
Submitted:
05 November 2023
Posted:
06 November 2023
You are already at the latest version
Abstract
The genus Colias Fabricius, 1807 includes numerous taxa and forms with uncertain status and taxonomic position. Among such taxa are Colias mongola Alphéraky, 1897 and Colias tamerlana Staudinger, 1897, interpreted in the literature either as conspecific forms, as subspecies of different but morphologically somewhat similar Colias species, or as distinct species-level taxa. Based on mitochondrial and nuclear DNA markers, we reconstruct a phylogeographic pattern of the taxa in question. We recover and include in our analysis DNA barcodes of the century-old type specimens, the lectotype of C. tamerlana deposited in the Natural History Museum (Museum für Naturkunde), Berlin, Germany (ZMHU), the paralectotype of C. tamerlana and the lectoptype of C. mongola deposited in the Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia (ZISP). Our analysis grouped all specimens within four (HP_I-HP-IV) deeply divergent but geographically poorly structured clades which did not support nonconspecifity of C. mongola–C. tamerlana. We also show that all studied females of the widely distributed haplogroup HP_II were infected with a single Wolbachia strain belonging to the supergroup B, while the males of this haplogroup, as well as all other investigated specimens, were not infected. Our data underline the relevance of large-scale sampling dataset analysis and the need of testing for Wolbachia infection to avoid erroneous phylogenetic reconstructions and species misidentification.
Keywords:
Colias
; DNA barcoding
; Lepidoptera
; molecular taxonomy
; PCR screening
; Wolbachia
1. Introduction
Colias Fabricius, 1807 (Lepidoptera, Pieridae) is one of the largest pierid genera, with approximately 90 described species [1,2,3,4]. The genus has a predominantly Holarctic distribution, with a few species occurring in the Afrotropical, Oriental and Neotropical biogeographical regions [4,5,6,7]. Although Colias butterflies are among the most spectacular and well-known Lepidoptera, which historically attracted much attention of researchers and collectors, their taxonomy is still poorly elucidated. Colias butterflies may exhibit a significant level of interspecific variation in the wing pattern. At the same time, many allopatric taxa traditionally treated as separate species may have very similar phenotypes. In addition, such taxonomically important characters as genitalia structures, commonly used for species delimitation in insects, are largely uniform in this group and do not possess reliable diagnostic features [1]. Consequently, systematics and real taxonomic status (species, subspecies or interspecific forms) of many Colias taxa are a matter of debate.
The issues mentioned above can be applied to Colias tamerlana Staudinger, 1897 (Figure 1a) and Colias mongola Alphéraky, 1897 (Figure 1b) distributed in Western Mongolia, Northewestern China and South Siberian part of Russia [8,9]. Contradictory taxonomic status has been suggested for this pair of taxa in literature, from synonyms or subspecies of several distinct species to separate species-level taxa (Figure 2). The former taxon was described as a separate species by Otto Staudinger in 1897 from China, Xinjiang, East Tian Shan, north of Hami, Karlik Shan Mts. ["im ostlichsten Thian-Schan-Gebirge (nordlich von Chamyl)"] [10]. The latter one was described by Alphéraky as a variety of Colias nastes Boisduval, 1832 [Colias Nastes B. var. Mongola Alph. var. nova] in 1897 [11]. The type locality of C. mongola has been the subject of considerable debates and contradictory interpreted in the taxonomic literature (Figure 3b). Korb [12] in his paper devoted to the taxonomy of Colias cocandica Erschoff, 1874 and allied taxa placed the type locality of C. mongola a few km south of Ulan-Bator (Mongolia). Grieshuber and co-authors [3] suggested that the type series of C. mongola originates from the vicinity of Uliastai (Mongolia), at a distance of ca. 750 km to the west from the locality indicated by Korb [12], but subsequently corrected the area to the SE Khangai Mts. [1]. Gorbunov [13] indicated the environs of the village Turan (Republic of Buryatia, Russia) as the type locality of C. mongola, an area located more than 500 km away from the localities suggested by Korb [12] and Greishuber [1,3]. Finally, Weiss [14] erroneously mentioned the South Altai (Kuray, Chuya, Kobdo) as the type locality of C. mongola.
The nomenclatural priority of the name C. tamerlana over C. mongola [13] and taxonomic status of these taxa are also disputable [2,12,13,14,15,16,17,18,19,20]. Staudinger treated C. tamerlana and C. mongola as synonyms [15]. Greishuber & Lamas [2] pointed out weakly expressed morphological differences between the two taxa and speculated that molecular analysis could provide evidences for synonymizing C. tamerlana and C. mongola. Talbot [16] in his “Lepidopterorum Catalogus” placed C. tamerlana as synonym of C. cocandica maja Grum-Grshimailo, 1891 without providing further explanations. Korb [12], based on examinations of genitalia, concluded that mongola and tamerlana represent distinct species. Tshikolovets and co-authors [17] regarded C. mongola as a subspecies of C. cocandica. These two taxa are thought to be biogeographically isolated (the eastern limit of C. cocandica distribution range lies in SE Kazakhstan (South Dzungaria) and NW China (Borohoro Mts.)) [3], thus the conclusion of their conspecificity is rather doubtful. It should be noted that butterflies from Lake Khovsgol area (Mongolia), currently considered as easternmost known population of C. tamerlana/mongola, were originally referred to much westerly distributed C. cocandica and were described as a subspecies of the latter, C. cocandica sidonia Weiss, 1968 [14]. Some authors regarded this taxon as subspecies of Colias nastes Boisduval, 1832, without considering the relationships between C. mongola and C. tamerlana and their taxonomic status [18,19,20,21], while mentioning that C. mongola was formerly treated as separate species.
To some extent, this assumption was based on the discovering of a large series of specimens, later described as Colias nastes jacutica Kurentsov 1970, which phenotypically somewhat resemble sidonia, the taxon that was provisionally considered a valid subspecies of C. tamerlana/mongola. In the original description [21], Colias nastes jacutica is interpreted as an intermediate link between nastes and mongola (sensu lato), yet this assumption requires further confirmation (in particular, by applying molecular methods), given that sidonia and nastes are separated by more than 2500 km without any record for the species in the gap. The second subspecies of C. tamerlana/mongola, C. mongola ukokana Korb & Yakovlev, 2000, was described from the Ukok Plateau (Republic of Altai, Russia)[22], but currently ukokana is considered a synonym of the nominotypical mongola [1,2,3].
The correct delineation and identification of species is not always possible based on characters of external morphology alone, especially in such taxonomically challenging groups as Colias [23,24,25,26]. Over the last decades, the rapid development and implementation of new molecular techniques and approaches validated mitochondrial (e.g. DNA barcodes) and nuclear DNA markers as a useful and efficient tool for species identification, detecting and analyzing cryptic diversity, revealing interspecific and deep-level relationships and phylogenetic structure in taxonomically challenging groups of insects, solving long-standing taxonomic problems [23,27,28,29,30,31,32,33,34,35,36,37,38,39]. Despite broad and constantly growing usage of DNA-based techniques in butterfly taxonomy and molecular systematics, surprisingly few studies focused so far on the genus Colias [6,42,43,44,45,46]. Moreover, all but a few research aimed at inferring Colias phylogeny or relating to molecular aspects of the genus were based on a limited set of taxa and/or poor specimen sampling. Some recent studies and our unpublished data suggested that morphologically well-differentiated Colias species may have nearly identical or even shared COI (cytochrome c oxidase subunit 1 gene) haplotypes. For instance, D’Ercole and co-authors [47] showed that all 22 North American Colias species shared at least one of the revealed barcode sequences with another species. Conversely, two or more deeply diverged mitochondrial lineages (e.g. DNA barcodes) may be found within the same species [42]. In such cases, imbalanced taxa sampling, and/or testing only a small number of specimens per species could obscure the true phylogenetic relationships within the genus, and lead to species misidentification and incorrect taxonomical conclusions. To overcome this problem, all Colias taxa should be analysed in detail to ascertain their actual taxonomic status and avoid erroneous interpretations.
Here, we analysed three nuclear genes, Ca-ATPase (sarco/endoplasmic reticulum calcium ATPase), H3 (Histone h3) and CAD (Carbamoyl-Phosphate Synthetase 2, Aspartate Transcarbamylase and Dihydroorotase), as well as the barcoding region of the mitochondrial COI gene for a large set of C. mongola and C. tamerlana samples, including recovered for the first time DNA barcodes of the century-old type specimens. We also conducted PCR screening for three Wolbachia genes of the collected C. mongola and C. tamerlana butterflies in order to reveal patterns of Wolbachia infection.
Basing on a large-scale sampling dataset, our study aimed to (i) reveal the phylogeographic structure of C. mongola and C. tamerlana, (ii) clarify their taxonomic status, contradictory interpreted in the literature, (iii) shed light on the role of Wolbachia in evolutionary history and observed biogeographic patterns of the taxa in question.
2. Materials and Methods
2.1. Taxon Sampling
Ninety-one specimens of C. mongola and C. tamerlana were collected in various localities of Western Mongolia, Northwestern China, and South Siberian part of Russia (Figure 3a), covering the known distribution ranges of these taxa. All sampling sites were partitioned into eighteen populations (Figure 3b). Samples either were preserved in 96% alcohol or were kept dry for subsequent molecular analysis. The lectotype of C. tamerlana deposited in the Natural History Museum (Museum für Naturkunde), Berlin, Germany (ZMHU), the paralectotype of C. tamerlana and the lectotype of C. mongola deposited in the Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia (ZISP), four specimens of C. tamerlana sidonia from the type locality deposited in the Zoological Museum of the Moscow State University, Moscow, Russia (ZMMU), and three samples mined from BOLD database (http://www.boldsystems.org, accessed on 03 September 2022) were included in the sampling dataset. Thus, the final dataset included 101 specimens. The list of the specimens used for the molecular analysis with identification codes and collection data is given in Table 1.
2.2. DNA Extraction
One leg from each specimen was taken for DNA extraction. For the samples more than 10 years old, total genomic DNA was extracted using QIAamp DNA Investigator Kit (Qiagen, Venlo, The Netherlands) following the manufacturer’s protocol. For the specimens up to 10 years old, DNA extraction was performed using the CTAB-based method [48] with some modifications [39,49].
2.3. Molecular Markers, PCR Amplification, and Sequencing
One mitochondrial (COI) and three nuclear (Ca-ATPase, H3, and CAD) genes were used as molecular markers. A 658 bp fragment of the COI gene (mitochondrial DNA barcode) was amplified using LCO1490/HCO2198 [50] and LepF/LepR primer pairs [51]. In case standard lepidopteran barcode primers failed to yield a sufficient product, we amplified full-length barcode fragment using the primer pair combinations LepF/MH-MR1 + MH-MF1/LepR and LCO1490/MH-MR1 + MH-MF1/HCO2198 [52]. Primers CAD743nF/CADmidR, CADmidF/CAD1028R [53], H3aF/H3aR [54], and Ca-ATPase_F/Ca-ATPase_R [55] were used for nucDNA amplification and resulted in 847 bp fragment of the CAD, 328 bp fragment of the H3, and 445 bp fragment of the Ca-ATPase genes, respectively.
The PCR amplifications were performed in a 15 µL reaction volume per sample. Each reaction contained 1 µL template DNA (ca. 10–50 ng genomic DNA), 0.9 µL of both forward and reverse primers diluted to a standard concentration of 10 µM, 3 µL of 5× ScreenMix (Evrogen, Moscow, Russia), and 9.2 µL of ddH2O. The temperature profile for COI, CAD, and Ca-ATPase genes was as follows: initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C (COI)/55 °C (CAD, Ca-ATPase)/60 °C (H3) for 30 s, and extension at 72 °C for 1 min 30 s, with a final extension at 72 °C for 10 min. The purified PCR products were subjected to the further sequencing. Sequencing of the double-stranded product was carried out at the Research Resource Center for Molecular and Cell Technologies (St. Petersburg State University, St. Petersburg, Russia) using ABI 3500xL analyzer (Applied Biosystems, Waltham, MA, USA).
2.4. Processing and Sequencing of Old Type Specimens
To obtain COI barcodes of the century-old type specimens we followed the protocol described in detail by Li et al. [56]. In brief, a single leg was used for DNA extraction. The DNA isolation protocol was non-destructive: the entire leg was soaked in a DNA extraction solution overnight and was preserved after the extraction. Genomic libraries were constructed from total DNA and sequenced for 150 bp from both ends on Illumina HiSeq x10 (Illumina, San Diego, CA, USA) in a pool with others. The sequences were demultiplexed to assign to each specimen by the index and indices removed. The barcodes were assembled using a reference sequence as a bait. All sequence reads matching the reference were mapped to it using DIAMOND [57]. The coverage of the barcode sequence is typically high (100 to 1000-fold or more), and overlapping reads result in an unambiguous sequence. Further details of experimental and computational protocols can be found in [56].
2.5. Detection of Wolbachia Endosymbionts
Colias specimens were screened for the presence of Wolbachia infection by amplifying three Wolbachia genes, 16S ribosomal RNA (16S), Wolbachia surface protein (wsp) and Filamentation temperature-sensitive protein Z (ftsZ). We used Wolbachia-specific primer pairs, W-Specf/W-Specr [58], wsp81F/wsp691R [59] and ftsZ-F/ftsZ-R [60], amplifying ~ 396 bp fragment of the 16S RNA gene, ~ 549 bp fragment of the wsp gene and ~ 510 bp fragment of the ftsZ gene (actual length of PCR fragments may vary, depending on the individual Wolbachia strain), respectively. The PCR amplifications were performed in a 15 µL reaction volume. Each reaction contained 1 µL template DNA (ca. 10–50 ng genomic DNA), 0.8 µL of both forward and reverse primers diluted to a standard concentration of 10 µM, 3 µL of 5× ScreenMix (Evrogen, Moscow, Russia) and 9.4 µL of ddH2O. The temperature profile for 16S, wsp, and ftsZ genes was as follows: initial denaturation at 95 °C for 5 min, followed by 40 cycles of 30 s denaturation at 95 °C, 1 min annealing at 50 °C and extension at 72 °C for 45 s, with a final extension at 72 °C for 5 min. Each PCR reaction contained two negative (PCR mix with ddH2O instead of DNA sample) and one positive (genomic DNA of a Wolbachia-infected Colias specimen, previously successfully amplified for 16S, wsp, and ftsZ genes) controls. PCR amplification was conducted twice for each specimen in order to avoid the technical errors. To ascertain the presence/absence of Wolbachia, each PCR product was checked on 1% standard agarose gel. Our personal unpublished data suggests that the standard screening procedure for Wolbachia allows detecting infection in host specimens up to 30-35 years old, but its sufficiency largely depends on the quality of the genomic DNA and storage conditions of the specimens. Thus, to avoid false-negative results, we excluded from the analysis samples collected more than twenty years ago (before the 2000s). Specimens positive for three Wolbachia genes were sequenced at the Research Resource Center for Molecular and Cell Technologies (St. Petersburg State University, St. Petersburg, Russia) using ABI3500xL Genetic Analyzer (Applied Biosystems, Waltham, MA, USA).
2.6. Molecular Data Analysis and Phylogenetic Reconstructions
All sequences were checked for errors, edited and aligned using Geneious v.8.1.6 [61] and BioEdit v.7.0.3 [62] software. Primer sequences were cropped. The final COI dataset (alignment length 658 bp) included 97 sequences of C. mongola/C. tamerlana. Sequences of pierids Leptidea juvernica (Linnaeus, 1758) and Colias croceus (Geoffroy, 1785) (GenBank accession numbers MT210323 and OR178497, respectively) obtained previously were included as an outgroup to root the phylogram. COI sequences were collapsed to unique haplotypes using online tool FaBox v.1.61 (https://birc.au.dk/~palle/php/fabox/, accessed on 05 November 2022) [63]. Concatenated nuclear data set (alignment length 1620 bp) included sequences of 17 C. mongola/C. tamerlana specimens representing all revealed COI haplotypes. Phylogenetic reconstructions placed C. croceus within basal lineages of the genus Colias, thus it was selected as the outgroup to root the nuclear phylogram [42,43]. Nucleotide substitution models for each dataset were estimated based on the Bayesian information criterion (BIC) using jModelTest v.2.1.10 [64]. The best-fitting models were as follows: JC for COI data set, HKY+I for concatenated Ca-ATPase+H3+CAD nuclear genes fragments. Bayesian analyses were performed using the MrBayes v.3.2.7a software [65]. Parameters were estimated using two independent runs of 10 million generations each with four simultaneous chains (one cold and three heated). The sampling of trees and parameters was set to every 1000 generations. The first 10% of trees were discarded as burn-in prior to computing a consensus phylogeny and posterior probabilities. TRACER v.1.6 was used for checking stationarity and convergence of Bayesian analyses between runs [66]. The consensuses of the obtained trees were visualized using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 05 November 2022). For the analysis of the phylogeographical structure of C. mongola and C. tamerlana, a median-joining haplotype network [67] was built using popART v.1.7 software [68]. Genetic distances among COI barcodes were calculated using MEGA v.7.0.14 [69]. The number of polymorphic and parsimony informative sites, the number of haplotypes, haplotype (h) and nucleotide (π) diversities were calculated in DnaSP v.6.12.03 [70]. DnaSP was further used to infer the demographic history of C. mongola and C. tamerlana with Tajima’s D [71], Fu and Li’s D [72], Fu and Li’s F [72[, and Fu’s Fs [73] statistical tests for neutrality.
2.6.1. Molecular Characterization of Wolbachia
The BLAST algorithm implemented in NCBI (https://blast.ncbi.nlm.nih.gov, accessed on 20 July 2023) was used to search for sequence similarities in GenBank database with known DNA (BLASTN) sequences. We mined 43 16S, 74 wsp, and 71 ftsZ sequences with the highest percentage identity match, which were included in the alignment datasets. In the final alignments, identical Wolbachia sequences of the same host species were limited to one record. To estimate relationships among Wolbachia alleles, phylogenetic analyses were conducted for each gene independently using the Bayesian inference (BI) approach, applying GTR+G (for 16S gene fragment) and GTR + I +G (for wsp and ftsZ genes fragments) substitution models, as suggested by jModelTest v.2.1.7 [64]. All other parameters of the Bayesian analyses were the same as for COI alignment.
2.7. Data Availability
All sequences obtained for COI, CAD, Ca-ATPase and H3 Colias genes, and for 16S, wsp and ftsZ Wolbachia genes were deposited to GenBank under accession numbers OP946559–OP946652, OR178498–OR178501, OR178497 (COI), OQ192178–OQ192194 (CAD), OQ1921161–OQ192177 (Ca-ATPase), OQ192144–OQ192151 (H3), OQ155222–OQ155235 (16S), OQ192116–OQ192129 (wsp) and OQ192130–OQ192143 (ftsZ) and listed in the electronic supplementary materials (Table S1).Voucher specimens were deposited in the Department of Karyosystematics of the Zoological Institute of the Russian Academy of Sciences and private collections of A. Kir’yanov, A. Krupitsky, A. Kurmaev, S. Churkin (Moscow, Russia), B. Khramov (Saint-Petersburg, Russia) and R. Yakovlev (Barnaul, Russia).
3. Results
3.1. Phylogeographic Structure of Colias mongola and Colias tamerlana
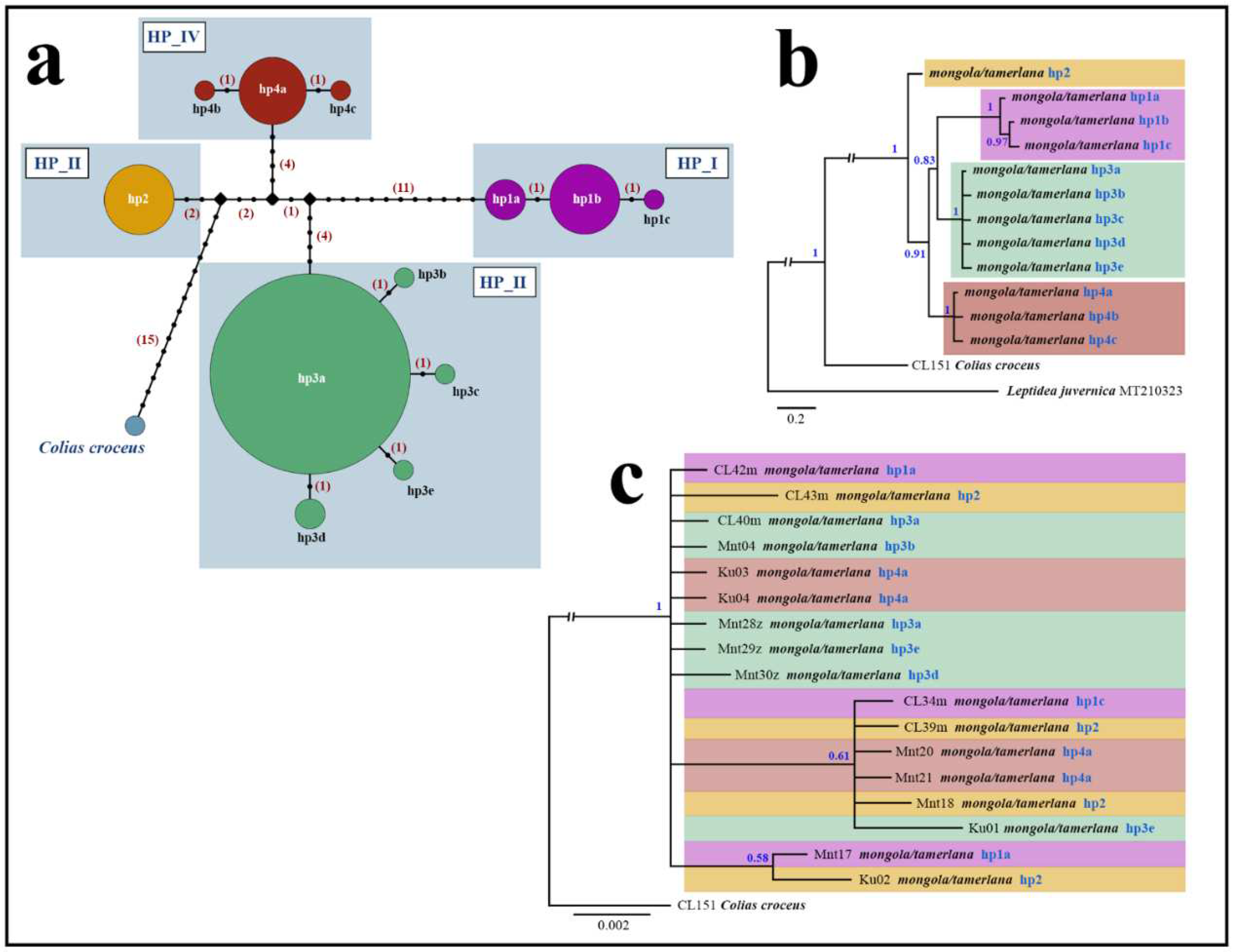
Haplotype analysis of a dataset of 101 C. mongola and C. tamerlana specimens revealed 12 COI haplotypes clustering in four distinct haplogroups (HP_I – HP_IV) (Figure 4a). Each haplogroup consists of one main haplotype and one to four satellites, differing from the main haplotypes in one nucleotide substitution. The only one exception is haplogroup HP_II, which consists of a single haplotype hp2. Low rate of genetic diversity ranging from 0% to 0.07%±0.05% detected within each haplogroup. On the contrary, sequence divergence between haplogroups is relatively high (1.23%±0.41% – 2.56%±0.62%), with a maximum p-distance of 2.89%± 0.62% between the most divergent haplotypes (Table 2).
Figure 3.
Maps showing location of the study area (red rectangle) (a), sampling localities of analysed specimens of C. mongola/C. tamerlana (b), and geographical distribution of reveled COI haplogroups of C. mongola/C. tamerlana (c-f). Question marks indicate suggested type localities of C. mongola (see text for explanation). Wolbachia infected populations indicated by "+”.
Figure 3.
Maps showing location of the study area (red rectangle) (a), sampling localities of analysed specimens of C. mongola/C. tamerlana (b), and geographical distribution of reveled COI haplogroups of C. mongola/C. tamerlana (c-f). Question marks indicate suggested type localities of C. mongola (see text for explanation). Wolbachia infected populations indicated by "+”.
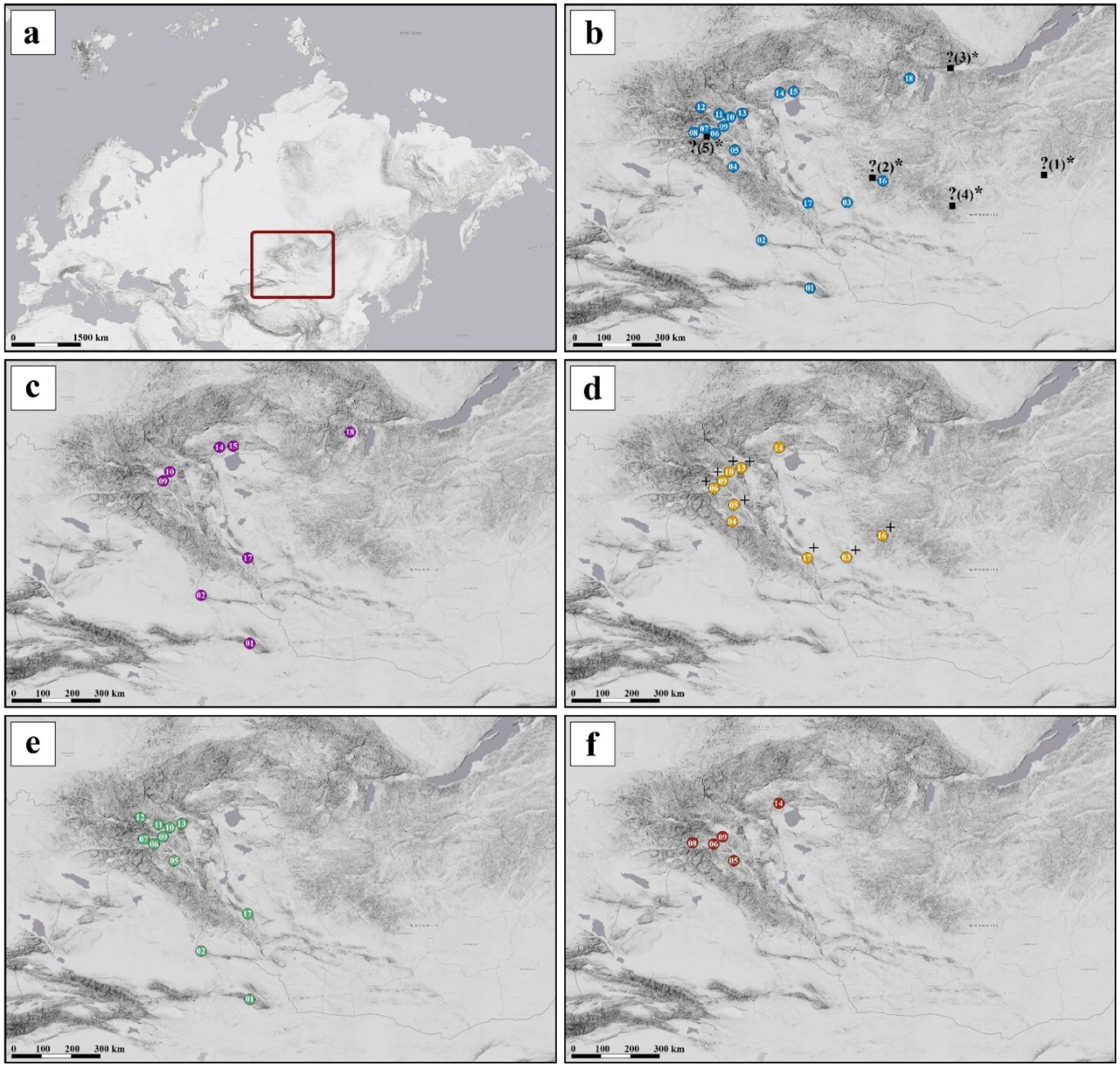
In general, haplotype distribution demonstrates no clear geographical structure: haplotypes of all four haplogroups are commonly occurred in sympatry; at the same time, they can be found in the geographically remote populations (Figure 3c-f). Twenty-six specimens clustered in three haplotypes of the haplogroup HP_I which was shared by 8 populations. Seventeen specimens constituted a single haplotype of the haplogroup HP_II which occurred in 10 sampling sites. Forty-two specimens were grouped into five haplotypes of the haplogroup HP_III and sixteen specimens constituted three haplotypes of the haplogroup HP_IV, which were found in 11 and 4 sampling sites, respectively. Four haplotypes, namely hp1b, hp2, hp3a and hp4a, were the most commonly observed haplotypes out of all screened individuals. On the contrary, haplotypes hp1c, hp3b, hp3c, hp4b, hp4c consisted of only one specimen each were found in single sampling sites. Detailed data on the distribution of the haplotypes among specimens and localities is given in the Table 3.
3.2. Phylogenetic Analyses of Mitochondrial and Nuclear Markers
The Bayesian phylogenetic tree for C. mongola/C. tamerlana COI haplotypes displayed four strongly supported (PP=1) lineages (Figure 4b) corresponding to four haplogroups revealed by haplotype network analysis. Within the C. mongola/C. tamerlana clade, a basal position is occupied by haplotype hp2, which is common and widespread across the geographical area studied. Further splitting recovers the clade containing specimens of HP_IV haplogroup. This clade in turn appeared as sister to the remaining two clusters, consisted of haplotypes hp3a-hp3e and hp1a-hp1c, respectively.
Combined analysis of three nuclear markers (Ca-ATPase, H3, and CAD) resulted in an unresolved tree for C. mongola/C. tamerlana that failed to recover clades revealed by BI analysis of COI barcodes (Figure 4c). In general, nuclear markers demonstrate very shallow divergence forming on the phylogenetic reconstructions two unsupported clades (PP=0.58; PP=0.61), each consisted of specimens bearing COI haplotypes of different haplogroups. Both intra-individual heterogeneities and single nucleotide substitutions were found in sequenced fragments of Ca-ATPase, CAD and H3 nuclear genes among the 17 analysed specimens of C. mongola/C. tamerlana. However, polymorphic sites were distributed inhomogeneously across the three genes fragments. The 847 bp fragment of the CAD was most variable with 23 heterogeneities/substitutions found, whereas 445 bp fragment of the Ca-ATPase gene and 328 bp fragment of the H3 gene were conserved, having four and one polymorphic sites, respectively. The detailed information on the nucleotide variability of the studied nuclear genes fragments is given in the electronic supplementary (Table S2).
Figure 4.
Phylogenetic patterns of C. mongola/C. tamerlana based on the analysis of mitochondrial (COI) and nuclear (Ca-ATPase, H3, and CAD) markers. (a) Median-joining haplotype network illustrating relationships of the revealed COI haplotypes; mutations are shown as 1-step edge; number of mutations separating haplotypes is given in parentheses. (b-c) The Bayesian consensus trees of C. mongola/C. tamerlana inferred from COI sequences (b) and concatenated alignment of three nuclear markers (c); numbers at nodes indicate Bayesian posterior probabilities.
Figure 4.
Phylogenetic patterns of C. mongola/C. tamerlana based on the analysis of mitochondrial (COI) and nuclear (Ca-ATPase, H3, and CAD) markers. (a) Median-joining haplotype network illustrating relationships of the revealed COI haplotypes; mutations are shown as 1-step edge; number of mutations separating haplotypes is given in parentheses. (b-c) The Bayesian consensus trees of C. mongola/C. tamerlana inferred from COI sequences (b) and concatenated alignment of three nuclear markers (c); numbers at nodes indicate Bayesian posterior probabilities.
3.3. Wolbachia Analysis
A total of 88 specimens of C. mongola/C. tamerlana, representing all four COI haplogroups recovered, were screened for the presence of Wolbachia infection. Screening for three Wolbachia genes (16S, wsp, and ftsZ) did not reveal any cases of dissimilar results (i.e., when specimen was positive for one Wolbachia gene, but negative for another gene/genes). In total, 14 specimens out of 88 tested were scored positive for Wolbachia infection (prevalence: 16%). Our analysis revealed strong sex-dependent congruence between a specific COI haplogroup and Wolbachia infection: all 14 infected specimens were females of COI haplogroup HP_II comprising a single haplotype hp2. Three analysed males of this haplogroup, as well as all other investigated specimens, were not infected (Figure 5). Our survey did not recover specific geographical pattern of Wolbachia incidence: infected specimens were randomly found in the geographically remote populations (Figure 3d).
3.3.1. Wolbachia Allele Identified in C. mongola/C. tamerlana
Wolbachia 16S, wsp, and ftsZ genes were sequenced for all infected specimens (14 females) of C. mongola/C. tamerlana. These specimens were infected by a single allele belonged to the supergroup B (grouping according to [74], which was designated as wTam (allele name follow the widely accepted abbreviation style [75,76]).
Comparison with Wolbachia alleles found in other Colias taxa (personal unpublished data) revealed that wTam allele isolated from C. mongola/C. tamerlana was most similar to one of the alleles found in C. palaeno (Linnaeus, 1761) having with the latter identical 16S and ftsZ Wolbachia sequences and differing in one nucleotide substitution in the wsp gene fragment. Infected C. mongola/C. tamerlana specimens share 16S Wolbachia sequence with several insect taxa, belonging to orders Lepidoptera (Pieridae, Nymphalidae, Tortricidae), Orthoptera (Acrididae), Hemiptera (Issidae, Lygaeidae) and wsp sequence with Phengaris nausithous (Bergsträsser, 1779) (Lepidoptera, Lycaenidae). The closely related Wolbachia alleles were found in Hemiptera families Aleyrodidae, Triozidae, Delphacidae, Aphididae, Liviidae for the gene 16S; in insect orders Lepidoptera, Orthoptera, Hemiptera, Diptera, Hymenoptera and spider mites (Trombidiformes) for the gene wsp; in Lepidoptera, Orthoptera, Hemiptera, Diptera, Hymenoptera, Coleoptera and Trombidiformes for the gene ftsZ (https://blast.ncbi.nlm.nih.gov, accessed on 20 July 2023).
3.3.2. Phylogenetic Inferences
Bayesian analysis of the Wolbachia 16S gene fragment based on 43 most similar sequences with known host species mined from GenBank recovered two clades with high (clade I, BS=0.96) and low (clade II, BS=0.78) support (Figure 6). All Wolbachia 16S sequences isolated from infected C. mongola/C. tamerlana specimens grouped within clade II together with Wolbachia alleles found in three insect orders: Lepidoptera, Hemiptera and Orthoptera. The clade I included Wolbachia alleles found in various Lepidoptera families, Diptera, Coleoptera, Hemiptera and mites (Parasitiformes, Trombidiformes). Maximum p-distances within clade I and clade II were as high as 1.5% and 0.8%, respectively; maximum p-distances between two clades – 1.8%.
Bayesian analysis of the Wolbachia wsp gene fragment based on 74 most similar sequences with known host species mined from GenBank revealed two clades: well-supported clade I (BS=1) and weakly supported clade II (BS=0.69). The latter, in turn, subdivided into several unsupported lineages (Figure 7). Obtained in the present study Wolbachia wsp sequences from C. mongola/C. tamerlana clustered within the subclade A of the wsp clade II, which unites Wolbachia wsp alleles isolated from members of insect orders Lepidoptera, Orthoptera, Hemiptera, Diptera, Hymenoptera and spider mites (Trombidiformes). Maximum p-distances within wsp clade I and wsp clade II were as high as 1.8% and 5.7%, respectively; between two clades – 7.1%; within subclade A – 3%.
For the phylogenetic analysis of the Wolbachia ftsZ gene fragment, we used additional 71 most similar sequences with known host species obtained from GenBank. Bayesian analysis revealed several highly supported clades (Bayesian posterior probability >0.95), however deeper nodes mostly remained poorly resolved (Figure 8). Wolbachia samples isolated from C. tamerlana specimens grouped within well-supported clade (BS=0.98) together with members of insect orders Lepidoptera, Orthoptera, Hemiptera, Diptera, Hymenoptera, Coleoptera and Trombidiformes mites. Maximum p-distances within this clade were as high as 1.77%; maximum p-distances among all analysed ftsZ samples reached 2.37%.
4. Discussion
4.1. Molecular Analysis and Taxonomy of C. mongola/C. tamerlana
Mitochondrial DNA is widely used in phylogenetic reconstructions, taxonomic studies and species identification and delimitation due to distinct advantages over other molecular markers: the mitochondrial genome is maternally inherited and generally does not undergo recombination; mtDNA usually evolves faster compared to nuclear DNA, potentially providing better resolution to recover low-level taxonomic relationships; mtDNA presented in cells in a high copy number, thereby allowing better and ease amplification of targeted fragment. However, numerous studies underline limited application of DNA barcoding in the context of incomplete lineage sorting, mitochondrial introgression and infection by endosymbionts, such as Wolbachia [77,78,79,80,81,82,83]. Our analysis of the mitochondrial barcodes alone revealed four differentiated clades within C. mongola/C. tamerlana with genetic distances reaching 2.89%. Such values are comparable to “standard” species-level COI divergence empirically estimated for Lepidoptera [84,85,86,87] and suggest relatively old separation of the recovered COI lineages. In fact, if we relied solely on COI barcodes, the possible conclusion would have been that observed lineages might represent unique cryptic species, especially taking into account comparatively young radiation of the genus Colias [43,44]. However, this conclusion was not supported by subsequent analyses of the nuclear genes and phenotypic traits. Recovered mtDNA phylogenetic structure was not corroborated by nuclear data: the specimens bearing different COI haplotypes were randomly distributed across the nucDNA phylogenetic tree. The nuclear sequences showed no signs of significant divergence between the clades, delimited by mitochondrial barcodes, forming a single, nearly unstructured entity. Furthermore, butterflies collected from the remote geographical localities show no traces of morphological differentiation despite its phenotypic variability. The only exception is C. tamerlana sidonia, the geographically isolated taxon from Lake Khovsgol area (Mongolia), which differs from typical C. mongola/C. tamerlana in larger size, less developed dark suffusion on the fore- and hindwings and large submarginal light spots. Such prominent morphological differences even force some authors to consider sidonia as a subspecies of much easterly distributed taxon, C. nastes. However, our data unequivocally show that sidonia undoubtedly belongs to C. mongola/C. tamerlana complex sharing common and geographically widespread COI haplotype hb1 with the latter. It should also be noted that in the case of cryptic species we should expect a geographic isolation (in case of allopatry), or niche separation (in case of sympatry) of revealed mtDNA clusters, which we consider as putative cryptic species [88,89,90]. Certainly, this is not the case for C. mongola/C. tamerlana, where butterflies of different COI haplogroups were found flying together syntopically (in the same habitat) and synchronously (at the same time), in other words in complete sympatry without any niche separation.
Taxonomic status of C. mongola and C. tamerlana and relationships of this pair of taxa and allied species are also a subject of longstanding debates. Analysis of the century-old type specimens, namely the lectotype of C. tamerlana, the paralectotype of C. tamerlana and the lectoptype of C. mongola, along with C. tamerlana sidonia specimens from the type locality, presumably belonging to the type series, allowed us to shed light on this very controversial issue of the Colias taxonomy. The type locality of C. mongola has been contradictory interpreted in the taxonomic literature and cannot be clearly ascertained. Thus, the limited number of specimens originated from the type series stored in the museum collections are the only reliable source of molecular data. Here, using NGS approach, we recover and analyse DNA barcodes of these old museum specimens. We confirm the conspecificity of C. tamerlana and C. mongola and show that the type specimens of these taxa share the same COI haplotype hb1, common and widely spread over a large geographical area from Northwestern China through South Siberian part of Russia to Western Mongolia.
4.2. Wolbachia Infection in C. mongola/C. tamerlana
Recent investigations have suggested that Wolbachia infection is common and widespread in Lepidoptera [91,92,93]; however, large-scale and comprehensive studies devoted to the incidence, patterns of Wolbachia infection and its impact on phylogenetic inferences of host species are still scarce [94,95,96,97,98,99,100,101,102,103]. To date, Wolbachia infection have been reported only for a few Colias species, namely C. palaeno, C. hyale (Linnaeus, 1758), C. erate poliographus Motschulsky, 1861 and C. croceus [46,93,104,105]. Here, we designated a new Wolbachia allele wTam, which had not been previously recorded in Colias. Surprisingly, none of the available Wolbachia wsp and ftsZ STs (sequence types) found in Colias and deposited in public databases GenBank (https://blast.ncbi.nlm.nih.gov, accessed on 20 July 2023) and PubMLST-Wolbachia (https://pubmlst.org/organisms/wolbachia-spp, accessed on 25 July 2023) fall within the sequences with the highest percentage identity much to wTam. However, our personal unpublished data suggests that alleles similar to wTam can be found in other Colias taxa.
The Wolbachia allele found in C. mongola/C. tamerlana is shared among different insect species, families and even orders. Wolbachia infection is mainly vertically transmitted to the progeny via maternal cytoplasm. Accordingly, one should expect the phylogeny of Wolbachia to be consistent with the phylogeny of their hosts. Notwithstanding, the horizontal transfer of Wolbachia between insect hosts have been suggested for many insect taxa [106,107,108,109,110,111], however the mechanisms of this phenomenon yet remain to be characterized. It has been suggested that Wolbachia can shift between distantly related hosts through host-parasitoid interactions [112,113,114,115], shared host plants [116,117,118,119,120], hybridisation events [98,121,122,123,124] and predator-prey associations [125].
Wolbachia 16S sequence obtained in our study is shared by two Lepidoptera species, nymphalid Neonympha mitchellii French, 1889 distributed in the eastern USA and tortricid moth Eucosma cana (Haworth, 1811), the meadow grasshopper Chorthippus parallelus (Zetterstedt, 1821) (Orthopthera, Acrididae), the false chinch bug Nysius expressus Distant, 1883 (Hemiptera, Lygaeidae) and the planthopper Agalmatium flavescens (Olivier, 1791) (Hemiptera, Issidae). Wolbachia wsp sequence is shared by lycaenid butterfly Phengaris nausithous (Lepidoptera, Lycaenidae). Our data evidenced that the same Wolbachia strain may occur in very distant, not closely related taxa. Thus, we confirm previous studies suggesting that horizontal transmissions events are quite common in nature [126,127]. Interestingly, the very similar wsp and ftsZ STs to C. mongola/C. tamerlana carry spider mites (Acari, Trombidiformes). Mites are known as common parasites for Lepidoptera and other insects and arthropods and have been suggested as potential vectors for Wolbachia transmission [128,129]. Occurrence of closely related Wolbachia strains in butterflies and Trombidiformes indicate that Wolbachia host switches in Lepidoptera might be caused by mites.
Screening for Wolbachia revealed infection in 14 specimens (out of 88 tested), suggesting relatively low infection rate in C. mongola/C. tamerlana (prevalence: 16%). One of the most intriguing results obtained in the present study is that we found a full congruence between sex, specific mitochondrial lineage and Wolbachia infection: all 14 infected specimens were females of COI haplogroup HP_II comprising a single haplotype hp2. To our knowledge, such sex-biased, selective and total infection of certain mitochondrial lineage have never been observed in Lepidoptera. Moreover, our personal unpublished data suggest that such patterns of selective infection can be found in other Colias taxa, presumably being a general characteristic of the genus. It also should be noted that surprisingly low number of publications devoted to analysis of Wolbachia infection in Lepidoptera consider Wolbachia prevalence in males and females independently [130,131], while sex-dependent impact of Wolbachia on its hosts is highly expected.
Interestingly, somewhat similar pattern has been observed in another pierid genus Eurema (Lepidoptera, Pieridae) belonging to the same subfamily Coliadinae, where sex-biased female lineages of Wolbachia were discovered in two Japanese species [132,133,134]. The Wolbachia allele wFem was found at low frequencies only in the Eurema females and has not been observed in the males. A causative role of wFem allele in feminization, a well-known manipulation effect deployed by Wolbachia [135], has been proposed. It has been shown that antibiotic treatment of infected larvae leads to occurrence of intersex individuals, while treatment of adult females results in all-male progeny. We cannot exclude that sex-biased Wolbachia infection observed in C. mongola/C. tamerlana is also a consequence of feminization, a phenomenon that has been rarely encountered within Lepidoptera [136]. However, further comprehensive studies based on the lab experiments and analysis of additional material are needed to confirm such assumptions.
5. Conclusions
Our study is the first large-scale investigation aimed at a detailed analysis of phylogeographical structure, geographical distribution and taxonomy of two enigmatic Colias taxa with the controversial taxonomic status, C. mongola and C. tamerlana. Our analysis cluster DNA barcodes obtained in the present study in four distinct haplogroups, however no association between nuclear genes and mitochondrial clusters, as well as between the distribution of mitochondrial haplotypes and geography have been revealed. Using NGS approach, we recover and analyse DNA barcodes of century-old C. mongola and C. tamerlana type specimens. We show that the type specimens of these taxa share the same COI haplotype. These results confirm the conspecificity of C. tamerlana and C. mongola, solving a longstanding question about their taxonomic status, also demonstrating that application of modern techniques is of a great importance in cases when the type locality is unknown or cannot be clearly ascertained, and old material originated from the type series is the only reliable source of molecular data. We analyse the presence and prevalence of Wolbachia in C. mongola/C. tamerlana and found strong correlation between sex, specific mitochondrial lineage and Wolbachia infection. Phylogenetic analysis placed Wolbachia strain of C. mongola/C. tamerlana together with members of different insect families and even orders, indicating multiple events of host shifts, thus being consistent with the former studies, evidencing that horizontal transmission is a common mechanism of Wolbachia expansion. We conclude that occurrence of deep intraspecific divergences of DNA barcodes do not necessarily is a consequence of cryptic speciation but instead can be a result of Wolbachia infection and some other, most likely environmental factors.
6. Taxonomic conclusion
The nomenclature of C. tamerlana and C. mongola is a subject of longstanding debates [1,3,13]. Here we follow the logic of Greishuber [1], who pointed out that under the International Code of Zoological Nomenclature (Article 8.1.2.) a work is regarded as having been published only when it becomes available. The earliest confirmed date for Romanoff‘s book is 18.12.1897, when the copy of 9th volume have been received by the Harvard University library [1]. Consequently, the name tamerlana has priority over the mongola. Taking into account the molecular data obtained in the present study we confirm conspecificity of the taxa in question and consider Colias nastes mongola Alphéraky, 1897 a subjective junior synonym of Colias tamerlana Staudinger, 1897, syn. nov. We did not find any molecular evidences supporting subspecies status of Colias mongola ukokana Korb & Yakovlev, 2000 and Colias cocandica sidonia Weiss, 1968, despite pronounced differences in the external morphological characters of the latter. These populations do not show any signs of divergence and should be considered as the nominotypical Colias tamerlana.
Thus, we suggest following rearrangements here:
- Colias nastes mongola Alphéraky, 1897 = Colias tamerlana Staudinger, 1897, syn. nov.
- Colias mongola ukokana Korb & Yakovlev, 2000 = Colias tamerlana Staudinger, 1897, syn. nov.
- Colias cocandica sidonia Weiss, 1968 = Colias tamerlana Staudinger, 1897, syn. nov.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: List of studied materials with GenBank accession numbers for sequence records. Table S2: A detailed summary of the nucleotide variability of the studied Ca-ATPase, CAD, and H3 nuclear genes fragments among the sequenced samples of C.mongola/C. tamerlana.
Author Contributions
Project design, N.A.S., G.N.S.; conceptualization, N.A.S., A.K., G.N.S.; methodology, N.A.S., N.V.G.; PCR amplification and sequencing, N.A.S., G.N.S., A.V.K., A.E.R., N.V.G.; molecular analysis, N.A.S., G.N.S., N.V.G.; writing—original draft preparation, N.A.S., G.N.S.; writing—review and editing, A.K., A.V.K., N.V.G.; figure preparation, N.A.S., G.N.S.; Collecting material, R.V.Y., A.K, A.V.K. All authors have read and agreed to the published version of the manuscript.
Funding
The financial support for this study was provided to N.A.S., G.N.S., A.V.K. and R.V.Y. by the Russian Science Foundation grant No. 22-24-01086 (collecting the material and molecular analysis) and to N.A.S. and G.N.S. by the state research project No. 122031100272-3 (morphological analysis).
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors are grateful to Boris Khramov (Saint-Petersburg, Russia), Alexander Kurmaev, Anatoly Marusov and Sergey Churkin (Moscow, Russia) for providing material for DNA analysis. We thank Alexander Lvovsky and Sergey Sinev (St. Petersburg, Russia) and Andrey Sviridov (Moscow, Russia) for their kind assistance during the work in the collections of the Zoological Institute, Russian Academy of Sciences and the Zoological Museum of the Moscow State University.
Conflicts of Interest
All authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Greishuber, J. Guide to the butterflies of the Palearctic region. Pieridae part II; Omnes Artes: Milano, Italy, 2014. [Google Scholar]
- Greishuber, J.; Lamas, G. A synonymic list of the genus Colias Fabricius, 1807 (Lepidoptera: Pieridae). Mitt. Münch. Ent. Ges. 2007, 97, 131–171. [Google Scholar]
- Greishuber, J.; Worthy, B.; Lamas, G. The genus Colias Fabricius, 1807. Jan Haugum’s annotated catalogue of the Old World Colias (Lepidoptera, Pieridae); Tshikolovets Publications: Pardubice, Czech Republic, 2012. [Google Scholar]
- Verhulst, J. Les Colias du Globe. Monograph of the genus Colias; Goecke & Evers: Keltern, Germany, 2000. [Google Scholar]
- Boyer, T. Barcode et Colias africains (Lepidoptera, Pieridae). Entomologia Africana 2018, 23, 19–28. [Google Scholar]
- Kir’yanov, A.V. DNA Barcoding and renewed taxonomy of South American Clouded Sulphurs (Lepidoptera: Pieridae: Colias). Southwest. Entomol. 2021, 46, 891–927. [Google Scholar] [CrossRef]
- Kir’yanov, A.V. A new species of Colias Fabricius, 1807 from eastern Sudan (Lepidoptera, Pieridae, Coliadinae). Entomologia Africana 2021, 26, 19–28. [Google Scholar]
- Tshikolovets, V.V.; Yakovlev, R.V.; Bálint, Z. The butterflies of Mongolia; Tshikolovets Publications: Pardubice, Czech Republic, 2009. [Google Scholar]
- Tshikolovets, V.V.; Yakovlev, R.V.; Kosterin, O.E. The butterflies of Altai, Sayan and Tuva (South Sibiria); Tshikolovets Publications: Pardubice, Czech Republic, 2009. [Google Scholar]
- Staudinger, O. Drei neue paläarktische Lepidopteren. Deut. ent. Zeit. Iris 1897, 10, 152–156. [Google Scholar]
- Alphéraky, S.N. Mémoire sur différents lépidoptères, tant nouveaux que peu connus, de la faune paléarctique. In Mémoires sur les Lépidoptères; Romanoff, N.M., Ed.; Imprimerie de P.P. Soïkine: Saint-Petersburg, Russia, 1897; Vol. 9, pp. 185–227. [Google Scholar]
- Korb, S.K. On the systematic of some Colias cocandica-like taxa (Lepidoptera: Pieridae). Phegea 2006, 34, 147–154. [Google Scholar]
- Gorbunov, O.G. On the nomenclature of Colias nastes mongola Alpheraky, 1897 and Colias tamerlana Staudinger, 1897. Entomol. Rev. 2012, 92, 559–564. [Google Scholar] [CrossRef]
- Weiss, D. Beitrag zur Kenntnis der Tagfalter-Fauna der Mongolei (Lepidoptera, Rhopalocera). Acta Entomol. Mus. Natl. Pragae 1968, 13, 109–118. [Google Scholar]
- Staudinger, O. Macrolepidoptera. In Catalog der Lepidopteren des palaearctischen Faunengebeites; Staudinger, O., Rebel, H., Eds.; Friedländer & Sohn: Berlin, Germany, 1901. [Google Scholar]
- Talbot, G. Pieridae. In Lepidopterorum Catalogus; Strand, E., Ed.; W. Junk: Berlin, Germany, 1935; Vol. 66, pp. 386–697. [Google Scholar]
- Tshikolovets, V.V.; Bidzilya, O.V.; Golovushkin, M.I. The butterflies of Transbaikal Siberia; Tshikolovets Publications: Brno–Kiev, Czech Republic–Ukraine, 2002. [Google Scholar]
- Tuzov, V.K.; Bogdanov, P.V.; Devyatkin, A.L.; Kaabak, L.V.; Korolev, V.A.; Murzin, V.S.; Samodurov, G.D.; Tarasov, E.A. Guide to the butterflies of Russia and adjacent territories (Lepidoptera, Rhopalocera). Volume I. Hesperiidae, Papilionidae, Pieridae, Satyridae; Pensoft: Sofia–Moscow, Bulgaria–Russia, 1997. [Google Scholar]
- Gorbunov, P.Y. The Butterflies of Russia: Classification, genitalia, keys for identification (Lepidoptera: Hesperioidea and Papilionoidea); Thesis: Ekaterinburg, Russia, 2001. [Google Scholar]
- Gorbunov, P.Y.; Kosterin, O.E. The Butterflies (Hesperioidea and Papilionoidea) of North Asia (Asian Part of Russia) in Nature; Rodina & Fodio and Gallery Fund: Moscow–Chelyabinsk, Russia, 2003. [Google Scholar]
- Kurentsov, A.I. Bulavousye cheshyekrylye Dal’nego Vostoka SSSR [Butterflies of the Far East USSR]; Nauka: Leningrad, USSR, 1970. [Google Scholar]
- Korb, S.K.; Yakovlev, R.V. Colias mongola ukokana nov. ssp. (Lepidoptera Pieridae). Alexanor 2000, 21, 3–6. [Google Scholar]
- Dincă, V.; Montagud, S.; Talavera, G.; Hernandez-Roldan, J.; Munguira, M.L.; Garcia-Barros, E.; Hebert, P.D.N.; Vila, R. DNA barcode reference library for Iberian butterflies enables a continental-scale preview of potential cryptic diversity. Sci. Rep. 2015, 5, 12395. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Dantchenko, A.V.; Vishnevskaya, M.S.; Saifitdinova, A.F. Detecting cryptic species in sympatry and allopatry: analysis of hidden diversity in Polyommatus (Agrodiaetus) butterflies (Lepidoptera: Lycaenidae). Biol. J. Linn. Soc. 2015, 116, 468–485. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Shapoval, N.A. Chromosomal identification of cryptic species sharing their DNA barcodes: Polyommatus (Agrodiaetus) antidolus and P. (A.) morgani in Iran (Lepidoptera, Lycaenidae). Comp. Cytogenet. 2017, 11, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Hammond, P.C.; McCorkle, D.V. A review of geographic variation and possible evolutionary relationships in the Colias scudderii-gigantea complex of North America (Pieridae). J. Lepid. Soc. 2008, 64, 201–215. [Google Scholar]
- Wahlberg, N.; Braby, M.F.; Brower, A.V.Z.; De Jong, R.; Lee, M.-M.; Nylin, S.; Pierce, N.E.; Sperling, F.A.H.; Vila, R.; Warren, A.D.; Zakharov, E. Synergistic effects of combining morphological and molecular data in resolving the phylogeny of butterflies and skippers. Proc. Royal Soc. B 2005, 272, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, N.; Rota, J.; Braby, M.F.; Pierce, N.E.; Wheat, C.W. Revised systematics and higher classification of pierid butterflies (Lepidoptera: Pieridae) based on molecular data. Zool. Scr. 2014, 43, 641–650. [Google Scholar] [CrossRef]
- Espeland, M.; Hall, J.P.W.; DeVries, P.J.; Lees, D.C.; Cornwall, M.; Hsu, Y.-F.; Wu, L.-W.; Campbell, D.L.; Talavera, G.; Vila, R.; Salzman, S.; Ruer, S.; Lohman, D.J.; Pierce, N.E. Ancient Neotropical origin and recent recolonisation: phylogeny, biogeography and diversification of the Riodinidae (Lepidoptera: Papilionoidea). Mol. Phylogenet. Evol. 2015, 93, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Espeland, M.; Breinholt, J.; Willmot, K.R.; Warren, A.D.; Vila, R.; Toussaint, E.F.A.; Maunsell, S.C.; Aduse-Poku, K.; Talavera, R.; Eastwood, R.; Jarzyna, M.A.; Guralnick, R.; Lohman, D.J.; Pierce, N.E.; Kawahara, A. A comprehensive and dated phylogenomic analysis of butterflies. Curr. Biol. 2018, 28, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Warren, A.D.; Wahlberg, N.; Brower, A.V.Z.; Lukhtanov, V.A.; Kodandaramaiah, U. Ten genes and two topologies: an exploration of higher relationships in skipper butterflies (Hesperiidae). PeerJ 2016, 4, e2653. [Google Scholar] [CrossRef]
- Mitter, C.; Davis, D.R.; Cummings, M.P. Phylogeny and evolution of Lepidoptera. Annu. Rev. Entomol. 2017, 62, 265–283. [Google Scholar] [CrossRef]
- Seraphim, N.; Kaminski, L.A.; Devries, P.J.; Penz, C.; Callaghan, C.; Wahlberg, N.; Silva-Brandao, K.L.; Freitas, A.V.L. Molecular phylogeny and higher systematics of the metalmark butterflies (Lepidoptera: Riodinidae). Syst. Entomol. 2018, 43, 407–425. [Google Scholar] [CrossRef]
- Toussaint, E.F.A.; Breinholt, E.W.; Earl, C.; Warren, A.D.; Brower, A.V.Z.; Yago, M.; Dexter, K.M.; Espeland, M.; Pierce, N.E.; Lohman, D.J.; Kawahara, A.Y. Anchored phylogenomics illuminates the skipper butterfly tree of life. BMC Evol. Biol. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cong, Q.; Shen, J.; Brockmann, E.; Grishin, N. Three new subfamilies of skipper butterflies (Lepidoptera, Hesperiidae). ZooKeys 2019, 861, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Allio, R.; Scornavacca, C.; Nabholz, B.; Clamens, A.-L.; Sperling, F.A.H.; Fabien, L. Whole genome shotgun phylogenomics resolves the pattern and timing of swallowtail butterfly evolution. Syst. Biol. 2020, 69, 38–60. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, M.; Chazot, N.; Wheat, C.W.; Schweiger, O.; Wahlberg, N. A complete time-calibrated multi-gene phylogeny of the European butterflies. ZooKeys 2020, 938, 97–124. [Google Scholar] [CrossRef] [PubMed]
- Vodă, R.; Dapporto, L.; Dincă, V.; Vila, R. Cryptic matters: overlooked species generate most butterfly beta-diversity. Ecography 2015, 38, 405–409. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Shapoval, N.A.; Dantchenko, A.V. Agrodiaetus shahkuhensis sp. n. (Lepidoptera, Lycaenidae), a cryptic species from Iran discovered by using molecular and chromosomal markers. Comp. Cytogenet. 2008, 2, 99–114. [Google Scholar]
- Shapoval, N.A.; Lukhtanov, V.A. On the generic position of Polyommatus avinovi (Lepidoptera: Lycaenidae). Folia Biol. (Krakow) 2016, 64, 267–273. [Google Scholar] [CrossRef]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.E.; Vila, R. Establishing criteria for higher-level classification using molecular data: the systematics of Polyommatus blue butterflies (Lepidoptera, Lycaenidae). Cladistics 2013, 29, 166–192. [Google Scholar] [CrossRef]
- Kramp, K.; Cizek, O.; Madeira, P.M.; Ramos, A.A.; Konvicka, M.; Castilho, R.; Schmitt, T. Genetic implications of phylogeographical patterns in the conservation of the boreal wetland butterfly Colias palaeno (Pieridae). Biol. J. Linn. Soc. 2016, 119, 1068–1081. [Google Scholar] [CrossRef]
- Laiho, J.; Ståhls, G. DNA barcodes identify Central Asian Colias butterflies (Lepidoptera, Pieridae). ZooKeys 2013, 365, 175–196. [Google Scholar] [CrossRef]
- Wheat, C.W.; Watt, W.B. A mitochondrial-DNA-based phylogeny for some evolutionary-genetic model species of Colias butterflies (Lepidoptera, Pieridae). Mol. Phylogenet. Evol. 2008, 47, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Porter, A.H. An AFLP-based interspecific linkage map of sympatric, hybridizing Colias butterflies. Genetics 2004, 168, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Dzurinka, M.; Šemeláková, M.; Panigaj, L. Taxonomy of hybridizing Colias croceus (Geoffroy, 1785) and Colias erate (Esper, 1805) (Lepidoptera, Pieridae) in light of mitochondrial and nuclear DNA, with occurrence and effects of Wolbachia infection. Zool. Anz. 2022, 299, 73–811. [Google Scholar] [CrossRef]
- D’Ercole, J.; Dincă, V.; Opler, P.A.; Kondla, N.; Schmidt, C.; Phillips, J.D.; Robbins, R.; Burns, J.M.; Miller, S.E.; Grishin, N.; Zakharov, E.V.; DeWaard, J.R.; Ratnasingham, S.; Hebert, P.D.N. A DNA barcode library for the butterflies of North America. PeerJ 2021, 9, e11157. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Lukhtanov, V.A.; Shapoval, N.A. Detection of cryptic species in sympatry using population analysis of unlinked genetic markers: A study of the Agrodiaetus kendevani species complex (Lepidoptera: Lycaenidae). Dokl. Biol. Sci. 2008, 423, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Marine Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. PNAS 2006, 103, 968–971. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Sourakov, A.; Zakharov, E.V.; Hebert, P.D.N. DNA barcoding Central Asian butterflies: Increasing geographical dimension does not significantly reduce the success of species identification. Mol. Ecol. Resour. 2009, 9, 1302–1310. [Google Scholar] [CrossRef]
- Wahlberg, N.; Wheat, C.W. Genomic outposts serve the phylogenomic pioneers: Designing novel nuclear markers for genomic DNA extractions of Lepidoptera. Syst. Biol. 2008, 57, 231–242. [Google Scholar] [CrossRef]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Wahlberg, N.; Peña, C.; Ahola, M.; Wheat, C. W.; Rota, J. PCR primers for 30 novel gene regions in the nuclear genomes of Lepi- doptera. ZooKeys 2016, 596, 129–141. [Google Scholar] [CrossRef]
- Li, W.; Cong, O.; Shen, J.; Zhang, J.; Hallwachs, W.; Janzen, D. H.; Grishin, N.V. Genomes of skipper butterflies reveal extensive convergence of wing patterns. PNAS 2019, 116, 6232–6237. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D. H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequency in insects: Evidence of a global equilibrium? Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neil, S. Phylogeny and PCR- based classification of Wolbachia strains using wsp gene sequences. Proc. Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; Thierer, T.; Ashton, B.; Meintjes, P.; Drummond, A. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Villesen, P. FaBox: an online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Meth- ods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1. 7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-X.; Li, W.-H. Statistical test of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Werren, J.H.; Zhang, W.; Guo, L.R. Evolution and phylogeny of Wolbachia: Reproductive parasites of arthropods. Proc. Biol. Sci. 1995, 261, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Rousset, F.; O’Neil, S. Phylogeny and PCR- based classification of Wolbachia strains using wsp gene sequences. Proc. Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; de Stordeur, E. Properties of Drosophila simulans strains experimentally infected by different clones of the bacterium Wolbachia. Heredity 1994, 72, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Funk, D.J.; Omland, K.E. Species-level paraphyly and polyphyly: frequency, causes, and consequences, with insights from animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 397–423. [Google Scholar] [CrossRef]
- Hurst, G.D.D.; Jiggins, F.M. Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: the effects of inherited symbionts. Proc. Royal Soc. B Biol. Sci. 2005, 272, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, M.; Fiedler, K. Does the DNA barcoding gap exist? - a case study in blue butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Zakharov, E.V.; Lobo, N.F.; Nowak, C.; Hellma, J.J. Introgression as a likely cause of mtDNA paraphyly in two allopatric skippers (Lepidoptera: Hesperiidae). Heredity 2009, 102, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Gerth, M.; Geißler, A.; Bledidorn, C. Wolbachia infections in bees (Anthophila) and possible implications for DNA barcoding. Syst. Biodivers. 2011, 9, 319–327. [Google Scholar] [CrossRef]
- Kvie, K.S.; Hogner, S.; Aarvik, L.; Lifjeld, J.T.; Johnsen, A. Deep sympatric mtDNA divergence in the autumnal moth (Epirrita autumnata). Ecol. Evol. 2013, 3, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Ritter, S.; Michalski, S.G.; Settele, J.; Wiemers, M.; Fric, Z.F.; Sielezniew, M.; Šašić, M.; Rozier, Y.; Durka, W. Wolbachia infections mimic cryptic speciation in two parasitic butterfly species, Phengaris teleius and P. nausithous (Lepidoptera: Lycaenidae). PloS ONE 2013, 8, e78107. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. Royal Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, A.; Godfray, H.C.J.; Huemer, P.; Mutanen, M.; Rougerie, R.; van Nieukerken, E.J.; Ratnasingham, S.; Hebert, P.D.N. Genetic patterns in European geometrid moths revealed by the Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e84518. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: the Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; deWaard, J.R. Counting animal species with DNA barcodes: Canadian insects. Philos. Trans. R. Soc. B, Biol. Sci. 2016, 371, 20150333. [Google Scholar] [CrossRef]
- Friberg, M.; Bergman, M.; Kullberg, J.; Wahlberg, N.; Wiklund, C. Niche separation in space and time between two sympatric sister species – a case of ecological pleiotropy. Evol. Ecol. 2008, 22, 1–18. [Google Scholar] [CrossRef]
- Friberg, M.; Olofsson, M.; Berger, D.; Karlsson, B.; Wiklund, C. Habitat choice precedes host plant choice-niche separation in a species pair of a generalist and a specialist butterfly. Oikos 2008, 117, 1337–1344. [Google Scholar] [CrossRef]
- Friberg, M.; Leimar, O.; Wiklund, C. Heterospecific courtship, minority effects and niche separation between cryptic butterfly species. J. Evol. Biol. 2013, 26, 971–979. [Google Scholar] [CrossRef]
- Salunkhe, R.C.; Narkhede, K.P.; Shouche, Y.S. Distribution and evolutionary impact of Wolbachia on butterfly hosts. Indian J Microbiol. 2014, 54, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Araujo-Jnr, E.V.; Welch, J.J.; Kawahara, A.Y. Wolbachia in butterflies and moths: geographic structure in infection frequency. Front. Zool. 2015, 12, 16. [Google Scholar] [CrossRef]
- Ilinsky, Y.; Kosterin, O.E. Molecular diversity of Wolbachia in Lepidoptera: Prevalent allelic content and high recombination of MLST genes. Mol. Phylogenet. Evol. 2017, 109, 164–179. [Google Scholar] [CrossRef]
- Smith, M.A.; Bertrand, C.; Crosby, K.; Eveleigh, E.S.; Fernandez-Triana, J.; Fisher, B.L.; Gibbs, J.; Hajibabaei, M.; Hallwachs, W.; Hind, K.; Hrcek, J.; Huang, D.W.; Janda, M.; Janzen, D.H.; Li, Y.; Miller, S.E.; Packer, L.; Quicke, D.; Ratnasingham, S.; Rodriguez, J.; Rougerie, R.; Shaw, M.R.; Sheffield, C.; Stahlhut, J.K.; Steinke, D.; Whitfield, J.; Wood, M.; Zhou, X. Wolbachia and DNA barcoding insects: patterns, potential, and problems. PloS ONE 2012, 7, e36514. [Google Scholar] [CrossRef]
- Kodandaramaiah, U.; Simonsen, T.J.; Bromilow, S.; Wahlberg, N.; Sperling, F. Deceptive single-locus taxonomy and phylogeography: Wolbachia-associated divergence in mitochondrial DNA is not reflected in morphology and nuclear markers in a butterfly species. Ecol. Evol. 2013, 3, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Lohman, D.J.; Wahlberg, N.; Müller, C.J.; Brattström, O.; Collins, S.C.; Peggie, D.; Aduse-Poku, K.; Kodandaramaiah, U. Evolution of Hypolimnas butterflies (Nymphalidae): Out-of-Africa origin and Wolbachia-mediated introgression. Mol. Phylogenet. Evol. 2018, 123, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Gaunet, A.; Dincă, V.; Dapporto, L.; Montagud, S.; Vodă, R.; Schär, S.; Badiane, A.; Font, E.; Vila, R. Two consecutive Wolbachia-mediated mitochondrial introgressions obscure taxonomy in Palearctic swallowtail butterflies (Lepidoptera, Papilionidae). Zool. Scr. 2019, 48, 507–519. [Google Scholar] [CrossRef]
- Duplouy, A.; Pranter, R.; Warren-Gash, H.; Tropek, R.; Wahlberg, N. Towards unravelling Wolbachia global exchange: a contribution from the Bicyclus and Mylothris butterflies in the Afrotropics. BMC Microbiol. 2020, 20, 319. [Google Scholar] [CrossRef] [PubMed]
- Arif, S.; Gerth, M.; Hone-Millard, W.G.; Nunes, M.D.S.; Dapporto, L.; Shreeve, T.G. Evidence for multiple colonisations and Wolbachia infections shaping the genetic structure of the widespread butterfly Polyommatus icarus in the British Isles. Mol Ecol. 2021, 30, 5196–5213. [Google Scholar] [CrossRef] [PubMed]
- Lucek, K.; Bouaouina, S.; Jospin, A.; Grill, A.; de Vos, J.M. Prevalence and relationship of endosymbiotic Wolbachia in the butterfly genus Erebia. BMC Ecol. Evol. 2021, 21, 95. [Google Scholar] [CrossRef]
- Sucháčková Bartoňová, A.; Konvička, M.; Marešová, J.; Wiemers, M.; Ignatev, N.; Wahlberg, N.; Schmitt, T.; Faltýnek Fric, Z. Wolbachia affects mitochondrial population structure in two systems of closely related Palaearctic blue butterflies. Sci Rep. 2021, 11, 3019. [Google Scholar] [CrossRef]
- Wendt, M.; Kulanek, D.; Varga, Z.; Rákosy, L.; Schmitt, T. Pronounced mito-nuclear discordance and various Wolbachia infections in the water ringlet Erebia pronoe have resulted in a complex phylogeographic structure. Sci Rep. 2022, 12, 5175. [Google Scholar] [CrossRef]
- Narita, S.; Shimajiri, Y.; Nomura, M. Strong cytoplasmic incompatibility and high vertical transmission rate can explain the high frequencies of Wolbachia infection in Japanese populations of Colias erate poliographus (Lepidoptera: Pieridae). Bull. Entomol. Res. 2009, 99, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.-H.; Gao, S. A prevalence survey of Wolbachia in butterflies from South China. Entomol. Exp. Appl. 2021, 169, 1157–1166. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Giordano, R.; Colbert, A.M.E.; Karr, T.L. 16S rRNA phylogenetic analysis of the bacterial endosymbionts associated with cytoplasmic incompatibility in insects. Proc. Natl. Acad. Sci. 1992, 89, 2699–2702. [Google Scholar] [CrossRef]
- Rousset, F.; Bouchon, D.; Pintureau, B.; Solignac, M. Wolbachia endosymbionts responsible for various alterations of sexuality in arthropods. Proc. Biol. Sci. 1992, 250, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, R.O.; Prezotto, L.F.; Perondini, A.L.P.; Marino, C.L.; Selivon, D. Wolbachia in guilds of Anastrepha fruit flies (Teph- ritidae) and parasitoid wasps (Braconidae). Genet. Mol. Biol. 2016, 39, 600–610. [Google Scholar] [CrossRef]
- Chu, C.; Hoffmann, M.; Braswell, W.E.; Pelz-Stelinski, K.S. Genetic variation and potential coinfection of Wolbachia among widespread Asian citrus psyllid (Diaphorina citri Kuwayama) populations. Insect Sci. 2019, 26, 671–682. [Google Scholar] [CrossRef]
- Sintupachee, S.; Milne, J.R.; Poonchaisri, S.; Baimai, V.; Kittayapong, P. Closely related Wolbachia strains within the pumpkin arthropod community and the potential for horizontal transmission via the plant. Microb. Ecol. 2006, 51, 294–301. [Google Scholar] [CrossRef]
- Gutzwiller, F.; Dedeine, F.; Kaiser, W.; Giron, D.; Lopez-Vaamonde, C. Correlation between the green-island phenotype and Wolbachia infections during the evolutionary diversification of Gracillariidae leaf-mining moths. Ecol. Evol. 2015, 5, 4049–4062. [Google Scholar] [CrossRef]
- Vavre, F.; Fleury, F.; Lepetit, D.; Fouillet, P.; Bouletreau, M. Phylogenetic evidence for horizontal transmission of Wolbachia in host-parasitoid associations. Mol. Biol. Evol. 1999, 16, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Haine, E.R.; Pickup, N.J.; Cook, J.M. Horizontal transmission of Wolbachia in a Drosophila community. Ecol. Entomol. 2005, 30, 464–472. [Google Scholar] [CrossRef]
- Johannesen, J. Tracing the history and ecological context of Wolbachia double infection in a specialist host (Urophora cardui) – parasitoid (Eurytoma serratulae) system. Ecol. Evol. 2017, 7, 986–996. [Google Scholar] [CrossRef]
- Hou, H.-Q.; Zhao, G.-Z.; Su, C.-Y.; Zhu, D.-H. Wolbachia prevalence patterns: horizontal transmission, recombination, and multiple infections in chestnut gall wasp-parasitoid communities. Entomol. Exp. Appl. 2020, 168, 752–765. [Google Scholar] [CrossRef]
- Mitsuhashi, W.; Saiki, T.; Wei, W.; Kawakita, H.; Sato, M. Two novel strains of Wolbachia coexisting in both species of mulberry leafhoppers. Insect Mol. Biol. 2002, 11, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Kittayapong, P.; Jamnongluk, W.; Thipaksorn, A.; Milne, J.R.; Sindhusake, C. Wolbachia infection complexity among insects in the tropical rice-field community. Mol. Ecol. 2003, 12, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Sintupachee, S.; Milne, J.R.; Poonchaisri, S.; Baimai, V.; Kittayapong, P. Closely related Wolbachia strains within the pumpkin arthropod community and the potential for horizontal transmission via the plant. Microb. Ecol. 2006, 51, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-H.; Zhu, D.-H.; Liu, Z.; Zhao, L.; Su, C.-Y. High levels of multiple infections, recombination and horizontal transmission of Wolbachia in the Andricus mukaigawae (Hymenoptera; Cynipidae) communities. PLoS ONE 2013, 8, e78970. [Google Scholar] [CrossRef]
- Li, S.-J.; Ahmed, M.Z.; Lv, N.; Shi, P.-Q.; Wang, X.-M.; Huang, J.-L.; Qui, B.-L. Plantmediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2017, 11, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; Solignac, M. The evolution of single and double Wolbachia symbioses during speciation in the Drosophila simulans complex. Proc. Natl. Acad. Sci. 1995, 92, 6389–6393. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M. Male-killing Wolbachia and mitochondrial DNA: selective sweeps, hybrid introgression and parasite population dynamics. Genetics 2003, 164, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.S.; Vanderpool, D.; Conner, W.R.; Matute, D.R.; Turelli, M. Wolbachia acquisition by Drosophila yakuba-clade hosts and transfer of incompatibility loci between distantly related Wolbachia. Genetics 2019, 212, 1399–1419. [Google Scholar] [CrossRef]
- Miraldo, A.; Duplouy, A. High Wolbachia strain diversity in a clade of dung beetles endemic to Madagascar. Front. Ecol. Evol. 2019, 7, 157. [Google Scholar] [CrossRef]
- Le Clec’h, W.; Chevalier, F.D.; Genty, L.; Bertaux, J.; Bouchon, D.; Sicard, M. Cannibalism and predation as paths for horizontal passage of Wolbachia between terrestrial isopods. PLoS ONE 2013, 8, e60232. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.; Albanese, D.; Tuohy, K.; Donati, C.; Segata, N.; Rota-Stabelli, O. Large scale genome reconstructions illuminate Wolbachia evolution. Nat. Commun. 2020, 11, 5235. [Google Scholar] [CrossRef] [PubMed]
- Bing, X.-L.; Zhao, D.-S.; Sun, J.-T.; Zhang, K.-J.; Hong, X.-Y. Genomic analysis of Wolbachia from Laodelphax striatellus (Delphacidae, Hemiptera) reveals insights into its “Jekyll and Hyde” mode of infection pattern. Genome Biol. Evol. 2020, 12, 3818–3831. [Google Scholar] [CrossRef]
- Conradt, L.; Corbet, S.A.; Roper, T.J.; Bodsworth, E.J. Parasitism by the mite Trombidium breei on four UK butterfly species. Ecol. Entomol. 2002, 27, 651–659. [Google Scholar] [CrossRef]
- Sanaei, E.; Charlat, S.; Engelstädter, J. Wolbachia host shifts: routes, mechanisms, constraints and evolutionary consequences. Biol. Rev. 2021, 96, 433–453. [Google Scholar] [CrossRef]
- Solovyev, V.I.; Ilinsky, Y.; Kosterin, O.E. Genetic integrity of four species of Leptidea (Pieridae, Lepidoptera) as sampled in sympatry in West Siberia. Comp. Cytogenet. 2015, 9, 299–324. [Google Scholar] [CrossRef]
- Makhov, I.A.; Gorodilova, Y. Yu.; Lukhtanov, V.A. Sympatric occurrence of deeply diverged mitochondrial DNA lineages in Siberian geometrid moths (Lepidoptera: Geometridae): cryptic speciation, mitochondrial introgression, secondary admixture or effect of Wolbachia? Biol. J. Linn. Soc. 2021, 134, 342–365. [Google Scholar] [CrossRef]
- Kato, Y. Overlapping distribution of two groups of the butterfly Eurema hecabe differing in the expression of seasonal morphs on Okinawa- jima Island. Zool. Sci. 2000, 17, 539–547. [Google Scholar] [CrossRef]
- Narita, S.; Kageyama, D.; Nomura, M.; Fukatsu, T. Unexpected mechanism of symbiont-induced reversal of insect sex: feminizing Wolbachia continuously acts on the butterfly Eurema hecabe during larval development. Appl. Environ. Microbiol. 2007, 73, 4332–4341. [Google Scholar] [CrossRef]
- Kern, P.; Cook, J.M.; Kageyama, D.; Riegler, M. Double trouble: Combined action of meiotic drive and Wolbachia feminization in Eurema butterflies. Biol. Lett. 2015, 11, 20150095. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Duplouy, A.; Hornett, E.A. Uncovering the hidden players in Lepidoptera biology: The heritable microbial endosymbionts. PeerJ 2018, 6, e4629. [Google Scholar] [CrossRef]
Figure 1.
Type specimens of the Colias species deposited in the Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia (ZISP), barcoded in the present study: (a) paralectotype (female) of C. tamerlana Staudinger, 1897; (b) lectotype (male) of C. mongola Alphéraky, 1897.
Figure 1.
Type specimens of the Colias species deposited in the Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia (ZISP), barcoded in the present study: (a) paralectotype (female) of C. tamerlana Staudinger, 1897; (b) lectotype (male) of C. mongola Alphéraky, 1897.

Figure 2.
Relationships of C. mongola, C. tamerlana and the related taxa suggested by different authors (see text for explanation).
Figure 2.
Relationships of C. mongola, C. tamerlana and the related taxa suggested by different authors (see text for explanation).
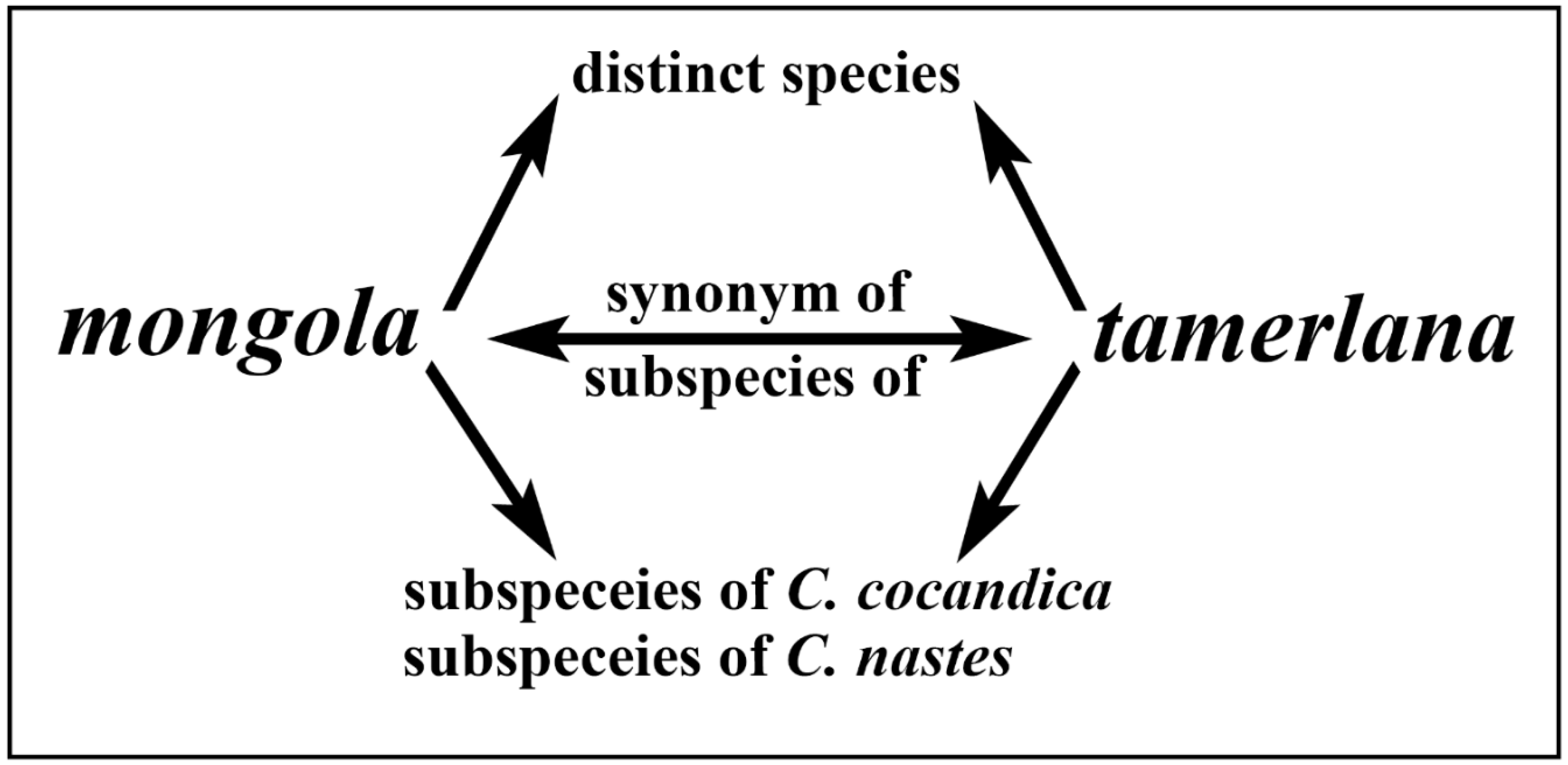
Figure 5.
Infection rates of Wolbachia in 88 specimens of C. mongola/C. tamerlana analysed in the present study. Values and symbols in each bar indicate the number and sex of individuals bearing COI haplotypes of certain haplogroup. Green and red bars indicate uninfected and infected specimens, respectively.
Figure 5.
Infection rates of Wolbachia in 88 specimens of C. mongola/C. tamerlana analysed in the present study. Values and symbols in each bar indicate the number and sex of individuals bearing COI haplotypes of certain haplogroup. Green and red bars indicate uninfected and infected specimens, respectively.
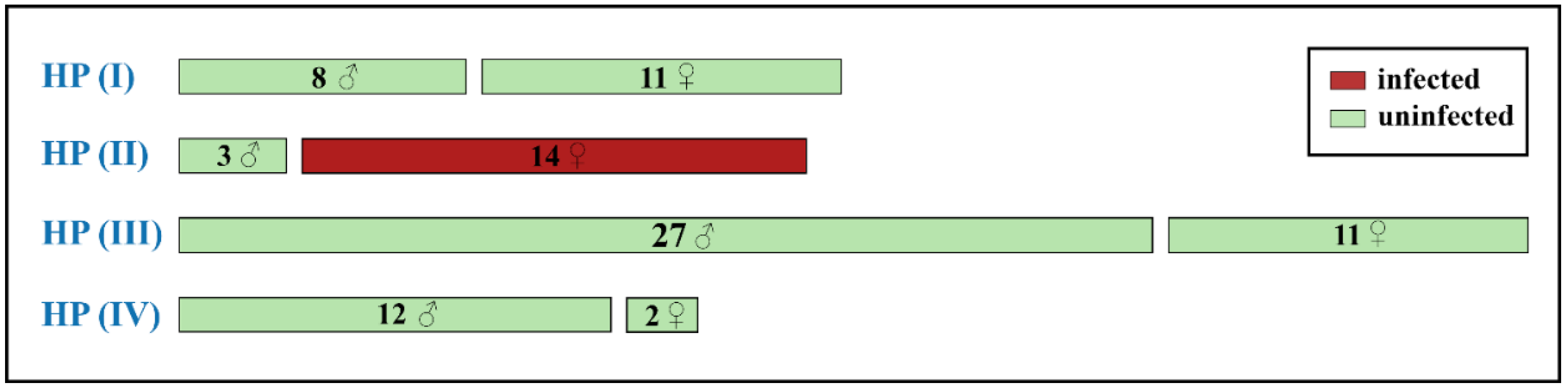
Figure 6.
The Bayesian tree of 16S gene fragment inferred from 57 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
Figure 6.
The Bayesian tree of 16S gene fragment inferred from 57 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
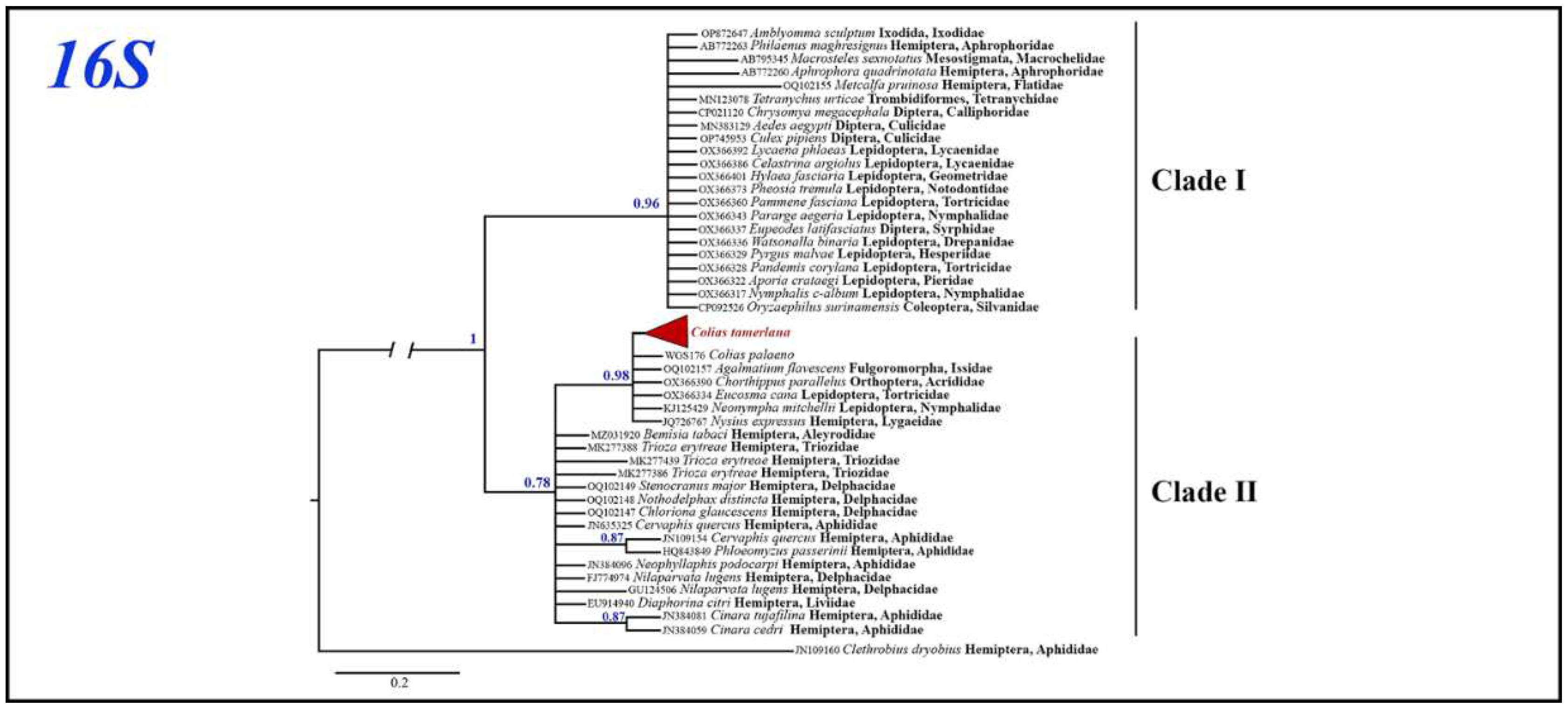
Figure 7.
The Bayesian tree of wsp gene fragment inferred from 88 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
Figure 7.
The Bayesian tree of wsp gene fragment inferred from 88 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
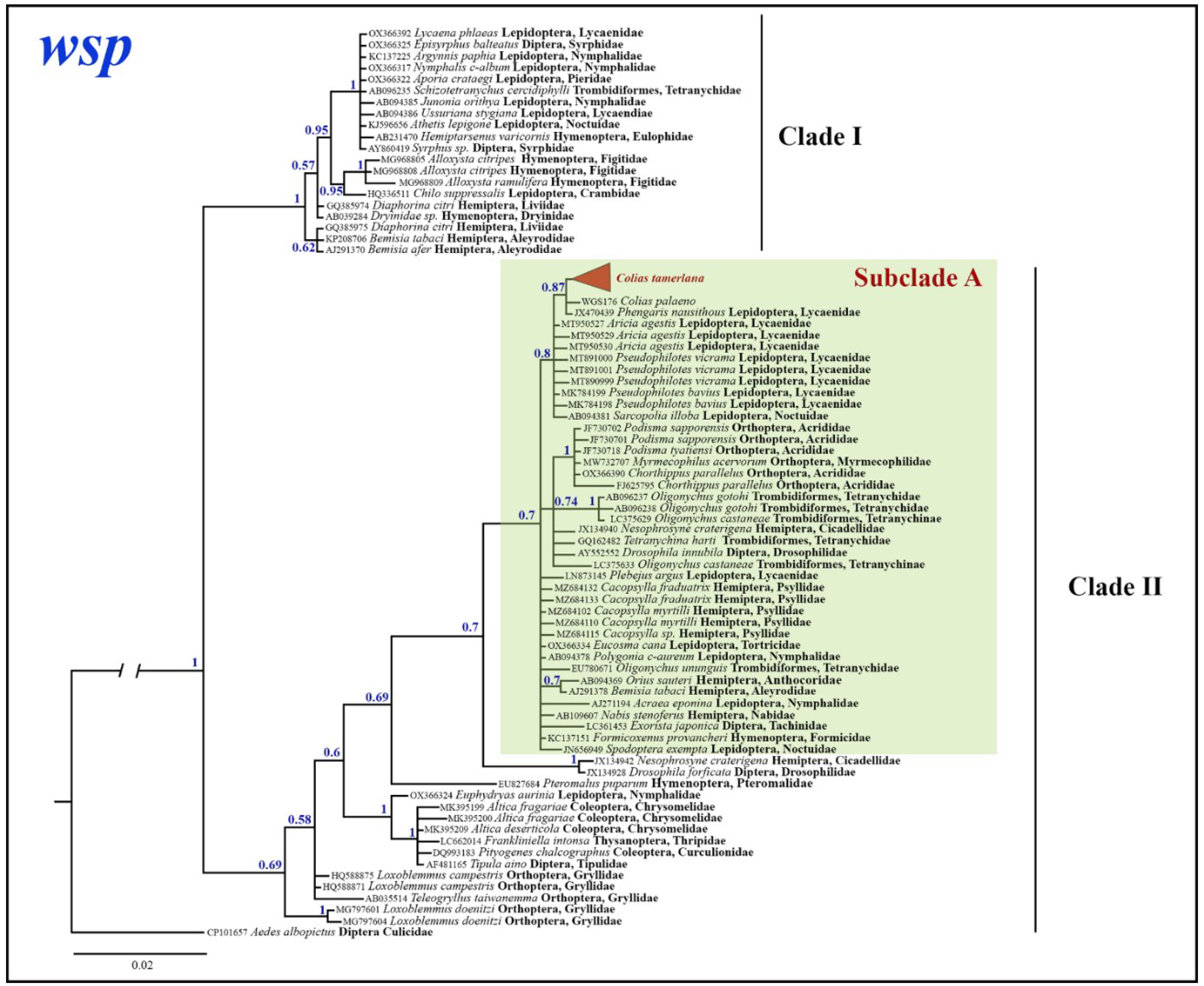
Figure 8.
The Bayesian tree of ftsZ gene fragment inferred from 85 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
Figure 8.
The Bayesian tree of ftsZ gene fragment inferred from 85 Wolbachia samples. Numbers at nodes indicate Bayesian posterior probability.
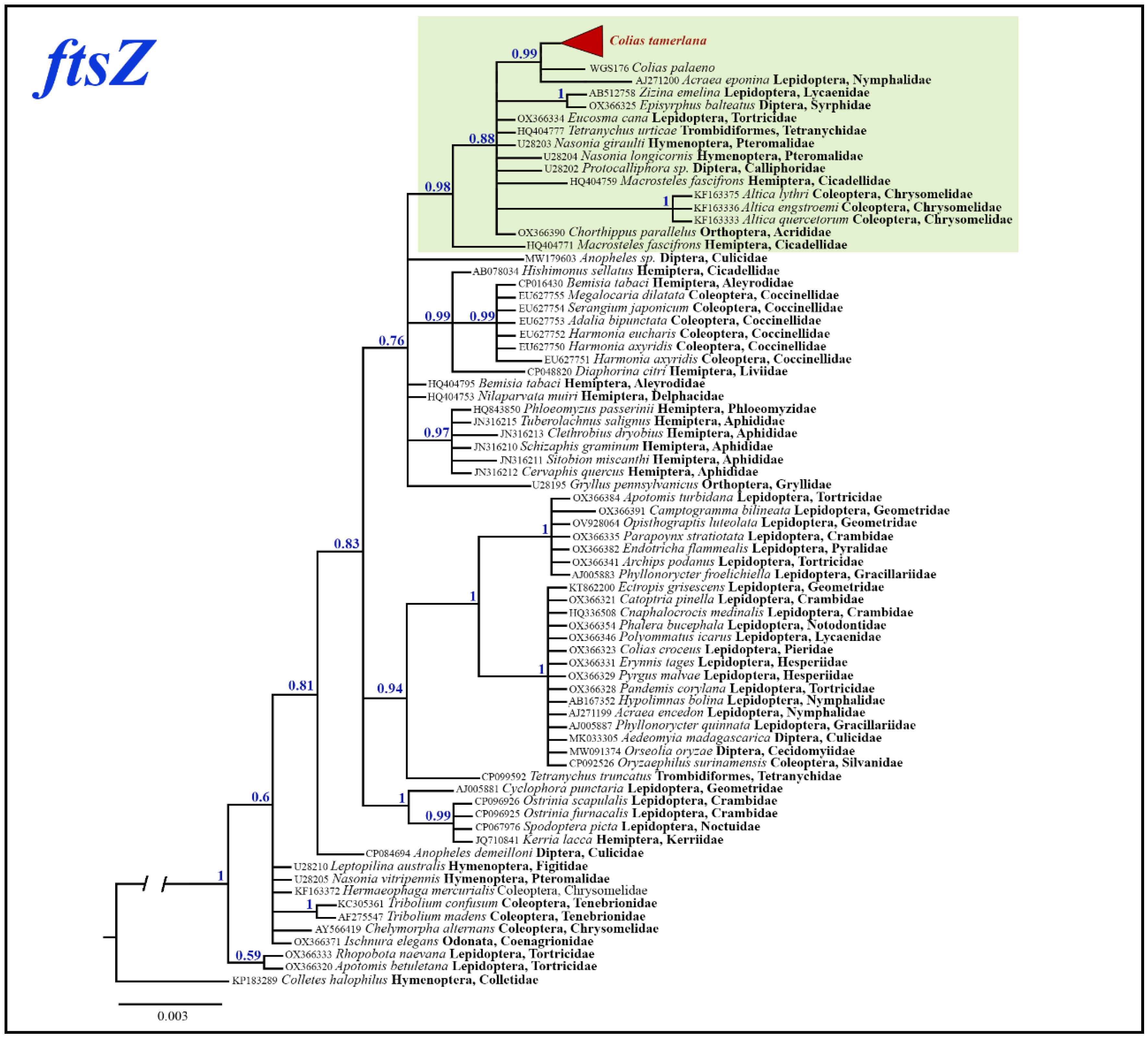

Table 1.
List of the studied specimens. LT – lectotype; PT – paratype; PLT – paralectotype; *— DNA barcodes, mined from BOLD; **—numbers in brackets correspond to sampling localities, shown in Figure 3b.
Table 1.
List of the studied specimens. LT – lectotype; PT – paratype; PLT – paralectotype; *— DNA barcodes, mined from BOLD; **—numbers in brackets correspond to sampling localities, shown in Figure 3b.
| Taxon | Sample ID | Sex | COI haplotype | Wolbachia | Year | Locality | ||
|---|---|---|---|---|---|---|---|---|
| 16S | wsp | ftsZ | ||||||
| tamerlana | Akr01 | ♂ | hp3a | - | - | - | 2016 | (01)** China, Xinjiang |
| tamerlana | Akr02 | ♂ | hp3a | - | - | - | 2016 | (01) China, Xinjiang |
| tamerlana | Akr03 | ♀ | hp3a | - | - | - | 2016 | (01) China, Xinjiang |
| tamerlana | Akr04 | ♀ | hp3a | - | - | - | 2016 | (01) China, Xinjiang |
| tamerlana | CL162 | ♂ | hp3a | - | - | - | 2016 | (01) China, Xinjiang |
| tamerlana LT | 21128B07 | ♂ | hp1b | n/a | n/a | n/a | 1896 | (01) China, Xinjiang |
| tamerlana PLT | 20022A02 | ♀ | hp1b | n/a | n/a | n/a | 1896 | (01) China, Xinjiang |
| mongola | Mnt03 | ♂ | hp1b | - | - | - | 2018 | (02) Mongolia, Hovd |
| mongola | Mnt04 | ♀ | hp3b | - | - | - | 2018 | (02) Mongolia, Hovd |
| mongola | CL34m | ♂ | hp1c | - | - | - | 2015 | (02) Mongolia, Hovd |
| mongola | Mnt54 | ♀ | hp1b | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt55 | ♀ | hp2 | + | + | + | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt56 | ♀ | hp1b | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt57 | ♀ | hp1b | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt58 | ♀ | hp2 | + | + | + | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt59 | ♀ | hp3a | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt60 | ♀ | hp2 | + | + | + | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt61 | ♂ | hp1b | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt62 | ♂ | hp2 | + | + | + | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt63 | ♂ | hp1b | - | - | - | 2003 | (17) Mongolia, Hovd |
| mongola | Mnt01 | ♀ | hp2 | + | + | + | 2013 | (03) Mongolia, Gobi-Altai |
| mongola | Mnt02 | ♀ | hp2 | + | + | + | 2013 | (03) Mongolia, Gobi-Altai |
| mongola | CL41m | ♀ | hp2 | + | + | + | 2017 | (03) Mongolia, Gobi-Altai |
| mongola | Nsk16 | ♂ | hp2 | - | - | - | 2005 | (04) Mongolia, Bayan-Ulegej |
| mongola | CL39m | ♀ | hp2 | + | + | + | 2017 | (05) Mongolia, Bayan-Ulegej |
| mongola | CL40m | ♀ | hp3a | - | - | - | 2017 | (05) Mongolia, Bayan-Ulegej |
| mongola | CL43m | ♀ | hp2 | + | + | + | 2017 | (05) Mongolia, Bayan-Ulegej |
| mongola | Mnt24z | ♀ | hp3a | - | - | - | 2017 | (05) Mongolia, Bayan-Ulegej |
| mongola | Mnt25z | ♀ | hp4a | - | - | - | 2017 | (05) Mongolia, Bayan-Ulegej |
| mongola | Mnt53 | ♀ | hp2 | + | + | + | 2004 | (16) Mongolia, Zavkhan |
| mongola ukokana | Mnt18 | ♀ | hp2 | + | + | + | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Mnt19 | ♀ | hp3c | - | - | - | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Mnt20 | ♂ | hp4a | - | - | - | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Mnt21 | ♂ | hp4a | - | - | - | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Mnt22 | ♂ | hp4a | - | - | - | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Mnt23 | ♂ | hp2 | - | - | - | 2022 | (06) Russia, Republic of Altai |
| mongola ukokana | Nsk014 | ♂ | hp3a | n/a | n/a | n/a | 1997 | (07) Russia, Republic of Altai |
| mongola ukokana | Mnt34z | ♂ | hp3a | n/a | n/a | n/a | 1997 | (07) Russia, Republic of Altai |
| mongola ukokana PT | Nsk015 | ♂ | hp4c | n/a | n/a | n/a | 1995 | (08) Russia, Republic of Altai |
| mongola ukokana | Mnt24 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt25 | ♀ | hp1a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt26 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt27 | ♀ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt28 | ♀ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt29 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt30 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt31 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt32 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt33 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt34 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt35 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt36 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt37 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt38 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt39 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt44 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt45 | ♂ | hp4a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt46 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt47 | ♂ | hp3d | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt48 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt49 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt50 | ♂ | hp3d | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt51 | ♂ | hp3a | - | - | - | 2001 | (09) Russia, Republic of Altai |
| mongola ukokana | Ku01 | ♀ | hp3e | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Ku02 | ♀ | hp2 | + | + | + | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Ku03 | ♂ | hp4a | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Ku04 | ♂ | hp4a | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt28z | ♂ | hp3a | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt29z | ♀ | hp3e | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt30z | ♂ | hp3d | - | - | - | 2022 | (09) Russia, Republic of Altai |
| mongola ukokana | LOWA141-06* | ♂ | hp3a | n/a | n/a | n/a | 1999 | (09) Russia, Republic of Altai |
| mongola ukokana | LOWA818-06* | ♂ | hp3a | n/a | n/a | n/a | 1999 | (09) Russia, Republic of Altai |
| mongola ukokana | LOWA142-06* | ♂ | hp4b | n/a | n/a | n/a | 1999 | (09) Russia, Republic of Altai |
| mongola ukokana | CL42m | ♀ | hp1a | - | - | - | 2017 | (09) Russia, Republic of Altai |
| mongola ukokana | Mnt17 | ♀ | hp1a | - | - | - | 2022 | (10) Russia, Republic of Altai |
| mongola ukokana | Mnt40 | ♀ | hp2 | + | + | + | 2003 | (10) Russia, Republic of Altai |
| mongola ukokana | Mnt41 | ♂ | hp3a | - | - | - | 2003 | (10) Russia, Republic of Altai |
| mongola ukokana | Mnt42 | ♂ | hp3a | - | - | - | 2003 | (10) Russia, Republic of Altai |
| mongola ukokana | Mnt43 | ♂ | hp3a | - | - | - | 2003 | (10) Russia, Republic of Altai |
| mongola ukokana | Mnt09 | ♂ | hp3a | - | - | - | 2002 | (11) Russia, Republic of Altai |
| mongola ukokana | Mnt10 | ♂ | hp3a | - | - | - | 2002 | (11) Russia, Republic of Altai |
| mongola ukokana | Mnt08 | ♂ | hp3a | - | - | - | 2003 | (12) Russia, Republic of Altai |
| mongola ukokana | Mnt12 | ♂ | hp3a | - | - | - | 2003 | (12) Russia, Republic of Altai |
| mongola | Mnt14 | ♀ | hp2 | + | + | + | 2010 | (13) Russia, Republic of Tyva |
| mongola | Mnt15 | ♀ | hp3a | - | - | - | 2010 | (13) Russia, Republic of Tyva |
| mongola | Mnt07 | ♂ | hp2 | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL35m | ♀ | hp1b | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL37m | ♂ | hp1b | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL38m | ♀ | hp1a | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL54m | ♂ | hp1b | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL55m | ♂ | hp4a | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL56m | ♂ | hp1b | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | CL57m | ♀ | hp1b | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | Mnt26z | ♂ | hp1a | - | - | - | 2002 | (14) Russia, Republic of Tyva |
| mongola | Mnt05 | ♀ | hp1b | - | - | - | 2001 | (15) Russia, Republic of Tyva |
| mongola | Mnt06 | ♀ | hp1b | - | - | - | 2001 | (15) Russia, Republic of Tyva |
| mongola LT | 20022A03 | ♂ | hp1b | n/a | n/a | n/a | ? | (??) see text for explanation |
| sidonia | S1-22064B02 | ♀ | hp1b | n/a | n/a | n/a | 1967 | (18) Mongolia, Khovsgol |
| sidonia | S2-22064B03 | ♀ | hp1b | n/a | n/a | n/a | 1966 | (18) Mongolia, Khovsgol |
| sidonia | S3-22064B04 | ♂ | hp1b | n/a | n/a | n/a | 1967 | (18) Mongolia, Khovsgol |
| sidonia | S4-22064B05 | ♂ | hp1b | n/a | n/a | n/a | 1967 | (18) Mongolia, Khovsgol |
Table 2.
Summary of haplotype diversity of C. mongola/C. tamerlana. The number of individuals sequenced (N), the number of haplogroups (HG) and haplotypes (H) revealed, the number of polymorphic sites (S), nucleotide (π) and haplotype (h) diversities are given. Tajima’s D, Fu and Li’s D, Fu and Li’s F, Fu’s Fs, max. p-distance, within and between group divergence values with standard deviation (in brackets) are shown.
Table 2.
Summary of haplotype diversity of C. mongola/C. tamerlana. The number of individuals sequenced (N), the number of haplogroups (HG) and haplotypes (H) revealed, the number of polymorphic sites (S), nucleotide (π) and haplotype (h) diversities are given. Tajima’s D, Fu and Li’s D, Fu and Li’s F, Fu’s Fs, max. p-distance, within and between group divergence values with standard deviation (in brackets) are shown.
| N | HG | H | S | π | h | Tajima’s D | Fu and Li’s D | Fu and Li’s F | Fu’s Fs | max. p-distance | within group divergence | between group divergence |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 101 | 4 | 12 | 28 | 0.0138656 | 0.7874 | 2.01246 | 1.21273 | 1.82127 | 7.422 | 2.89% | HP_I 0.04% (± 0.02%) | HP_I / HP_II 2.56% (± 0.59%) |
| 0.1 > p > 0.05 | p>0.1 | p<0.05 | p=0.001 | (± 0.62%) | HP_II 0.00% (±0.00%) | HP_I / HP_III 2.40% (± 0.57%) | ||||||
| HP_III 0.07% (± 0.05%) | HP_I / HP_IV 2.56% (± 0.58%) | |||||||||||
| HP_I 0.04% (± 0.03%) | HP_II / HP_III 1.39% (± 0.42%) | |||||||||||
| HP_II / HP_IV 1.23% (± 0.41%) | ||||||||||||
| HP_III / HP_IV 1.41% (± 0.43%) |
Table 3.
Haplogroup and haplotype composition (the number of individuals constituted a haplogroup/haplotype (n), and the number of sampling sites where haplogroup/haplotype was found (Sn) are given) and its geographical distribution (the number of individuals (n), the number of haplogroups (HGn), the number of haplotypes (Hn) found in each locality and haplotype constitution are shown). * - sampling sites correspond to Figure 3b.
Table 3.
Haplogroup and haplotype composition (the number of individuals constituted a haplogroup/haplotype (n), and the number of sampling sites where haplogroup/haplotype was found (Sn) are given) and its geographical distribution (the number of individuals (n), the number of haplogroups (HGn), the number of haplotypes (Hn) found in each locality and haplotype constitution are shown). * - sampling sites correspond to Figure 3b.
| Haplogroup | n | Sn | COI haplotype | n | Sn | Samplingsite* | n | HGn | Hn | COI haplotype (n) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HP_I | 26 | 9 | hp1a | 5 | 4 | 01 | 7 | 2 | 2 | hp1b (2), hp3a (5) | |||
| hp1b | 20 | 6 | 02 | 3 | 2 | 3 | hp1a (1), hp1c (1), hp3b (1) | ||||||
| hp1c | 1 | 1 | 03 | 3 | 1 | 1 | hp2 (3) | ||||||
| HP_II | 17 | 10 | hp2 | 17 | 10 | 04 | 1 | 1 | 1 | hp2 (1) | |||
| HP_III | 42 | 11 | hp3a | 35 | 9 | 05 | 5 | 3 | 3 | hp2 (2), hp3a (2), hp4a (1) | |||
| hp3b | 1 | 1 | 06 | 6 | 3 | 3 | hp2 (2), hp3c (1), hp4a (3) | ||||||
| hp3c | 1 | 1 | 07 | 2 | 1 | 1 | hp3a (2) | ||||||
| hp3d | 3 | 1 | 08 | 1 | 1 | 1 | hp4c (1) | ||||||
| hp3e | 2 | 1 | 09 | 35 | 4 | 7 | hp1a (2), hp2 (1), hp3a (17), hp3d (3), hp3e (2), hp4a (9), hp4b (1) | ||||||
| HP_IV | 16 | 4 | hp4a | 14 | 4 | 10 | 5 | 3 | 3 | hp1a (1), hp2 (1), hp3a (3) | |||
| hp4b | 1 | 1 | 11 | 2 | 1 | 1 | hp3a (2) | ||||||
| hp4c | 1 | 1 | 12 | 2 | 1 | 1 | hp3a (2) | ||||||
| 13 | 2 | 2 | 2 | hp2 (1), hp3a (1) | |||||||||
| 14 | 9 | 3 | 4 | hp1a (2), hp1b (5), hp2 (1), hp4a (1) | |||||||||
| 15 | 2 | 1 | 1 | hp1b (2) | |||||||||
| 16 | 1 | 1 | 1 | hp2 (1) | |||||||||
| 17 | 10 | 3 | 3 | hp1b (5), hp2 (4), hp3a (1) | |||||||||
| 18 | 4 | 1 | 1 | hp1b (4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



