
Submitted:
03 November 2023
Posted:
03 November 2023
You are already at the latest version
Abstract
Previous studies detail that the different blood groups are associate to incidence of oxidative stress-related diseases such as certain carcinomas. Bioactive compounds represent an alternative to prevent this oxidative-stress. The aim of this study was to elucidate the impact of blood groups on the erythroprotective potential of Fucoxanthin, β-Carotene, gallic acid, quercetin, and ascorbic acid as therapeutic agents against oxidative-stress. The impact of ABO blood groups on the erythroprotective potential was evaluate by the elucidated antioxidant capacity, blood biocompatibility, blood susceptibility and erythroprotective potential (membrane stabilization, in vitro photostability and antihemolytic activity). All antioxidant tested exhibited a highest potent antioxidant capacity and presented the ability to inhibit ROO•-induced oxidative-stress without compromising the cell membrane, providing an erythroprotective effect dependent on the blood group. Effect that increased in the presence of antigen A. This result could be key to the prevention of carcinomas associated with antigen A, such as breast and skin cancer. These results revealed a probable relationship between the different erythrocyte antigens with the erythroprotective potential, highlighting the importance of bio-targeted drugs for groups mostly susceptible to certain chronic-degenerative pathologies. These compounds could be applied as additive, nutraceutical or encapsulated to improve its bioaccessibility.
Keywords:
Blood groups
; Antioxidant
; erythroprotective potential
; oxidative-stress
1. Introduction
Reactive oxygen species (ROS), such as superoxide anion (O2•-), hydroxyl radical (•OH), and peroxyl radical (ROO•), are highly reactive unstable by-products of cellular metabolism, originally produced in mitochondria (1). Free radicals are generally produced through electron transfer reactions in cells with or without enzymatic intervention. These reactions can be mediated by metal ions in transition, e.g., the •OH radical is generated in the presence of H2O2 with copper (Cu+2) or Iron (Fe+2) ions (2). In adequate concentrations, reactive oxygen species are essential for cellular and immunological functions (3). The respiratory discharge of phagocytes activated by bacterial stimulation is directed to produce oxygen metabolites. These free radicals are intended to degrade phagocytosed parasitic bacteria (2). The bactericidal capacity of free radicals is very efficient; nevertheless, a considerable increase can not only damage the bacteria, as well as the environment in which the phagocytes act, resulting in damage to the body's cells. Uncontrolled respiratory discharge is associated with chronic inflammation that can stimulated neoplastic processes since free radicals accumulate in cells and damage molecules such as DNA, lipids, and proteins (4).
The involvement of uncontrolled respiratory discharges stimulates the degradation of biomolecules, this action is associated with non-communicable diseases such as cancer. Chronic-degenerative diseases caused 74% of deaths around the world in 2019 with more than 40,805 million deaths recorded. Therefore, the intervention of the health and research sector for the prevention of neoplasia’s associated with cellular oxidative stress is urgent. Antioxidants such as carotenoids, phenols, flavonoids, and vitamins represent an alternative to reduce oxidative stress (5). These biomolecules maintain a low redox state in cells through mechanisms of direct action on reactive species. The free radical scavenging capacity of antioxidants is related to Sequential Proton Loss Electron Transfer (SPLET), Single Electron Transfer (SET) and Hydrogen Atom Transfer (HAT) mechanisms (6,5). The antiradical activity of carotenoids is mainly due to the highly unsaturated chromophore and the functional groups in the case of xanthophylls. β-Carotene specifically presents a SET mechanism, due to its chemical nature incapable of donating protons. While Fucoxanthin has the ability to transfer both hydrogen atoms and electrons through its double bonds and the -OH and epoxy functional groups (Müller, Fröhlich, & Böhm, 2011). The free radical scavenging process of phenols and flavonoids is linked to successive deprotonations in their aromatic rings (6; 7). For some phenolic acids, the thermodynamically preferred deprotonation sites are the 4′-OH, 5-OH, and 7-OH groups, a mechanism presented by SPLET compared to HAT or SET (6, 8). Vitamin C (L-ascorbic acid) is an important reducing agent, capable of donating electrons and protons to receptor molecules such as free radicals. In relation to the reducing potential of antioxidants, they play a very important role in inhibiting oxidation and cytoprotective effect (neuroprotective, hepatoprotective and erythroprotective effect), preventing the development of chronic diseases related to oxidative stress, such as neurological(9), hemolytic (10, 11, 12) and carcinogenic processes (11,13).
Some carcinogenic processes are associated with different ABO phenotypes presence in the cell and tissues. Recent epidemiological research has reported the association between the risk of contracting non-communicable and communicable chronic diseases with the different ABO and Rh blood groups of human erythrocytes. Blood group A is associated with the incidence of different types of cancer (breast, gastrointestinal, liver, lung, pancreas, stomach, and non-melanoma skin cancer) and virally transmitted diseases (COVID-19) (14). In our research, blood susceptibility studies were carried out on ultraviolet radiation-induced oxidative stress in order to promote research on the role of antigen A and B of skin epithelial cells. It is expected that the results of the study will help us understand the possible effect of ultraviolet radiation (UVR) metastatic potential, pathological development, and prevention in subjects at higher risk of developing non-melanoma skin cancer associated with blood groups (15). It is known that UVR causes damage to genetic material, classifying it as an important factor for the development of skin cancer, mainly non-melanoma, and squamous cell carcinoma of the skin. Previously, a possible association between blood groups and the risk of skin cancer induced by prolonged exposure to ultraviolet radiation has been demonstrated (15). To date, studies suggest focusing on finding photoprotective compounds for groups A RhD-ve, AB RhD+ve and O (RhD not specified) that inhibit oxidative damage induced by UV radiation. Since a higher incidence of cancer has been reported in these blood groups (15). UVR induces peroxyl and alkoxyl radicals that react with the lipids and proteins of the erythrocyte membrane. Respiratory discharge AAPH-free radical initiator stimulated could be the key to elucidating the blood susceptibility of different blood phenotypes to chemical oxidation (16).
Recent research suggests that microalgae extract act as important erythroprotective agents, reducing oxidative stress depending on the surface antigen of the erythrocytes (10, 11, 12). Fucoxanthin and β-Carotene are among the most important and abundant marine carotenoids in algae and microalgae since they contribute more than 10% of the total estimated production of carotenoids in the marine environment. These pigments have generated great interest in the pharmaceutical-food industry due to their various health-beneficial properties, promoting pharmacological effects related to oxidative stress (13,17,18). Previous studies have shown that phenolic extracts from beans(19), chickpea (20) and sorghum (21) present high antioxidant activity with erythroprotective properties (22). However, the impact of ABO and RhD blood groups on the evaluation of the erythroprotective potential of fucoxanthin, β-carotene, gallic acid, quercetin and ascorbic acid as therapeutic agents against oxidative stress has not been reported. The use of human erythrocytes as a cellular model of ABO antigens could contribute to the development of functional foods designed for people with a specific blood type, preventing chronic-degenerative diseases that affect a more susceptible group.
2. Materials and Methods
2.1. Reagents
All chemical reagents such as Fucoxanthin (FXN), β-Carotene (β-Car), Gallic Acid (GA), Quercetin (QUE), Ascorbic Acid (AA), dimethyl sulfoxide (DMSO), DPPH (1,1-diphenyl-2-picrylhydrazyl), ABTS [2,2'-azinobis (3-ethylbenzothiazolin)-6-sulfonic acid], sodium acetate buffer, FeCl3 (ferric chloride), TPTZ (2,4,6-tripyridyl-s-triazine), hydrochloric acid (HCl), Triton X-100, AAPH [2,2′-Azobis (2-methylpropionamidine) dihydrochloride] and PBS (phosphate buffer saline) were purchased from Sigma-Aldrich Co. (USA). All other chemicals and solvents used for this study are of the highest commercial quality.
2.2. Biological Material
All methodologies that employed human red blood cell (RBC) were performed following the Mexican (NOM-253-SSA1-2012) and international (FDA: CFR - Code of Federal Regulations Title 21, part 640 Additional Standards for human blood and blood product, Support. B Red blood cells, Sec. 640.14 Testing the blood [21CFR640.14]) regulations. The membrane of RBCs with different ABO and RhD blood types were generously donate by the clinical analysis laboratory of the University of Guadalajara. The clinical laboratory is accredited by ISO-IEC 17,025 (NMX-EC-17025) and ISO 15,189 elaborated by technical committee ISO/TC 212 (Clinical Laboratory Testing and In vitro Diagnostic Systems) taking as reference ISO/ IEC 17,025 and ISO 9001. The human erythrocytes were collected from healthy adult volunteers (25 to 40 years old) that contained approximately 4.7 to 6.1 x106 cells/μL. Previously, the information the procedure was provided obtaining each one’s informed consent. The venipuncture technique was applied to collect human RBCs using a sterile vial with anticoagulant (EDTA). For these studies, the RBC samples were processed immediately after extraction for further experimental analysis. The purpose of these studies is to use RBCs as a membrane model to assess the impact of the ABO and RhD blood groups on erythroprotective potential of FXN, therefore, the object of study does not focus on volunteers (11, 12).
2.3. Preparation of Samples
FXN, β-Car, GA, QUE, and AA were dissolved previously in DMSO. Once the samples were solubilized in DMSO, aliquots were taken to prepare the samples at different concentrations (0.5-10 µM) in a DMSO/PBS (1:9) solution. The purpose of preparing the solutions in DMSO/PBS is to prevent erythrocyte hemolysis (Chisté et al., 2014). After a detailed search, the literature reports that these compound concentrations in blood plasma range from 0.1 to 0.9 µM.
2.4. Determination of Antioxidant Capacity
The antioxidant capacity was determined in FXN, β-Car, GA, QUE, and AA using three recognized methods. The 2,2-diphenyl-1-picrylhydrazyl (DPPH•) radical scavenging assay was performed following the procedure outlined by Brand-Williams et al. (23) The 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS•+) radical scavenging assay was conducted according to the method described by Re et al. (24) The ferric reducing antioxidant power (FRAP) assay was carried out as reported by Benzie and Strain (25). The results were expressed as mmol of μmol TE (Trolox equivalents)/g for each antioxidant compound.
2.5. Erythroprotective Potential
2.5.1. Blood Biocompatibility
Blood biocompatibility of FXN, β-Car, GA, QUE, and AA were confirmed by direct hemolysis assay (11) on erythrocytes of the ABO and RhD blood system. RBCs samples were collected by venipuncture in EDTA tubes as described in the biological material section. A suspension of 10% erythrocytes was prepare through three washed times with PBS (phosphate-buffered saline; 0.15 M; pH 7.4) removing the total plasma by centrifugation (2000 x g for 10 min) and recovered globular package. A quantity of 100 μL of erythrocytes suspension (10%) + 150 μL of each sample (v/v) at different concentration (0.5 to 100 µM) + 100 μL of PBS were incubated at 37 ºC. The erythrocytes were exposed for 6 hours to the samples to evaluate direct hemolysis with measured at 2 hours intervals. After concluding the incubation, each sample was centrifuged after adding 1 mL of PBS. A 300 µL volume of supernatant was measured at 540 nm in a 96-well microplate reader per triplicate. PBS buffer and Triton X-100 1% were used as negative and positive controls, respectively. Results were expressed as percentage of hemolysis and calculated using the following equation (1):
2.5.2. Membrane Stabilization Assay
Heat-induced hemolysis and hypotonicity-induced hemolysis assays were used to evaluate membrane stabilization (%) of all tested compounds and diclofenac sodium (DS) as a standard drug on erythrocyte membrane with different antigen types. Heat-induced hemolysis assay carried out according to the methodology by Agarwal et al. (26) with slight modification. The reaction mixture contained 150 μL of erythrocytes suspension (2%) + 150 μL of each sample (10 µM) was incubated in a water bath at 55 °C for 30 min. After time of incubation, 1 mL of PBS was added to each sample vial and centrifuged at 2000 x g for 10 min. Subsequently, the supernatant was recovered; where, 300 µL was taken to a microplate of 96-well and read at an absorbance of 540 nm. An amount of 300 µL of erythrocytes suspension were used as positive control in the same controlled condition. The membrane stabilization (%) it was reported by following equation (2):
For the hypotonicity-induced hemolysis assay, the methodology of Agarwal et al. (26) it was employed with some modification. The reaction mixture was carried out used 50 μL of RBC suspension (2%) + 100 μL of PBS + 100 μL of each sample (10 µM) + 200 μL of hyposaline solution in a water bath at 37 °C for 30 min. After incubation is complete, 850 μL of PBS was added to each sample vial and centrifuged at 2000 x g for 10 min. As a negative control was used 50 μL of RBC suspension (2 %) + 400 μL PBS and as positive control was used 50 μL of RBC suspension (2 %) + 200 μL PBS + 200 μL hyposaline solution. The membrane stabilization (%) was calculated using Equation 2.
2.5.3. Blood Susceptibility Test against Oxidative Stress
Blood susceptibility was determined on all erythrocyte blood phenotypes. The cell oxidative damage was induced by ultraviolet radiation (UVR) and chemically by the free radical generator 2,2′-azobis-[2-methylpropionam idine] (AAPH). Photohemolysis was induced using the Petri dish monolayer technique; where, a volume of 5 mL of each erythrocyte suspension (1%) was placed in the UV-A (315-395 nm) and UV-B (280-315 nm) chamber under controlled temperature (18 ± 1 °C) and different exposure times (60 and 120 min) (27,28). After irradiation, the RBC suspension was centrifuged at 2000 x g for 10 min. Two milliliters of supernatant were taken and mixed with 2 mL of Drabkin's solution (KCN, 0.05 g; K3Fe(CN)6 0.2 g in distilled water). After 10 minutes of rest, 300 µL were measured at 540 nm in a microplate reader (Hetherington and Johnson, 1984). In addition to this, blood susceptibility was also analyzed by AAPH-induced hemolysis. This molecule was used as a free-radical initiator to induce hemolysis on human erythrocytes. To carry out this part, we mixed 100 μL of RBC (2%) + 100 μL of PBS (0.15 M; pH 7.4) + 100 μL of AAPH (40 mM at pH 7.4). After the incubation period for three hours, 1 mL of PBS was added to each sample and centrifuged for 10 min at 2000 x g. One milliliters of supernatant were taken and mixed with 1 mL of Drabkin's solution. The reaction was allowed to stand for at 10 min under dark room condition. Later, 300 µL were measured at 540 nm in a microplate reader (Hetherington and Johnson, 1984). Blood susceptibility results are reported as released hemoglobin concentration (g/dL) calculated by the following equation (3):
2.5.4. In Vitro Photostability Studies
Ultraviolet-Visible (UV-Vis) Spectrophotometric Analysis
The photostability of the FXN and β-Car were evaluated by exposing the compound directly to UVR for 150 min under a flat horizontal array of two UV light-emitting tubes with an intensity of 0.85 mW/cm2 at a distance of 10 cm from the lamps. The changes in the structure were observed using the UV-Vis spectroscopic method (29).
Photoprotector Assay
The photoprotective effect of FXN, β-Car, GA, QUE, and AA against photooxidation induced by UV-A and UV-B radiation was evaluated on human erythrocytes with different blood groups, using the monolayer technique in a petri dish (11,30). One milliliter of blood (1%) was poured into a petri dish; then, 1 mL of physiological solution and 1 mL of each bioactive compound was added (in different petri dishes). Samples were subjected to preincubation at 37 ºC for 30 min. Subsequently, the petri dishes with the mixtures were placed inside the UVR chambers to evaluate the photoprotective effect. As a negative control, 1 mL of erythrocyte suspension (1%) + 2 mL of PBS without exposure to UVR was used. As a positive control, 1 mL of erythrocyte suspension (1%) + 2 mL of PBS (positive control) exposed to UVR was used. The positive control and the samples were exposed to different times (0, 30, 60, 120 and 150 min) of irradiation. After UVR exposure, controls and samples were centrifuged (2000 x g at 4°C for 10 min). Subsequently, 300 µL of supernatant was read at 540 nm in a 96-well microplate reader. Results were reported as photohemolysis inhibition (%) calculated by the following equation (4):
where A+ve control= Absorbance UVR-induced hemolysis. A-ve control = Absorbance without hemolysis. Asample = optical density of the photohemolysis inhibition by each treatment.
Finally, to observe RBC´s cellular membrane changes induced by UVR, an optical microscopy (100x Eclipse FN1 microscope) was used. For this study, the supernatant samples were performed immediately after reading. Approximately 50 μL of fresh plasma is added to the globular package (from inhibition-hemolysis test) and carefully mixed to avoid mechanical damage to the membranes. A blood suspension drop was spread over the slide creating a thin layer of RBC. To visualize the structure of erythrocyte membrane was necessary stain the RBC by wright method (12). The micrographs were observed with a 100x magnification and presented whit a scale bars of 5 µm to compare the RBC size and the images was capture used the software NIS-Elements F.
2.5.5. Antihemolytic Activity Assay
The AAPH-induced hemolysis inhibition was determined by the Antihemolytic activity assay (31) on different blood group types. The AAPH molecule was used as a free-radical initiator to induce hemolysis on human erythrocytes. Three different reactions were prepared: as a sample reaction, an amount of 100 μL of RBC (2%) + 100 μL of antioxidant + 100 μL of AAPH (40 mM at pH 7.4) were mixed; as a negative control, an amount of 100 μL of RBC (2%) + 200 μL of PBS (0.15 M; pH 7.4) were mixed; as a positive control, an amount of 100 μL of RBC (2%) + 100 μL of PBS (0.15 M; pH 7.4) + AAPH (40 mM at pH 7.4) were mixed. All reactions were incubated for 3 hours at 37 ºC. After the incubation, 1 mL of PBS was added to each sample and centrifuged for 10 min at 2000 x g. A volume of 300 µL of supernatant was read at 540 nm on a 96-well microplate (Multiskan Go, Thermo Scientific, Waltham, MA, USA). The results were report as hemolysis inhibition (%) calculated by following equation (5):
where AAPH1= optical density of the hemolysis caused per radical AAPH. % HI = percentage of hemolysis inhibition. HS = optical density of the hemolysis inhibition by each treatment.
2.6. Statistical Analysis
For this study, all data were analyzed by the statistical program JMP software v16 for Mac and expressed as mean ± SD (standard deviation). One and two-way ANOVA was performed to observe interaction between different factors evaluated. Tukey test was applied at a p < 0.05 confidence interval. The study was carried out under controlled conditions with a minimum of three repetitions (n ≥ 3) for each analysis.
3. Results and Discussion
3.1. Antioxidant Capacity Assay
In order to determine the antioxidant properties of FXN, β-Car, GA, QUE, and AA, three in vitro assays were employed specifically FRAP, DPPH• and ABTS•+, which can be seen in Table 1. By the FRAP method, the GA showed the highest antioxidant activity (5970.45 ±230.07 µmol TE/g) than all bioactive compound tasted including fucoxanthin (2687.03 ±173.26). Meanwhile, AA presented a high antioxidant potential to inhibit ABTS•+ free radicals scavenging (3412.22 a ±21.94 µmol TE/g). In this case, the antioxidant capacity of FXN was below the AA with values of 2638.53 µmol TE/g. Similar results are presented with DPPH• method, showing that AA and QUE (4190.9 ±155.16 and 3980.61 ±222.58 µmol TE/g) have a most antioxidant activity that FXN (303.15 ±42.66). Despite the results described above, the FXN is considered as a potent antioxidant as the β-Car. Nevertheless, previous studies have shown that fucoxanthin is not very reactive on the DPPH• radical (Nomura, 1997; Takaharu 1997).
Some methodologies conducted to analyze the antioxidant capacity of commercial fucoxanthin. In this study, FRAP, ABTS•+ and DPPH• assays were used to corroborate the antioxidant properties and compared to standard compound (β-Car; carotene, GA; phenol, QUE; flavonoid, and AA; vitamin) with the purpose of relating the results with the erythroprotective potential. Fucoxanthin, have a system of conjugated double bonds (Figure 1) with the ability to transfer electrons to reactive molecules such as the ABTS•+ radical and reduce molecules in an oxidized state to a reduced state (Habeebullah, 2018). The diverse mechanisms to inhibit free radicals and reducing power of FXN are attributed to its unique chemical structure such as to their system of conjugated double bonds, allenic bond, acetyl, and hydroxyl groups, that provide the multi-potent health effects and being key to the antioxidant activity (32). Electron transfer is carrying out in place during the second step of the single electron transfer followed by proton transfer (SET-PT) and sequential proton loss electron transfer (SPLET) mechanisms. Both mechanisms are carried out in two steps, which are processes in which an electron (e-) is lost and a less reactive cation-radical is formed, while in the second step, the cation-radical is deprotonated to form the corresponding radical (5,6).
It is important to mention, the use of different methodology for measure antioxidant capacity are a very useful tool. However, ABTS•+ and DPPH• are synthetic radicals, while the AAPH molecule is a generator of free radicals, that being thermolabile, breaks down into alkoxyl and peroxyl radicals. These radicals directly impact cell membranes, inducing lipid peroxidation and proteolysis (16). Therefore, the results obtained in this section is very importance to explain the erythroprotective potential of FXN. In later studies, fucoxanthin could form an active part of a drug, a nutraceutical or added to a functional food. For this reason, it is necessary to study its erythroprotective potential (blood biocompatibility, membrane stabilization, photoprotector and antihemolytic effect).
3.2. Erythroprotective Potential
3.2.1. Blood Biocompatibility
FXN, β-Car, GA, QUE, and AA are very important bioactive compound that promote essential health benefit. All compounds exhibit important antioxidant properties that inhibit cellular oxidation and premature aging induced by exposure to free radicals. Therefore, determining the blood biocompatibility of these compounds could be the key to future research in food and health.
The blood biocompatibility of FXN decreases at higher concentrations and prolonged exposure times on ABO and RhD blood groups (Figure 1). In general, FXN presented values around ≈ 97 to 100% of blood biocompatibility in the blood groups RhD+ve during three exposure times (Figure 1A). The most notable differences between blood groups with RhD+ve were observed at concentrations of 10 µM, specifically at the sixth hour of exposure, group O RhD+ve decreased significantly (p<0.05) with values of 87.20 %. This same trend occurred in RhD-ve blood groups (Figure 1B). Fucoxanthin is more biocompatible with blood groups A and O RhD-ve maintained values above 90%. Fucoxanthin significantly (p<0.05) affects to the blood groups AB and B RhD-ve. The decrease in blood biocompatibility of FXN on group AB RhD-ve begins at a concentration of 0.5 µM (4 h) with values of 83.50% to 42.70% at 10 µM (6 h). While percentage of blood biocompatibility of FXN decrease at high concentration (10 µM) on blood groups B RhD-ve with values of 83.75, 73.17, and 54.19 at exposure time of 2, 4, and 6 h, respectively.
A similar behavior was observed in the blood biocompatibility values of β-Car on erythrocytes with different blood groups in Figure 2. Figure 2A shows a decrease in biocompatibility with a concentration of 10 µM of β-Car at the sixth hour of exposure on RhD+ve erythrocytes. The values oscillate between 91 to 88% (A>AB>B>O) of biocompatibility, values considered innocuous(Gong et al., 2016). β-Car significantly affected (p<0.05) groups B and AB RhD-ve (Figure 2B), being directly affected by the concentration and exposure times. Figure 2B shows a significant decrease in the AB RhD-ve group at the sixth hour at all concentrations and the lowest biocompatibility is observed drastically decreased at 10 µM at 6 h (19.06%). Meanwhile, the decrease in group B RhD-ve is not so pronounced. This indicates that erythrocytes are more susceptible to hemolysis at 10 µM of β-Car after 2 h of exposure, decreasing from 81 to 66% biocompatibility.
Unlike the blood biocompatibility shown by FXN and β-Car, GA fluctuated from the start of the study (Figure 3). The blood groups non-B RhD+ve present the highest biocompatibility during the six hours of exposition at three concentration tasted. At the same time as the concentration of gallic acid increase, the percentage of blood biocompatibility decreases, significantly affecting at group B RhD+ve from 4 hour at concentration of 1 µM and 10 µM (83 at 81%, respectively) (Figure 3A). However, the blood biocompatibility of GA in the RhD-ve groups (Figure 3B) was different from the behavior in the RhD+ve groups (Figure 3A). GA showed higher biocompatibility values in non-O RhD-ve groups, being less toxic. Unlike the groups with the D antigen present in the membrane, the O RhD-ve group was more susceptible to the presence of gallic acid (82.46 %). Derived from this, the B RhD+ve and O RhD-ve groups were the most susceptible to hemolysis due to the presence of gallic acid. Probably, the presence or absence of the D antigen is important for the blood biocompatibility of bioactive compounds. In the case of blood biocompatibility of quercetin, the results are shown in Figure 4. The results indicate that the blood biocompatibility of Quercetin depends on the concentration and the exposure time. This trend was observed on RhD+ve erythrocytes (Figure 4A). Throughout the study, the non-B RhD+ve groups presented the highest values for each concentration (94-100% for 0.5 µM; 81-91% for 1 µM and 66-71% for 10 µM). In this case, the group most susceptible to hemolysis is B RhD+ve with the greatest decrease in biocompatibility after the fourth hour at 10 µM. On the other hand, non-O RhD-ve blood groups (Figure 4B) presented values of 92 to 100% at concentrations of 0.5 µM of QUE. Values fluctuate from 1 to 10 µM after four hours of exposure in these blood groups. However, it was observed that the O RhD-ve group is the most susceptible to oxidation due to the pro-oxidant actions of Quercetin, decreasing its biocompatibility up to 53%.
Figure 5 shows the blood biocompatibility of ascorbic acid on the different erythrocyte phenotypes. The effect of ascorbic acid on RhD+ve red blood cells was different from the previously mentioned compounds. Ascorbic acid is considered innocuous and biocompatible on the four phenotypes of the RhD+ve blood groups at the concentrations and exposure time tested. The values oscillated between 92-100% biocompatibility (Figure 5A). In contrast, the effects of ascorbic acid concentration and exposure time on RhD-ve blood phenotypes were different (Figure 5B). Group B RhD-ve was more susceptible to concentrations of 1 and 10 µM of AA, decreasing up to 50% of its blood biocompatibility. While the non-B RhD-ve groups maintained around 82 to 94% biocompatibility under the same conditions. Ascorbic acid can protect against oxidative damage caused by oxidative stress in cells. Studies by Chuyen et al. (33) have confirmed the damage by direct exposure of ascorbic acid on human T lymphocytes at concentrations of 1 mM. At this concentration, AA generates 85-161 mM H2O2 and induces programmed cell death. While in our study, exposure to low concentrations (<10 µM) of ascorbic acid does not affect the cellular integrity of RhD+ve red blood cells. However, AA exposure on RhD-ve red blood cells, specifically B RhD-ve, compromises cell integrity at the same concentrations. Despite the fact that the literature indicates the non-toxicity of AA on human lymphocytes at high concentrations, it does affect other types of cells according to their blood type.
Studies report that the use of FXN for human consumption could be safe, since it does not present toxicity either mutagenicity at low doses in animal models. Fucoxanthin levels in human plasma range from 0.1 to 0.9 µM (3435,36). However, these concentrations may not be sufficient in blood plasma to fully exert the biological power of this molecule. Probably, future research will design FXN carrier vehicles that control its release, increasing its absorption, bioaccessibility, and bioavailability in the gastrointestinal tract. This would lead to an increase in FXN the blood. Therefore, more studies are needed to evaluate its blood biocompatibility at higher concentration ranges. In our research, FXN concentration ranges from 0.5 to 10 µM were evaluated at different exposure times in order to study its biocompatibility on erythrocytes with different blood groups. The FXN presented acceptable blood biocompatibility (>95%) at concentrations tested of 0.5, 1 and 10 µM, specifically for non-AB and B RhD-ve blood groups. Less biocompatible B RhD-ve and AB RhD-ve erythrocytes were found at concentrations higher than those found in plasma. These results could be useful to understand the blood biocompatibility of FXN when incorporated into nano-liposomes vehicles as an oral administration route to improve and increase its biocompatibility and bioavailability. Like fucoxanthin, β-Car is a very important antioxidant pigment within the carotenoid family. Therefore, it is necessary to carry out toxicological tests for subsequent applications in the food industry. In several studies, the toxicological results made to β-Car do not exhibit mutagenicity or embryotoxicity. Toxicological tests carried out through the Ames test, mouse bone marrow cell micronucleus test and embryotoxicity in rats and rabbits (37,38). However, the impact of blood groups associated with blood biocompatibility of carotenoids such as FXN and β-Car has not been reported. Therefore, our study is one of the first to document the blood biocompatibility of β-Car on erythrocytes with different surface antigens of the ABO and RhD system. Like the previous carotenoids, GA is part of a series of antioxidants responsible for reducing oxidized compounds.
In our study, gallic acid was evaluated at 0.5, 1 µM and 10 µM and not at 0.6 µM. Therefore, its toxicity on groups B RhD+ve and O RhD-ve could be due to its pro-oxidant effects at high concentrations. Probably, the administration of high doses of GA could cause adverse health effects. The benefits provided by quercetin to human health maintain to the researchers interested in this flavonoid (39). The available literature states that dietary intake does not produce adverse health effects. QUE is a flavonoid used commonly as an antioxidant that provide cellular protection again oxidative damage. This characteristic reducing cell death induced by free radical (40). However, the use of quercetin becomes challenging due to its relative toxicity and possible low biocompatibility. Recently studies showed that the quercetin possess the ability to modify membrane penetrate lipidic bilayers (41,42). On the contrary, studies suggest that ascorbic acid has no cytotoxic effect at concentrations of 60 µM (43). Ascorbic acid has a protective effect on human erythrocytes against oxidative damage (75% inhibition of hemolysis), stabilizes the membrane by interacting with tocopheroxyl radicals. Unfortunately, studies have been observed that contradict the safety of ascorbic acid. The purpose of the cytotoxic study is to clarify and confirm the safety of FXN consumption prior to its incorporation into food, a toxic effect associated with different blood groups. The results obtained in our study indicate that blood biocompatibility decreases depending on the bioactive compound evaluated, exposure time and finally the blood group studied. The predominant blood biocompatibility occurred mainly in group A regardless of the RhD. Where the most affected groups were blood group B RhD+ve and O RhD-ve. The increase in antioxidant concentrations promotes an increase in their pro-oxidant potential, acting as oxidizing agents and inducers of cell death.
3.2.2. Membrane Stabilization Assay
The membrane stabilization assay was used as part of the evaluation of the erythroprotective potential of fucoxanthin on ABO and RhD blood groups and evaluated by Heat-induced hemolysis and Hypotonicity-induced hemolysis assay. The erythrocytes were exposed to different hemolysis conditions, thermal and osmotic disruption to determine the percentage of membrane stabilization conferred by FXN and its impact on the different blood groups (Figure 6).
Heat-induced hemolysis assay was carried out to evaluate the membrane stabilization (%) of FXN, four standard compound and diclofenac sodium (DS) as a standard for membrane stabilization (Figure 6A). Fucoxanthin, β-Car, and QUE were more effective in stabilizing the membrane of AB RhD+ve erythrocytes (67.34 ±2.65, 68.82 ±2.43, and 56.05 ±1.07 %, respectively) inducing an erythroprotective effect against heat-induced oxidative damage. Gallic acid provided a protective effect on groups A and B RhD+ve (61-62%) than on AB and O RhD+ve (57-55%). Meanwhile, ascorbic acid mostly stabilized non-O RhD +ve blood groups (55-57%). Despite the fact that diclofenac sodium is a potent inflammatory agent capable of stabilizing the lysosomal membrane, the erythroprotective effect was the lowest among all the compounds tested. Resulting in the O RhD+ve group (44%) with the highest membrane stabilization values than the non-O RhD+ve blood groups (31-40%). Figure 6B shows the membrane stabilization (%) on ABO RhD-ve blood groups. The increase in membrane stabilization in ABO RhD-ve blood types is evident. The antioxidant compounds FXN, β-Car, GA, QUE, and AA had a significantly higher protective effect against heat-induced oxidative damage in blood groups B and AB RhD-ve with values of 55-98%. The erythroprotective effect of diclofenac sodium was diminished compared to antioxidants. The values varied between 42-44% in the non-AB RhD-ve groups.
A hypotonicity-induced hemolysis assay was carried out to evaluate the membrane stabilization (%) properties of the FXN, β-Car, GA, QUE, AA, and DS on ABO blood groups (Figure 7). The Figure 7A show that the membrane stabilization (%) depends on both the blood groups and the bioactive compounds. FXN, β-Car, GA, and QUE stabilized the erythrocyte membrane efficiently in O RhD+ve groups (97-100%). The biological activity of FXN decreased in the AB RhD+ve group (61 %). A notable decrease was observed in group A RhD+ve with GA (41%). The erythroprotective effect of AA increased in the AB RhD+ve group (91%). Finally, the DS drug was effective in the AB and O RhD+ve groups with approximate values of 81%. The ability of FXN and β-Car to stabilize the membrane was found to be significantly higher in the O RhD-ve group (100%), while in the non-O RhD-ve groups values of 63-69% were observed for FXN and 84-92 % for β-Car (Figure 7B), significantly higher in the O RhD-ve group (100%), while in the non-O RhD-ve groups values of 63-69% were observed for FXN and 63-69%. for beta carotene. In contrast, gallic acid did not provide potent stabilizing activity, resulting in a significantly lower value of α in the O RhD-ve group (27.78%). However, its protective effect stabilized the membranes in non-O RhD-ve groups (27.78%) was high, with values around 93 and 100%. The percentage of membrane stabilization was similar for QUE, AA, and DS in all RhD-ve blood groups. The minimum values presented by these compounds were 84 % and the maximum 98 %. Therefore, FXN, β-Car, and GA were the bioactive compounds that varied the most in their protective activity according to the blood group evaluated. In general, according to the results obtained, the evaluated compounds presented a higher percentage of membrane stabilization in the O RhD+ve and O RhD-ve groups, with the exception of gallic acid.
FXN as a bioactive compound is emerging as a powerful therapeutic drug or food additive used against a wide range of chronic-degenerative pathologies associated with the recurrent appearance of free radicals (36,44). Red blood cell membranes serve as membrane models, where its ABO and RhD antigens play a very important role. Epidemiological research dictates that blood groups are related to the occurrence of various pathologies. Therefore, using membranes with different antigens within the ABO and RhD system is extremely important to find drugs and/or nutraceuticals capable of preventing diseases associated with blood groups (45,46). Therefore, plasma membranes have the function of target cells used to measure the inhibition of indicial lipid peroxidation by various factors (heat, hypotonicity and/or ultraviolet radiation). Establishing the interaction between FXN and cell membranes is of great interest (46). In previous investigations, through absorption and fluorescence spectroscopy, studies have been carried out on the effects of various flavonoids (e.g., quercetin, chrysin, morin, among others) on goat erythrocyte membranes (42,43). Membrane exploration with different functional surface antigens serves as a more realistic system to evaluate the action of new drugs or nutraceuticals that could increase therapeutic action and counteract pathologies associated with blood groups such as cancer (47,48). In some studies it has been demonstrated that the binding of flavonoids to erythrocyte membranes significantly inhibits lipid peroxidation, improving their integrity against osmotic disruption. In addition, they express that the antioxidant and antihemolytic activity of the bioactive compounds are related to the stabilization of the erythrocyte membrane. This indicate that the flavonoid binding site could be interacting very close to tryptophan residues in transmembrane proteins. This union plays a very important role in stabilizing membranes and inhibiting oxidation, providing important antioxidant potential. Membrane integrity is closely related to the antihemolytic activity of flavonoids. These studies are really interesting, since they demonstrate the importance of the incorporation of antioxidants for the prevention of osmotic disruption and inhibition of lipid peroxidation. However, in our study, human erythrocytes with different blood groups in the ABO and RhD system were used, which gives us a wide range of results. These indicate the impact of blood groups to evaluate membrane stability in the face of various factors that produce cell lysis. Therefore, the evaluation of the antihemolytic activity is widely justified for its study as part of the erythroprotective potential of FXN.
3.2.3. Blood Susceptibility Test Against Oxidative Stress
Since the discovery of the ABO blood group system, interest has remained in its potential involvement in chronic-degenerative diseases such as cancer and infectious diseases such as COVID-19 (SARS-CoV-2) (49). The difference in the expression of the ABO blood group polymorphic antigens can increase or decrease the susceptibility of the host to different pathologies and infections, due to their diverse functions in cell biology. Free radicals are produced during cell metabolism, as well as by the exposure or presence of various environmental or dietary factors (6,50). Oxidative damage induced by free radicals plays a very important role in premature aging and in many pathologies and degenerative disorders, mainly cancer. Antioxidants have been shown to reduce and inhibit oxidative stress and cell damage; in addition to reducing the chances of acquiring cancer (51,52). One of the aims of this research is focused on blood susceptibility test against ultraviolet radiation-induced oxidative stress and AAPH-induced oxidative stress. This information will be useful to clarify the erythroprotective potential of fucoxanthin. Therefore, erythrocytes were used as a model cell system to delineate the effects of ultraviolet radiation-induced oxidative stress and AAPH-induced oxidative stress on human cells with different surface antigens of the ABO system (6,13,16).
To evaluate the effects of ultraviolet radiation-induced oxidative stress on human erythrocyte was used monolayer Petri dish technique. It was observed that oxidative denaturation of the erythrocyte membrane by UVR leads to the release of hemoglobin (Hb) that depended on three factors: type of erythrocyte antigen, type of UV radiation and exposure time (Figure 8). It was established that the groups most susceptible to hemolysis induced by UV-B radiation are the B RhD+ve and B RhD-ve groups with values of 5.8-6.3 [Hb] g/dL at 120 min of exposure. Values significantly higher than in non-B blood groups, while the B RhD+ve and AB RhD-ve groups were more susceptible to hemolysis against UV-A radiation at 120 minutes of exposure with values ranging between 1.2 and 1.4 [Hb] g/dL; where, susceptibility is reported as the amount of hemoglobin released by exposure to ultraviolet radiation. At present, the incidence, morbidity, and mortality rates of skin cancer are increasing, representing a major public health problem (Narayanan et al., 2010). An exhaustive search has revealed that studies point to ultraviolet radiation as the main etiological agent in the development of skin cancer (15). Ultraviolet radiation (UVR) causes damage to genetic material, causing DNA mutations, which later lead to the development of neoplasms, in this case skin cancer. Previous studies have examined the association between ABO blood groups and skin cancer risk induced by prolonged UVR exposure (15). Specifically, UVR has been shown to be one of the main risk factors for developing non-melanoma skin cancer and squamous cell carcinoma. Studies to date suggest focusing on finding photoprotective compounds for groups A RhD-ve, AB RhD+ve and O (RhD not specified) that inhibit oxidative damage induced by ultraviolet radiation. It has been reported that there is a higher incidence of these types of cancers in populations with the aforementioned blood groups (15). By detecting in our study that the blood groups B RhD+ve and AB RhD-ve are susceptible to UV-A radiation and B RhD+ve and B RhD-ve are susceptible to UV-B radiation (UV-B being of higher risk) it could be considered that free radicals and the oxidative stress generated by UVR participate in a great way for the development of skin cancer in the risk groups mentioned above, being these groups more susceptible to oxidative damage. Our research is intended to stimulate the prevention of cancer induced by UVR in subjects at higher risk or more susceptible and promote basic research addressing this social problem. The photoprotective effect of the antioxidants studied in our research could help prevent the incidence of skin cancer for the risk groups most affected by UVR. Ultraviolet radiation generates free radicals by exposing tissues to high doses of radiation for a long time. Generating consequences that affect cellular integrity leading to the appearance of certain chronic pathologies such as skin cancer, tissue injuries and eye damage. However, ultraviolet radiation is not the only source of free radical formation; there are different pathways that generate oxidative stress. Food additives, metals in transition, or pesticides, induce the formation of free radicals that chemically oxidize lipids and proteins found in the cell membranes of tissues. In this step of the research, the AAPH molecule was used to induce lipid peroxidation and proteolysis on the different erythrocyte phenotypes to evaluate blood susceptibility (53,54).
It can be seen in Figure 9, to the AAPH molecul disturbs the integrity of the erythrocyte cell, an action that releases hemoglobin as a product of cellular oxidation and which also depends on the blood group. Blood susceptibility to AAPH was as follows: B RhD+ve; B RhD-ve; AB RhD+ve < A RhD+ve; To RhD-ve; O RhD+ve < B RhD-ve; AB RhD+ve (1.10-1.50 [Hb] g/dL < 1.75-2.05 [Hb] g/dL < 2.50-2.62 [Hb] g/dL, respectively). These last two groups being the most susceptible to oxidation induced by AAPH. The hemolytic oxidative effect promoted by the free radical initiator AAPH is the key to clarifying the blood susceptibility of different blood phenotypes to chemical oxidation. These results will promote the search for antioxidant compounds with greater potential to inhibit oxidative stress in those groups with phenotypes more susceptible to degradation. In this case, prevent lipid peroxidation and proteolysis of RhD-ve groups B; AB RhD+ve due to chemical agents that induce free radicals such as food additives, heavy metals, pesticides, food preservatives, among others. Thus, contributing to the prevention of chronic-degenerative pathologies associated with the blood groups of most susceptible groups.
Peroxyl and alkoxyl radicals react with the lipids and proteins of the erythrocyte membrane (16). The exact mechanisms of interaction between erythrocyte surface antigens and free radicals are unknown. However, it can be observed that the presence of these antigens contributes to reducing or increasing cellular oxidation. The increase in reactive oxygen species leads to the formation of holes in the erythrocyte membrane, causing the release of hemoglobin. Especially H2O2, which, being a product of physiological cellular metabolism during enzymatic and non-enzymatic reactions, triggers the oxidation of membrane components such as lipids and proteins. The chain reactions of ROS promote morphological changes on the cell membrane, compromising the structure and leading to eryptosis. Therefore, cytoprotective antioxidants could be a source of antioxidants capable of reducing oxidative stress associated with blood groups, highlighting their antihemolytic capacity.
3.2.4. In vitro Photostability Studies
In view of the increasing demand for oxidative stress suppressors, there is a general need for cytoprotective antioxidants that support the side effects of free radicals generated chemically and by exposure to sunlight (55,56). Prolonged exposure to sunlight has been linked to several types of skin damage, including sun injury, skin inflammation, photoimmunosuppression, photoaging, photocarcinogenesys, include irritation, inflammation, erythema, and photocarcinogenesys associated with UVR exposure. These pathologies occur through the formation of reactive oxygen species (ROS) (55,56). Fortunately, the skin has endogenous antioxidants that it uses as defense mechanisms to inhibit or delay the oxidative damage generated by UVR. Undoubtedly, this defense system decreases with chronic exposure to ultraviolet rays, therefore, a strategy to remedy this deficiency would be the use of antioxidant phytochemicals capable of delaying or inhibiting cell damage caused by free radicals, providing photoprotection to human skin (57,58,59). It has been observed that bioactive compounds such as flavonoids and carotenoids are capable of inhibiting cellular photo-oxidation. However, in the same way as endogenous antioxidants, exogenous antioxidants could be degraded by chronic UVR exposure. A probable hypothesis to reduce the development of pathologies associated with the effects of UVR is the application of oral, skin, or ocular antioxidants that eliminate reactive oxidants and modulate the cellular redox state. For this reason, part of our study focused on evaluating the photostability of fucoxanthin, blood susceptibility, and in vitro photoprotection efficacy. This study allows obtaining information on the use of antioxidants as photoprotective agents, inhibiting the effects of UVR on different erythrocyte blood groups as part of the erythroprotective potential (51,58,59).
Ultraviolet Spectrum of Fucoxanthin before UVR Exposure
Firstly, this assay aims to report on the effects of exposure to ultraviolet radiation on fucoxanthin to determine if there are changes in its structure that affect its photoprotective potential. A comprehensive review on photoprotective compounds has revealed that carotenoids are excellent antioxidants capable of inhibiting free radicals and oxidative stress induced by ultraviolet radiation. This quality highlights its potential use as photoprotective agents applicable in the food and pharmacological industries. However, it has not been reported whether the ultraviolet spectrum of Fucoxanthin changes due to exposure to UV-A and UV-B. Therefore, in this assay, fucoxanthin was exposed to UV-A (315-395 nm) and UV-B (280-315 nm) radiation for 120 min with an irradiation intensity of 0.85 mW/cm2. Furthermore, β-carotene was used as a standard of a known photoprotective agent. At the same time, the results were compared with the UV spectrum of both compounds without irradiation.
The spectral behavior of Fucoxanthin and β-carotene before and after UV irradiation is reported in Figure 10. The main objective of this assay was to determine its photoprotective effect despite the probable photoisomerization at extended periods of UV irradiation (60-64)Their profiles are in accordance with those observed directly in studies of Kumar et al. (64), Greul et al. (62) in non-irradiated samples (Figure 10A). From the spectra recorded, one can observe the differences in spectral profiles for the fucoxanthin and β-carotene after UV-A (315-395 nm) and UV-B (280-315 nm) irradiation for 120 min exposure. From the profiles obtained under UV radiation, one can deduce that the Fucoxanthin is less photostable than β-carotene after irradiation with UV-A and UV-B (Figure 10B and 10C). Fucoxanthin has some degree of photoinstability above β-carotene after UV-A and UV-B irradiation. This instability could be due to photoisomerization that occurs after exposure to photooxidative ultraviolet light. Probably, reactions are initiated that affect the structural integrity of Fucoxanthin, generating free radicals of this antioxidant or less biologically active isomers. Studies were found that described dehydrated product ions of protonated molecules for hydroxylated carotenoids (65,66). Therefore, these structural changes in Fucoxanthin focus directly on the hydroxylated terminal functional groups of the photoisomers and even the loss of one of the rings at the terminal ends. Furthermore, it directly affects the epoxy functional group, reactions that could trigger reactive isomers. However, it is necessary to delve deeper and carry out studies on the biological activity of the photoisomers formed by photooxidation. Once the photostability of Fucoxanthin was observed, its photoprotective effect against different erythrocyte blood groups was evaluated.
In Vitro Photoprotection Efficacy
Ultraviolet radiation (UVR) induced a substantial decrease in the integrity of the erythrocyte membrane of the eight blood phenotypes, measured through hemoglobin release (Figure 11). After preincubation with Fucoxanthin, the levels of photoprotection against UV-A radiation increased significantly on non-A RhD-ve blood groups in the first 60 min of exposure with values around 75-99% inhibition of photohemolysis (Figure 11A). However, after 120 minutes, photoprotection decreased in most blood groups (22-80%), except in groups A RhD+ve and B RhD+ve (90-95%). Undoubtedly, the groups most affected by UV-A radiation were groups A RhD-ve and B RhD-ve, presenting a greater susceptibility to cellular oxidation. Particularly interesting, those erythrocytes with the absence of antigen D showed an increase in their photooxidation, specifically in antigens A and B, being the phenotypes that provided the least photoprotection from fucoxanthin. Meanwhile, it can be seen from Figure 11B that the photoprotection of fucoxanthin was different against UV-B radiation. Here photooxidation was more evident after 30 minutes of exposure. Despite this, the inhibition of photohemolysis ranged between 60-97% at 30 and 60 min of exposure. In this exposure period, fucoxanthin provided greater photoprotection on blood groups A and B RhD+ve (94-98%). Unlike the results reported for UV-A radiation, UV-B radiation did not drastically affect group A RhD-ve, but rather group AB RhD+ve. At the end of the exposure period (120 min), photoprotection decreased drastically for group B RhD-ve compared to the previous two periods. In general, all groups decreased in this period, with a greater photoprotection present in the O RhD-ve group by Fucoxanthin (85.34 %).
The photoprotective effect exhibited by β-carotene against hemolysis induced by UV-A radiation (Figure 12A) presented similar values in the three exposure periods on the eight erythrocyte phenotypes. β-Carotene conferred a greater photoprotective effect on A RhD+ve erythrocytes compared to the remaining 7 phenotypes during the three periods. The values ranged between 95-100% inhibition of photohemolysis. While the AB RhD-ve group was the most affected by UV-A radiation despite the photoprotection provided by β-carotene. This means that erythrocytes with AB antigens with absent RhD probably present an antagonistic effect (17-38%). Presenting a significant decrease (p>0.05) with respect to the values presented for groups A and B RhD+ve, A and B RhD-ve, and AB RhD+ve. This same trend occurred in blood groups exposed to UV-B radiation (Figure 12B). The most notable difference is found under the 120-minute period where the value of the O RhD-ve group decreased to 18.54%.
Figure 13 also shows a notable variability in the photoprotective effect of gallic acid on the different erythrocyte phenotypes as well as the previous bioactive compounds. This variability can be observed in the affinity of gallic acid by the different phenotypes seen in Figure 13A. This trend indicates that as the exposure time increases, the photoprotective effect of gallic acid decreases. At a time of 120 minutes, erythrocyte hemolysis due to exposure to UV-A radiation is accentuated. Revealing a decrease in the protective effect in groups A RhD-ve, AB RhD+ve and O RhD-ve, with values ranging between 43-51%. Gallic acid showed greater effectiveness in inhibiting photooxidation induced by UV-B radiation (Figure 13B) on RhD+ve erythrocytes A and B, with notable photoprotective stability in all exposure periods (80-98%). However, free radicals and oxidative stress induced at 120 minutes caused cellular degradation, lipid peroxidation and proteolysis on the erythrocyte membrane of the AB and O RhD-ve groups (≈20%). This result indicates that gallic acid probably did not have such a strong affinity with these blood groups, since this decrease in the photoprotective effect was observed from the first 30 minutes of exposure.
The photoprotective efficacy of quercetin (Figure 14) showed a behavior dependent on blood groups. Which means that the inhibitory potential of quercetin on free radicals induced by UV-A and UV-B radiation varied depending on the erythrocyte surface antigen. Figure 13A shows the variation of the effect of quercetin on erythrocyte phenotypes with a very similar behavior for the three exposure periods. A greater response was obtained in groups A, B, and O, where antigen D is present in all three group types (90-98%). In Figure 14B, a greater photooxidation can be seen with UV-B radiation as opposed to UV-A radiation. Here, the intensity of ultraviolet radiation significantly decreased the photoprotective efficacy of quercetin in the three established time periods. In the first 60 minutes of exposure, quercetin provide the AB RhD+ve group an efficacy between 90 to 95%, which decreased after 120 minutes (58%). Photoprotective efficacy decreased below 60% for all erythrocyte phenotypes.
The results of the efficacy of ascorbic acid showed that blood groups B RhD-ve, AB RhD+ve, and AB RhD-ve were mostly susceptible to degradation by UV-A radiation (45 and 74%) in the three periods (Figure 15A). The oxidative stress present in these blood groups was evident, when compared to the remaining blood groups. Once again, groups A RhD+ve and B RhD+ve showed a better response to the protection provided by ascorbic acid. Not only this antioxidant, but also the previous antioxidants exhibited significant photoprotection against photooxidation induced by UV-A radiation on these same blood groups. The evaluation of the sensitivity to UV-B radiation of erythrocytes preincubated with ascorbic acid showed statistically significant differences between each blood group tested (Figure 15B). Ascorbic acid seemed to suppress the effects of UV-B radiation on the erythrocyte membrane of the AB RhD+ve group in the first and second periods (30 and 60 min) of exposure. However, at 120 min, the effects of radiation suppressed the photoprotection provided by ascorbic acid, reducing its effect on all blood groups.
Figure 16 shows the morphological changes of group A RhD+ve erythrocytes exposed to ultraviolet radiation and the photoprotective effect provided by the carotenoids tested. This phenotype was selected for the photoprotective efficacy of these antioxidants. Micrographs A, B, and C are red blood cells not exposed to UV radiation and are used as a control without hemolysis. These erythrocytes show a typical morphology of healthy human erythrocytes, with characteristics such as a well-defined biconcave central zone, measuring between 5 to 6.5 nm. Micrographs D and G are shown to describe the oxidative stress lethality generated by UV-A and UV-B radiation, respectively. These micrographs show erythrocytes exposed to UVR to generate oxidative stress and evaluate its effects on changes in their cellular structure, a model of photohemolysis control. In this case, radiation altered the membrane of the erythrocytes, inducing hemolysis. This alteration releases hemoglobin, a result that is interpreted as possible cell death or eryptosis in the particular case of erythrocytes. UVR wavelengths directly affect the erythrocyte membrane, causing lipid peroxidation and proteolysis. This action promotes the loss of the physical integrity of the membrane, resulting in a typical hemolytic activity, accompanied by release of hemoglobin. On some occasions the biconcave characteristic of the erythrocyte can be lost. Membrane oxidation is characterized by the presence of an intense purple and red color at the periphery of the biconcave disk of the erythrocyte (67-70). Additionally, two groups of micrographs are observed where the photoprotective effect of the carotenoids studied is expressed. Group of erythrocytes exposed to UV-A radiation with photoprotective antioxidants (Figure E and F). Group of erythrocytes exposed to UV-B radiation with photoprotective antioxidants (Figure H and I).
No significant morphological changes are observed in erythrocytes exposed to UV-A radiation in the presence of Fucoxanthin and β-carotene compared to the control (Figure 16 E and F). Photooxidation inhibited by erythroprotective carotenoids is evident by the preservation of the physical integrity and morphology typical of a healthy cell. In this case, the antioxidants tested largely counteracted all the alterations induced by UV-A rays described above. These same characteristics were observed in the erythrocyte sample exposed to UV-B radiation with β-Carotene (Figure 16 I) as a photoprotective agent, which maintains the cellular integrity of a healthy erythrocyte. In this case, it was observed that all the effects of photooxidation induced by UV-B rays are counteracted. However, Fucoxanthin partially prevented the damage to the erythrocyte membrane induced by UV-B rays (Figure 16 H). Therefore, photostability decreased, compromising the stability of erythrocytes. The morphological changes showed partial damage to the erythrocyte, without apparent damage to the integrity of the membrane as a consequence of a decrease in photoprotection. Further analysis is needed to confirm whether this damage could be significant and later contribute to possible eryptosis.
The evaluation of photoprotection with Fucoxanthin and β-Carotene can be observed in micrographs E, F (UV-A), H and I (UV-B), respectively. No significant morphological changes are observed in erythrocytes exposed to UV-A radiation in the presence of Fucoxanthin and β-Carotene. Photooxidation inhibited by erythroprotective antioxidants is evident by the preservation of the physical integrity and the typical morphology of a healthy cell. In this case, the antioxidants tested counteracted all of the above-described UVA-induced alterations to a significant extent. These same characteristics were observed in micrograph I with β-Carotene. In this case, it was observed that all the effects UV-B ray-induced-photooxidation is counteracted. However, fucoxanthin only partially prevented damage to the erythrocyte membrane induced by UV-B rays. Therefore, photostability decreased, compromising erythrocyte stability. The morphological changes made damage to the erythrocyte evident, causing damage to the integrity of the membrane, disruption and lipid oxidation that occurred as a consequence of a decrease in fucoxanthin photoprotection at 120 min with photoinhibition values of ≈70%. More analysis is needed to confirm that this damage could be significant and mitigate eryptosis. Ultraviolet radiation interacts with DNA and human cells in a very complex way. This interaction and the pathological effects have been the introduction for therapeutic advances in the formulation of treatments against eye and skin diseases induced by ultraviolet radiation. Currently, protective mechanisms against UV damage and bioactive photoprotective compounds are the key to reducing the processes that take place when human skin is exposed to sunlight (64). Through of cellular models, it has been possible to study the intervention of drugs and bioactive compounds that could be directly involved in the intervention of photooxidation and consequently cellular mutagenesis. Additionally, study the photoprotective effect, susceptibility, and the association of different blood phenotypes with photooxidation (30).
Sunlight is the main source of human exposure to ultraviolet radiation, which is classified into three main types: UV-A (315-395 nm), UV-B (280-315 nm) and UV-B (100-280 nm). Studies have revealed that overexposure without adequate sun protection has been associated with epidermal diseases. Ultraviolet radiation has adverse effects on the eyes and skin cells, causing sunburn, photodermatitis, premature aging, cataracts formation, mutagenesis, and skin cancer. The main biomolecules affected by UVR are proteins and DNA; this cellular damage stimulates the proliferation of neoplastic cells. There are two pathways of cellular damage mediated by UV radiation: (a) the direct absorption of radiation by cellular components, altering cellular chemistry and the formation of an excited state; and (b) the absorption of radiation by endogenous or exogenous cells that are excited to their triplet states, a mechanism called photosensitization (30,71,72).
Endogenous photosensitization is responsible for inducing reactive oxygen species by UVR, generating superoxide anion and singlet oxygen. Over time, through different mechanisms, it is possible to induce the synthesis of other reactive oxygen species, such as peroxyl radicals, hydrogen peroxide and hydroxyls. In this context, various studies associate the generation of free radicals induced by UVR with skin pathologies. Fortunately, the skin and the human body have an elaborate defense system of endogenous antioxidants capable of mitigating the effects of oxidative stress induced by chemical agents, cellular metabolism, inflammatory processes, and external agents such as UVR. However, excessive exposure to ultraviolet radiation can reduce the skin's antioxidant capacity, limiting its biological action. These antioxidant systems are limited after chronic exposure, reaching damaging levels of free radicals. In this case, it causes oxidative damage, skin lesions, inflammation in the affected area, immunosuppression, premature aging, and skin cancer. Therefore, the intervention of exogenous antioxidants is an interesting strategy to limit and inhibit photooxidation, providing a photoprotective potential to support the endogenous antioxidant system. This can be achieved through the intervention of exogenous antioxidants, such as carotenoids, phenols, flavonoids, and vitamins, expanding options for photoprotective products that could include supplementation with oral, topical and ocular antioxidants (73,74).
Exogenous antioxidants such as carotenoids are fundamental pieces for systemic photoprotection in humans. An exhaustive search indicates that carotenoids such as Astaxanthin, Fucoxanthin, β-Carotene, and lycopene have acted as sunscreens, reducing the risk of inducing pathologies related to oxidative stress induced by UVR (40,71-74). However, there is not enough scientific information about its properties when applied to the different blood phenotypes, which are not only found in erythrocytes, but also in skin cells and other cell lines. Fucoxanthin and β-Carotene have high antioxidant activity, capable of acting as a photoprotective agent on human cells. Despite presenting photoinstability, this carotenoid is considered a promising blood group-dependent photoprotective agent. Since it managed to effectively protect groups A RhD+ve, A RhD-ve (UV-A and UV-B). These results could contribute to generating targeted drugs, nutraceuticals, or functional foods for a population highly susceptible to developing conditions related to overexposure to UVR. The results are promising, since group A RhD-ve was the most susceptible to phototoxicity induced by UV-A and UV-B radiation. Napoleão-Tavares et al. (2020) demonstrated that Fucoxanthin has a photoprotective effect on skin cells exposed to UVA, UVB and visible radiation. The ability to inhibit the formation of reactive oxygen species induced by UVR on fibroblasts and keratinocytes reduces phototoxicity. When human dermal fibroblasts are exposed to moderate doses of UV-A radiation, an apoptotic process is triggered; where, the levels of reactive oxygen species and thiobarbituric acid compromise the integrity of the cell membrane (74,75). Furthermore, it has been reported that during the digestive process, enzymes metabolize fucoxanthin mainly to Fucoxanthinol. This isomer has antimutagenic, anti-inflammatory, antineoplastic and photoprotective properties. The modulating effect of fucoxanthinol intervenes in the arrest of the cell cycle of cancer cells. This property could prevent skin cancer induced by UVR in oral drugs (74,76,77)
In addition to carotenoids, antioxidants such as phenols, flavonoids and vitamins participate in the prevention of skin cancer associated with ultraviolet radiation. Polyphenols such as gallic acid are ideal chemopreventive agents to counteract skin disorders. Particularly, recent advances suggest that gallic acid can suppress, slow, and reverse the process of skin carcinogenesis (28,42,77). The photoprotective potential of these polyphenols has reduced oxidative stress, DNA damage and suppressed the immune response. Additionally, they can nullify the various biochemical processes mediated by ultraviolet radiation at in vitro and in vivo levels. Polyphenols can act as photoprotective agents since they have the ability to absorb radiation in the length spectrum of UV-A and UV-B radiation. However, this characteristic has only been detected in those polyphenols that are yellow, red, or purple. Since gallic acid is a transparent compound, the photoprotective mechanism is probably different (Nichols and Katiyar, 2010). In our study, the photoprotective effect of gallic acid depended on the blood groups tested; where, the most protected group was A RhD+ve (UV-A); A RhD+ve, B RhD+ve, and AB RhD-ve (UV-B). Therefore, gallic acid can be used as an inhibitor of ultraviolet radiation-induced oxidative damage in people with type B RhD+ve, since they are the blood type most susceptible to UVR according to the results obtained.
In view of the increasing demand for oxidative stress suppressors, there is a general need for cytoprotective antioxidants that support the side effects of free radicals generated chemically and by exposure to sunlight. Prolonged exposure to sunlight has been linked to several types of skin damage, including sun injury, skin inflammation, photoimmunosuppression, photoaging, and photocarcinogenesys. With increasing awareness of these potentially harmful effects, there is a need to search for safe and effective photoprotective antioxidant compounds. The development of skin pathologies associated with ultraviolet radiation (UVR) is due to the formation of reactive oxygen species (ROS) and the deterioration of the skin antioxidant system. A probable hypothesis to reduce the development of pathologies associated with the effects of UVR is the application of oral, skin, or ocular antioxidants that eliminate reactive oxidants and modulate the cellular redox state (30,64).
Unlike carotenoids and polyphenols, ascorbic acid (Vitamin C) is not considered a sunscreen due to its chemical nature, because vitamin C does not absorb light in the UV-A or UV-B spectrum. However, vitamin C has a potential antioxidant that is praised for its anti-aging benefits. For this reason, vitamin C has been shown in studies to protect against oxidative damage induced by UV radiation by acting as a sunscreen. There are proteins that transport vitamin C to the site of inflammation that has come into contact with ultraviolet light in harmful amounts. As a consequence, vitamin C increases in keratinocytes, resulting in the need to absorb a higher concentration of vitamin C for adequate protection. In cultured keratinocytes exposed to UVR, vitamin C addition reduces oxidative damage to DNA and lipid peroxidation, prevents lysosomal degradation, inhibiting the release of pro-inflammatory cytokines and protecting against cell death by apoptosis. While in animal models exposed to UV radiation, the addition of vitamin C to the diet of rodents through oral supplementation has reduced the size of skin tumors and the development of dermal neoplastic diseases. However, studies suggest that the combination of vitamin C, vitamin E, RRR-alpha-tocopherol and β-Carotene effectively reduce the damage induced by UV radiation. The synergistic effect would expand the properties of vitamin C as a photoprotective agent in oral supplementation (79). Our study used erythrocytes as a cellular model for the evaluation of the photoprotective effect of vitamin C on the different blood phenotypes exposed to UV-A and UV-B radiation. The addition of vitamin C inhibited the effects of UV-A in groups A RhD+ve and B RhD+ve, preserving cellular integrity without visible oxidative damage, conferring adequate photoprotection. This effect decreased when erythrocytes were exposed to UV-B light, radiation considered more harmful, in this case the photoprotective effect decreased in all blood phenotypes. The AB RhD-ve blood group stood out among the others, being the group mostly protected by vitamin C. The photoprotective potential of vitamin C depended on the surface antigen on the erythrocytes, more studies would have to be carried out in combination with photoprotective antioxidants to evaluate its effect on the groups most susceptible to degradation in the presence of UVR (Anne-Katrin et al., 2002; Chen et al., 2012).
3.2.5. Antihemolytic Activity Assay
The antioxidant mechanisms of Fucoxanthin, β-Carotene, gallic acid, quercetin, and ascorbic acid, mainly HAT, SET-PT, and SPLET, inhibit AAPH-induced oxidative hemolysis. A conjugated double bonds system, benzene rings, and hydroxyl groups contribute to the ability to transfer electrons and protons to reactive molecules such as the peroxyl and alkoxyl radicals. Antioxidants act as reducing agents against the radicals generated by the AAPH compound before contact with the erythrocyte membrane, through the interaction of glycoproteins and membrane proteins, depending on the blood phenotype. The antioxidant-protein interaction could form a protective complex against membrane oxidation promoted by AAPH-produced free radical. Another proposed inhibition pathway could result in an increase in the surface area-volume ratio, resulting in membrane expansion or cell contraction, modifying the interaction with membrane anchoring proteins and providing protection to the cell against hemolytic oxidation (5,6,16).
The antihemolytic response of antioxidants directly depended on blood groups with slight differences between antioxidant compounds. The antioxidants tested exerted an erythroprotective effect on RhD+ve blood erythrocytes above 80% inhibition of hemolysis (Table 2). Carotenoids inhibited total hemolysis in the A RhD+ve phenotype (≈100%). Gallic acid, quercetin and ascorbic acid generated a similar erythroprotective potential, around 97-100% inhibition of hemolysis on the AB RhD+ve and AB RhD+ve phenotypes. However, this antihemolytic effect exerted by antioxidants decreased in the O RhD+ve phenotype until it decreased by 80%. This could denote that erythrocytes with the absence of antigens A and B with the presence of antigen D affect the erythroprotective potential of these antioxidants. On the other hand, it is possible that the presence of antigens A and B (RhD+ve) increases its antihemolytic activity. The inhibitory effect of erythroprotective antioxidants decreased on non-O RhD-ve groups, even below 80% in some cases (Table 3). Increasing this effect in the RhD-ve group O using Fucoxanthin and β-carotene as erythroprotective agents. The cellular integrity of the B RhD-ve phenotype was preserved above 80% using quercetin and ascorbic acid as erythroprotective agents. In this case, the results are more heterogeneous than for the phenotypes with RhD-ve.
In an exhaustive review of the scientific literature, the involvement of blood groups with the severity of certain chronic-degenerative and infectious pathologies has been found. Research suggests that blood groups are associated with the antioxidant potential of biomolecules (11,13). These studies indicate that the erythroprotective potential of biomolecules such as proteins, phycobiliproteins and carotenoids is associated with the presence of blood groups. The protective effect varies depending on the surface antigens. The results are relevant, since this blood system is associated with inflammatory responses without pathological stimulus, that is, systemic inflammation that could lead to the development of a neoplastic process. The susceptibility of phenotype A to certain pathologies is recurrent. This characteristic could occur in people suffering from different types of cancer (maternal cancer) and infectious diseases (HIV infection, hepatitis B virus and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) related to this blood group. Recently, signs of association have been detected between the 9q34.2 locus of SARS-CoV-2 (COVID-19) with the ABO blood group locus (14). A specific analysis showed a greater risk of acquiring this infection in blood group A than in non-A groups, causing a greater risk of respiratory failure. Therefore, a possible participation of ABO blood groups in oxidative stress induced by the AAPH molecule is confirmed.
For this reason, our study has focused on determining the erythroprotective potential of Fucoxanthin, β-Carotene, gallic acid, quercetin, and ascorbic acid, representing biomolecules that belong to different families of antioxidants (carotenes, xanthophylls, phenolic acids, flavonoids and vitamins) with the aim of preventing or reducing oxidative stress induced by different sources. The results indicate that the different antioxidants tested in our research could be potential candidates to provide protection, mainly to cells with antigen A on their membrane. In subsequent research, functional foods, nutraceuticals and even effective micro or nano transporters loaded with potentially cytoprotective antioxidants could be developed to reduce the incidence of pathologies related to blood groups.
4. Conclusions
All antioxidants tested exhibited potent antioxidant capacity in FRAP and ABTS. The results indicate that the erythroprotective antioxidant are suitable for use at the concentrations tested, since they did not show toxicity in blood biocompatibility tests. Therefore, the integrity of the erythrocyte membrane is not compromised, and antioxidants even confer an erythrocyte effect on the stability of the membrane against hemolysis induced by heat and hypotonicity, factors that could be induce the synthesis of free radicals that promote cellular oxidation. The impact of blood groups on the erythroprotective potential of the antioxidants FXN, β-Car, GA, QUE, and AA is evident, providing an erythroprotective effect dependent on blood group. Photoprotective effect and antihemolytic activity increased with the presence of antigen A, inhibiting uncontrolled respiratory discharges. These results could be key to preventing carcinomas associated with the A antigen, such as breast and skin cancer. Erythroprotective antioxidants stabilized redox state of the erythrocytes, stimulating the inhibition of lipid peroxidation and proteolysis, an action that could prevent chronic-degenerative pathologies associated with prolonged exposure to ultraviolet radiation or free radicals generated by chemical reactions. These compounds could be applied as additives, nutraceuticals or encapsulated to improve their bioaccessibility, highlighting the importance of developing bio-targeted drugs for groups mostly susceptible to certain chronic-degenerative pathologies related to cancer. Beyond the impact of blood groups with the erythroprotective potential of different antioxidants, the susceptibility of the eight blood phenotypes to oxidative stress is reported. It was found that group B is the most susceptible to degradation by free radicals induced by ultraviolet radiation, while groups AB RhD-ve and O RhD-ve to oxidative stress induced by AAPH. These results could help understand the mechanisms of association between blood groups with susceptibility to certain types of cancer.
Author Contributions
R.I.G.-V: conceptualization and investigation; M.A.R-G: project administration; C.L.D.-T.-S.: supervisor; L.Y.M-U and K.N.C-E: methodology and investigation; S.R-C and F.V. V-V: writing-review; MG-L and M.G.A.-N: formal analysis; RDI-G: data curation.
Funding
This study did not receive any specific funding agencies in the public, commercial, or not-for-profit sectors.
Institutional Review Board Statement
The study was carried out in accordance with the Declaration of Helsinki of 1975. The clinical laboratory is accredited by ISO-IEC 17.025 (NMX-EC-17025) and ISO 15.189 prepared by the technical committee ISO/TC 212 (Clinical Laboratory Testing and In vitro Diagnostic Systems) taking as reference the ISO/IEC 17.025 and ISO 9001 standards. Ethical approval: As per University procedure, approval of for the study was taken by the authors (CI 2023-47).
Informed Consent Statement
All subjects gave their informed consent for inclusion in the study prior to their participation.
Data Availability Statement
The original contributions data presented in this research are included in the article; further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors are pleased to acknowledge the Department of Research and Postgraduate in Food, Universidad de Sonora, and Department of Medical and Life Sciences, Cienega University Center (CUCIÉNEGA), Universidad de Guadalajara, for the facilities granted to provide resources and infrastructure. Ricardo Iván González-Vega acknowledges the Consejo Nacional de Ciencia y Tecnología (CONAHCyT, Mexico) for a posdoctoral scholarship.
References
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress, and the biology of ageing. Nature. 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants, and animals. Comparative Biochemistry and Physiology Part C. Toxicol. Pharmacol. 2010, 153(2), 175–190. [Google Scholar]
- Goiris, K.; Van Colen, W.; Wilches, I.; Tamariz, F.; De Cooman, L.; Muylaert, K. Impact of nutrient stress on antioxidant production in three species of microalgae. Algal Research. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Achar,A.;Myers,R.;Ghosh, C. Drug Delivery Challenges in Brain Disorders across the Blood–Brain Barrier: Novel Methods and Future Considerations for Improved Therapy. Biomedicines. 2021, 9, 1834. [CrossRef]
- Biela, M.; Rimarčík, J.; Senajová, E.; Kleinová, A.; Klein, E. Antioxidant action of deprotonated flavonoids: Thermodynamics of sequential proton-loss electron-transfer. [CrossRef]
- Xue, Y.; Liu, Y.; Luo, Q.; Wang, H.; Chen, R.; Liu, Y.; Li, Y. Antiradical activity and mechanism of coumarin-chalcone hybrids: theoretical insights. J. Phys. Chem. 2018, 122, 8520–8529. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-Z.; Chen, D.-F.; Deng, G.; Guo, R.; Lai, R.-C. The influence of C2=C3 double bond on the antiradical activity of flavonoid: different mechanisms analysis. Phytochem. 2019, 157, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zemlicka, L.; Fodran, P.; Lukes, V.; Vaganek, A.; Slovakova, M.; Stasko, A.; Dubaj, T.; Liptaj, T.; Karabín, M., Bírosova, L., Rapta, P. Physicochemical and biological properties of luteolin-7-O-β-d-glucoside (cynaroside) isolated from Anthriscus sylvestris (L.) Hoffm. Mon. 2014, 145, 1307–1318. [CrossRef]
- Alghazwi, M.; Smid, S.; Musgrave, I.: Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta toxicity and aggregation. Neurochem. Int. 2019, 124, 215-224. [CrossRef]
- González-Vega, R.I.; Cárdenas-López, J.C.; López-Elías, J.A.; Ruiz-Cruz, S.; Reyes-Díaz, A.; Perez-Perez, L.M.; Cinco-Moroyoqui, F.J.; Robles-Zepeda, R.E.; Borboa-Flores, J.; Del-Toro-Sánchez, C.L. Optimization of growing conditions for pigments production from microalga Navicula incerta using response surface methodology and its antioxidant capacity. Saudi J. Biol. Sci. 2021, 28; 1401–1416. [CrossRef]
- Ruiz-Cruz, S.; González-Vega, R.I.; Robles-Zepeda, R.E.; Reyes-Díaz, A.; López-Elías, J.A.; Alvarez-Ainza, M.L.; Cinco-Moroyoqui, F.J.; Moreno-Corral, R.A.; Wong-Corral, F.J.; Borboa-Flores, J.; et al. Association of Different ABO and Rh Blood Groups with the Erythroprotective Effect of Extracts from Navicula Incerta and Their Anti-Inflammatory and Antiproliferative Properties. Metabolites 2022, 12, 1203. [Google Scholar] [CrossRef] [PubMed]
- Peña-Medina, R.L.; Fimbres-Olivarría, D.; Enríquez-Ocaña, L.F.; Martínez-Córdova, L.R.; Del-Toro-Sánchez, C.L.; López-Elías, J.A.; González-Vega, R.I. Erythroprotective potential of phycobiliproteins extracted from Porphyridium cruentum. Metabolites. 2023, 13, 366. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Yan-jun, C.; Jia, Q.; Min-min, Z.; Tian-liang, Z.; Min, C.; Shun-mei, L.; Guangce, W. Fucoxanthin Exerts Cytoprotective Effects against Hydrogen Peroxide-induced Oxidative Damage in L02 Cells. BioMed Res. Int. 2018, x(x), 1–11. [Google Scholar] [CrossRef]
- Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl J Med. 2020, 1–13. [CrossRef]
- Celic, D.; Lipozencic, J.; Kolaric, B.; Ferencak, G.; Kanižaj Rajkovic, J.; Borlinic, T. Association between Blood Group and Nonmelanoma Skin Cancers (Basal Cell Carcinoma and Squamous Cell Carcinoma). Int. J. Environ. Res. Public Health. 2019, 16, 2267; [Google Scholar] [CrossRef]
- Zhang, J.; Hou, X.; Ahmad, H.; Zhang, H.; Zhang, L.; Wang, T. Assessment of free radicals scavenging activity of seven natural pigments and protective effects in AAPH-challenged chicken erythrocytes. Food Chem. 2014, 145, 57–75. [Google Scholar] [CrossRef]
- Rostamabadi, H.; Reza, Falsafi, S.; Mahdi, Jafari, S. Nanoencapsulation of carotenoids within lipid-based nanocarriers. JCR 2019, 298, 38–67. [Google Scholar] [CrossRef] [PubMed]
- González-Vega, R.I.; Del-Toro-Sánchez, C.L.; Moreno-Corral, R.A.; López-Elías, J.A.; Reyes-Díaz, A.; García-Lagunas, N.; Carvajal-Millán, E.; Fimbres-Olivarría, D. Sulfated polysaccharide-rich extract from Navicula incerta: physicochemical characteristics, antioxidant activity, and anti- hemolytic property. AIMS Bioengineering, 9(4), 364–382. [CrossRef]
- Rodríguez-Roque, M.J.; Del-Toro-Sánchez, C.L.; Chávez-Ayala, J.M.; González-Vega, R.I.; Pérez-Pérez, L.M.; Sánchez-Chávez, E.; Salas-Salazar, N.A.; Soto-Parra, J.M; Iturralde-García, R.D.; Flores-Córdova, M.A. Digestibility, Antioxidant and Anti-Inflammatory Activities of Pecan Nutshell (Carya illioinensis) Extracts. Journal of Renewable Materials, 2022. Impact Factor: 2.115. 2570-2580. [CrossRef]
- Perez-Perez, L.M.; Huerta-Ocampo, J.A.; Ruiz-Cruz, S.; Cinco-Moroyoqui, F.J.; Wong-Corral, F.J.; Rascón-Valenzuela, L.A.; Robles-García, M.A.; González-Vega, R.I; Rosas-Burgos, E.C.; Corella-Madueño, M.A.G.; et al. Evaluation of Quality, Antioxidant Capacity, and Digestibility of Chickpea (Cicer arietinum L. cv Blanoro) Stored under N2 and CO2 Atmospheres. Molecules, 2021. Impact Factor: 4.927. 26:2773. [CrossRef]
- Reyna-Reyna, L.Y.; Montaño-Leyva, B.; Valencia-Rivera, D.E.; Cinco-Moroyoqui, F.J.; González-Vega, R.I.; Bernal-Mercado, A.T.; Ballesteros-Monrrea, M.G.; Méndez-Encinas, M.A.; Del-Toro-Sánchez, C.L. Antioxidant, antibacterial, anti-inflammatory, and antiproliferative activity of sorghum lignin (Sorghum bicolor) treated with ultrasonic pulses. “Metabolites” 2023. [CrossRef]
- Alghazwi, M.; Smid, S.; Musgrave, I.: Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta toxicity and aggregation. Neurochem. Int. 2019, 124, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, H.; Shanmugam, V.K. Anti-inflammatory activity screening of Kalanchoe pinnata methanol extract T and its validation using a computational simulation approach. Inform. Med. Unlocked. 2019, 14, 6–14. [Google Scholar] [CrossRef]
- Takebayashi, J.; Chen, J.; Tai, A. A method for evaluation of antioxidant activity based on inhibition of free radical-induced erythrocyte hemolysis. Methods Mol. Biol. 2010, 594, 287–296. [Google Scholar]
- Rizvi, S.I.; Jha, R.; Pandey, K.B. Activation of erythrocyte plasma membrane redox system provides a useful method to evaluate anti-oxidant potential of plant polyphenols. Methods Mol. Biol. 2010, 594, 341–348. [Google Scholar]
- Dere, S.; Gűnes, T.; Sivaci, R. Spectrophotometric determination of chlorophyll - A, B and total carotenoid contents of some algae species using different solvents. Turkish J. Botany. 1998, 22, 13–17. [Google Scholar]
- Simioni, C.; Schmidt, E.; Felix, M.R.L.; Polo, L.K.; Rover, T.; Kreusch, M.; Pereira, D.T.; Chow, F.; Ramlov, F.; Maraschin, M.; Bouzon, Z.L. Effects of Ultraviolet Radiation (UVA+UVB) on Young Gametophytes of Gelidium floridanum: Growth Rate, Photosynthetic Pigments, Carotenoids, Photosynthetic Performance, and Ultrastructur. Photochem. and Photobiol. 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, K.L.; Ruiz-Cruz, S.; Cira-Chávez, L.A.; Gassos-Ortega, L.E.; Ornelas-Paz, J.J.; Del-Toro-Sánchez, C.L.; Márquez-Ríos, E.; López-Mata, M.A.; Rodríguez-Félix, F. Evaluation of Antioxidant Capacity, Protective Effect on Human Erythrocytes and Phenolic Compound Identification in Two Varieties of Plum Fruit (Spondias spp.) by UPLC-MS. Moelcyles. 2018, 23, 3200; [Google Scholar] [CrossRef]
- Lourenço-Lopes, C.; Fraga-Corral, M.; Jimenez-Lopez, C.; Carpena, M.; Pereira, A.G.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Biological action mechanisms of fucoxanthin extracted from algae for application in food and cosmetic industries. Trends Food Sci. Technol. 2021, 117, 163–181. [Google Scholar] [CrossRef]
- Chuyen, V.H.; Eun, J.-B. Marine carotenoids: bioactivities and potential benefits to human health. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Emi, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK-Ay mice. Arch. Biochem. Biophys. 2010, 504, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Jian-Ping, Y.; Chou-Fei, W.; Jiang-Hai, W. Fucoxanthin, a Marine Carotenoid Present in Brown Seaweeds and Diatoms: Metabolism and Bioactivities Relevant to Human Health. Mar. Drugs. 2011, 9:1806-1828. [CrossRef]
- Peng, C.H.; Chang, C.H.; Peng, R.Y.; Chyau, C.C. Improved membrane transport of astaxanthin by liposomal encapsulation. Eur. J. Pharm. Biopharm. 2010, 75, 154–161. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, N.; Liao, L.; Yu, K.; Shu, X.-O.; Zheng, W.; Yuan, J.-M.; Koh, W.-P.; Qiao, Y.-L.; Fan, J.-H.; et al. ABO Genotypes and the Risk of Esophageal and Gastric Cancers. BMC Cancer 2021, 21, 589. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotecnol. Adv. 2016, 34:1396-1412. [CrossRef]
- Zheng, Y.-Z.; Chen, D.-F.; Deng, G.; Guo, R.; Lai, R.-C. The influence of C2=C3 double bond on the antiradical activity of flavonoid: different mechanisms analysis. Phytochem. 2019, 157, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Masek, A., Latos, M., Piotrowska, M., Zaborski, M. 2018. The potential of quercetin as an effective natural antioxidant and indicator for packaging materials. Food Packaging and Shelf Life, 16, 51-58. [CrossRef]
- Miltonprabu, S., Tomczyk, M., Skalicka-Woźniak, K., Rastrelli, L., Daglia, M., NabavI, S.F., Alavian, S.M., Nabavi, S.M. 2017. Hepatoprotective effect of quercetin: From chemistry to medicine. Food and Chemical Toxicology, 108, 365-374. [CrossRef]
- Mu, Y., Fu, Y., Li, J., Yu, X., Li, Y., Wang, Y., Wu, X., Zhang, K., Kong, M., Feng, C., Chen, X. 2019. Multifunctional quercetin conjugated chitosan nano-micelles with P-gp inhibition and permeation enhancement of anticancer drug. Carbohydrate Polymers, 203, 10-18. [CrossRef]
- Chisté, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids are Effective Inhibitors of in vitro Hemolysis of Human Erythrocytes, as Determined by a Practical and Optimized Cellular Antioxidant Assay. J. Food Sci. 2014, 79, 9. [Google Scholar] [CrossRef]
- Miyashita, K.; Maeda, H.; Okada, T.; Abe, M.; Hosokawa, M. Anti-obesity, and anti-diabetic effects of allenic carotenoid, fucoxanthin. Agro. Food Ind. Hi Tech 2010, 21, 24–27. [Google Scholar]
- Niki, E.; Komuro, E.; Takahashi, M.; Urano, S.; Ito, E.; Terao, K. Oxidative hemolysis of erythrocytes and its inhibition by free radical scavengers. J. Biol.Chem. 1988, 263, 19809–19814. [Google Scholar] [CrossRef] [PubMed]
- Parentini, I.; Bergamini, E.; Cecchi, L.; Cavallini, G.; Donati, A.; Maccheroni, M.; Tamburini, I.; Gori, Z. The effect of carbon tetrachloride and ultraviolet radiation on dolichol levels in liver cells isolated from 3- and 24-month-old male Sprague–Dawley rats. Biogerontology 2003, 4, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Ananthi, S.; Raghavendran, H.R.B.; Sunil, A.G.; Gayathri, V.; Ramakhrisnan, G.; Vasanthi, H.R. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from Turbinaria ornate (Marine Brown Alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Moualek, I.; Iratni Aiche, G.; Mestar Guechaoui, N.; Lahcene, S.; Houali, K. Antioxidant and anti-inflammatory activities of Arbutus unedo aqueous extract. Asian Pac. J. Trop. Biomed. 2016, 6, 937–944. [Google Scholar] [CrossRef]
- Fernández-Botrán, R. Association of ABO blood group with COVID-19 susceptibility. Ciencia, Tecnología y Salud. 2020, 7(3), 325–332. [Google Scholar] [CrossRef]
- Medithi, S., Jonnalagadda, P.R.; Jee, B. Predominant role of antioxidants in ameliorating the oxidative stress induced by pesticides. Arch. Environ. Occup. Health. 2021, 76(2):61-74. [CrossRef]
- Fuchs, J. Potentials and limitations of the natural antioxidants RRR-alpha-tocopherol, L-ascorbic acid, and beta-carotene in cutaneous photoprotection. Free Radic. Biol. Med. 1998, 25, 848–873. [Google Scholar] [CrossRef] [PubMed]
- Steenvoorden, D.P.; van Henegouwen, G.M. The use of endogenous antioxidants to improve photoprotection. J. Photochem. Photobiol. B. 1997, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Medithi, S., Jonnalagadda, P.R.; Jee, B. Predominant role of antioxidants in ameliorating the oxidative stress induced by pesticides. Arch. Environ. Occup. Health. 2021, 76(2):61-74. [CrossRef]
- Benítez-Leite, S.; Ml, M.; Fernández, V.; Franco, D.; Ea, F.; Cuevas, M.A.; Alfonso, J.; Sales, L. Daño celular en una población infantil potencialmente expuesta a pesticidas Cell Damage in a Pediatric Population Potentially Exposed to Pesticides.
- Pattison, D.I.; Davies, M.J. Actions of ultraviolet light on cellular structures. Cancer: Cell Structures, Carcinogens and Genomic Instability. 2006, 131-157. [CrossRef]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet radiation-mediated damage to cellular DNA. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis. 2005, 571(1-2), 3-17. [CrossRef]
- Jacobson, M.K.; Kim, H. Coyle, W. R. et al. Effect of myristyl nicotinate on retinoic acid therapy for facial photodamage. Exp. Dermatol. 2007, 16, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Greul, A.K.; Grundmann, J.U.; Heinrich, F.; et al. Photoprotection of UV-irradiated human skin: an antioxidative combination of vitamins E and C, carotenoids, selenium and proanthocyanidins. Skin Pharmacol. Appl. Skin. Physiol. 2002; 15, 307–315. [Google Scholar]
- Greul, A.-K.; Grundmann, J.-U.; Heinrich, F.; Pfitzner, I.; Bernhardt, J.; Ambach, A.; Biesalski, H.-K.; Gollnick, H. Photoprotection of UV-Irradiated Human Skin: An Antioxidative Combination of Vitamins E and C, Carotenoids, Selenium and Proanthocyanidins. Skin Pharmacol. Appl Skin Physiol. 2002, 15, 307–315. [Google Scholar] [CrossRef]
- Chen, L.; Judy Y. Hu, J.J.; Steven Q. Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67:1013-24. [CrossRef]
- Simioni, C.; Schmidt, E.C. Felix, M.R.L.; Polo, L.P.; Rover, T.; Kreusch, M.; Pereira, D.T.; Chow, F.; Ramlov, F.; Maraschin, M.; Bouzon, A.L. Effects of Ultraviolet Radiation (UVA+UVB) on Young Gametophytes of Gelidium floridanum: Growth Rate, Photosynthetic Pigments, Carotenoids, Photosynthetic Performance, and Ultrastructure. Photochemistry and Photobiology. 2014. [CrossRef]
- Greul, A.-K.; Grundmann, J.-U.; Heinrich, F.; Pfitzner, I.; Bernhardt, J.; Ambach, A.; Biesalski, H.-K.; Gollnick, H. Photoprotection of UV-Irradiated Human Skin: An Antioxidative Combination of Vitamins E and C, Carotenoids, Selenium and Proanthocyanidins. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 307–315. [Google Scholar] [CrossRef]
- Nichols, J.A., Katiyar, S.K. Skin photoprotection by natural polyphenols: anti-inflammatory, antioxidant, and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302: 71–83 (2010). [CrossRef]
- Kumar, S.; Devi, S.; Misra, P. ; Priyanka. Solar and artificial ultraviolet-B induced erythrocytes hemolysis with photosensitizers. Ind. J. Experimen. Biol. 2009, 47, 906–910. [Google Scholar]
- Rostamabadi, H.; Reza, Falsafi, S.; Mahdi, Jafari, S. Nanoencapsulation of carotenoids within lipid-based nanocarriers. JCR 2019, 298, 38–67. [Google Scholar] [CrossRef]
- Salvia-Trujillo, L.; Verkempinck, S.; Rijal, SK.; Van Loey, A.; Grauwet, T.; Hendrickx, M. Lipid nanoparticles with fats or oils containing β-carotene: Storage stability and in vitro digestibility kinetics. Food Chem. 2019, 278(1), 396–405. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam R.; Ki, J-S.A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar Drugs. 2018, 16(1):26. [CrossRef]
- Zmijewski, C.M. Inmunohematology. Eherd Edition; Appleton Century Crojts: New York, NY, USA, 1978. [Google Scholar]
- Hou, J.; Cui, H.L. In Vitro Antioxidant, Antihemolytic, and Anticancer Activity of the Carotenoids from Halophilic Archaea. Curr. Microbiol. 2018, 75, 266–271. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Carotenoids and Flavonoids Contribute to Nutritional Protection against Skin Damage from Sunlight. Mol. Biotechnol. 2007, 37:26–30. [CrossRef]
- Carini, M.; G. Aldini, G.; Bombardelli, E.; Morazzon, P.; R. Maffei Facino, R.M. UVB-induced hemolysis of rat erythrocytes: Protective effect of procyanidins from grape seeds. Life Sciences. 2000, 67, 1799–1814. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. b-Carotene and other carotenoids in protection from sunlight. Am. J. Clin. Nutr. 2012, 96. [Google Scholar] [CrossRef] [PubMed]
- Camera, E.; Mastrofrancesco, A.; Fabbri, C.; Daubrawa, F.; Picardo, M.; Sies, H.; Stahl, W. Astaxanthin, canthaxanthin and b-carotene differently affect UVA-induced oxidative damage and expression of oxidative stress-responsive enzymes. Exp. Dermatol. 2009, 18: 222–231. [CrossRef]
- Lourenço-Lopes, C.; Fraga-Corral, M.; Jimenez-Lopez, C.; Carpena, M.; Pereira, A.G.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Biological action mechanisms of fucoxanthin extracted from algae for application in food and cosmetic industries. Trends Food Sci. Technol. 2021, 117, 163–181. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Zakharova, L.V.; Daurtseva, A.V.; Flisyuk, E.V.; Shikov, A.N. Efficacy of Natural Deep Eutectic Solvents for Extraction of Hydrophilic and Lipophilic Compounds from Fucus vesiculosus. Molecules 2021, 26, 4198. [Google Scholar] [CrossRef]
- Akhavan, S.; Assadpour, E.; Katouzian, I.; Jafari, S.M. Lipid nanoscale cargos for the protection and delivery of food bioactive ingredients and nutraceuticals. Trends Food Sci. Technol. 2018, 74, 132–146. [Google Scholar] [CrossRef]
- Aly, R.; Yousef, A.; Elbably, O. Association of ABO blood group and risk of breast cancer. J. Blood Disord. Transfus. 2014, 5: 241. [CrossRef]
- Abella, A.; Messaoudi, C.; Laurent, D.; Marot, D.; Chalas, J.; Breux, J.; Claise, C.; Lindenbaum, A. A method for simultaneous determination of plasma and erythrocyte antioxidant status. Evaluation of the antioxidant activity of vitamin E in healthy volunteers. Br. J. Clin. Pharmacol. 1996, 42, 737–741. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of the direct hemolysis from Fucoxanthin (FXN) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0P < 0.01 in comparison to the most affected blood group by concentration.
Figure 1.
Effect of the direct hemolysis from Fucoxanthin (FXN) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0P < 0.01 in comparison to the most affected blood group by concentration.
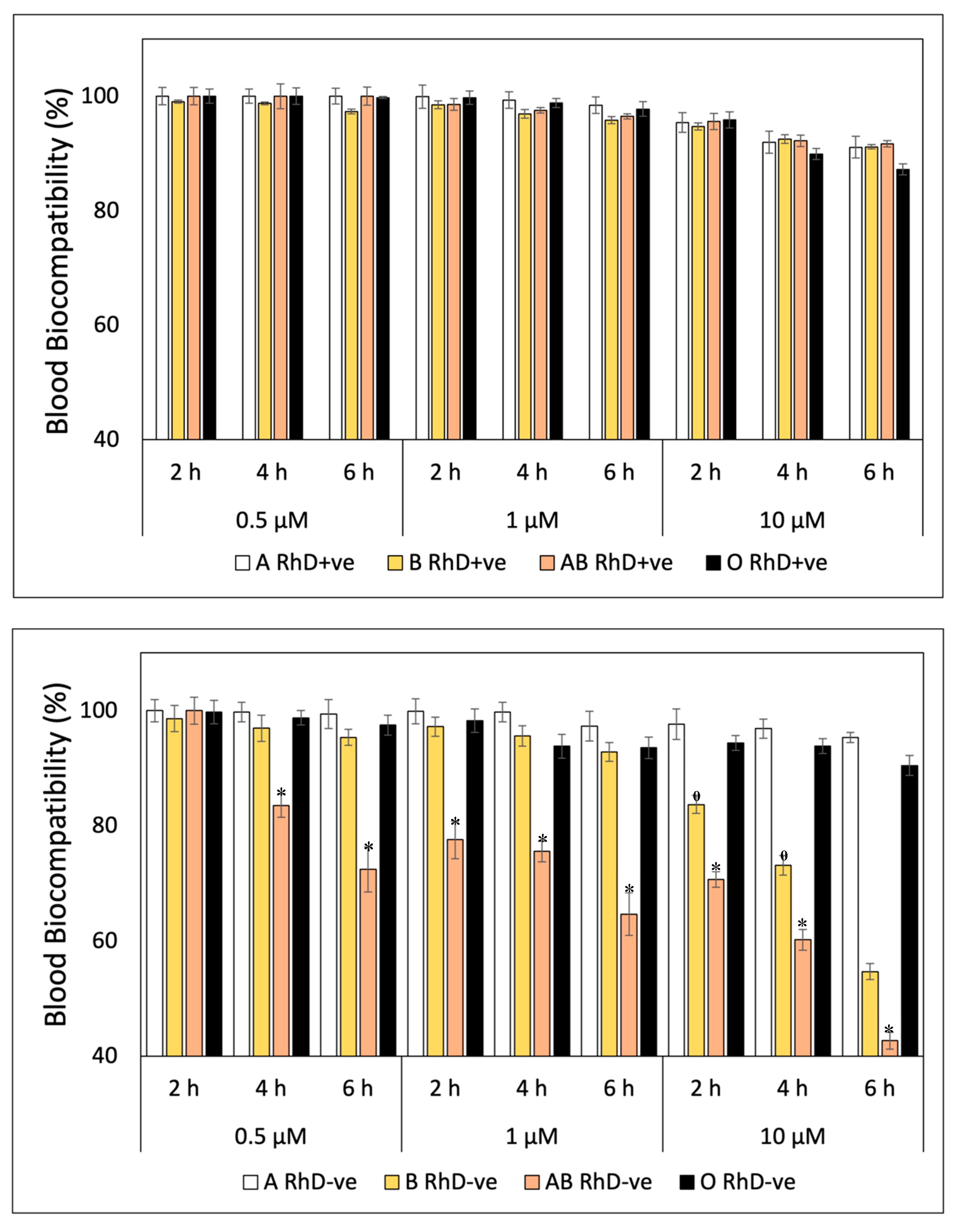
Figure 2.
Effect of the direct hemolysis from β-Carotene (β-Car) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 2.
Effect of the direct hemolysis from β-Carotene (β-Car) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
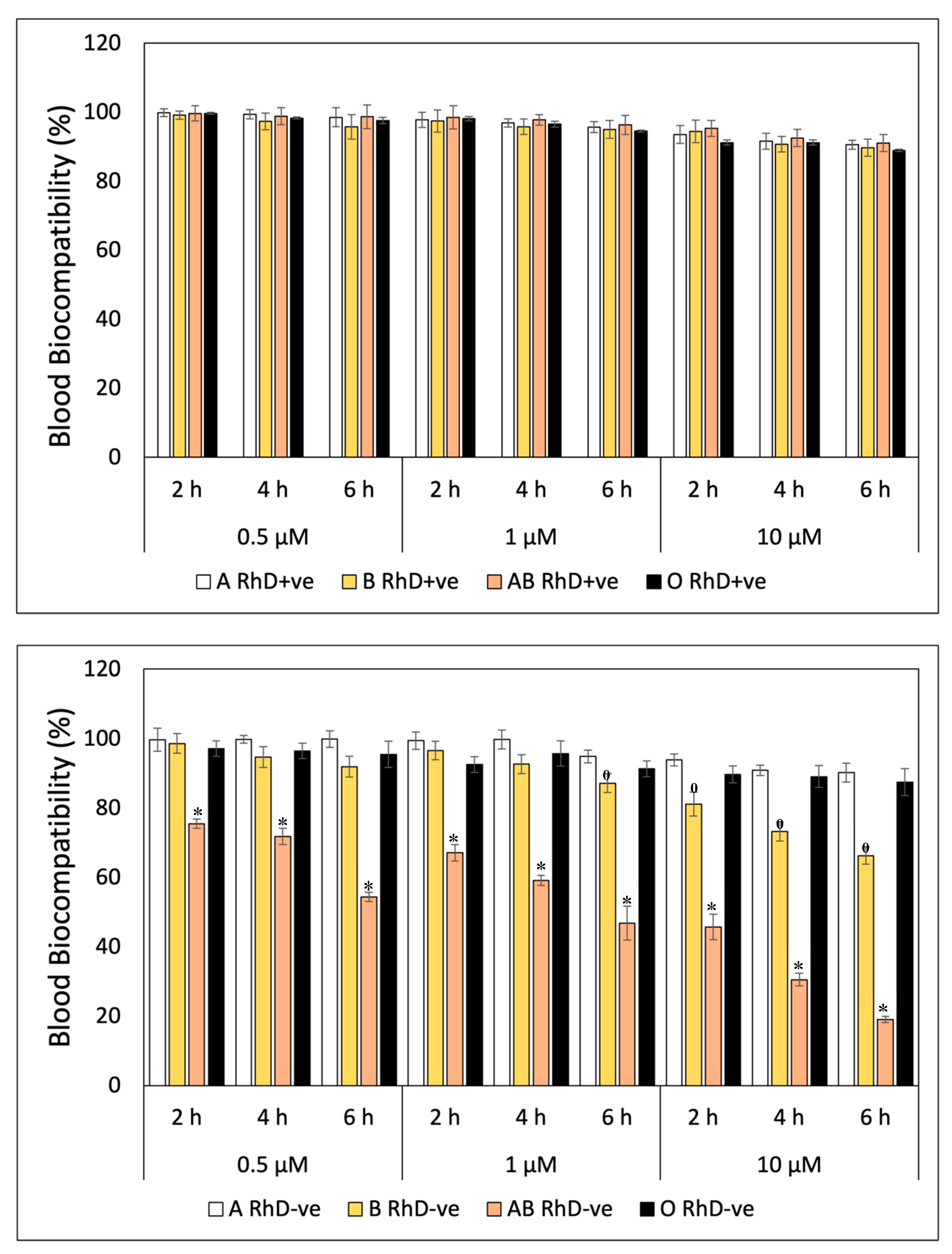
Figure 3.
Effect of the direct hemolysis from Gallic acid (GA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. Different lowercase letter in the same concentration is different at a significance level of 0.05 (Tukey post hoc test). *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration.
Figure 3.
Effect of the direct hemolysis from Gallic acid (GA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. Different lowercase letter in the same concentration is different at a significance level of 0.05 (Tukey post hoc test). *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration.
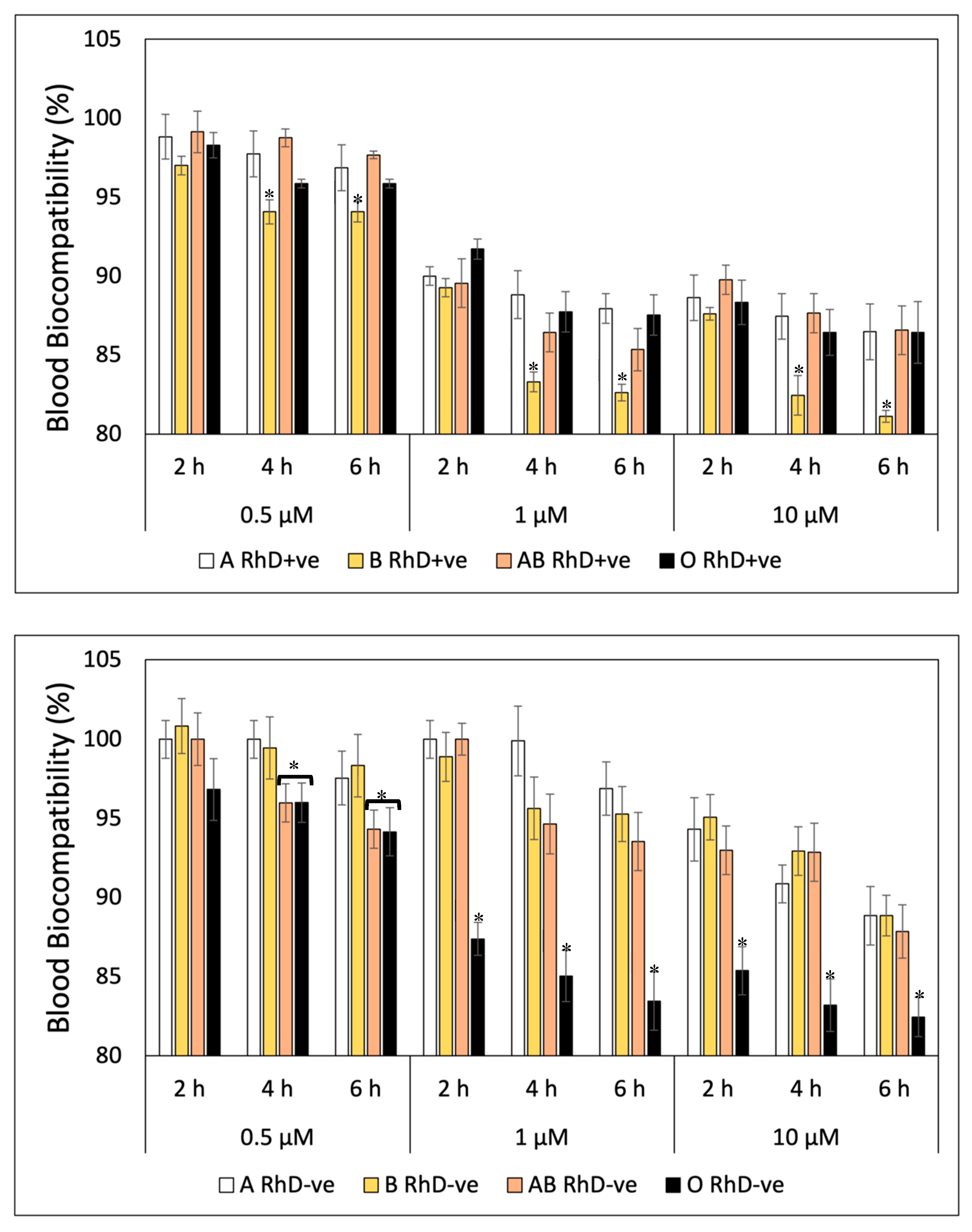
Figure 4.
Effect of the direct hemolysis from Quercetin (QUE) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 4.
Effect of the direct hemolysis from Quercetin (QUE) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
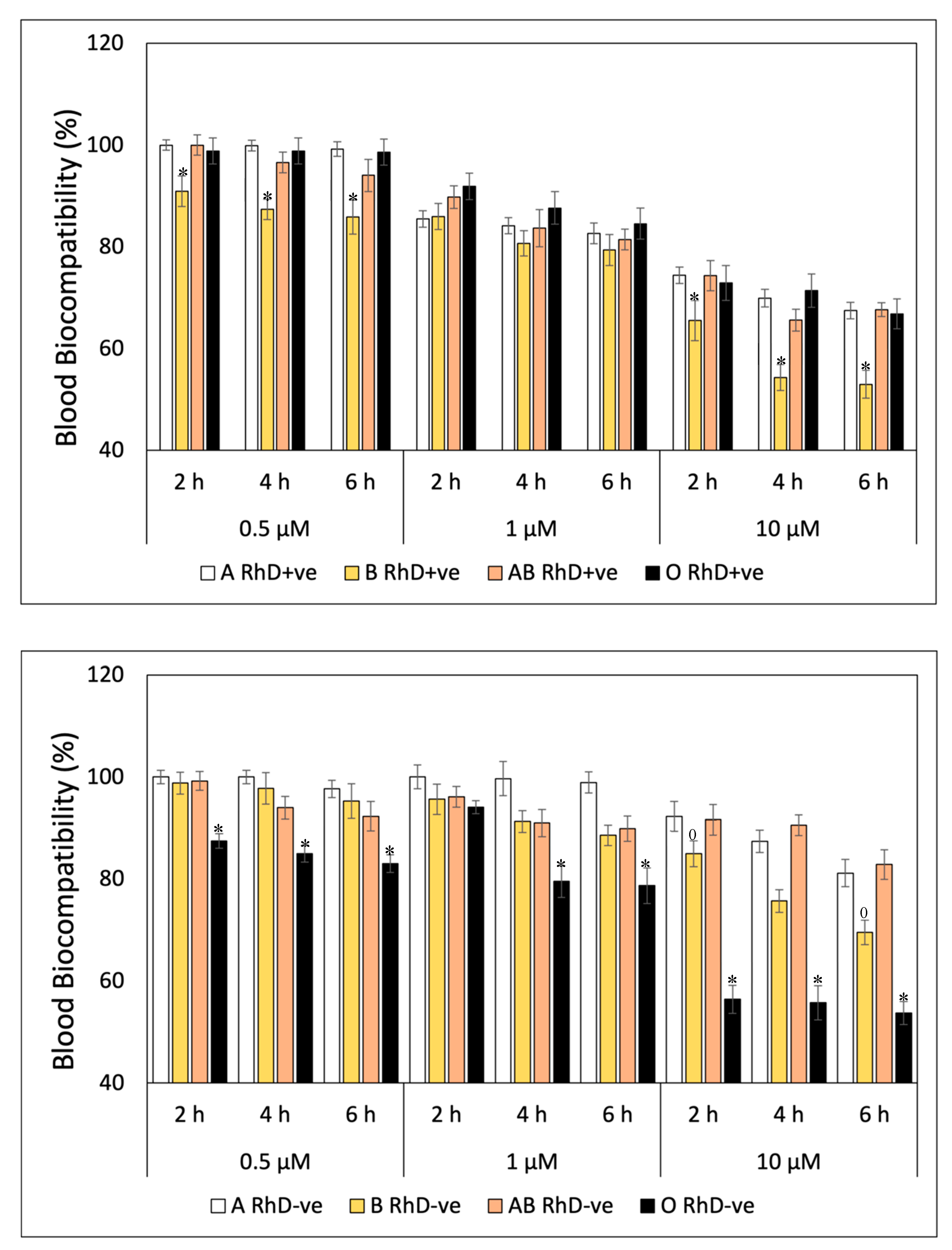
Figure 5.
Effect of the direct hemolysis from Ascorbic acid (AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 5.
Effect of the direct hemolysis from Ascorbic acid (AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at different concentration (0.5, 1.0 and 10 µM) and at variance exposure times (2, 4 and 6 h). Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
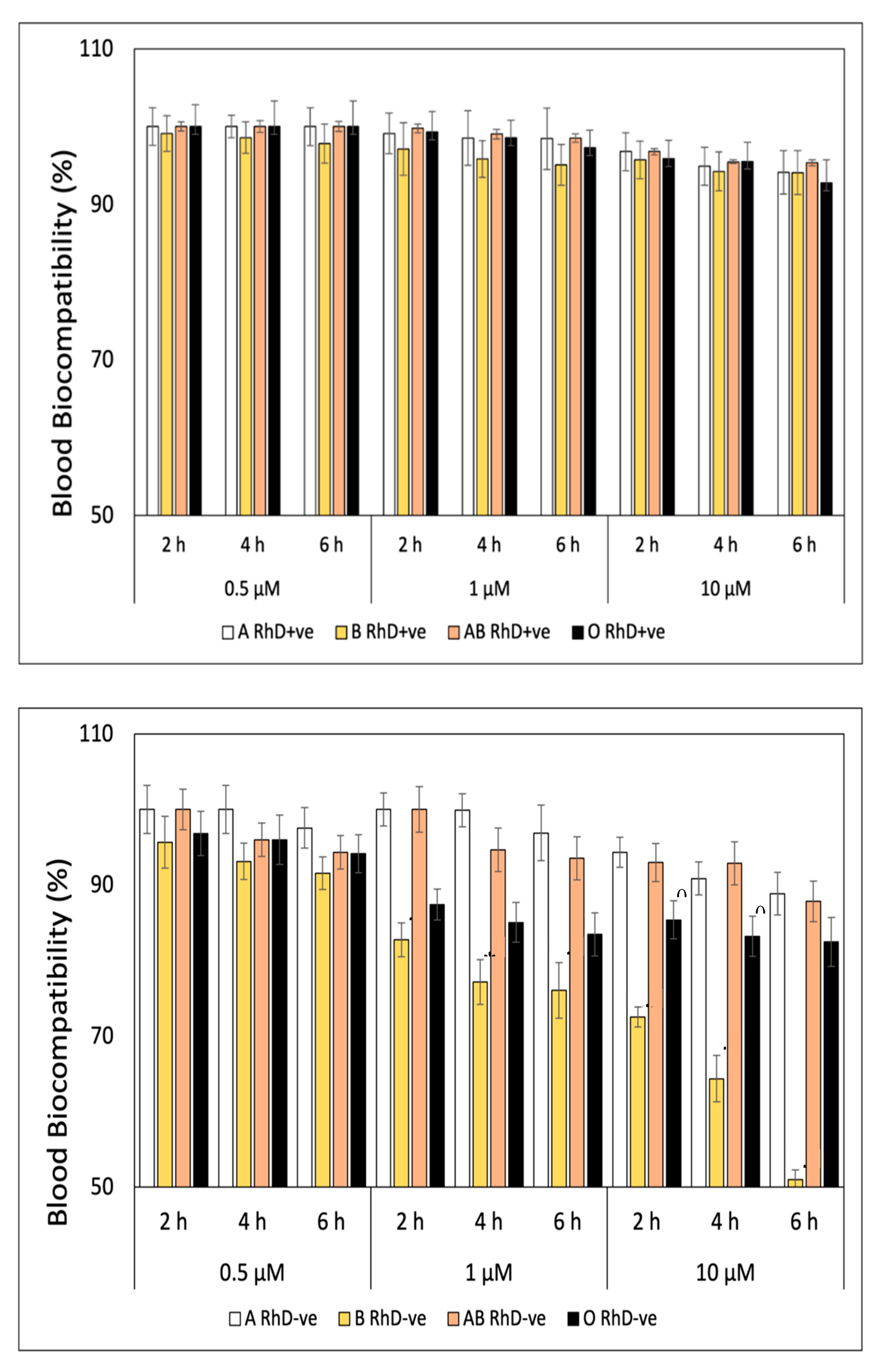
Figure 6.
Effect of the heat-induced hemolysis to evaluate membrane stabilization (%) from Fucoxanthin (FXN) and standard compound (β-Car, GA, QUE, and AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at a concentration of 10 µM. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group mostly protected from heat-induced hemolysis for each compound tested.
Figure 6.
Effect of the heat-induced hemolysis to evaluate membrane stabilization (%) from Fucoxanthin (FXN) and standard compound (β-Car, GA, QUE, and AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at a concentration of 10 µM. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group mostly protected from heat-induced hemolysis for each compound tested.
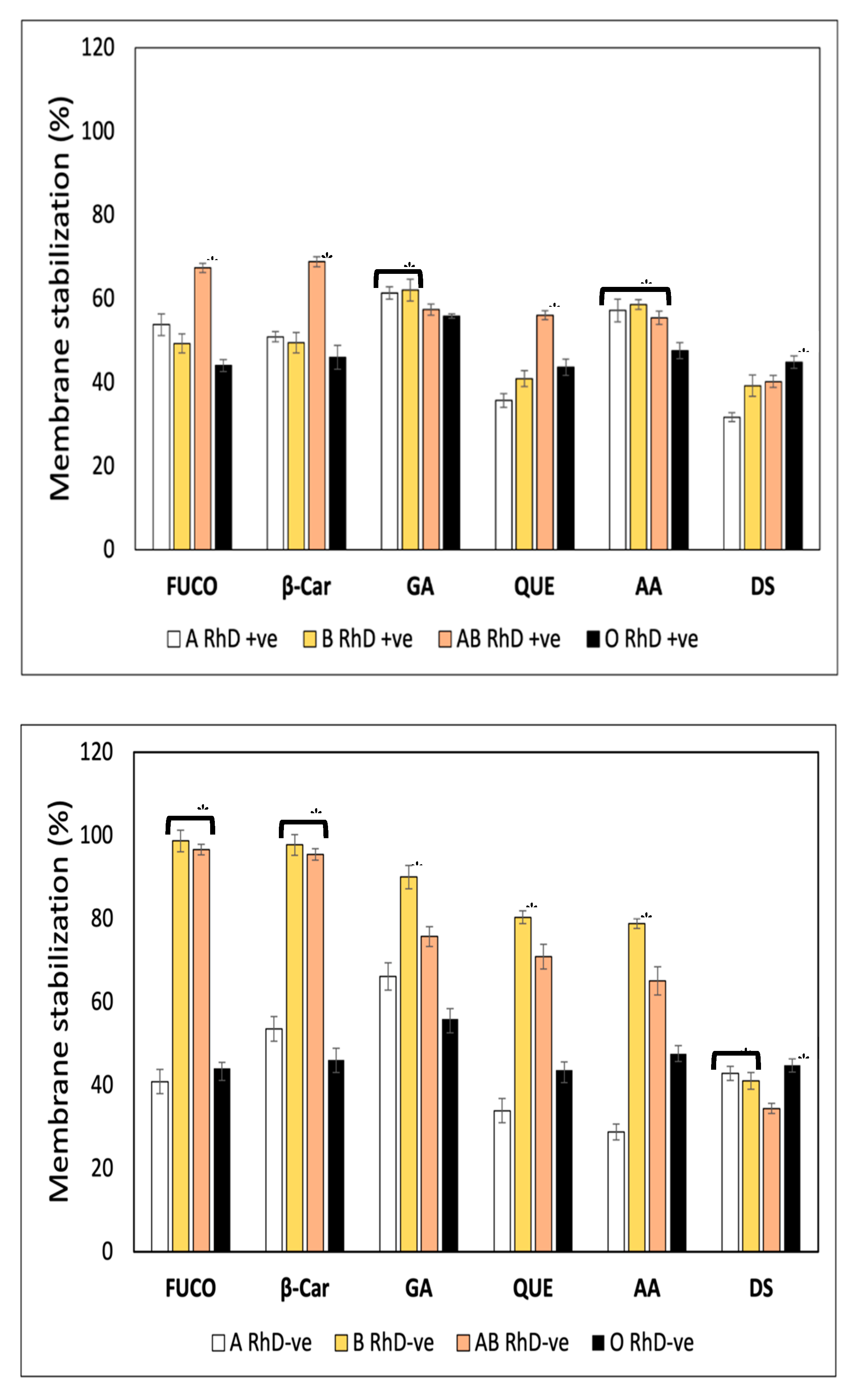
Figure 7.
Effect of the hypotonicity-induced hemolysis to evaluate membrane stabilization (%) from Fucoxanthin (FXN) and standard compound (β-Car, GA, QUE, and AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at a concentration of 10 µM. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 7.
Effect of the hypotonicity-induced hemolysis to evaluate membrane stabilization (%) from Fucoxanthin (FXN) and standard compound (β-Car, GA, QUE, and AA) on different ABO RhD+ve (A) and RhD-ve (B) blood groups at a concentration of 10 µM. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
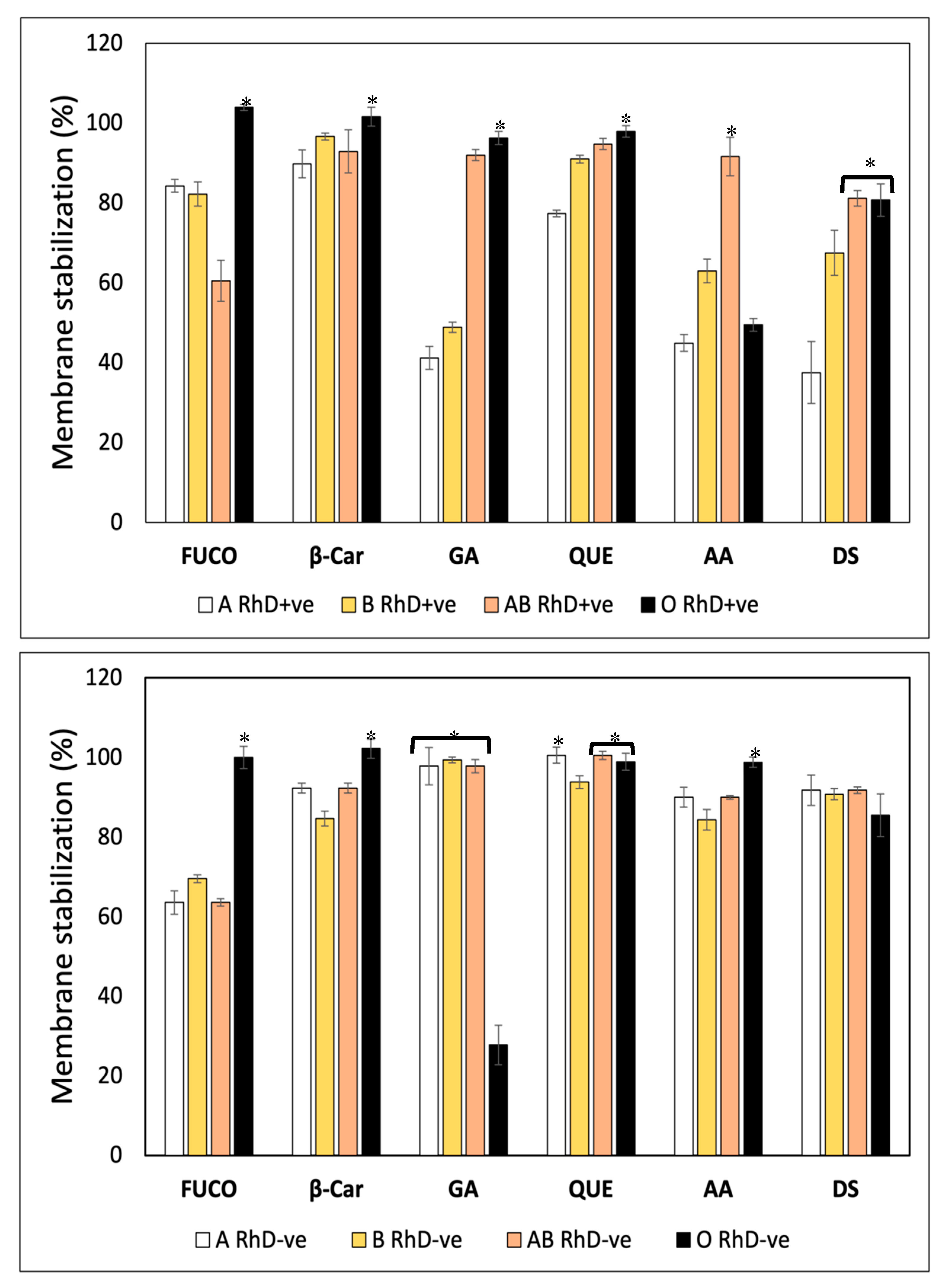
Figure 8.
Blood Susceptibility study. Effect of UV-A (315-395 nm) and UV-B (280-315 nm) irradiation-induced hemoglobin released at different exposure time (30, 60 and 120 min) on ABO and RhD different blood types to evaluate blood susceptibility. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration.
Figure 8.
Blood Susceptibility study. Effect of UV-A (315-395 nm) and UV-B (280-315 nm) irradiation-induced hemoglobin released at different exposure time (30, 60 and 120 min) on ABO and RhD different blood types to evaluate blood susceptibility. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration.
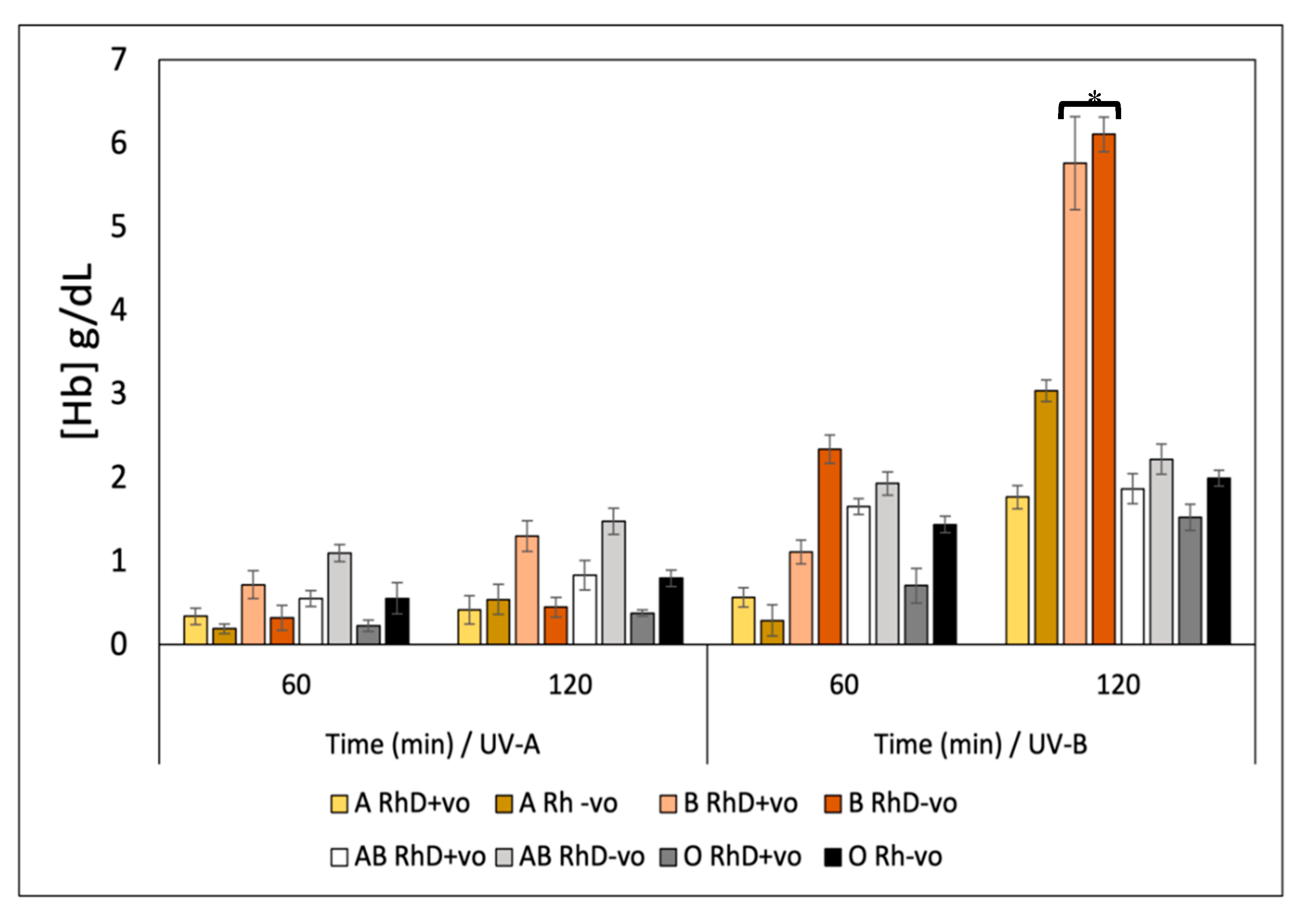
Figure 9.
Effect of the oxidative-hemolysis by the free radical generator AAPH as an inducer hemoglobin release on ABO and RhD different blood types to evaluate blood susceptibility. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. Different lowercase letter in the same concentration is different at a significance level of 0.05 (Tukey post hoc test).
Figure 9.
Effect of the oxidative-hemolysis by the free radical generator AAPH as an inducer hemoglobin release on ABO and RhD different blood types to evaluate blood susceptibility. Barr represents standard deviation of at least three repetitions (n > 3). All data was analyzed by one-way ANOVA. Different lowercase letter in the same concentration is different at a significance level of 0.05 (Tukey post hoc test).
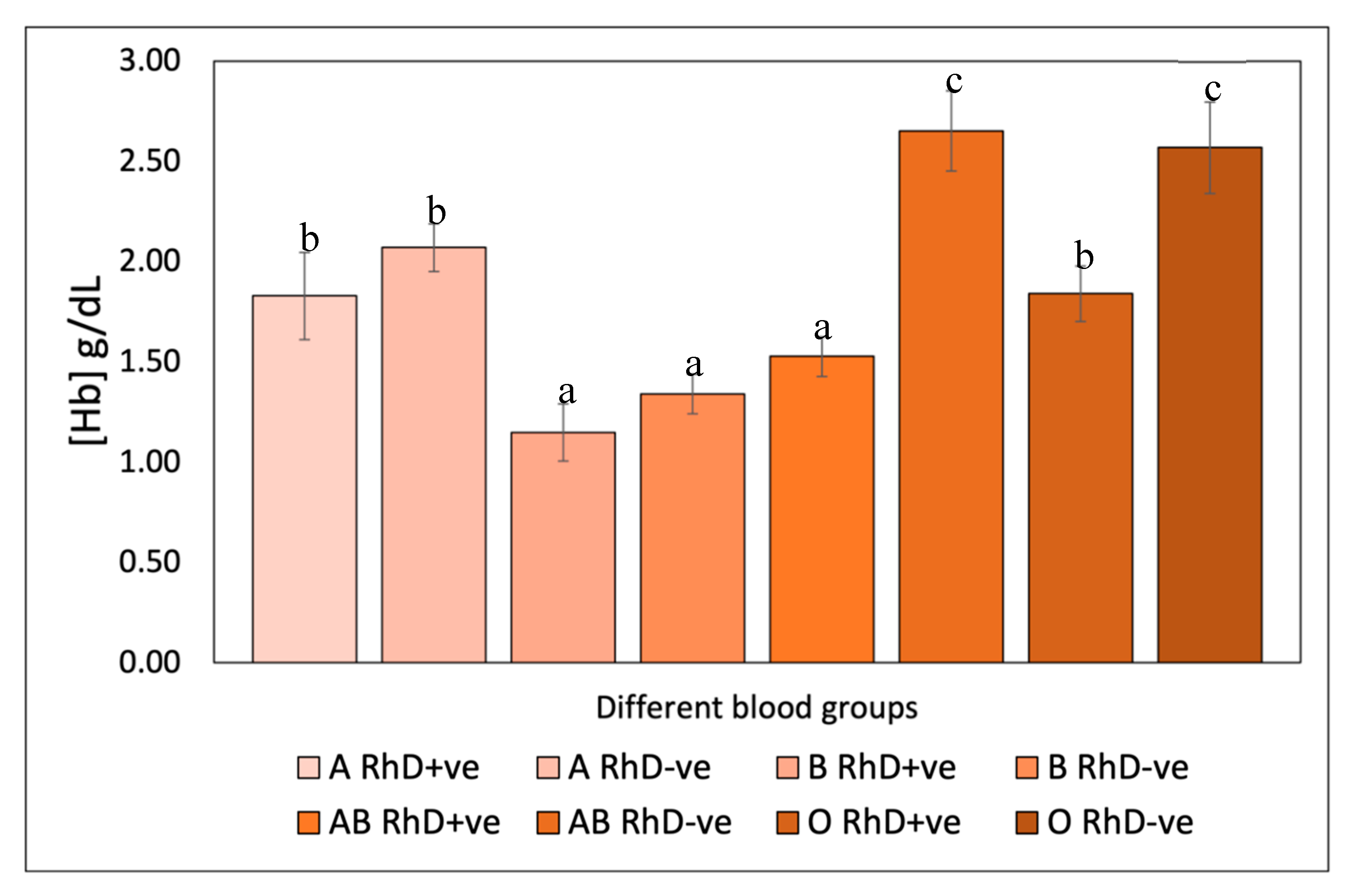
Figure 10.
UV absorption spectra of fucoxanthin and β-Carotene as a sun filter before and after UV-A (315-395 nm) and UV-B (280-315 nm) exposure. Exposure time of 120 min with an irradiation intensity of 0.85 mW/cm2. (A) UV absorption spectra of Fucoxanthin and β-Carotene photostability without UVR exposure. (B) UV absorption spectra of Fucoxanthin and β-Carotene photostability under UV-A exposure. (C) UV-Vis analysis of Fucoxanthin and β-Carotene photostability under UV-B light exposure.
Figure 10.
UV absorption spectra of fucoxanthin and β-Carotene as a sun filter before and after UV-A (315-395 nm) and UV-B (280-315 nm) exposure. Exposure time of 120 min with an irradiation intensity of 0.85 mW/cm2. (A) UV absorption spectra of Fucoxanthin and β-Carotene photostability without UVR exposure. (B) UV absorption spectra of Fucoxanthin and β-Carotene photostability under UV-A exposure. (C) UV-Vis analysis of Fucoxanthin and β-Carotene photostability under UV-B light exposure.
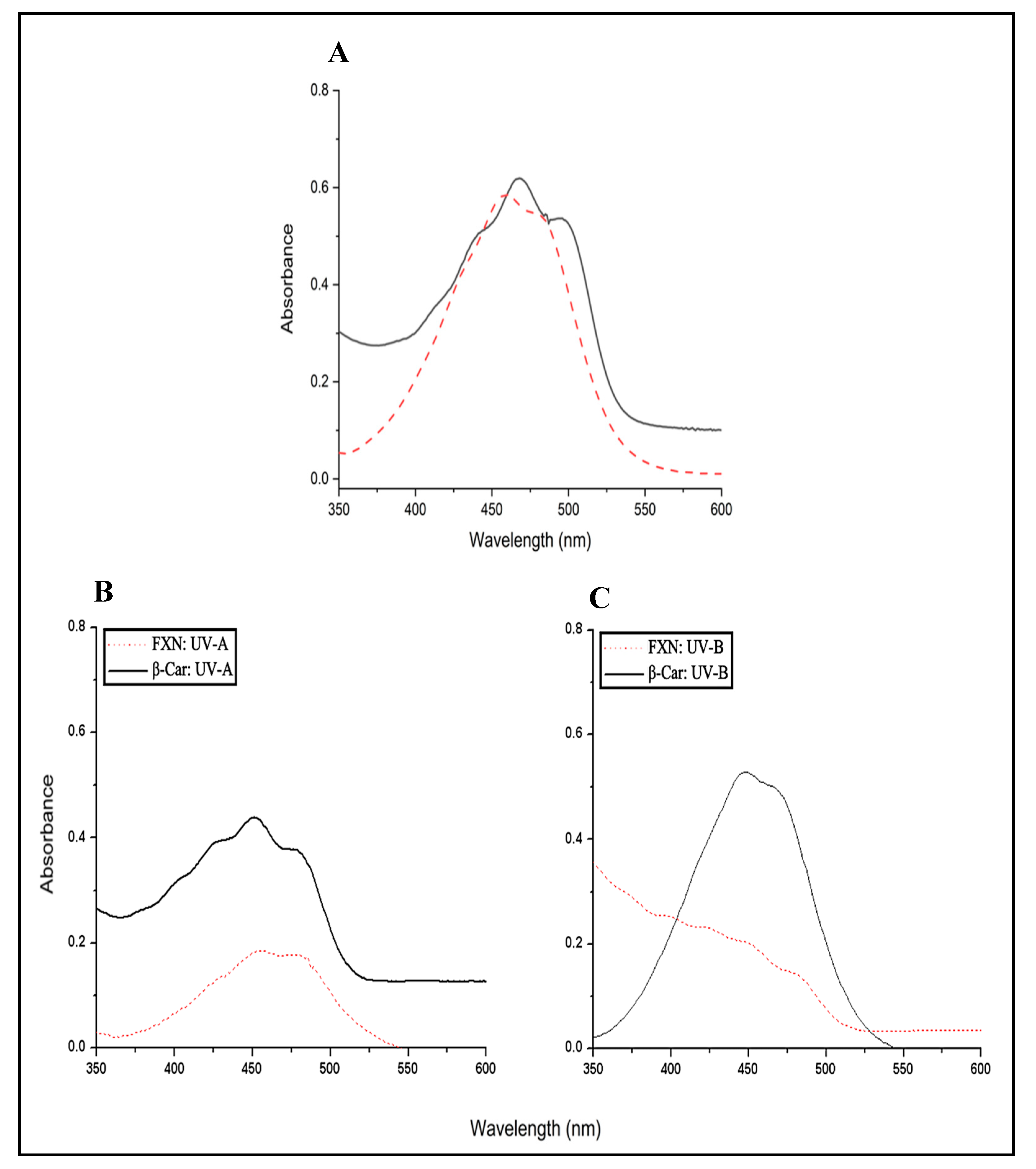
Figure 11.
Photoprotector effect of Fucoxanthin (FXN) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 11.
Photoprotector effect of Fucoxanthin (FXN) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
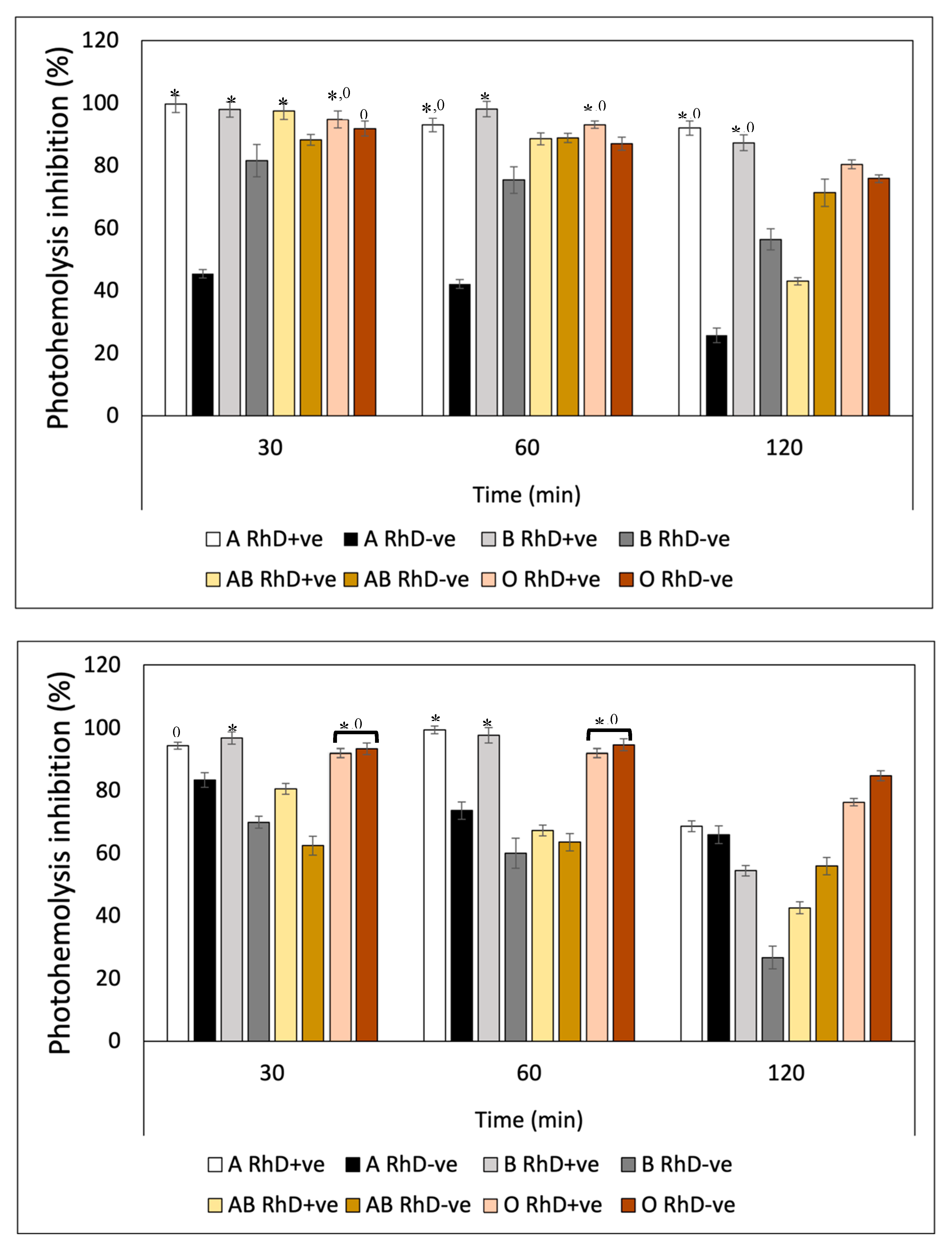
Figure 12.
Photoprotector effect of β-Carotene (β-Car) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 12.
Photoprotector effect of β-Carotene (β-Car) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
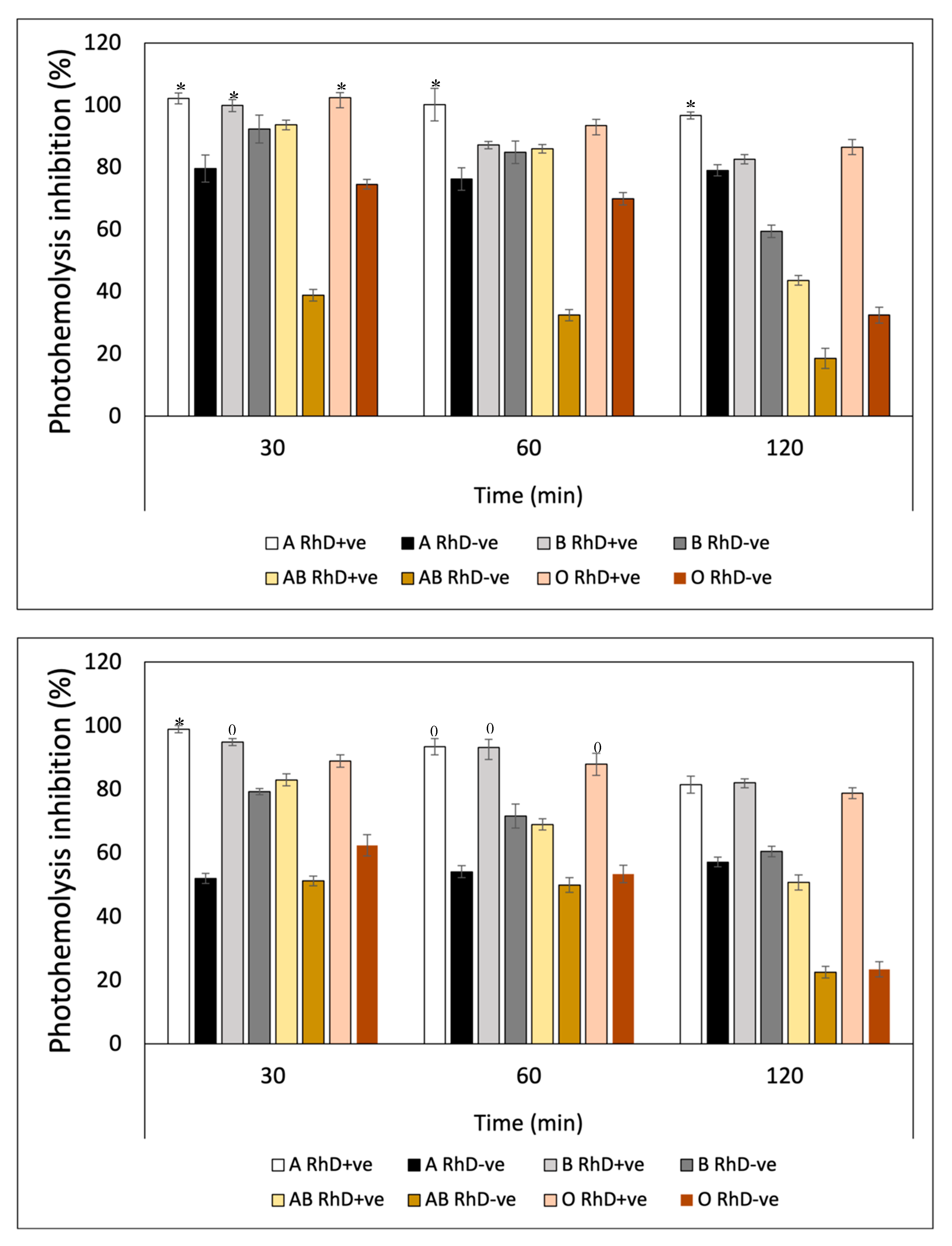
Figure 13.
Photoprotector effect of Gallic acid (GA) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 13.
Photoprotector effect of Gallic acid (GA) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
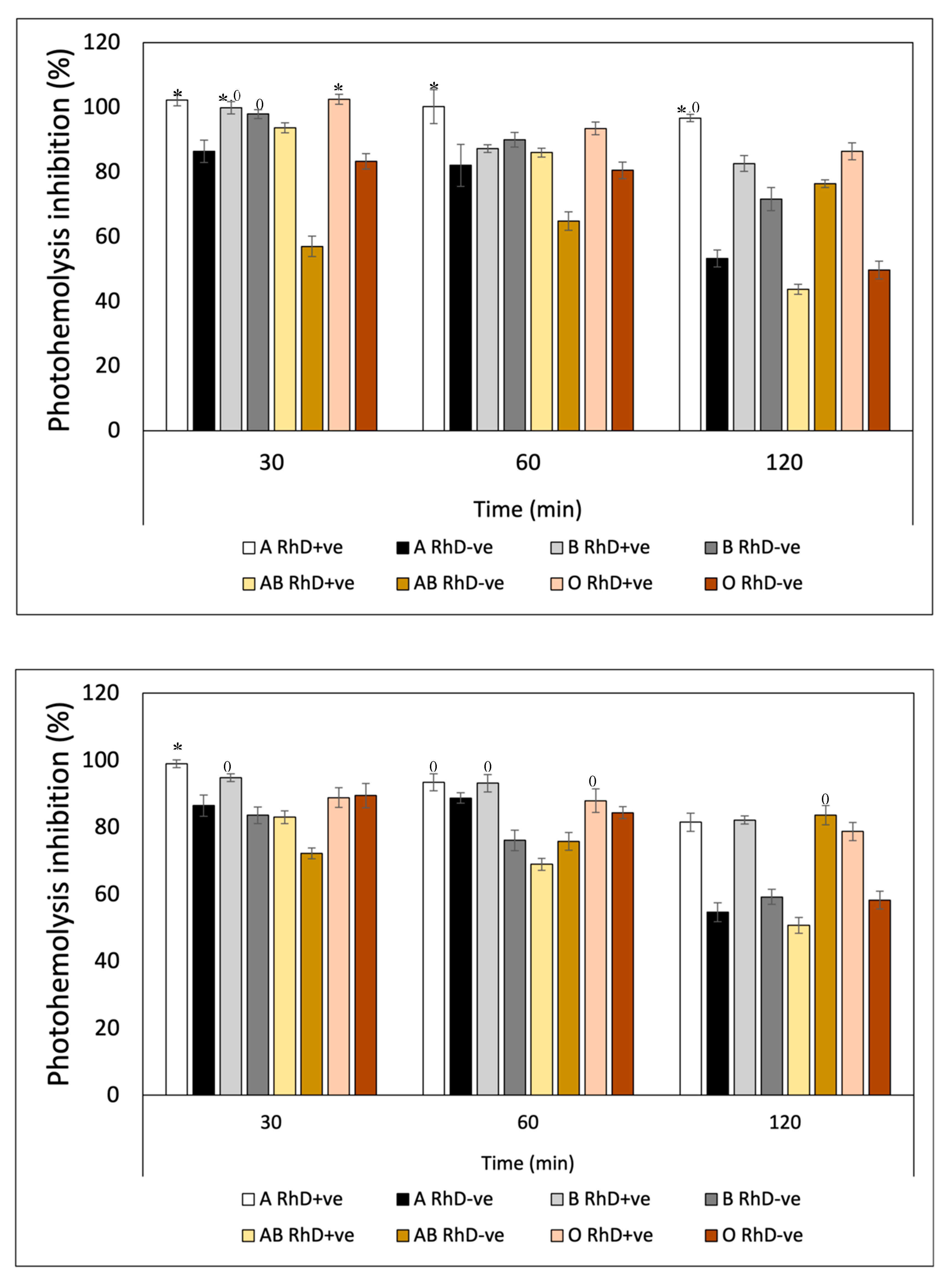
Figure 14.
Photoprotector effect of Quercetin (QUE) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 14.
Photoprotector effect of Quercetin (QUE) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
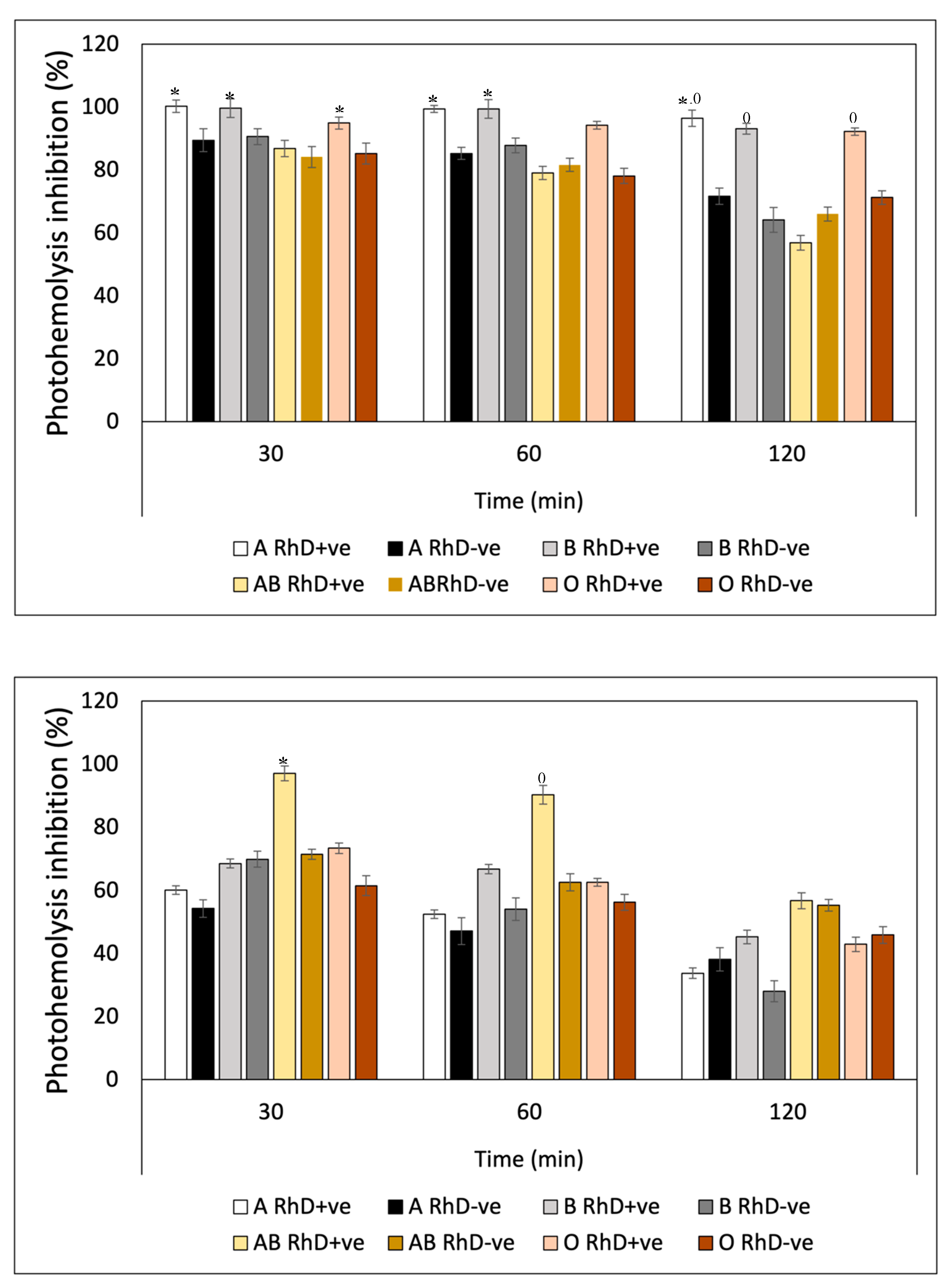
Figure 15.
Photoprotector effect of Ascorbic acid (AA) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
Figure 15.
Photoprotector effect of Ascorbic acid (AA) against UV-A (A) and UV-B (B) irradiation-induced hemolysis on human erythrocytes with different ABO and RhD blood groups at different exposure time (30, 60 and 120 min). A univariate ANOVA each exposure time was applied. *P < 0.01 Blood group most affected compared to other blood phenotypes by concentration. 0 P < 0.01 in comparison to the most affected blood group by concentration.
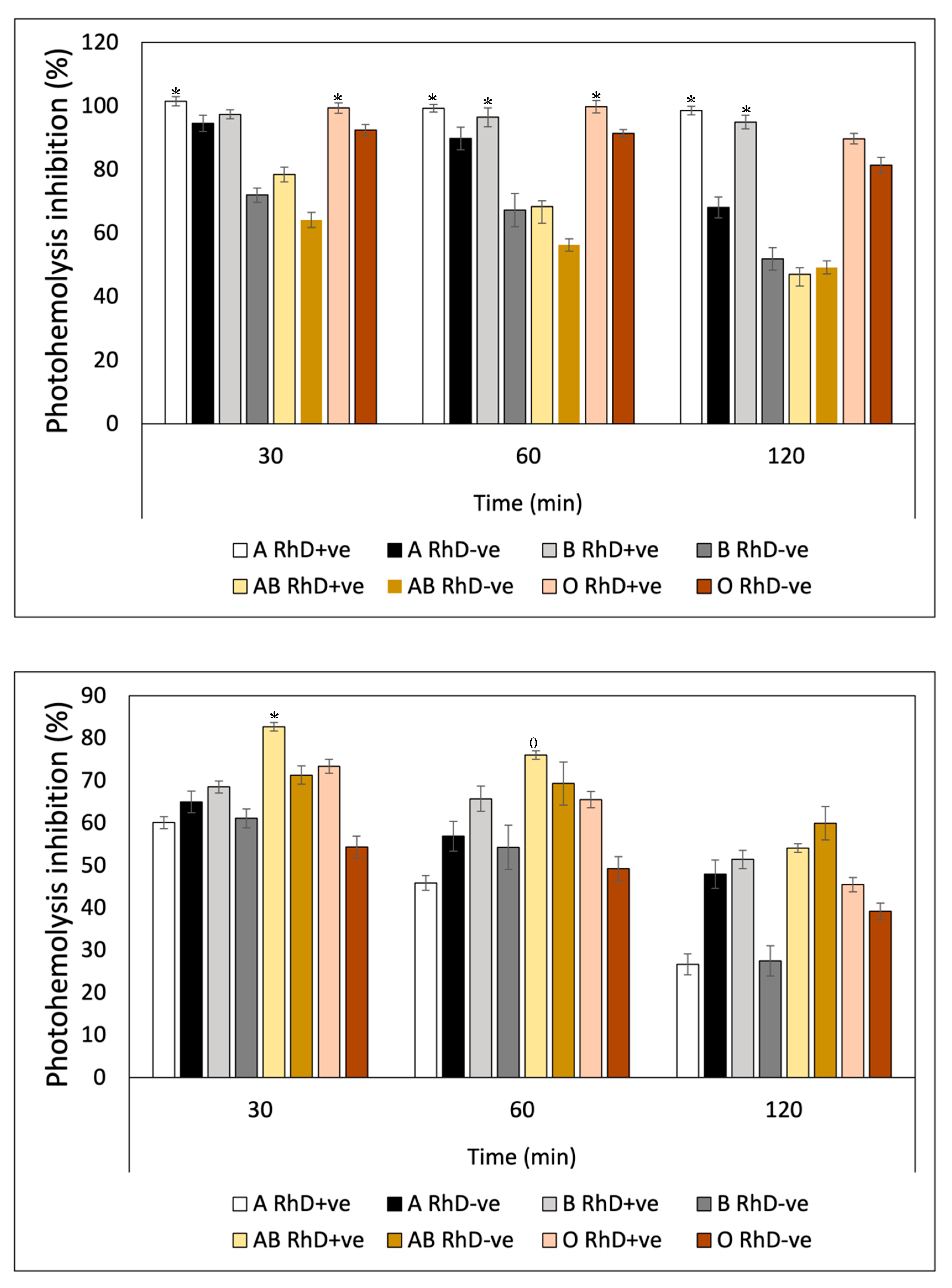
Figure 16.
Erythrocyte morphological changes associated with cellular photooxidation caused by ultraviolet radiation at 120 min and the photoprotective effect of Fucoxanthin and β-carotene exposed to a concentration of 10 µM. Control erythrocyte group: (A, B, and C) Erythrocytes not exposed to UV radiation. Group of erythrocytes exposed to UV-A radiation: (D, E, and F) Erythrocyte exposed to UV-A radiation without antioxidant; Erythrocyte exposed to UV-A radiation with Fucoxanthin; Erythrocyte exposed to UV-A radiation with β-carotene; Erythrocyte exposed to UV-A radiation with Astaxanthin. Group of erythrocytes exposed to UV-B radiation: (G, H, and I) Erythrocyte exposed to UV-B radiation without antioxidant; Erythrocyte exposed to UV-B radiation with Fucoxanthin; Erythrocyte exposed to UV-B radiation with β-carotene; Erythrocyte exposed to UV-B radiation with Astaxanthin. Micrograph at 100×. The bar represents a 5 μm scale. Exposure time 120 min.
Figure 16.
Erythrocyte morphological changes associated with cellular photooxidation caused by ultraviolet radiation at 120 min and the photoprotective effect of Fucoxanthin and β-carotene exposed to a concentration of 10 µM. Control erythrocyte group: (A, B, and C) Erythrocytes not exposed to UV radiation. Group of erythrocytes exposed to UV-A radiation: (D, E, and F) Erythrocyte exposed to UV-A radiation without antioxidant; Erythrocyte exposed to UV-A radiation with Fucoxanthin; Erythrocyte exposed to UV-A radiation with β-carotene; Erythrocyte exposed to UV-A radiation with Astaxanthin. Group of erythrocytes exposed to UV-B radiation: (G, H, and I) Erythrocyte exposed to UV-B radiation without antioxidant; Erythrocyte exposed to UV-B radiation with Fucoxanthin; Erythrocyte exposed to UV-B radiation with β-carotene; Erythrocyte exposed to UV-B radiation with Astaxanthin. Micrograph at 100×. The bar represents a 5 μm scale. Exposure time 120 min.
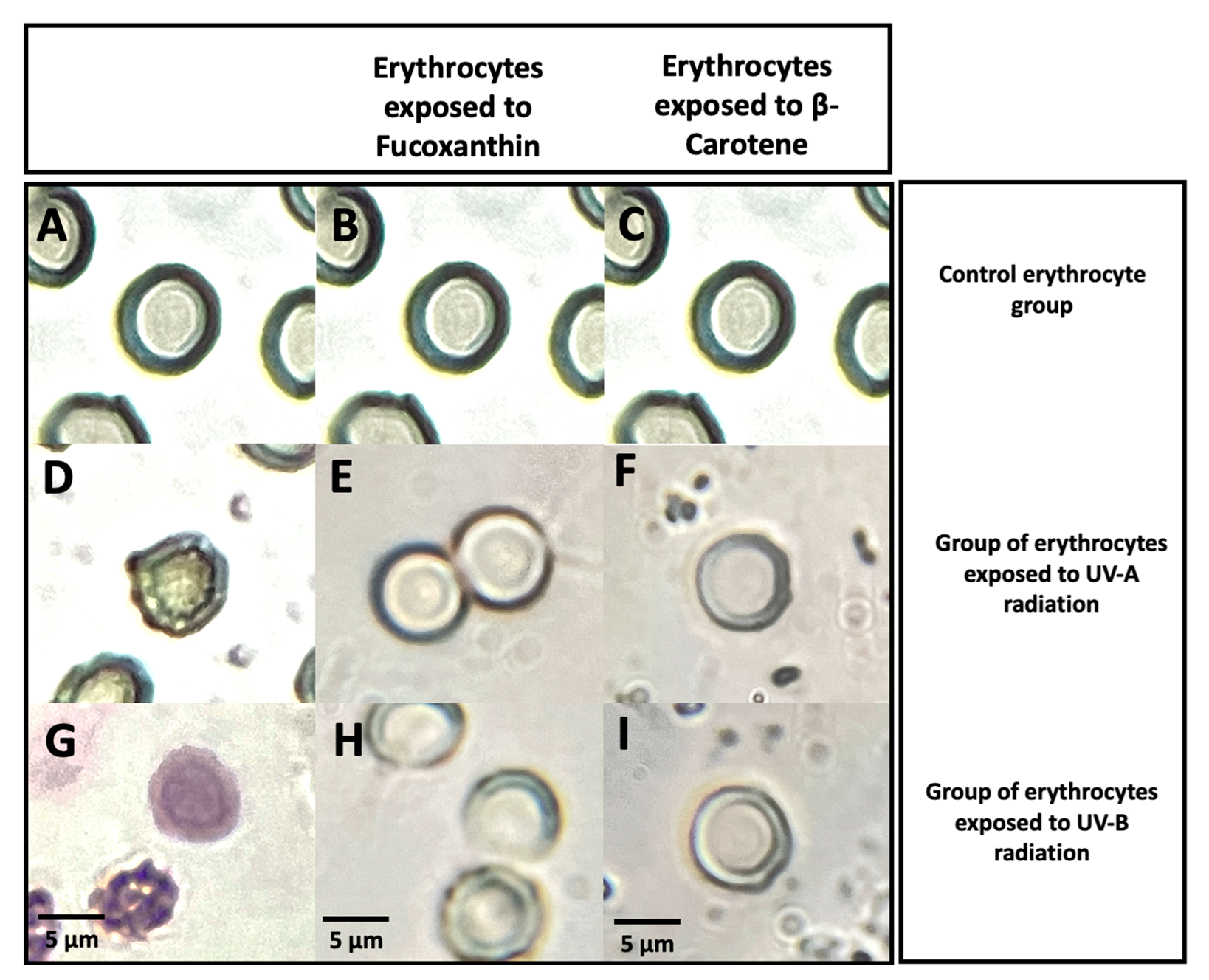

Table 1.
Antioxidant activities (µmol TE/g) of FXN and standard compound (β-Car, GA, QUE, and AA).
| µmol TE/g ± SD | |||
|---|---|---|---|
| Compounds | FRAP | ABTS | DPPH |
| FXN | 2687.03 b ±173.26 | 2638.53c ±244.87 | 303.15 c ±42.66 |
| β - Car | 2780.87 b ±92.23 | 2371.99 c ±26.79 | 348.12 c ±38.49 |
| GA | 5970.45 a ±230.07 | 3124.72 b ±133.22 | 614.74 b ±99.77 |
| QUE | 896.70 c ±19.06 | 2037.62 d ±80.62 | 3980.61 a ±222.58 |
| AA | 2684.54 b ±104.13 | 3412.22 a ±21.94 | 4190.9 a ±155.16 |
FXN, fucoxanthin; β–Car, β–carotene; GA, gallic acid; QUE, quercetin; AA, ascorbic acid. The data represent the mean ± standard deviation of at least three analyses. Different letters mean significant differences (p ≤ 0.05). One-way analysis of variance (ANOVA) with multiple comparisons of Tukey test.
Table 2.
Erythroprotective effect of Fucoxanthin, β–carotene, Gallic acid, Quercetin, Ascorbic acid. against the oxidative hemolysis induced by AAPH on different ABO RhD+ve blood types.
Table 2.
Erythroprotective effect of Fucoxanthin, β–carotene, Gallic acid, Quercetin, Ascorbic acid. against the oxidative hemolysis induced by AAPH on different ABO RhD+ve blood types.
| Inhibition of Hemolysis (%) | ||||
|---|---|---|---|---|
| Different blood groups (RhD+ ve) | ||||
| Compounds | A | B | AB | O |
| FXN | 100.70 Aa ±0.71 | 79.57 Bd ±1.13 | 97.30 Bb ±1.37 | 91.15 Bc ±1.04 |
| β - Car | 101.05 Aa ±3.78 | 84.82 Ac ±2.56 | 98.87 ABb ±0.49 | 97.05 Ab ±0.31 |
| GA | 94.42 Bb ±1.08 | 77.16 Bd ±0.49 | 99.43 Aa ±1.37 | 81.94 Dc ±1.08 |
| QUE | 95.61 Bb ±0.41 | 70.78 Cc ±0.64 | 100.85 Aa ±1.43 | 83.51 Cb ±0.61 |
| AA | 93.12 Bb ±3.55 | 62.84 Dd ±3.08 | 99.29 Aa ±1.07 | 83.33 CDc ±3.26 |
FXN, fucoxanthin; β – Car, β – carotene; GA, gallic acid; QUE, quercetin; AA, ascorbic acid. The compounds’ concentration was 10 µM. 1Values are mean ± standard deviation of at least three repetitions. Values followed by different uppercase letter in the same row indicate significant differences (p <0.05) between the reaction time tested of each phytochemical compound.
Table 3.
Erythroprotective effect of Fucoxanthin, β–carotene, Gallic acid, Quercetin, Ascorbic acid. against the oxidative hemolysis induced by AAPH on different ABO RhD-ve blood types.
Table 3.
Erythroprotective effect of Fucoxanthin, β–carotene, Gallic acid, Quercetin, Ascorbic acid. against the oxidative hemolysis induced by AAPH on different ABO RhD-ve blood types.
| Inhibition of Hemolysis (%) | ||||
|---|---|---|---|---|
| Different blood groups (RhD-ve) | ||||
| Compounds | A | B | AB | O |
| FXN | 79.93 Ab ±3.28 | 92.41 Aa ±1.34 | 81.28 Bb ±1.28 | 91.15 Ba ±1.43 |
| β - Car | 73.72 Bd ±1.47 | 90.86 Ab ±2.63 | 78.30 Bc ±1.53 | 97.05 Aa ±2.43 |
| GA | 75.30 Bd ±2.34 | 89.76 Ab ±1.18 | 95.60 Aa ±1.30 | 81.94 Cc ±2.66 |
| QUE | 50.85 Cc ±2.56 | 82.44 Ba ±2.45 | 62.55 Db ±4.60 | 83.51 Ca ±1.65 |
| AA | 80.66 Ab ±3.39 | 91.02 Aa ±1.11 | 74.75 Cc ±1.30 | 83.33 Cb ±4.21 |
FXN, fucoxanthin; β – Car, β – carotene; GA, gallic acid; QUE, quercetin; AA, ascorbic acid. The compounds’ concentration was 10 µM. 1Values are mean ± standard deviation of at least three repetitions. Values followed by different uppercase letter in the same row indicate significant differences (p <0.05) between the reaction time tested of each phytochemical compound.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



