
Submitted:
05 November 2023
Posted:
06 November 2023
You are already at the latest version
Abstract
Toxicological studies have shown that the cockroach Periplaneta americana is a classical model for studying the mode of action of commonly used insecticides. In a previous study, we demonstrated that thiamethoxam and clothianidin decreased locomotor activity and decreased the locomotor activity in an open-field-like apparatus. Here, we tested the effect of the neonicotinoid acetamiprid when applied orally, topically, or injected into the haemolymph. We found that acetamiprid decreased locomotor activity in the open-field-like apparatus. When treated with acetamiprid, a strong alteration in locomotor activity was observed 1 h, 24 h and 48 h after haemolymph and topical applications. Oral application of acetamiprid demonstrated that it impaired locomotor activity at 24 h and 48 h. A comparison of the present data with our previously published results showed that neonicotinoids were more active when injected into the haemolymph compared to oral and topical applications. These findings increased our understanding of the effect of neonicotinoid insecticides on insect locomotor activity, and demonstrated that the cyano-substituted neonicotinoid, acetamiprid, was able to alter cockroach locomotor activity.
Keywords:
Insect
; cockroach
; locomotor activity
; insecticide
; neonicotinoid
; acetamiprid
1. Introduction
The cockroach Periplaneta americana is an insect pest in urban entomology, and a potential vector of various pathogenic organisms, and also produces allergens that are responsible for allergies and asthma [1,2]. It is currently used as a model for studying the mode of action of several compounds, including neonicotinoid insecticides [3,4,5,6,7]. Neonicotinoids are agrochemical compounds, which were discovered in the 1980s [8]. They are strongly efficient in controlling sap-feeding insects in agriculture [9,10]. The seven major commercial neonicotinoids are divided into three structural compounds including chloropyridinyl (imidacloprid, nitempyram, thiacloprid and acetamiprid), clorothiazolyl (clothianidin and thiamethoxam), and tetrahydrofuryl (dinotefuran) [8,11]. At the present, their mode of action are thought to be related to their capacity to bind to nicotinic acetylcholine receptors as agonists [12,13,14,15,16]. Neonicotinoids were split into two different subgroups, depending on their action on cockroach Periplaneta americana thoracic neurons [7]. Those with a heterocyclic ring in their electronegative pharmacophore moiety (imidacloprid and thiacloprid) and the open chain compounds (acetamiprid, dinotefuran, nitenpyram and clothianidin), which were much more effective agonists compared to acetylcholine [7]. Moreover, different neonicotinoids also caused two distinct types of intoxication, which were associated to their chemical structures. For example, imidacloprid and thiacloprid, which have a heterocyclic electronegative moiety, caused strong excitatory symptoms, with uncoordinated quivering, hyper-excitability and rapid spontaneous movements. By contrast, the open chain compounds elicited different poisoning symptoms. In this group, acetamiprid was the exception, causing excitation symptoms rather than depression and paralysis [7]. Acetamiprid is formed with chloropyridinylmethyl and N-cyanoimine groups [17]. In a recent study comparing dinotefuran, imidacloprid and acetamiprid, it was demonstrated that acetamiprid and dinotefuran had a stronger hydrogen-bond acceptor site than imidacloprid due to their cyano and nitro moieties, respectively [18]. The crystal structure of acetamiprid pointed out cooperative π-π and hydrogen-bond interactions important for the binding with nicotinic acetylcholine receptors [18]. Electrophysiological studies performed on cockroach dorsal unpaired median (DUM) neurons, located in the sixth abdominal ganglion, showed that a pressure application of acetamiprid onto isolated DUM neuron somata induced a biphasic dose-response curve. These results demonstrated that it acted as an agonist on both cockroach nicotinic acetylcholine receptor 1 and 2 subtypes, whereas imidacloprid only activated the subtype 1 [19]. At the synaptic level, using the synapse between the cercal afferent giant interneuron and the nerve XI, it was demonstrated that acetamiprid induced a strong dose-dependent increase in the ganglionic depolarisation [18]. Thus, acetamiprid was able to activate synaptic and extra-synaptic nicotinic acetylcholine receptor subtypes.
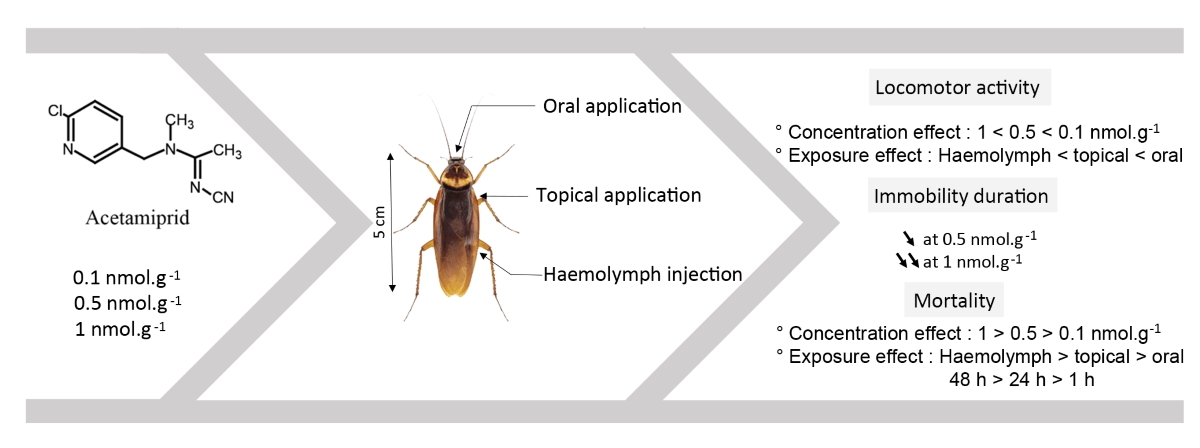
In the present study, we explored the effect of acetamiprid on cockroach locomotor activity and discussed the results regarding our previous studies on the effects of other neonicotinoids. Indeed, using an open-field-like apparatus, we demonstrated that at higher doses (0.5 nmol.g-1 and 1 nmol.g-1), clothianidin led to uncoordinated movements, leg shakings and prostration in intoxicated cockroaches. However, when exposed to thiamethoxam, cockroaches only demonstrated excitatory signs with characteristic shaking of legs [19]. Moreover, we found that the two major nitro-imines (thiamethoxam [20] and clothianidin [21]) decreased cockroach locomotor activity. The percentage of cockroaches displaying locomotor activity was significantly reduced one hour after haemolymph application. No significant effect was found after topical and oral administration of thiamethoxam and clothianidin. Thiamethoxam levels remained persistent 48 h after application, and the amount of clothianidin in cockroach tissues was persistent with the toxicity of thiamethoxam [22]. The main conclusion was that the effect of thiamethoxam was partly due to its metabolization into clothianidin. Here, we studied the effect of the cyano-imine acetamiprid using the same experimental conditions.
2. Materials and Methods
2.1. Animals
Cockroaches Periplaneta americana used for the experiments were obtained from Arbiotech (Saint Gilles, France). Until use, they were kept under a 12 h : 12 h dark : light cycle, with food and water provided ad libitum. Before acetamiprid administration, cockroaches were anesthetized on ice (4°C) during 10 min [23]. Then, all experiments were done at room temperature. The experimental procedures were in compliance with the European Laws (86/609/CEE) on the use of animals.
2.2. Chemicals
All chemicals were purchased from Sigma-Aldrich (Saint Quentin, France). Acetamiprid (Figure 1A) was dissolved in dimethylsulfoxide (DMSO) to prepare a concentrated stock solution. The stock solution was extemporaneously diluted to get a working solution of acetamiprid at 0.01% DMSO). The control consisted in DMSO diluted at a final concentration of 0.01% in a saline solution. The saline solution contained (in mmol-1): NaCl 208; KCl 3.1; CaCl2 5.4; NaHCO3 2; sucrose 26; and had a pH of 7.4 [24].
2.3. Exposure protocol
Acetamiprid was applied to cockroaches in three different ways at concentrations of 0.1, 0.5 and 1 nmol.g-1 [22]. For topical application, 1 µl of the final solution was applied to the thorax of the cockroach using a Hamilton syringe. As previously described, for the haemolymph treatment, 10 µl of solution were injected between the third and fourth sternites [25]. Oral application consisted to a starvation for the day before the experiment, and then cockroaches were fed with 5 µl of the working solution (either acetamiprid or control solution) [22].
2.4. Evaluation of locomotor activity
The locomotor activity of cockroaches was individually evaluated using an open-field-like apparatus (32 x 6.5 x 3 cm), according to the method used by Lambin et al.[26], with slight adaptations of the device. Two series of 14 LEDs divided the device into seven different levels and allowed the observation of cockroaches’ horizontal displacements (Figure 1B). The cockroaches were tested 1 hour after acetamiprid or control exposure. Each individual was introduced to the open-field-like apparatus, via the cylinder (a), which remained closed by the trap door [22]. At the beginning of the recording, the door opened and the cockroach walked from the left to the right freely during 5 min. Several parameters such as the position and the crossing time of the device (average time between the trap door and the last LED) were automatically scored. The proportion of cockroaches displaying locomotor activity were evaluated 1 h, 24 h and 48 h after application of acetamiprid or control solutions.
2.5. Evaluation of cockroaches’ mortality
To evaluate the direct toxicity of acetamiprid, the total number of dead cockroaches was determined for each acetamiprid concentration (0.1, 0.5 and 1 nmol.g-1) applied either by injection into the haemolymph, by topical or oral application. The mortality was scored at 1 h, 24 h and 48 h after acetamiprid exposure. The number of dead cockroaches observed in the corresponding control condition was used to calculate the corrected mortality percentage, using Henderson Tilton’s formula.
2.6. Mannitol-gap recordings
Mannitol-gap experiments were performed according to the previously published protocol [27,28,29,30]. Cockroaches were dissected and opened along the longitudinal dorsal-median line with a fine pair of forceps. The sixth abdominal ganglion, nerve cord, one circus and the corresponding cercal nerve XI were isolated in saline solution containing (in mM) : NaCl, 208; KCl, 3.1; CaCl2, 5.4; NaHCO3, 2; Sucrose, 26; pH 7.4 [24,29,30]. The preparation was transferred to the recording chamber and superfuzed with mannitol solution (87 g/l). The excitatory postsynaptic potentials and action potentials were evoked using electrical stimulation of cercal nerve XI with a dual pulse stimulator (Campden 915, USA) [24,29,30].
2.7. Statistical analysis
We used a generalized linear model (GLM) with repeated measures to compare three factors: concentration (0.1, 0.5 and 1 nmol.g-1), application (haemolymph, topical and oral), and treatment (acetamiprid and control), using R [31]. The variable was the binomial family in which ‘1’ corresponded to a cockroach eliciting locomotor activity and ‘0’ to a cockroach without locomotor activity. We also used Student t-test and Welch’s correction to illustrate the relationship between control and tested cockroaches (GraphPad Prism 9).
3. Results
3.1. Percentage of cockroaches displaying locomotor activity
We used a set of simulation scenarios to estimate the probability of detecting a significant difference between the acetamiprid and control groups (Using R. Program). Three factors were evaluated: application (haemolymph, topical and oral), concentration (0.1, 0.5 and 1 nmol.g-1) and treatment (acetamiprid and control). Each cockroach tested belonged to one of two categories: (1) normal cockroaches (normal locomotor activity) and (2) those, which were not able to present locomotor activity after acetamiprid application. Over the 180 cockroaches tested, a significant difference appeared between each application (χ² = 44.48, p < 0.001), concentration (χ2 = 38.50; p < 0.001) and treatment (χ2 = 22.27; p < 0.001). Globally, acetamiprid was more active when injected into the haemolymph compared to oral or topical applications (χ2 = -3.4, p < 0.05). The proportion of cockroaches eliciting locomotor activity decreased under haemolymph application, after 1 h, for the following doses, 0.5 nmol.g-1 (t = 96.0, df = 4, p < 0.05, n = 20) and 1 nmol.g-1 (t = 1105, df = 4, p < 0.001, n = 20, Figure 2), compared to control group. The decrease in the locomotor activity was also found for the same doses at 24 h (0.5 nmol.g-1: t = 39.1, df = 4, p < 0.05, n = 20; 1 nmol.g-1: t = 724.8, df = 4, p < 0.001, n = 20) and 48 h (0.5 nmol.g-1: t = 60.9, df = 4, p < 0.05, n = 20; 1 nmol.g-1: t = 226.2, df = 4, n = 20) after exposure to acetamiprid.
For the topical application of acetamiprid (Figure 3), a decrease in locomotor activity was only found at concentrations of 1 nmol.g-1, and was observed at all time delays, 1 h (t = 36.3, df = 4, p < 0.05, n = 20), 24 h (t = 43.5, df = 4, p < 0.001, n = 20) and 48 h (t = 43.4, df = 4, , p < 0.05, n = 20).
Oral application of acetamiprid led to a significant lowering of locomotor activity at 0.5 nmol.g-1 for 48 h (t = 46.5, df = 4, p < 0.05, n = 20). Similarly, a decrease was also found at 1 nmol.g-1 at 24 h (t = 30.2, df = 4, p < 0.05, n = 20) and 48 h (t = 78.0, df = 4, p < 0.05, n = 20) (Figure 4).
3.2. Cockroach mortality after acetamiprid application
We assessed the direct toxicity of acetamiprid for the different tested concentration and for each acetamiprid exposure method. The mortality scores correspond to corrected mortality percentage (with n = number of cockroaches tested). No mortality was observed at 1 h after exposure to acetamiprid for the different concentrations (0.1, 0.5 and 1 nmol.g-1) and exposure methods (haemolymph, topical or oral application). At 24 h, no mortality was found after topical application of acetamiprid at the different concentrations. After heamolymph injection, acetamiprid led to 5% mortality at 0.5 nmol.g-1 and 1 nmol.g-1; for both concentrations the mortality was not significant compared to the corresponding control conditions (p > 0.05, n = 50). We did not observed any dead individual at the lowest concentration (0.1 nmol.g-1). At this delay, an increase in mortality was found in orally intoxicated cockroaches. In particular, oral application of acetamiprid at 1 nmol.g-1, induced 10% mortality at 24 h, although this percentage was not statistically different from control condition (p > 0.05, n = 50). The lower concentrations (0.1 and 0.5 nmol.g-1) did not induced any mortality, 24 h after oral exposure.
At 48 h after acetamiprid application, no mortality was observed for haemolymph, topical and oral applications of 0.1 nmol.g-1. However, for haemolymph application, a significant toxicity was observed with 20% mortality at 0.5 nmol.g-1 (p < 0.05, n = 50) and 35% mortality at 1 nmol.g-1 (p < 0.05, n = 20). A similar concentration dependent toxicity was observed 48 h after topical application, with 10% (p > 0.05, n = 50) and 20% (p < 0.05, n = 50) mortality at 0.5 nmol.g-1 and 1 nmol.g-1 respectively. We also found that acetamiprid applied orally induced a significant toxicity with 35% mortality at the concentration of 1 nmol.g-1 (p < 0.05, n = 50 cockroaches), whereas no mortality was observed at 0.5 nmol.g-1. In complement, we observed some particularities concerning cockroaches that were more sensitive and thus died after application of acetamiprid. We first found that they were unable to walk correctly, and they presented an increase of their body length compared to control cockroaches. Normal and treated cockroaches were between 3.5 – 4 cm, and 5.5 - 6 cm lenght, respectively (Figure 5).
3.3. Characterization of cockroach locomotor activity: immobility and time of exploration
For the cockroaches displaying locomotor activity, we evaluated the locomotor behavior more precisely. In particular, we determined the duration of immobility and the time of exploration. These measurements were done 1 h after acetamiprid application to avoid any bias due to direct toxicological effect. We observed that the duration of immobility strongly increased when acetamiprid was applied at higher concentrations (Figure 6). Indeed, no significant difference was found with control groups, at 0.1 nmol.g-1 after haemolymph (t = 1.14, df = 85.9, p > 0.05, n = 50), topical (t = 1.52, df = 90.3, p > 0.05, n = 50) and oral (t = 0.65, df = 90.3, p > 0.05, n = 50) applications. After exposure to acetamiprid at 0.5 nmol-g-1 significant difference was revealed between control and treated cockroaches, with an exposure through haemolymph (t = 5.4, df = 51.2, p < 0.05), topical (t = 5.9, df = 63.2, p < 0.05, n = 50), or oral (t = 6.5, df = 68, p < 0.05, n = 50) applications. At 1 nmol.g-1, a significant difference was found after haemolymph (t = 5.0, df = 49.2, P < 0.05, n = 50), topical (t = 4.8, df = 49.6, p < 0.05, n = 50), and oral (t = 6.1, df = 50.7, p < 0.05, n = 50) applications.
To assess the exploration speed of the cockroaches, we measured the time needed for each individual from the trap door of the cylinder (starting area) to the last LED of the open-field-like apparatus. The average time spent to cross the experimental device is reported in the Table 1, for the different way of exposure and the different concentrations of acetamiprid. We found that the time of exploration was increased in a similar way as the immobility duration. In fact, the time needed to reach the last LED was increased at higher concentration.
3.4. Effect of acetamiprid on cockroach sixth abdominal ganglion
To complete our study, we aimed to evaluate if excitatory postsynaptic potentials or action potentials could be recorded in treated cockroaches, which presented a decrease of locomotor activity. We focused our study on cockroaches treated with 1 nmol.g-1 acetamiprid after haemolymph injection, topical and oral applications, because major effect on locomotor activity was found with 1 nmol.g-1. Thus, using mannitol-gap recording, the sixth abdominal ganglion from treated cockroaches were compared, as used in our previous studies [18].
Our results showed that the sixth abdominal ganglia from cockroaches treated with 1 nmol.g-1 acetamiprid, presented action potentials after electrical stimulation of the nerve XI, similarly to control cockroaches (figure 7). Thus, in control cockroaches, the electrical stimulation led to action potentials of 3.82 ± 0.10 mV, whereas they were 3.21 ± 0.14 mV (n = 6 ganglia, Figure 7A), 3.20 ± 0.16 mV (n = 5, Figure 7B) and 3.93 ± 0.10 mV (n = 6, Figure 7C), after haemolymph, topical and oral applications of 1 nmol.g-1 acetamiprid. These results suggested that synaptic transmission of cockroaches which had a decrease of locomotor activity was not affected by acetamiprid.
4. Discussion
The present study focused on the effect of acetamiprid on the locomotor activity of the cockroach Periplaneta americana. Our results show an effect of the concentration and application methods of acetamiprid, and the locomotor activity. We first found that at higher concentrations, the mean duration of immobility increased irrespective of the application method. A similar effect had previously been found with thiamethoxam and clothianidin [22], confirming that at higher doses, exposure to neonicotinoids increased the mean duration of immobility in the open-field-like apparatus. This experiment showed that when acetamiprid was administered to the haemolymph or applied topically, a decrease in the locomotor activity was found one hour after application, whereas no effect was seen after oral administration. After 24 h and 48 h, a strong effect occurred at higher concentrations, for all application methods. Similar discrepancies were found in our previous study using thiamethoxam and clothianidin [22]. This suggests that the three neonicotinoids altered the locomotor activity differently, depending on the application method. Indeed, a comparison of the present data with our previously published results showed that neonicotinoids were more active when injected into the haemolymph compared to oral and topical applications. We proposed that acetamiprid is more active than clothianidin and thiamethoxam one hour after application. Similar analysis performed at 24 h showed a significant effect when thiamethoxam was applied topically. We also highlighted a concentration dependent effect of acetamiprid, which was more effective at higher concentration. Moreover, we found that despite its effect on locomotor activity, acetamiprid did not affect synaptic transmission in treated cockroaches. Using mannitol-gap recording, we found that electrical stimulation of the nerve XI, which is the largest and the most posterior of the nerves in the sixth ganglion, induced excitatory postsynaptic potentials and action potentials. We propose that the underlying circuitry of cockroach locomotor activity affected by acetamiprid are independent to sensitive interneurons in the abdominal nerve cord, in particular giant interneurons in the sixth abdominal ganglion. Indeed, for insecticides there are several potential sites of action located at the presynaptic terminal and/or on the postsynaptic side. Our results demonstrated that acetamiprid did not bind to nicotinic acetylcholine receptors expressed in the sixth abdominal ganglion. The effect of neonicotinoid insecticides on locomotor activity was previously demonstrated in the honeybee Apis mellifera using an open-field apparatus [32]. In naive honeybees, it was demonstrated that neonicotinoids increase the time spent immobile, and the distance covered was reduced in treated honeybees [32]. Moreover, thiamethoxam and acetamiprid had no significant effect on the parameters of locomotor activity of treated honeybee compared to control groups. The same lack of effect was observed whatever the method used to apply the neonicotinoids [32]. This result was in accordance with our observations. The most significant finding in our studies was that as with acetamiprid and clothianidin, thiamethoxam altered cockroach locomotor activity, despite the fact that we did not observe uncoordinated movements or leg tremors. Consequently, we confirmed that thiamethoxam had its own effect, which was not associated to its metabolite clothianidin as previously proposed [33]. In fact, pharmacokinetic studies have revealed that thiamethoxam was metabolized to clothianidin and was the predominant neonicotinoid in both plants and insects [33]. In this previous experiment, one explanation could be that thiamethoxam and clothianidin were differently metabolized in cockroaches and/or had a different action on their targets. This explanation is consistent with the finding that the metabolism of the neonicotinoid either increases or decreases its potency depending on the compound and specificity of the nicotinic acetylcholine receptor subtypes [7,8,21,34,35]. The link between neonicotinoids and cockroach nicotinic receptors has been demonstrated in several studies [36,37,38,39,40]. The neonicotinoid agonist actions vary significantly, including partial, full and super activities. Clothianidin, which has an acyclic moiety corresponding to the imidazolidine moiety of imidacloprid, was referred to as a super or full agonist [7,41], and acetamiprid was the least efficacious analog in the acyclic group [7]. These agonist activities appeared to be related both to their structure and the cockroach nicotinic acetylcholine receptor subtypes expressed in the insect central nervous system [7]. These discrepancies between the three neonicotinoids were also observed in other studies. Several nicotinic acetylcholine receptor subtypes have been identified between the cercal afferent giant interneuron synapses and in the fast coxal depressor neurons [24,42]. Clothianidin, acetamiprid and thiamethoxam differently acted as agonists of the cockroach nicotinic receptors expressed in the ventral nerve cord [7,43]. Given this background information, we suggested for that neonicotinoids, which alter the cholinergic neurons through their action on insect neuronal nicotinic acetylcholine receptors, could also impair locomotor behavior of the cockroach Periplaneta americana.
5. Conclusions
In the present study, we demonstrated that the cyano-substituted neonicotinoid, acetamiprid, which differently acted as an agonist of cockroach nicotinic acetylcholine receptors, could also impair locomotor activity, as found with the nitro-subtituted, thiamethoxam and clothianidin.
Author Contributions
E.T. and S.H.T conceived and designed the research. S.H.T., A.C. and E.T., analyzed the data and wrote de manuscript. E.T conducted the experiments. S.H.T. and E.T. edited the manuscript. S.H.T. received the funding.
Funding
This research was funded by the Region Centre-Val de Loire “Electro-CELL funding”N°21060LBL. Emiliane Taillebois received a BQR funding from the University of Orleans.
Data Availability Statement
Data presented in this study are available on request from the corresponding author.
Acknowledgments
Authors would like to thanks all master students for their technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fang, Y.; Long, C.; Bai, X.; Liu, W.; Rong, M.; Lai, R.; An, S. Two new types of allergens from the cockroach, Periplaneta americana. Allergy 2015, 70, 1674-1678. [CrossRef]
- Gore, J.C.; Schal, C. Cockroach allergen biology and mitigation in the indoor environment. Annu Rev Entomol 2007, 52, 439-463. [CrossRef]
- Kagabu, S.; Ishihara, R.; Hieda, Y.; Nishimura, K.; Naruse, Y. Insecticidal and neuroblocking potencies of variants of the imidazolidine moiety of imidacloprid-related neonicotinoids and the relationship to partition coefficient and charge density on the pharmacophore. J Agric Food Chem 2007, 55, 812-818. [CrossRef]
- Kagabu S., K.C., Nishimura K., . Insecticidal and neuroblocking activities toward American cockroach (Periplaneta americana L.) of imidacloprid metabolites, 5-hydroxy-, 4,5-dihydroxy- and 4,5-dehydroimidacloprid. J Pestic Sci 2004, 29, 376-379.
- Kagabu S., M.N., Hibino R., Hanzawa M., Nishimura K. Insecticidal and neuroblocking activities of thiamethoxam-type compounds in the American cockroach (Periplaneta americana L.). J Pestic Sci 2005, 30, 111-115. [CrossRef]
- Kiriyama, K.; Nishiwaki, H.; Nakagawa, Y.; Nishimura, K. Insecticidal activity and nicotinic acetylcholine receptor binding of dinotefuran and its analogues in the housefly, Musca domestica. Pest Manag Sci 2003, 59, 1093-1100. [CrossRef]
- Tan, J.; Galligan, J.J.; Hollingworth, R.M. Agonist actions of neonicotinoids on nicotinic acetylcholine receptors expressed by cockroach neurons. Neurotoxicology 2007, 28, 829-842. [CrossRef]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: mechanisms of selective action. Annu Rev Pharmacol Toxicol 2005, 45, 247-268. [CrossRef]
- Jeschke, P.; Nauen, R. Neonicotinoids-from zero to hero in insecticide chemistry. Pest Manag Sci 2008, 64, 1084-1098. [CrossRef]
- Jeschke, P.; Nauen, R.; Beck, M.E. Nicotinic acetylcholine receptor agonists: a milestone for modern crop protection. Angew Chem Int Ed Engl 2013, 52, 9464-9485. [CrossRef]
- Casida, J.E. Neonicotinoids and Other Insect Nicotinic Receptor Competitive Modulators: Progress and Prospects. Annu Rev Entomol 2018, 63, 125-144. [CrossRef]
- Honda, H.; Tomizawa, M.; Casida, J.E. Insect nicotinic acetylcholine receptors: neonicotinoid binding site specificity is usually but not always conserved with varied substituents and species. J Agric Food Chem 2006, 54, 3365-3371. [CrossRef]
- Tomizawa, M.; Casida, J.E. Minor structural changes in nicotinoid insecticides confer differential subtype selectivity for mammalian nicotinic acetylcholine receptors. Br J Pharmacol 1999, 127, 115-122. [CrossRef]
- Tomizawa, M.; Lee, D.L.; Casida, J.E. Neonicotinoid insecticides: molecular features conferring selectivity for insect versus mammalian nicotinic receptors. J Agric Food Chem 2000, 48, 6016-6024. [CrossRef]
- Thany, S.H.; Lenaers, G.; Raymond-Delpech, V.; Sattelle, D.B.; Lapied, B. Exploring the pharmacological properties of insect nicotinic acetylcholine receptors. Trends Pharmacol Sci 2007, 28, 14-22. [CrossRef]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid Insecticides: Molecular Targets, Resistance, and Toxicity. Annu Rev Pharmacol Toxicol 2020, 60, 241-255. [CrossRef]
- Yamada T., T.H., Hatano R. A novel insecticide, acetamiprid In Nicotinoid insecticides and the nicotinic acetylcholine receptor, Yamamoto L;, Casida J.E. In Eds; 1999; pp. 149-176.
- Le Questel, J.Y.; Graton, J.; Ceron-Carrasco, J.P.; Jacquemin, D.; Planchat, A.; Thany, S.H. New insights on the molecular features and electrophysiological properties of dinotefuran, imidacloprid and acetamiprid neonicotinoid insecticides. Bioorg Med Chem 2011, 19, 7623-7634. [CrossRef]
- Bodereau-Dubois, B.; List, O.; Calas-List, D.; Marques, O.; Communal, P.Y.; Thany, S.H.; Lapied, B. Transmembrane potential polarization, calcium influx, and receptor conformational state modulate the sensitivity of the imidacloprid-insensitive neuronal insect nicotinic acetylcholine receptor to neonicotinoid insecticides. J Pharmacol Exp Ther 2012, 341, 326-339. [CrossRef]
- Maienfisch, P.; Huerlimann, H.; Rindlisbacher, A.; Gsell, L.; Dettwiler, H.; Haettenschwiler, J.; Sieger, E.; Walti, M. The discovery of thiamethoxam: a second-generation neonicotinoid. Pest Manag Sci 2001, 57, 165-176. [CrossRef]
- Honda, H.; Tomizawa, M.; Casida, J.E. Neo-nicotinoid metabolic activation and inactivation established with coupled nicotinic receptor-CYP3A4 and -aldehyde oxidase systems. Toxicol Lett 2006, 161, 108-114. [CrossRef]
- Benzidane, Y.; Touinsi, S.; Motte, E.; Jadas-Hecart, A.; Communal, P.Y.; Leduc, L.; Thany, S.H. Effect of thiamethoxam on cockroach locomotor activity is associated with its metabolite clothianidin. Pest Manag Sci 2010, 66, 1351-1359. [CrossRef]
- Schaefer, P.L.; Ritzmann, R.E. Descending influences on escape behavior and motor pattern in the cockroach. J Neurobiol 2001, 49, 9-28. [CrossRef]
- Buckingham, S.; Lapied, B.; Corronc, H.; Sattelle, F. Imidacloprid actions on insect neuronal acetylcholine receptors. J Exp Biol 1997, 200, 2685-2692. [CrossRef]
- Rosenberg, L.A.; Glusman, J.G.; Libersat, F. Octopamine partially restores walking in hypokinetic cockroaches stung by the parasitoid wasp Ampulex compressa. J Exp Biol 2007, 210, 4411-4417. [CrossRef]
- Lambin, M.; Armengaud, C.; Raymond, S.; Gauthier, M. Imidacloprid-induced facilitation of the proboscis extension reflex habituation in the honeybee. Arch Insect Biochem Physiol 2001, 48, 129-134. [CrossRef]
- Callec, J.J.; Sattelle, D.B. A simple technique for monitoring the synaptic actions of pharmacological agents. J Exp Biol 1973, 59, 725-738. [CrossRef]
- Callec, J.J.; Sattelle, D.B.; Hue, B.; Pelhate, M. Central synaptic actions of pharmacological agents in insects : oil-gap and mannitol-gap studies. In Neurotox 79, Sherwood, M., Ed.; Plenum Press: New York, 1980; pp. 93-100.
- Thany, S.H. Agonist actions of clothianidin on synaptic and extrasynaptic nicotinic acetylcholine receptors expressed on cockroach sixth abdominal ganglion. Neurotoxicology 2009, 30, 1045-1052. [CrossRef]
- Thany, S.H. Thiamethoxam, a poor agonist of nicotinic acetylcholine receptors expressed on isolated cell bodies, acts as a full agonist at cockroach cercal afferent/giant interneuron synapses. Neuropharmacology 2011, 60, 587-592. [CrossRef]
- Ihaka R., G.R. R: a language for data analysis and graphics. J Comput Graph Stat 1996, 5, 299-314. [CrossRef]
- Aliouane, Y.; El Hassani, A.K.; Gary, V.; Armengaud, C.; Lambin, M.; Gauthier, M. Subchronic exposure of honeybees to sublethal doses of pesticides: effects on behavior. Environ Toxicol Chem 2009, 28, 113-122. [CrossRef]
- Nauen, R.; Salgado, V.; Kaussmann, M. Thiamethoxam is a neonicotinoid precursor converted to clothianidin in insects and plants. Pestic Biochem Physiol 2003, 76, 55-69. [CrossRef]
- Nishimura, K.; Kanda, Y.; Okazawa, A.; Ueno, T. relationship between insecticidal and neurophysiological activities of imidacloprid and related compounds. Pest Biochel Physiol 1994, 50, 51-59. [CrossRef]
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu Rev Entomol 2003, 48, 339-364. [CrossRef]
- Tan, J.; Salgado, V.L.; Hollingworth, R.M. Neural actions of imidacloprid and their involvement in resistance in the Colorado potato beetle, Leptinotarsa decemlineata (Say). Pest Manag Sci 2008, 64, 37-47. [CrossRef]
- Benzidane, Y.; Goven, D.; Abd-Ella, A.A.; Deshayes, C.; Lapied, B.; Raymond, V. Subchronic exposure to sublethal dose of imidacloprid changes electrophysiological properties and expression pattern of nicotinic acetylcholine receptor subtypes in insect neurosecretory cells. Neurotoxicology 2017, 62, 239-247. [CrossRef]
- Houchat, J.N.; Taillebois, E.; Thany, S.H. Effects of the DAG analogue 1,2-dioctanoyl-sn-glycerol (DiC8) on nicotine- and clothianidin-evoked currents through alpha-bungarotoxin-insensitive nicotinic acetylcholine receptors expressed on cockroach neurosecretory cells. Neurotoxicology 2020, 78, 143-151. [CrossRef]
- Salgado, V.L. Antagonist pharmacology of desensitizing and non-desensitizing nicotinic acetylcholine receptors in cockroach neurons. Neurotoxicology 2016, 56, 188-195. [CrossRef]
- Bantz, A.; Goven, D.; Siegwart, M.; Maugin, S.; Raymond, V. Exposure to a sublethal dose of imidacloprid induces cellular and physiological changes in Periplaneta americana: Involvement of alpha2 nicotinic acetylcholine subunit in imidacloprid sensitivity. Pestic Biochem Physiol 2022, 181, 105014. [CrossRef]
- Ihara, M.; Matsuda, K.; Shimomura, M.; Sattelle, D.B.; Komai, K. Super agonist actions of clothianidin and related compounds on the SAD beta 2 nicotinic acetylcholine receptor expressed in Xenopus laevis oocytes. Biosci Biotechnol Biochem 2004, 68, 761-763. [CrossRef]
- David, J.A.; R.M., P. The pharmacology of alpha-bungarotoxin-resistant acetylcholine receptors on an identified cockroach motoneurone. J Comp Physiol 1993, 172, 359-368. [CrossRef]
- Salgado, V.L.; Saar, R. Desensitizing and non-desensitizing subtypes of alpha-bungarotoxin-sensitive nicotinic acetylcholine receptors in cockroach neurons. J Insect Physiol 2004, 50, 867-879. [CrossRef]
Figure 1.
(A) Structure of the chloropyridylmethyl compound acetamiprid. (B) Schematic representation of the open-field-like apparatus. 28 infrared light-emitting diodes (black circles) enable the detection of cockroaches’ position through the device. At the beginning of the experiment, the tested cockroach was placed in the cylinder (a) and blocked by closed trap door in this starting area. When the door open, the cockroach’s movements were recording continuously using the numerical counter, allowing its position, speed, and the duration of immobility to be determined.
Figure 1.
(A) Structure of the chloropyridylmethyl compound acetamiprid. (B) Schematic representation of the open-field-like apparatus. 28 infrared light-emitting diodes (black circles) enable the detection of cockroaches’ position through the device. At the beginning of the experiment, the tested cockroach was placed in the cylinder (a) and blocked by closed trap door in this starting area. When the door open, the cockroach’s movements were recording continuously using the numerical counter, allowing its position, speed, and the duration of immobility to be determined.
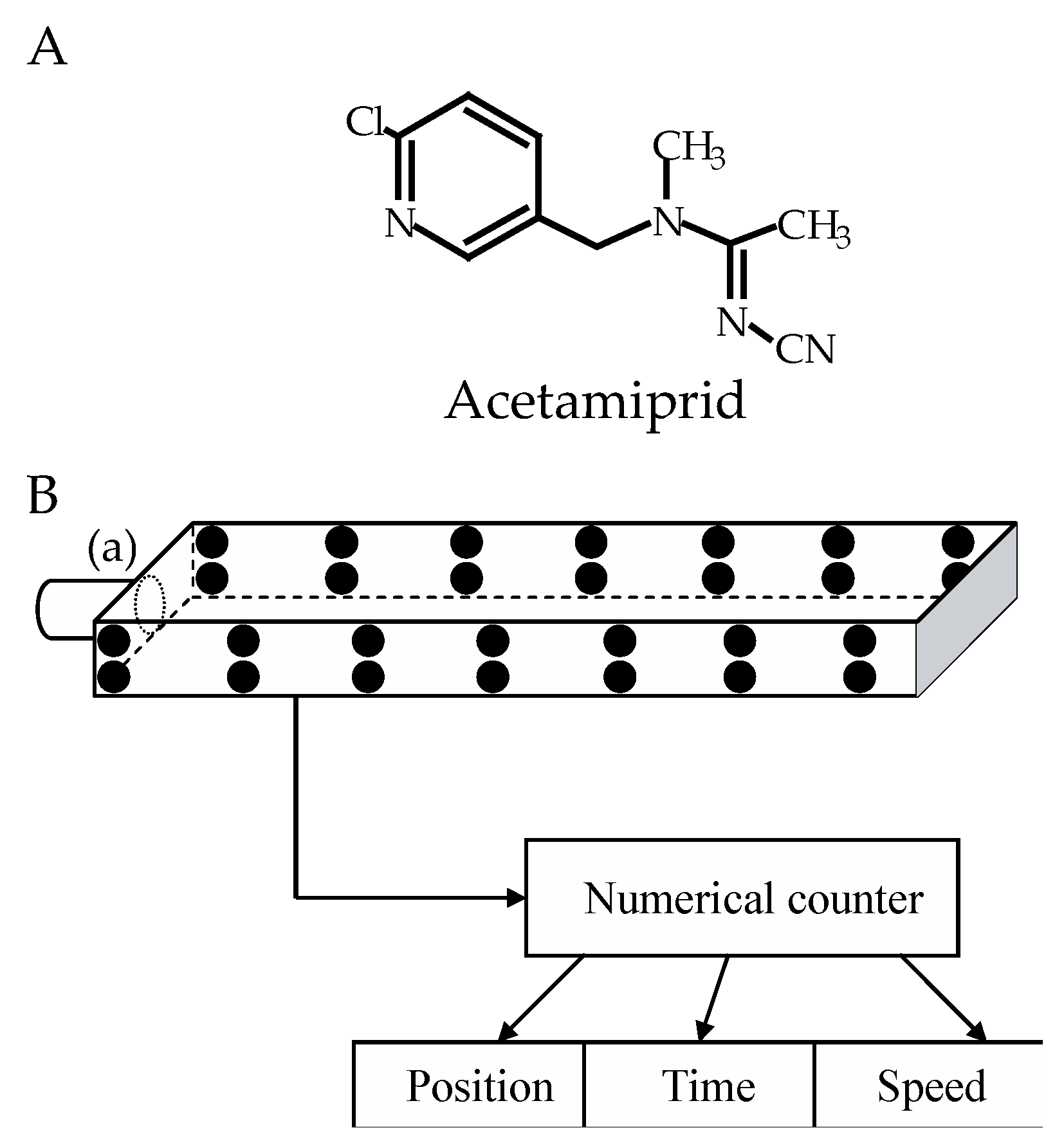
Figure 2.
Proportion of cockroaches eliciting locomotor activity 1 h, 24 h, and 48 h after acetamiprid injection into the haemolymph. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
Figure 2.
Proportion of cockroaches eliciting locomotor activity 1 h, 24 h, and 48 h after acetamiprid injection into the haemolymph. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
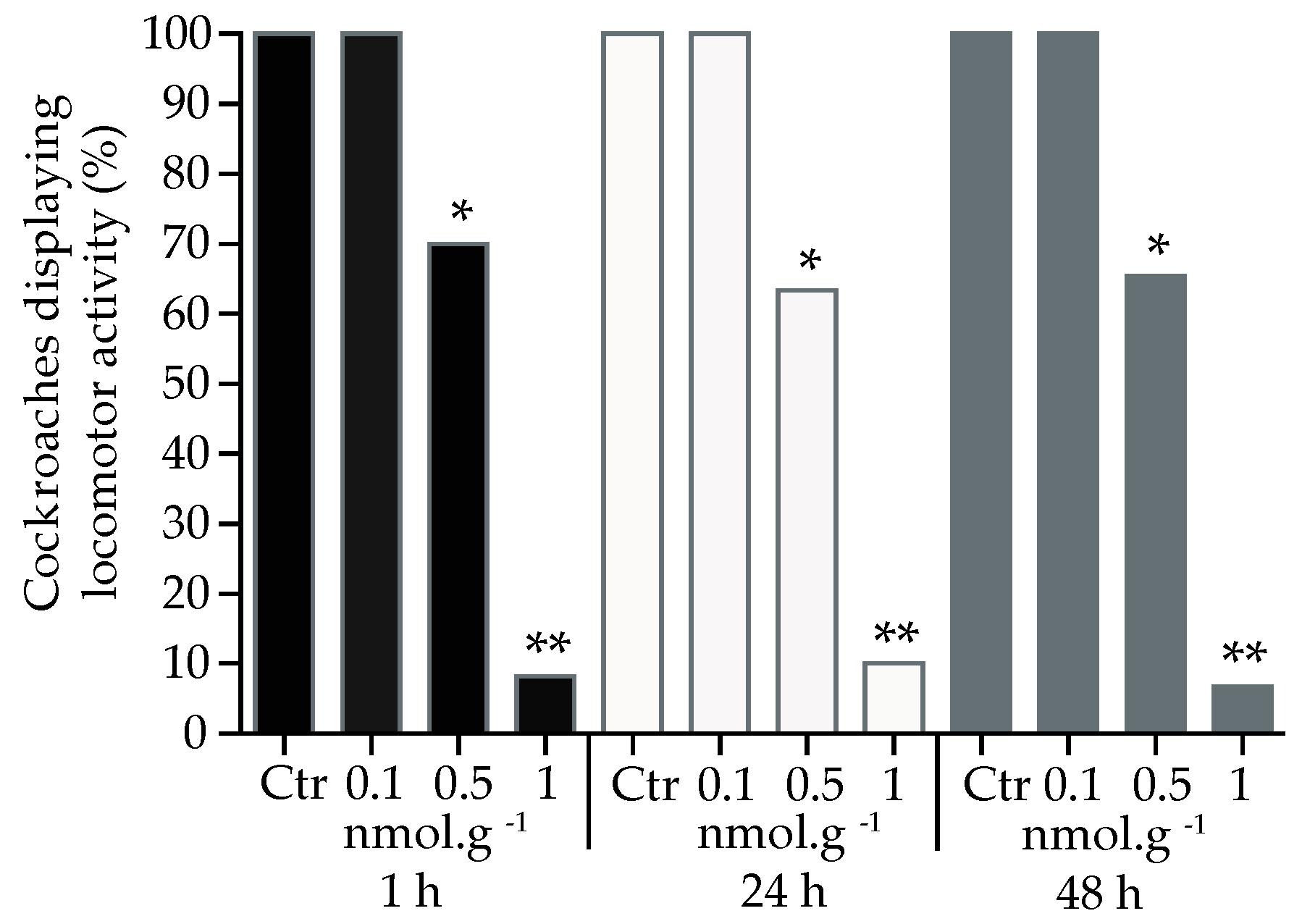
Figure 3.
Proportion of cockroaches eliciting locomotor activity 1 h, 24 h and 48 h after topical application of acetamiprid. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
Figure 3.
Proportion of cockroaches eliciting locomotor activity 1 h, 24 h and 48 h after topical application of acetamiprid. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
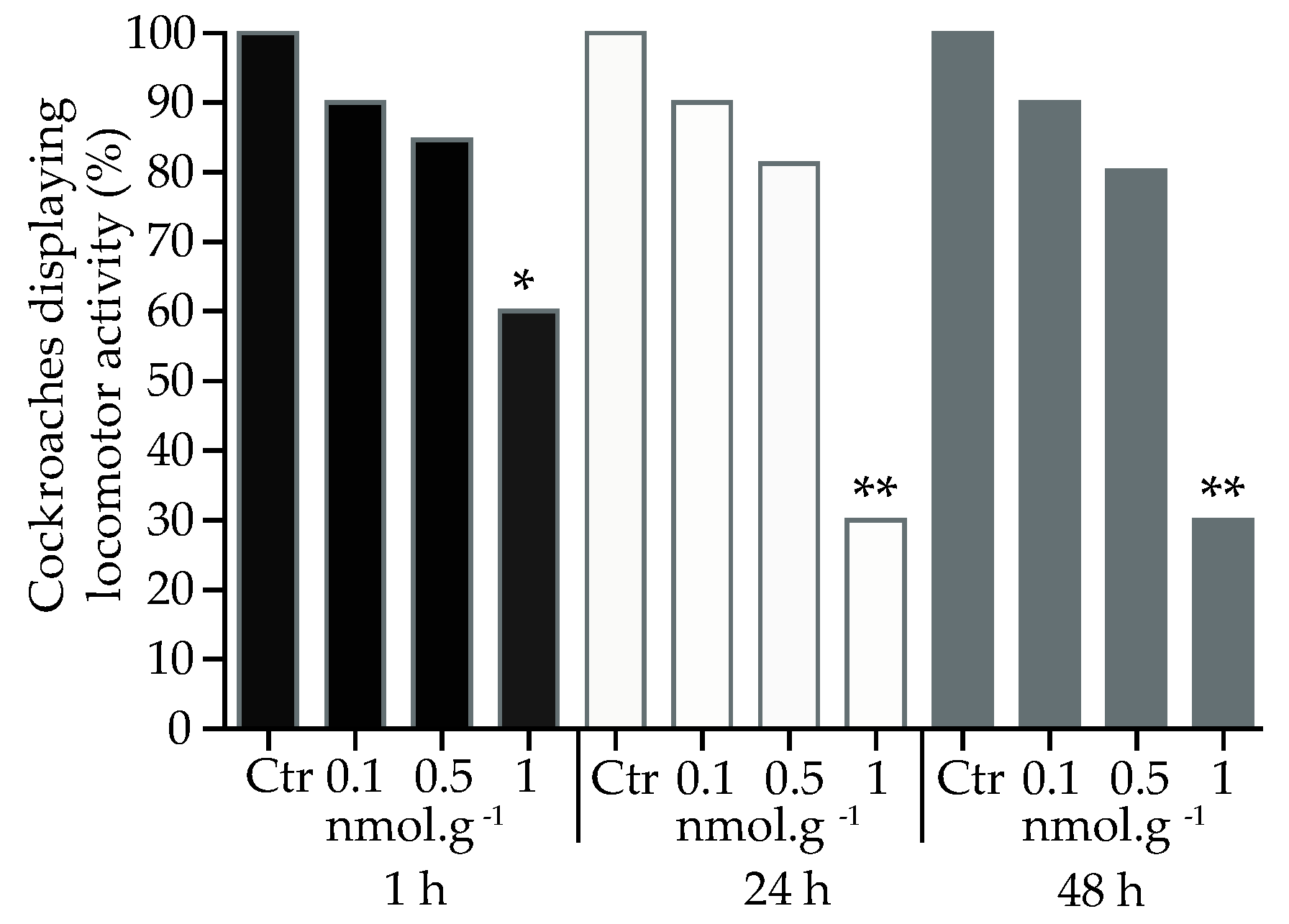
Figure 4.
Proportion of cockroaches eliciting locomotor activity after 1 h, 24 h, and 48 h oral application of acetamiprid. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
Figure 4.
Proportion of cockroaches eliciting locomotor activity after 1 h, 24 h, and 48 h oral application of acetamiprid. For each condition, 20 cockroaches were tested. Control group (Ctr). Significant differences between acetamiprid treatment and control, using Student t-test, are indicated with asterisks (* p < 0.05 and ** p < 0.001).
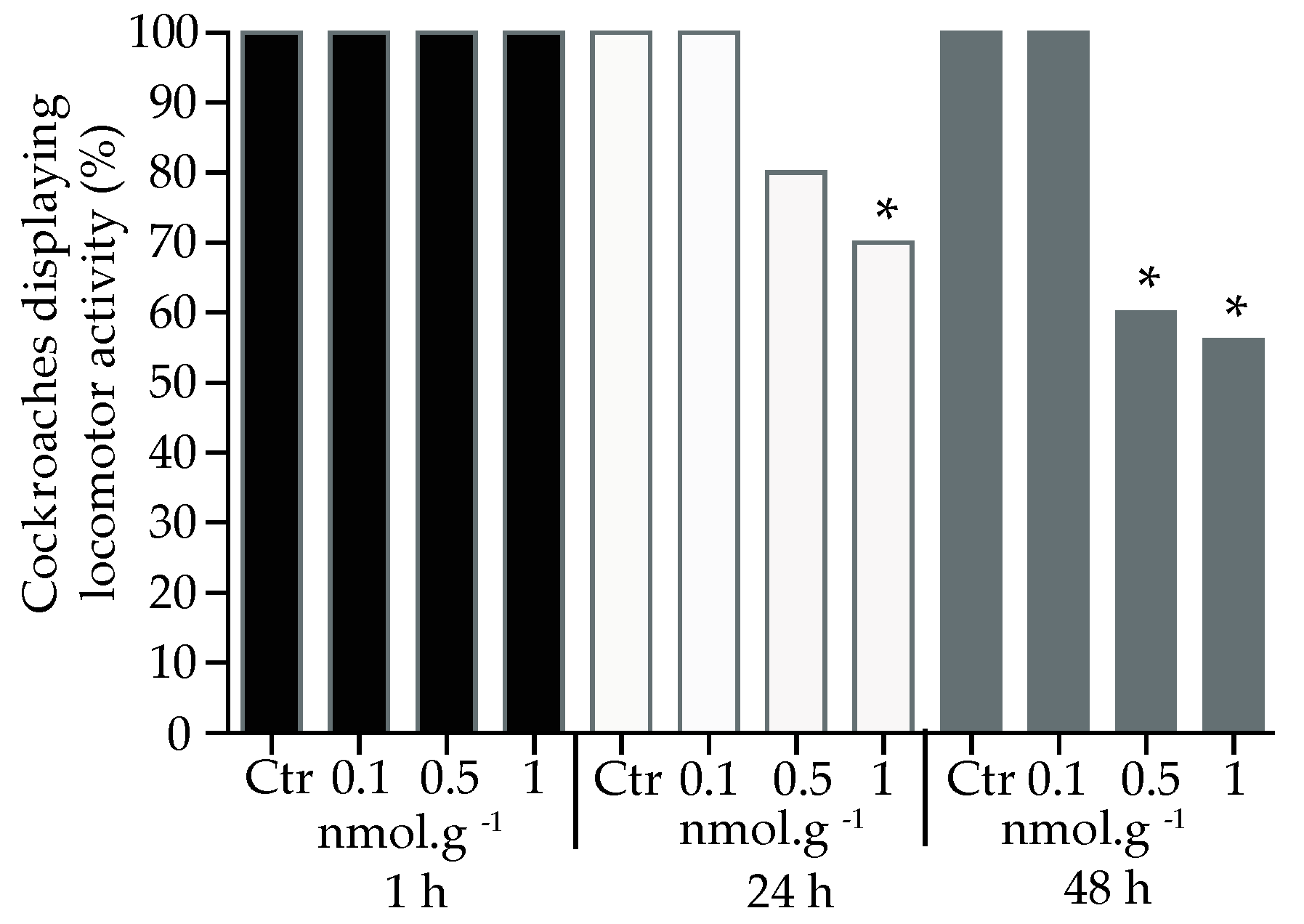
Figure 5.
Illustration of the morphological aspect of cockroach after 1 nmol.g-1 haemolymph application of acetamiprid (on the left) compared to control cockroach (on the right).
Figure 5.
Illustration of the morphological aspect of cockroach after 1 nmol.g-1 haemolymph application of acetamiprid (on the left) compared to control cockroach (on the right).

Figure 6.
Mean duration of immobility of cockroaches after acetamiprid treatment. The duration of immobility was measured during 5 min of observation in the open-field-like apparatus. Recordings were obtained 1 h after acetamiprid exposure through, (A) haemolymph, (B) topical and (C) oral applications. Significant differences between treated and control conditions are marked with asteriks (* p < 0.05, ** p < 0.01, Student t-test with Welch’s correction).
Figure 6.
Mean duration of immobility of cockroaches after acetamiprid treatment. The duration of immobility was measured during 5 min of observation in the open-field-like apparatus. Recordings were obtained 1 h after acetamiprid exposure through, (A) haemolymph, (B) topical and (C) oral applications. Significant differences between treated and control conditions are marked with asteriks (* p < 0.05, ** p < 0.01, Student t-test with Welch’s correction).
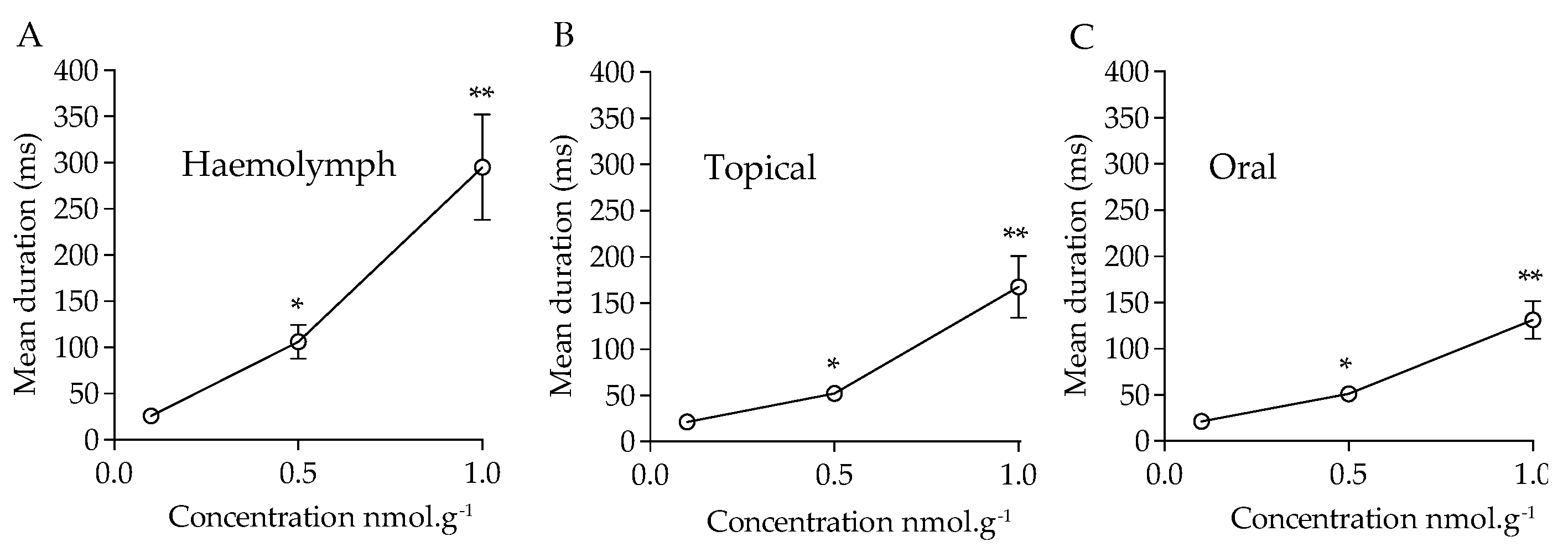
Figure 7.
Mannitol-gap recording studies. Electrical stimulation of the nerve XI on treated cockroaches with 1 nmol.g-1 acetamiprid through haemolymph injection (A), topical (B) and oral (C) applications. Electrical stimulation of the nerve XI were recorded 20 min after locomotor activity in the open-field-like apparatus.
Figure 7.
Mannitol-gap recording studies. Electrical stimulation of the nerve XI on treated cockroaches with 1 nmol.g-1 acetamiprid through haemolymph injection (A), topical (B) and oral (C) applications. Electrical stimulation of the nerve XI were recorded 20 min after locomotor activity in the open-field-like apparatus.
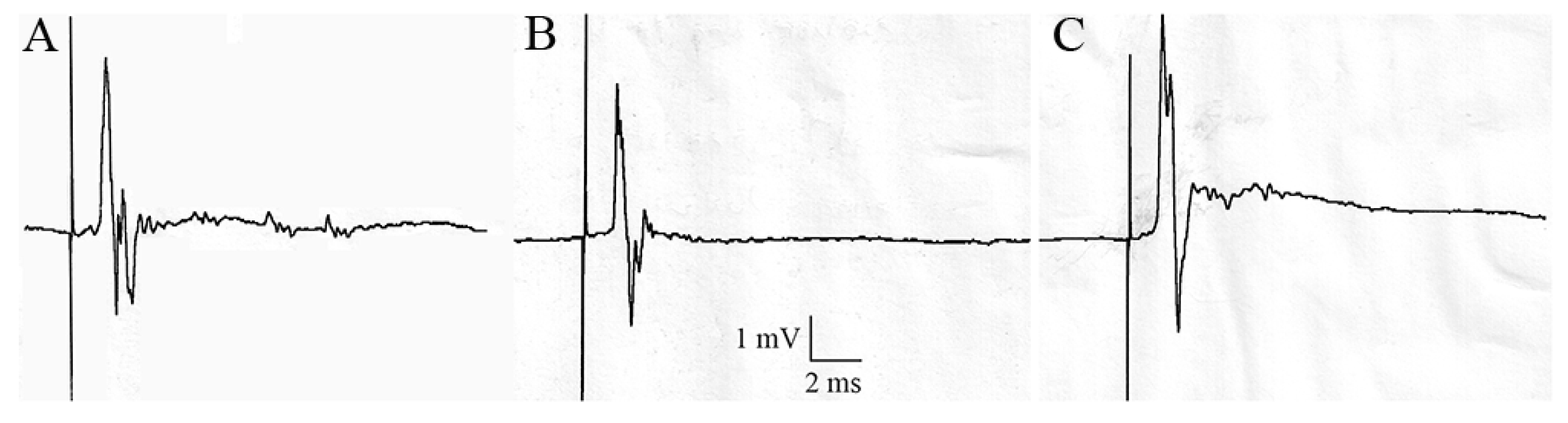

Table 1.
Time needed for the cockroaches to to cross the experimental device, 1h after acetamiprid treatment.
Table 1.
Time needed for the cockroaches to to cross the experimental device, 1h after acetamiprid treatment.
| Acetamiprid (nmol.g-1) | |||
| 0.1 | 0.5 | 1 | |
| Haemolymph injection | 6.4 ± 1.7 s | 129.3 ± 25.0 s | 319.0 ± 67.0 s |
| Topical application | 6.4 ± 2.5 s | 74.4 ± 2.0 s | 189.0 ± 3.0 s |
| Oral application | 6.0 ± 2.0 s | 73.4 ± 17.0 s | 137.7 ± 38.0 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



