
Submitted:
06 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
Antimicrobial resistance (AMR) has become a worldwide health problem. Consequently, there is a global interest in the study of the antimicrobial activity of natural compounds as an alternative to antibiotics. One of the possibilities studied during the last years is the use of bacteriophages and their derived proteins. More specifically, phage lytic proteins or endolysins are specific enzymes that cause cell lysis and can be easily produced and purified after overexpression in bacteria. However, one of the major disadvantages of these proteins is their sensitivity to some environmental conditions that can reduce their efficiency. The use of vesicles for endolysin encapsulation could help overcome this disadvantage, offering an additional protection to the protein and allowing its controlled release and long-term stability.
Keywords:
Antimicrobial activity
; vesicles
; endolysin
; encapsulation
; gelatine films
1. Introduction
The widespread use of antimicrobial agents boosts the ability of bacteria to acquire resistance mechanisms, and has led to a rapid increase in the overall antibiotic resistance of microbial populations. In turn, a lower efficacy of antibiotics is resulting in substantial morbidity and mortality, representing a public health threat with significant social and economic costs. In this context, the World Health Organization (WHO) has published the list of bacteria for which new antibiotics are urgently needed, which include the pathogen Staphylococcus aureus (methicillin-resistant MRSA, vancomycin-intermediate and resistant strains) [1]. In fact, MRSA percentages equal to or above 25% were found in 10 (25%) out of 40 countries/areas in 2020 [2] .
S. aureus is a Gram-positive bacterium that possesses multiple virulence factors, including toxins, immune system evasion factors, and the ability to form biofilms [3] This arsenal makes S. aureus one of the most important human pathogens, causing a variety of diseases, especially in the hospital environment. Additionally, this species is a major causative agent of food-borne diseases in humans due to the production of heat-resistant enterotoxins. Also, the presence of MRSA in farm animals is a serious concern, since animals can acquire and disseminate antibiotic-resistant strains.
Biofilm formation is a complex process involving the initial adhesion of bacterial cells to a biotic or inert surface, followed by the production of a self-produced extracellular matrix, mostly containing a combination of exopolysaccharides, proteins and DNA. Biofilm cells are known for their greater ability to resist antibiotics and disinfectants compared to planktonic cells. Therefore, prevention and destruction of biofilms is a challenging task, making it necessary to develop new strategies [4].
Bacteriophages and phage-derived lytic proteins have been proposed as an alternative or complementary strategy to antibiotics and disinfectants to fight against antibiotic resistant bacteria and biofilms in both clinical settings and the food industry. Typically, phages degrade the structural peptidoglycan present in the bacterial cell wall using two classes of lytic proteins: virion-associated peptidoglycan hydrolases (VAPGHs), which degrade peptidoglycan in the initial steps of the infection, and endolysins, which help release the phage progeny during the late phase of the lytic cycle. The modular structure of lytic proteins facilitates the design of new chimeric proteins via domain shuffling, which frequently leads to new proteins displaying improved lytic activity. These enzymes (both endolysins and their derived chimeric proteins) can be used as antibacterial agents by targeting bacteria from the outside, accessing the peptidoglycan, and destroying the cell walls, ultimately leading to cell lysis. Lytic proteins are relatively easy to produce, can be target-specific, and do not easily select for resistant mutants [5]. However, in order to successfully use lytic proteins, it is essential to ensure their stability.
Encapsulation in vesicles is a commonly used method for drug delivery. Vesicles are good carriers formed by an aqueous core surrounded by a lipid layer in which it is possible to encapsulate both hydrophilic and hydrophobic drugs, enhancing their pharmacodynamic properties and reducing some potential side effects [6]. Other advantages are their high biocompatibility, physical and chemical stability, good affinity towards drugs, and easy route of administration. Moreover, in terms of increasing the activity of phage lytic proteins, liposome-mediated endolysin encapsulation systems allow these proteins to penetrate the outer membrane of Gram-negative bacteria and to reduce the viable cell numbers without with the need for a membrane permeabilizer [7].
Previous studies about staphylococcal phage lytic proteins have shown their antimicrobial and antibiofilm activity [5]. Moreover, some phage lytic proteins such as LysH5 and CHAPSH3b are effective for both biofilm removal and inhibition of biofilm formation [8]. The chimeric protein CHAPSH3b is derived from PG hydrolase HydH5 (encoded by the S. aureus phage vB_SauS-phiIPLA88) and was obtained by fusion of the HydH5 CHAP domain and the SH3b cell wall binding domain from lysostaphin [9]. This protein shows antistaphylococcal activity in growth medium and milk. Studies about the stability of this protein and the influence of temperature and pH on its lytic activity showed that CHAPSH3b maintains its activity intact at 40°C and remains active even at higher temperatures. It can also withstand pH values that range from 3 to 11. These two parameters are fundamental for application of this protein as an antimicrobial and, therefore, one goal is to improve these stability values for specific applications. [9,10]. An interesting approach to successfully use endolysins in the food industry consists in embedding them into food packaging films. Active food packaging systems are designed to address the challenge of pathogens that exhibit resistance to traditional food processing methods [11] . In this regard, the inclusion of endolysins within the packaging matrix can offer an innovative approach to enhance food safety as an antimicrobial packaging system. This strategy can provide an additional layer of protection against spoilage and pathogenic microorganisms, thereby improving the safety and quality of packaged food products.
In the present work, the lytic protein CHAPSH3b was encapsulated into positively-charged vesicles (niosomes). The prepared vesicles were characterized in terms of size, zeta potential, encapsulation efficiency (EE) and antibiofilm activity. Moreover, we incorporated the vesicles into gelatine films and characterized the antimicrobial properties of the resulting material. Both free vesicles and gelatine films are intended to stabilize the antimicrobial activity of CHAPSH3b keeping in mind its future application in clinical and food settings.
2. Results
2.1. Protein Encapsulation and Characterization of the Prepared Vesicles
Previous studies had demonstrated the promising antistaphylococcal activity of the lytic protein CHAPSH3b as well as its ability as an antibiofilm agent (Fernández et al.,). In this study, we aimed to explore whether these properties could be improved through increasing protein stability by nanoencapsulation. The lytic protein CHAPSH3b was encapsulated in vesicles that contained the non-ionic surfactant Span 60 as a main compound, cholesterol as a stabilizer and CTAB as positively charged co-stabilizer. The most appropriate molar ratio between the vesicle components was 2:1:2 (Span 60 (Sp60): Cholesterol (Cho): CTAB), with a total membrane compounds concentration in the organic phase of 15 g/L. The protein used for encapsulation was purified at 1.6-2.9 mg/ml and then diluted at 8 µM in pure MQ water and PBS, used as aqueous phases. This value was chosen based on previous results showing that this was an effective dose for antibiofilm activity [12]. In parallel, empty vesicles were obtained and used as negative control in all assays.
The size and surface charge of the resulting vesicles, with and without protein, are summarized in Table 1. The particle size distribution for both aqueous phases tested is presented in Figure 1. As can be observed in Figure 1 and Table 1, nanovesicles in ultrapure water without protein have a size of 100 nm and a highly positive surface charge, 55.2 mV, due the presence of the cationic surfactant CTAB. In contrast, encapsulation of CHAPSH3b resulted in a reduced vesicle size, approximately half of that observed for the empty vesicles, and a narrower particle size distribution. However, zeta potential was similar to that of empty vesicles (46±5 mV). Interestingly, we observed that the average vesicle size doubled when using PBS buffer as aqueous hydration medium instead of water. Nevertheless, the size of vesicles prepared in PBS decreased after loading the protein, following the same pattern found in water. In general, the presence of the protein did not affect the positive charge of the vesicles, with just a slight reduction observed when protein encapsulation was carried out in MQ water, probably by the partially interaction of the protein with the CTAB molecules located at the vesicle membrane surface (Figure 1 and Table 1).
Moreover, the morphology of empty and protein-loaded vesicles was observed by TEM microscopy (Figure 2). Images show that vesicles exhibit a spherical shape with a mean size around 50-200 nm, confirming the data measured by DLS.
2.2. Efficiency of Protein Encapsulation (EE) Determined by Size Exclusion Chromatography
The concentration of encapsulated protein was determined by size exclusion chromatography, using a wavelength of 280 nm (commonly used for protein detection) to quantify absorbance. Prior to assessing the EE, CHAPSH3b protein solutions in ultrapure water and PBS were tested by size exclusion chromatography in concentrations ranging from 2 to 12 µmol (Figure 3). The void volume of the column resulted in an exclusion time of 14.5 min (measured with Blue Dextran 2000) while the smallest molecule used for column calibration (acetone) had an elution time of 54 min. The protein solution contained a significant amount of imidazole (MW 68 g/mol) which appeared at 55 min. In this medium, and despite the ionic strength used in the mobile phase, there are solute-agarose interactions that result in delayed elution times with respect to the proteins and peptides usedfor column calibration (BSA and its dimer, carbonic anhydrase, alfa albumin, citochrome C and aprotinin). To confirm the correct allocation of the protein, given the numerous peaks found, fractions of the eluate were collected at times between 26 and 55 min and further analysed by UHPLC-ESI-Q/TOF MS. The protein was identified in the peak found at an elution time of 44.6 min, with a possible aggregate eluting at 42.5 min. The rest of the UV-absorbing matter (other proteins, remains of the culture and small molecules) present in the protein solution also showed a different behaviour depending on the medium. The ionic strength of PBS appears to aid protein stabilization in solution, which results in larger absorbance values (please notice that the scale is 4x in the case of PBS). To some extent, PBS also shields the protein from aggregation, as there were fewer peaks visible at earlier elution times. The number of peaks found after the exclusion time were also significantly larger in ultrapure water, indicating that more molecules are interacting with the column matrix, thereby delaying their elution.
The presence of CHAPSH3b and its relative concentration in the vesicles was also analysed by size exclusion chromatography, following rupture of the vesicles with a mixture of ethanol and water (25:75). Solvent and empty vesicles were also tested for control purposes. Since some of the vesicle components also absorb UV light, a wider range of wavelengths were tested, from 224 to 280 nm. Protein absorbance showed a maximum peak at 254 nm, while the peak corresponding to imidazole became the predominant one at lower wavelengths (Figure 4). At 280 and 254 nm, the protein was easily identified and quantified in the presence of vesicles (and solvent) (Figure 4). Indeed, analysis of the empty vesicles and solvent revealed that no overlapping peak was detected at the protein elution time (44.6 min). In contrast, the protein solution exhibited a large peak at the aforementioned elution time. In order to analyse the effect of the proteins and vesicles together, mixtures of empty vesicles and protein solutions were prepared (1/3 and 2/3 of protein solution in 2/3 and 1/3 of empty vesicles suspension, respectively). As can be seen in Figure 5, the peaks corresponding to the protein can be easily identified in the prepared mixtures.
To assess the efficiency of protein encapsulation (EE), the loaded vesicles were separated from the supernatant and broken by mixing them with ethanol. Aliquots from the supernatant of the broken vesicles containing the released protein and the overall mixture (without supernatant separation) were analysed by HPSEC (Figure 5). The encapsulation efficiency was estimated from the relative absorbance of the peak observed between 44 and 45 min, which is marked with a green arrow in Figure 5. While it seems that ultrapure water yields more encapsulated protein according to the chromatography peak dimensions (approximately 62 % EE in water and 51% in PBS, respectively), PBS resulted in better protein stability, as previously indicated.
2.3. Antimicrobial Activity of CHAPSH3b-Loaded Vesicles
Based on the above results, we decided to test the antimicrobial activity of vesicles formed using PBS as a solvent. S. aureus 15981 was chosen as a target strain for these tests because it forms strong biofilms, rich in polysaccharides [13]. The antimicrobial activity of the protein-loaded vesicles was tested against 24-hour biofilms formed by this strain. As can be seen in Table 2, a significant reduction in biofilm cells compared with control biofilms (3.74 log units versus 8.42 log units) was observed after the first hour of treatment when using the total suspension of CHAPSH3b-containing vesicles. The highest reduction in viable cells occurred after 4 h of incubation, when the number of viable cells was below the detection level. Empty vesicles also displayed antimicrobial activity and curtailed growth thanks to the presence of CTAB, an ammonium salt with known antibacterial properties. However, in this treatment bacterial removal was always below that obtained with protein-loaded vesicles. It is worth mentioning that biofilms treated with free protein also showed a reduction in viable cells but this reduction was always lower than that obtained using vesicles. The reduction continues to be clear after 6 h of treatment; however, a regrowth of the bacterial population was observed after 24 h of incubation in both treatments, probably indicating the loss of antimicrobial activity of the protein.
2.4. Characterization of Gelatine Films Containing Encapsulated or Free CHAPSH3b
With the aim of enhancing the possibilities of encapsulated CHAPSH3b in the food industry, we prepared films based on pork skin gelatine containing free protein, empty vesicles or protein-loaded vesicles. Then, these films were morphologically characterized in order to observe the physical interactions between protein and vesicles with the gelatine matrix itself. Samples of the different films prepared were observed by SEM and pictures were collected to compare their morphology. No significant differences were observed between the gelatine films prepared with PBS alone (control films) (Figure 6A) and the ones containing empty vesicles resuspended in PBS (Figure 6B). The films appeared to be homogeneous at a magnification of x150. It must be noted that films containing free protein (Figure 6C) or encapsulated protein (Figure 6D) exhibited dots that may indicate the presence of protein agglomerates inside the gelatine matrix when observed at a magnification of x50 000.
2.5. Antimicrobial Activity of CHAPSH3b-Containing Gelatine Films
The antimicrobial activity of protein-loaded films was tested using a suspension of S. aureus Sa9 cells in TSB medium. Gelatine films containing empty vesicles and free protein were also tested. In all cases the protein concentration added to the gelatine was 8 μM and the mixtures were incubated at 37°C for 16 h, resulting in a final concentration of 2 μM in the incubation medium during the assay. The results demonstrated that CHAPSH3b was successfully incorporated into the gelatine films, since its antimicrobial activity was still noticeable after the film preparation process. However, as shown in Table 3, it is important to point out the large antimicrobial activity displayed by the empty vesicles, which results in inhibition of bacterial growth below the detection level, in comparison with the free protein, which reduced but did not completely prevent bacterial growth (Table 3).
After preparation, some films were stored for 14 days and then used to treat a cell suspension for 4 hours in order to test their long-term stability. It can be noticed that the film-embedded free protein displayed lower antibacterial activity than films containing encapsulated protein or empty vesicles, both of which led to a complete inhibition of bacterial growth (Table 3).
3. Discussion
Encapsulation of phage lytic proteins could be a suitable strategy to increase their stability and enhance their antimicrobial potential. Indeed, several recent studies have shown the advantages of encapsulating endolysins in either vesicles [7]or solid nanoparticles [14]. In recent years, some studies have even shown the effectiveness of this methodology in therapeutic applications [15,16] However, there are no studies regarding the use of encapsulated endolysins in the food industry.
In this study, the antistaphylococcal lytic protein CHAPSH3b was successfully encapsulated in non-ionic vesicles by adding Span60, cholesterol and CTAB. Vesicles were obtained at 45°C, temperature at which the protein is not denatured and does not lose its activity [17]. The presence of the cationic surfactant, CTAB, in the vesicle membrane layer results in positively-charged vesicles, which is less noticeable in PBS as the salt shields the CTAB positive charge [18], although it is still high enough to ensure colloidal stability [19]. The presence of CTAB was selected to arise a positive charged vesicles which is well known that enhance antibacterial activity [20]. The concentration of membrane compounds was chosen based on the final nanovesicle size. The estimated charge of the vesicle surface (25-55 mV) is similar to that found in other studies involving encapsulation with cationic surfactants [21].
Regarding size, the protein-loaded vesicles exhibit a two-fold decrease (from 100-200 nm to 40-80 nm) when compared to the free vesicles. This trend has also been found in other works, in which the presence of an encapsulated biocompound exerted a high influence on the final vesicle size [21]. In contrast to the results observed here, other biocompounds generally increase vesicle size [22].This behaviour may indicate the tendency of lytic proteins to be located at the vesicle membrane, probably acting as costabilizers, and reducing the critical packing parameter of the self-assembly surfactant, similar to the behaviour exhibited by high hydrophylic-lipophylic balance surfactants when they are added to the vesicle membrane. [23]. In other works, the molecular weight of the encapsulated biocompound was found to influence vesicle size, observing larger final vesicle size when encapsulating high molecular weight bicompounds [24]. Other studies demonstrate that the presence of certain additives, such as some costabilizers, leads to the formation of much larger vesicles. This is the case, for instance, of glycerol and polyethylene glycol, which have been used in several works in order to increase the EE [23,25,26]. A similar effect on the final vesicle size has been observed when lipophilic costabilizers such as cholesterol are used [27].
Analysis of protein encapsulation by size exclusion chromatography shows that the use of PBS as aqueous hydration medium during vesicle formulation results in better protein stability, as revealed by the presence of a more defined main peak around 45 min in those samples (Figure 5).
CHAPSH3b has already been shown to be an effective antibiofilm agent [8,12]. Here, we demonstrate that encapsulation of this protein leads to even better results, although it must be noted that part of the antimicrobial activity observed is due to the vesicles themselves. The cationic surfactant Cetyltrimethylammonium Bromide (CTAB) is an ammonium salt with antimicrobial activity that provokes cell lysis [28]. Moreover, the use of CTAB confers the prepared nanovesicles with a positive charge, which is known to have beneficial effects on biofilm penetration against bacterial resistance [29]. Nonetheless, the highest antimicrobial efficacy was observed when testing endolysin-loaded vesicles, which resulted in no detectable bacterial growth after 4 and 6 hours of treatment. Previous studies had already explored endolysin encapsulation. For instance, the endolysin LysRODI, encoded by phage vB_SauM_phiIPLA-RODI was encapsulated in pH-sensitive liposomes (Encapsulation of the Antistaphylococcal Endolysin LysRODI in pH-Sensitive Liposomes [30], and then tested against S. aureus biofilms. The results revealed a significant reduction in viable cells after 24 h of treatment when treated with the liposome encapsulated protein, showing that the encapsulation of these antimicrobials opens new possibilities for their delivery.
An important aspect to consider is that biofilms have microchannels that allow the entry of water and the contact with the external environment. Their diameters are 200 µm wide, which facilitates the entry of small-sized vesicles and the delivery of antibiotics or antimicrobials in order to kill bacteria inside the biofilm [31]. Moreover, different studies confirmed that positively-charged vesicles could electrostatically interact with bacterial cells, which have a negatively charged surface, and facilitate their entrance. The best range of zeta potential to achieve this effect is around 40-50 mV [32].
Similarly to our results, liposomes (DMPC:DOPE:CHEMS, molar ratio 4:4:2) designed to encapsulate lysins Pa7 and Pa119, also showed a lytic effect (when they were empty) against Pseudomonas aeruginosa cultures, probably due to their ability for membrane destabilization [33].
It is worth mentioning that only a few staphylococcal endolysins have been successfully encapsulated to date. LysMR-5, an endolysin derived from phage MR-5 was encapsulated in alginate-chitosan nanoparticles showing no change in structural integrity and bioactivity after entrapment [34]. Also, endolysin LysRODI encapsulated in pH-sensitive liposomes effectively reduced S. aureus cell counts by > 2 log units in both planktonic cultures and biofilms upon incubation at pH 5 [30].
Several strategies have been proposed to increase the stability of endolysins and allow their controlled release. Nanoparticles of chitosan derivatized with diethylaminoethyl groups act as ligands for the lytic protein Cpl-711 (ChiDENPs-711) improving its stability and releasing more than 90% of the active enzybiotic in approximately 2 h [35].
Gelatine is a natural biopolymer that is biodegradable, biocompatible, affordable and easy to procure. In the food industry, it is also used in the form of films to protect food against oxidation and microbial contamination, allowing long-term preservation [36]. Here, we also tested encapsulation of CHAPSH3b in biodegradable gelatine films suitable for food packaging purposes, as well as clinical or pharmaceutical applications. For instance, food packaging is an area of interest due to its high impact on food product quality and, over time, edible films have become widely used. If these films can serve as vehicles for transporting bioactive compounds their applicability can be extended even further.
Lytic protein CHAPSH3b was incorporated into gelatine films either directly (as free protein) or encapsulated into vesicles. To the best of our knowledge, this is the first time that a lytic protein has been embedded in this type of films, while bacteriophages have already been successfully tested as part of biodegradable films [37] and gelatin films [38]..
Our results prove that CHAPSH3b retains its antimicrobial activity after the film preparation procedure, which involves several temperature changes. However, we also observed that the empty vesicles have a high antimicrobial activity when added to gelatine films, and unfortunately no difference was detected between empty vesicles and lytic protein-loaded vesicles in gelatine films. In both cases, no bacterial growth was detected even after 14 days of films storage. Further studies would therefore be required to demonstrate if vesicle encapsulation prior to film preparation can further improve protein stability. A previous work in which phage PBSE191 was embedded in a PVA film [39] demostrated that the activity of the encapsulated phage against Salmonella was prolonged for 24 hours compared to that of the free phage. To our knowledge no studies have explored the activity of phages or endolysins within a film matrix after storage.
Overall, the results obtained in this work seem to confirm that encapsulation of lytic proteins is a promising strategy for enhancing their efficacy, and highlight the need to further explore the possibilities offered by nanovesicles in the field of phage-derived antimicrobials.
4. Materials and Methods
4.1. Chemical Compounds, Bacterial Strains and Proteins
The surfactants used as membrane components of nanovesicles were Span® 60 (Sigma-Aldrich, San Louis, Misuri, USA), cholesterol stabilized at 96% (Acros Organics, New Jersey, US) and cetyltrimethylammonium bromide (CTAB) (Sigma-Aldrich). All these components were dissolved in absolute ethanol (J.T. Baker, Avantor, Allentown, PA, USA). For the aqueous phase, phosphate buffer saline (PBS)(137 mM NaCl, 2.7 mM KCI, 8 mM Na2HPO4 2 mM KH2PO4 , pH 7.4) was prepared using phosphate buffered saline tablets (Oxoid, Hampshire, UK) according to the manufacturer's instructions. Gelatine films were prepared using gelatine from porcine skin (Sigma-Aldrich ref. G1890) and glycerol (Sigma-Aldrich 99.5% ref. G7893).
The lytic protein CHAPSH3b was extracted and purified according to the method described previously [17]. The bacterial strains used in this study were S. aureus 15981 [40] and S. aureus Sa9 [41], which were routinely grown at 37°C in TSB (tryptic soy broth, Scharlau Microbiology, Barcelona, Spain) with shaking or on plates containing TSB supplemented with 2% (w/v) agar (Roko, S.A., Llanera, Spain) (TSA) or Baird-parker agar plates (AppliChem, Germany).
4.2. Synthesis and Characterization of Nanovesicles
4.2.1. Synthesis of Nanovesicles
Preliminary experiments were made to determine the most appropriate combination of non-ionic surfactant Span 60, cholesterol as a membrane stabilizer and the cationic surfactant CTAB. Different ratios of Span 60 and CTAB were tested until the desired vesicle size was registered (30-80 nm).
The synthesis of nanovesicles was carried out using the thin film hydration (TFH) method that consists in the dissolution of the membrane components of the vesicles (total membrane compounds: 0.25 g, with mass ratio 1:1:0.5 for Span 60, CTAB and cholesterol) in an organic phase (10 ml of ethanol), followed by vacuum evaporation of the latter with a rotary evaporator. To obtain a homogeneous, transparent, and dry film, temperature (controlled at 40°C using a thermostatic water bath), pressure (70 mbar), rotation speed (135 rpm) and time were chosen according to previous works [24]. The lytic protein was firstly dissolved in the aqueous phase (25 ml) at 8 μM and then added to the flask to hydrate the thin layer. Two different solutions were tested as aqueous phase: pure MQ water and PBS. Finally, the mixture was incubated at 45°C (above the surfactants transition temperature) in a water bath (Figure 7). The same procedure was followed to obtain empty vesicles without addition of lytic protein.
4.1.2. Size and Morphology Characterization
The size and zeta potential of nanovesicles were measured (100 µl and 2 mL respectivetly) by dynamic light scattering (DLS) on a Zetasizer NanoZS series (Malvern Instruments Ltd., Malvern, UK) and the morphology was assessed by transmission electron microscopy (TEM) with a JEOL-2000 Ex II transmission electron microscope (Tokyo, Japan). For microscope observation a sample drop was placed on a carbon-coated copper grid and sample excess was removed with filter paper. Then, a drop of 2% (w/v) Phosphotungstic Acid solution (PTA) was applied to the carbon grid and left to stand for 1 minute. Once the excess of staining agent was removed with filter paper, the sample was air-dried and the thin film of stained and fixed vesicles was observed.
4.1.3. Vesicles Purification
Purification was carried out to remove the non-encapsulated material by ultracentrifugation at 35,000 rpm for 50 min) at 4˚C (centrifuge OptimaTMMAX ultacentrifuge 130,000 RPM, Beckman Coulter). The supernatant was filtered using 0.22 µm pore diameter PES syringe filters for further analysis by Reverse phase High Performance Liquid Chromatography (RP-HPLC).
4.1.4. Determination of the Protein Encapsulation Efficiency (EE)
In order to evaluate the encapsulation efficiency, two ml of vesicles suspension, in pure MQ water or PBS, were treated with a mixture of ethanol (25% in water). This was done for purified and non-purified systems. This step breaks the membrane bilayer and releases the encapsulated endolysin.
Samples were analysed and quantificated by chromatography by the measurement of the peak area. Sample separation was performed at room temperature with an HPLC 1100 series chromatograph (Agilent Technologies) equipped with a Superose™ 12 10/300 GL column (Cytiva, Buckinghamshire, UK), by injecting between 5 and 100 µl per sample. The eluents were 0.5 M NaCl and 0.2 M Tris(hydroxymethyl)aminomethane (Sigma-Aldrich) prepared in a solution with ultrapure water, buffered at pH 8 using HCl and filtered through a 0.2 μm nylon disc filter. The eluent flow rate was 0.5 mL/min, and a diode array detector was used recording absorbances at 224, 234, 244, 254 and 280 nm. Calibration was carried out using proteins of known molecular weight and the exclusion limits were determined with Blue Dextran 2000 (Cytiva, Buckinghamshire, UK) and acetone (Panreac, Barcelona, Spain), respectively.
An ESI-Q/TOF type mass spectrometer (IMPACT-II, Bruker) coupled to a UHPLC liquid chromatograph (DIONEX ULTIMATE3000, ThermoScientific) was used to determine the molecular weight of the eluted proteins in the HPSEC- chromatrography described above. A BIOshellTM A400 Protein C4 column (150 X 2.1 mm, 3.4 um GE HEalthCare), and the SUPELCOR LC Program (with the column oven at 70 degrees) were used. A gradient using two mobile phases: 0.1% formic acid in ultrapure water and 0.1% formic acid in acetonitrile was used. EE was calculated according to equation 1, using the areas measured from the chromatograms.
4.1.5. Antimicrobial Activity of Vesicles
For the antimicrobial tests, 25-ml samples containing empty and protein-loaded vesicles were ultracentrifuged at 35,000 rpm, for 50 min at 4°C (OpticaTM MAX ultracentruge, Beckman Coulter). The supernatant containing free non-encapsulated protein was removed and the pellet was resuspended with 250 µl of PBS or MQ water in order to purify the suspension from surfactants, empty vesicles and contaminants [24,42] .
An overnight culture of S. aureus 15981 was diluted (1:100 v/v) in fresh TSB medium supplemented with 0.25% (v/v) glucose (Merk, Darmstadt, Germany) (TSBg) and 1 ml-aliquots were inoculated into each well of a 24-well polystyrene microtiter plate (Thermo Scientific, Nunclon Delta Surface). These plates were incubated for 24 h at 37°C and, afterwards, the planktonic phase was removed, and the biofilms were washed twice with PBS. Biofilms were then treated with 0.5 ml of PBS (control), with empty vesicles, or with vesicles containing protein, and allowed to act for 1, 2, 4, 6 and 24 hours. After incubation, the planktonic phase was removed, and the adhered biofilms were washed twice with PBS. To assess the efficacy of the different treatments, the number of viable attached cells was quantified using the spot test. For that purpose, biofilms were scraped and resuspended in PBS. Afterwards, 10 µl droplets from tenfold serial dilutions of these cell suspensions were spotted onto TSA plates and allowed to dry. These plates were then incubated at 37°C for 24 h. All experiments were performed in triplicate. The cell counts obtained in these experiments were used to determine the number of colony forming units (CFU/cm2) per area unit using eq. 2:
4.2. Synthesis and Characterization of Gelatine Films
Gelatine films were prepared by direct mixing of 0.7 g of gelatine from porcine skin with 0.245 g of glycerol in 10 mL of water suspensions containing empty vesicles or protein-loaded vesicles. The mixtures were then placed in a water bath at 40°C for 25-30 minutes to dissolve the gelatine. Once dissolved, the solutions were poured into Petri dishes and allowed to dry in an oven at 35°C for 24 hours. Finally, the films were peeled off from the dishes with tweezers. Four different gelatine films were prepared: (i) plain gelatine films as a control, (ii) gelatine films with free lytic protein, (iii) films with empty vesicles and (iv) films with loaded vesicles. . All films prepared with endolysin contain a protein concentration of 0.057 µM/g.
4.2.1. Films Characterization
The morphology of the different films was observed using a scanning electron microscope (SEM) (JSM-5600, JEOL, Peabody, MA, USA) analyzing the surface and cross-sections. Films were cut into (0.2x0.2 mm) squares and placed on stubs, coated with gold.
4.2.2. Films Antimicrobial Activity
The antimicrobial activity of films was tested on S. aureus Sa9. Each film was divided equally in 4, immersing each piece in 10 mL of TSB medium with an initial concentration of 107 CFU/mL of S. aureus Sa9 and incubated for 4 h or 24 h at 37°C with shaking. After incubation, the number of viable cells was determined by serial dilution and plating on Baird-Parker medium and incubated at 37°C for 16 h. The films were also stored for 14 days at 4°C and tested again for residual antimicrobial activity. Data obtained from at least three independent biological replicates were analysed with a two-tailed Student’s t-test where p-values < 0.05 were considered significant.
4.3. Statistical Analysis of Data
All experiments were performed in triplicate. Results are shown as arithmetic mean values. To analyse differences between test groups, analysis of variance (ANOVA) was performed with IBM® SPSS® Statistics 25 statistical software. Significant differences between groups were determined using Fischer’s least significant difference (LSD) test. Statistical analysis of the biofilm data was carried out by a two-way ANOVA, using the dunnett’s multiple comparations test, using GraphPad Prism 6 software. In all tests, p-values < 0.05 were considered significant.
5. Conclusions
The present study demonstrates the feasibility of encapsulating endolysins with antimicrobial activity against S. aureus in non-ionic nanovesicles. The antimicrobial test indicates that vesicles loaded with endolysins had higher antimicrobial activity than free endolysins or empty vesicles. However, vesicular empty systems also exhibit inhibition activity.
This study also shows that it is possible to produce gelatine films containing the endolysin, free or encapsulated in vesicles, since the gelation process did not inhibit the antimicrobial activity of the lytic protein. Loaded and empty vesicles incorporated into gelatine films had high inhibition activity against films even after 14 days of storage. Nevertheless, the inhibition activity of the empty vesicles was so high that it did not allow us to discriminate between the activity of the encapsulated protein and that of the vesicles themselves.
The present study indicates the possibility of producing gelatine films with natural antimicrobial activity for food packaging, clinical or pharmaceutical applications.
Author Contributions
Conceptualization, SL, LF and PG., methodology, MM, ACD, SA, IM.; software, VM, ACD and SA.; validation, MM, LF and GG.; formal analysis, ACD, SA, LF, PG, SL, GG.; investigation, VM, SA and ACD.; resources, PG, MCB, MM and GG.; data curation, SL, LF, MM, and GG; writing—original draft preparation, VM and ACD; writing—review and editing, ACD, SA, LF, PG, MCB, SL, MM, and GG.; visualization, IM, MCB and PG.; supervision, LF, GG, PG and MCB,.; project administration, MCB and PG.; funding acquisition, MCB.
Funding
This work is part of a project that has received funding from the European Union’s Horizon 2020 research and innovation program under the Marie Sklodowska-Curie Grant Agreement No. 813439 (Break Biofilms). This research was also funded by the Ministerio de Economía y Competitividad (MINECO, Spain), grant number PID2019-105311RB-I00 and was also co-financed by Consejería de Educación y Ciencia del Principado de Asturias (AYUD/2021/52132).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis; 2017;
- European Centre for Disease Prevention and Control Antimicrobial Resistance Surveillance in Europe; 2022;
- Pinto, R.M.; Lopes-De-Campos, D.; Martins, M.C.L.; Van Dijck, P.; Nunes, C.; Reis, S. Impact of Nanosystems in Staphylococcus Aureus Biofilms Treatment. FEMS Microbiol. Rev. 2019, 43, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.W. Biofilms and Antibiotic Therapy: Is There a Role for Combating Bacterial Resistance by the Use of Novel Drug Delivery Systems? Adv. Drug Deliv. Rev. 2005, 57, 1539–1550. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Fernández, L.; Rodríguez, A.; García, P. Are Phage Lytic Proteins the Secret Weapon to Kill Staphylococcus Aureus? Am. Soc. Microbiol. 2018, 9, e01923–17. [Google Scholar] [CrossRef] [PubMed]
- Forier, K.; Raemdonck, K.; De Smedt, S.C.; Demeester, J.; Coenye, T.; Braeckmans, K. Lipid and Polymer Nanoparticles for Drug Delivery to Bacterial Biofilms. J. Control. Release 2014, 190, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Yang, E.; Chang, P.S.; Ryu, S. Preparation and Characterization of Endolysin-Containing Liposomes and Evaluation of Their Antimicrobial Activities against Gram-Negative Bacteria. Enzyme Microb. Technol. 2019, 128, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; González, S.; Campelo, A.B.; Martínez, B.; Rodríguez, A.; García, P. Downregulation of Autolysin-Encoding Genes by Phage-Derived Lytic Proteins Inhibits Biofilm Formation in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2017, 61, e02724–16. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; Donovan, D.M.; García, P. Enhanced Staphylolytic Activity of the Staphylococcus Aureus Bacteriophage VB_SauS-PhiiPla88 HydH5 Virion-Associated Peptidoglycan Hydrolase: Fusions, Deletions, and Synergy with LysH5. Appl. Environ. Microbiol. 2012, 78, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- De Maesschalck, V.; Gutiérrez, D.; Paeshuyse, J.; Lavigne, R.; Briers, Y. Advanced Engineering of Third-Generation Lysins and Formulation Strategies for Clinical Applications. Crit. Rev. Microbiol. 2020, 1–17. [Google Scholar] [CrossRef]
- Irkin, R.; Esmer, O.K. Novel Food Packaging Systems with Natural Antimicrobial Agents. J. Food Sci. Technol. 2015, 52, 6095–6111. [Google Scholar] [CrossRef]
- Duarte, A.C.; Fernández, L.; De Maesschalck, V.; Gutiérrez, D.; Campelo, A.B.; Briers, Y.; Lavigne, R.; Rodríguez, A.; García, P. Synergistic Action of Phage PhiIPLA-RODI and Lytic Protein CHAPSH3b: A Combination Strategy to Target Staphylococcus Aureus Biofilms. npj Biofilms Microbiomes 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Toledo-Arana, A.; Merino, N.; Vergara-Irigaray, M.; Débarbouillé, M.; Penadés, J.R.; Lasa, I. Staphylococcus Aureus Develops an Alternative, Ica-Independent Biofilm in the Absence of the ArlRS Two-Component System †. J. Bacteriol. 2005, 187, 5318–5329. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Khanal, D.; Alreja, A.B.; Yang, H.; YK Chang, R.; Tai, W.; Li, M.; Nelson, D.C.; Britton, W.J.; Chan, H.K. Bacteriophage Endolysin Powders for Inhaled Delivery against Pulmonary Infections. Int. J. Pharm. 2023, 635, 122679. [Google Scholar] [CrossRef] [PubMed]
- Gondil, V.S.; Chhibber, S. Bacteriophage and Endolysin Encapsulation Systems: A Promising Strategy to Improve Therapeutic Outcomes. Front. Pharmacol. 2021, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.; Paris, J.L.; Gama, F.M.; Silva, B.F.B.; Sillankorva, S. Sustained Release of a Streptococcus Pneumoniae Endolysin from Liposomes for Potential Otitis Media Treatment. ACS Infect. Dis. 2021, 7, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Garrido, V.; Fernández, L.; Portilla, S.; Rodríguez, A.; Grilló, M.J.; García, P. Phage Lytic Protein LysRODI Prevents Staphylococcal Mastitis in Mice. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Mal, A.; Bag, S.; Ghosh, S.; Moulik, S.P. Physicochemistry of CTAB-SDS Interacted Catanionic Micelle-Vesicle Forming System: An Extended Exploration. Colloids Surfaces A 2018, 553, 633–644. [Google Scholar] [CrossRef]
- Sobral, C.N.C.; Soto, M.A.; Carmona-Ribeiro, A.M. Characterization of DODAB/DPPC Vesicles. Chem. Phys. Lipids 2008, 152, 38–45. [Google Scholar] [CrossRef]
- Qian, Y.; Hu, X.; Wang, J.; Li, Y.; Liu, Y.; Xie, L. Polyzwitterionic Micelles with Antimicrobial-Conjugation for Eradication of Drug-Resistant Bacterial Biofilms. Colloids Surfaces B Biointerfaces 2023, 231, 113542. [Google Scholar] [CrossRef]
- Marchianò, V.; Matos, M.; López, M.; Weng, S.; Serrano-Pertierra, E.; Luque, S.; Blanco-López, M.C.; Gutiérrez, G. Nanovesicles as Vanillin Carriers for Antimicrobial Applications. Membranes (Basel). 2023, 13, a93. [Google Scholar] [CrossRef]
- Estupiñan, O.; García Manrique, P.; Blanco-lopez, M.; Matos, M.; Gutierrez, G. Vitamin d3 loaded niosomes and transfersomes produced by ethanol injection method: Identification of the critical preparation step for size control. Foods 2020, 10, 1367. [Google Scholar] [CrossRef]
- Estupiñan, O.R.; Garcia-Manrique, P.; del Carmen Blanco-Lopez, M.; Matos, M.; Gutiérrez, G. Vitamin D3 Loaded Niosomes and Transfersomes Produced by Ethanol Injection Method: Identification of the Critical Preparation Step for Size Control. Foods 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- García-Manrique, P.; Machado, N.D.; Fernández, M.A.; Blanco-López, M.C.; Matos, M.; Gutiérrez, G. Effect of Drug Molecular Weight on Niosomes Size and Encapsulation Efficiency. Colloids Surfaces B Biointerfaces 2020, 186, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Marchianò, V.; Matos, M.; Serrano, E.; Álvarez, J.R.; Marcet, I.; Carmen Blanco-López, M.; Gutiérrez, G. Lyophilised Nanovesicles Loaded with Vitamin B12. J. Mol. Liq. 2022, 365, 120129. [Google Scholar] [CrossRef]
- Melis, V.; Letizia Manca, M.; Bullita, E.; Tamburini, E.; Castangia, I.; Cardia, M.C.; Valenti, D.; Fadda, A.M.; Peris, J.E.; Manconi, M. Inhalable Polymer-Glycerosomes as Safe and Effective Carriers for Rifampicin Delivery to the Lungs. Colloids Surfaces B Biointerfaces 2016, 143, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Audia Triani, O.; Akhmad Kharis, Nugroho Ronny, M. ; Sugeng, R. Effect of Ratio Span 60 - Cholesterol on the Characteristic of Niosomes Vitamin D3. Res. J. Pharm. Technol. 2022, 15, 5551–5554. [Google Scholar] [CrossRef]
- Bucci, A.R.; Marcelino, L.; Mendes, R.K.; Etchegaray, A. The Antimicrobial and Antiadhesion Activities of Micellar Solutions of Surfactin, CTAB and CPCl with Terpinen-4-Ol: Applications to Control Oral Pathogens. World J. Microbiol. Biotechnol. 2018, 34, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Qiao, J.; Xiong, M.P. Antibacterial and Biofilm-Eradicating Activities of PH-Responsive Vesicles against Pseudomonas Aeruginosa. Mol. Pharm. 2022, 19, 2406–2417. [Google Scholar] [CrossRef] [PubMed]
- Portilla, S.; Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Encapsulation of the Antistaphylococcal Endolysin Lysrodi in Ph-Sensitive Liposomes. Antibiotics 2020, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dai, X.; Wei, X.; Yu, Y.; Chen, X.; Zhang, X.; Li, C. Near-Infrared Light-Activated Thermosensitive Liposomes as Efficient Agents for Photothermal and Antibiotic Synergistic Therapy of Bacterial Biofilm. ACS Appl. Mater. Interfaces 2018, 10, 14426–14437. [Google Scholar] [CrossRef]
- Selvamani, V. Stability Studies on Nanomaterials Used in Drugs. Charact. Biol. Nanomater. Drug Deliv. 2019, 425–444. [Google Scholar]
- Morais, D.; Tanoeiro, L.; Marques, A.T.; Gonçalves, T.; Duarte, A.; Matos, A.P.A.; Vital, J.S.; Cruz, M.E.M.; Carvalheiro, M.C.; Anes, E.; et al. Liposomal Delivery of Newly Identified Prophage Lysins in a Pseudomonas Aeruginosa Model. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Kour, A.; Panda, J.J.; Harjai, K.; Chhibber, S. Exploring Endolysin-Loaded Alginate-Chitosan Nanoparticles as Future Remedy for Staphylococcal Infections. AAPS PharmSciTech 2020, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, R.; Caro-León, F.J.; Nakal, A.; Ruiz, S.; Doñoro, C.; García-Fernández, L.; Vázquez-Lasa, B.; San Román, J.; Sanz, J.; García, P.; et al. DEAE-Chitosan Nanoparticles as a Pneumococcus-Biomimetic Material for the Development of Antipneumococcal Therapeutics. Carbohydr. Polym. 2021, 273, 118605. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Ma, Q.; Li, S.; Wang, W.; Ma, Y.; Zhao, H.; Sun, J.; Wang, J. Preparation and Characterization of Biodegradable Composited Films Based on Potato Starch/Glycerol/Gelatin. J. Food Qual. 2021, 2021, 1–11. [Google Scholar] [CrossRef]
- de Dicastillo, C.L.; Settier-Ramírez, L.; Gavara, R.; Hernández-Muñoz, P.; Carballo, G.L. Development of Biodegradable Films Loaded with Phages with Antilisterial Properties. Polymers (Basel). 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.; López, A.; Sáez-Orviz, S.; Marcet, I.; García, P.; Rendueles, M.; Díaz, M. Effectiveness of Bacteriophages Incorporated in Gelatine Films against Staphylococcus Aureus. Food Control 2021, 121, 1–8. [Google Scholar] [CrossRef]
- Kim, S.; Chang, Y. Anti-Salmonella Polyvinyl Alcohol Coating Containing a Virulent Phage PBSE191 and Its Application on Chicken Eggshell. Food Res. Int. 2022, 162, 111971. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Toledo-Arana, A.; Berasain, C.; Ghigo, J.-M.; Amorena, B.; Penadés, J.R.; Lasa, I. SarA and Not ΣB Is Essential for Biofilm Development by Staphylococcus Aureus. Mol. Microbiol. 2003, 48, 1075–1087. [Google Scholar] [CrossRef]
- Obeso, J.M.; Martínez, B.; Rodríguez, A.; García, P. Lytic Activity of the Recombinant Staphylococcal Bacteriophage ΦH5 Endolysin Active against Staphylococcus Aureus in Milk. Int. J. Food Microbiol. 2008, 128, 212–218. [Google Scholar] [CrossRef]
- Marchianò, V.; Matos, M.; Serrano-Pertierra, E.; Gutiérrez, G.; Blanco-López, M.C. Vesicles as Antibiotic Carrier: State of Art. Int. J. Pharm. 2020, 585, 119478. [Google Scholar] [CrossRef]
Figure 1.
Particle size distribution of nanovesicles in (A) pure MQ water and (B) PBS obtained by DLS in number.
Figure 1.
Particle size distribution of nanovesicles in (A) pure MQ water and (B) PBS obtained by DLS in number.
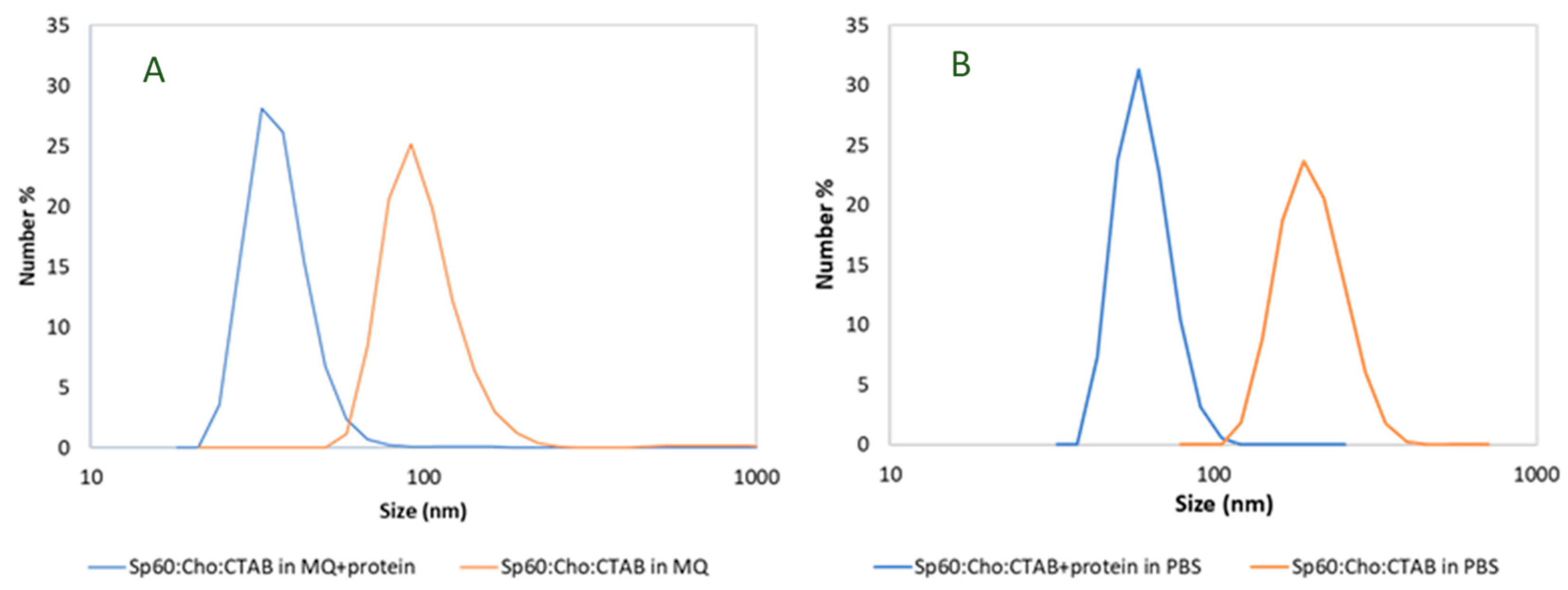
Figure 2.
TEM images of nanovesicles stained with... A) empty vesicles in water; B) protein-loaded vesicles in water; C) empty vesicles in PBS; D) protein-loaded vesicles in PBS.
Figure 2.
TEM images of nanovesicles stained with... A) empty vesicles in water; B) protein-loaded vesicles in water; C) empty vesicles in PBS; D) protein-loaded vesicles in PBS.

Figure 3.
Protein elution profiles obtained after measurement (A 280 nm) of fractions eluted from molecular exclusion chromatography of CHAPSH3b (4, 8, 12 µM) in ultrapure water (A) and in PBS (B).
Figure 3.
Protein elution profiles obtained after measurement (A 280 nm) of fractions eluted from molecular exclusion chromatography of CHAPSH3b (4, 8, 12 µM) in ultrapure water (A) and in PBS (B).
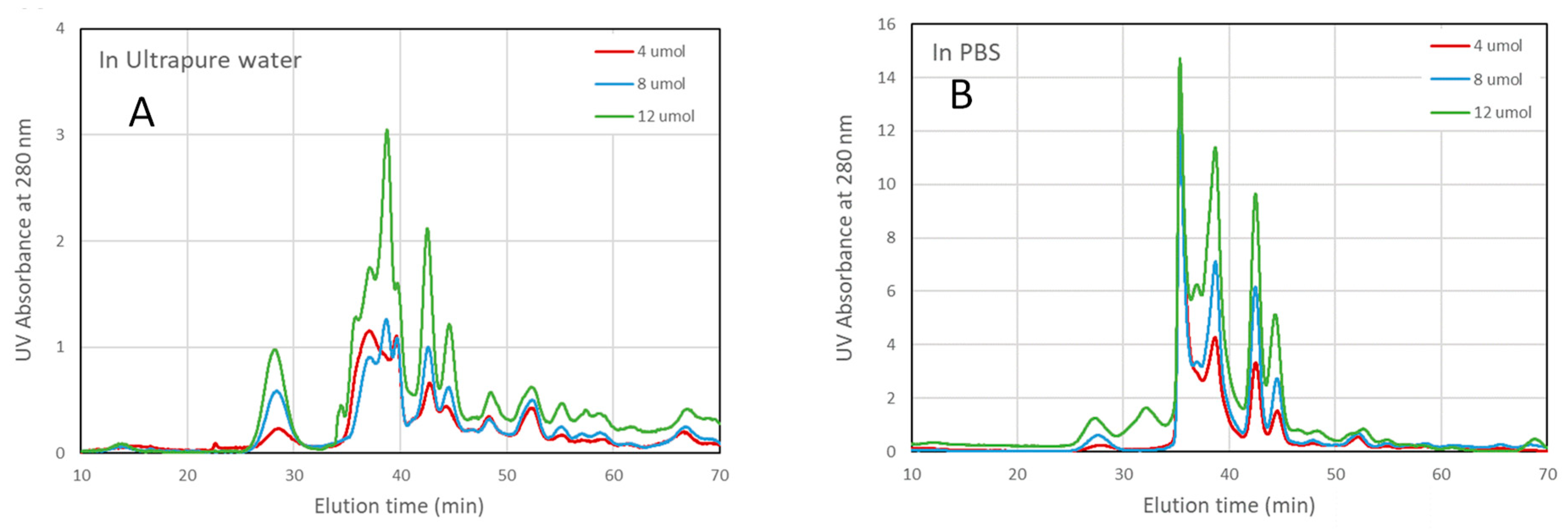
Figure 4.
Elution profiles of fractions obtained by molecular exclusion chromatography of the solvent (ethanol), empty vesicles, and protein/vesicles/solvent after measurement at A) different wavelengths (blue line: 224, green line: 254, and red line: 280 nm), B) at 254 nm. Red line: protein CHAPSH3b, green line: mixture of empty vesicles (1/3) and protein (2/3) solution; blue line: mixture of empty vesicles (2/3) and protein (1/3) solution. The green arrow indicates the CHAPSH3b fraction.
Figure 4.
Elution profiles of fractions obtained by molecular exclusion chromatography of the solvent (ethanol), empty vesicles, and protein/vesicles/solvent after measurement at A) different wavelengths (blue line: 224, green line: 254, and red line: 280 nm), B) at 254 nm. Red line: protein CHAPSH3b, green line: mixture of empty vesicles (1/3) and protein (2/3) solution; blue line: mixture of empty vesicles (2/3) and protein (1/3) solution. The green arrow indicates the CHAPSH3b fraction.
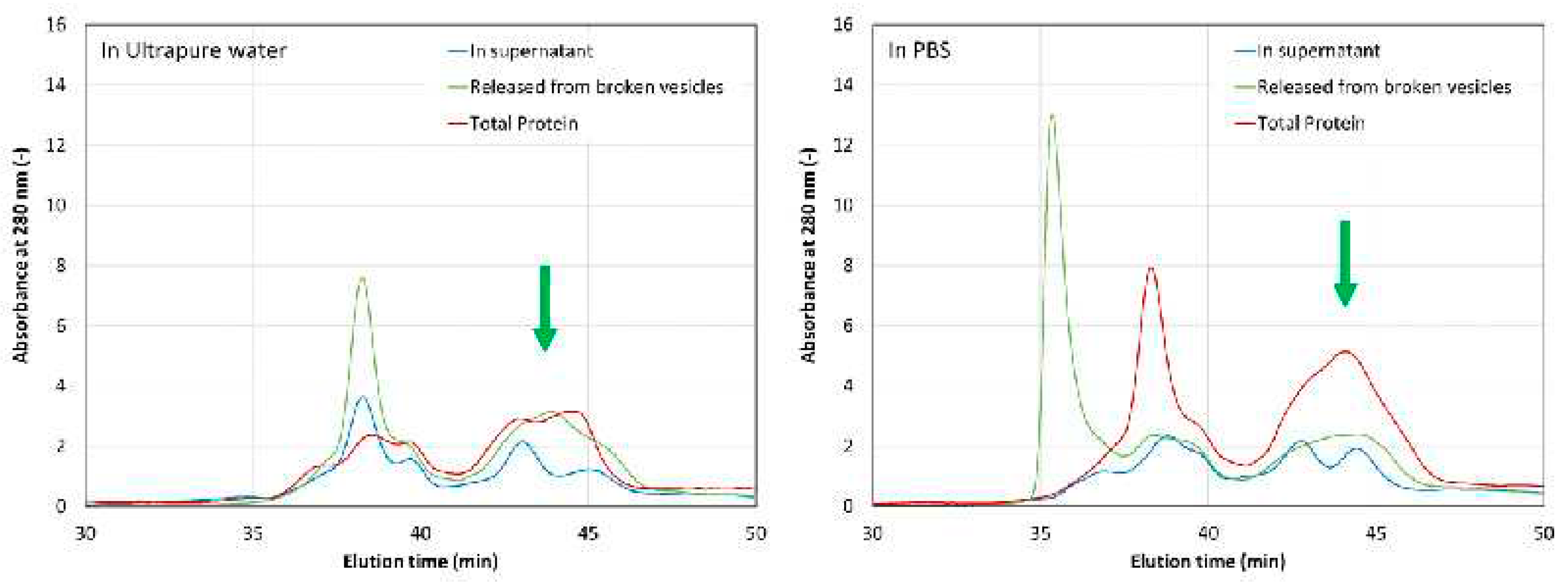
Figure 5.
HPSEC chromatographs (at 280 nm) for different samples eluted in ultrapure water (A) and in PBS (B). Protein-loaded vesicles were separated from the supernatant (blue line) and then the vesicles were broken (green line). A mixture of broken vesicles (total protein) was also analysed (red line).
Figure 5.
HPSEC chromatographs (at 280 nm) for different samples eluted in ultrapure water (A) and in PBS (B). Protein-loaded vesicles were separated from the supernatant (blue line) and then the vesicles were broken (green line). A mixture of broken vesicles (total protein) was also analysed (red line).

Figure 6.
Analysis by SEM of gelatine films obtained with A) PBS, B) empty vesicles, C) free protein, and D) vesicles loaded with protein.
Figure 6.
Analysis by SEM of gelatine films obtained with A) PBS, B) empty vesicles, C) free protein, and D) vesicles loaded with protein.
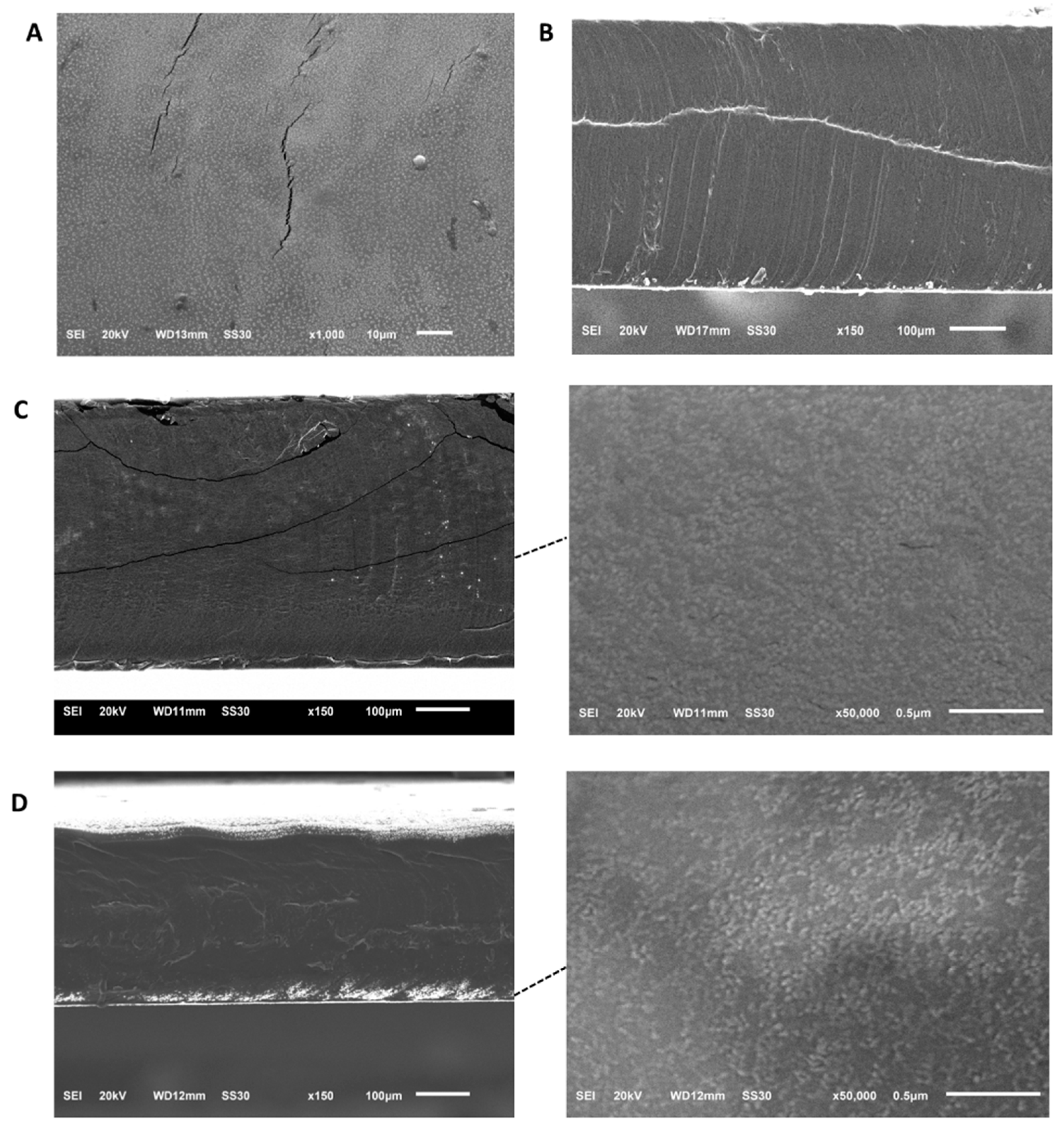
Figure 7.
Thin film hydration method used for the synthesis of vesicles containing the lytic protein CHAPSH3b.
Figure 7.
Thin film hydration method used for the synthesis of vesicles containing the lytic protein CHAPSH3b.
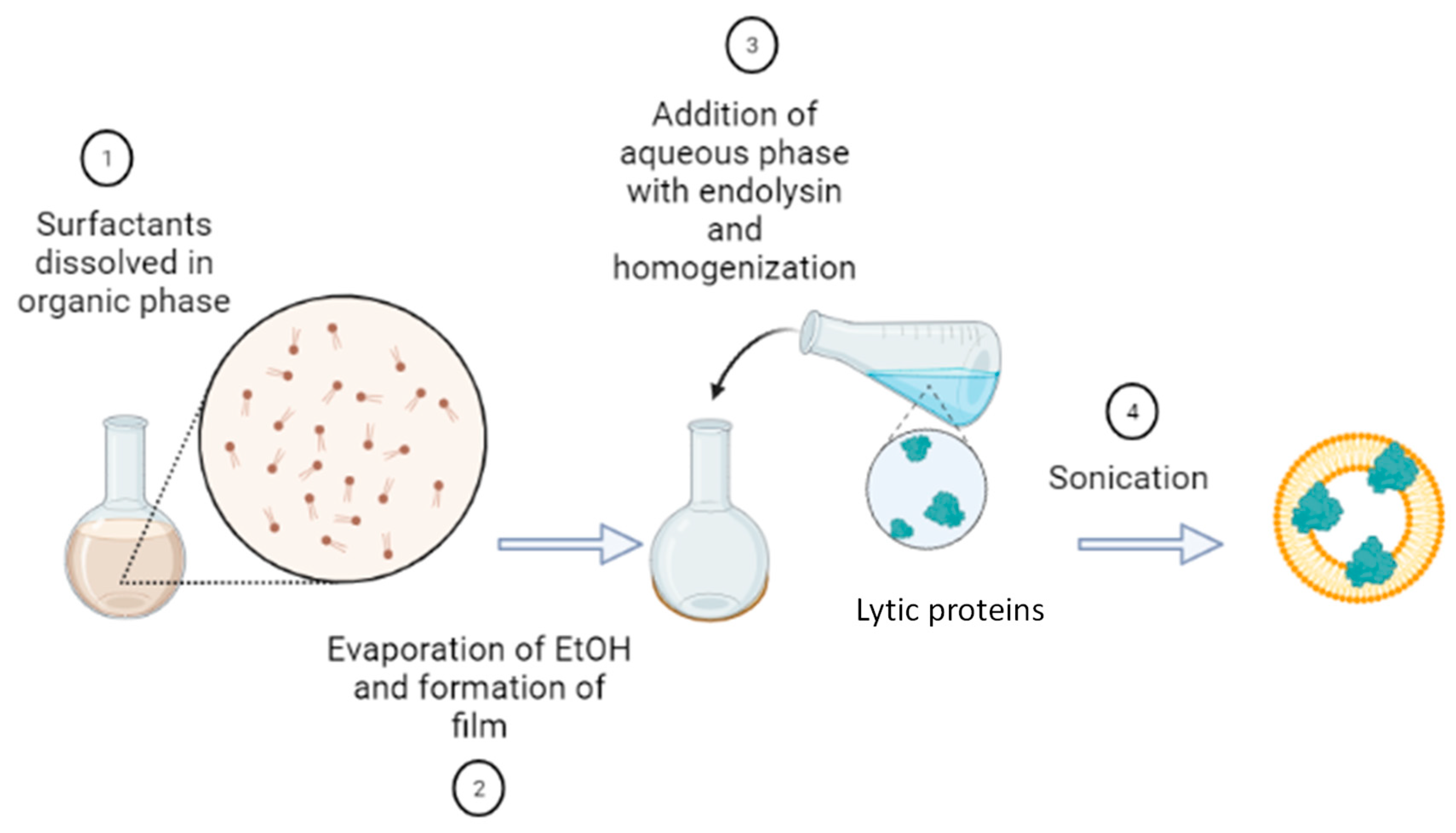

Table 1.
Mean particle size and zeta potential of nanovesicles prepared in different aqueous media.
| Formulation | Aqueous phase | Size (nm) | Zeta potential (mV) |
| Sp60 : Cho : CTAB (no protein) | MQ water | 100±27 | 55±2 |
| Sp60 : Cho : CTAB + CHAPSH3b (8 µM) | MQ water | 38±18 | 46±5 |
| Sp60 : Cho : CTAB (no protein) | PBS buffer | 205±46 | 28±2 |
| Sp60 : Cho : CTAB + CHAPSH3b (8 µM) | PBS buffer | 77±21 | 30±4 |
Table 2.
24-h-old biofilms of strain S. aureus 15981 treated with free CHAPSH3b, empty and protein-loaded vesicles.
Table 2.
24-h-old biofilms of strain S. aureus 15981 treated with free CHAPSH3b, empty and protein-loaded vesicles.
| Time | Control | CHAPSH3b (8 µM) | Empty vesicles | CHAPSH3b (8 µM) loaded vesicles |
| 1 h | 8.42 ± 0.06 | 7.85 ± 0.02 | 5.27 ± 0.90 | 3.74 ± 1.71** |
| 2 h | 8.24 ± 0.13 | 7.28 ± 0.60 | 4.13 ± 3.60* | 3.90 ± 3.40** |
| 4 h | 7.62 ± 0.71 | 6.50 ± 0.46 | 1.33 ± 2.30**** | 0.00 ± 0.00**** |
| 6 h | 8.72 ± 0.58 | 7.32 ± 0.06 | 1.15 ± 1.99**** | 0.00 ± 0.00**** |
| 24 h | 6.81 ± 0.30 | 6.36 ± 0.68 | 3.22 ± 2.79* | 3.07 ± 2.66* |
Values correspond to Log (CFU/cm2). Data represent the means ± standard deviations of three independent experiments. Values with an asterisk are statistically different from the untreated control of each time point with two-way ANOVA using Dunnett’s multiple comparisons test. *p-value < 0.05, **p-value < 0.01 and ****p-value <0.0001.
Table 3.
Antimicrobial activity of gelatine films (obtained with empty and protein-loaded vesicles) against a S. aureus Sa9 suspension. S. aureus Sa9 cultures (control) and treated with gelatine films containing free protein CHAPSH3b were also tested. .
Table 3.
Antimicrobial activity of gelatine films (obtained with empty and protein-loaded vesicles) against a S. aureus Sa9 suspension. S. aureus Sa9 cultures (control) and treated with gelatine films containing free protein CHAPSH3b were also tested. .
| Incubation Time | Control | CHAPSH3b (8 μM) | CHAPSH3b (8 µM) loaded vesicles | Empty vesicles |
|---|---|---|---|---|
| 4 h | 8.6±0.30 | 5.18±0.17* | 0.00±0.00* | 0.00±0.00* |
| 24 h | 9.50±0.17 | 7.43±0.32* | 0.00±0.00* | 0.00±0.00* |
| After 14 days of storage | ||||
| 4 h | 9.02±0.12 | 8.57±0.22 | 0.00±0.00* | 0.00±0.00* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



