Submitted:
03 November 2023
Posted:
03 November 2023
You are already at the latest version
Abstract
Aluminum (Al) toxicity is the most common factor limiting the growth of alfalfa in acidic soil conditions. Melatonin (MT), a significant pleiotropic molecule present in both plants and animals, has shown promise in mitigating Al toxicity in various plant species. This study aims to elucidate the underlying mechanism by which melatonin alleviates Al toxicity in alfalfa through a combined physiological and transcriptomic analysis. The results reveal that 5 μM MT significantly alleviates Al-induced growth inhibition and preserves the morphology of root tip cells. Moreover, MT application reduces Al accumulation in alfalfa roots and shoots by 28.6% and 27.6%, respectively. Additionally, MT plays a crucial role in scavenging Al-induced excess H2O2 by enhancing the activities of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT), consequently reducing malondialdehyde (MDA) levels. The RNA-seq results reveal that MT application significantly upregulates the expression of xyloglucan endotransglucosylase/hydrolase (XTH) and carbon metabolism-related genes, including those involved in the glycolysis process, as well as sucrose and starch metabolism, suggesting that MT application may mitigate Al toxicity by facilitating the binding of Al to the cell walls, thereby reducing intracellular Al accumulation, and by improving respiration and the content of sucrose and trehalose. Taken together, our study demonstrates that MT alleviates Al toxicity in alfalfa by restoring redox homeostasis, enhancing Al binding to the cell wall, and modulating carbon metabolism. This research advances our understanding of the mechanisms underlying MT's effectiveness in mitigating Al toxicity, offering potential strategies for reducing Al accumulation in alfalfa.
Keywords:
Al toxicity
; Melatonin
; Redox homeostasis
; RNA sequencing
; Cell wall
; Carbon metabolism
1. Introduction
It is estimated that approximately 40-50% of the world's arable lands had become acidic by the year 2015, with nearly 60% of these acidic lands concentrated in tropical and subtropical regions, which are the primary grain-producing areas [1]. Recent societal developments have exacerbated soil acidification due to anthropogenic factors such as the indiscriminate application of synthetic fertilizers, accumulation of organic matter, and disruptions in the natural nutrient cycling within the soil [2]. Aluminum (Al), the third most abundant element in the Earth's crust, assumes a paramount role in this effect as it represents the most crucial and widespread factor limiting crop yields in acidic soils. In such acidic environments, aluminum dissolves into a toxic ionic form, notably Al3+, originating from less harmful alumino-silicates or aluminum oxides [3].
Numerous studies have indicated that primary symptoms of Al toxicity in plants include inhibited root growth and biomass, resulting in altered root morphology [4,5]. The root apical transition zone (TZ), located between the meristem and elongation zone, is particularly sensitive to Al exposure [5-9]. Another significant consequence of Al toxicity in plants is the rapid production and accumulation of reactive oxygen species (ROS), such as hydrogen peroxide (H2O2), superoxide (O2·−), and hydroxyl (OH·−) radicals. This ROS buildup leads to membrane lipid peroxidation, intensifying damage to the membrane system, protein degradation, and ultimately programmed cell death [5,10-12]. To counteract the excessive ROS accumulation induced by Al, plants activate an enzyme-mediated antioxidant defense system, which includes superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT) and peroxidases (POD) [13,14]. SOD plays a pivotal role in the oxidative stress response by catalyzing the dismutation of O2·− into oxygen and H2O2, marking the initial step in this process, after which H2O2 is further converted into water through the catalytic actions of APX, CAT and POD [15]. Furthermore, non-enzymatic antioxidants, including proline, ascorbate and glutathione, have been documented to effectively detoxify ROS and enhance Al tolerance [16,17], whose production and activation are integral components of the internal tolerance mechanism that plants employ to combat Al toxicity. In addition to this internal mechanism, plants also combat Al toxicity through intracellular Al chelation and the compartmentalization of Al-organic acid complexes within vacuoles [5,18,19]. Beyond intracellular Alisolation, the Al exclusion mechanisms have received substantial attention in research. This mechanism involves the secretion of organic acids (OAs) as a well-characterized and extensively studied response to Al stress, including citrate, malate and oxalic acids, which serve to chelate Al and form non-toxic complexes [1,5]. Citrate and malate are the main types of OA secreted in many plant species. For example, crops such as rice [20], maize [21] and pea [22] have been shown to release citrate in response to Al stress. Malate secretion has been identified in plants like rape [23] and sorghum [24]. Additionally, some species, including Arabidopsis [25], soybean [26] and barley [27], can secrete both citrate and malate when confronted with Al stress. In contrast, oxalic acid secretion has been detected in buckwheat and taro, with no known transporters identified for its secretion in response to Al toxicity. Numerous experiments have pinpointed the involvement of gene families such as multidrug and toxic compound extrusion (MATE), responsible for encoding citrate transporters on the plasma membrane, and Al-activated malate transporter (ALMT), an anion channel activated by Al3+ on the plasma membrane, in facilitating the release of the respective OAs from plant root cells into the rhizosphere under Al stress [20-27]. However, transporters for oxalic acid secretion in response to Al toxicity remain unidentified. In certain Al-tolerant species like Melastoma malabathricum and Melaleuca cajuputi, besides exuding OAs for Al chelation, they also produce phenolic compounds in their roots that can chelate Al [17]. Hence, the extent of Al tolerance varies among different plant species. Alfalfa (Medicago sativa L.), a widely cultivated forage crop renowned for its nutritional value and high yield capacity, is highly sensitive to Al stress and has been associated with severely curtailing alfalfa productivity in acidic soils [28]. Therefore, it is important to investigate alfalfa's response to Al stress and adopt appropriate strategies to enhance alfalfa yield.
Melatonin (N-acetyl-5-methoxytryptamine) is an indole derivative that was initially discovered in vascular plants in 1995 [29]. It has been identified as a key regulator in various physiological processes, including the regulation of circadian rhythms [30], promotion of seed germination, root development, enhancements in photosynthesis [31], and the postponement of leaf and flower senescence [32]. One of the most extensively characterized and studied roles of melatonin is its ability to enhance plant tolerance to various abiotic stresses. For instance, exogenous melatonin has been shown to improve salt tolerance in sunflower [33] and rice [34], play a crucial role in mitigating the toxicity of heavy metals, and alleviate cadmium toxicity in Chinese cabbage seedlings [35], chromium toxicity in maize [36], and copper toxicity in cucumber [37]. Researchers have also explored the potential alleviating effects of melatonin on Al toxicity. Its application has been found to enhance resistance to Al toxicity by increasing the antioxidant defense system and promoting the exudation of Al-induced malate and citrate in soybean [38] and wheat [39]. Ren et al. [40] demonstrated that melatonin mitigates Al-induced growth inhibition in maize [41] by modulating carbon and nitrogen metabolism and regulating redox homeostasis. However, it remains poorly understood whether melatonin can alleviate the inhibitory effects of Al treatment on alfalfa, and the molecular mechanisms underlying how melatonin mitigates Al-induced growth inhibition in alfalfa have not been adequately elucidated.
In recent years, RNA sequencing (RNA-seq) has emerged as an indispensable tool for transcriptome analysis because it can generate a vast number of reads, providing a comprehensive overview of the transcriptomic landscape. Consequently, RNA sequencing has been applied to investigate Al response mechanisms in various plant species, including Pinus massoniana [42], sugarcane [43], aspen [44], peanut [45] and alfalfa [46], uncovering numerous physiological and metabolic processes involved in Al stress such as oxidative stress, organic acid exudation, defense against cell wall toxicity, and hormone signal transduction. Despite extensive information on Al stress responses, the specific mechanisms via which melatonin regulates Al toxicity in alfalfa remain unexplored. In our study, we integrate physiological and transcriptomic analyses to investigate the regulatory mechanisms through which melatonin might mitigate Al toxicity in alfalfa and provide valuable insights into a theoretical foundation for enhancing alfalfa yield in acidic soil conditions.
2. Results
2.1. Melatonin applications significantly alleviate Al-induced growth inhibition in alfalfa
Previous research has demonstrated that the optimal concentration of exogenous melatonin required to mitigate Al toxicity varies among different plant species. For instance, 1μM exogenous melatonin is most effective for soybeans [38], while 50 μM exogenous melatonin is optimal for maize [41]. To determine the optimal concentration of exogenous melatonin for alleviating Al toxicity in alfalfa, we conducted an experiment in which we observed the growth of alfalfa under different melatonin concentrations (1 μM, 5 μM, 10 μM, or 50 μM) while exposed to 10 μM AlCl3. We assessed root length and fresh weight as indicators of plant growth and found that melatonin significantly mitigates Al toxicity, which was most pronounced at lower melatonin concentrations (1 μM and 5 μM) but diminished at higher concentrations (10 μM and 50 μM) (Supplementary Figure S1). Thus, 5 μM melatonin was selected for further investigation as it exhibited the most effective alleviation of Al toxicity (Figure 1A).
In the absence of melatonin, exposure to 10 μM AlCl3 resulted in a substantial reduction of 41.2% in alfalfa root length and a 33.3% decrease in fresh weight. However, 5 μM melatonin showed a remarkable improvement, with alfalfa exhibiting a 48% increase in root length and a 45.4% increase in fresh weight (Figure 1B, C). Importantly, the primary symptoms of Al toxicity extend beyond the inhibition of root growth and encompass disruptions in root morphology [4,5]. To assess cell morphology, transverse sections of the root tip cells were examined. In the absence of Al stress (control), the root apical cells exhibited a normal rectangular shape and close arrangement. Under the influence of Al stress, the cells became enlarged and assumed irregular shapes with a loose arrangement. However, melatonin applications alleviated this cell damage. The cells treated with melatonin appeared smaller than those exposed to Al alone, exhibited a more normal shape and had a relatively compact arrangement-conditions conducive to promoting root growth (Figure 1D). These results collectively demonstrate that the application of exogenous melatonin effectively alleviates Al-induced growth inhibition and preserves the morphology of root tip cells in alfalfa.
2.2. Melatonin could reduce the accumulation of Al in alfalfa roots under Al stress
To investigate the mechanism of how melatonin alleviated Al toxicity in alfalfa, the Al content of alfalfa seedings treated with and without melatonin for 3 days was qualitatively and quantitatively analyzed. Morin staining, a sensitive fluorescent dye for Al, was used to visualize Al presence in the roots, which showed that Al treatment significantly increased green fluorescence intensity, indicating extensive Al accumulation in roots. However, when alfalfa was exposed to both Al and melatonin, the strong green fluorescence was barely detectable (Figure 2A). These findings were further confirmed by quantitative analysis of Al content. Furthermore, melatonin treatment reduced Al accumulation in alfalfa roots and shoots by 28.6% and 27.6%, respectively, compared to Al treatment alone (Figure 2B, C). These results confirm that melatonin mitigates Al toxicity in alfalfa by reducing Al content.
2.3. Melatonin could alleviate Al-induced oxidative damage in root apex cells
Melatonin is well-known for its ability to enhance plant resistance to various abiotic stresses, including heavy metal stress [35-37], drought [41] and salt stress [34], primarily due to its potent free radical scavenging capabilities. Therefore, we sought to validate the mitigating effects of melatonin on cellular oxidative damage by assessing H2O2 levels, Malondialdehyde (MDA) content and cell vitality, which are indicative of Al-induced oxidative damage. Following Al treatment, the H2O2 content in alfalfa roots increased by 22.6%. However, the application of melatonin significantly reduced Al-induced H2O2 levels by 15.7%, although they did not return to control levels (Figure 3B). Furthermore, it is known that the accumulation of free radicals in plants can result in membrane lipid peroxidation. MDA, a final product of lipid peroxidation, is positively correlated with the extent of this peroxidation. Al treatment can lead to an approximately 1.6-fold increase in MDA content in roots compared to the untreated control. Nevertheless, the addition of melatonin to Al-treated plants reduced MDA accumulation, indicating that melatonin applications could mitigate Al-induced lipid peroxidation in alfalfa roots (Figure 3C). Additionally, we evaluated root apical cell vitality using Evans blue staining, which accumulates in dying and dead cells. In alfalfa treated with Al alone, root tip cells displayed extensive blue patches, indicative of a large number of dying or dead cells. However, simultaneous treatment with Al and melatonin significantly reduced the number of dead cells, resulting in reduced blue staining (Figure 3A). These findings collectively suggest that melatonin can partially alleviate Al-induced oxidative damage in alfalfa root tip cells by reducing H2O2 levels, mitigating lipid peroxidation, and preserving membrane integrity.
2.4. Melatonin may enhance the activity of antioxidant enzymes
The primary strategy for mitigating oxidative damage involves the elimination of ROS through the action of antioxidant enzymes. We hypothesized that melatonin could enhance the activity of these antioxidant enzymes, thus maintaining ROS homeostasis. Thus, we assessed the activities of SOD, POD, CAT and APX. The results showed that compared to the control, Al treatment alone had minimal impact on SOD activity but reduced POD activity by 22.8%. Furthermore, it significantly inhibited the activities of CAT and APX in the roots. This inhibition of POD, CAT and APX activities led to the accumulation of H2O2, which could not be effectively catalyzed into H2O and O2, consistent with our previous findings (Figure 3). Conversely, melatonin application, when compared to Al treatment alone, resulted in a substantial increase in the activities of SOD (approximately 1.58-fold), POD (approximately 1.91-fold), CAT (approximately 5.27-fold), and APX (approximately 5.27-fold), indicating that melatonin application could significantly enhance the ROS clearance capacity of alfalfa. Moreover, there was no significant difference in the activity of these enzymes between the control and melatonin-treated plants in the roots (Figure 4). Overall, these data suggest that melatonin application can effectively maintain ROS homeostasis by increasing the activity of antioxidant enzymes in alfalfa.
2.5. Treatments with Al and melatonin trigger transcriptome reprogramming in alfalfa roots
To explore the mechanisms underlying melatonin's alleviation of Al toxicity, we conducted transcriptome analysis on alfalfa roots exposed to Al and Al + MT (melatonin) treatments, each with three biological replicates. Control (CK) and MT treatments were performed as experimental controls. Principal component analysis (PCA) and a heatmap of sample correlations (Figure 5A, B) indicated strong correlations among the biological replicates, affirming data accuracy and reliability. Then, we applied statistical analysis with criteria of |log2fold change| > 1 and padj < 0.05 to identify DEGs in each comparison, which showed that when comparing Al vs. CK, 993 DEGs were identified, comprising 422 upregulated and 571 downregulated genes. In the Al + MT vs. Al comparison, 1,937 DEGs were detected, with 1,000 upregulated and 937 downregulated genes (Figure 5C). The Venn diagram shows the number of DEGs among different treatment comparisons and highlights common and unique DEGs in each comparison. The results indicate that upon comparing Al + MT vs. Al and Al vs. CK, 265 DEGs were found to be shared between them. Furthermore, the Al + MT vs. Al comparison revealed 1,672 DEGs unique to it, while the Al vs. CK comparison had 728 exclusive DEGs (Figure 5D). These findings suggest that the addition of melatonin elicited the differential expression of a broader set of genes in response to Al stress.
2.6. Gene Ontology (GO) classification and KEGG pathway analysis.
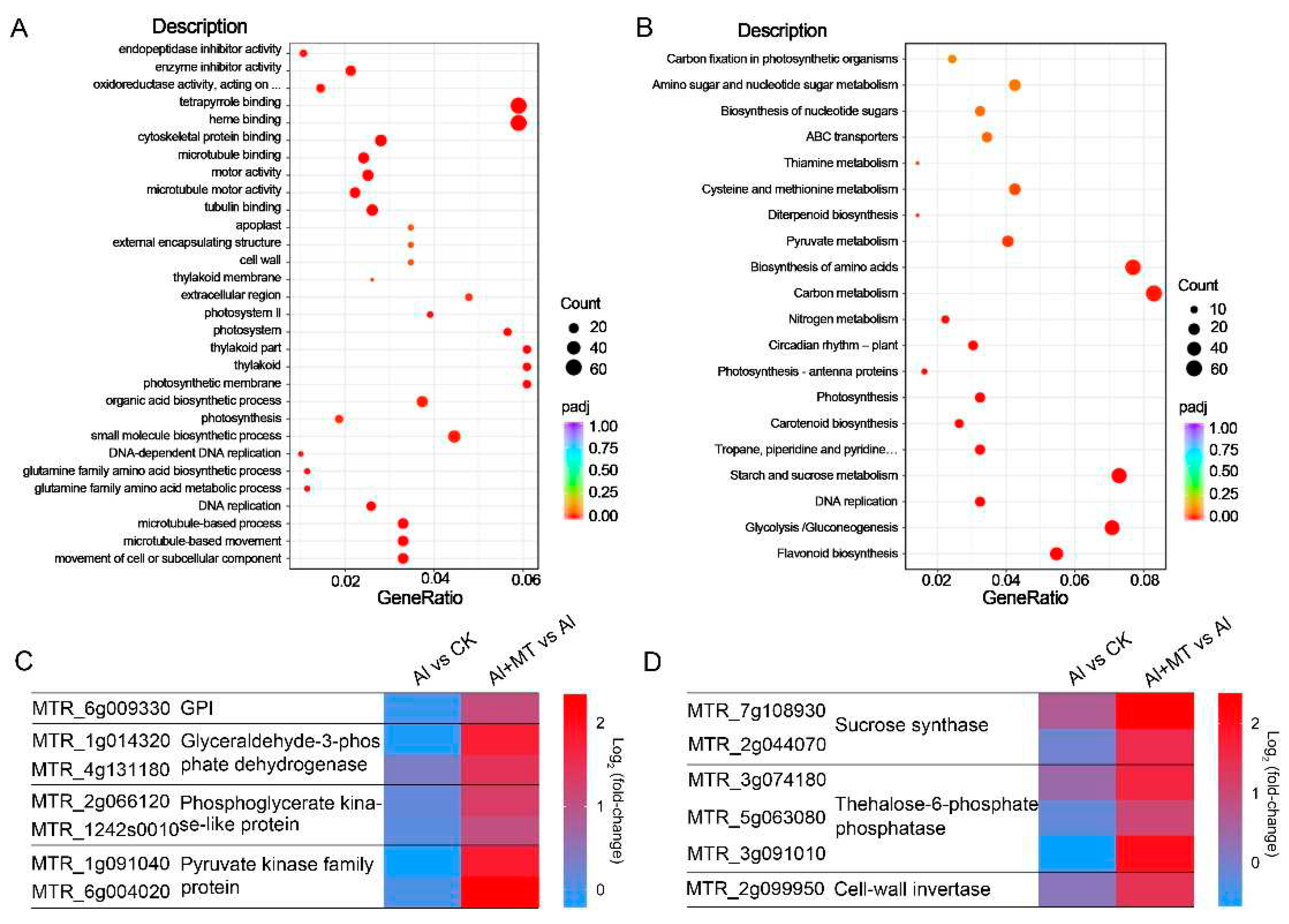
To determine the functions of these DEGs, we conducted gene ontology (GO) enrichment analysis with a threshold of padj < 0.05. In the Al + MT vs. Al comparisons, the GO analysis categorized the DEGs into three domains: 380 Biological Process (BP) categories, 82 Cellular Component (CC) categories, and 261 Molecular Function (MF) categories. In the BP category, the DEGs were primarily associated with processes such as organic acid biosynthetic processes, cell wall organization or biogenesis, and the movement of cells or subcellular components. Concerning CC, a substantial proportion of DEGs were linked to the extracellular region and cell walls. In the MF category, the DEGs were predominantly related to activities involving oxidoreductase and microtubule motor functions (Figure 6A). These results suggest that the supplementation of exogenous melatonin might alleviate Al toxicity by not only mitigating Al-induced oxidative damage but also potentially regulating processes such as organic acid synthesis, cell wall organization, and cellular movement and remodeling as mechanisms for mitigating Al toxicity. Additionally, we conducted Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis to identify metabolic pathways associated with the effects of melatonin on Al stress in alfalfa. In the Al + MT vs. Al comparisons, the KEGG enrichment analysis revealed that these DEGs were predominantly enriched in metabolic pathways such as carbon metabolism, glycolysis/gluconeogenesis, and starch and sucrose metabolism (Figure 6B).
In the carbon metabolism and glycolysis/gluconeogenesis, most DEGs that were associated with glycolysis exhibited notable upregulation in Al + MT-treated plants (Figure 6C). These upregulated genes included pyruvate kinase, glyceraldehyde-3-phosphate dehydrogenase, phosphopyruvate hydratase, and 6-phosphofructokinase, confirming that melatonin supplementation enhances carbon metabolism in alfalfa roots under Al stress. In regard to pathways related to starch and sucrose metabolism, numerous DEGs annotated as sucrose synthase, trehalose-6-phosphate phosphatase, and cell-wall invertase showed significant upregulation in the Al + MT vs. Al comparison (Figure 6D). These results suggest that the application of exogenous melatonin may boost the synthesis capacity of sucrose and trehalose in alfalfa, thereby enhancing their stress resistance under Al stress.
3. Discussion
In acidic soils, Al stress can significantly hinder plant growth, resulting in reduced crop yields, particularly in Al-sensitive crops such as alfalfa and soybean. Melatonin, an endogenous molecule with high conservation across eukaryotic evolution, plays a pivotal role as a signaling molecule in various biological processes and response to abiotic stress in plants. These stresses include salt stress [33,34], drought stress [47], and heavy metal stress in certain species [35-37]. Recent studies have also revealed that melatonin mitigates Al toxicity in soybean [38], wheat [39] and maize [40,41] through diverse mechanisms. Our present study mainly investigated the mechanisms underlying the application of exogenous melatonin in alleviating Al stress in alfalfa.
3.1. Exogenous melatonin reduced Al concentration in both alfalfa roots and shoots.
Zhang et al. [38] previously demonstrated that melatonin alleviated Al toxicity in soybeans by enhancing the exudation of malate and citrate to chelate aluminum. In wheat, melatonin modified the polysaccharide content of cell walls, thereby reducing Al accumulation in the roots [39]. In line with these findings, our study showed that the application of exogenous melatonin significantly reduced Al content in both alfalfa roots and shoots under aluminum stress (Figure 2). Additionally, melatonin mitigated the growth inhibition of alfalfa induced by aluminum treatment (Figure 1). Next, we investigated whether melatonin reduced aluminum accumulation through the above-mentioned mechanisms. Firstly, our analysis of citrate content in the rhizosphere indicated no significant differences in the presence or absence of melatonin under aluminum treatment (results not shown). Furthermore, our transcriptome sequencing results revealed that the transcription levels of citrate transporters were not upregulated following the addition of melatonin (results not shown). These findings collectively suggest that melatonin activated alternative exclusion mechanisms, rather than citrate secretion, to reduce Al accumulation in alfalfa.
The cell wall, a dynamic architectural structure primarily comprising cellulose, hemicellulose and pectin, plays a crucial role in sensing and responding to Al toxicity [1]. When plant roots are exposed to Al, the cell wall, serving as the initial point of contact, acts as a significant reservoir for Al. For instance, nearly 85% of the total Al accumulates in the cell wall of maize roots [48], while more than 70% of Al has been detected in the cell wall of wheat [39]. Among the cell wall components, pectic polysaccharides and hemicellulose are recognized as major binding sites for aluminum, with a specific emphasis on xyloglucan in hemicellulose, which has been found to be particularly sensitive to Al stress in recent studies [49,50]. Furthermore, research has revealed that one of the primary reasons for aluminum-induced inhibition of root growth is its binding to the cell wall, resulting in increased rigidity, reduced cell expansion, and diminished mechanical extensibility of the cell wall. This alteration has detrimental effects on the structure and function of the cell wall [51,52]. Theoretically, xyloglucan also plays a pivotal role in cell wall extensibility. The process of cell wall extension, which involves the cleavage of the xyloglucan backbone followed by the synthesis of a newly formed xyloglucan chain, is catalyzed by enzymes from the xyloglucan endotransglucosylase/hydrolase (XTH) family [53,54]. Our transcriptome sequencing results revealed a significant increase in the transcription levels of XTH upon the application of melatonin under Al stress (Supplementary Figure S2), suggesting that melatonin may upregulate the activity of XTH, which in turn regulates the content of xyloglucan, a hemicellulose capable of binding Al to reduce intracellular Al accumulation. Moreover, the alleviation of Al-induced growth inhibition in alfalfa by melatonin may be attributed to the modulation of XTH activity, thereby restoring cell wall extensibility that is otherwise reduced by Al.
3.2. Exogenous melatonin enhanced the antioxidant capacity in alfalfa.
Under normal growth conditions in alfalfa, a dynamic equilibrium exists between the generation and elimination of ROS. However, Al toxicity can disrupt this balance, leading to the excessive accumulation of ROS, which in turn, can interact with lipids and proteins, causing protein degradation and lipid peroxidation, ultimately compromising the integrity of the cell membrane [10-12]. Melatonin, widely known as an important animal hormone, primarily participates in regulating circadian rhythms, improving sleep quality and combating free radicals to delay the aging process [55]. It is a highly conserved indole molecule with similar physiological functions to indole-3-acetic acid in plants, which include involvement in plant root growth, promotion of seed germination, and increasing crop yield [56]. Furthermore, numerous studies have confirmed that melatonin enhances the stress tolerance of plants. For instance, it can improve salt tolerance in rice [34], mitigate cadmium stress in Chinese cabbage [35], alleviate chromium stress in maize [36], and combat aluminum stress in soybean [38] and wheat [39], owing to its potent antioxidant capacity. In this study, we observed a significant increase in the H2O2 levels in alfalfa roots under Al stress compared to the control. The increased content of MDA, an indicator of membrane lipid peroxidation, and together with Evans staining under Al stress indicated that ROS accumulation induced by Al led to membrane superoxidation and the disruption of membrane integrity (Figure 3). However, the use of melatonin significantly mitigated these adverse effects induced by Al, highlighting the crucial role of melatonin in scavenging ROS, alleviating membrane superoxidation, and preserving membrane integrity (Figure 3).
Plants have developed both enzymatic and non-enzymatic antioxidant systems to counteract the effects of ROS. Vital antioxidant enzymes include SOD, POD, CAT and APX, while glutathione plays a crucial role as a non-enzymatic antioxidant in plants [57]. Our findings revealed that Al treatment had a minor impact on the activity of SOD but significantly reduced the activities of POD, CAT and APX (Figure 4), indicating that Al stress reduced the activities of catalases responsible for H2O2 breakdown, resulting in the accumulation of H2O2 and disrupting the balance of the oxidative defense system. However, the application of melatonin significantly enhanced the activities of SOD, POD, CAT and APX under Al exposure (Figure 4). These results suggest that melatonin can alleviate the imbalance in the oxidative defense system caused by Al by enhancing the activities of these enzymatic antioxidants.
3.3. Exogenous melatonin elevated C metabolism under Al stress in alfalfa.
The process of carbon metabolism in plants involves photosynthesis and respiration, which provide carbon compounds and serve as an energy source for various biological activities, respectively [58]. Previous studies have demonstrated that melatonin can enhance photosynthesis in various plant species under different abiotic stresses. For instance, melatonin application has been shown to improve photosynthesis in rice under salt stress [34], in tobacco under cadmium stress [59], and in maize under Al stress [60]. Given the characteristic root growth inhibition and morphological disruption observed in many plants under Al stress [4,5], we conducted a transcriptome sequencing analysis of alfalfa root tips. The results of RNA-Seq analysis revealed that exogenous melatonin application significantly increased the transcript levels of enzymes related to glycolysis and gluconeogenesis in alfalfa root tips under Al stress. These enzymes include pyruvate kinase, glyceraldehyde-3-phosphate dehydrogenase, phosphopyruvate hydratase, and 6-phosphofructokinase (Figure 7A). These findings suggest that melatonin not only enhances photosynthesis in plant leaves but also improves respiration in alfalfa roots, leading to increased energy production, which can then be utilized for plant growth, thereby promoting the growth of plants exposed to Al stress.
Plant growth and carbohydrate metabolism, including sucrose and starch, are closely intertwined, as carbohydrates serve as both structural components and energy reservoirs for biomass synthesis and maintenance [61]. In higher plants, leaves (sources) are responsible for synthesizing sucrose and starch, which are subsequently transported to roots (sinks) to support root growth [61]. For instance, Zhao et al. [62] found that foliar application of melatonin significantly increased starch and sucrose synthesis in maize leaves under drought stress. Additionally, it has been reported that melatonin application could enhance the activity of sucrose phosphate synthase (SPS) and sucrose content in maize under Al stress [60]. In this present study, RNA-seq results of alfalfa roots revealed significant alterations in the starch and sucrose metabolism pathways following the addition of melatonin under Al stress (Figure 7B). Many DEGs were annotated as sucrose synthase and trehalose-6-phosphate phosphatase, and they were markedly upregulated in the Al + MT vs. Al comparison. The increase in sucrose synthase transcript levels suggests an elevated sucrose content in alfalfa roots, indicating that melatonin may allocate more carbohydrates to the roots under Al stress, a role consistent with its function in maize under Al stress [60]. Trehalose-6-phosphate phosphatase catalyzes the conversion of trehalose-6-phosphate to trehalose, a compound known to protect cells from various stresses and adversities in many organisms [63-65]. Therefore, based on the RNA-Seq results, the exogenous application of melatonin may enhance the synthesis capacity of sucrose and trehalose in alfalfa, thereby improving stress resistance under Al stress.
4. Materials and Methods
4.1. Plant culture and measurements of Al treatments.
Seeds of alfalfa were from forage Engineering Center of Southwest University. To initiate germination, 5-6 alfalfa seeds were placed on individual moist cotton balls in a plastic container filled with 6 liters of distilled water. These containers were then kept in a greenhouse under controlled conditions: 22°C temperature, 100μmol m-2s-1 white light intensity, and a photoperiod of 16 hours of light and 8 hours of darkness for one day. After three days, when two cotyledons had emerged, healthy seedlings were carefully selected to ensure that each cotton ball retained four alfalfa seedlings with uniform growth. These 4-day-old seedlings were then subjected to treatment with a 10μM AlCl3 solution containing 0.5mM CaCl2 for either 3 or 5 days at a pH of 4.2, as previously described [38]. Control groups included untreated seedlings and those treated with only 5μM melatonin, both maintained in a 0.5mM CaCl2 solution at pH 4.2. To maintain the pH at 4.2, the treatment solution was renewed daily. After the treatment period, the alfalfa seedlings were divided into two groups. The first group was used for measuring root length and fresh weight, with each treatment having four biological replicates, each containing four seedlings. The root tips from this group were used for cell anatomical observations. The second group of seedlings was separated into root and shoot sections, quickly frozen in liquid nitrogen, and stored at -80°C for subsequent analyses, including transcriptome sequencing, Al content determination, oxidative damage indicator assessment, and enzyme activity assays.
4.2. Anatomical observation of root tip cells
The root tips of alfalfa seedlings, which had been treated for 3 days, were fixed in 70% FAA fixative. After fixation, they were dehydrated using a series of alcohol concentrations and embedded in paraffin. The paraffin-embedded blocks were then sectioned into 5μm slices using a microtome. To prepare the sections for analysis, paraffin was removed with xylene and ethanol, and the slices were stained with safranin for 1 hour, followed by rinsing with water. A brief immersion in fast green staining solution for 2 minutes followed. After dehydration with anhydrous ethanol and transparency achieved with xylene, the root tip slice images were examined under a microscope in brightfield conditions.
4.3. Moring staining
Morin staining was used to assess Al accumulation, following the procedure outlined by Zhang et al. [38]. After applying various treatments, approximately 2 cm-long fresh alfalfa roots were cut and immersed in 100μM morin for 1 hour at room temperature. Subsequently, the roots were washed with water to remove any excess morin. The Al-morin complex exhibited green fluorescence when excited at a wavelength of 420 nm, and the emitted light was observed at a wavelength of 510 nm.
4.4. Determination of Al content
The concentrations of Al in the leaves and roots (0-1 cm) were determined using a graphite furnace atomic absorption spectrophotometer (Agilent Technologies, GTA 120 Graphite Tube Atomizer), following the method described by Riaz et al. [66]. After the respective treatments, the alfalfa shoots and root segments (0-1 cm) were excised after three washes with distilled water. Approximately 1 g of each sample was weighed into a graphite crucible and subjected to ashing at 500°C for 2 hours to completely ash them. Subsequently, 4 g of sodium hydroxide and 1 g of sodium peroxide were added, and the mixture was melted in a muffle furnace at 650°C for 15 minutes. The mixture was then heated with 50 ml of HCl to a slight boil and diluted to a final volume of 250 ml. Finally, an Al reagent was added, and the volume was adjusted to 50 ml. The spectrophotometer measured the absorbance at 530 nm. By comparing the standard curve generated using Al standard solutions of varying concentrations, the total Al content of the sample was calculated.
4.5. Determination of H2O2 content
The H2O2 content in root tips was determined by measuring the formation of the titanium-peroxide complex at 415 nm using an enzyme-linked immunosorbent assay (ELISA) instrument (TECAN SPARK, Austria) [39]. Briefly, we added 1 mL of acetone to 0.1 g of tissue. The mixture was then homogenized in an ice bath and centrifuged, after which 2.0 mL of a solution containing 20% sulfuric acid (v/v) and 0.1% titanium tetrachloride (v/v) was added to the 200 μL supernatant. The supernatants obtained after centrifugation were analyzed at 415 nm using the ELISA instrument (TECAN SPARK, Austria). The H2O2 concentration in the samples was determined by comparing the absorbance values to a standard curve of known H2O2 concentrations.
4.6. Analysis of oxidative damage
To assess lipid peroxidation, the levels of 2-thiobarbituric acid-reactive metabolites (TBA) were measured, with these TBA-reactive substances quantified as malondialdehyde (MDA), which serves as an end product of lipid peroxidation [67].
Firstly, 1 mL of extraction solution was added to approximately 0.1 g of root tips. The mixture was homogenized in an ice bath and then centrifuged at 4℃ and 12,000 rpm for 10 minutes. Next, 300 μL of a reaction solution containing 0.5% thiobarbituric acid (TBA) was added to 200 μL of the supernatant, followed by incubation at 95℃ for 30 minutes. After centrifugation, the MDA content was determined and calculated by measuring the absorbance at 532 nm and 600 nm using an ELISA instrument (TECAN SPARK, Austria). The integrity of the plasma membrane was assessed using a 0.25% Evans blue solution, which is capable of entering dead cells [67]. The root apices were immersed in a solution of 0.25% Evans blue dye for 20 minutes and then thoroughly washed with water 3-5 times. Finally, the root tips were examined using a microscope under brightfield conditions.
4.7. Determination of antioxidant enzyme activity
For enzyme activity assays, approximately 0.1 g of frozen root samples were homogenized and extracted in PBS buffer, followed by centrifugation at 4℃ and 12,000 rpm for 10 minutes. Subsequently, the supernatant was collected via pipetting for enzyme activity determination. The activity of superoxide dismutase (SOD; EC 1.15.1.1) was assessed using the total superoxide dismutase assay kit with WST-8 (Geruisi, China) [68]. In the coupled reaction system of WST-8 and xanthine oxidase (XO), the SOD enzyme activity in the reaction system was defined as one unit of enzyme activity (U/mL) when the inhibition percentage reached 50%. The activity of peroxidase (POD; EC 1.11.1.7) was determined by monitoring the oxidation of guaiacol upon the addition of H2O2 [39]. One unit (U) of POD activity was defined as the increase of 0.5 in absorbance at 470 nm per minute per gram of root sample in the reaction system. The activity of catalase (CAT; EC 1.11.1.6) was measured by observing the decrease in absorbance at 240 nm, reflecting the reduction in H2O2 concentration [67]. One unit (U) of CAT activity was defined as the degradation of 1 μmol H2O2 per minute per gram of root sample. The activity of ascorbate peroxidase (APX; EC 1.11.1.11) was quantified by measuring the decrease in absorbance at 290 nm, which resulted from the oxidation of ascorbic acid by the complex of APX and H2O2 [66]. The unit (U) of APX activity was defined as the oxidation of 1μmol of ascorbate per minute per gram of root sample.
4.8. RNA extraction and transcriptome sequencing
After 3 days of treatment with Al alone or in combination with melatonin, we collected 2 cm of alfalfa roots for transcriptome sequencing. The groups treated only with acid and only with melatonin were used as controls. Each treatment was replicated three times. Total RNA from each sample was extracted using the Polysaccharide Polyphenol RNA Extraction Kit (QIAGEN, Germany) following the manufacturer's instructions. The concentration and integrity of RNA were assessed using the Nanodrop spectrophotometer (Thermo Scientific, USA) and the 2100 bioanalyzer (Agilent Technologies, Santa Clara, USA), respectively. Samples with RNA integrity >8 and OD260/280 >8 were selected for library construction [69]. Subsequently, the library was constructed using the NEBNext Ultra RNA Library Prep Kit for Illumina (NEB, Ipswich, USA). The concentration and insert size of the library were detected using the Qubit 2.0 Fluorometer and the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, USA), respectively. Once the concentration and insert size met the expected range, Q-PCR was performed to accurately quantify the effective concentration of the library, ensuring that the library's effective concentration was higher than 2 nM to ensure library quality. Finally, the libraries were sequenced using the Illumina HiSeq platform (Novogene, Beijing, China) [60].
5. Conclusions
In this study, we conducted comprehensive physiological and transcriptomic analysis to determine the mechanisms underlying melatonin's ability to alleviate Al toxicity in alfalfa. Our results demonstrated that melatonin could effectively mitigate the negative effects of Al on alfalfa growth, including growth inhibition and damage to root tip cells, which was associated with a reduction in Al accumulation in both root and shoot tissues. Additionally, melatonin played a role in scavenging excess H2O2, reducing lipid peroxidation as indicated by decreased MDA levels, and enhancing the activities of antioxidant enzymes such as SOD, POD and CAT. Our RNA-seq data revealed that melatonin treatment upregulated the expression of genes involved in key processes such as XTH, glycolysis and sucrose and starch metabolism, suggesting that melatonin could enhance the binding of aluminum to the cell wall, improve respiration and increase the content of sucrose and trehalose. These molecular and physiological changes collectively contribute to the alleviation of Al toxicity in alfalfa. Taken together, our research provides valuable insights into the mechanisms through which melatonin mediates aluminum stress in alfalfa, and the findings hold the potential to inform strategies for improving alfalfa production in acidic soils, which could benefit agriculture and forage production.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Effects of different concentrations of melatonin on the phenotype of alfalfa. Figure S2: Transcript changes in cell wall-related genes induced by Al and melatonin.
Author Contributions
Project administration and supervision, D.Y., Y.W.; funding acquisition, Y.W.; investigation, C.L., H.C.; S.W. and Y.W.; data curation, C.L., H.C., D.Y. and Y.W.; writing—original draft, C.L.; writing—review and editing, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Henan province (222300420395).
Institutional Review Board Statement
Ethical review and approval were waived for this study, because it did not involve humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J. V Plant Adaptation to Acid Soils: The Molecular Basis for Crop Aluminum Resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Sade, H.; Meriga, B.; Surapu, V.; Gadi, J.; Sunita, M.S.L.; Suravajhala, P.; Kavi Kishor, P.B. Toxicity and Tolerance of Aluminum in Plants: Tailoring Plants to Suit to Acid Soils. Biometals 2016, 29, 187–210. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, C.; Gunsé, B.; Corrales, I.; Barceló, J. A Glance into Aluminum Toxicity and Resistance in Plants. Sci. Total Environ. 2008, 400, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yan, X.; Han, X.; Tang, R.; Chu, M.; Yang, Y.; Yang, Y.H.; Zhao, F.; Fu, A.; Luan, S.; et al. A Defective Vacuolar Proton Pump Enhances Aluminum Tolerance by Reducing Vacuole Sequestration of Organic Acids. Plant Physiol. 2019, 181, 743–761. [Google Scholar] [CrossRef] [PubMed]
- Ofoe, R.; Thomas, R.H.; Asiedu, S.K.; Wang-Pruski, G.; Fofana, B. Abbey, Lord Aluminum in Plant: Benefits, Toxicity and Tolerance Mechanisms. Front. Plant Sci. 2022, 13, 1085998. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.B.; Horst, W.J. Aluminum-Induced Inhibition of Root Growth: Roles of Cell Wall Assembly, Structure, and Function. 2015, 253-274.
- Zhu, X.F.; Wan, J.X.; Wu, Q.; Zhao, X.S.; Zheng, S.J.; Shen, R.F. PARVUS Affects Aluminum Sensitivity by Modulating the Structure of Glucuronoxylan in Arabidopsis Thaliana. Plant. Cell Environ. 2017, 40, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Cao, X.C.; Zhu, L.F.; Hu, W.J.; Hu, A.Y.; Abliz, B.; Bai, Z.G.; Huang, J.; Liang, Q.D.; Sajid, H.; et al. Boron Reduces Cell Wall Aluminum Content in Rice (Oryza Sativa) Roots by Decreasing H2O2 Accumulation. Plant Physiol. Biochem. 2019, 138, 80–90. [Google Scholar]
- Wu, Q.; Tao, Y.; Huang, J.; Liu, Y.S.; Yang, X.Z.; Jing, H.K.; Shen, R.F.; Zhu, X.F. The MYB Transcription Factor MYB103 Acts Upstream of TRICHOME BIREFRINGENCE-LIKE27 in Regulating Aluminum Sensitivity by Modulating the O-Acetylation Level of Cell Wall Xyloglucan in Arabidopsis Thaliana. Plant, J. 2022, 111, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Rabe, S.; Lanzoni, M.; Adriana, R.; Martinelli, P.; Antunes, R. Aluminum-Induced Stress Differently Modifies Urochloa Genotypes Responses on Growth and Regrowth: Root-to-Shoot Al-Translocation and Oxidative Stress. 2018, 141-152.
- Huang, W.; Yang, X.; Yao, S.; LwinOo, T.; He, H.; Wang, A.; Li, C.; He, L. Reactive Oxygen Species Burst Induced by Aluminum Stress Triggers Mitochondria-Dependent Programmed Cell Death in Peanut Root Tip Cells. Plant Physiol. Biochem. 2014, 82, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y. Aluminum Toxicity in Plant Cells: Mechanisms of Cell Death and Inhibition of Cell Elongation. Soil Sci. Plant Nutr. 2019, 65, 41–55. [Google Scholar] [CrossRef]
- Awasthi, J.P.; Saha, B.; Panigrahi, J.; Yanase, E.; Koyama, H.; Panda, S.K. Redox Balance, Metabolic Fingerprint and Physiological Characterization in Contrasting North East Indian Rice for Aluminum Stress Tolerance. Sci. Rep. 2019, 9, 8681. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Yang, Z.M. Nitric Oxide Reduces Aluminum Toxicity by Preventing Oxidative Stress in the Roots of Cassia Tora, L. Plant Cell Physiol. 2005, 46, 1915–1923. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Du, H.; Huang, Y.; Qu, M.; Li, Y.; Hu, X.; Yang, W.; Li, H.; He, W.; Ding, J.; Liu, C.; et al. A Maize ZmAT6 Gene Confers Aluminum Tolerance via Reactive Oxygen Species Scavenging. Front. Plant Sci. 2020, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Maejima, E.; Osaki, M.; Wagatsuma, T.; Watanabe, T. Contribution of Constitutive Characteristics of Lipids and Phenolics in Roots of Tree Species in Myrtales to Aluminum Tolerance. Physiol. Plant. 2017, 160, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Furlan, F.; Borgo, L.; Rabêlo, F.H.S.; Rossi, M.L.; Linhares, F.S.; Martinelli, A.P.; Azevedo, R.A.; Lavres, J. Aluminum-Induced Toxicity in Urochloa Brizantha Genotypes: A First Glance into Root Al-Apoplastic and -Symplastic Compartmentation, Al-Translocation and Antioxidant Performance. Chemosphere 2020, 243, 125362. [Google Scholar] [CrossRef] [PubMed]
- Bojórquez-Quintal, E.; Escalante-Magaña, C.; Echevarría-Machado, I.; Martínez-Estévez, M. Aluminum, a Friend or Foe of Higher Plants in Acid Soils. Front. Plant Sci. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Su, Q.; Wu, Z.; Huang, Z.; Bao, J.; Li, J.; Tu, H.; Zeng, C.; Fu, J.; He, H. Genome-Wide Characterization of MATE Gene Family and Expression Profiles in Response to Abiotic Stresses in Rice (Oryza Sativa). BMC Ecol. Evol. 2021, 21, 141. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Ryan, P.R.; Liu, C.; Li, H.; Hu, W.; Yan, W.; Huang, Y.; He, W.; Luo, B.; Zhang, X.; et al. ZmMATE6 from Maize Encodes a Citrate Transporter That Enhances Aluminum Tolerance in Transgenic Arabidopsis Thaliana. Plant Sci. 2021, 311, 111016. [Google Scholar] [CrossRef] [PubMed]
- Kichigina, N.E.; Puhalsky, J.V.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Loskutov, S.I.; Safronova, V.I.; Tikhonovich, I.A.; Vishnyakova, M.A.; Semenova, E.V.; et al. Aluminum Exclusion from Root Zone and Maintenance of Nutrient Uptake Are Principal Mechanisms of Al Tolerance in Pisum Sativum, L. Physiol. Mol. Biol. Plants 2017, 23, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Ligaba, A.; Katsuhara, M.; Ryan, P.R.; Shibasaka, M.; Matsumoto, H. The BnALMT1 and BnALMT2 Genes from Rape Encode Aluminum-Activated Malate Transporters That Enhance the Aluminum Resistance of Plant Cells. Plant Physiol. 2006, 142, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Melo, J.O.; Martins, L.G.C.; Barros, B.A.; Pimenta, M.R.; Lana, U.G.P.; Duarte, C.E.M.; Pastina, M.M.; Guimaraes, C.T.; Schaffert, R.E.; Kochian, L. V.; et al. Repeat Variants for the SbMATE Transporter Protect Sorghum Roots from Aluminum Toxicity by Transcriptional Interplay in Cis and Trans. Proc. Natl. Acad. Sci. USA. 2019, 116, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Magalhaes, J.V.; Shaff, J.; Kochian, L. V Aluminum-Activated Citrate and Malate Transporters from the MATE and ALMT Families Function Independently to Confer Arabidopsis Aluminum Tolerance. Plant, J. 2009, 57, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, Y.; Wang, W.; Gai, J.; Li, Y. Genome-Wide Analysis of MATE Transporters and Expression Patterns of a Subgroup of MATE Genes in Response to Aluminum Toxicity in Soybean. BMC Genomics 2016, 17, 223. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Ryan, P.R.; Richardson, A.E.; Tyerman, S.D.; Ramesh, S.; Hebb, D.M.; Howitt, S.M.; Delhaize, E. HvALMT1 from Barley Is Involved in the Transport of Organic Anions. J. Exp. Bot. 2010, 61, 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Lv, A.; Wen, W.; Fan, N.; Li, J.; Gao, L.; Zhou, P.; An, Y. MsMYB741 Is Involved in Alfalfa Resistance to Aluminum Stress by Regulating Flavonoid Biosynthesis. Plant, J. 2022, 112, 756–771. [Google Scholar] [CrossRef] [PubMed]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by High Performance Liquid Chromatography-Mass Spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin as a Regulatory Hub of Plant Hormone Levels and Action in Stress Situations. Plant Biol. (Stuttg). 2021, 23 Suppl 1, 7–19. [Google Scholar] [CrossRef]
- MB, A.; J, H.R. Protective Effect of Melatonin against Chlorophyll Degradation during the Senescence of Barley Leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in Flowering, Fruit Set and Fruit Ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; David, A.; Yadav, S.; Baluška, F.; Bhatla, S.C. Salt Stress-Induced Seedling Growth Inhibition Coincides with Differential Distribution of Serotonin and Melatonin in Sunflower Seedling Roots and Cotyledons. Physiol. Plant. 2014, 152, 714–728. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhang, J.; Li, W.; Ding, Y.; Zhong, Q.; Xu, X.; Wei, H.; Li, G. Exogenous Melatonin Alleviates Salt Stress by Improving Leaf Photosynthesis in Rice Seedlings. Plant Physiol. Biochem. 2021, 163, 367–375. [Google Scholar] [CrossRef]
- Wang, T.; Song, J.; Liu, Z.; Liu, Z.; Cui, J. Melatonin Alleviates Cadmium Toxicity by Reducing Nitric Oxide Accumulation and IRT1 Expression in Chinese Cabbage Seedlings. Environ. Sci. Pollut. Res. Int. 2021, 28, 15394–15405. [Google Scholar] [CrossRef]
- Yang, X.; Ren, J.; Lin, X.; Yang, Z.; Deng, X.; Ke, Q. Melatonin Alleviates Chromium Toxicity in Maize by Modulation of Cell Wall Polysaccharides Biosynthesis, Glutathione Metabolism, and Antioxidant Capacity. Int. J. Mol. Sci. 2023, 24, 3816. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.Y.; Qi, C.D.; Li, S.; Wang, Z.; Wang, X.; Wang, J.; Ren, S.; Li, X.; Zhang, N.; Guo, Y.D. Melatonin Alleviates Copper Toxicity via Improving Copper Sequestration and ROS Scavenging in Cucumber. Plant Cell Physiol. 2019, 60, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zeng, B.; Mao, Y.; Kong, X.; Wang, X.; Yang, Y.; Zhang, J.; Xu, J.; Rengel, Z.; Chen, Q. Melatonin Alleviates Aluminum Toxicity through Modulating Antioxidative Enzymes and Enhancing Organic Acid Anion Exudation in Soybean. Funct. Plant Biol. 2017, 44, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Lv, T.; Huang, L.; Liu, X.; Jin, C.; Lin, X. Melatonin Ameliorates Aluminum Toxicity through Enhancing Aluminum Exclusion and Reestablishing Redox Homeostasis in Roots of Wheat. J. Pineal Res. 2020, 68, e12642. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, X.; Zhang, N.; Feng, L.; Ma, C.; Wang, Y.; Yang, Z.; Zhao, J. Melatonin Alleviates Aluminum-Induced Growth Inhibition by Modulating Carbon and Nitrogen Metabolism, and Reestablishing Redox Homeostasis in Zea Mays, L. J. Hazard. Mater. 2022, 423, 127159. [Google Scholar] [CrossRef]
- Huang, B.; Chen, Y.E.; Zhao, Y.Q.; Ding, C.B.; Liao, J.Q.; Hu, C.; Zhou, L.J.; Zhang, Z.W.; Yuan, S.; Yuan, M. Exogenous Melatonin Alleviates Oxidative Damages and Protects Photosystem II in Maize Seedlings Under Drought Stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, Y.; Chen, H.; Tan, J.; Xu, H.; Li, P.; Wu, D.; Jia, J.; Yang, Z. Transcriptome Analysis of Response to Aluminum Stress in Pinus Massoniana. Forests 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Rosa-Santos, T.M.; da Silva, R.G.; Kumar, P.; Kottapalli, P.; Crasto, C.; Kottapalli, K.R.; França, S.C.; Zingaretti, S.M. Molecular Mechanisms Underlying Sugarcane Response to Aluminum Stress by Rna-Seq. Int. J. Mol. Sci. 2020, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Grisel, N.; Zoller, S.; Künzli-Gontarczyk, M.; Lampart, T.; Münsterkötter, M.; Brunner, I.; Bovet, L.; Métraux, J.P.; Sperisen, C. Transcriptome Responses to Aluminum Stress in Roots of Aspen (Populus Tremula). BMC Plant Biol. 2010, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.; Li, S.; Zhou, Q.; Ashraf, U.; Qiao, J.; Li, X.; Wan, X.; Zheng, Y. Transcriptomic Analysis Provides Insights into the Differential Effects of Aluminum on Peanut (Arachis Hypogaea, L.). Genes (Basel). 2022, 13, 1830. 13.
- Liu, W.; Xiong, C.; Yan, L.; Zhang, Z.; Ma, L.; Wang, Y.; Liu, Y.; Liu, Z. Transcriptome Analyses Reveal Candidate Genes Potentially Involved in Al Stress Response in Alfalfa. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic Insights on Melatonin-Mediated Drought Stress Mitigation in Plants. Physiol. Plant. 2021, 172, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stass, A.; Horst, W.J. Apoplastic Binding of Aluminum Is Involved in Silicon-Induced Amelioration of Aluminum Toxicity in Maize. Plant Physiol. 2004, 136, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.L.; Li, Y.Y.; Zhang, Y.J.; Zhang, S.S.; Wu, Y.R.; Wu, P.; Zheng, S.J. Cell Wall Polysaccharides Are Specifically Involved in the Exclusion of Aluminum from the Rice Root Apex. Plant Physiol. 2008, 146, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, J.L.; He, L.S.; Li, Y.Y.; Zheng, S.J. Effect of Aluminum on Cell Wall, Plasma Membrane, Antioxidants and Root Elongation in Triticale. Biol. Plant. 2008, 52, 87–92. [Google Scholar] [CrossRef]
- Ma, J.F.; Shen, R.; Nagao, S.; Tanimoto, E. Aluminum Targets Elongating Cells by Reducing Cell Wall Extensibility in Wheat Roots. Plant Cell Physiol. 2004, 45, 583–589. [Google Scholar] [CrossRef]
- Yang, Z.B.; Eticha, D.; Rao, I.M.; Horst, W.J. Alteration of Cell-Wall Porosity Is Involved in Osmotic Stress-Induced Enhancement of Aluminium Resistance in Common Bean (Phaseolus Vulgaris, L.). J. Exp. Bot. 2010, 61, 3245–3258. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell Wall Hemicellulose Contributes Significantly to Aluminum Adsorption and Root Growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- XF, Z.; YZ, S.; GJ, L.; SC, F.; BC, Z.; YH, Z.; J, B.; T, J.; XY, X.; CZ, M.; et al. XTH31, encoding an in Vitro XEH/XET-Active Enzyme, Regulates Aluminum Sensitivity by Modulating in Vivo XET Action, Cell Wall Xyloglucan Content, and Aluminum Binding Capacity in Arabidopsis. Plant Cell 2012, 24. [Google Scholar]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. (Lausanne). 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of Melatonin in Abiotic Stress Resistance in Plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Asgher, M.; Ahmed, S.; Sehar, Z.; Gautam, H.; Gandhi, S.G.; Khan, N.A. Hydrogen Peroxide Modulates Activity and Expression of Antioxidant Enzymes and Protects Photosynthetic Activity from Arsenic Damage in Rice (Oryza Sativa, L.). J. Hazard. Mater. 2021, 401, 123365. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Yin, L.; Wang, B.; Ke, Q.; Deng, X.; Wang, S. Melatonin Promotes Plant Growth by Increasing Nitrogen Uptake and Assimilation under Nitrogen Deficient Condition in Winter Wheat. Plant Physiol. Biochem. 2019, 139, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Duan, S.; Zhou, Z.; Chen, S.; Wang, D. Foliar Spraying of Melatonin Confers Cadmium Tolerance in Nicotiana Tabacum, L. Ecotoxicol. Environ. Saf. 2019, 170, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, X.; Zhang, N.; Feng, L.; Ma, C.; Wang, Y.; Yang, Z.; Zhao, J. Melatonin Alleviates Aluminum-Induced Growth Inhibition by Modulating Carbon and Nitrogen Metabolism, and Reestablishing Redox Homeostasis in Zea Mays L. J. Hazard. Mater. 2022, 423, 127159. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Liu, Y.; Pang, J.; Yong, J.W.H.; Chen, Y.; Bai, C.; Gille, C.; Shi, Q.; Wu, D.; Han, X.; et al. Supplementary Calcium Restores Peanut (Arachis Hypogaea) Growth and Photosynthetic Capacity Under Low Nocturnal Temperature. Front. Plant Sci. 2019, 10, 1637. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Guo, H.; Wang, J.; Wang, Y.; Zhang, R. Melatonin Enhances Drought Tolerance by Regulating Leaf Stomatal Behavior, Carbon and Nitrogen Metabolism, and Related Gene Expression in Maize Plants. Front. Plant Sci. 2021, 12, 779382. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the Trehalose-6-Phosphate Phosphatase Gene OsTPP1 Confers Stress Tolerance in Rice and Results in the Activation of Stress Responsive Genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef]
- Zhu, F.; Li, M.; Sun, M.; Jiang, X.; Qiao, F. Plant Hormone Signals Regulate Trehalose Accumulation against Osmotic Stress in Watermelon Cells. Protoplasma 2022, 259, 1351–1369. [Google Scholar] [CrossRef] [PubMed]
- Kosmas, S.A.; Argyrokastritis, A.; Loukas, M.G.; Eliopoulos, E.; Tsakas, S.; Kaltsikes, P.J. Isolation and Characterization of Drought-Related Trehalose 6-Phosphate-Synthase Gene from Cultivated Cotton (Gossypium Hirsutum, L. ). Planta 2006, 223, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Wang, Y.; Imran, M.; Jiang, C. Boron Alleviates the Aluminum Toxicity in Trifoliate Orange by Regulating Antioxidant Defense System and Reducing Root Cell Injury. J. Environ. Manage. 2018, 208, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Xu, H.; Zhang, Y.; Chen, G. Silicon Mediated Redox Homeostasis in the Root-Apex Transition Zone of Rice Plays a Key Role in Aluminum Tolerance. Plant Physiol. Biochem. 2023, 201, 107871. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Zhang, X.; Jin, Q.; Li, J. Phenolic Profiles, Antioxidant Activities, and Neuroprotective Properties of Mulberry (Morus Atropurpurea Roxb.) Fruit Extracts from Different Ripening Stages. J. Food Sci. 2016, 81, 2439–2446. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Lan, P.; Ding, C.; Wang, J.; Zhang, T.; Wang, X. A New Perspective on the Toxicity of Arsenic-Contaminated Soil: Tandem Mass Tag Proteomics and Metabolomics in Earthworms. J. Hazard. Mater. 2020, 398, 122825. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Melatonin alleviates aluminum-induced phenotypic changes in alfalfa. (A) Phenotypic comparison after 3 days of AlCl3 and melatonin treatment. Scale bar, 1 cm. (B) Alfalfa root length; data are shown as mean ± SE (n=16). (C) Fresh weight of four alfalfa seedlings on a cotton ball; data are shown as mean ± SE (n=4). (D) Cross-sectional view of alfalfa root tips.
Figure 1.
Melatonin alleviates aluminum-induced phenotypic changes in alfalfa. (A) Phenotypic comparison after 3 days of AlCl3 and melatonin treatment. Scale bar, 1 cm. (B) Alfalfa root length; data are shown as mean ± SE (n=16). (C) Fresh weight of four alfalfa seedlings on a cotton ball; data are shown as mean ± SE (n=4). (D) Cross-sectional view of alfalfa root tips.
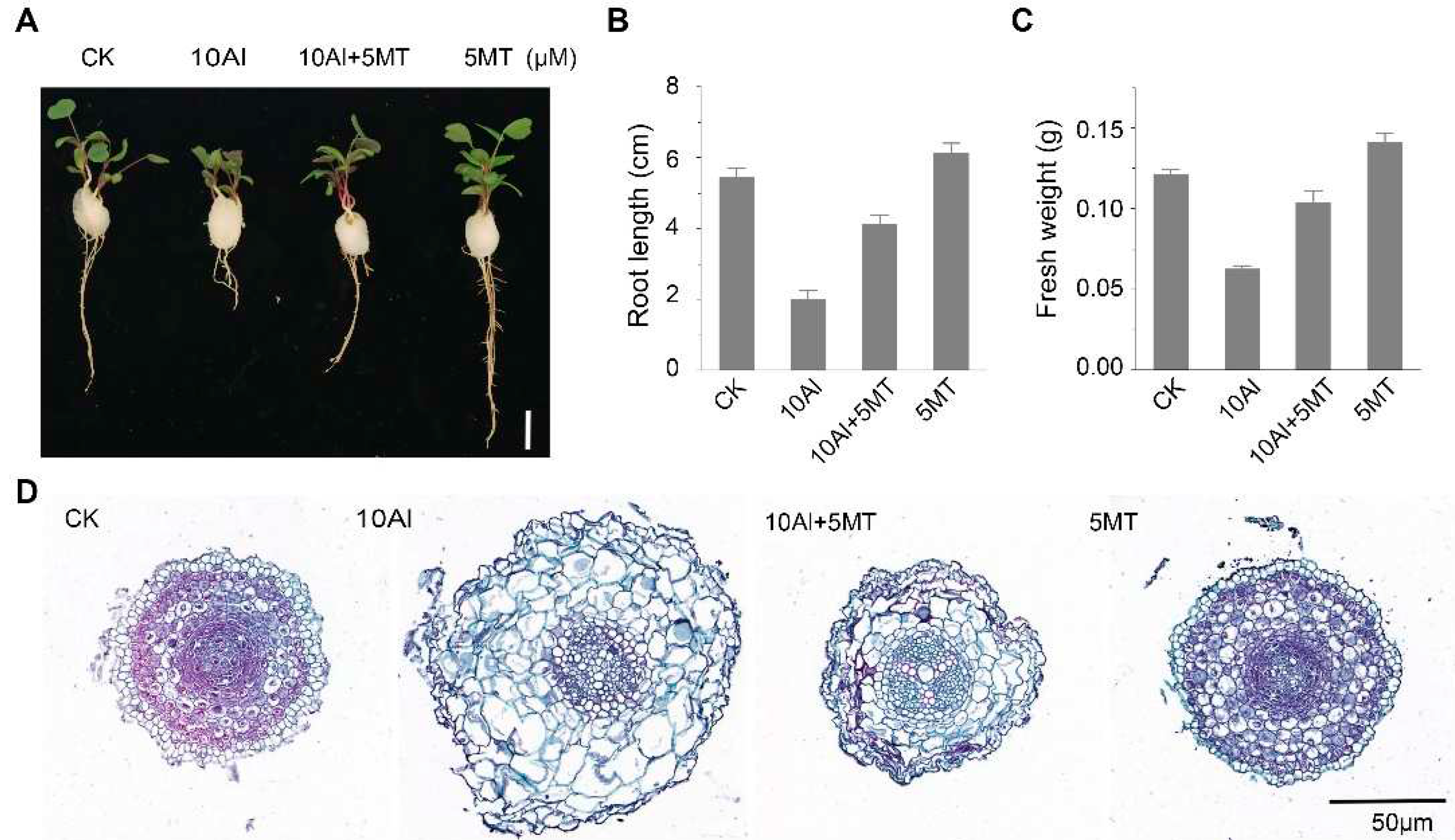
Figure 2.
Effects of melatonin on Al accumulation in alfalfa roots. (A) Morin staining after 3 days of AlCl3 and melatonin treatment. Scale bar, 100 μm. The Al content in alfalfa shoot (B) and root (C) after 3 days of AlCl3 and melatonin treatment. Data are shown as mean ± SD (n=3).
Figure 2.
Effects of melatonin on Al accumulation in alfalfa roots. (A) Morin staining after 3 days of AlCl3 and melatonin treatment. Scale bar, 100 μm. The Al content in alfalfa shoot (B) and root (C) after 3 days of AlCl3 and melatonin treatment. Data are shown as mean ± SD (n=3).
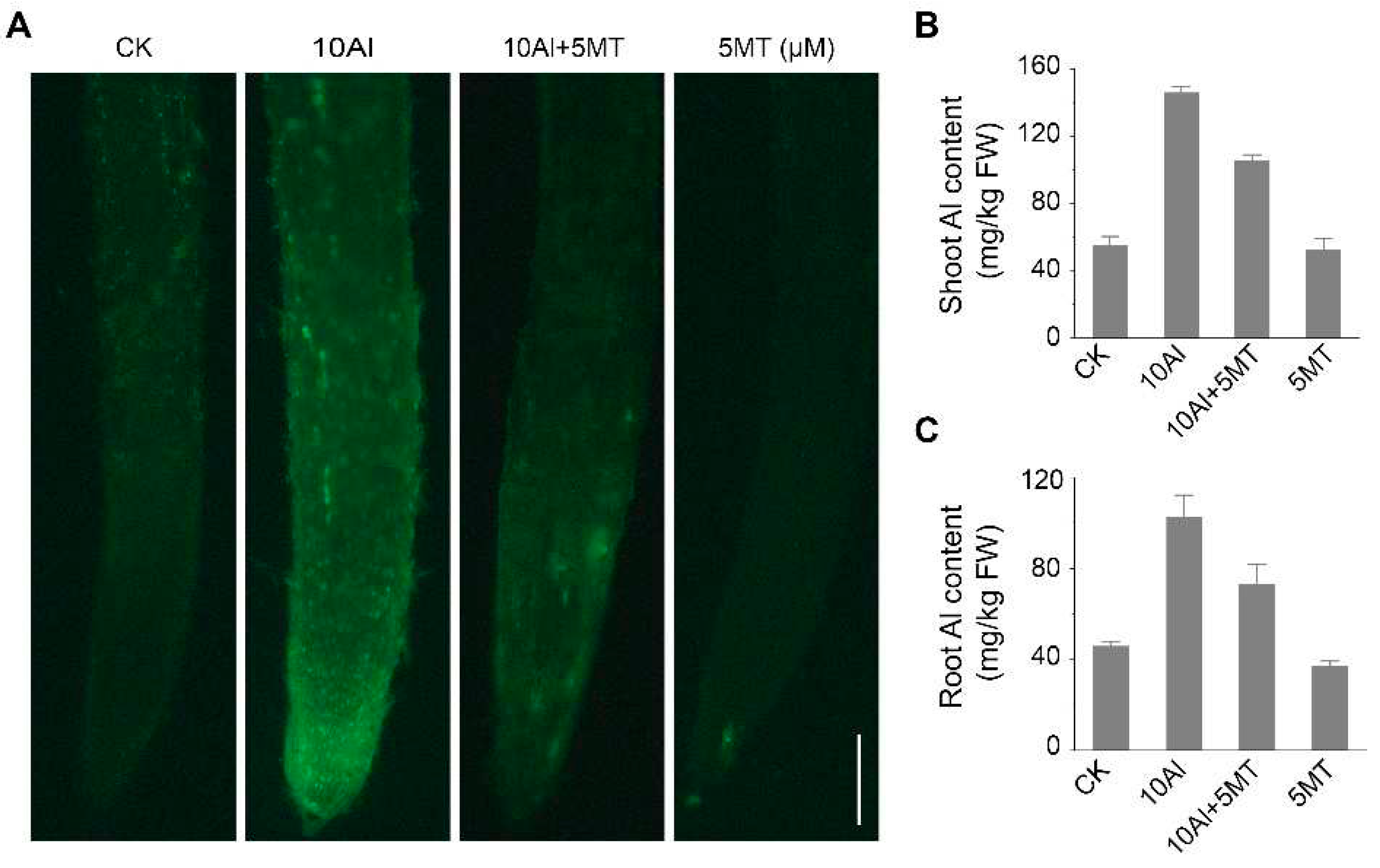
Figure 3.
Effects of melatonin on Al-induced oxidative damage in alfalfa roots. (A) Evans blue staining after 3 days of AlCl3 and melatonin treatment. Scale bar, 100μm. The content of H2O2 (B) and MDA (C) in alfalfa root after 3 days of treatment. Data are shown as mean ± SD (n=3).
Figure 3.
Effects of melatonin on Al-induced oxidative damage in alfalfa roots. (A) Evans blue staining after 3 days of AlCl3 and melatonin treatment. Scale bar, 100μm. The content of H2O2 (B) and MDA (C) in alfalfa root after 3 days of treatment. Data are shown as mean ± SD (n=3).
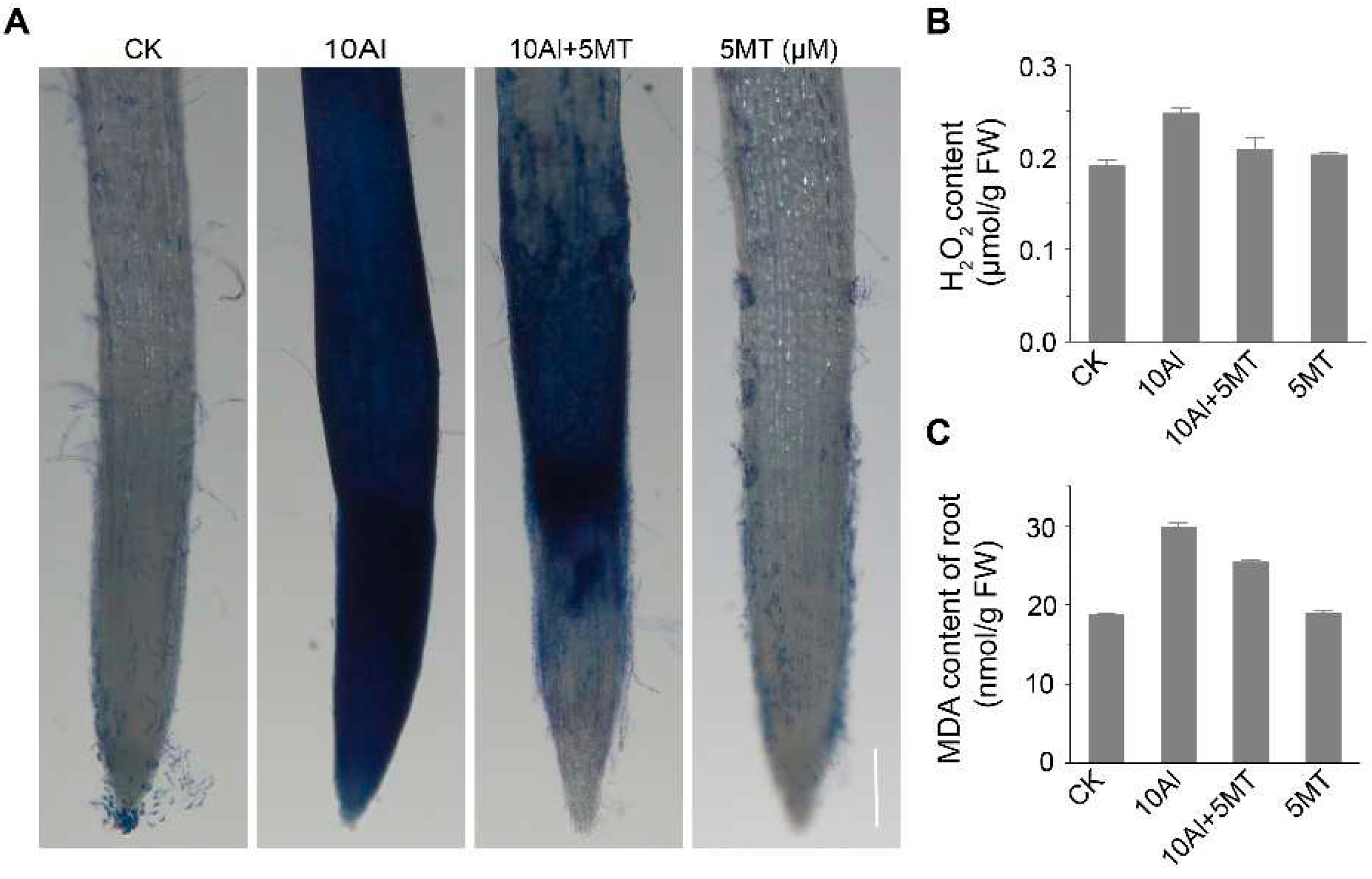
Figure 4.
Effects of Al and melatonin on the activity of antioxidant enzymes in alfalfa roots. (A) Superoxide dismutase (SOD) activity, (B) peroxidase (POD) activity, (C) catalase (CAT) activity, and (D) ascorbate peroxidase (APX) activity in alfalfa roots after 3 days of treatment. Data are shown as mean ± SD (n=3).
Figure 4.
Effects of Al and melatonin on the activity of antioxidant enzymes in alfalfa roots. (A) Superoxide dismutase (SOD) activity, (B) peroxidase (POD) activity, (C) catalase (CAT) activity, and (D) ascorbate peroxidase (APX) activity in alfalfa roots after 3 days of treatment. Data are shown as mean ± SD (n=3).
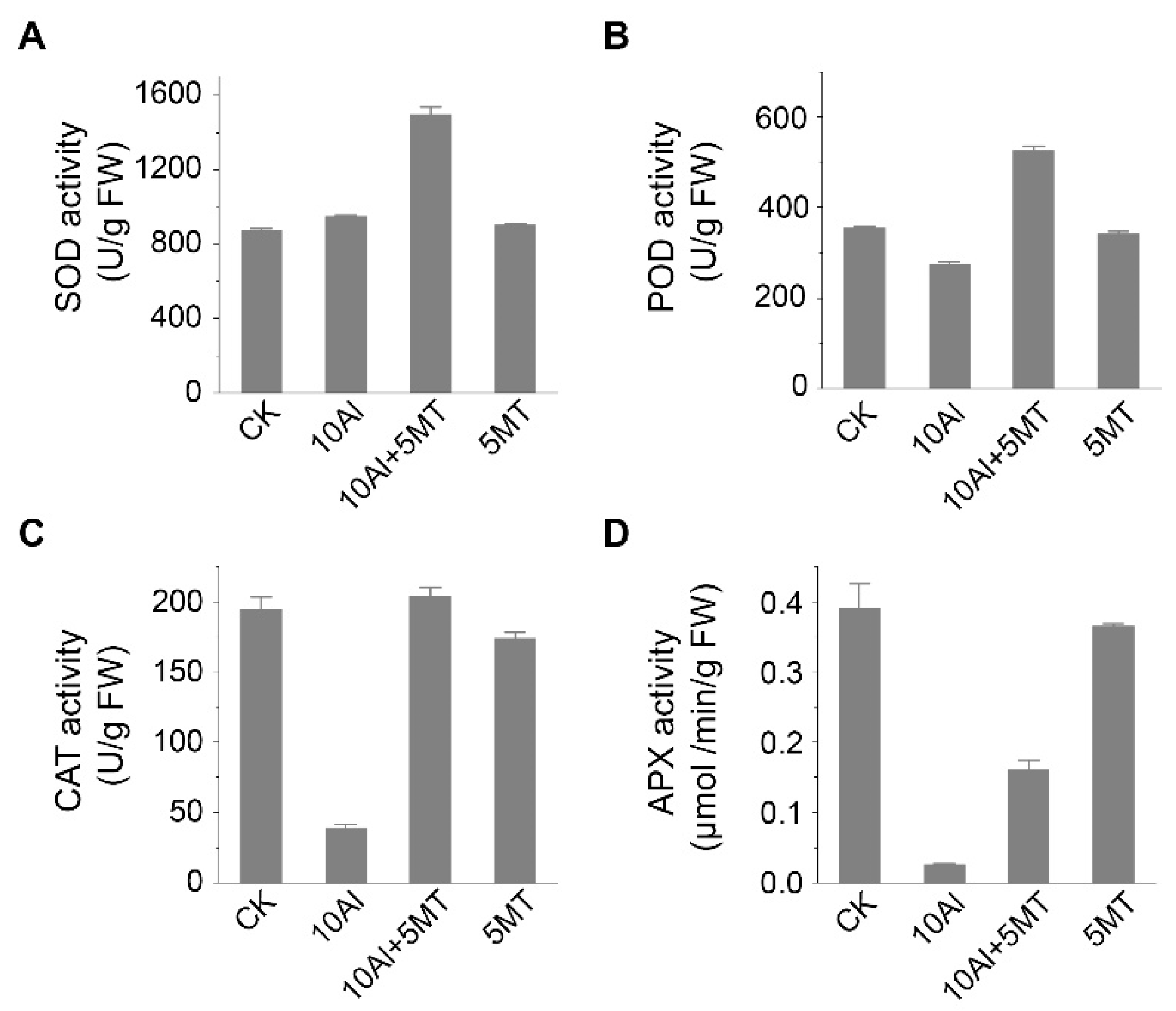
Figure 5.
Global analysis of the transcriptome data of alfalfa roots. (A) Principal Component Analysis (PCA) of the RNA-seq data. (B) Correlation heatmap among the samples, with the horizontal and vertical coordinates representing the square of the correlation coefficients for each sample. (C) Number of differentially expressed genes (DEGs) for each comparison. (D) Venn diagram illustrating DEGs in different comparisons. MT, Al represents the treatment of Al and melatonin (MT).
Figure 5.
Global analysis of the transcriptome data of alfalfa roots. (A) Principal Component Analysis (PCA) of the RNA-seq data. (B) Correlation heatmap among the samples, with the horizontal and vertical coordinates representing the square of the correlation coefficients for each sample. (C) Number of differentially expressed genes (DEGs) for each comparison. (D) Venn diagram illustrating DEGs in different comparisons. MT, Al represents the treatment of Al and melatonin (MT).
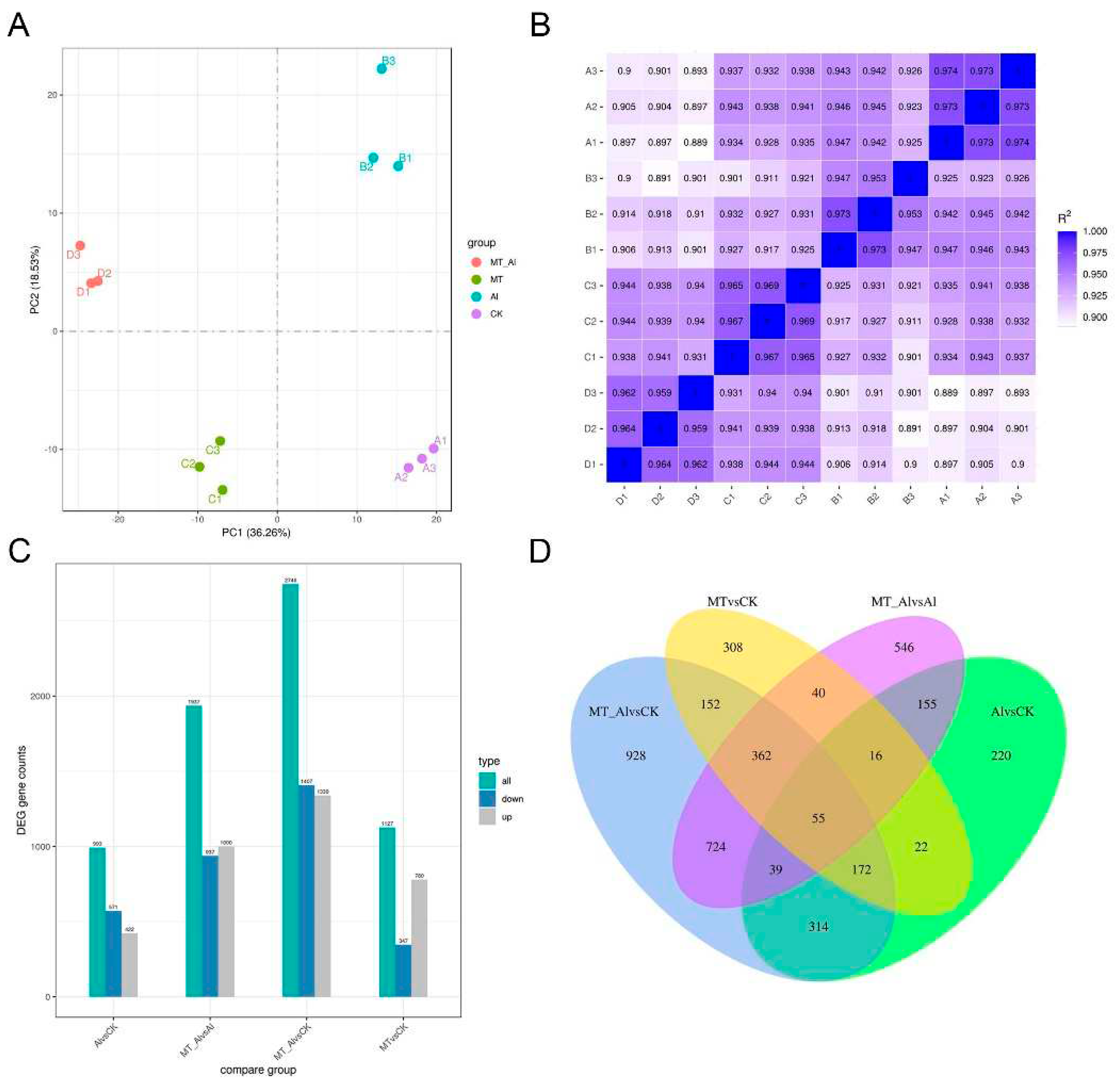
Figure 6.
Enrichment analysis of differentially expressed genes (DEGs) on GO terms and KEGG pathways. (A) GO term enrichment analysis of the Al + MT vs. Al pair, showing the top 30 terms. (B) KEGG pathway enrichment analysis of the Al + MT vs. Al pair, showing the top 30 pathways. (C) Transcript changes in glycolysis-related genes. (D) Transcript changes in starch and sucrose metabolism-related genes.
Figure 6.
Enrichment analysis of differentially expressed genes (DEGs) on GO terms and KEGG pathways. (A) GO term enrichment analysis of the Al + MT vs. Al pair, showing the top 30 terms. (B) KEGG pathway enrichment analysis of the Al + MT vs. Al pair, showing the top 30 pathways. (C) Transcript changes in glycolysis-related genes. (D) Transcript changes in starch and sucrose metabolism-related genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



