro on (A) soluble sugar content, (B) free amino acid. content, (C) proline content and (D) glutathione content of peach aphid under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 7.
Effects of Met and Pro on (A) soluble sugar content, (B) free amino acid. content, (C) proline content and (D) glutathione content of peach aphid under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
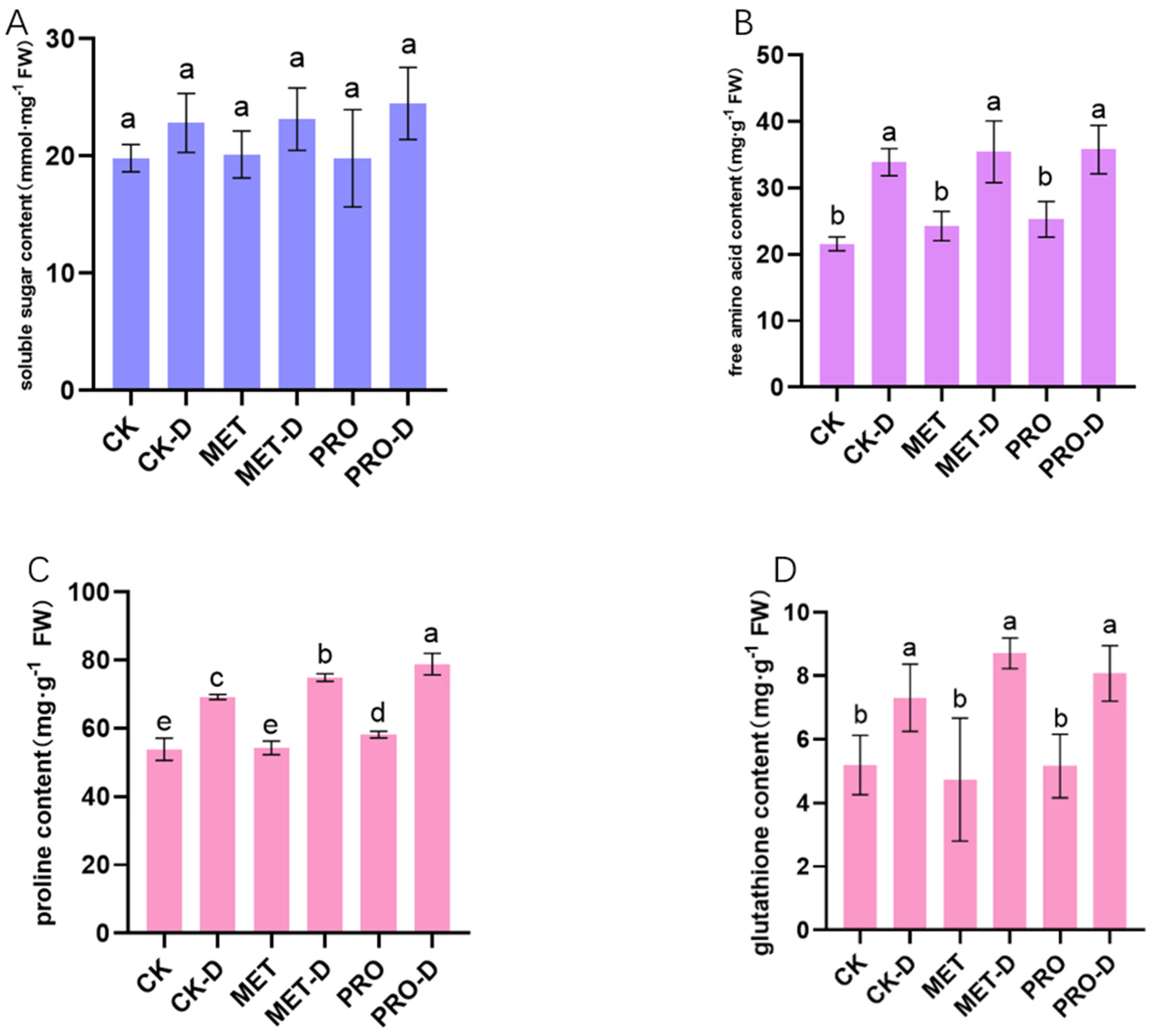
Figure 8.
Effects of exogenous amino acids on active oxygen content and antioxidant enzyme activity in leavesThe data of (A) O2-, (B) H2O2 production, (C) MAD, (D)CAT, (E)POD and (F) SOD content were mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 8.
Effects of exogenous amino acids on active oxygen content and antioxidant enzyme activity in leavesThe data of (A) O2-, (B) H2O2 production, (C) MAD, (D)CAT, (E)POD and (F) SOD content were mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
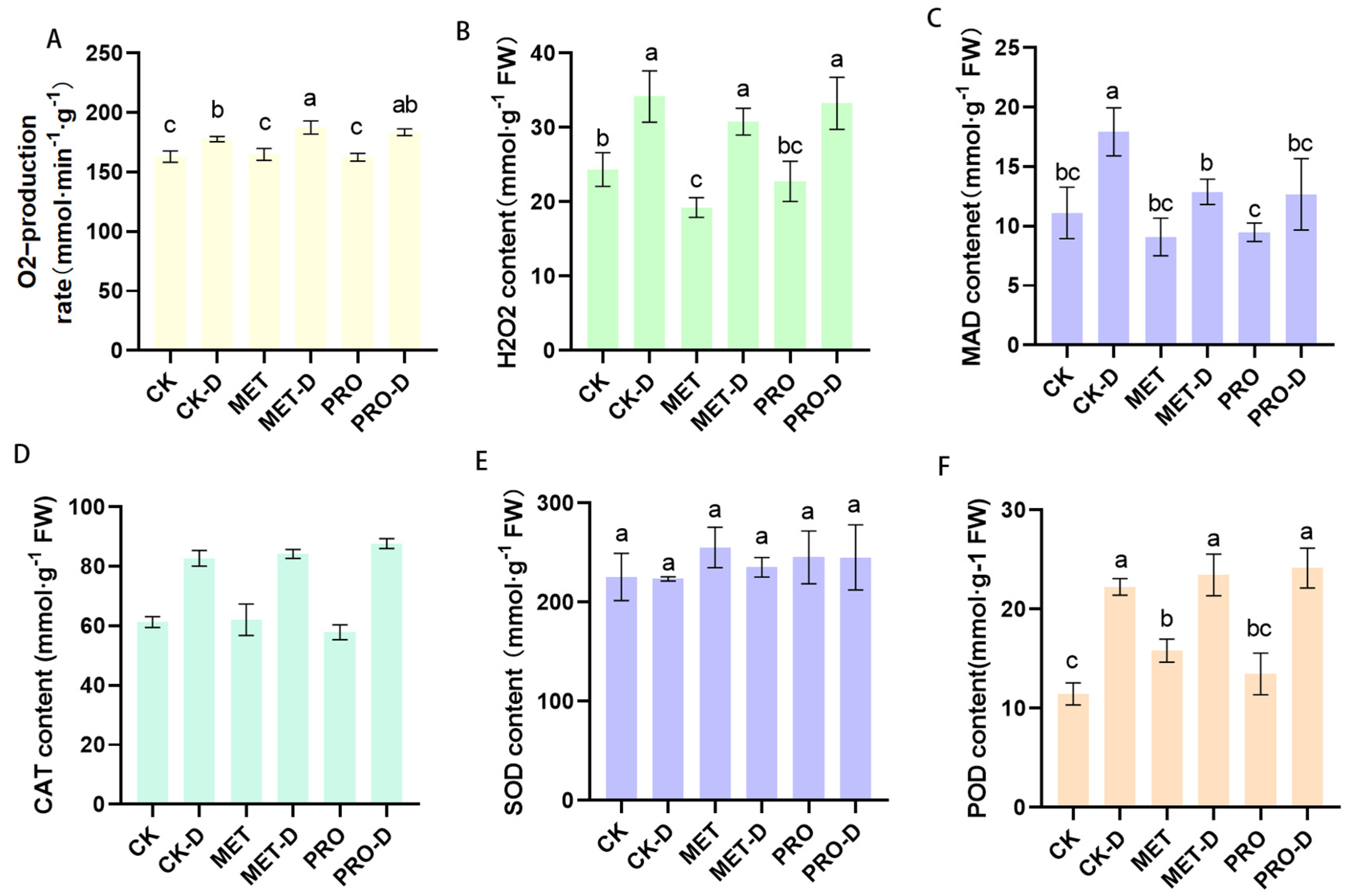
Figure 9.
Effects of Met and Pro on root (A) root activity (B) root configuration in peach under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Figure 9.
Effects of Met and Pro on root (A) root activity (B) root configuration in peach under drought stress. Data are mean ± standard deviation (n = 3). Duncan's test and ANOVA were used. The difference was significant (P < 0.05) are indicated by different letters.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



