
Submitted:
03 November 2023
Posted:
06 November 2023
You are already at the latest version
Abstract
The circadian clock is a 24-hour cycle within the body that regulates various factors, including sleep, body temperature, and hormone secretion. It allows the body to adjust to the light and dark cycle. Disruptions to the circadian rhythm are important risk factors for many diseases, including neurodegenerative illnesses. The central clock is controlled by the clock genes in the suprachiasmatic nucleus (SCN) of the brain, and one of the main functions of the circadian clock is to regulate lipid metabolism. There has been a lack of investigation on the circadian regulation of lipid metabolism-related apolipoprotein genes in the brain; thus, this study sought to summarize the rhythmic expression of clock genes and lipid metabolism-related apolipoprotein genes within the SCN of Mus musculus. Out of the 20 apolipoprotein genes that were studied by the published database, all 9 genes were highly expressed in the SCN. Most of the 6 genes showed rhythmic expression in the brain of mice, suggesting that the master clock might regulate them. The results, therefore, reveal a potential target for preventing and treating circadian disorders. We summarized most studies of lipid-related apolipoprotein genes within limited locations of the SCN and brain to further understand how disordered cerebral lipid metabolism causes multiple brain diseases and disorders. We reviewed recent studies, determined the unexplored questions, and identified a potential direction for further research.
Keywords:
lipid
; apolipoprotein
; circadian clock
; brain
; neuron
; hormone
1. Introduction
The circadian rhythm is a 24-h cycle in which the body adjusts physically and behaviorally to the light and dark phases throughout the day. When the eye receives light from the surrounding environment, the body controls various factors, including the sleep-wake cycle, body temperature, hormone secretion, and blood pressure [1,2,3]. However, there are several common disruptions to the circadian rhythm, such as clock genes mutations, medications, stress, aging, irregular work schedules, changes in time zones, and vision impairment [4]. Frequent disruptions to the internal clock are a risk factor for multiple disorders: cancer, sleeping disorders (i.e., jet lag), atherosclerosis, obesity, diabetes, viral infections (i.e., COVID-19), and neurodegenerative diseases (i.e., Alzheimer’s disease) [5,6,7,8,9,10,11,12].
Apolipoproteins are major constituents of lipoprotein, which serve as lipid transporters by binding to lipids and secreting them into the blood, lymph, and cerebrospinal fluid (CSF). Apolipoprotein synthesis is controlled by varying factors, such as age and gender; these varying controls are exercised differently depending on the tissue type. Studies have reported that plasma levels of all apolipoproteins, except apolipoprotein (Apo)H, were higher in females than in males [13]. Women only had significant negative associations of ApoB, ApoE, ApoH, and ApoJ in mid-life, while associations at older age were non-significant or positive. Plasma levels of apolipoproteins, including in ApoA1, ApoA2, ApoB, ApoC3, and ApoH, play a negative relationship with increasing age starting from mid-life; however, plasma levels of ApoE and ApoJ were found to observe U-shaped curves with age from mid-life, reacting to higher levels of ApoE and ApoJ in oldest aged male [13].
Many physiological functions and many types of disease have been ascribed to apolipoproteins. For example, we have shown that the circadian clock regulates the ApoB-containing lipoprotein pathway to control lipid absorption and plasma lipids in clock circadian regulator (Clock)-mutant mice [14,15]. Global or liver-specific basic helix-loop-helix ARNT like 1 (Bmal1)-deficient mice increase ApoB-containing very low-density lipoprotein (VLDL) assembly and secretion as well as the development of atherosclerosis [16]. In addition, we also have identified that ApoA-IV is mainly targeted by Bmal1, when combined with nuclear receptor subfamily 1 group D member 1 (Nr1d1), to control VLDL lipoprotein assembly and secretion in the liver via the cyclic adenosine monophosphate (cAMP)-responsive element-binding protein H [17]. This directly governs ApoA4 expression in the liver and intestine to increase VLDL lipoprotein particle size in the circulation blood [17]. ApoA4 is primarily synthesized in the small intestine, which plays a vital role in the circadian regulation of intestinal lipid absorption and mediates lipoprotein production [18]. High concentration of total cholesterol in serum at middle aged is a risk factor for AD and other dementia types in later life and is directly related with higher risk of dementia mortality. Recently studies have suggested that there is a strong relationship between Alzheimer’s disease (AD) and cardiovascular disease (CVD) risk factors such as high density lipoprotein (HDL) levels, low density lipoprotein (LDL) levels and the presence of atherosclerosis and hypertension [19]. Studies have shown that lowering cholesterol agents such as statins, is able to reduce the incidence of AD [20,21], although the mechanism is still unknown.
In peripheral tissues, many genes involved in lipid biosynthesis and metabolism are rhythmically controlled by circadian clock genes [6,22,23,24], suggesting that the dysfunction of clock gene results in abnormal lipid metabolic and impaired lipid absorption. For example, nocturnin (gene name Ccrn4l) plays an important role in the trafficking of dietary lipid in the intestinal enterocytes by optimizing efficient absorption of lipids [25]. Nocturnin-/- mice have shown to decrease chylomicron (CM)/VLDL triglyceride and cholesterol levels [25]. Several studies have determined that circadian clock regulation is essential for normal physiology and pathology mechanism.
The circadian clock may also play a significant role in the regulation of cerebral lipid metabolism and help prevent apolipoprotein-associated neurodegenerative diseases, such as AD [26,27,28,29,30,31]. Several apolipoproteins have been identified to be directly implicated in the etiopathology of AD, including APOE, clusterin (CLU or apoJ), APOC1, and APOB [32,33,34,35].
In this focused review, we first introduce the SCN, circadian clock, apolipoproteins in the SCN, and then discuss recent known or hypothesized roles in Brain physiology and diseases. This review does not cover all of the apolipoproteins’ functions and modulations of different lipid metabolism pathways or those involved in the different diseases of apolipoproteins; other reviews can glean this information [35,36,37,38].
2. SCN Systemic
2.1. SCN
The brain suprachiasmatic nucleus (SCN) is a minor area within the brain’s hypothalamus. Each unilateral SCN has about 10,000 neurons in two anatomic subdivisions. SCN is the main input and output location for light, neuronal, and hormonal activities [39,40,41],. The SCN has many different cell types, such as neuropeptides, neurotransmitters, and input or output connections. Wen et al. have identified eight major cell types, with each type displaying a specific pattern of circadian gene expression by single-cell RNA-sequencing [42]. Interestingly, several studies have shown that astrocytes contribute to the circadian rhythm in the SCN [43,44,45,46]. These studies have suggested that the SCN is the most critical region for sleep, mood, control of circadian timing, and non-neural circadian gene expression and regulation. Studies have shown that removal of the SCN results in the ablation of circadian rhythm in mammals [47,48,49], suggesting that the SCN is the primary center for circadian regulation in mammals. Also, individual neurons of SCN can produce independent circadian oscillations of circadian clock gene expression and nerve reflex by ex vivo culture.
2.2. Neurons and Hormone in SCN
There are several different peptide hormones such as vasopressin (AVP), vasoactive intestinal peptide (VIP), peptide histidine-isoleucine (PHI), and neurotransmitters found within the SCN [50]. In mice, VIP makes up 10% of all SCN cells while AVP neurons make up 20% of all SCN cells [51]. While AVP regulates blood pressure [52,53], VIP regulates the release of various bodily chemicals during digestion [54]. PHI regulates food consumption behavior [55]. These hormones are essential to control energy and glucose homeostasis. Studies have shown that the circadian rhythms of AVP and VIP secretion were affected by gap junction (which is cell-cell junction) blockers through rat SCN slice cultures [56]. When the gap junction blockers were taken out of the culture, the circadian rhythms of AVP and VIP reappeared, suggesting that the hormones may be regulated by cell-cell interaction in SCN, such as neuron and astrocytes cross-talking [56]. In addition, the circadian clock may control hormones, which might possibly be controlled by cell-cell interaction. The central circadian clock may regulate physiological and pathological functions in peripheral tissues by hormones and peripheral tissue cross-talking.
Sex hormone receptors, such as estrogen receptor-ɑ (ERɑ), estrogen receptor-ꞵ (ERꞵ), progesterone receptors, and androgen receptors are expressed in the SCN [57]. The SCN regulates circadian rhythms in sex hormone secretion; alternately, sex hormone feedback also influences SCN functions. ERɑ expression was more statistically significant in the SCN of females, which may explain the differences between the male SCN and the female SCN [58]. Pre-astroglia (astrocytes and ependymocytes) have a gender-related role in the organization of the SCN but not the sexually dimorphic area, and neuroglia also has a gender-related role in the mature SCN of Mongolian gerbils [59].
Glucagon-like peptide 1 (GLP-1) is mainly synthesized in the intestine, but it is also synthesized in the brain, especially in the nucleus tractus solitarius, which is located in the brainstem [60]. Also, GLP-1 is expressed in neurons of the hypothalamus, especially in the paraventricular nucleus and arcuate nucleus [61]. Additionally, the GLP-1 receptor is expressed in neurons of the central and peripheral nervous system. Synthetic GLP-1 receptor agonists are recently approved by the FDA for the treatment of type-2 diabetes mellitus, and to reduce overweight in obesity [62,63]. Several studies have shown in animal models, GLP-1 receptor agonists may decrease motor, behavioral, and cognitive symptoms in Parkinson’s disease (PD) and AD [64], thus, may slowing development of neurodegeneration.
2.3. Immune Factors in SCN
The SCN is of immense biological importance, functioning as the internal clock of the brain that guides sleep patterns, ovulation cycles, and other of the sort. The glial fibrillary acidic protein, a specific marker for astrocytes, is located in the SCN [65]. The glial cells of the SCN can regulate input signals to the mouse’s circadian system by receiving them from the immune system via NF-kappaB signaling [65,66]. Interleukin-1β (IL-1β) and Interleukin-1 Type 1 Receptor (IL-1R1) were expressed in the SCNs of young and old groups of mice. However, IL-1β was more highly expressed in younger mice, and IL-1R1 showed rhythmic expression only in younger mice. The rhythmic expressions of both IL-1β and IL-1R1 in the paraventricular nucleus of the brain were changed by age [67]. The study then found when lipopolysaccharide (LPS; 5mg/kg) is added to stimulate an immune response, only the expression of IL-1R1 increases after 6 and 24 h of treatment [67]; these data suggest that diurnal, age, and immune regulation of IL-1β and IL-1R1 in the mouse SCN. The other markers of immune factors include interferon-γ, tumor necrosis factor-α (TNFα), showed a reduction in excitatory SCN activity after being treated with LPS [68]. Thus, alterations to the synaptic mechanisms of SCN neurons might also have an integral role in sleeping disorders, and proinflammatory cytokines share in these changes.
3. Circadian Rhythm of Clock Genes in SCN
Within the body, the brain’s SCN controls the central clock, yet peripheral clocks are located in every tissue. The two nuclei are located near the hypothalamus, with approximately 10,000 neurons controlling a spectrum of changes occurring within the body throughout the day. Numerous clock genes regulate the circadian rhythm, specifically including Clock, Bmal1, period circadian regulators (Per1, Per2, Per3), and cryptochrome circadian regulators (Cry1, Cry2). The Clock gene encodes a protein part of a heterodimer and a Bmal1-encoded protein. This heterodimer binds to the E-box enhancers, binding sites, that increase the transcription process upstream of the Per1, Per2, Per3, Cry1, and Cry2 genes. After the genes undergo transcription, the Per encoded proteins and the Cry encoded proteins combine to produce a heterodimer to repress their self’s transcription by interacting with the BMAL1:CLOCK/NPAS2 complex [69].
To date, only one study has used targeted lipidomic approaches to examine the circadian course of lipids in the human brain [70]. Using the SCNseq database, we found that most of the studied circadian rhythm-related genes were present in the SCN of male mice, and Figure 1 shows that all seven of the studied genes were expressed in similar proportions. Cry2 had the highest expression when compared with the other 6 genes. All of the studied circadian clock-related genes showed rhythmic expression over a 24-h cycle. The shaded parts of the graph represent the dark phases of the 24-h cycle, and similar patterns were observed and organized in Figure 2. Bmal1 (Arntl), Per1, Per2, Per3, and Cry2 all showed their highest peaks during the daytime while showing their lowest expressions in the nighttime. Further, because these expression levels were measured in mice, their nocturnal behavior must also be considered. Their inactive phase is during the day, and their active phase is during the night.
The circadian cock regulates the immune system; in turn, the immune system also plays an essential role on the circadian rhythm of circadian clock genes in the SCN. Adults with prenatal LPS exposure appeared to have increased anxiety-like behavior along with an altered rhythm of clock genes in the SCN [71]: the most significant impact was on Nr1d1 expression, while the least impacted was Per2. However, the developing SCN also showed adaptive flexibility to immune challenges in the early stages of development. While Nr1d1 in the LPS-treated group was arrhythmic at postnatal day 3, it showed positive improvement in the form of a higher amplitude by postnatal day 20 [71], suggesting that the features of adaptive flexibility of the circadian clock, which may help the immune system to cope with challenges later in life.
When the circadian clock is disrupted by the deletion of Bmal1 in the SCN of mice brain, amyloid-β starts to build up within the brain. Once the circadian rhythm is disrupted locally in the brain parenchyma, the expression of ApoE increases, and fibrillar plaque, starts to accumulate [72,73,74], suggesting the circadian clock plays an essential role in the regulation of amyloid-β and ApoE dynamics and pathology. As we known, AD is characterized by plaque accumulation in the brain, including amyloid-β buildup, and changes in sleep cycle are proven to be symptoms of AD.
4. Circadian Rhythm of Apolipoprotein Genes in SCN
One of the circadian clock’s functions is regulating lipid synthesis and transport. We and other groups have shown that circadian clock genes regulate lipid metabolism in peripheral tissues, such as the intestine and liver [12,23,75,76,77,78,79]. Apolipoproteins are essential for regulating tissue and plasma lipid levels in peripheral tissues and the central brain [80,81]. Lipids can be transported throughout the body through lipoproteins, and are produced when apolipoprotein molecules and lipids, like cholesterol and triglycerides, bind together [82]. Specific apolipoprotein genes encode these apolipoproteins; specifically, apolipoprotein E (ApoE), clusterin (Clu), apolipoprotein A1 (ApoA1), apolipoprotein C1 (ApoC1), apolipoprotein H (ApoH), and apolipoprotein M (ApoM) were closely examined in different studies. There are 22 known apolipoproteins. 9 apolipoproteins were found in the central nervous system (CNS), and 8 apolipoprotein messenger RNA (mRNA) were found in the brain. Twenty years ago, Stuerenburg et al. determined that there are several apolipoproteins in 13–20 nm particles of lipoprotein fraction, such as ApoE, ApoA-I, ApoA-IV, ApoD, ApoH, and ApoJ [83]. Importantly, ApoE is usually presented to the largest particles. ApoA-I and ApoA-II are found on smaller particles, and ApoJ is allocated across the particle size range. Other apolipoproteins such as ApoA-IV, ApoD, and ApoH are also found in the CSF [83]. ApoE and ApoA1 are found in higher concentrations in the CSF compared with ApoJ, ApoD, ApoA-II and ApoA-IV [83,84].
HDL, Apo AI, ApoJ, and some free fatty acids and triglycerides may cross the blood brain barrier (BBB), while others, such as ApoE do not [85]. The same principle applies to cholesterol: some forms may cross the barrier while others cannot. Additionally, lipids may have an interactive role with the BBB regarding its functionality: HDL can protect the barrier during multiple sclerosis, cholesterol may disrupt the barrier, and triglycerides can inhibit the transportation of leptin across the barrier [86]. While the aforementioned ApoE cannot cross the BBB, the human APOE4 allele can have damaging effects on BBB’s function [85]. As demonstrated, lipids, lipoproteins and apolipoproteins relation to the BBB range from functional to detrimental and are integral to the regulation of health and the study of disease.
The topic of cholesterol is a relevant neurological issue, and its metabolism is largely controlled by astrocytic activity; adult neurons import cholesterol through ApoE-abundant particles via astrocytes in place of natural biosynthesis. Cholesterol is necessary for a multitude of neural functions; in addition, cholesterol dyshomeostasis impacts amyotrophic lateral sclerosis, a common neurodegenerative disease in adults [87]. Neurons regulate cholesterol synthesis and metabolism through ApoE, cholesterol is then secreted from glial cells, specifically astrocytes. Using human U373 (glioblastoma astrocytoma cells) astrocyte-derived cell line, cholesterol metabolism-related proteins were increased when the glial cells were treated with nerve growth factors (NGF). ApoE secretion and the amount of cholesterol increased when the cells were treated with NGF [88]. Importantly, neuroprotection was suppressed when N1E-115 (an adrenergic cell line derived from mouse neuroblastoma C1300) mouse neuroblastoma cells were cultured with ApoE-silenced cells unlike when it was granted with NGF treatment. NGF was also necessary to activate neuroprotection in N1E-115 neurons [88].
Lipoproteins are not only responsible for the transportation and processing of lipophilic substances within aqueous bodily fluids, but also involved in other physiological functions, such as immune regulation [89]. Relatedly, neurons are very sensitive to immune responses and the supply of lipids, which further supports the possible relationship between lipoproteins and neurodegenerative diseases.
HDL and plasma concentrations of certain apolipoproteins can also be a potential biomarker of covert brain infarcts. Manja Koch et al. found that in subspecies with lower concentrations of ApoC3, ApoJ, or ApoE, higher ApoA1 concentrations were correlated with fewer brain infarcts [90]. Similarly, in subspecies with lower concentrations of ApoC3 or ApoJ, higher apoE concentrations were correlated with greater covert infarcts [90]. It is suggesting that HDL is a potential method of drug delivery to tumors even across the BBB, specifically with poorly soluble or unstable therapeutics [91].
To understand how apolipoprotein genes are expressed in the SCN, we checked several apolipoprotein genes from the SCNseq and CircaDB. We found that ApoJ (Clu) has the highest expression in SCN, compared with ApoE and ApoC1 (Figure 3), suggesting that these apolipoproteins are expressed and synthesized within the brain tissue. It is possible that these apolipoproteins were expressed in locations other than in the SCN. The other apolipoprotein genes were lowly expressed, indicating that these apolipoproteins might be synthesized in peripheral tissues and brought to the brain through circulation. However, ApoM, ApoB, ApoA4 did not show any expression within the SCN of mice. Interestingly, ApoA1, ApoC3, and ApoH are also expressed in SCN. In addition, we found that, like the circadian clock-related genes, all the studied apolipoprotein genes also showed rhythmic expression (Figure 4). Although ApoM did not show expression in the SCN, it did show changes in expression levels over a 24-h cycle in the overall cerebral tissue. ApoA1, ApoC1, and ApoE all showed their highest peaks in the daytime when the mice were inactive. On the other hand, ApoH, ApoM, and Clu showed the highest activity in their active phase.
In the following discussion, we have not discussed all apolipoproteins that may bind to/transport lipids; instead, we have focused on those genes that have both been in recent years of research studies and have been able to be detected in the SCN of the brain at the genetic level by SCNseq. (http://www.wgpembroke.com/shiny/SCNseq/).
4.1. ApoA1
ApoA1 can be synthesized in brain endothelial cells and conduce to the CNS pool [92,93]. ApoA1 protein was detected in the CSF; however, mRNA was not expressed in brain tissue. ApoA1 is a major component of HDL and acts in the transportation of cholesterol from peripheral tissues to the liver. Despite being synthesized in the lower abdominal region (liver, intestine and kidney), ApoA1 can enter the brain at the choroid plexus, crossing the BBB, therefore, might be implicated in brain lipid homeostasis [94]. ApoE is a well-known risk-factor for the development of AD with a protective allele variant (ε2) and a risk-increasing allele variant (ε4); however, ApoA1 has not been explored with depth. As a major component of HDL, it acts in the transportation of cholesterol from peripheral tissues to the liver.
When ApoA1 enters the brain, it can co-localize with occasional Aß plaques in the cortex of AD patients [89], suggesting that ApoA1 might affect amyloid precursor protein (APP) cleavage and Aß aggregation and/or clearance. However, this needs to be further clarified. Vollbach et al. have shown that polymorphism in the promoter region of the APOA1 gene (-75 A/G) is associated with an earlier age of AD onset. They found that individuals homozygous for the A allele (at this location) develop AD, on average 8 years earlier than heterozygotes [95,96]. ApoA1 is also linked with a variety of neurodegenerative diseases, such as PD, and Schizophrenia [97].
4.2. ApoA4
Apolipoprotein A4 (also known as ApoA-IV), is a plasma protein, mainly in HDL as well as on the surface of newly synthesized CMs, to control lipid absorption, plasma lipids, and transport cholesterol from peripheral cells back to the liver via several steps of the reverse cholesterol transport pathway [17,18,98].
ApoA-IV is found to be synthesized in the hypothalamus and colocalizes mainly with neurons and with glia in the brain region [99,100]. It is detected in astrocytes of rat [101]. Expression of ApoA-IV is increased following a high-fat meal, and ApoA-IV has been demonstrated to play a central role in satiety [102,103,104]. Evidence suggests that a polymorphic version of ApoA-IV, which is more efficient at activating lecithin cholesterol acyl transferase, may be associated with increased AD risk [105]. In mice studies, the genetic ablation of APOA4 may promote AD pathogenesis in an APP/presenilin 1 (PS1) transgenic mouse model [106]. In human study, the APOA4 codon 360 mutation (C > T) is related with the increased risk of AD patients [107,108].
4.3. ApoB
ApoB in the liver is controlled by several factors such as hormones (i.e., insulin), dietary factors (fatty acid), and variation therapies drugs (i.e., statins, niacin, and fibric acids) [109]. As mentioned before, cerebral ApoB mRNA is easily detected with RNA-Seq techniques. ApoB-containing lipoproteins are detected in the CSF and are found at a concentration consistent with porous diffusion enhanced by CSF secretion. Pre-symptomatic participants have been shown to elevate CSF ApoB levels as well as t-tau, p-tau, and the four aforementioned synaptic markers in pre-symptomatic participants. Additionally, the CSF ApoB levels of those without AD symptoms correlated with positron emission tomography flortaucipir binding in the entorhinal, parahippocampal, and fusiform regions. Thus, CSF ApoB levels have a correlation to early tau dysregulation and can help identify those predisposed to developing visuospatial cognitive decline [83,110], suggesting that ApoB is a novel marker for early tau pathology in AD.
4.4. ApoD
ApoD is expressed and detected in several regions of the brain, including in the SCN, cortex, and cerebellum [111]. Cerebral ApoD is synthesized and excreted mainly by oligodendrocytes and astrocytes and secreted as lipoproteins. ApoD has gained attention from its expression in various disease conditions and its wide-ranging presence in tissues and subcellular locations. Based on ApoD activity, its nature is better described as lipid-management rather than lipid-transport [111]. Examples of this activity include ApoD ligand binding in the Lipocalin pocket and antioxidant activities occurring at the rim of the pocket. Additionally, ApoD can regulate a large variety of health and disease processes by controlling redox states of such lipid structures [112]. In normal conditions, neuronal ApoD protein, not mRNA, is observed in Purkinje neurons and, to a lesser extent, in cortical neurons; however, in pathological conditions, CSF ApoD protein may be increased [113] and affected in brain different regions [114,115,116].
4.5. ApoE
ApoE has been explored with much depth. The human APOE has three major alleles, including APOE2, APOE3, and APOE4, but mouse ApoE has only one isoform. The variants of the human APOE gene may cause the link between microglial cells and synaptic dysfunction and be the cause of malfunctioning lipid metabolism [117]. Understanding how risk factors affect neurons and glia is at the forefront of US public health. Over 20% of the population carry the APOE4 gene and 40% are obese, suggesting that people with ApoE4 variants in their genotype may be at a higher risk for neurodegenerative diseases such as AD, late-stage AD, Lewy body dementia, depression, traumatic brain injury (TBI), spinal cord injury (SCI) and stroke. In addition, the APOE2 allele is associated with a higher disease risk of PD [118].
There are three human ApoE alleles and six phenotypes with different metabolic implications and plasma cholesterol levels. Defective APOE2 is associated with type III dysbetalipoproteinemia due to inefficient clearance of triglyceride-rich lipoproteins. Defective APOE4 is known to increase the risk of AD as well as the age of onset and progression. Much researches have been centered on deciphering the role of APOE in the neural β-amyloid deposition to further understand why APOE4 is considered a risk factor for AD and Lewy body dementia [119]. Recently, studies used human iPSC-derived cerebral organoid models, found the lack of cerebral APOE expression increased α-synuclein buildup and lipid buildup, which are characteristics of AD and dementia [120]. Interestingly, the expression of APOE2 and APOE3 can prevent these accumulations; however, APOE4 may enhance the influence of genetics on Lewy body dementia [121]. α-synuclein buildup was higher in APOE4 carriers than APOE3 carriers.
APOE4 is a well known risk factor for the development of AD, specifically for the development of late-stage AD, and that abnormal sphingolipid levels have been implicated as well. The impairment of endolysosomal trafficking, disruption of synaptic homeostasis, and reduced amyloid clearance are due to the isoelectric point of APOE4 matching the pH of the early endosome, which induces delayed dissociation from ApoE receptors [122]. Recently, Montagne et al. have shown that breakdown of the BBB contributes to APOE4 -related cognitive decline [123].
To study how human APOE polymorphism plays a role in atherosclerosis, lipid metabolism, and AD at the animal level, humanized mouse models were discovered and used with the replacement of the mouse Apoe gene with human APOE4 [124]. APOE4 is also known to cause neuronal hyperexcitability in knock-in mice susceptible to AD, suggesting that APOE4 carriers maybe a therapeutic target during the breakdown of the BBB contributes to APOE4-related cognitive decline.
APOE is also required for a properly functioning hippocampus and for hippocampal neurogenesis after TBI in mice [125]. Female mice presented a significant APOE genotype and TBI correlation regarding alcohol consumption. APOE4 carrier status and reduced hippocampal volume were both linked with symptoms of depression [126]. However, there was a lack of data to show that decreased cognitive ability is associated with depression, suggesting that depression is associated with APOE status and hippocampal volume but not cognitive decline in older adults aging with TBI [127]. APOE levels in the amygdala and prefrontal cortex can predict relative regional brain volumes in irradiated Rhesus macaques [128]. Disease risk caused by APOE4 also depends on its effects on β-amyloid deposition, regardless of diagnosis, the gene is associated with increased β-amyloid levels [119].
Inflammatory pathways are impaired in AD and differentially associated with APOE Statue. Multiple neurological pathways are damaged in APOE4 carriers with AD when compared with APOE3 carriers [129]. APOE4 mice developed a leaky BBB [130], increased Matrix Metallopeptidase 9, damaged tight junctions, and decreased astrocyte end-foot coverage of blood vessels, whereas APOE2 and APOE3 mice did not [130]. Removal of APOE4 led to improvements in all aforementioned areas while the removal of APOE3 did not affect BBB integrity. Thus, APOE4 is a main proprietor of BBB dysfunction, and in turn, the astrocytic production of the said allele is responsible for regulating its integrity [130].
APOE4-oeverexpression mice had higher ceremide levels in the cortex than APOE3-overexpression mice; mice with a familial history of AD also showed higher ceremide levels in the cortex than mice without. The older mice (>5 months) showed higher sphingosine-1-phosphate (S1P) levels in all three brain regions compared to younger mice (<3 months). Another differentiating factor was sex, as ceramide levels were lower in the hippocampus but higher in the cortex of female mice than of male mice [131]. Thus, one can determine that sex has a more substantial influence on neural ceramide levels in mice than the APOE genotype, familial AD history, or age.
While APOE4 is mainly generated by astrocytes, it can also be generated by neurons when the brain is under unusual stress, such as aging. APOE isoforms show different effects on neural activity using different cellular sources, including astrocytes and neurons, in control and AD-like in vitro cultured neuron models [131]. While astrocyte-originated APOE4 appears to increase neuronal activity, neuron-expressed ApoE appears to induce a higher firing rate in APOE3. Moreover, APOE isoforms have different effects on neuronal activity in APP/PS1 AD transgenic compared to wild-type mice neurons [132]. These studies suggested that ApoE has different implications and effects on neuronal activity depending on whether it is produced by astrocytes or neurons: astrocytic ApoE is associated with a stronger firing rate of ApoE4, while the most active neuronal activity induced by neuronal ApoE is caused by APOE3.
APOE3-overexpression mice regulated a neural response to high-fat diet while APOE4- overexpression mice did not, supporting a model wherein early dysregulation of inflammation in APOE4 brains precipitates increased CNS damage [133]. Sortilin is a receptor that works with ApoE to stimulate the polyunsaturated fatty acids to become neuromodulators and create anti-inflammatory genes within the brain, building up cerebral protection. However, while this process occurs in APOE3 carriers, it is absent in APOE4 carriers, suggesting that impaired cerebral lipid metabolism is a characteristic of AD in APOE4 carriers [134].
ApoE is a lipid-transporting protein, known to control the inflammatory characteristics of activated microglia in various neurodegenerative diseases. Microglia are known to activate early in prion pathogenesis, but, result in microglia-driven neuroinflammation and deterioration of neural networks. There is a marked upregulation of ApoE expression in activated microglia, but reduced expression in activated astrocytes in the infection of wild-type mice with 22L prions. Ablating ApoE showed an exaggeration of prion protein (PrP)-modulated neurodegeneration [135].
ApoE-deficient (Apoe−/−) mice provide an excellent model to study atherosclerosis—the buildup of fats in artery walls—due to their similarilarities to human pathology and diet [136]. Apoe−/− mice have shorter disease incubation periods, increased spongiform lesions, increased neurodegeneration, and increased astrocytes and microgliosis. Astrocytes of Apoe−/− mice demonstrate upregulation of markers indicative of neurotoxic A1 reactive astrocytes, and microglia show upregulation of markers indicative of microglial neurodegenerative phenotype. Thus, ApoE-deficient mice result in impaired clearance of normal cellular PrP and dying neurons by microglia—the excess neural debris leads to neuroinflammation and neuronal death [135].
There is a strong association between ApoE and Aβ production in vivo within the brains of mice. ApoE regulates Aβ synthesis with lipid clusters in vitro system studies [137]. ApoE uses cholesterol molecules from astrocytes to regulate and interact with APP and synthesizes Aβ peptide molecules. When astrocyte-derived cholesterol was deleted, the impact of amyloid plaque accumulation on AD decreased. The amount of cholesterol within the neurons decreased, whereas the protective product of APP, APP-ɑ, was synthesized after being treated with cholesterol-free ApoE in neurons.
ApoE has also shown many physiological and pathophysiological functions, such as immunomodulation, oxidative stress, stabilization of neuronal microtubules, synaptic plasticity, and apoptosis. The unknown contribution of ApoE may still need to be established.
4.6. ApoC1
ApoC1 is small apolipoprotein molecule. ApoC-I mRNA and protein have been expressed in brain astrocytes, which decreases with age [138,139,140]. ApoC1 is mainly expressed in the liver, macrophage, but also expressed in SCN of brain. Polymorphisms of the human APOC1 gene are related with the altered transcription, which increased the development of AD and blood total cholesterol and triglyceride [141,142]. ApoC1 has shown play a central role in the metabolism of HDL and VLDL. One important function of apoC1 is activated when monocytes differentiate in macrophages. ApoC1 works on a lipoprotein receptors by several factors, such as: inhibiting binding mediated by ApoE, reducing lipoprotein lipase (LPL) and hepatic lipase to enhance blood triglyceride levels, decreasing phospholipase A2, downregulating cholesterylester transfer protein and activating lecithin-cholesterol transferase to control blood cholesterol levels [38]. Several studied have shown that ApoC1 linked to several pathways including plasma lipoprotein assembly, remodeling, and clearance and liver X receptors LXRα and LXRβ-related signaling [143,144,145]. More important function of ApoC1 is play important role in triglyceride-rich lipoprotein by inhibiting the binding of VLDL to VLDL-receptor, to LDL-receptor and to LDL-receptor related protein (LRP) and downregulating the activity of LPL.
Studies have shown that Global apoC1 knockout (apoC1−/−) mice showed impaired hippocampal-dependent memory and increased expression of TNFα. Mice overexpressing human APOC1 also had damages in learning and memory. These studies showed that the loss or gain of ApoC1 function both decrease learning and memory [146]. ApoC-I is known to block the ApoE receptor interaction, and in turn, the expression of APOE4 allele usually results in increased expression of ApoC1 [147]. There are several functions of ApoC1 -mediated lipid metabolism in the brain as shown in peripheral tissue and play an impartment role in CVD, however, future research is necessary to determine whether ApoC1 that showed rhythmic expressions are associated with brain lipid metabolism.
4.7. ApoH
ApoH is expressed in astrocytes and neurons as a phospholipid-binding (e.g., cardiolipin) plasma protein. ApoH is synthesized within the brain. ApoH proteins have been detected in the CNS and CSF; ApoH is like ApoA4, as they are able to enter the brain as “lipid-poor” particles, or as free apolipoproteins [83].
Brain and plasma ApoH levels have also been linked with cognitive aging, predementia syndrome, mild cognitive impairment, and AD [148]. Therefore, it is important to study ApoH to better understanding the factors influencing its levels of impact on brain disease. Plasma ApoH levels are associated with cognitive status; while APOH Single Nucleotide Polymorphisms (SNPs) are linked to diabetes type 2, chronic inflammatory disease, and age-related cognitive performance [149]. In peripheral tissues, ApoH has been related to different physiological pathways such as lipoprotein metabolism, coagulation, and antiphospholipid autoantibodies production. Genome-wide association study has shown that APOH is a novel locus for lipoprotein (a) levels [150]. From lupus and primary antiphospholipid syndrome patient samples, ApoH may be a required cofactor for anionic phospholipid binding by the antiphospholipid autoantibodies [151].
4.8. ApoJ
ApoJ, also called Clusterin, is found in the brain, synthesized by astrocytes, is found on HDL. ApoJ- lipoproteins are generally small and contain less lipids. ApoJ expression has been found in astrocytes, neurons, and the ependymal cells, lining the ventricle of the brain [148]. ApoJ has also been shown to play an important role as an essential determinant of CSF cholesterol efflux capacity, which in turn could mitigate the risk of mild cognitive impairment and AD via assistance of cellular efflux of cholesterol or other lipids [148]. ApoJ can also act as a molecular chaperone in the cellular stress response [89,152,153]. ApoJ is able to bind to Aß fibrils and is correlated with Aß plaques, neuropil threads, and cerebrovascular amyloid deposits in the AD brain [89,154,155]. ApoJ can suppress Aß aggregation and facilitate its pass through the BBB, suggesting that ApoJ may exert a neuroprotective effect. ApoJ has been identified as playing an essential role in neurodegenerative conditions, such as Lewy bodies, and prion deposits, and pathological conditions, such as AD, multiple sclerosis, TBI, SCI, gliomas, ischemia, epilepsy, chemically induced lesions, and aging [89,156,157]. ApoJ knock out (ApoJ-/-) mice displaedy significantly impaired recovery from cerebral ischemic insult, suggesting that ApoJ provides protection from these injurious states.
4.9. ApoM
ApoM is also included in HDL, LDL, triglyceride-rich lipoproteins, and VLDL, being involved in lipid transport. Studies have determined that ApoM is secreted by tubular epithelial cells and is reabsorbed in a megalin-dependent manner [158]; however, it is not found in urine under normal physiological condition.
Single nucleotide polymorphisms in the ApoM gene are related to several prevalent diseases, including type 2 diabetes mellitus [159], chronic obstructive pulmonary disease, and systemic lupus erythematosus [160,161]. Yet, the regulation and possible roles of ApoM in the brain and SCN are poorly understood. In animal studies, the blood glucose levels in ApoM-deficient mice were decreased compared with those in control wild-type mice, and ApoM was suggested to promote insulin secretion and sensitivity [159,162,163]. ApoM is essential for the formation of pre-β-HDL and reverse cholesterol transport [164]. Interestingly, ApoM levels were decreased in ApoA1- deficient mice compared with those in wild-type mice, suggesting that ApoM might be associated with the pathogenesis of several cardiometabolic diseases. Janiurek et al. showed that ApoM-bound S1P maintains low paracellular BBB permeability in all cerebral microvessels causing low levels of vesicle-mediated transport in penetrating arterioles [165].
5. Conclusions
The SCN controls apolipoprotein and lipid metabolism-related genes. 9 of apolipoprotein genes showed rhythmic expression in the brain tissue, suggesting a correlation between the circadian master clock and cerebral lipid metabolism. Past studies have proved that disrupted circadian clock is a high-risk factor for neurodegenerative diseases, specifically AD. AD is characterized by impaired lipid metabolism and cholesterol efflux capacity, eventually accumulating plaque within the brain. The rhythmic expression of lipid metabolism-related genes proves the presence of an association between AD and SCN. Thus, disruptions to the circadian clock causes impaired lipid metabolism. Additionally, ApoE and Clu showed relatively high expressions in the brain, suggesting that they are produced within the brain. ApoE showed a high correlation with AD; for ApoE4 status, a form of ApoE increases the risk for AD [166]. Therefore, future research is necessary to determine whether the genes that showed similar rhythmic expressions are associated. Identifying the other genes affected circadian rhythms through the use of conditional knockout mice is essential.
While extensively studied in peripheral tissues of the body, there needs to be more investigation done on the rhythmic expression of the mentioned clock and apolipoproteins, in the brain. Thus, this study intended to determine the expression levels of the mentioned genes in the SCN and whether they showed rhythmic expression over a 24-h cycle in the brains of mice. It was hypothesized that all the studied genes, vital to the circadian clock and lipid metabolism, are regulated by the SCN in the brains of mice and thus will show rhythmic expression, reacting to the light and dark phases throughout 24 h. We proposal that there are two pathway apolipoproteins that are present in the brain (Figure 5). The first hypothesis is that apolipoproteins are expressed and synthesized in the brain, and are directly regulated by the central circadian clock and indirectly linked with the peripheral circadian clock. The second hypothesis is that apolipoproteins are able to enter the brain through the BBB from the blood, which may be directly associated with the peripheral circadian clock and indirectly related with the central circadian clock. The two pathways are able to control the apolipoprotein-associated physiological function and pathology of diseases.
As our attention extends to more and more complicated life forms, circadian rhythm as well as brain research scientists must be convinced to include the circadian rhythm as a crucially important variable in their experimental protocols. It is becoming clear that the role of the apolipoproteins to maintain health, prevent brain disease, and treat illnesses will be increasingly affected by research that expands our understanding of the brain SCN hormone, immune factors, and circadian clock, which affect apolipoprotein’s expression. However, in spite of the different pathological effects of apolipoproteins, there remains a capacity for a therapeutic target for AD, atherosclerosis and diabetes mellitus.
To the best of our knowledge, the roles of the apolipoprotein signaling pathway in SCN as well as other location of brain related with numerous diseases and under normal physiology have not been completely elucidated. Further studies are necessary to fully investigate the mechanism of apolipoproteins in the pathogenesis of brain diseases and determine their capacity as a novel therapeutic target.
Author Contributions
conceptualization and writing first draft, X.P.; writing original draft preparation, X.P., H.L. and E.M.C.; editing, H.L. and X.P.; reading, H.N., E.M.C., A.C. and X.P.
Funding
This work was supported in part by NIH National Heart, Lung, and Blood Institute Grant R56 HL137912-01 and American Heart Association Grant-In-Aid 16GRNT30960027 to X. Pan.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AD: Alzheimer’s disease; APP: amyloid precursor protein; AVP: vasopressin; BBB: blood brain barrier; Bmal1: basic helix-loop-helix ARNT like 1 (ARNTL); cAMP: cathelicidin antimicrobial peptide; Clock: clock circadian regulator; Clu: clusterin or ApoJ; CM: chylomicron; CNS: central nervous system; Cry: cryptochrome circadian regulator; CVD: cardiovascular disease; CSF: cerebrospinal fluid; ERα: estrogen receptor-α; ERβ: estrogen receptor-β; GLP-1: glucagon-like peptide 1; HDL: high-density lipoprotein; IL1R1: Interleukin-1 Type 1 Receptor; IL-1B: Interleukin-1b; LDL: low density lipoprotein; LPS: lipopolysaccharide; mRNA: messenger RNA; NGF: nerve growth factor; Nr1d1: nuclear receptor subfamily 1 group D member 1; PD: Parkinson’s disease; Per: period circadian regulators; PHI: peptide histidine-isoleucine; PrP: prion protein; PS1: presenilin 1; S1P: sphingosine-1-phosphate; SCI: spinal cord injury; TBI: traumatic brain injury; TNF-α: tumor necrosis factor-α; VIP: vasoactive intestinal peptide; VLDL: very low-density lipoprotein
References
- Roenneberg, T.; Merrow, M. The Circadian Clock and Human Health. Curr. Biol. 2016, 26, R432–R443. [CrossRef]
- Green, C.B.; Takahashi, J.S.; Bass, J. The Meter of Metabolism. Cell 2008, 134, 728–742. [CrossRef]
- Bass, J.T. The circadian clock system’s influence in health and disease. Genome Med. 2017, 9, 1–5. [CrossRef]
- Lananna, B.V.; Musiek, E.S. The wrinkling of time: Aging, inflammation, oxidative stress, and the circadian clock in neurodegeneration. Neurobiol. Dis. 2020, 139, 104832–104832. [CrossRef]
- Prasai MJ, George JT, and Scott EM. Molecular clocks, type 2 diabetes and cardiovascular disease. Diab Vasc Dis Res. 2008;5(2):89-95. [CrossRef]
- Maury E, Ramsey KM, and Bass J. Circadian rhythms and metabolic syndrome: from experimental genetics to human disease. Circ Res. 2010;106(3):447-62. [CrossRef]
- Firsov, D.; Bonny, O. Circadian rhythms and the kidney. Nat. Rev. Nephrol. 2018, 14, 626–635. [CrossRef]
- Pan X, Mota S, and Zhang B. Circadian Clock Regulation on Lipid Metabolism and Metabolic Diseases. Adv Exp Med Biol. 2020;1276:53-66. [CrossRef]
- Nassan, M.; Videnovic, A. Circadian rhythms in neurodegenerative disorders. Nat. Rev. Neurol. 2021, 18, 7–24. [CrossRef]
- Zhang, D.; Pollock, D.M. Circadian regulation of kidney function: finding a role for Bmal1. Am. J. Physiol. Physiol. 2018, 314, F675–F678. [CrossRef]
- Levi, F.; Schibler, U. Circadian Rhythms: Mechanisms and Therapeutic Implications. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 593–628. [CrossRef]
- Gooley, J.J. Circadian regulation of lipid metabolism. Proc. Nutr. Soc. 2016, 75, 440–450. [CrossRef]
- Muenchhoff, J.; Song, F.; Poljak, A.; Crawford, J.D.; Mather, K.A.; Kochan, N.A.; Yang, Z.; Trollor, J.N.; Reppermund, S.; Maston, K.; et al. Plasma apolipoproteins and physical and cognitive health in very old individuals. Neurobiol. Aging 2017, 55, 49–60. [CrossRef]
- Pan, X.; Zhang, Y.; Wang, L.; Hussain, M.M. Diurnal Regulation of MTP and Plasma Triglyceride by CLOCK Is Mediated by SHP. Cell Metab. 2010, 12, 174–186. [CrossRef]
- Pan, X.; Jiang, X.-C.; Hussain, M.M. Impaired Cholesterol Metabolism and Enhanced Atherosclerosis in Clock Mutant Mice. Circ. 2013, 128, 1758–1769. [CrossRef]
- Pan, X.; Bradfield, C.A.; Hussain, M.M. Global and hepatocyte-specific ablation of Bmal1 induces hyperlipidaemia and enhances atherosclerosis. Nat. Commun. 2016, 7, 13011–13011. [CrossRef]
- Pan X, and Hussain MM. Bmal1 regulates production of larger lipoproteins by modulating cAMP-responsive element-binding protein H and apolipoprotein AIV. Hepatology. 2022;76(1):78-93. [CrossRef]
- Pan, X.; Munshi, M.K.; Iqbal, J.; Queiroz, J.; Sirwi, A.A.; Shah, S.; Younus, A.; Hussain, M.M. Circadian Regulation of Intestinal Lipid Absorption by Apolipoprotein AIV Involves Forkhead Transcription Factors A2 and O1 and Microsomal Triglyceride Transfer Protein. J. Biol. Chem. 2013, 288, 20464–20476. [CrossRef]
- Martins IJ, Hone E, Foster JK, Sunram-Lea SI, Gnjec A, Fuller SJ, et al. Apolipoprotein E, cholesterol metabolism, diabetes, and the convergence of risk factors for Alzheimer’s disease and cardiovascular disease. Mol Psychiatry. 2006;11(8):721-36. [CrossRef]
- Dufouil C, Richard F, Fievet N, Dartigues JF, Ritchie K, Tzourio C, et al. APOE genotype, cholesterol level, lipid-lowering treatment, and dementia: the Three-City Study. Neurology. 2005;64(9):1531-8. [CrossRef]
- Sparks DL, Sabbagh MN, Connor DJ, Lopez J, Launer LJ, Browne P, et al. Atorvastatin for the treatment of mild to moderate Alzheimer disease: preliminary results. Arch Neurol. 2005;62(5):753-7. [CrossRef]
- Reppert, S.M.; Weaver, D.R. Molecular Analysis of Mammalian Circadian Rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [CrossRef]
- Hussain, M.M.; Pan, X. Clock genes, intestinal transport and plasma lipid homeostasis. Trends Endocrinol. Metab. 2009, 20, 177–185. [CrossRef]
- Froy O. The circadian clock and metabolism. Clin Sci (Lond). 2011;120(2):65-72. [CrossRef]
- Douris, N.; Kojima, S.; Pan, X.; Lerch-Gaggl, A.F.; Duong, S.Q.; Hussain, M.M.; Green, C.B. Nocturnin Regulates Circadian Trafficking of Dietary Lipid in Intestinal Enterocytes. Curr. Biol. 2011, 21, 1347–1355. [CrossRef]
- Schnell A, Chappuis S, Schmutz I, Brai E, Ripperger JA, Schaad O, et al. The nuclear receptor REV-ERBalpha regulates Fabp7 and modulates adult hippocampal neurogenesis. PLoS One. 2014;9(6):e99883. [CrossRef]
- Landgraf, D.; Long, J.E.; Proulx, C.D.; Barandas, R.; Malinow, R.; Welsh, D.K. Genetic Disruption of Circadian Rhythms in the Suprachiasmatic Nucleus Causes Helplessness, Behavioral Despair, and Anxiety-like Behavior in Mice. Biol. Psychiatry 2016, 80, 827–835. [CrossRef]
- Wolff SEC, Wang XL, Jiao H, Sun J, Kalsbeek A, Yi CX, et al. The Effect of Rev-erbalpha Agonist SR9011 on the Immune Response and Cell Metabolism of Microglia. Front Immunol. 2020;11:550145. [CrossRef]
- McKee, C.A.; Lee, J.; Cai, Y.; Saito, T.; Saido, T.; Musiek, E.S. Astrocytes deficient in circadian clock gene Bmal1 show enhanced activation responses to amyloid-beta pathology without changing plaque burden. Sci. Rep. 2022, 12, 1796. [CrossRef]
- Yang, G.; Zhang, J.; Jiang, T.; Monslow, J.; Tang, S.Y.; Todd, L.; Puré, E.; Chen, L.; FitzGerald, G.A. Bmal1 Deletion in Myeloid Cells Attenuates Atherosclerotic Lesion Development and Restrains Abdominal Aortic Aneurysm Formation in Hyperlipidemic Mice. Arter. Thromb. Vasc. Biol. 2020, 40, 1523–1532. [CrossRef]
- Lee, J.; Kim, D.E.; Griffin, P.; Sheehan, P.W.; Kim, D.H.; Musiek, E.S.; Yoon, S.Y. Inhibition of REV-ERBs stimulates microglial amyloid-beta clearance and reduces amyloid plaque deposition in the 5XFAD mouse model of Alzheimer’s disease. Aging Cell 2019, 19, e13078. [CrossRef]
- Hofman A, Ott A, Breteler MM, Bots ML, Slooter AJ, van Harskamp F, et al. Atherosclerosis, apolipoprotein E, and prevalence of dementia and Alzheimer’s disease in the Rotterdam Study. Lancet. 1997;349(9046):151-4. [CrossRef]
- Wingo, T.S.; Cutler, D.J.; Wingo, A.P.; Le, N.-A.; Rabinovici, G.D.; Miller, B.L.; Lah, J.J.; Levey, A.I. Association of Early-Onset Alzheimer Disease With Elevated Low-Density Lipoprotein Cholesterol Levels and Rare Genetic Coding Variants of APOB. JAMA Neurol. 2019, 76, 809–817. [CrossRef]
- Marchant NL, Reed BR, Sanossian N, Madison CM, Kriger S, Dhada R, et al. The aging brain and cognition: contribution of vascular injury and abeta to mild cognitive dysfunction. JAMA Neurol. 2013;70(4):488-95. [CrossRef]
- Power, M.C.; Rawlings, A.; Sharrett, A.R.; Bandeen-Roche, K.; Coresh, J.; Ballantyne, C.M.; Pokharel, Y.; Michos, E.D.; Penman, A.; Alonso, A.; et al. Association of midlife lipids with 20-year cognitive change: A cohort study. Alzheimer’s Dement. 2017, 14, 167–177. [CrossRef]
- Blue, M.L.; Williams, D.L.; Zucker, S.; A Khan, S.; Blum, C.B. Apolipoprotein E synthesis in human kidney, adrenal gland, and liver.. Proc. Natl. Acad. Sci. 1983, 80, 283–287. [CrossRef]
- Wang H, and Eckel RH. What are lipoproteins doing in the brain? Trends Endocrinol Metab. 2014;25(1):8-14. [CrossRef]
- Fuior, E.V.; Gafencu, A.V. Apolipoprotein C1: Its Pleiotropic Effects in Lipid Metabolism and Beyond. Int. J. Mol. Sci. 2019, 20, 5939. [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164–179. [CrossRef]
- Earnest, D.J.; A Olschowka, J. Circadian regulation of c-fos expression in the suprachiasmatic pacemaker by light.. 1993, S65–71.
- Brancaccio, M.; Enoki, R.; Mazuski, C.N.; Jones, J.; Evans, J.A.; Azzi, A. Network-Mediated Encoding of Circadian Time: The Suprachiasmatic Nucleus (SCN) from Genes to Neurons to Circuits, and Back. J. Neurosci. 2014, 34, 15192–15199. [CrossRef]
- Wen, S.; Ma, D.; Zhao, M.; Xie, L.; Wu, Q.; Gou, L.; Zhu, C.; Fan, Y.; Wang, H.; Yan, J. Spatiotemporal single-cell analysis of gene expression in the mouse suprachiasmatic nucleus. Nat. Neurosci. 2020, 23, 456–467. [CrossRef]
- Barca-Mayo, O.; Pons-Espinal, M.; Follert, P.; Armirotti, A.; Berdondini, L.; Tonelli, D.D.P. Astrocyte deletion of Bmal1 alters daily locomotor activity and cognitive functions via GABA signalling. Nat. Commun. 2017, 8, 14336. [CrossRef]
- Tso, C.F.; Simon, T.; Greenlaw, A.C.; Puri, T.; Mieda, M.; Herzog, E.D. Astrocytes Regulate Daily Rhythms in the Suprachiasmatic Nucleus and Behavior. Curr. Biol. 2017, 27, 1055–1061. [CrossRef]
- Brancaccio, M.; Patton, A.P.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Astrocytes Control Circadian Timekeeping in the Suprachiasmatic Nucleus via Glutamatergic Signaling. Neuron 2017, 93, 1420–1435.e5. [CrossRef]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-autonomous clock of astrocytes drives circadian behavior in mammals. Science 2019, 363, 187–192. [CrossRef]
- Granados-Fuentes, D.; Prolo, L.M.; Abraham, U.; Herzog, E.D. The Suprachiasmatic Nucleus Entrains, But Does Not Sustain, Circadian Rhythmicity in the Olfactory Bulb. J. Neurosci. 2004, 24, 615–619. [CrossRef]
- Ralph, M.R.; Foster, R.G.; Davis, F.C.; Menaker, M. Transplanted Suprachiasmatic Nucleus Determines Circadian Period. Science 1990, 247, 975–978. [CrossRef]
- Lehman, M.; Silver, R.; Gladstone, W.; Kahn, R.; Gibson, M.; Bittman, E. Circadian rhythmicity restored by neural transplant. Immunocytochemical characterization of the graft and its integration with the host brain. J. Neurosci. 1987, 7, 1626–1638. [CrossRef]
- Harmar, A.J.; Marston, H.M.; Shen, S.; Spratt, C.; West, K.M.; Sheward, W.; Morrison, C.F.; Dorin, J.R.; Piggins, H.D.; Reubi, J.-C.; et al. The VPAC2 Receptor Is Essential for Circadian Function in the Mouse Suprachiasmatic Nuclei. Cell 2002, 109, 497–508. [CrossRef]
- Welsh, D.K.; Takahashi, J.S.; Kay, S.A. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2010, 72, 551–577. [CrossRef]
- Pittman, Q.J. Vasopressin and central control of the cardiovascular system: A 40-year retrospective. J. Neuroendocr. 2021, 33, e13011. [CrossRef]
- Smith, M.C.; Dunn, M.J. The Role of Prostaglandins in Human Hypertension. Am. J. Kidney Dis. 1985, 5, A32–A39. [CrossRef]
- Reghunandanan, V. Vasopressin in circadian function of SCN. J. Biosci. 2020, 45, 1–11. [CrossRef]
- Olszewski PK, Wirth MM, Shaw TJ, Grace MK, and Levine AS. Peptides that regulate food intake: effect of peptide histidine isoleucine on consummatory behavior in rats. Am J Physiol Regul Integr Comp Physiol. 2003;284(6):R1445-53. [CrossRef]
- Shinohara, K.; Funabashi, T.; Mitushima, D.; Kimura, F. Effects of gap junction blocker on vasopressin and vasoactive intestinal polypeptide rhythms in the rat suprachiasmatic nucleus in vitro. Neurosci. Res. 2000, 38, 43–47. [CrossRef]
- Kruijver, F.P.; Swaab, D.F. Sex Hormone Receptors Are Present in the Human Suprachiasmatic Nucleus. Neuroendocrinology 2002, 75, 296–305. [CrossRef]
- Kruijver FP, Zhou JN, Pool CW, Hofman MA, Gooren LJ, and Swaab DF. Male-to-female transsexuals have female neuron numbers in a limbic nucleus. J Clin Endocrinol Metab. 2000;85(5):2034-41. [CrossRef]
- O’Brien, E.R.; Howarth, C.; Sibson, N.R. The role of astrocytes in CNS tumors: pre-clinical models and novel imaging approaches. Front. Cell. Neurosci. 2013, 7, 40. [CrossRef]
- Brierley, D.I.; Holt, M.K.; Singh, A.; de Araujo, A.; McDougle, M.; Vergara, M.; Afaghani, M.H.; Lee, S.J.; Scott, K.; Maske, C.; et al. Central and peripheral GLP-1 systems independently suppress eating. Nat. Metab. 2021, 3, 258–273. [CrossRef]
- Rowlands, J.; Heng, J.; Newsholme, P.; Carlessi, R. Pleiotropic Effects of GLP-1 and Analogs on Cell Signaling, Metabolism, and Function. Front. Endocrinol. 2018, 9, 672. [CrossRef]
- Nauck MA, and Meier JJ. GLP-1 receptor agonists and SGLT2 inhibitors: a couple at last? Lancet Diabetes Endocrinol. 2016;4(12):963-4. [CrossRef]
- Hussein H, Zaccardi F, Khunti K, Davies MJ, Patsko E, Dhalwani NN, et al. Efficacy and tolerability of sodium-glucose co-transporter-2 inhibitors and glucagon-like peptide-1 receptor agonists: A systematic review and network meta-analysis. Diabetes Obes Metab. 2020;22(7):1035-46. [CrossRef]
- Cheng, D.; Yang, S.; Zhao, X.; Wang, G. The Role of Glucagon-Like Peptide-1 Receptor Agonists (GLP-1 RA) in Diabetes-Related Neurodegenerative Diseases. Drug Des. Dev. Ther. 2022, ume 16, 665–684. [CrossRef]
- Leone, M.J.; Marpegan, L.; Bekinschtein, T.A.; Costas, M.A.; Golombek, D.A. Suprachiasmatic astrocytes as an interface for immune-circadian signalling. J. Neurosci. Res. 2006, 84, 1521–1527. [CrossRef]
- Hainich, E.C.; Pizzio, G.A.; Golombek, D.A. Constitutive activation of the ERK-MAPK pathway in the suprachiasmatic nuclei inhibits circadian resetting. FEBS Lett. 2006, 580, 6665–6668. [CrossRef]
- Liu, X.; Quan, N. Microglia and CNS Interleukin-1: Beyond Immunological Concepts. Front. Neurol. 2018, 9, 8. [CrossRef]
- Lundkvist, G.; Hill, R.; Kristensson, K. Disruption of Circadian Rhythms in Synaptic Activity of the Suprachiasmatic Nuclei by African Trypanosomes and Cytokines. Neurobiol. Dis. 2002, 11, 20–27. [CrossRef]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [CrossRef]
- Chua, E.C.-P.; Shui, G.; Lee, I.T.-G.; Lau, P.; Tan, L.-C.; Yeo, S.-C.; Lam, B.D.; Bulchand, S.; Summers, S.A.; Puvanendran, K.; et al. Extensive diversity in circadian regulation of plasma lipids and evidence for different circadian metabolic phenotypes in humans. Proc. Natl. Acad. Sci. 2013, 110, 14468–14473. [CrossRef]
- Spišská, V.; Pačesová, D.; Míková, H.; Pohanová, P.; Telenský, P.; Novotný, J.; Bendová, Z. Prenatal exposure to lipopolysaccharide induces changes in the circadian clock in the SCN and AA-NAT activity in the pineal gland. Brain Res. 2020, 1743, 146952. [CrossRef]
- Huo, M.; Huang, Y.; Qu, D.; Zhang, H.; Wong, W.T.; Chawla, A.; Huang, Y.; Tian, X.Y. Myeloid Bmal1 deletion increases monocyte recruitment and worsens atherosclerosis. FASEB J. 2016, 31, 1097–1106. [CrossRef]
- Ma, Z.; Jiang, W.; Zhang, E.E. Orexin signaling regulates both the hippocampal clock and the circadian oscillation of Alzheimer’s disease-risk genes. Sci. Rep. 2016, 6, 36035. [CrossRef]
- Chen, Q.; Peng, X.; Huang, C.; Hu, X.; Zhang, X. Association between ARNTL (BMAL1) rs2278749 polymorphism T >C and susceptibility to Alzheimer disease in a Chinese population. Evolution 2015, 14, 18515–18522. [CrossRef]
- Shimba, S.; Ishii, N.; Ohta, Y.; Ohno, T.; Watabe, Y.; Hayashi, M.; Wada, T.; Aoyagi, T.; Tezuka, M. Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis. Proc. Natl. Acad. Sci. 2005, 102, 12071–12076. [CrossRef]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [CrossRef]
- Pan, X.; Hussain, M.M. Clock is important for food and circadian regulation of macronutrient absorption in mice. J. Lipid Res. 2009, 50, 1800–1813. [CrossRef]
- Shimba, S.; Ogawa, T.; Hitosugi, S.; Ichihashi, Y.; Nakadaira, Y.; Kobayashi, M.; Tezuka, M.; Kosuge, Y.; Ishige, K.; Ito, Y.; et al. Deficient of a Clock Gene, Brain and Muscle Arnt-Like Protein-1 (BMAL1), Induces Dyslipidemia and Ectopic Fat Formation. PLOS ONE 2011, 6, e25231. [CrossRef]
- Nikolaeva, S.; Ansermet, C.; Centeno, G.; Pradervand, S.; Bize, V.; Mordasini, D.; Henry, H.; Koesters, R.; Maillard, M.; Bonny, O.; et al. Nephron-Specific Deletion of Circadian Clock Gene Bmal1 Alters the Plasma and Renal Metabolome and Impairs Drug Disposition. J. Am. Soc. Nephrol. 2016, 27, 2997–3004. [CrossRef]
- Kress GJ, Liao F, Dimitry J, Cedeno MR, FitzGerald GA, Holtzman DM, et al. Regulation of amyloid-beta dynamics and pathology by the circadian clock. J Exp Med. 2018;215(4):1059-68.
- Vitali, C.; Wellington, C.L.; Calabresi, L. HDL and cholesterol handling in the brain. Cardiovasc. Res. 2014, 103, 405–413. [CrossRef]
- Feingold KR. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al. eds. Endotext. South Dartmouth (MA); 2000.
- Koch, S.; Donarski, N.; Goetze, K.; Kreckel, M.; Stuerenburg, H.-J.; Buhmann, C.; Beisiegel, U. Characterization of four lipoprotein classes in human cerebrospinal fluid. J. Lipid Res. 2001, 42, 1143–1151. [CrossRef]
- Borghini, I.; Barja, F.; Pometta, D.; James, R.W. Characterization of subpopulations of lipoprotein particles isolated from human cerebrospinal fluid. Biochim. et Biophys. Acta (BBA)–Lipids Lipid Metab. 1995, 1255, 192–200. [CrossRef]
- Rhea, E.M.; Banks, W.A. Interactions of Lipids, Lipoproteins, and Apolipoproteins with the Blood-Brain Barrier. Pharm. Res. 2021, 38, 1469–1475. [CrossRef]
- Button, E.B.; Robert, J.; Caffrey, T.M.; Fan, J.; Zhao, W.; Wellington, C.L. HDL from an Alzheimer’s disease perspective. Curr. Opin. Infect. Dis. 2019, 30, 224–234. [CrossRef]
- Hartmann, H.; Ho, W.Y.; Chang, J.; Ling, S. Cholesterol dyshomeostasis in amyotrophic lateral sclerosis: cause, consequence, or epiphenomenon?. FEBS J. 2021, 289, 7688–7709. [CrossRef]
- Colardo, M.; Petraroia, M.; Lerza, L.; Pensabene, D.; Martella, N.; Pallottini, V.; Segatto, M. NGF Modulates Cholesterol Metabolism and Stimulates ApoE Secretion in Glial Cells Conferring Neuroprotection against Oxidative Stress. Int. J. Mol. Sci. 2022, 23, 4842. [CrossRef]
- A Elliott, D.; Weickert, C.S.; Garner, B. Apolipoproteins in the brain: implications for neurological and psychiatric disorders. Clin. Lipidol. 2010, 5, 555–573. [CrossRef]
- Koch, M.; Aroner, S.A.; Fitzpatrick, A.L.; Longstreth, W.; Furtado, J.D.; Mukamal, K.J.; Jensen, M.K. HDL (High-Density Lipoprotein) Subspecies, Prevalent Covert Brain Infarcts, and Incident Overt Ischemic Stroke: Cardiovascular Health Study. Stroke 2021, 53, 1292–1300. [CrossRef]
- Delk, S.C.; Chattopadhyay, A.; Escola-Gil, J.C.; Fogelman, A.M.; Reddy, S.T. Apolipoprotein mimetics in cancer. Semin. Cancer Biol. 2021, 73, 158–168. [CrossRef]
- Möckel, B.; Zinke, H.; Flach, R.; Weiß, B.; Gassen, H.G.; Weiler-Güttler, H. Expression of Apolipoprotein A-I in Porcine Brain Endothelium In Vitro. J. Neurochem. 1994, 62, 788–798. [CrossRef]
- Weiler-Güttler, H.; Sommerfeldt, M.; Papandrikopoulou, A.; Mischek, U.; Bonitz, D.; Frey, A.; Grupe, M.; Scheerer, J.; Gassen, H.G. Synthesis of Apolipoprotein A-1 in Pig Brain Microvascular Endothelial Cells. J. Neurochem. 1990, 54, 444–450. [CrossRef]
- Endres K. Apolipoprotein A1, the neglected relative of Apolipoprotein E and its potential role in Alzheimer’s disease. Neural Regen Res. 2021;16(11):2141-8. [CrossRef]
- Smach, M.A.; Edziri, H.; Charfeddine, B.; Ben Othman, L.; Lammouchi, T.; Ltaief, A.; Nafati, S.; Dridi, H.; Bennamou, S.; Limem, K. Polymorphism in apoA1 Influences High-Density Lipoprotein Cholesterol Levels but Is Not a Major Risk Factor of Alzheimer’s Disease. Dement. Geriatr. Cogn. Disord. Extra 2011, 1, 249–257. [CrossRef]
- Vollbach, H.; Heun, R.; Morris, C.M.; Edwardson, J.A.; McKeith, I.G.; Jessen, F.; Schulz, A.; Maier, W.; Kölsch, H. APOA1 polymorphism influences risk for early-onset nonfamiliar AD. Ann. Neurol. 2005, 58, 436–441. [CrossRef]
- Swanson, C.R.; Li, K.; Unger, T.L.; Gallagher, M.D.; Van Deerlin, V.M.; Agarwal, P.; Leverenz, J.; Roberts, J.; Samii, A.; Gross, R.G.; et al. Lower plasma apolipoprotein A1 levels are found in Parkinson’s disease and associate with apolipoprotein A1 genotype. Mov. Disord. 2014, 30, 805–812. [CrossRef]
- Wang, F.; Kohan, A.B.; Lo, C.-M.; Liu, M.; Howles, P.; Tso, P. Apolipoprotein A-IV: a protein intimately involved in metabolism. J. Lipid Res. 2015, 56, 1403–1418. [CrossRef]
- Shen, L.; Pearson, K.J.; Xiong, Y.; Lo, C.-M.; Tso, P.; Woods, S.C.; Davidson, W.S.; Liu, M. Characterization of apolipoprotein A-IV in brain areas involved in energy homeostasis. Physiol. Behav. 2008, 95, 161–167. [CrossRef]
- Liu, M.; Doi, T.; Shen, L.; Woods, S.C.; Seeley, R.J.; Zheng, S.; Jackman, A.; Tso, P.; Weng, J.; Lou, D.; et al. Intestinal satiety protein apolipoprotein AIV is synthesized and regulated in rat hypothalamus. Am. J. Physiol. Integr. Comp. Physiol. 2001, 280, R1382–R1387. [CrossRef]
- Fagan, A.M.; Holtzman, D.M.; Munson, G.; Mathur, T.; Schneider, D.; Chang, L.K.; Getz, G.S.; Reardon, C.A.; Lukens, J.; Shah, J.A.; et al. Unique Lipoproteins Secreted by Primary Astrocytes From Wild Type, apoE (−/−), and Human apoE Transgenic Mice. J. Biol. Chem. 1999, 274, 30001–30007. [CrossRef]
- Okumura, T.; Fukagawa, K.; Tso, P.; Taylor, I.L.; Pappas, T.N. Intracisternal injection of apolipoprotein A-IV inhibits gastric secretion in pylorus-ligated conscious rats. Gastroenterology 1994, 107, 1861–1864. [CrossRef]
- Tso, P.; Chen, Q.; Fujimoto, K.; Fukagawa, K.; Sakata, T. Apolipoprotein A-IV: A Circulating Satiety Signal Produced by the Small Intestine. Obes. Res. 1995, 3, 689S–695S. [CrossRef]
- Roula D, Theiler A, Luschnig P, Sturm GJ, Tomazic PV, Marsche G, et al. Apolipoprotein A-IV acts as an endogenous anti-inflammatory protein and is reduced in treatment-naive allergic patients and allergen-challenged mice. Allergy. 2020;75(2):392-402. [CrossRef]
- Császár, A.; Kálmán, J.; Szalai, C.; Janka, Z.; Romics, L. Association of the apolipoprotein A-IV codon 360 mutation in patients with Alzheimer’s disease. Neurosci. Lett. 1997, 230, 151–154. [CrossRef]
- Cui, Y.; Huang, M.; He, Y.; Zhang, S.; Luo, Y. Genetic Ablation of Apolipoprotein A-IV Accelerates Alzheimer’s Disease Pathogenesis in a Mouse Model. Am. J. Pathol. 2011, 178, 1298–1308. [CrossRef]
- Fan, Y.; Gao, J.; Li, Y.; Chen, X.; Zhang, T.; You, W.; Xue, Y.; Shen, C. The Variants at APOA1 and APOA4 Contribute to the Susceptibility of Schizophrenia With Inhibiting mRNA Expression in Peripheral Blood Leukocytes. Front. Mol. Biosci. 2021, 8. [CrossRef]
- Deng, X.; Walker, R.G.; Morris, J.; Davidson, W.S.; Thompson, T.B. Role of Conserved Proline Residues in Human Apolipoprotein A-IV Structure and Function. J. Biol. Chem. 2015, 290, 10689–10702. [CrossRef]
- Haas, M.E.; Attie, A.D.; Biddinger, S.B. The regulation of ApoB metabolism by insulin. Trends Endocrinol. Metab. 2013, 24, 391–397. [CrossRef]
- Picard, C.; Nilsson, N.; Labonté, A.; Auld, D.; Rosa-Neto, P.; Ashton, N.J.; Zetterberg, H.; Blennow, K.; Breitner, J.C.; Villeneuve, S.; et al. Apolipoprotein B is a novel marker for early tau pathology in Alzheimer’s disease. Alzheimer’s Dement. 2021, 18, 875–887. [CrossRef]
- Dassati, S.; Waldner, A.; Schweigreiter, R. Apolipoprotein D takes center stage in the stress response of the aging and degenerative brain. Neurobiol. Aging 2014, 35, 1632–1642. [CrossRef]
- Sanchez, D.; Ganfornina, M.D. The Lipocalin Apolipoprotein D Functional Portrait: A Systematic Review. Front. Physiol. 2021, 12. [CrossRef]
- Belloir, B.; Kövari, E.; Surini-Demiri, M.; Savioz, A. Altered apolipoprotein D expression in the brain of patients with Alzheimer disease. J. Neurosci. Res. 2001, 64, 61–69. [CrossRef]
- Kalman, K.; McConathy, W.; Araoz, C.; Kasa, P.; Lacko, A.G. Apolipoprotein D in the aging brain and in Alzheimer’s dementia. Neurol. Res. 2000, 22, 330–336. [CrossRef]
- A Thomas, E.; Laws, S.M.; Sutcliffe, J.; Harper, C.; Dean, B.; McClean, C.; Masters, C.; Lautenschlager, N.; E Gandy, S.; Martins, R.N. Apolipoprotein D levels are elevated in prefrontal cortex of subjects with Alzheimer’s disease: no relation to apolipoprotein E expression or genotype. Biol. Psychiatry 2003, 54, 136–141. [CrossRef]
- Terrisse, L.; Poirier, J.; Bertrand, P.; Merched, A.; Visvikis, S.; Siest, G.; Milne, R.; Rassart, . Increased Levels of Apolipoprotein D in Cerebrospinal Fluid and Hippocampus of Alzheimer’s Patients. J. Neurochem. 1998, 71, 1643–1650. [CrossRef]
- Paasila PJ, Aramideh JA, Sutherland GT, and Graeber MB. Synapses, Microglia, and Lipids in Alzheimer’s Disease. Front Neurosci. 2021;15:778822. [CrossRef]
- Li J, Luo J, Liu L, Fu H, and Tang L. The genetic association between apolipoprotein E gene polymorphism and Parkinson disease: A meta-Analysis of 47 studies. Medicine (Baltimore). 2018;97(43):e12884. [CrossRef]
- Jung JH, Jeon S, Baik K, Lee YH, Chung SJ, Yoo HS, et al. Apolipoprotein E4, amyloid, and cognition in Alzheimer’s and Lewy body disease. Neurobiol Aging. 2021;106:45-54. [CrossRef]
- Zhao J, Lu W, Ren Y, Fu Y, Martens YA, Shue F, et al. Apolipoprotein E regulates lipid metabolism and alpha-synuclein pathology in human iPSC-derived cerebral organoids. Acta Neuropathol. 2021;142(5):807-25. [CrossRef]
- Dickson DW, Heckman MG, Murray ME, Soto AI, Walton RL, Diehl NN, et al. APOE epsilon4 is associated with severity of Lewy body pathology independent of Alzheimer pathology. Neurology. 2018;91(12):e1182-e95. [CrossRef]
- Pohlkamp T, Xian X, Wong CH, Durakoglugil MS, Werthmann GC, Saido TC, et al. NHE6 depletion corrects ApoE4-mediated synaptic impairments and reduces amyloid plaque load. Elife. 2021;10. [CrossRef]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; et al. APOE4 leads to blood–brain barrier dysfunction predicting cognitive decline. Nature 2020, 581, 71–76. [CrossRef]
- Kotredes KP, Oblak A, Pandey RS, Lin PB, Garceau D, Williams H, et al. Uncovering Disease Mechanisms in a Novel Mouse Model Expressing Humanized APOEepsilon4 and Trem2*R47H. Front Aging Neurosci. 2021;13:735524. [CrossRef]
- Yu, T.-S.; Tensaouti, Y.; Stephanz, E.P.; Chintamen, S.; Rafikian, E.E.; Yang, M.; Kernie, S.G. Astrocytic ApoE underlies maturation of hippocampal neurons and cognitive recovery after traumatic brain injury in mice. Commun. Biol. 2021, 4, 1–12. [CrossRef]
- Vervoordt, S.M.; Arnett, P.; Engeland, C.; Rabinowitz, A.R.; Hillary, F.G. Depression associated with APOE status and hippocampal volume but not cognitive decline in older adults aging with traumatic brain injury.. Neuropsychology 2021, 35, 863–875. [CrossRef]
- Main, B.S.; Villapol, S.; Sloley, S.S.; Barton, D.J.; Parsadanian, M.; Agbaegbu, C.; Stefos, K.; McCann, M.S.; Washington, P.M.; Rodriguez, O.C.; et al. Apolipoprotein E4 impairs spontaneous blood brain barrier repair following traumatic brain injury. Mol. Neurodegener. 2018, 13, 1–18. [CrossRef]
- Kundu, P.; Zimmerman, B.; Perez, R.; Whitlow, C.T.; Cline, J.M.; Olson, J.D.; Andrews, R.N.; Raber, J. Apolipoprotein E levels in the amygdala and prefrontal cortex predict relative regional brain volumes in irradiated Rhesus macaques. Sci. Rep. 2021, 11, 1–14. [CrossRef]
- Kloske, C.M.; Wilcock, D.M. The Important Interface Between Apolipoprotein E and Neuroinflammation in Alzheimer’s Disease. Front. Immunol. 2020, 11, 754. [CrossRef]
- Jackson, R.J.; Meltzer, J.C.; Nguyen, H.; Commins, C.; E Bennett, R.; Hudry, E.; Hyman, B.T. APOE4 derived from astrocytes leads to blood–brain barrier impairment. Brain 2021, 145, 3582–3593. [CrossRef]
- den Hoedt S, Crivelli SM, Leijten FPJ, Losen M, Stevens JAA, Mane-Damas M, et al. Effects of Sex, Age, and Apolipoprotein E Genotype on Brain Ceramides and Sphingosine-1-Phosphate in Alzheimer’s Disease and Control Mice. Front Aging Neurosci. 2021;13:765252. [CrossRef]
- Konings, S.C.; Torres-Garcia, L.; Martinsson, I.; Gouras, G.K. Astrocytic and Neuronal Apolipoprotein E Isoforms Differentially Affect Neuronal Excitability. Front. Neurosci. 2021, 15. [CrossRef]
- Jones, N.S.; Watson, K.Q.; Rebeck, G.W. High-fat diet increases gliosis and immediate early gene expression in APOE3 mice, but not APOE4 mice. J. Neuroinflammation 2021, 18, 1–13. [CrossRef]
- Asaro, A.; Sinha, R.; Bakun, M.; Kalnytska, O.; Carlo-Spiewok, A.-S.; Rubel, T.; Rozeboom, A.; Dadlez, M.; Kaminska, B.; Aronica, E.; et al. ApoE4 disrupts interaction of sortilin with fatty acid-binding protein 7 essential to promote lipid signaling. J. Cell Sci. 2021, 134. [CrossRef]
- Pankiewicz, J.E.; Lizińczyk, A.M.; Franco, L.A.; Diaz, J.R.; Martá-Ariza, M.; Sadowski, M.J. Absence of Apolipoprotein E is associated with exacerbation of prion pathology and promotes microglial neurodegenerative phenotype. Acta Neuropathol. Commun. 2021, 9, 1–30. [CrossRef]
- Zhang, S.H.; Reddick, R.L.; Piedrahita, J.A.; Maeda, N. Spontaneous Hypercholesterolemia and Arterial Lesions in Mice Lacking Apolipoprotein E. Science 1992, 258, 468–471. [CrossRef]
- Wang, H.; Kulas, J.A.; Wang, C.; Holtzman, D.M.; Ferris, H.A.; Hansen, S.B. Regulation of beta-amyloid production in neurons by astrocyte-derived cholesterol. Proc. Natl. Acad. Sci. 2021, 118. [CrossRef]
- Weisgraber, K.H.; Mahley, R.W.; Kowal, R.C.; Herz, J.; Goldstein, J.L.; Brown, M.S. Apolipoprotein C-I modulates the interaction of apolipoprotein E with beta-migrating very low density lipoproteins (beta-VLDL) and inhibits binding of beta-VLDL to low density lipoprotein receptor-related protein.. J. Biol. Chem. 1990, 265, 22453–22459. [CrossRef]
- Kowal, R.C.; Herz, J.; Weisgraber, K.H.; Mahley, R.W.; Brown, M.S.; Goldstein, J.L. Opposing effects of apolipoproteins E and C on lipoprotein binding to low density lipoprotein receptor-related protein. J. Biol. Chem. 1990, 265, 10771–10779. [CrossRef]
- Sehayek, E.; Eisenberg, S. Mechanisms of inhibition by apolipoprotein C of apolipoprotein E-dependent cellular metabolism of human triglyceride-rich lipoproteins through the low density lipoprotein receptor pathway.. J. Biol. Chem. 1991, 266, 18259–18267. [CrossRef]
- Petit-Turcotte, C.; Stohl, S.M.; Beffert, U.; Cohn, J.S.; Aumontb, N.; Tremblayd, M.; Deab, D.; Yangc, L.; Poirierab, J.; Shachter, N.S. Apolipoprotein C-I Expression in the Brain in Alzheimer’s Disease. Neurobiol. Dis. 2001, 8, 953–963. [CrossRef]
- Drigalenko, E.; Poduslo, S.; Elston, R. Interaction of the apolipoprotein E and CI loci in predisposing to late-onset Alzheimer’s disease. Neurology 1998, 51, 131–135. [CrossRef]
- Jong MC, Hofker MH, and Havekes LM. Role of ApoCs in lipoprotein metabolism: functional differences between ApoC1, ApoC2, and ApoC3. Arterioscler Thromb Vasc Biol. 1999;19(3):472-84. [CrossRef]
- van der Hoogt CC, Berbee JF, Espirito Santo SM, Gerritsen G, Krom YD, van der Zee A, et al. Apolipoprotein CI causes hypertriglyceridemia independent of the very-low-density lipoprotein receptor and apolipoprotein CIII in mice. Biochim Biophys Acta. 2006;1761(2):213-20. [CrossRef]
- Jong, M.C.; E Dahlmans, V.; van Gorp, P.J.; van Dijk, K.W.; Breuer, M.L.; Hofker, M.H.; Havekes, L.M. In the absence of the low density lipoprotein receptor, human apolipoprotein C1 overexpression in transgenic mice inhibits the hepatic uptake of very low density lipoproteins via a receptor-associated protein-sensitive pathway.. J. Clin. Investig. 1996, 98, 2259–2267. [CrossRef]
- Abildayeva, K.; Berbée, J.F.P.; Blokland, A.; Jansen, P.J.; Hoek, F.J.; Meijer, O.; Lütjohann, D.; Gautier, T.; Pillot, T.; De Vente, J.; et al. Human apolipoprotein C-I expression in mice impairs learning and memory functions. J. Lipid Res. 2008, 49, 856–869. [CrossRef]
- Cudaback, E.; Li, X.; Yang, Y.; Yoo, T.; Montine, K.S.; Craft, S.; Montine, T.J.; Keene, C.D. Apolipoprotein C-I is an APOE genotype-dependent suppressor of glial activation. J. Neuroinflammation 2012, 9, 192–192. [CrossRef]
- Song, F.; Poljak, A.; Crawford, J.; Kochan, N.A.; Wen, W.; Cameron, B.; Lux, O.; Brodaty, H.; Mather, K.; Smythe, G.A.; et al. Plasma Apolipoprotein Levels Are Associated with Cognitive Status and Decline in a Community Cohort of Older Individuals. PLOS ONE 2012, 7, e34078. [CrossRef]
- Mather, K.A.; Thalamuthu, A.; Oldmeadow, C.; Song, F.; Armstrong, N.J.; Poljak, A.; Holliday, E.G.; McEvoy, M.; Kwok, J.B.; Assareh, A.A.; et al. Genome-wide significant results identified for plasma apolipoprotein H levels in middle-aged and older adults. Sci. Rep. 2016, 6, 23675–23675. [CrossRef]
- Hoekstra M, Chen HY, Rong J, Dufresne L, Yao J, Guo X, et al. Genome-Wide Association Study Highlights APOH as a Novel Locus for Lipoprotein(a) Levels-Brief Report. Arterioscler Thromb Vasc Biol. 2021;41(1):458-64. [CrossRef]
- Vogrinc, Ž.; Trbojević-Čepe, M.; Coen, D.; Vitale, K.; Stavljenić-Rukavina, A. Apolipoprotein H (apoH)-dependent autoantibodies and apoH protein polymorphism in selected patients showing lupus anticoagulant activity. cclm 2005, 43, 17–21. [CrossRef]
- Gelissen IC, Hochgrebe T, Wilson MR, Easterbrook-Smith SB, Jessup W, Dean RT, et al. Apolipoprotein J (clusterin) induces cholesterol export from macrophage-foam cells: a potential anti-atherogenic function? Biochem J. 1998;331 (Pt 1)(Pt 1):231-7. [CrossRef]
- Wyatt, A.R.; Yerbury, J.J.; Wilson, M.R. Structural Characterization of Clusterin-Chaperone Client Protein Complexes. J. Biol. Chem. 2009, 284, 21920–21927. [CrossRef]
- Giannakopoulos, P.; Kövari, E.; French, L.E.; Viard, I.; Hof, P.R.; Bouras, C. Possible neuroprotective role of clusterin in Alzheimer’s disease: a quantitative immunocytochemical study. Acta Neuropathol. 1998, 95, 387–394. [CrossRef]
- DeMattos RB, Cirrito JR, Parsadanian M, May PC, O’Dell MA, Taylor JW, et al. ApoE and clusterin cooperatively suppress Abeta levels and deposition: evidence that ApoE regulates extracellular Abeta metabolism in vivo. Neuron. 2004;41(2):193-202. [CrossRef]
- Freixes, M.; Puig, B.; Blanco, R.; Ferrer, I. Clusterin solubility and aggregation in Creutzfeldt-Jakob disease. Acta Neuropathol. 2004, 108, 295–301. [CrossRef]
- Sasaki, K.; Doh-Ura, K.; Ironside, J.W.; Iwaki, T. Increased clusterin (apolipoprotein J) expression in human and mouse brains infected with transmissible spongiform encephalopathies. Acta Neuropathol. 2001, 103, 199–208. [CrossRef]
- Pan, X. The Roles of Fatty Acids and Apolipoproteins in the Kidneys. Metabolites 2022, 12, 462. [CrossRef]
- Liu, D.; Pan, J.-M.; Pei, X.; Li, J.-S. Interaction Between Apolipoprotein M Gene Single-Nucleotide Polymorphisms and Obesity and its Effect on Type 2 Diabetes Mellitus Susceptibility. Sci. Rep. 2020, 10, 1–6. [CrossRef]
- Katz, P.; Pedro, S.; Trupin, L.; Yelin, E.; Michaud, K. The Impact of Asthma and Chronic Obstructive Pulmonary Disease (COPD) on Patient-Reported Outcomes in Systemic Lupus Erythematosus (SLE). ACR Open Rheumatol. 2021, 3, 221–230. [CrossRef]
- Du, W.; Shen, T.; Li, H.; Liu, Y.; He, L.; Tan, L.; Hu, M.; Ren, Y. Low apolipoprotein M serum levels correlate with Systemic lupus erythematosus disease activity and apolipoprotein M gene polymorphisms with Lupus. Lipids Heal. Dis. 2017, 16, 1–6. [CrossRef]
- Liu, M.; Frej, C.; Langefeld, C.D.; Divers, J.; Bowden, D.W.; Carr, J.J.; Gebre, A.K.; Xu, J.; Larsson, B.; Dahlbäck, B.; et al. Plasma apoM and S1P levels are inversely associated with mortality in African Americans with type 2 diabetes mellitus. J. Lipid Res. 2019, 60, 1425–1431. [CrossRef]
- Christoffersen C. Apolipoprotein M-A Marker or an Active Player in Type II Diabetes? Front Endocrinol (Lausanne). 2021;12:665393. [CrossRef]
- Christoffersen, C.; Jauhiainen, M.; Moser, M.; Porse, B.; Ehnholm, C.; Boesl, M.; Dahlbäck, B.; Nielsen, L.B. Effect of Apolipoprotein M on High Density Lipoprotein Metabolism and Atherosclerosis in Low Density Lipoprotein Receptor Knock-out Mice. J. Biol. Chem. 2008, 283, 1839–1847. [CrossRef]
- Mathiesen Janiurek M, Soylu-Kucharz R, Christoffersen C, Kucharz K, and Lauritzen M. Apolipoprotein M-bound sphingosine-1-phosphate regulates blood-brain barrier paracellular permeability and transcytosis. Elife. 2019;8. [CrossRef]
- Raulin, A.-C.; Doss, S.V.; Trottier, Z.A.; Ikezu, T.C.; Bu, G.; Liu, C.-C. ApoE in Alzheimer’s disease: pathophysiology and therapeutic strategies. Mol. Neurodegener. 2022, 17, 1–26. [CrossRef]
Figure 1.
Circadian Clock Pathway Genes Expression Levels in the SCN; data retrieved from SCNseq. Gene information, including the expression levels of each mentioned gene, was found through the SCNseq database. The expression levels in the SCN were then organized, and the gene that is highly or lowly expressed was then analyzed by GraphPad Prism 9.
Figure 1.
Circadian Clock Pathway Genes Expression Levels in the SCN; data retrieved from SCNseq. Gene information, including the expression levels of each mentioned gene, was found through the SCNseq database. The expression levels in the SCN were then organized, and the gene that is highly or lowly expressed was then analyzed by GraphPad Prism 9.
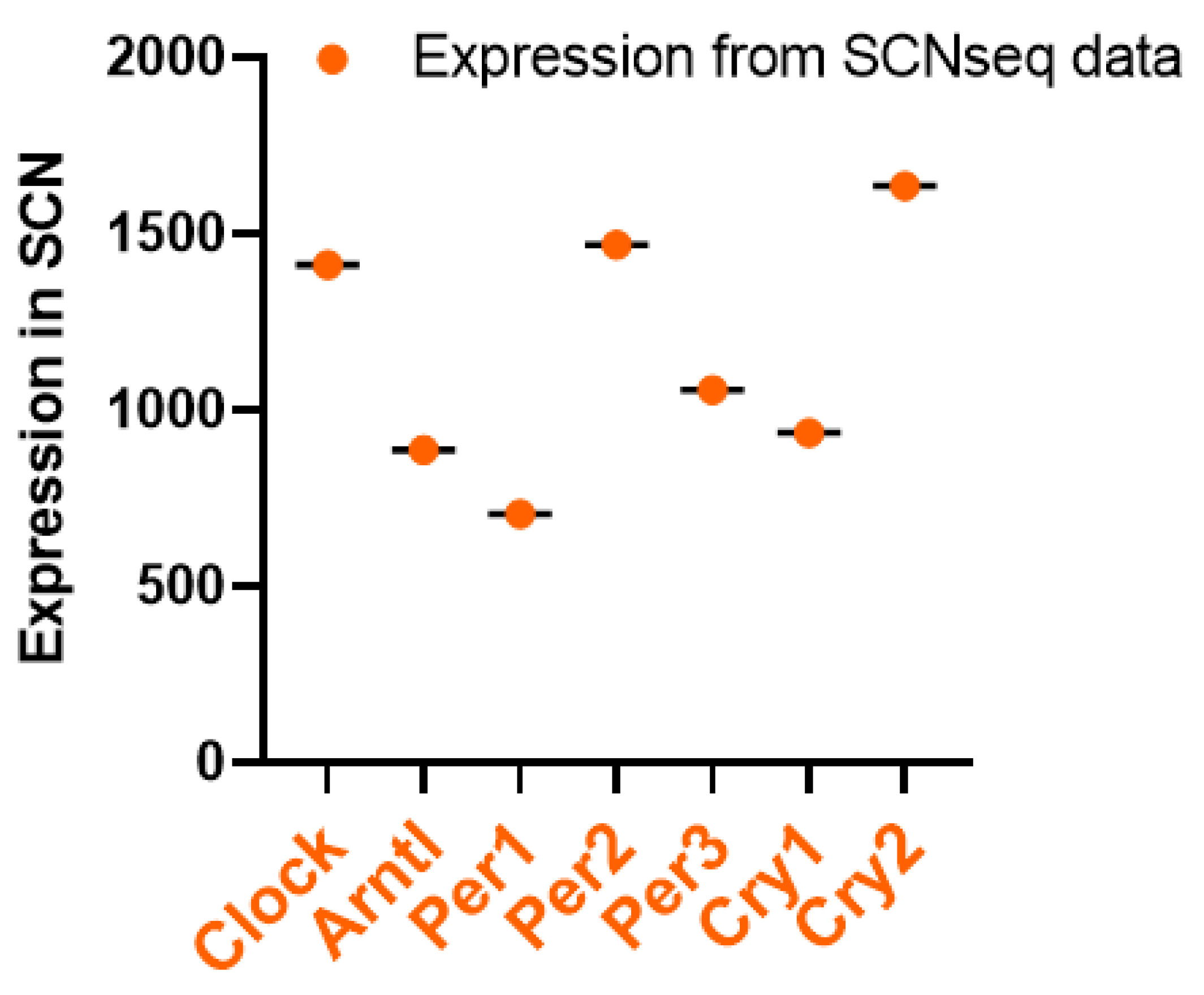
Figure 2.
Mouse gene expression levels in the brain over two 24-h periods; data retrieved from CircaDB (a) Arntl (Bmal1) expression in the brain stem; Mouse 1. OST Brain Stem (Affymetrix) (b) Per1 expression in the SCN; Mouse SCN MAS4 Panda 2002 (Affymetrix) (c) Per2 expression in the SCN; Mouse 1. OST SCN 2014 (Affymetrix) (d) Per3 expression in the brain stem; Mouse 1. OST (Affymetrix) (e) Cry2 expression in the SCN; Mouse 1. OST SCN 2014 (Affymetrix). Through the same database Figure 1′s gene information, we then obtained whether the gene was rhythmically expressed throughout 24 h. The obtained graphs were divided into two sections: light and dark, with light representing the daytime and dark representing the nighttime. The graphs were compared to see when the genes were most highly expressed throughout the day. Similar patterns between the circadian rhythms were identified. Then, past studies done on the brain tissue of Mus musculus were collected through the Circadian Expression Profiles Database (CircaDB), were compared with the graphs from SCNseq, and checked for consistency. The graphs from CircaDB were also show the available graphs of the circadian rhythmic expression of the genes. Similar to the graphs from SCNseq, the light and dark phases were indicated on the CircaDB graphs. High and low peaks were identified. All collected data were obtained from the Affymetrix assay, allowing analysis of gene expression levels.
Figure 2.
Mouse gene expression levels in the brain over two 24-h periods; data retrieved from CircaDB (a) Arntl (Bmal1) expression in the brain stem; Mouse 1. OST Brain Stem (Affymetrix) (b) Per1 expression in the SCN; Mouse SCN MAS4 Panda 2002 (Affymetrix) (c) Per2 expression in the SCN; Mouse 1. OST SCN 2014 (Affymetrix) (d) Per3 expression in the brain stem; Mouse 1. OST (Affymetrix) (e) Cry2 expression in the SCN; Mouse 1. OST SCN 2014 (Affymetrix). Through the same database Figure 1′s gene information, we then obtained whether the gene was rhythmically expressed throughout 24 h. The obtained graphs were divided into two sections: light and dark, with light representing the daytime and dark representing the nighttime. The graphs were compared to see when the genes were most highly expressed throughout the day. Similar patterns between the circadian rhythms were identified. Then, past studies done on the brain tissue of Mus musculus were collected through the Circadian Expression Profiles Database (CircaDB), were compared with the graphs from SCNseq, and checked for consistency. The graphs from CircaDB were also show the available graphs of the circadian rhythmic expression of the genes. Similar to the graphs from SCNseq, the light and dark phases were indicated on the CircaDB graphs. High and low peaks were identified. All collected data were obtained from the Affymetrix assay, allowing analysis of gene expression levels.
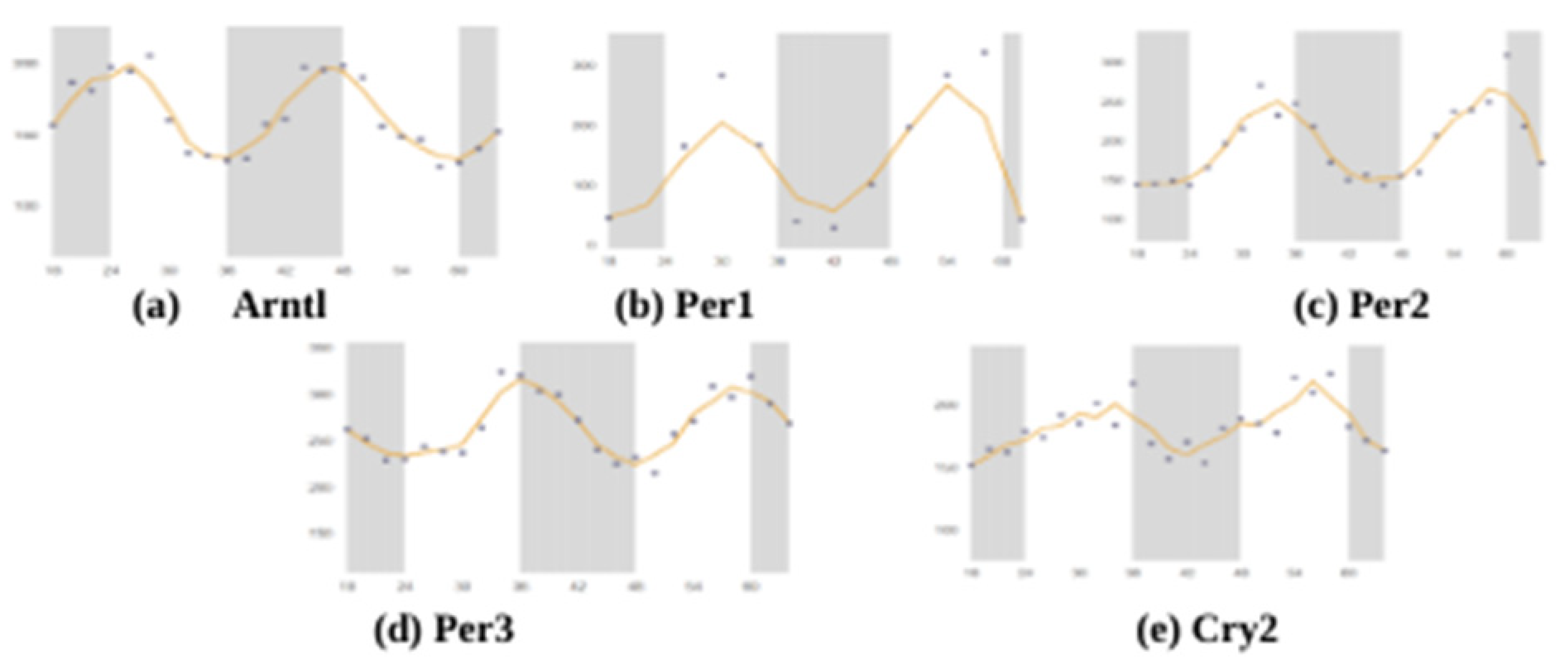
Figure 3.
Apolipoprotein Genes Expression Levels in the SCN; data retrieved from SCN seq, as descriptive method in Figure 1.
Figure 3.
Apolipoprotein Genes Expression Levels in the SCN; data retrieved from SCN seq, as descriptive method in Figure 1.
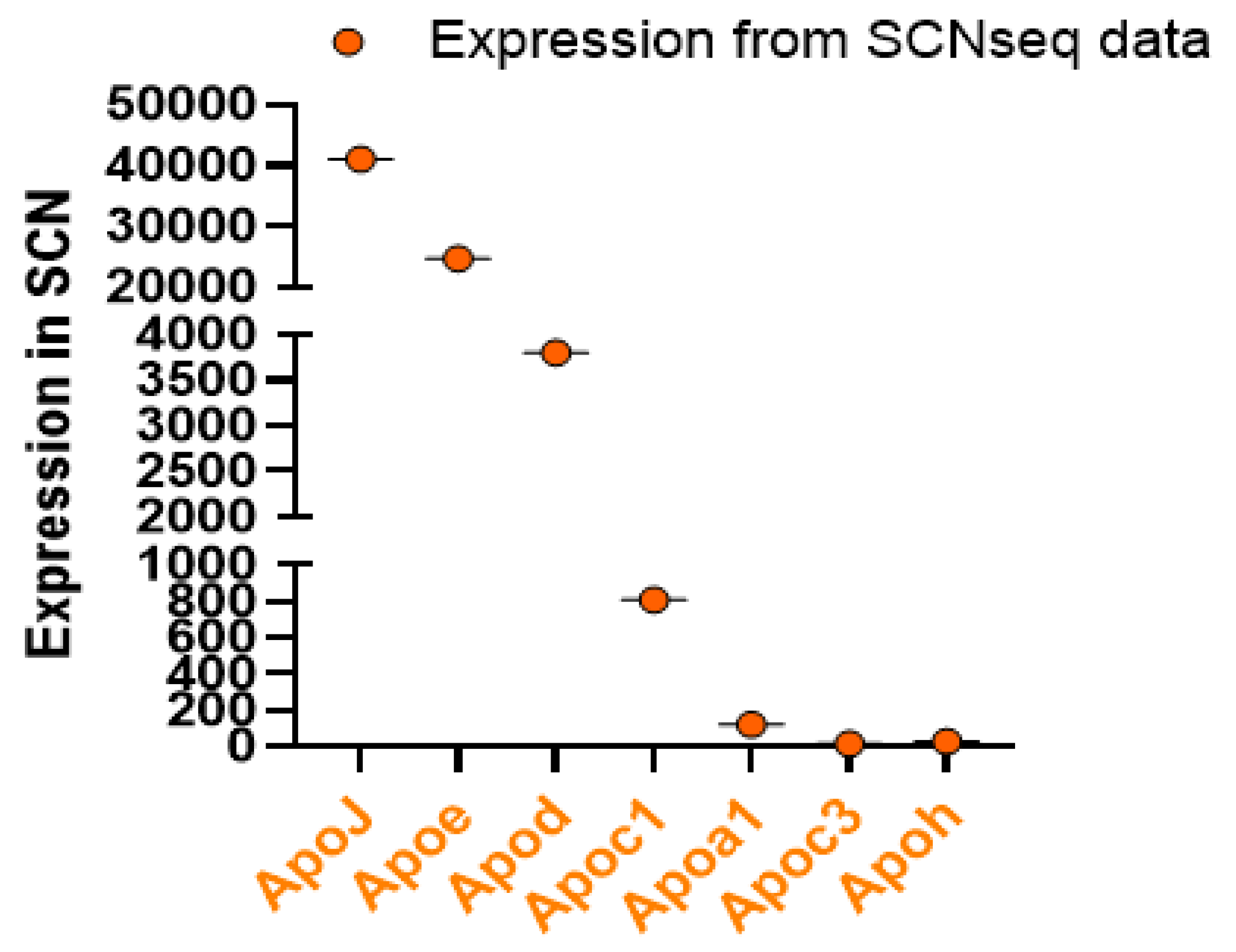
Figure 4.
Mouse gene expression levels over 24 h in the brain; data received from SCNseq (a) Apoa1 expression levels (b) Apoc1 expression levels (c) Apoe expression levels (d) Apoh expression levels (e) Apom expression levels (f) Apoj/Clu expression levels as the descriptive method in Figure 2.
Figure 4.
Mouse gene expression levels over 24 h in the brain; data received from SCNseq (a) Apoa1 expression levels (b) Apoc1 expression levels (c) Apoe expression levels (d) Apoh expression levels (e) Apom expression levels (f) Apoj/Clu expression levels as the descriptive method in Figure 2.

Figure 5.
There are two pathway apolipoproteins that are present in the brain. One hypothesis is that apolipoproteins are expressed and synthesized in the brain, being directly regulated by the central circadian clock and indirectly linked with the peripheral circadian clock; and the second hypothesis is that apolipoproteins are able to enter the brain through BBB from blood, which may be directly associated with the peripheral circadian clock and indirectly related with the central circadian clock. The two pathways are able to control the physiological function and pathology of diseases.
Figure 5.
There are two pathway apolipoproteins that are present in the brain. One hypothesis is that apolipoproteins are expressed and synthesized in the brain, being directly regulated by the central circadian clock and indirectly linked with the peripheral circadian clock; and the second hypothesis is that apolipoproteins are able to enter the brain through BBB from blood, which may be directly associated with the peripheral circadian clock and indirectly related with the central circadian clock. The two pathways are able to control the physiological function and pathology of diseases.
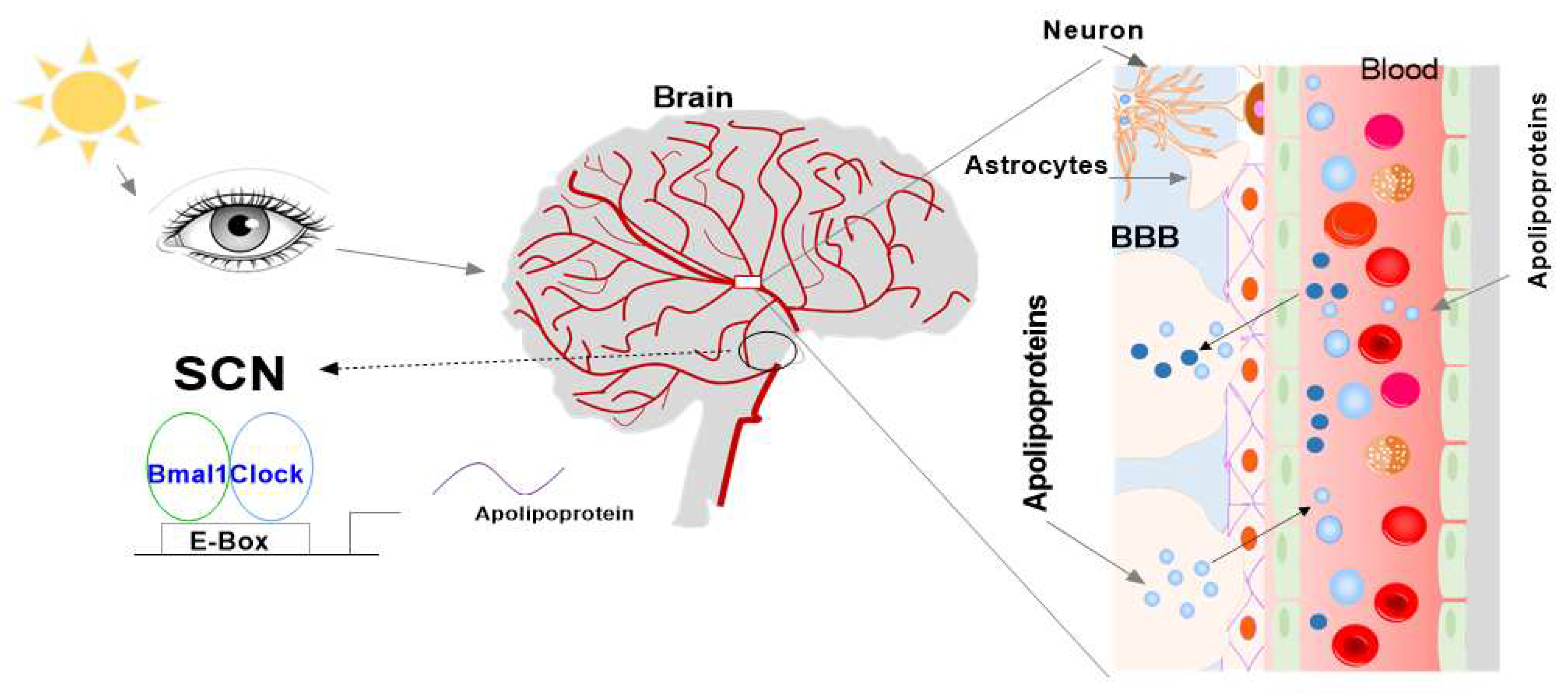
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



