
Submitted:
06 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
We propose a silviculture with a purely biological logic that takes soil into account. The article is divided into three parts: 1) presentation of the issue in the context of biodiversity and forest pro-tection; 2) impact of the soil functioning on forestry; 3) possible implementation of soil functioning in forestry practice, to support and protect a non-or minimally anthropized evolution of the forest.
The first chapter illustrates the importance of the forest for the future of the planet and our human species, referring to recently published works on the topic of climate warming and biodiversity. The second illustrates the aspects that link soil dynamics to the silvogenetic cycle, focusing on the humipedon (organic and organo-mineral surface horizons, approximately the first 30 cm of forest soil), and more significant carbon storage in old forests since up to 2/3 is found in the soil. In the third chapter we consider the possibility of lengthening the forestry cycle, to allow the return of nutrients to the soil. Theoretical graphs and distribution models of the number of trees as a func-tion of diameter and age are reported, with examples of logging in uneven Alpine forests, with reference to Swiss fir-beech forests published on the Pro-Silva website.
Keywords:
Sylviculture
; forestry
; forest management
; forest dynamics
; humipedon
; humus system
; carbon cy-cle
; global change
; soil biodiversity
1. Introduction
1.1. The Forbidden Apple
Well-known biblical recommendation (Figure 1): “You can do whatever you want, planet Earth is yours, plants, animals, mountains, sea, clouds, everything. That apple tree you see in the middle of the meadow, only that one, behold, don’t eat its apples.”
6-7 million years ago, the lineage we share with chimpanzees bifurcated, generating a branch of hominids. They differed from their ancestors because they preferred to stand upright position, on two feet instead of four [1]. Until then, and for a few million years more, evolution depended on environmental conditions, first astronomical, chemical, physical, then increasingly biological. Little by little, the hominids changed the cards on the table: for 10 thousand years now, but above all and definitely in the last century, evolution depends directly on the needs of this new species of living beings that we are.
1.2. The Behavior of Living Beings
List of biological truths that indirectly support the content and conclusions of this article (these are synthetic lines of thought - therefore incomplete - but scientifically shared in the essential):
Nothing in Biology Makes Sense Except in the Light of Evolution. Dobzhansky (1973) [2]: “Without that light biology becomes a pile of miscellaneous facts, some of them interesting or curious but which do not create a meaningful picture as a whole. This is not to say that we know everything that can and should be known about biology and evolution. Every competent biologist is aware of a multitude of unsolved problems and unanswered questions. After all, biological research shows no sign of nearing completion; exactly the opposite is true.”
The first microorganisms were present on our planet already 3.7 billion years ago (https://naturalhistory.si.edu/education/teaching-resources/life-science/early-life-earth-animal-origins). The first unicellular organisms capable of photosynthesis appeared 2.4 billion years ago, while the first multicellular animals arrived 800 million years ago. Coniferous and deciduous trees appeared 400 and 200 million years ago respectively, flowering plants began to populate the planet only 150 million years ago. Progressively, living organisms took control of the planetary environment [3]. The most obvious example was the increase in the concentration of oxygen in the air and the decrease in that of CO2 following the development of plants, with consequent radical modification of the planet’s climate (summarized in Dahl et al. 2020) [4].
Predators in a food web cannot eliminate all their prey because if they did, they would starve. The energy extractable from prey and useful to the predator is worth approximately 1/10 of that ingested by eating the prey [5]; there is therefore a minimum threshold of prey density below which t predator cannot survive. When the energy contained in the remaining prey is no longer sufficient to support the number of individuals in the predator’s population, that number automatically decreases [6,7].
Each species is linked to the others in an almost mathematical way (ecological pyramid: https://en.wikipedia.org/wiki/Ecological_pyramid ) and together they co-evolve in ecosystems that cover the planet with a biodiversity that has increased over time, although still poorly understood major crises every 62 (145) million years characterize it [8].
A species becomes extinct when it can no longer reproduce in the system that contains it. It is like one of the countless wheels in the gear of a living system. When “the species no longer revolves in harmony with the others”, when it prevents the movement of the whole, it is “abandoned to itself”, and being isolated, it dies. On average this happens every 5-7 million years [9]. In general, it is replaced by species that are better adapted to the motor gear that evolves with the system in which the species live. Leaving aside the numerous variations, today two theories are currently competing to explain the evolution of the natural world on our planet:
- -
- competition between species for the use of resources: the individuals of the species that reproduce most successfully in this historical period and in this environment are selected; the individuals that make up the populations of a species are constantly changing and in constant competition in the exploitation of the environment with individuals of many other species [10,11,12];
- -
- the cooperation of individuals in the use of resources: whatever the species, it is a complex system of individuals adapted to each other in order to grow, a kind of supra-organism contained in other larger systems, making the best possible use of resources and the available energy of an environment that is changing with it [3,13,14].
Practically the evolving system unit is like a sports team: it wins the race when: 1) it is composed of individuals with peculiar characteristics that are complementary to those of the others; 2) the players “team up”, that is, they functionally cooperate with others to win; 3) the individuals and team know how to develop an unpredictable and effective game during the match; 4) the environment in which the game is played must also come out on top, that is, it must have more resources at the end than at the start of the game, in order to continue supporting new races in the long term. This statement was published indirectly with the documented natural history of our planet (https://en.wikipedia.org/wiki/History_of_Earth ).
1.3. Toumaï and the Anthropocene
The hominid who lived 6-7 million years ago (these are the oldest hominid fossil remains ever found) was given the name “Toumaï”. The scientific classification of this species is as follows [15]:
Order Primates L., 1758; Spttorder Anthropoidea Mivart, 1864; Superfamily Hominoidea Gray, 1825; Family Hominidae Gray, 1825; Genus Sahelanthropus gen. Nov., Species Sahelanthropus tchadiensis sp. Nov.
Using their innate intelligence to live better in their environment, Tomaï’s offspring more or less voluntarily triggered the new geological era of the Anthropocene (not yet officially recognized as of 2022: https://en.wikipedia.org/wiki/Anthropocene). Three million years later, the skeleton linked to the upright position of this distant relative was perfected in Lucy (Australopithecus afarensis) [16]. The still relatively small brain revealed that the upright position preceded (it was the spark that ignited) the development of the size of the brain, which we find much more voluminous in modern Homo sapiens. Standing upright, they could see further, above the grasses of the savannah; erect and stable on two feet, they had their hands free to make new tools for more effective experiences and easier conquests [16].
If theoretically the Anthropocene began when a previously quadrupedal and arboreal animal “became” bipedal and terrestrial, as long as this new living being remained a hunter-gatherer, the influence on natural evolution was that of a species that respected the laws presented above. In a noticeable way, the replacement of the natural forest by the artificial one began only 10 thousand years ago, with the change of land use imposed by agriculture. We are talking about populations of Homo sapiens similar in genetic inheritance to modern ones, with a brain capable of using natural resources to produce abundant food, dwelling houses and monuments of organized societies. The division of tasks in the social fabric left free time for groups of dominant individuals to scientifically conceive the exploitation of other living beings and the environment [17].
It is no longer possible to continue to consider the environment (living and non-living) as something external and not connected to the living structure of human society for the following reasons:
1) Every human organism hosts numerous microorganisms that it needs to live [18]. These microorganisms are in equilibrium with those external to us and widespread in the environment. In surface soil these microorganisms number billions of cells per gram of soil; an equivalent number is found in 1 m3 of air we breathe.
2) Every human organism eats to obtain from food the energy it needs to live. What we eat is made of mineral elements that make up the earth’s crust (https://www.weforum.org/agenda/2021/12/abundance-elements-earth-crust/). From a nutritional point of view, living organisms are chemical elements (a concentrate of elements useful for the development of living cells and other systems), and energy, which move and are maintained on the surface of the planetary sphere, and which thanks to the process of recycling are available for future generations [19].
3) Like all other living things, the human species modifies the environment in which it develops and reproduces. The difference with other species lies in the magnitude of the change that our species is causing. The impact is decidedly greater than that of other species since humans began extracting energy from fossil organic matter to turn engines and produce mechanical and electrical energy. We are practically talking about pratically all modern industry and agriculture, with all means of transportation, lighting, information transmission, construction, education, cultivation, in short the list includes everything that has made human life easier in the last century [20]. The impact on the environment is so strong that it is dangerously influencing the characteristics of the planet’s climate [21].
In the coming decades, the species that are more resistat to the new climate will be selected along with the microorganisms that reproduce better in a warmer environment [22]. The phenomenon will be global, even if zonal (different in intensity and consequences depending on the geographical position), but it will also affect the human species, with the microorganisms associated with it. The idea of solving the problem by generalizing the air conditioning of environments in the areas most sensitive to the phenomenon involves nothing less than the risk of the extinction of our species [23].
1.4. The Forests of the World
1.4.1. State of the Art
The surface of the planet (51 billion ha = 51 x 109 ha consists of 71% oceans and 29% of emerged lands (14.9 x 109 ha). Of these emerged lands, 71% (10.6 x 109 ha) is habitable (it is not bare rock, or desert or ice). Ten thousand years ago, forests covered 57% (6 x 109 ha) of habitable land; today 38% (about 4.02 x 109 ha), half (2 x 109 ha) are almost intact and a third (1.33 x 109 ha) are in the state of primary forest, i.e. naturally regenerating with primary species [24]. The accuracy of the estimate depends on the definition of the term forest. These calculations are operated from satellite with timely checks in the field. A recent definition [25] uses the term area covered by trees, i.e. “woody vegetation with a height of at least 5 meters and a canopy density of at least 20-25%, with a resolution of 30 meters. This definition includes unmanaged and managed natural forests, tree plantations, and urban forests. Our planet has 4.02 x 109 ha of tree cover, which correspond to 30% of the surface of the emerged lands and 38% of the habitable lands. A minimum size of 0.5 hectares not enforced by these authors is often included in many forest definitions. Forest cover is reduced by 3-4% if plots < 0.5 ha in size are not considered.
In 1700 the area devoted to agriculture occupied 3% of the habitable land, and grazing 6%; in 2020 these values became 15 and 31% respectively; the part occupied by ungrazed prairies and shrublands reaches 14%.
Gradually, cultivation, grazing and urbanization have occupied 1/3 of the forest area [26]. As human populations living in forests are relatively disarmed and declining [27], and urban populations continue to grow and require resources and food, the replacement process is called for to continue to meet the needs of humans and domestic animals [28]
1.4.1. Can Humans Live far from Earth?
Let’s try to argue in the affirmative. It would be enough to take with us greenhouses full of plants and animals that will produce food for us in space, when we set out in search of other worlds as in science fiction movies. Is this journey in the company of a small productive part of the planet really possible? Some astronauts have lived in space for more than a year. Valeri Polyakov was the most resilient of all, and remained in orbit 437 days and 18 hours (https://en.wikipedia.org/wiki/Valeri_Polyakov). Everyone else returns first, not out of nostalgia, but for health reasons (https://en.wikipedia.org/wiki/Effect_of_spaceflight_on_the_human_body). Our body have adapted to gravity. In space, our blood no longer circulates in our body as it does on Earth. Muscles (even the heart) and bones atrophy. The ear functions poorly, and is linked to orientation in space, with sensations of dizziness and vomiting. Back on Earth, it takes astronauts several months to be able to walk like before. Furthermore, and closer to the subject of this article, on Earth we are surrounded by a cloud of microorganisms, that we breathe and eat all the time. In the air of a spacecraft, the microorganisms are much fewer and not connected to all the others that are prevalent and evolving in the environment on Earth. There are dangers of infection, the ecological balance between groups of microorganisms being broken; it is necessary to filter the air continuously, as and better than a sterile hospital room [29]. Considering that current techniques allow us to stem the problem but not to solve it completely [30], we still don’t know microorganisms well.
The reality is very different from what they show us on television: most of us would die during a space journey that would take us to the moon. And the moon is one light second away from us (if we want to go around another sun, the distances are at least 3 light years minimum). The moon is worse, much worse than the depths of the Sahara in midsummer or the top of Mont Blanc in midwinter: there is no air no water, not even microorganisms live there; if you stick your nose out of the diving suit, it turns to stone in a few seconds and freezes your brain forever. It takes 6 months to go to Mars with the means not yet available today (https://mars.nasa.gov/mars2020/timeline/cruise/). The Moon is a piece of Earth; in comparison you can easily come back home; Mars is really another planet, which requires knowledge that we don’t yet have. In Ridley Scott’s 2015 film “The Martian” (https://en.wikipedia.org/wiki/The_Martian_(film)), based on the book by Andy Weir [31], Matt Damon attempts to produce potatoes on Mars using astronaut’s excrements as a source of energy and minerals, after having produced terrestrial air in a greenhouse hermetically sealed from the outside environment. The story makes you dream and ends well, as in all American films. But the reality is closer to that told in another film work entitled “First man”, by Damien Chazelle in 2018 (https://en.wikipedia.org/wiki/First_Man_(film), where Rayan Gosling in the role by Neil Armstrong manages to set foot on the moon. The difficulties are impressive, only a young Reinhold Messner, the man who went from one over 8000 to another without oxygen, would perhaps be able to reach Mars if well equipped.
We have to resign ourselves: we are forced to live on our planet, at least until we manage to find a way to migrate with a large part of the planet’s biodiversity, or until we are able to manufacture and breed it along, modifying ourselves accordingly to stay alive.
Since almost all of us live outside of forests, and do well, it seems possible that forests are not essential. We have seen that they occupy about 1/3 of the planet’s habitable land surface. The other 2/3 are deserts+rocks+perennial snow (30%), prairies+pastures+scrubland+agricultural land (65%) and cities (1%). Let’s assume that forests become agricultural land and/or pastures. What would that change for us?
It would change the biodiversity of the planet. In the largest primary forest in the Amazon, there are 427 species of mammals, more than 1300 species 378 species of reptiles, and more than 400 species of amphibians (www.amazonconservation.org - last access: 28 October 2023). This density of species is also found in other primary forests (1/3 of all forests). For comparison, throughout Italy (not only in its forests) we have about 123 species of mammals, 550 birds, 9150 plant taxa [32,33].
In total, the world’s forests contain 80% of all terrestrial species of animals, plants and insects [34]. If we lose this biodiversity, we will experience inconveniences similar to those encountered by astronauts in space with reduced biodiversity of their shuttle. 80% less biodiversity occurred around the middle of the Cretaceous, around 100 million years ago, when the thousands of genera were 1/5 of the current ones (Figure 2). Part of the biodiversity will take refuge in the remaining green areas, but that of the forests will disappear. Without this biodiversity we do not know what would happen to our species, since we are still ecologically connected to it.
To get an idea of the climate that could characterize an Earth without forests, and keeping in mind that the activity of living organisms affects the planet’s climate, we can hypothesize that the average global temperature and CO2 concentration could be similar to those of 100 million years ago, when biodiversity was 1/5 of that of today.
Tierney et al. 2020 [38] compared the air temperature and CO2 concentration forecasts of the IPCC scenarios with those of the period from the present to minus 100 million years ago. The worst IPCC scenario would correspond to the planet’s climate in the mid-Cretaceous, with an average air temperature 5°C higher than present and disastrous for humans.
If we consider biodiversity as a macro-system, in which all living are more or less connected to each other (like the cells of our brain and our body), in balance in what we call the biosphere (with us becoming the main actors of change ), and which governs the parameters essential to life on the planet (climate, quality of water and air, cycles of the elements, ocean currents and winds, ecological potential, number of living beings and all the vital processes linked to these essential parameters), with the loss of the forests, a large part of this biodiversity is lost, making the evolution of the planet unpredictable. There are no models capable of predicting what could happen to humanity if it were confronted with conditions of poor biodiversity. The fact that we are among the species born at the end of 3.7 billion years of evolution raises fears of a profound functional dependence on living things that existed long before us. Perhaps, the most pressing need is to learn much about the world of microorganisms [39,40,41,42]. The soil, main source of microorganisms, will be treated accordingly in chapter 3.
Science tells us that we cannot live without forests, we must find a way to live with those that remain [43]. As a precaution, any attempt to reduce the area of forests (now equal to two thirds or half of the original area) should be avoided. We don’t yet have the knowledge to try to manage a forest-free biosphere that includes humans. We do not know what and how much biodiversity is necessary to sustain the human species in the biosphere. From a practical point of view, this means that forests are a common good that must be safeguarded. It is necessary to think about a planetary management of this resource (as well as other related to it, water, air, climate, etc...). Maybe, the administrative division of the planet into independent and uncoordinated states no longer corresponds to the ecological needs of the planet. All citizens of the world should be governed by the same laws of mutual respect and regard for the planet’s resources. This is a utopian discourse, but without this progress, which still seems impossible, we will not emerge from the selective and adaptive processes underlying natural evolution.
1.4.1. What If “Forest Ecosystem Services” Didn’t Exist?
The word “services” is not appropriate. That the action of a forest existence and survival can also have beneficial consequences for humankind is acceptable and certainly true [44,45,46,47]. But that the forest should provide “services” to humans is an anthropocentric way of thinking that must be abandoned. The forest does not exist to provide services to humans, although it cannot do without them. The forest and humanity are called to coexist. It should not happen that the forest continues to decline so that humanity can grow. This is what has happened in the past.
We can make a conscious decision (weighing the economic consequences) that the remaining forest can continue to live as a forest and independently of the ecosystem services that it provides for humans. The forest is an autopoietic living system that is called by design to exist, to increase its complexity/biodiversity, to increase its efficiency in the use of resources, to contribute to the balance of the systems around it, to ensure sustainability to the Earth system which includes them all, in the conceptual scheme of Lovelock-Margulis and Gaia (Figure 3). In doing so, it responds to laws that Darwin [10] had interpreted as competition between living things and that Khakhina also brought back to the path of symbiosis [13,14,48]. Life as a whole is an unstoppable process and therefore human activities on living systems should necessarily consider the consequences on the inevitable movement of the living whole, which includes humanity.
That the forest might “want” something seems like a decidedly unscientific point of view. Perhaps it is easier to understand this concept if we consider our body as a living being: our body also “wants” something that is beyond our control, our body also wants to grow and age; we cannot stop growing; since the fertilization of the egg cell in the mother’s womb, <we all continue to grow, and there is no way to stop this intrinsic movement. Even death is nothing but a different growth with the transformation of matter in harmony with the universal movement: we will become mineral dust that will later be integrated into other growing bodies. Our body “wants something”, it wants to grow and “complexify”, increase its “harmonious and functional diversity”, it wants to reproduce and perpetuate life. We cannot stop this universal and intrinsic movement of the living [49]. A forest carries out a similar process on the scale of a larger system composed of numerous co-evolving living beings. A forest does not live to provide ecosystem services, but grows and reproduces, changes and evolves.
Humans belong to the world of the living and not vice versa. It was the forest system that generated humans who then began to live outside of it, until they succeeded in changing the evolution of the planet. However, what recently happened with Covid-19 should incite prudence and humility in the face of a universe that is still too large and unknown for us. The limits of what lives in the infinitely small (viruses and microorganisms) touch us closely, and correspond in the infinitely large to what our space telescopes see in the distance (primordial galaxies and giant black holes).
The forest (functional set of living things) needs what all living things (including humans) need. Therefore, it is mandatory that humans share space, air, water, biodiversity, nature’s products and the future on the same planet with the forest. These “elements” (space…future, planet) belong to an evolving whole, not just humans. Outside of this living whole that surrounds them, humans may not be able to exist. Many foresters/humans are thinking: “we are already trying to coexist with the forest; silviculture was born with the intention of perennializing forestry production” [50,51,52,53,54,55]. A question then arises spontaneously: “Is it true that we think that our future depends on that of other living beings and on the forest?”
1.4.1. Indisputable Facts
1) For reasons essentially related to food and the contingent needs of humanity, humans have taken over half of the area that once belonged to the forest to make cultivation fields and cities [26]. About 4 billion hectares of forest remain on the planet;
2) A quarter (about 1 billion ha) of the forest surface is still in the natural state of Old-growth forest (a forest that has reached a very advanced age without having undergone major disturbances by human activities - https://en.wikipedia.org/wiki/Old-growth_forest ) [56,57,58] and 20% of this can be classified as Intact forest landscape (forest area of at least 500 km2 without remote human action - https:// intactforests.org ) [59].
3) About half of the planet’s biodiversity is found in tropical forests. As their surface area has decreased, so has biodiversity. Overall estimated biodiversity losses outside forests in the period between 1970 and 2022 range from 18 to 94% depending on the region of the planet (https://livingplanet.panda.org) [60].
4) Human needs for wood will increase by 37% between now and 2050 (http://www.fao.org/documents/card/en/c/cc2265en ) [61].
5) In 2021 we lost another 3.75 million primary forests (https://research.wri.org/gfr/latest-analysis-deforestation-trends).
If we continue at this rate, we will lose all primary forests. Is that a big deal? If all forests were managed according to the rules of the latest close-to-nature [50], would this be a problem for the planet and for us humans? Why would soil-conscious forestry be more suitable for forest management than nature-friendly forestry? What does soil do that is important in practice that is not covered by close-to-nature silviculture?
2. The Soil
2.1. In the Future of the Living Forest, There Is a Dead Forest
To imagine the underground world of a forest, we suggest that you turn the trees upside down in your mind to give them two tips, one outside and one inside the ground, as shown in Figure 4. The forest doubled, as if there were a nice big one existed even under the feet. The new double forest is closer to reality than you might think. Physically it is not exactly like that, symmetrical as in the photo, but in reality, it is even more complex underfoot than outdoors, because the soil matrix is denser, richer and different from that of the air. The light phase of photosynthesis takes place in the air and outdoors. Without water, which comes from the soil, photosynthesis would also stop; the first step of the process is the splitting of water molecules to recover the energetic electron of hydrogen. The products of photosynthesis are transported everywhere, including in the roots and the soil. To bring light to the solar panel-like leaves, the tree must grow, and to do so it also needs the mineral elements that are present in the soil. These elements are not only those of the parent rock, but above all those of the parts continuously recycled by the tree itself. We are talking about wood, leaves, bark, bud perulae, flowers, bracts, cones, or exudates inside and outside the ground, but also about dead animals or parts of animals that live on the tree or their excrements. These are hundreds of grams per m2 per year that continuously “feed” the microorganisms and animals or fungi in the soil. In turn, these organisms die and are recycled, releasing the mineral elements that make up their tissues and molecules. In simple words, the soil is the belly of the forest, a belly fed by the living system of which it is a part.
Down: European forests ant their typical humipedons, from the left: high mountain forests in periodically dry climate and base-rich parent material with Tangel humus system; mountain, submountain forest with Amphi; low mountain, temperate plain forest on neutral substrate and Mull; mountain forest on acidic substrate and Moder, high mountain in cold climate, base-poor parent material and Mor.
Much of the forest is therefore under the forester’s feet. Water, energy, nutrients, biodiversity, forest production and renewal all depend on the soil. We know that it is useful to distinguish at least three layers (herbaceous, shrubby and arboreal) to be able to understand the dynamics of the vegetation. From a functional point of view, to better study its complexity, the soil can also be divided vertically into three sections (Figure 5):
1- The Humipedon, with the organic horizons OL, OF, OH, and the surface organo-mineral A horizons. Much of the soil’s biodiversity lives these horizons. These organisms evolve with the soil and are the key to understanding the litter recycling processes and the relationships between plants and soil;
2- the Copedon, in more developed soils, composed of newly formed mineral horizons E and B). In this layer, the processes of formation of new clays take place which migrate biologically and not in the profile and which can be related to the water cycle in the soil;
3- and the Lithopedon including the horizon of fragmented rock C and the superficial part of the rock more or less altered but still in place R.
A good knowledge of the soil is essential for forest management. Particular attention should be paid to the humipedon, whose relationships with the evolution of the forest are illustrated below.
2.2. Either It Is Alive, in Which Case It Is Really “Soil”, or It Is “Rock”
There are no living organisms on the moon (Figure 6). For this reason, soil like that of our planet has not formed on this natural satellite of ours. The lunar rock (identical to that of the earth’s crust) which is constantly exposed to the disaggregating action of the physical and chemical agents of space, the lunar rock (identical to that of the earth’s crust) has and continues to alter on the surface, remaining inert. On Earth, however, rock has been transformed into soil by living beings. You only have to imagine the process to realize the importance of this phenomenon. Living things did not emerge, as biologists have stated so far, but ecosystems did, first elementary and without walls, and then enclosed in even larger systems, composed of systems of systems. On figure 8 the plants do not appear, but rather the most obvious organisms involved in the process. We know that there are also animals and that the most important and numerous are the barely visible organisms such as microarthropods [62] or the invisible ones such such as microorganisms [63]
2.3. Humipedon and Forest Soil Functioning
The evolution of the soil is linked to the evolution of the living beings that inhabit it. To understand this phenomenon, it is necessary to carefully analyze the humipedon. In it, living beings build two types of structures that are extremely important for the functioning of the soil: the organic OH horizon and the organo-mineral A horizon. They are functionally related structures: when one increases, the other decreases, and vice versa. The OH thickness increases under difficult environmental conditions. Essentially these are low temperatures, drought, frost, or harmful/polluted/asphyxiated substrate for living beings. The A horizon, on the other hand, increases in thickness (to the detriment of the OH horizon if present, which disappears) when conditions are favorable for the development of life in the soil, such as water and temperature in the right doses, digestible and transformable organic matter. in order to understand the humipedon, it is therefore necessary to look at the profile and keep an eye on these two diagnostic horizons (Figure 7 and Figure 9). In general, they are produced by different pedofauna and microorganisms: the OH horizon is the result of arthropods, enchytraeids and epigeal earthworms; A is instead produced by endogeous and anecic earthworms. There are two easy to find exceptions: 1) when conditions in the soil are difficult for living things, the OH horizons are almost devoid of animals and the biodegradation of the organic substrate is carried out by fungi. The resulting horizon is a nozOF (a non-zoogenic OF horizon matted with fungal hyphae; the OF is not transformed into OH as usual because it is not eaten by animals but digested by fungi); 2) earthworms periodically abandon periodically dry environments, leaving the organic matter to be ingested and transformed by arthropods; these are ants or termites, or insect larvae that can produce an A horizon morpho-functionally similar to that of endogean and anecic earthworms (savannas, steppes, scrublands and Mediterranean forests).
Figure 7 illustrates the dynamics of these horizons in an Alpine spruce forest environment. The phenomenon was studied for the first time in France [64,65] and then ecologically circumscribed and confirmed in Italy [66]. The opening of the forest draws light and water at the soil surface; large earthworms arrive and, feed in the OH horizon expelling organo-mineral droppings. As the trees grow, they deplete the soil of nutrients and water, the earthworms retreat and the OH horizon reforms.
When trees are young, they feed on soil resources. When the trees are adults and vigorous, they can return nutrients to the soil. Finally, when the trees are old, they fall and feed the soil with all their mass [67]. In general, the soil releases nutrients in the first stages of the life of a forest, then gradually rebuilt its capital immobilizing nutrients in shallow organic layers. And towards the end, it surpasses in nutrients to fuel the natural renewal of the forest and start the cycle again. The duration of the periods of depletion, restoration and preparation for renewal change with the tree species and the forest environment.
In addition to the stage of the silvogenetic cycle, this alternation of soil horizons depends on the parent rock and the climatic conditions, which we know are particularly linked to altitude [68]. This last aspect was carefully studied by Bernier [69] (Figure 8).
In a forest, the thickness of the superficial organic (OL, OF and OH) and organo-mineral (A) horizons change in space and time:
1) when the climate becomes harsher for soil organisms, or when the soil becomes nutrient-poor during the growth of young trees, the biodegradation of the organic horizons slows down. The organic horizons are fed from above (loss by young trees of the leaves of the low branches in the shade) and are also digested more slowly by the soil fauna. At the same time, the organo-mineral horizon A loses the entry of nutrients, which instead escape more rapidly because nourishing the roots of the rapidly growing trees; the A horizon becomes lighter in color and its glomerular structure breaks down and gives way to particular or massive structure;
2) at the end of the forest cycle when the canopy opens, sun, water, fauna arrive and the organic horizons can be eaten to become the organo-mineral horizon A; this horizon regains its glomerular structure and its storage capacity to store water and nutritive elements for the plants.
These two situations of the humipedon are called Moder and Mull humus systems (Figure 11). Named Amphi, a third system also develops between the two [70]; called humus “gemellare” (twin humus) by Franz Hartman [71] and Amphimus in France [72], also known in Germany [73], this humus system helps to understand how forest soil works, with interdependent and alternating organic and organo-mineral horizons.
Forests in extremely cold and acidic environments have humipedons that are particularly rich in organic horizons and poor in pedofauna; the humus system is a Mor. Forests of extremely calcareous and dry environments have humipedons that are rich in organic horizons but in which a relatively active fauna is present in favorable periods; such a humus system is called Tangel. Even in these extreme environments the soil changes with the age of the trees but the renewal is not linked to the transformation of the organic horizons by earthworms but can involve a larger number of fungi and animals that feed on wood in humus systems called Ligno (Figure 9 below, left and right corners).
Humipedon changes coinciding with the spruce forest stages studied in France were also observed in Trentino (Italy). A doctoral thesis was developed to study the physicochemical causes that led to soil modifications during the development stages of the alpine spruce forest [66] and recently published [74]. Summarized in Figure 10, the general characteristics reported on the plane of the two axes of the multivariate analysis are, from left to right, a decrease in temperature and an increase in soil moisture; from bottom to top we read an increase in pH and the thickness of the A horizons, a decrease in total organic carbon and the thickness of the organic horizons. Acid-North and Acid-South separate well for these characters and along the diagonal that divides the plane in two from the point on the upper left (South exposure) and on the lower right (North). Formations on basic substrate separate on the horizontal axis, along a soil moisture gradient. In the 4 series, we observe an evolution of the cyclic humipedon, consistent with what was illustrated in the previous figures (Figure 8 and Figure 9), with an increase in organic layers in the stages of rapid growth of the forest and a return towards an environment inhabited by earthworms in the opening stages and the first period of forest renewal.
In the herbaceous opening (yellow background) we find a Mull three times and only in the most difficult condition (AN = Acidic North) an Amphi. In the two basic ones, the second stage of renewal (green background) is still characterized by the Mull, while in the acidic environment we find an Amphi to the south and a Moder to the north. The third stage is the critical one (orange background), in which the soil empties itself of its nutritive elements and water, giving them up to the rapidly growing trees, it is Amphi on basic and Moder or Mor on acid. The fourth (blue background), which in our forests rarely exceeds 200 years, is a Moder everywhere except in the best condition of BS (Basic Sud).
What can be seen in Figure 10 regarding the inversely correlated determinants reported on opposite axes (two opposite horizontal and two vertical), could be interpreted as general trends in the functioning of the soil in high mountain Alpine spruce forest:
- -
- the thicknesses of meA and maA (from earthworms) oppose the thicknesses of OF, OH and miA from arthropods;
- -
- the temperature at 10 cm depth opposes the humidity of the soil;
- -
- the total organic carbon of the soil increases under conditions of difficult bio-degradation of the litter.
From a general point of view, we can note two phases can be identified in the forest soil of an Alpine spruce forest (Figure 11): a) a period of “extraction” of energy and matter to nourish the future of the forest (first 150 years) = the autotrophic phase [67]; b) a period of storage of matter and energy necessary to replenish soil reserves and prepare new foundations for the future generation (second 150 years) = the heterotrophic phase. In addition, the renewal of spruce appears to be favored by wood in the final stages of decomposition, at least in non-megaforbiae areas [75]. The role of the mosses layer has also been discussed for some time, as it seems to favor regeneration in the long term but not in the short term [76]. Generally, the circumscribed humus system that develops around rotting wood (limited to the stump, branch or root transforming in the soil) is called “Ligno”; the foam system is instead called Bryo [77,78]. Bryo and Ligno can develop as a single system directly on the rock, or even on top of each other (Bryo on Ligno), or superimpose on Mull and Moder. It is important to note that the humus systems succeed each other in time in the forest stages; that the Mull system develops in clearings and under older trees, while the Moder develops in the intermediate stages. The Bryo and Ligno systems develop on Mull (at the beginning and end of the cycle) and contribute favorably to the success of forest renewal. In contrast, Moder formation (intermediate stages of a non-overmature forest) slow down and prevent renewal (while still protecting the seeds?). The rotation shifts of forest production depend on the cultivated species. However, it rarely goes over 100-150 years in Europe. A part of the necessary return on investments in the soil is thus lost in this way [67]. Leaving aside the other compartments for simplification, let’s take dead wood as an example of return to the soil. Ultimately, this mass corresponds to an enormous quantity of energy coming from the sun via the autotrophic compartment. It is made available to heterotrophs with the creation of new trophic chains from the activities of saproxylophagous fungi and countless categories of organisms which develop on dead wood. It’s a kind of little new forest in the big one. The same internal complexation process occurs each time organic matter returns to the soil. And the pyramids thus formed all return to the ground.
3. A Soil-Conscious Forestry
3.1. Autopoiesis
Autopoiesis is the property that Maturana and Valera [81] attributed to living systems in 1980, regardless of scale. This is a very interesting concept for foresters because it explains well what a living system is. It is an entity that is capable of preparing the building blocks needed for its construction and that manages to re-produce itself over time, adapting to the environment in which the vital process occurs. When the living beings that inhabit a forest die, they are recycled in the soil, to reuse part of their remains in the form of ions or more or less complex molecules reused at the level of living organisms in and out of the soil.
A forest that turns and grows, like a wheel that recycles more and more of the material it is made of, to reinvest them in a new and more complex forest. Growth is partially reinvested in the soil, which acts as an investment bank for the forest system (Figure 11).
3.2. Foundations of Practical Action Soil-Conscious Forestry
A managed forest ecosystem corresponds to a forest landscape (the set of forest parcels of a single potential phytocoenosis) that can be programmed for a complete forest cycle, in compliance with a prefigured potential evolution. This forest ecosystem is called a forest type [82,83]. The forest type contains a relatively homogeneous environment in terms of biological, geopedological and climatic characteristics, delimited by man for forest management reasons.
In general, forest planning has always been based on a prior study of the environment. This involves defining the historical framework of past interventions, the climatic, geological, pedological and vegetation environment, with the delineation of forest types (in Italy for example [84,85,86]). Each of these is classically divided into forest parcels to proceed with the inventory of the soil and topsoil, with detection of the structure, age, height and distribution of trees by species in each parcel (even recent historical studies like these [83,87,88,89,90,91,92]). Development plans now include an important chapter on wildlife. For some time now there has been a reference in Italy to the rise of phytoclimatic zones due to ongoing climate change [93]. For an article dedicated to the importance of the uneven-aged structure of the forest so supported by Susmel [94], especially when aiming at the conservation of biodiversity, we refer to the recent work by Savilaakso et al. 2021 [95].
Forestry that pays attention to the soil should include an inventory of humipedons by structural type, and an inventory of the animal biodiversity of the soil. Studies on microorganisms would also be desirable, since the decomposition process depends on the relationship between known categories of fungi and bacteria [96] and on the soil type [97]. Below are some points to consider for the formulation of a soil-conscious forestry, i.e. a silvicultural technique that takes into account the functional potential of the soil:
3.2.1. The Renewal of the Forest
The first main question that the forester asks himself is generally this: what is the natural evolution of the forest on which I am called to intervene? Where is it going, is it changing? To search for signs of forest natural renewal helps to answer this question. In order to survive, the forest system (plants+animals+microorganisms of that environment) must renew itself. The signs of renewal correspond to areas that have a humipedon, i.e. the sequence of organic (OL, OF, OH) and organo-mineral (A) horizons at the surface (first 30 cm) of the soil. The humipedon is different in regeneration sites compare to that found in the areas covered by intermediate age and non-perishable trees [70,98]. It is necessary to study the humipedon in clearings, under old decaying trees with more or less open foliage, near stumps or decaying wood and in all uncovered areas with herbaceous/shrub vegetation which often contrasts with that built under the areas covered by intermediate age and non-perishable trees. The humipedon changes in the areas where the forest prepares its rebirth. Recent studies have partially clarified the role of light, temperature and also herbaceous cover on seed development [96]. The size of the canopy opening appears to play a key role. There are even-aged forests that are renewed due to catastrophic events (wind, avalanches, fire). In these cases, the change of the humipedon occurs a posteriori throughout the area opened by the incident.
If there are no signs of renewal, it is necessary to find the causes of this lack by studying the ecology of the forest. The main characteristics of a healthy and balanced forest are the presence of all the stages of the forest (renewal, growth, stasis and decay), occupying the surface of the whole in due proportions and all with their own specific humipedon, and a biodiversity correlated with the stages of the forest. All this can be verified through phytosociological surveys (list of plant species by layers), census of wild fauna and pedological surveys (recognition of systems of humus and, if necessary, list of soil animals and microorganisms), in the different stages of the forest. If a forest has a balanced distribution of forest stages, then it will also show an adequate renewal stage; however, if this stage is absent, the reasons for this lack may be related to the imbalance of the system, in terms of specific composition, excessive homogeneity of the age and structure of the forest, the absence or deficit of light on the ground and/or or decaying woody material. In a forest that lacks natural renewal it is very likely that the older stage of decay does not exist, or is not advanced enough; it is then necessary to recreate the conditions for its progressive and protected aging, in order to lead it over time to the next stage of renewal. The mistake of the silvicultural past was not to recognize the importance of this stage of forest aging. It corresponds to the stage of reconstitution of the forest floor, from which the renewal of the forest ecosystem begins. At this stage, the soil is biologically and physic-chemically the most favorable for seed germination.
3.2.2. Supporting the Becoming of the Forest
Each type of forest has its own stage of regeneration characterized by typical vegetation and humipedon. The study of the specific composition of the vegetation and the characteristics of the humipedon of the aging stage (deterioration of the topsoil and revitalization of the soil) of the forest provides valuable indications on the evolution of the system. By knowing the trend of climate change and the phytoclimatic bands of each region, the forester will be able to interpret the dynamics observed and glimpse the future of the forest. Aware of the difficulty of the long-term predictions required by forestry, the forester will adopt a precautionary attitude: he will intervene cautiously (with light cuts) in order to accommodate the movement, he has perceived in the forest and will periodically check the result of his action. If the result is an increase in the vigor of renewal and in the state of the forest in general, the forester will continue this line of treatments (for example cuts in the oldest stage of the forest); if the evolution of the forest is not as expected, he will carry out new studies to understand the reaction of the forest, and on new bases he will try a different intervention capable of better accommodating the recent, even if unexpected, movement of the forest (widening the holes, starting from the margins, not cutting species that you thought were invasive...).
3.2.3. Forest Biodiversity
Respect for the evolution of the forest concerns above all its biodiversity. Even the microbial one [99]. Biodiversity should be understood as the functional biological whole of the forest [100,101]. It is well known that old-growth forests contain old trees of maximum height for each species, and that the highest levels of biodiversity are found in these forests [52,56,95,102,103,104]. It is also true that the richest in terms of biodiversity are the humid tropical forests, where radiant energy and water are never lacking [24] and where the trees occupy multiple layers and reach impressive heights. It is therefore likely that the stature of the forest (height of the dominant trees) is related to the biodiversity of each formation [100], in accordance with the climatic region in which the trees grow. From this point of view, Lucio Susmel’s formulas which relates height to other parameters of uneven-aged forests also have an ecological meaning that should be re-evaluated [94].
3.2.4. Exotic Species
Out of respect for the original forest, the forester should not introduce new species. If species over than from the local ones have arrived in the forest, or have been introduced in the past, the forester takes care of the endemic species so that they can resist the entry of the new species. If the new species manage to replace the local ones, the forester will support the formation of the new ecosystem.
This applies to all species both plants or animals. When the bark beetle kills spruce forests, it means that this species that became food for the bark beetle was no longer in the natural balance of the system. If the loss of this species represents a significant economic damage to the community, it will be necessary to intervene with the awareness that in the long term the replacement of the attacked species with a new species in equilibrium in the new environment must be envisaged. Similarly, if the pressure of ungulates prevents the renewal of the forest, it is important to promote the reintroduction of natural predators to restore the forest to its lost level of complexity. Functional biodiversity is the best way to guarantee the homeostasis of the system and to combat the arrival of exotic species and diseases.
3.2.5. The Growth of the Forest
The growth of the forest is continuous. Because the trees that make it up never stop growing [105]. For the future of the forest, what matters is the balance of the entire forest for all the mineral elements that compose it and in terms of biodiversity [106]. Those that remain in the forest as mineral elements useful for the development of life are contained in the living and dead organic matter [100]. An ecosystem is in a state of growth when the dead organic substance is greater than the live one, since it is in the dead organic matter that all the living beings in the ecosystem fish to grow. Since there are irremediably losses in the transition from dead to living organic matter (second principle of thermodynamics), the amount of dead matter is always greater than the amount of living matter. The growth of a living system depends on its biodiversity: if it continues to grow, it ensures the evolution and health of the whole [107]. It is not possible to stop the growth of the forest as a whole except by reducing its functional biodiversity and its necromass.
It is known that in old forests, 2/3 of organic carbon is found in the soil [108] (see also in French, “Parutions”, “Carbone forestiers en movement”, in: http://refora.online.fr/ ). We have not addressed the issue of carbon storage in forest soil because there is still a lack of a real connection between the quality of the humipedon (humus system) and the quantity and quality of the stored carbon. An Eumull can contain as much carbon as a Dysmoder, but the molecules in which the carbon is fixed are very different: in organo-mineral excrements from earthworms in the first case, in slightly decomposed organic substance and very organic excrements of enchytraeids and arthropods in the other.
3.2.6. Human Economy and Harvesting
From an economic point of view, the forester must decide what percentage of current growth that he can take from the forest to meet the needs of the human populations that depends on this resource. The lower that percentage, the better for the forest. It is not true that the forest would die, or burn, or become diseased without human intervention. This is a big misunderstanding that only applies to such artificial forests that they have lost even their semi-natural balance. There is no “right” intervention measure; there is only a withdrawal that interferes as little as possible with the growth of the forest. Withdrawal greater than the current average growth will inevitably leads to a decrease in biomass and a loss of biodiversity in the long term (biodiversity feeds on and is therefore correlated with the primary production of the forest). The removal of biomass (unless it is infected biomass that can reinfect the forest) corresponds to a loss of resource for the forest. The reduction in the number of species in the forest is also harmful to the balance of the forest; the change in the percentage of tree species which tends to favor the woody species more suitable for the human market is a mistake if practiced on a large scale and in the long term.
The entire cycle of the forest must be kept active at all its stages. And with all species. The forester wishes to keep the entire forest intact as possible, respecting “reasonable” human needs, to be discussed from time to time, depending on the regions in which one intervenes, and taking into account the global, social and long-term needs of the forest. The forester should only carry out only cautious “thinning” in all stages of the forest, with the aim of not changing the structural and specific composition of the forest and aware that the more he cuts and removes, the more he will influence the future of the forest, and the more difficult it will be to predict the future of the forest [109,110,111,112,113].
The key points of a silvicultural technique that gives importance to the soil are:
- -
- minimum harvesting: cutting and removing the minimum possible (instead of removing the maximum while respecting the vitality of the forest);
- -
- the maximum prolongation of the life of the trees: maintaining even the decaying stage of the forest (instead of interrupting the cycle of production in the moment of stasis of growth to obtain a high productivity – which, unfortunately, as we know is inevitably short-term, as happened with the depletion of agricultural land due to intensive agriculture);
- -
- respect for the structure that the forest has and will have in the future: careful thinning at all stages of the forest (instead of thinning to plan the growth and final cutting of a mature stand);
- -
- -
- replenishment tor energy reserves and mineral elements in the forest soil: it is recognized that the decline stage of the forest floor must be maintained because it ensures the reconstruction of the soil’s nutrient capital of the soil.
3.2.2. Examples in Alpine Environment
On page 55 of the Technical Lines for Corporate Forestry Planning of the Province of Trento (https://forestefauna.provincia.tn.it/content/download/13091/233986/file/linee%20tecniche_27_12_16.pdf ) [114], in the chapter dedicated to ’Thematic inventory and the description of the forests, the point of view of an eminent forester on the duration of the life cycle of a forest is reported (Thomasius 1996) [115]:
“Just as ecologically oriented silviculture distances itself from the spatial ordering of schematic cuts (cultivation ordering) and moves towards a subdivision of the forest based on natural-type spatial units, it must also move away from the mechanistic categories of the typical temporal ordering of the clear-cut forest and move towards biological time scales. In relation to this, it must be observed that the different stages of development of organisms (juvenile, growth, maturity and articulation stages), which are to be considered temporal indicators of their life course, are reached at different times depending on the biotope (faster in case of good fertility, less in case of poor fertility) of the tree species (earlier by pioneer species, later by definitive species) and by the environment (first by dominant species and later by dominated species) “.
To attempt to associate these principles with a practical case, let’s take the easily imaginable example of a mixed Alpine fir forest (fir, spruce and beech), montane, with an irregular structure and a rotation of 150-200 years, which we want to normalize. Today there are very sophisticated tools and techniques for both inventory and forecasting harvesting. For the purpose of this article, the method that was once taught (Prof. Mario Cappelli in Padua, for example) [116], with the real and normal curves and the humps of the first to be cut to “normalize” the structure, is hasty but gives a good idea of how a forester works in practice on the forest with a chainsaw (Figure 12 and Figure 13).
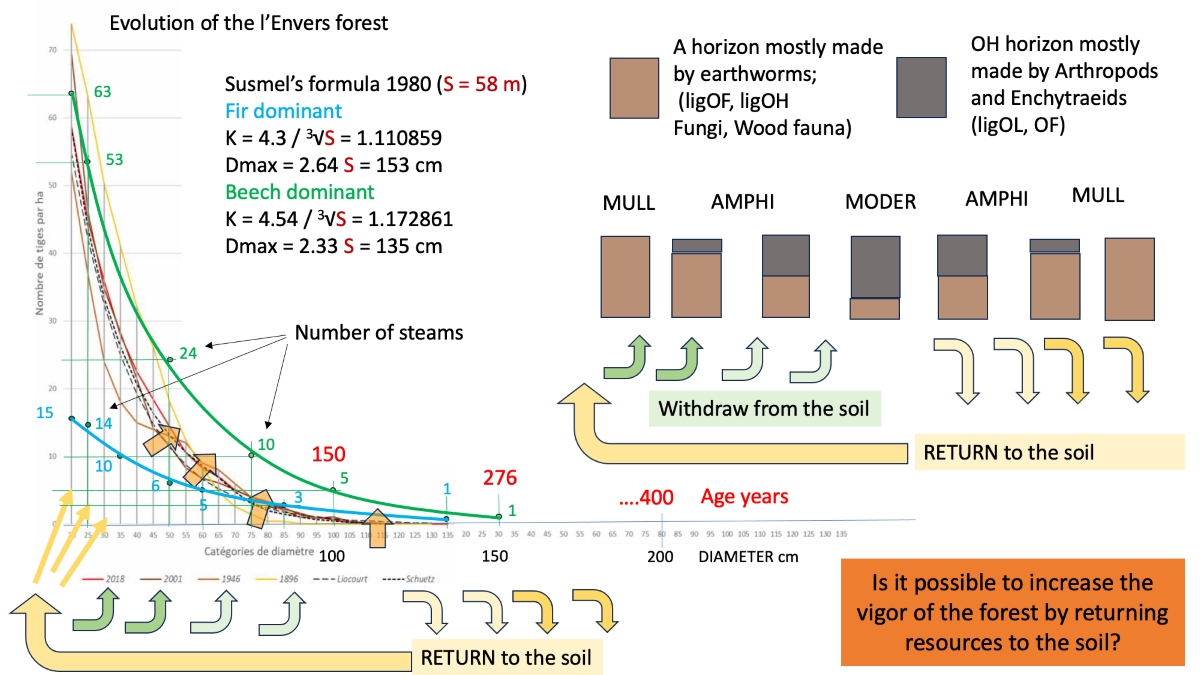
The universal law of the forest, and common to many living systems, is that an adult tree produces a population of many young trees. These new individuals decrease in number over the course of their lives, until they reach the single, last dying, tree of their generation. When that tree falls, the story of a new generation begins again. On the curves of figure 14 the phenomenon can be seen by moving along the x-axis from left to right (traditionally, foresters count the trees starting from the 20 cm diameter class; the part missing from the graph, those with diameters < 17.5 cm, would count in dozens of thousands of trees per hectare, a number that is drastically reduced in the first 50 years of the forest’s life). Darwin’s theory of evolution is based on this evidence [10]: over time, the trees best adapted to the environment in which they evolve (and which changes with them) will survive. The individuals that know how to best cooperate with each other and with the other species in the system to use of available resources survive [117]. Pro-Silva Helvetica offers real models of uneven-aged forests online. The first of these is the forest of “l’Envers à Couvet/NE” (https://www.pro-silva-helvetica.ch/pdf/Portrait01_f.pdf), with 70-80 trees/ha with a diameter of 20 cm which ending in a single tree with a diameter of 150 cm, 58 m high and 276 years old.
In order to mimic intervention, we propose a more irregular and more frequent structure when sliding upwards in formations with dominant spruce, defined as “multiplane” in the Forestry Typologies [118]. Due to climate warming, these spruce forests will experience the entry of broad-leaved trees and silver firs from below, as a consequence of the upward movement of the vegetation bands corresponding to the beech forest. Moving the structure of spruce forests towards a diameter’s distribution closer to that of mixed fir forests, similar to that of Pro Sylva mentioned above, could be a step that many foresters could share. Figure 1 shows two identical distributions of the number of trees in the forest stages were followed. In figure 13, however, the two curves have been grouped together to form a single cycle of double duration. The grouping assumes that the forest not subjected to a silvicultural regime can have a shift that lasts twice as long as the current one.
The nutrients are found in different “reservoirs”: the A horizon of the Mull system (macroaggregates of organo-mineral substance produced by earthworms); the organic OH horizon (microaggregates and minute particles of organic substance produced by enchytraeids and arthropods) of the Moder system; the decomposing wood of the Ligno system (ligOF and ligOH horizons) [77]. The groups of animals in each system associate the humus systems with the dynamics of the forest, allowing the release or fixation of nutrients at the right time. The A horizon is ready to release the ions that bind the organic and mineral substance in the macro-aggregates; the OH horizon is instead made up of blocked organic matter awaiting to be processed. It can be transformed into the A horizon by earthworms in a favorable environment for such animals. Dead wood is also very important in the forest [119]. The transformation of wood into aggregates rich in nutrients available for plants is due to a specific fauna and fungi [78,120,121], that transform branches, trunks and stumps into spongy horizons that release nutrients for the roots of the plants. The humipedon evolves with the stages of the forest in this way:
- -
- in the senescent stage and in the early juvenile stages, the A horizon of the Mull system (earthworms) and the ligOF and ligOH horizons of the Ligno system (fungi and wood fauna) develop;
- -
- in the intermediate stages the OH horizon increases in thickness at the expense of an A horizon which tends to disappear (from a thick A from earthworms in the Amphi of the first stages of development of the tall forest, to a thin A not from earthworms in the Moder of the dense perches and in the adult tall forest still growing strongly;
- -
- in the adult stages of relative stability, a redistribution of nutrients takes place outside and inside the soil, with a consequent decrease in the thickness of the OH horizon, storage of nutrient elements in the soil in the form of macroaggregates of the A horizon, or in any case of more organic substance easily recyclable in dead wood and partially biodegraded.
All these stages, the allocation of the resources produced by photosynthesis evolves accordingly and should be studied in more detail. It is known that trees invest more in trunks than in branches or leaves as they age [122]; it is also known that they grow less tall once they reach sexual maturity [123]; and we are also beginning to understand the process of rhizodeposition and estimate its importance in the carbon cycle in the storage of this element in the form of organic molecules of different complexity in the soil [124]; it has been found that such deposition is associated with selected soil microorganisms [125], and that these microorganisms are often symbionts of plant roots that activate a new process called rhizophagy [126]. I would not be surprised to read that different qualities of soil organic matter and microbiomes correspond to forest humus systems and their horizons; Rendzina in different environments are different plant-soil systems and produce humic acids with different molecular weight and functionality [127].
Regarding the productivity of mixed uneven-aged forests such as that of l’Envers, we read on page 7 of their factsheet (https://www.pro-silva-helvetica.ch/pdf/Portrait01_f.pdf) that from 1974 to 2008 (35 years), the volume of the forest measured in 1975 has been removed. This means a stand renewal rate of 35 years. On page 12 we read that this corresponds to a huge cutting rate of 10 m3 per hectare per year (1000 m3/100 years).
It seems possible to remove in 35 years what a forest produces in a cycle of a few hundred years. Even if the numbers look good on paper, there is probably something missing in terms of how the system works. We took the liberty of taking the figure on page 4 which corresponds to the evolution of the distribution curve of the stems in 1896, 1946, 2001 and 2018 (figure 16). The graph shows two new distribution curves of the number of stems with age, using two growth coefficients (K) and two maximum diameters (Dmax) obtained with the Susmel7 formulas starting from the height, for two high forests in the mountainous area of the Abieti-Fagetum, one dominated by fir (blue line) and one by beech (green line). In both cases, the height of the tallest tree in l’Envers (a 58 m silver fir) was taken as the height. The values calculated with the formulas of Susmel 1980 (for S = 58 m) are the following:
- with fir dominant: K = 4.3 / 3√S = 1.110859; Dmax = 2.64 S = 153 cm;
- with beech dominant: K = 4.54 / 3√S = 1.172861; Dmax = 2.33 S = 135 cm.
By multiplying the coefficients, the maximum is 150 cm, while with beech dominance it is 135 cm.
Susmel’s formulas were constructed on the basis of measurements in Italian relict nuclei and primary (virgin) forests in Croatia [94,128,129,130,131]. In the spirit of Susmel they should serve as a potential ecological model for Italian alpine forests. The l’Envers forest lies between the two curves of Susmel.
It is interesting to observe that the formation of dominant fir with beech delimits the model downwards, while that of dominant beech with fir places an upper limit (figure 14). From an ecological point of view, this means that with continued climate warming and the rise of the beech belt, we must expect a shift upwards and to the right of the real curves.
The Susmel curves are already shifted further to the right on the graph, and deviate in a curious way from the real ones, as if the young stages were closer to those of the beech forest and the old ones closer to those of the fir forest. Another puzzling is the distance of the dominant beech curve from those of the real distribution: it is as if medium and large sized trees were missing. These are probably the trees removed by foresters with production cuts.
Figure 14 also shows a cycle of returning nutrients to the soil. The balance between extraction and return is in favour of the former in the juvenile stages and instead becomes in favour of the latter in the final part of the cycle. Extending the shift could allow more nutrients to be returned to the forest by making the distribution curves evolve upwards and to the right, as indicated by the orange arrows in figure 16. The increase in necromass due to the extension of the shift would eventually feed the renewal, which could return to the historical values of 1896. At the bottom of page 12 of the report on the forest of l’Envers we find written: 1000 cubic meters of wood cut per hectare in a hundred years...not to mention that the most important thing is not what we harvest, but what we leave. Couvet, division 109, never 2009).
Always in the spirit of Pro-Silva, on page 19 of the description of the uneven-aged forest of Tscheppa Verda (https://www.pro-silva-helvetica.ch/pdf/Portrait15_f.pdf ), under the picture of a beautiful specimen of Silver fir, we can read an observation by Walter Trepp dated 26.8.1986: “S-chanf, Tscheppawald, divisions 48/50. Typical garden forest spruce, with an extraordinarily long crown. Is this tree sick? I bet not! Every year, at the end of fall, part of the needles turns yellow. This is how tree get rid of the dead needles. Trees with wide crowns - typical of garden forests - produce more dry needles than trees with narrow crowns in regular high forests. The “scientists” have obviously not yet integrated these observations!”
This is probably a way of restoring the nutrient potential of the soil which needs to be re-evaluated in silvicultural terms, allowing the trees to age beyond what seems to us economically invalid.
Old trees produce more seeds (more pine cones or more fruit) [132] than younger ones, as if in the last period of their life they “desired” to pass on and distribute the maximum of their genetic potential as an inheritance.
3.2.3. Planet Forest Management
From the point of view of forest management on a planetary scale, we know that even primary forests are affected by anthropogenic climate change and therefore can no longer be considered strictly...primary.
However, we believe that the time has come to decide at a global level (FAO?) to prevent the direct exploitation of a percentage of these forests (for example today’s 20% of primary forests called “Intact Forest Landscape”), as if these forests they were an inviolable asset of Planet Earth. For these “untouched” forests, an income could be established equal to that which would result from their hypothetical naturalistic exploitation. This sum would be paid to the countries where these forests grow, in order to preserve them as an asset of the Planet and of humanity.
We believe that soil-sensitive forestry should be applied to all the other forests in the world since it is the only one capable of guaranteeing the general conservation of the planet’s biodiversity.
3.2.4. Soil-Conscious Forestry and New Forestry Plantings in Climate Change
At the level of the current phytocoenosis, the plant species of this phytocoenosis but also species of the lower phytoclimatic band, so that the system can follow the ongoing climate warming, gradually losing the less suitable species. In the plains and in the low mountains on the ground in the areas where the dry period of the year is prolonged, an OH horizon appears above the A horizon with a consequent shift of the Mull system towards the Amphi; in the mountains or in a more continental environment, when the rainfall is good, the transformation of the Moder into Amphi and Mull is observed, with the reduction in thickness or disappearance of the OH horizon, and the integration of its organic matter content by the earthworms in an underlying A horizon. This would mean that in environments that tend to become drier, the earthworm fauna gives way to that of the arthropods; in the mountains, however, where there is no lack of water, the arthropods give way to the earthworms. Obviously, the change in the soil will soon correspond to the arrival of different plant species. These are personal observations widespread in France and Italy that should be confirmed by scientific investigations.
3.2.5. Coppice Forest, Wood Arboriculture, Urban Parks
From the point of view of a forestry that pays attention to the soil, coppice, wood arboriculture and urban parks are anthropized systems that are similar to agroforestry. These are ecosystems that belong to anthropic evolutionary lines and create new biodiversity. Although interesting from a biological point of view, these recent populations which are recent in terms of evolution do not have a forestry function and do not enjoy autonomy. If they are located in phytoclimatic zones that allow the complexity of ecosystems to grow up to the climatic stage, and if they are left to evolve without human intervention, these systems become new forests, hybridizing with existing ones. From an evolutionary point of view, the coppice is a mutilated forest: humans have decided to bring the foliage to the ground, removing the trunks of all the trees. By doing so, they limited the forest’s biospace, dramatically lowering its potential stature; they then reduced the shift to a minimum for economic reasons, linked to the use of wood as a source of energy; they have often reduced the specific tree composition making it monospecific, as has been done in agriculture. In wood arboriculture the species are selected by man to maximize wood production; a system layout is used that facilitates mechanization; even if polycyclic, with the presence of shrubby species, these populations have nothing to do with real forests.
The coppice has a long history of coexistence with man. In the Italian mountain regions (which is almost the entire country) these woods have played a fundamental role in the development of human societies, substantially supporting the economy with everything that goes with it. For this reason, these forests have also been well regulated so as not to be lost. With the oil revolution they experienced widespread abandonment, triggering a series of ecological dynamics more complex than assumed in the past, and today much studied and debated. To date, depending on the socio-economic developments of the last decade, these formations have seen their managerial restoration in various areas. These formations are now used again mainly the production of firewood and poles, while the inland populations are rediscovering and adapting uses (craftsmanship, family-scale woodworking industries) that were thought to be forgotten forever. There is a biodiversity linked to the coppice forest [133,134], just as there is that linked to the mowing of secondary prairies, often subject to protection at community level. As with the latter, the coppice forest has also characterized the traditional landscape [135,136] and is at the basis of ecological, landscape aspects, traditions, uses and customs and like these it should therefore be safeguarded [133,137,138].
Urban forests are often exotic trees, juxtaposed with criteria linked to the functional aesthetics of cities, interspersed with meadows which are also very simplified and linked to human recreational activities. Their management is entirely dictated by principles of safety and practicality that are not those applied in the forest. These “tree systems” are human deviations and manipulations, practical and convenient from an economic and social point of view, but they are not forests. They will evolve along with the rest, providing the planet with new anthropogenic plant covers.
The soil is also different from the natural one, and resembling agricultural soil, often tilled before planting, or urban soil often containing the remains of buildings and debris of all kinds. The dynamics of living beings try to return these soils back to a more natural stratification. Being in the plains or in the city, the humipedons very often tend to become earthworm Mulls, especially when irrigated.
3.2.6. Agroforestry
From the perspective of soil-conscious forestry, we propose to considering agroforestry as territory lost to the real forest. Agroforestry areas are hybrids between forest and agriculture and do not respect the laws of the natural forest. With agroforestry, agriculture is brought into the forest (or vice versa in the case of forestry revaluation of areas abandoned by agriculture). The focus is on some human consumption. It is not honest to classify these areas as forests. If the biodiversity of the forest is protected, the area dedicated to agroforestry cannot also be included in the “forest” category. Agroforestry creates a new clade of natural evolution, which may be economically interesting, but which must be scientifically separated from forest biodiversity. A salami made with pork from free-range pigs in the forest is obviously tastier and healthier than the one produced with pork from confined and in battery-fed pigs. Is it possible to create forest-like environments for these animals, starting from non-forest environments?
4. Conclusions
Soil-conscious forestry is the application of forest sciences research to principles that at first glance seem to contradict those of the past: as little interventions as long a cutting cycle as possible. In reality, if we have come to these resolutions, it is because things are not going as hoped: if we continue as before, the forests will disappear. Perhaps we do not yet have the knowledge necessary to manage natural ecosystems in the long term. Since the future of humanity depends on the health of the forest and on the ability of human to coexist with the greatest possible number of living beings on the planet, it is good to make a conscious decision not to touch a good part of the planet’s forests. Within the limits of a worthy coexistence, with enough food and comfort for humans as well... is that possible?
Soil-centered forestry means helping all forests to evolve towards a stage very close to “old growth-forest”.
The metaphor of the forbidden apple applies to aspects of forestry that are not obvious. They deserve the full attention of science: soil reserves and biodiversity have limits that must not be exceeded.
Soil cannot be exploited beyond a certain point. It’s not just a matter of quantity. Soil also recycles the DNA of the ecosystem, which is the DNA of all living biota that inhabit that volume of the ecosphere. It is from the ground that the life cycle of the forest begins again. Biodiversity is not only the number of species, but also the functional web of relationships between species: when an ecosystem is destroyed, the network of communication and exchange that kept it alive is also lost. When parents or grandparents are lost, the foundations on which the family rested are lost. The feelings that have been passed down to us, which work with the physical genes that are also the result of inheritance, and which constantly build living bodies and make them evolve, remain alive. All the components of a human society are necessary for the good functioning of the whole. The same is true for the forest. In order to live and evolve, it is also necessary to age and even to die.
Author Contributions
Conceptualization, A.Z. and R.G.; methodology, J.A., N.B., J-F.P., A.Z., R.Z.; software, R.Z., J-F.P., N.B., A.Z.; validation, A.Z., N.B., J-F.-P., R.G., J.A.; formal analysis, N.B., J-F.P., R.Z., A.Z.; investigation, N.B., R.Z., A.Z., G.M.; resources, A.Z., N.B., R.Z.; data curation, A.Z., N.B., R.Z.; writing—original draft preparation, A.Z.; writing—review and editing, A.Z., N.B., G.M.; visualization, A.Z., N.B.; supervision, A.Z., N.B.; project administration, A.Z.; funding acquisition, A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by UNIVERSITY OF PADUE, through the Ordinary Endowment Funds for Research (DOR, Dotazione Ordinaria Ricerca) attributed to prof. Augusto Zanella from 2007 to 2022.
Data Availability Statement
Not applicable here.
Acknowledgments
This article also arises from a long discussion involving Professors Orazio Ciancio and Susanna Nocentini. The aim of the discussion was to find practical indications of systemic sylviculture. We thank the founders of this modern forestry for the numerous suggestions and for the evolution of thinking that they have provoked in this year of fruitful exchange of ideas. Gaétan Zanella furnished the link to graphical issues made by artificial intelligence (https://www.bing.com/images/create).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brunet, M.; Beauvilain, A.; Coppens, Y.; Heintz, E.; Moutaye, A.H.E.; Pilbeam, D. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature 1995, 378, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Dobzhansky, T. Nothing in Biology Makes Sense except in the Light of Evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Lovelock, J.E.; Margulis, L. Atmospheric homeostasis by and for the biosphere: the gaia hypothesis. Tellus 1974, 26, 2–10. [Google Scholar] [CrossRef]
- Dahl, T.W.; Arens, S.K. The impacts of land plant evolution on Earth’s climate and oxygenation state – An interdisciplinary review. Chem. Geol. 2020, 547, 119665. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Volterra, V. Fluctuations in the Abundance of a Species considered Mathematically1. Nature 1926, 118, 558–560. [Google Scholar] [CrossRef]
- http://10.0.4.14/118558a0.
- Lotka, A.J. Analytical Note on Certain Rhythmic Relations in Organic Systems. Proc. Natl. Acad. Sci. USA 1920, 6, 410–415. [Google Scholar] [CrossRef]
- Rohde, R.A.; Muller, R.A. Cycles in fossil diversity. Nature 2005, 434, 208–210. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef]
- Darwin, C.M.A. On the Origin of Species by Means of Natural Selection or the Preservation of favoured races in the struggle for life; John Murray: Albemarle Street, London, UK, 1859; p. 502. [Google Scholar]
- Safran, R.J.; Nosil, P. Speciation: The Origin of New Species. Nat. Educ. Knowl. 2012, 3. Available online: https://www.nature.com/scitable/knowledge/library/speciation-the-origin-of-newspecies-26230527/.
- Dawkins, R. The Selfish Gene, Oxford Uni ed.; Oxford, 1976. [Google Scholar]
- Margulis, L. Symbiotic Planet {A new Look at Evolution}, Science Ma ed.; Perseus Books Group: New York, NY, USA, 1998; pp. 147–147. [Google Scholar]
- Khakhina, L.A.N.C.; Nikolaevna, L.; Margulis, R.C.L. (Eds.) Concepts of symbiogenesis: A Historical and Critical Study of the Researchof the Russian Botanists (Bio-Origins Series); Yale University Press: New Haven, CT, USA, 1992; p. 177. [Google Scholar]
- Brunet, M.; Guy, F.; Pilbeam, D.; Mackaye, H.T.; Likius, A.; Ahounta, D.; Beauvilain, A.; Blondel, C.; Bocherens, H.; Boisserie, J.-R.; et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature 2002, 418, 145–151. [Google Scholar] [CrossRef]
- Coppens, Y. Lucy’s Knee: The Story of Man and the Story of His Story (Paperback). Protea Boekhuis; English ed. edition (March 15, 2012), 2012, p. 176.
- Diamond, J.J. Guns, germs and steel - A short history of Everybody for the last 13,000 years; Vintage, 1998. [Google Scholar]
- Grice, E.A.; Segre, J.A. The Human Microbiome: Our Second Genome. Annu. Rev. Genom. Hum. Genet. 2012, 13, 151–170. [Google Scholar] [CrossRef]
- Vela, I.C.; Vilches, T.B.; Berndes, G.; Johnsson, F.; Thunman, H. Co-recycling of natural and synthetic carbon materials for a sustainable circular economy. J. Clean. Prod. 2022, 365. [Google Scholar] [CrossRef]
- Smil, V. Energy in World History—Energy and Civilization: A History; The MIT Press: Cambridge, MA, USA, 2017; p. 552. [Google Scholar]
- I. W. G. I, IPCC Sixth Assessment Report, Climate Change 2021: The physical Science Basis. Technical Summary. 2022. Available online: https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_FrontMatter.pdf.
- Condamine, F.L.; Rolland, J.; Morlon, H. Assessing the causes of diversification slowdowns: temperature-dependent and diversity-dependent models receive equivalent support. Ecol. Lett. 2019, 22, 1900–1912. [Google Scholar] [CrossRef]
- Lynas, M. Our Final Warning - Six Degrees of Climate Emergency; Fourth Estate, 2020. [Google Scholar]
- Fao and Unep, The State of the World’s Forests 2020. Forest, Biodiversity and People; Food and Agriculture Organization: Rome, Italy, 2020. [Google Scholar]
- Potapov, P.; Hansen, M.C.; Pickens, A.; Hernandez-Serna, A.; Tyukavina, A.; Turubanova, S.; Zalles, V.; Li, X.; Khan, A.; Stolle, F.; et al. The Global 2000-2020 Land Cover and Land Use Change Dataset Derived From the Landsat Archive: First Results. Front. Remote. Sens. 2022, 3. [Google Scholar] [CrossRef]
- Fao, Global Forest Resources Assessment 2020: Main report. Rome, 2020.
- Zhouri, A. “Adverse Forces” in the Brazilian Amazon: Developmentalism Versus Environmentalism and Indigenous Rights. J. Environ. Dev. 2010, 19, 252–273. [Google Scholar] [CrossRef]
- Fao, Ifad, Unicef, Wfp, and Who. The State of Food Security and Nutrition in the World 2022. Repurposing food and agricultural policies to make healthy diets more affordable; FAO: Rome, Italy, 2022. [Google Scholar]
- Jon, C. The International Space Station is home to potentially dangerous bacteria. An in-depth analysis of an air filter and vacuum bags discovers a unique microbiome. Science 2015. [Google Scholar] [CrossRef]
- Tršan, M.; Seme, K.; Srčič, S. The environmental monitoring in hospital pharmacy cleanroom and microbiota catalogue preparation. Saudi Pharm. J. 2019, 27, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Weir, A. Tha Martian, Penguin Ra ed.; Del Rey, 2011; p. 369. [Google Scholar]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.A.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian Vascular Flora: New Findings, Updates and Exploration of Floristic Similarities between Regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- Loy, A.; et al. Mammals of Italy: an annotated checklist. Hystrix 2019, 30, 87–106. [Google Scholar] [CrossRef]
- F. a. UNEP. The State of the World’s Forests 2020; ed: FAO and UNEP; 2020. [Google Scholar]
- Mannion, P.D.; Upchurch, P.; Benson, R.B.; Goswami, A. The latitudinal biodiversity gradient through deep time. Trends Ecol. Evol. 2014, 29, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–10. [Google Scholar] [CrossRef] [PubMed]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal Gradients of Biodiversity: Pattern, Process, Scale, and Synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Tierney, J.E.; Poulsen, C.J.; Montañez, I.P.; Bhattacharya, T.; Feng, R.; Ford, H.L.; Hönisch, B.; Inglis, G.N.; Petersen, S.V.; Sagoo, N.; et al. Past climates inform our future. Science 2020, 370, 680. [Google Scholar] [CrossRef] [PubMed]
- Mo, L.; Zanella, A.; Bolzonella, C.; Squartini, A.; Xu, G.-L.; Banas, D.; Rosatti, M.; Longo, E.; Pindo, M.; Concheri, G.; et al. Land Use, Microorganisms, and Soil Organic Carbon: Putting the Pieces Together. Diversity 2022, 14, 638. [Google Scholar] [CrossRef]
- Soong, J.L.; Fuchslueger, L.; Marañon-Jimenez, S.; Torn, M.S.; Janssens, I.A.; Penuelas, J.; Richter, A. Microbial carbon limitation: The need for integrating microorganisms into our understanding of ecosystem carbon cycling. Glob. Chang. Biol. 2019, 26, 1953–1961. [Google Scholar] [CrossRef]
- Pennanen, T.; Fritze, H.; De Boer, W.; Baldrian, P. Editorial: special issue on the ecology of soil microorganisms. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Hartmann, M.; Six, J. Soil structure and microbiome functions in agroecosystems. Nat. Rev. Earth Environ. 2023, 4, 4–18. [Google Scholar] [CrossRef]
- Bologna, M.; Aquino, G. Deforestation and world population sustainability: a quantitative analysis. Sci. Rep. 2020, 10, 7631. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Barrios, E.; Valencia, V.; Jonsson, M.; Brauman, A.; Hairiah, K.; Mortimer, P.E.; Okubo, S. Contribution of trees to the conservation of biodiversity and ecosystem services in agricultural landscapes. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 14, 1–16. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Guirado, E.; Reich, P.B.; Ochoa-Hueso, R.; Berdugo, M.; Sáez-Sandino, T.; Blanco-Pastor, J.L.; Tedersoo, L.; Plaza, C.; Ding, J.; et al. The global contribution of soil mosses to ecosystem services. Nat. Geosci. 2023, 16, 430–438. [Google Scholar] [CrossRef]
- Holland, R.; Eigenbrod, F.; Muggeridge, A.; Brown, G.; Clarke, D.; Taylor, G. A synthesis of the ecosystem services impact of second generation bioenergy crop production. Renew. Sustain. Energy Rev. 2015, 46, 30–40. [Google Scholar] [CrossRef]
- Sapp, J. Evolution by Association. A historyof Symbiosis; Oxford University Press, Inc.: New York, NY, USA, 1994; p. 255. [Google Scholar]
- Michaelian, K. Entropy Production and the Origin of Life. J. Mod. Phys. 2011, 02, 595–601. [Google Scholar] [CrossRef]
- Aszalós, R.; Thom, D.; Aakala, T.; Angelstam, P.; Brūmelis, G.; Gálhidy, L.; Gratzer, G.; Hlásny, T.; Katzensteiner, K.; Kovács, B.; et al. Natural disturbance regimes as a guide for sustainable forest management in Europe. Ecol. Appl. 2022, 32, e2596. [Google Scholar] [CrossRef] [PubMed]
- Cagliero, E.; et al. Exploring the long-term vegetation and fire-disturbance history of the Biogradska Gora old-growth forest (Montenegro); EGU: Vienna, Austria, 2020; Available online: https://meetingorganizer.copernicus.org/EGU2020/EGU2020-9910.html. [CrossRef]
- Motta, R. Why do we have to increase deadwood in our forests? How much deadwood does the forest need? For. Riv. Di Selvic. Ed Ecol. For. 2020, 17, 92–100. [Google Scholar] [CrossRef]
- Motta, R.; Ascoli, D.; Corona, P.; Marchetti, M.; Vacchiano, G. Silviculture and wind damages. The storm “Vaia”. For. Riv. Di Selvic. Ed Ecol. For. 2018, 15, 94–98. [Google Scholar] [CrossRef]
- Motta, R.; Larsen, J. A new paradigm for sustainable forest management: closeR to nature forest management. For. Riv. Di Selvic. Ed Ecol. For. 2022, 19, 52–62. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Keeton, W.S.; Lindner, M.; Svoboda, M.; Verkerk, P.J.; Bauhus, J.; Bruelheide, H.; Burrascano, S.; Debaive, N.; Duarte, I.; et al. Protection gaps and restoration opportunities for primary forests in Europe. Divers. Distrib. 2020, 26, 1646–1662. [Google Scholar] [CrossRef]
- Adamic, M.; Diaci, J.; Rozman, A.; Hladnik, D. Long-term use of uneven-aged silviculture in mixed mountain Dinaric forests: a comparison of old-growth and managed stands. For. Int. J. For. Res. 2016, 90, 279–291. [Google Scholar] [CrossRef]
- Hilbert, J.; Wiensczyk, A. Old-growth definitions and management: A literature review. J. Ecosyst. Manag. 2007, 8, 15–31. Available online: http://www.forrex.org/publications/jem/ISS39/vol8_no1_art2.pdf. [CrossRef]
- Leckie, S.; Vellend, M.; Bell, G.; Waterway, M.J.; Lechowicz, M.J. The seed bank in an old-growth, temperate deciduous forest. Can. J. Bot. 2000, 78, 181–192. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef]
- Almond, R.E.A.; Grooten, M.; Bignoli, D.J.; Petersen, T. World Wildlife Fund (WWF) and Zoological Society of London (2022). Living Planet Report 2022—Building a nature-positive society. Available online: https://ourworldindata.org/biodiversity.
- Fao, Global forest sector outlook 2050: Assessing future demand and sources of timber for a sustainable economy – Background paper for The State of the World’s Forests 2022; FAO Forestry Working Paper, No. 31; FAO, 2022.
- Jernigan, A.; Kao-Kniffin, J.; Pethybridge, S.; Wickings, K. Soil microarthropod effects on plant growth and development. Plant Soil 2022, 483, 27–45. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: diversity, complexity and dynamics. FEMS Microbiol. Rev. 2016, 41, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Bernier, N.; Ponge, J. Humus form dynamics during the sylvogenetic cycle in a mountain spruce forest. Soil Biol. Biochem. 1994, 26, 183–220. [Google Scholar] [CrossRef]
- Bernier, N. Earthworm feeding activity and development of the humus profile. Biol. Fertil. Soils 1998, 26, 215–223. [Google Scholar] [CrossRef]
- Zampedri, R. Relazione tra clima, forme di humus e dinamica forestale in ambiente di pecceta altimontana. Tesi di Dottorato. Ph.D. Thesis, Territorio e Sistemi Agro-Foestali (TESAF), Padova, Italy, 2005. [Google Scholar]
- André, J. Activité et diversité des organismes hétérotrophes: les clés du bouclage des cycles biogéochimiques et sylvigénétiques; Vallauri, D., André, J., Dodelin, B., Eynard-Machet, R., Rambaud, D., Eds.; Lavoisier, 2005; p. 404. [Google Scholar]
- Bernier, N. Fonctionnement biologique des humus et dynamique des pessières alpines. Le cas de la forêt de Macot-la-Plagne (Savoie). Ecologie 1997, 28, 23–44. Available online: https://www.researchgate.net/publication/233758152_Fonctionnement_biologique_des_humus_et_dynamique_des_pessieres_alpines_Le_cas_de_la_foret_de_Macot-La-Plagne_SavoieHumus_biological_organization_and_alpine_spruce_forest_dynamics_The_example_of_Macot-#ful.
- Bernier, N. Fonctionnement biologique des humus et dynamique des pessières alpines. Le cas de la forêt de Macot-La-Plagne (Savoie). [Humus biological organization and alpine spruce forest dynamics. The example of Macot-La-Plagne forest (Savoie, France)]. Ecologie 1997, 28, 23–44. [Google Scholar]
- Zampedri, R.; Zanella, A.; Giannini, R. Soil, humipedon and forest management. For. - Riv. di Selvic. ed Ecol. For. 2023, 20, 13–19. [Google Scholar] [CrossRef]
- Hartmann, F.; Marcuzzi, G. Diagnosi degli humus forestali su basi biomorfologiche; CEDAM Padova: Padova, Italy, 1970; pp. 284–284. [Google Scholar]
- Zanella, A.; et al. Humus Forestali - Manuale di ecologia per il riconoscimento e l’interpretazione - Applicazione alle faggete, CEA ed.; San Michele all’Adige, Trento (Italia): 321 p.: Centro Ecologia Alpina, Fondazione Edmund Mach; 2001; p. 320. [Google Scholar]
- Graefe, U. Gibt es in Deutschland die Humusform Amphi? Mitteilungen der Deutschen Bodenkundlichen Gesellschaft 2007, 110, 459–460. Available online: https://www.researchgate.net/profile/Ulfert_Graefe/publication/261286541_Gibt_es_in_Deutschland_die_Humusform_Amphi/links/00463533c43b6e792b000000.pdf.
- Zampedri, R.; Bernier, N.; Zanella, A.; Giannini, R.; Menta, C.; Visentin, F.; Mairota, P.; Mei, G.; Zandegiacomo, G.; Carollo, S.; et al. Soil, Humipedon, Forest Life and Management. Int. J. Plant Biol. 2023, 14, 571–592. [Google Scholar] [CrossRef]
- Gensac, P. Régénération en altitude de l’épicéa (Picea abies (L) Karst) sur les souches dans les Alpes françaises. Ann. For. Sci. 1990, 47, 173–182. [Google Scholar] [CrossRef]
- Motta, R.; Brang, P.; Frehner, M.; Ott, E. Copertura muscinale e rinnovazione di abete rosso (Picea abies L.) nella pecceta subalpina di Sedrum (Grigioni, Svizzera). Monti e Boschi 1994, 3, 49–56. [Google Scholar]
- Zanella, A.; Ponge, J.-F.; Fritz, I.; Pietrasiak, N.; Matteodo, M.; Nadporozhskaya, M.; Juilleret, J.; Tatti, D.; Le Bayon, R.-C.; Rothschild, L.; et al. Humusica 2, article 13: Para humus systems and forms. Appl. Soil Ecol. 2018, 122, 181–199. [Google Scholar] [CrossRef]
- Tatti, D.; Fatton, V.; Sartori, L.; Gobat, J.-M.; Le Bayon, R.-C. What does ‘lignoform’ really mean? Appl. Soil Ecol. 2018, 123, 632–645. [Google Scholar] [CrossRef]
- Lugato, E.; Bampa, F.; Panagos, P.; Montanarella, L.; Jones, A. Potential carbon sequestration of European arable soils estimated by modelling a comprehensive set of management practices. Glob. Chang. Biol. 2014, 20, 3557–3567. [Google Scholar] [CrossRef] [PubMed]
- Montanarella, L.; Panagos, P. The relevance of sustainable soil management within the European Green Deal. Land Use Policy 2020, 100, 104950. [Google Scholar] [CrossRef]
- Maturana, H.R.; Varela, F.J. Autopoiesis and Cognition (Boston Studies in the Philosophy and History of Science); Springer: Dordrecht, The Netherlands, 1980; p. 146. [Google Scholar]
- Zanella, A. Tipologia dei boschi: uno strumento per l’interpretazione e la gestione su basi ecologiche dei sistemi forestali. Parchi 1999, 3, 2–15. [Google Scholar]
- Del Favero, R. I boschi delle regioni alpine italiane. Tipologia, funzionamento, selvicoltura; CLEUP: Padova, Italy, 2004; p. 600. [Google Scholar]
- Del Favero, R. I boschi delle regioni alpine italiane. Tipologia, funzionamento, selvicoltura. Con CD-ROM; Edizione 2, Scienze forestali ed.; CLEUP, 2013; p. 602. [Google Scholar]
- Del Favero, R. I boschi delle regioni dell’Italia centrale. Tipologia, funzionamento, selvicoltura. Con CD-ROM, Scienze agrarie ed.; CLEUP, 2010; p. 430. [Google Scholar]
- Del Favero, R. I boschi delle regioni meridionali e insulari d’Italia. Tipologia, funzionamento, selvicoltura, Scienze Agrarie ed.; CLEUP, 2018; p. 469. [Google Scholar]
- Berretti, R.; Lingua, E.; Motta, R.; Piussi, P. Classificazione strutturale dei popolamenti forestali nella Riserva forestale integrale della Valbona a Paneveggio (TN). L’ Italia Forestale e Montana 2004, 59, 98–118. [Google Scholar]
- Dellagiacoma, F.; Motta, R.; Piussi. Ricerche sull’ecologia della pecceta subalpina nella foresta di Paneveggio. Dendronatura 1996, 17, 77–86. [Google Scholar]
- Piussi, P; Sulli, A.; MMencuccini. 30 Years of seed production in a subalpine Norway Spruce forest. Patterns of temporal and spatial variation. For. Ecol.Manag. 1995, 76, 109–125. [Google Scholar]
- Motta, R.; Berretti, R.; Castagneri, D.; Lingua, E.; Nola, P.; Vacchiano, G. Stand and coarse woody debris dynamics in subalpine Norway spruce forests withdrawn from regular management. Ann. For. Sci. 2010, 67, 803–803. [Google Scholar] [CrossRef]
- Motta, R.; Nola, P.; Piussi, P. Structure and stand development in three subalpine Norway spruce (Picea abies (L.) Karst.) stands in Paneveggio (Trento, Italy). Glob. Ecol.Biodivers. 1999, 8, 455–473. [Google Scholar] [CrossRef]
- Motta, R.; Nola, P.; Piussi, P. Long-term investigations in a strict forest reserve in the eastern Italian Alps: spatio-temporal origin and development in two multi-layered subalpine stands. J. Ecol. 2002, 90, 495–507. [Google Scholar] [CrossRef]
- Pignatti, G. Forest vegetation in view of some scenarios of climate change in Italy. For. Riv. Di Selvic. Ed Ecol. For. 2011, 8, 1–12. [Google Scholar] [CrossRef]
- Susmel, L. Normalizzazione delle foreste Alpine: basi ecosistemiche, equilibrio, modelli colturali, produttività: con applicazione alle foreste del Trentino; Liviana: Padova, Italy, 1980; p. 437. [Google Scholar]
- Savilaakso, S.; Johansson, A.; Häkkilä, M.; Uusitalo, A.; Sandgren, T.; Mönkkönen, M.; Puttonen, P. What are the effects of even-aged and uneven-aged forest management on boreal forest biodiversity in Fennoscandia and European Russia? A systematic review. Environ. Évid. 2021, 10, 1–38. [Google Scholar] [CrossRef]
- Xu, C.; De Frenne, P.; Blondeel, H.; De Pauw, K.; Landuyt, D.; Lorer, E.; Sanczuk, P.; Verheyen, K.; De Lombaerde, E. Light more than warming impacts understory tree seedling growth in a temperate deciduous forest. For. Ecol. Manag. 2023, 549. [Google Scholar] [CrossRef]
- Brown, R.W.; Jones, D.L. Plasticity of microbial substrate carbon use efficiency in response to changes in plant carbon input and soil organic matter status. Soil Biol. Biochem. 2023. [Google Scholar] [CrossRef]
- Zanella, A.; Ponge, J.-F.; Jabiol, B.; Van Delft, B.; De Waal, R.; Katzensteiner, K.; Kolb, E.; Bernier, N.; Mei, G.; Blouin, M.; et al. A Standardized Morpho-Functional Classification of the Planet’s Humipedons. Soil Syst. 2022, 6, 59. [Google Scholar] [CrossRef]
- Zhou, J.; Wen, Y.; Rillig, M.C.; Shi, L.; Dippold, M.A.; Zeng, Z.; Kuzyakov, Y.; Zang, H.; Jones, D.L.; Blagodatskaya, E. Restricted power: Can microorganisms maintain soil organic matter stability under warming exceeding 2 degrees? Glob. Ecol. Biogeogr. 2023, 32, 919–930. [Google Scholar] [CrossRef]
- Ette, J.-S.; Sallmannshofer, M.; Geburek, T. Assessing Forest Biodiversity: A Novel Index to Consider Ecosystem, Species, and Genetic Diversity. Forests 2023, 14, 709. [Google Scholar] [CrossRef]
- Kooch, Y.; Zarei, F.D. Soil function indicators below shrublands with different species composition. CATENA 2023, 227, 107111. [Google Scholar] [CrossRef]
- Della Rocca, F.; Stefanelli, S.; Pasquaretta, C.; Campanaro, A.; Bogliani, G. Effect of deadwood management on saproxylic beetle richness in the floodplain forests of northern Italy: some measures for deadwood sustainable use. J. Insect Conserv. 2014, 18, 121–136. [Google Scholar] [CrossRef]
- Lachat, T.; et al. Bois mort en forêt. Formation, importance et conservation; Institut fédéral de recherches WSL CH-8903 Birmensdorf: Birmensdorf, CH, USA, 2014; pp. 1–12. [Google Scholar]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef]
- Zanella, A.; Berg, B.; Ponge, J.-F.; Kemmers, R.H. Humusica 1, article 2: Essential bases—Functional considerations. Appl. Soil Ecol. 2018, 122, 22–41. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.-D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Ciancio, O.; Nocentini, S. The forest and man: the evolution of forestry thought from modern humanism to the culture of complexity. Systemic silviculture and management on natural bases; Ciancio, O., Ed.; Accademia Italiana di Scienze Forestali, Firenze (Italia): Florence, Italy, 1997; p. 115. [Google Scholar]
- Nocentini, S.; Ciancio, O.; Portoghesi, L.; Corona, P. Historical roots and the evolving science of forest management under a systemic perspective. Can. J. For. Res. 2021, 51, 163–171. [Google Scholar] [CrossRef]
- Ciancio, O. La teoria della selvicoltura sistemica, le “sterili disquisizioni” e il sonnambulismo dell’intellighenzia forestale; Accademia Italiana di Scienze Forestali, 2010; ISBN 9788887553192. [Google Scholar]
- Ciancio, O. Storia del pensiero forestale. Selvicoltura, filosofia, etica; Rubettino, 2014; p. 546. [Google Scholar] [CrossRef]
- Ciancio, O.; Iovino, F.; Nocentini, S. The theory of the normal forest. La teoria del bosco normale. In collaborazione con O. Ciancio e F. Iovino. L’Italia For. Emont. 1994, 49, 446–462. [Google Scholar]
- F. Provincia Autonoma di Trento - Servizio Foreste e, Linee Tecniche per la Pianificazione Forestale Aziendale - Versione approvata con determinazione del Dirigente del Servizio Foreste e Fauna n. 56 di data 19 dicembre 2016 Art. 9 del D.P.G.P. n. 35 del 26 agosto 2008. Provincia Autonoma di Trento - Servizio Foreste e Fauna, 2016, pp. 135-135.
- Thomasius, H. Geschichte, Theorie und Praxis des Dauerwaldes: erweiterte Fassung eines Vortrages anläßlich der Jahrestagung 1996 des Landesforstvereins Sachsen-Anhalt am 14.05.1996 in Garitz bei Dessau. Salzland Druck GmbH, 1996.
- Cappelli, M. Selvicoltura Generale. Edagricole 1987, 300. [Google Scholar]
- Margulis, L.; Sagan, D.; Eldredge, N. What is Life? University of California Press: Berkeley and Los Angeles, CA, USA, 2000. [Google Scholar]
- A. Ripartizione per le Foreste Ufficio Pianificazione Forestale Provincia Autonoma di Bolzano Alto; Tipologie forestali dell’Alto Adige - VOLUME 1: tipi forestali, regioni forestali, chiave dei tipi forestali; Provincia Autonoma di Bolzano-Alto Adige Ripartizione per le foreste Ufficio Pianificazione forestale: Bolzano, Italy, 2010; p. 310.
- Szewczyk, J.; Szwagrzyk, J. Tree regeneration on rotten wood and on soil in old-growth stand. Plant Ecol. 1996, 122, 37–46. [Google Scholar] [CrossRef]
- Rodriguez, A.; Perestelo, F.; Carnicero, A.; Regalado, V.; Perez, R.; Fuente, G.; Falcon, M. Degradation of natural lignins and lignocellulosic substrates by soil-inhabiting fungi imperfecti. FEMS Microbiol. Ecol. 1996, 21, 213–219. [Google Scholar] [CrossRef]
- Hardersen, S.; Zapponi, L. Wood degradation and the role of saproxylic insects for lignoforms. Appl. Soil Ecol. 2018, 123, 334–338. [Google Scholar] [CrossRef]
- Hu, M.; Lehtonen, A.; Minunno, F.; Mäkelä, A. Age effect on tree structure and biomass allocation in Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies [L.] Karst.). Ann. For. Sci. 2020, 77, 1–15. [Google Scholar] [CrossRef]
- Suzuki, M.; Umeki, K.; Orman, O.; Shibata, M.; Tanaka, H.; Iida, S.; Nakashizuka, T.; Masaki, T. When and why do trees begin to decrease their resource allocation to apical growth? The importance of the reproductive onset. Oecologia 2019, 191, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Villarino, S.H.; Pinto, P.; Jackson, R.B.; Piñeiro, G. Plant rhizodeposition: A key factor for soil organic matter formation in stable fractions. Sci. Adv. 2021, 7, eabd3176. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Reverdy, A.; She, Q.; Sun, B.; Chai, Y. The role of rhizodeposits in shaping rhizomicrobiome. Environ. Microbiol. Rep. 2019, 12, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Lowenfels, J. Teaming with bacteria. The Organic Gardener’s Guide to Endophytic Bacteria and the Rhizophagy Cycle; Timber Press, Inc.: Portland, OR, USA, 2022. [Google Scholar]
- Radmanović, S.; Nikolić, N.; Đorđević, A. Humic Acids Optical Properties Of Rendzina Soils In Diverse Environmental Conditions Of Serbia. Arch. Tech. Sci. 2018, 1, 63–70. [Google Scholar] [CrossRef]
- Susmel, L. Leggi di variazione dei parametri della foresta disetanea normale. L’Italia For. Emont. 1956, 3, 105–116. [Google Scholar]
- Klepac, D. Corsi di cultura in ecologia 1976-1977. Pianifucazione e governo delle abetine disetanee e dei rovereti della Croazia; Padova, Italy, 1978. [Google Scholar]
- Susmel, L. Conservazione e miglioramento delle abetine delle Alpi Orientali. In Atti del Congresso Nazionale di Selvicoltura; Accademie Nazionale di Scienze Forestali: Firenze, Italy, 1954; pp. 331–372. [Google Scholar]
- Susmel, L. Riordinamento su basi bioecologiche delle abetine di S. Vito di Cadore; Pubbl. n. 9.; Stazione Sperimentale di Selvicoltura: Firenze, Italy, 1955. [Google Scholar]
- Pesendorfer, M.B.; Bogdziewicz, M.; Szymkowiak, J.; Borowski, Z.; Kantorowicz, W.; Espelta, J.M.; Fernández-Martínez, M. Investigating the relationship between climate, stand age, and temporal trends in masting behavior of European forest trees. Glob. Chang. Biol. 2020, 26, 1654–1667. [Google Scholar] [CrossRef]
- Mei, G.; Pesaresi, S.; Corti, G.; Cocco, S.; Colpi, C.; Taffetani, F. Changes in vascular plant species composition, top-soil and seed-bank along coppice rotation in an Ostrya carpinifolia forest. Plant Biosyst. - Int. J. Deal. all Asp. Plant Biol. 2019, 154, 259–268. [Google Scholar] [CrossRef]
- Altman, J.; Hédl, R.; Szabó, P.; Mazůrek, P.; Riedl, V.; Müllerová, J.; Kopecký, M.; Doležal, J. Tree-Rings Mirror Management Legacy: Dramatic Response of Standard Oaks to Past Coppicing in Central Europe. PLoS ONE 2013, 8, e55770. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Buckley, G.P. Ecology and Management of Coppice Woodlands; Chapman and Hall: London, UK, 1992; p. 366. [Google Scholar] [CrossRef]
- Piussi, P.; Alberti, G. General Forestry. Woods, society and management techniques, Collana Sc ed.; Compagnia delle Foreste S.r.l.: Arezzo, 2015; p. 432. [Google Scholar]
- Buckley, P. Coppice restoration and conservation: a European perspective. J. For. Res. 2020, 25, 125–133. [Google Scholar] [CrossRef]
- Mairota, P.; Manetti, M.; Amorini, E.; Pelleri, F.; Terradura, M.; Frattegiani, M.; Savini, P.; Grohmann, F.; Mori, P.; Terzuolo, P.; et al. Opportunities for coppice management at the landscape level: the Italian experience. iForest - Biogeosciences For. 2016, 9, 775–782. [Google Scholar] [CrossRef]
Figure 1.
Adam and Eve in the Trauttmansdorff Castle Park, Merano, Italy. From that day, contravening a hypothetical divine warning, we entered the Anthropocene (We know that Hominids were an integral part of the forest in the Paleolithic and also in a large part of the Neolithic, of course. Currently, the consideration of non-log products, “ecosystem services” and biodiversity means that man is turning to the forest with a more conscious view of the value of this ecosystem.
Figure 1.
Adam and Eve in the Trauttmansdorff Castle Park, Merano, Italy. From that day, contravening a hypothetical divine warning, we entered the Anthropocene (We know that Hominids were an integral part of the forest in the Paleolithic and also in a large part of the Neolithic, of course. Currently, the consideration of non-log products, “ecosystem services” and biodiversity means that man is turning to the forest with a more conscious view of the value of this ecosystem.

Figure 2.
Biodiversity during the Phanerozoic. From: Mannion et al. 2014 [35], with data from Jenkins et al. 2013 [36] and Willig et al. 2003 [37]. The growth of biodiversity is noted in the long term, with moments of crisis every 62 million years. In: Wikimedia Commons. .
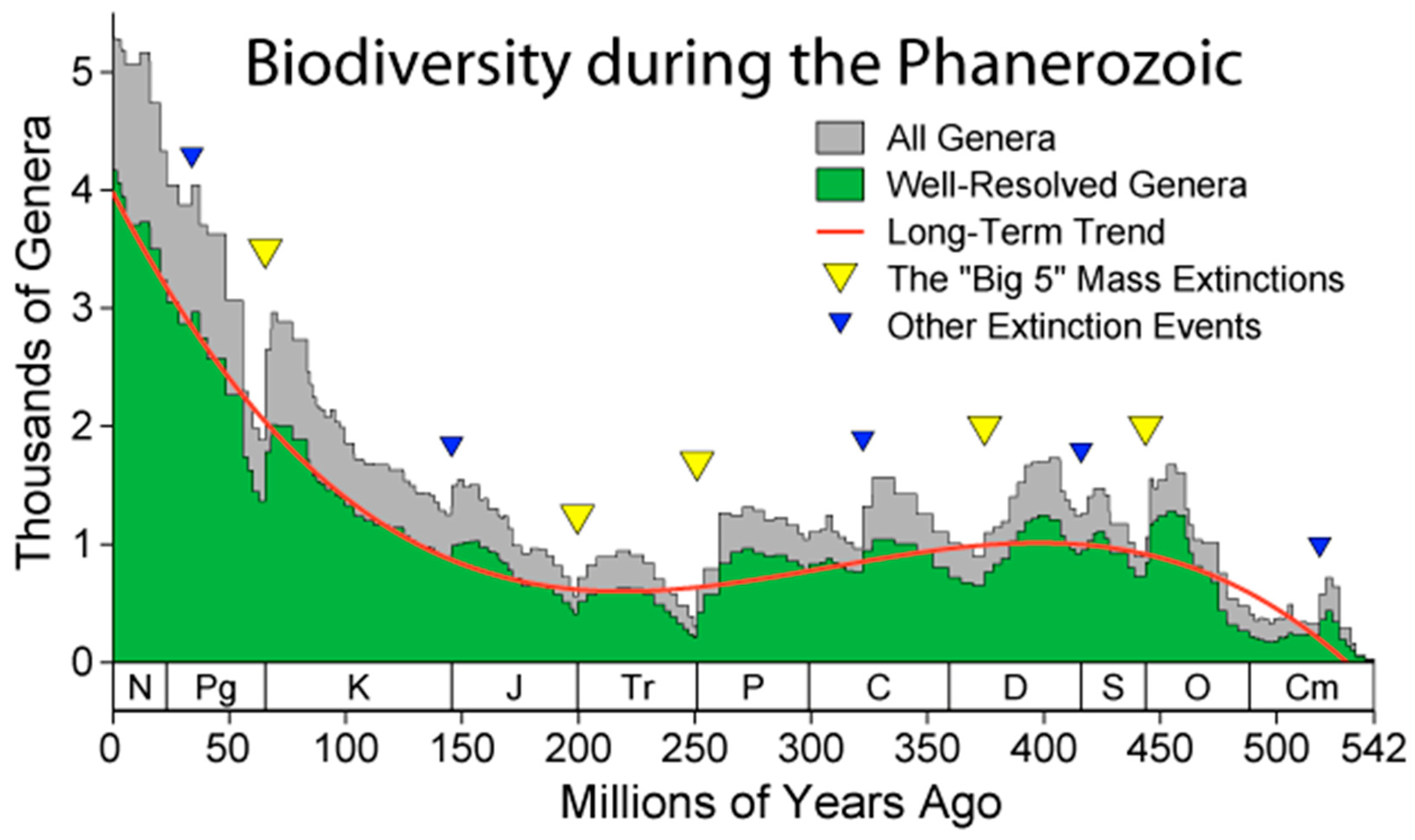
Figure 3.
Gaia hypothesis. In 1974, James Lovelock and Lynn Margulis published an article [3] in which they reported scientific evidence that once started, life acquired control of the planetary environment and produced the homeostasis by and for the biosphere that persists ever since. For fun, we used the artificial intelligence (AI) available online (https://www.bing.com/images/create) to create two images, using as “prompt” the words written under the figures. Up: To create the left figure, we didn’t use the world “soil”, but AI “understood” that the forest is made of two layers, with one very rich and partially unknown under our feet. Down: Noteworthy is the place that in the two figures the program assigns to the ground. Furthermore, the stratification of the latter into horizons, as in real construction, is highlighted with an orizontal organization which is not far from what we see on the ground. In the figure on the right, in the basement there are living animals from outside the ground, with green plants, as if to emphasize the rapprochement of these two worlds, between the earth and the earth, which we appreciate very much: this is where the whole that makes up the forest, in a vision that comes close to the concept of Gaia.
Figure 3.
Gaia hypothesis. In 1974, James Lovelock and Lynn Margulis published an article [3] in which they reported scientific evidence that once started, life acquired control of the planetary environment and produced the homeostasis by and for the biosphere that persists ever since. For fun, we used the artificial intelligence (AI) available online (https://www.bing.com/images/create) to create two images, using as “prompt” the words written under the figures. Up: To create the left figure, we didn’t use the world “soil”, but AI “understood” that the forest is made of two layers, with one very rich and partially unknown under our feet. Down: Noteworthy is the place that in the two figures the program assigns to the ground. Furthermore, the stratification of the latter into horizons, as in real construction, is highlighted with an orizontal organization which is not far from what we see on the ground. In the figure on the right, in the basement there are living animals from outside the ground, with green plants, as if to emphasize the rapprochement of these two worlds, between the earth and the earth, which we appreciate very much: this is where the whole that makes up the forest, in a vision that comes close to the concept of Gaia.

Figure 4.
Up: The flat, still mirror of a pond’s water and the dawning light of day can produce an unexpected effect: that the upside-down image of trees reflected in the water is sharper than that of their shape in the air. In the very brief hesitation moment in which one wonders which of the two images corresponds to the real world, one tries to evaluate the fact that both figures can correspond to reality. Here, precisely in this moment of meditative stasis, invaded by the desire to investigate to understand, also to defend oneself from the fog that prevents progress, connections open towards the hidden part of the forest where the roots of the trees fish and which we call soil. The clearest part of the forest is perhaps the one we don’t see. In the soil of a European forest one can easily count one million of individuals of soil animals per m2, as the ones collected by students in a Fago-Quercetum and reported on a band in the middle of the figure.
Figure 4.
Up: The flat, still mirror of a pond’s water and the dawning light of day can produce an unexpected effect: that the upside-down image of trees reflected in the water is sharper than that of their shape in the air. In the very brief hesitation moment in which one wonders which of the two images corresponds to the real world, one tries to evaluate the fact that both figures can correspond to reality. Here, precisely in this moment of meditative stasis, invaded by the desire to investigate to understand, also to defend oneself from the fog that prevents progress, connections open towards the hidden part of the forest where the roots of the trees fish and which we call soil. The clearest part of the forest is perhaps the one we don’t see. In the soil of a European forest one can easily count one million of individuals of soil animals per m2, as the ones collected by students in a Fago-Quercetum and reported on a band in the middle of the figure.

Figure 5.
Soil horizons and functional stratification of the soil in aerated conditions. Soil organisms are more present in the humipedon.
Figure 5.
Soil horizons and functional stratification of the soil in aerated conditions. Soil organisms are more present in the humipedon.
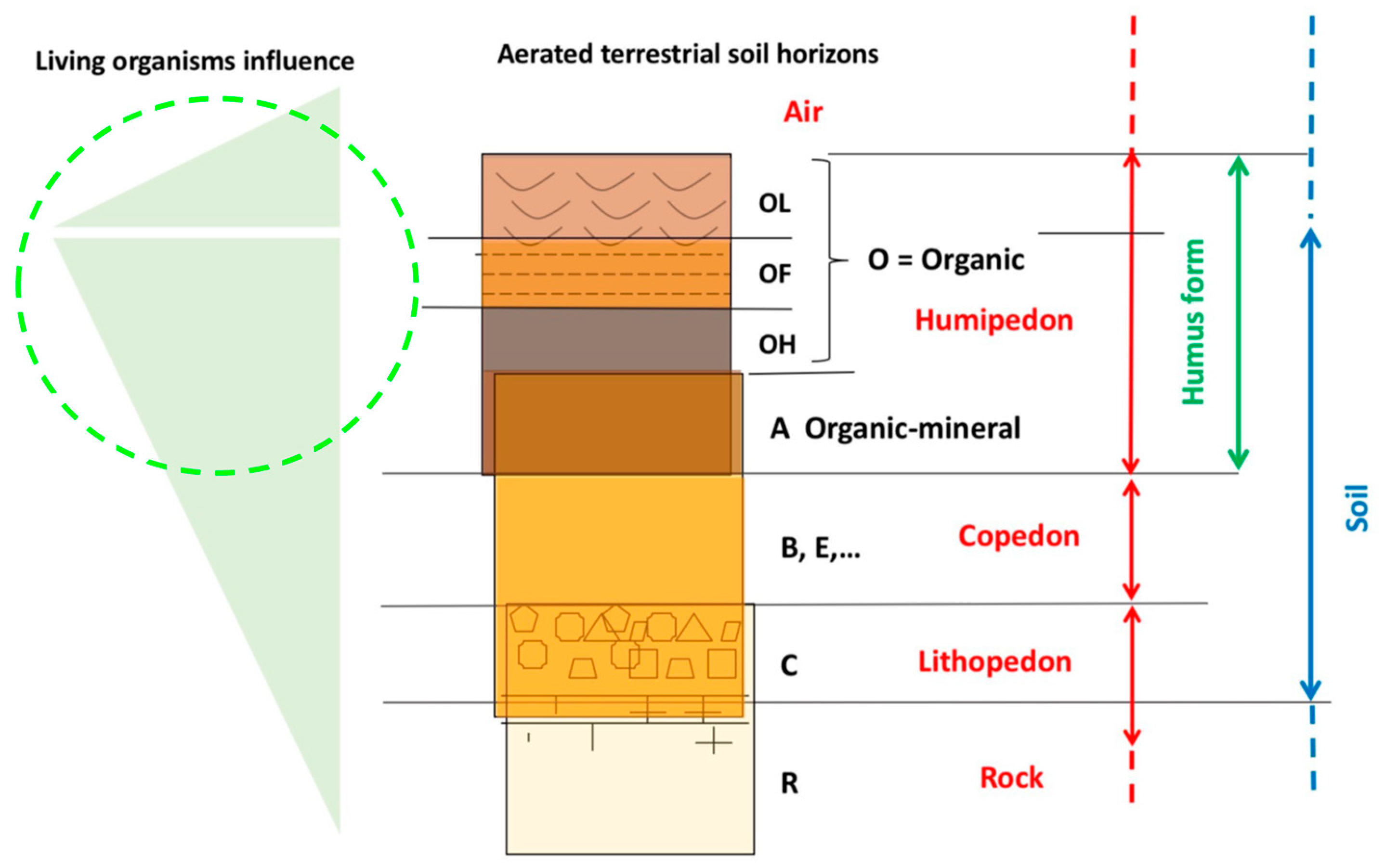
Figure 6.
Soil has a history. On our planet the soil is undergoing an evolution that follows that of the rest of living beings. Crusto humipedons are thin and cover a range of very young soils ranging from biofilms to desert crusts. They are formed by microorganisms or very small plants and animals. In the forest they are very common on rock outcrops or on tree trunks. Bryo are humipedons of greater thicknesses made of mosses and lichens, and pedofauna. Rhizo are humipedons that are formed by the strong action of the roots of herbaceous or shrubby plants, in cooperation with pedofauna. Along a gradient that runs from the high mountains to the plains, Mor and Moder or Tangel and Amphi humipedons form, respectively on acidic or basic parent rock respectively. In milder, lowland environments the dominant humipedon is the Mull. Moving through the figure from left to right we travel through the time of natural evolution and soil formation.
Figure 6.
Soil has a history. On our planet the soil is undergoing an evolution that follows that of the rest of living beings. Crusto humipedons are thin and cover a range of very young soils ranging from biofilms to desert crusts. They are formed by microorganisms or very small plants and animals. In the forest they are very common on rock outcrops or on tree trunks. Bryo are humipedons of greater thicknesses made of mosses and lichens, and pedofauna. Rhizo are humipedons that are formed by the strong action of the roots of herbaceous or shrubby plants, in cooperation with pedofauna. Along a gradient that runs from the high mountains to the plains, Mor and Moder or Tangel and Amphi humipedons form, respectively on acidic or basic parent rock respectively. In milder, lowland environments the dominant humipedon is the Mull. Moving through the figure from left to right we travel through the time of natural evolution and soil formation.

Figure 7.
Amphi humipedon in a montane spruce forest in the Alps. Left: a black organic OH horizon overlies an ochre organo-mineral A horizon. Arthropods or enchytraeids develop in the OH horizon; target earthworms inhabit and produce the A horizon. On the right: Graph from Bernier (1998) [65]. There are many earthworms in the young or old stages of the forest and here the A horizon is very present and rather thick; in the central part of the graph corresponding to the stage of intermediate age of the forest, the A horizon tends to disappear in favor of a more organic OH horizon.
Figure 7.
Amphi humipedon in a montane spruce forest in the Alps. Left: a black organic OH horizon overlies an ochre organo-mineral A horizon. Arthropods or enchytraeids develop in the OH horizon; target earthworms inhabit and produce the A horizon. On the right: Graph from Bernier (1998) [65]. There are many earthworms in the young or old stages of the forest and here the A horizon is very present and rather thick; in the central part of the graph corresponding to the stage of intermediate age of the forest, the A horizon tends to disappear in favor of a more organic OH horizon.
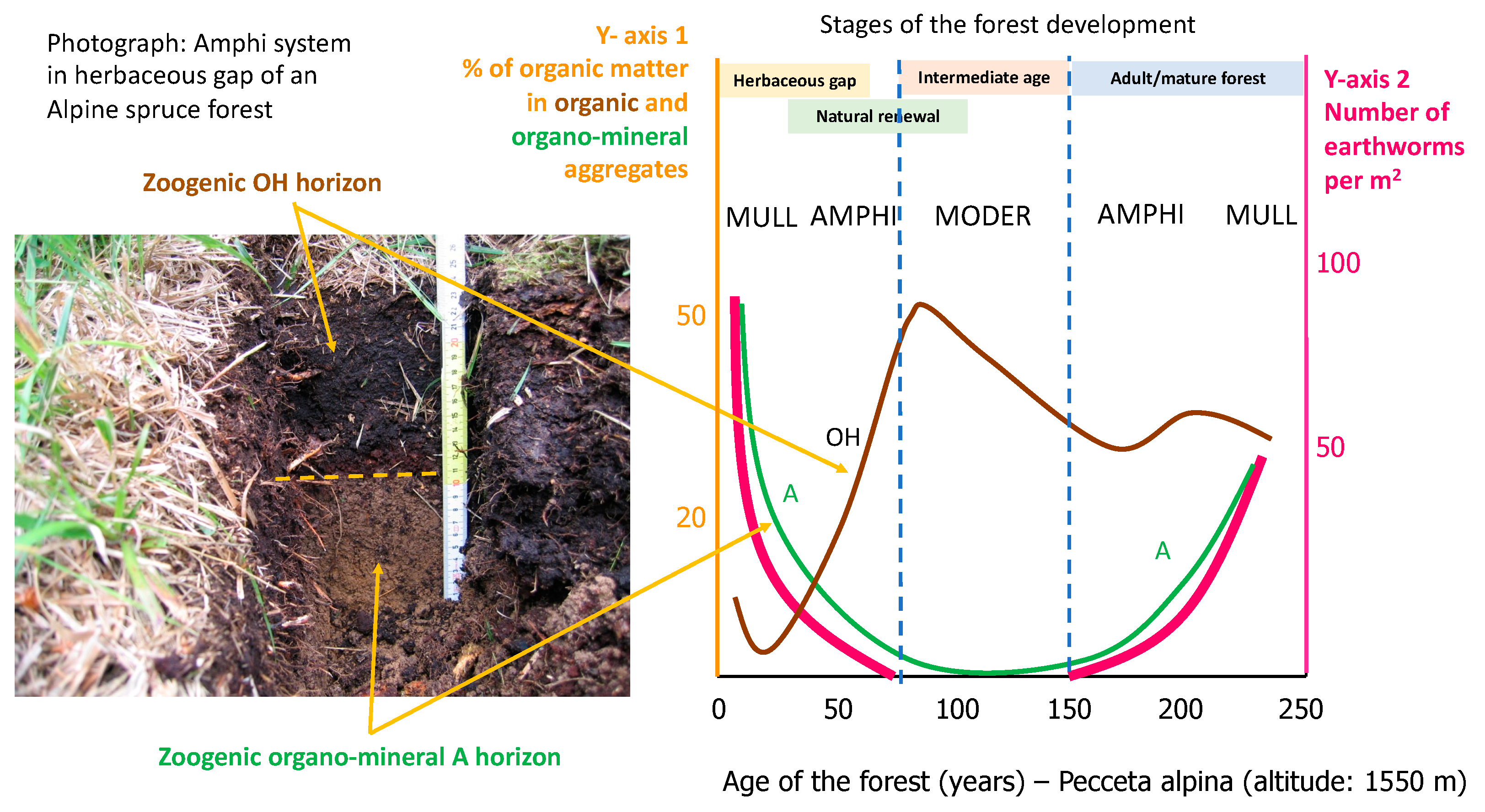
Figure 8.
Variations in the humipedon of a spruce forest as a function of tree age and elevation. From Bernier [68], modified to make it compatible with the modern classification of forest humus. With increasing altitude, the climate changes, becoming less conducive to the biodegradation of litter (the main factor is the lowering of the average air temperature). This favors the increase in thickness of the more superficial organic horizons (yellow and purple) to the detriment of the underlying organo-mineral horizons.
Figure 8.
Variations in the humipedon of a spruce forest as a function of tree age and elevation. From Bernier [68], modified to make it compatible with the modern classification of forest humus. With increasing altitude, the climate changes, becoming less conducive to the biodegradation of litter (the main factor is the lowering of the average air temperature). This favors the increase in thickness of the more superficial organic horizons (yellow and purple) to the detriment of the underlying organo-mineral horizons.
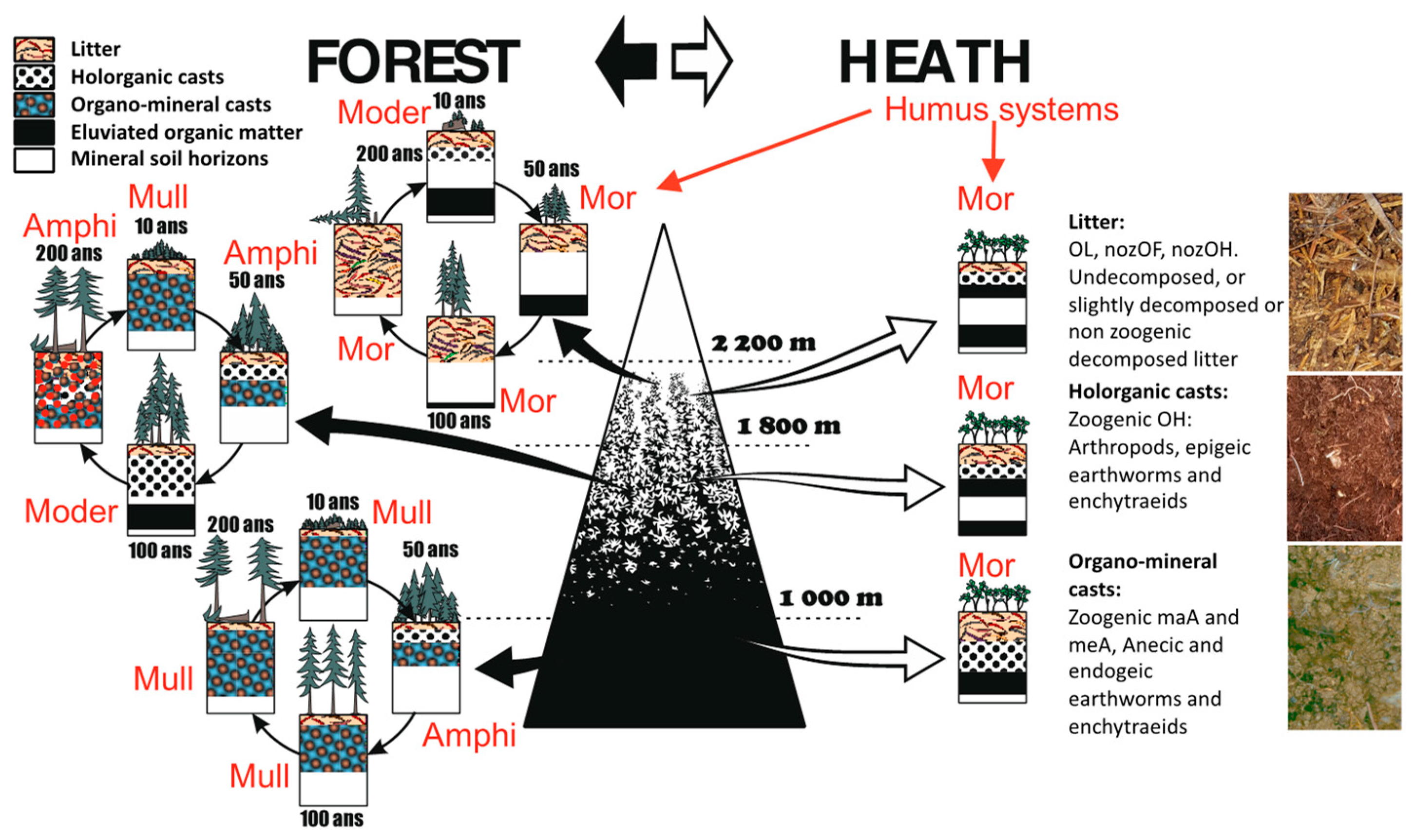
Figure 9.
Forest stages and corresponding change in humipedon. The 300-year cycle of an Alpine spruce forest can be divided down into two important periods for the soil: the first 150 years of progressive impoverishment of the soil, with energy and mass migrating from it to the trees, followed by another 150 years of soil enrichment as energy and mass that returns from the trees to the soil. Three initial Para humus systems have been illustrated in the above figure 8: Crusto, Bryo and Rhizo. The Ligno system at the bottom of figure 11, which corresponds to the biodegradation of wood, is also classified as Para system. Generated by animals and fungi specific to this organic substrate, this biological process of wood recycling is particularly evident on the stumps and branches left to decompose-freely after the removal of the tree trunk. Decaying spruce stumps are often and vigorously colonized by the renewal of the same species. The Bryo system often precedes the spruce seedlings on the stumps. .
Figure 9.
Forest stages and corresponding change in humipedon. The 300-year cycle of an Alpine spruce forest can be divided down into two important periods for the soil: the first 150 years of progressive impoverishment of the soil, with energy and mass migrating from it to the trees, followed by another 150 years of soil enrichment as energy and mass that returns from the trees to the soil. Three initial Para humus systems have been illustrated in the above figure 8: Crusto, Bryo and Rhizo. The Ligno system at the bottom of figure 11, which corresponds to the biodegradation of wood, is also classified as Para system. Generated by animals and fungi specific to this organic substrate, this biological process of wood recycling is particularly evident on the stumps and branches left to decompose-freely after the removal of the tree trunk. Decaying spruce stumps are often and vigorously colonized by the renewal of the same species. The Bryo system often precedes the spruce seedlings on the stumps. .
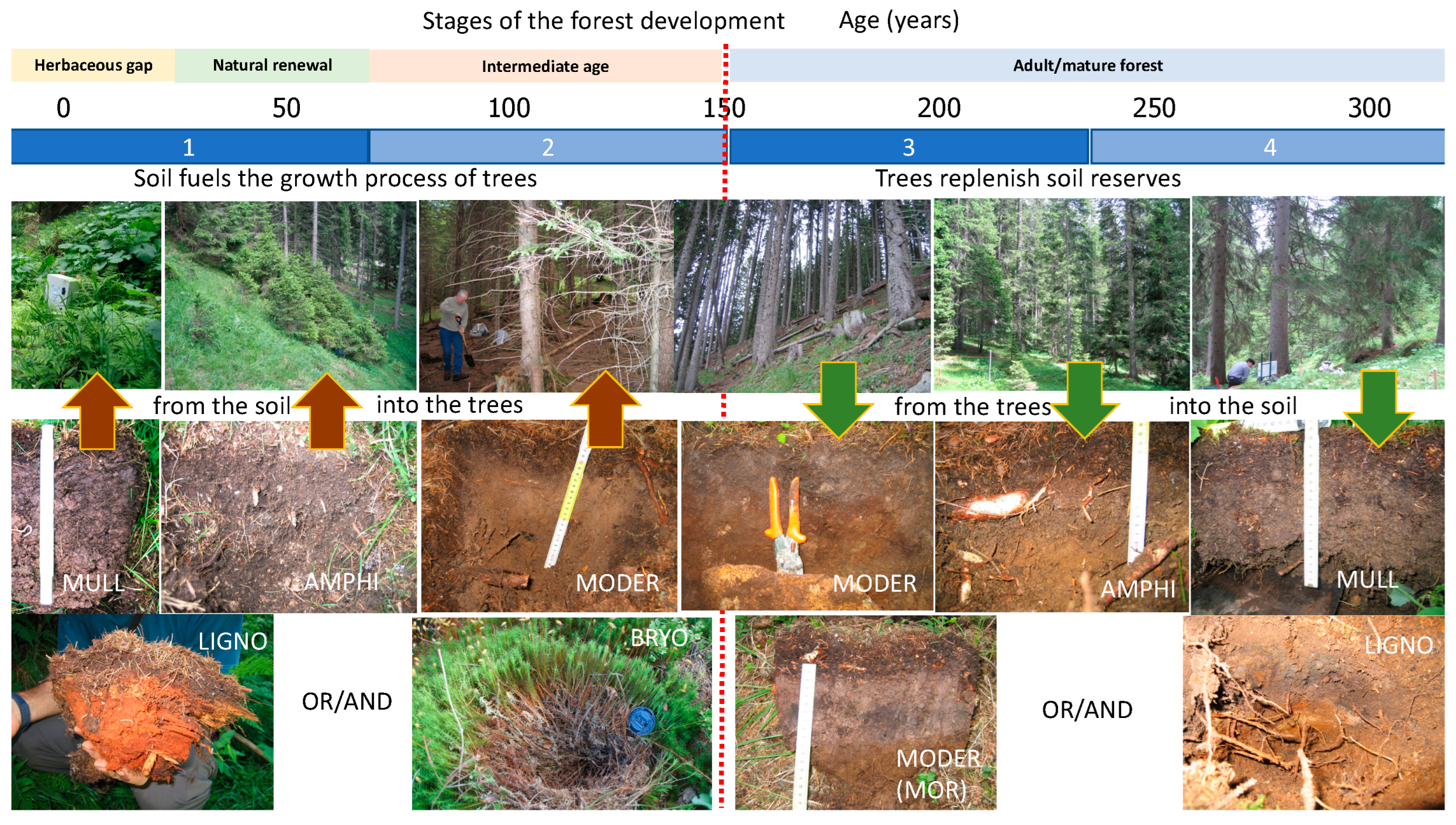
Figure 10.
The development stages of the Alpine spruce forest, the corresponding humus systems, arranged by an ACP in the space defined by climatic and pedological variables. Figure constructed from the work of Zampedri [66], published in [74]. Cutting the graph along the diagonal that starts from the upper left corner and reaches the lower right, the stations on carbonate rock (B, basic: limestone and dolomite) are positioned in the upper half on the right, while those on acidic rock (A, acidic: granites and porphyries) end in the lower half on the left. High pH, CEC%, thickness of the maA and meA horizons, average temperature for the month of May, soil humidity and number of days with temperatures below zero push the stations into the upper left box; High organic carbon, thickness of organic horizons OF and OH and thickness of organo-mineral horizon miA, average soil temperature at 10 cm depth of months 2, 3, 4, 5, 6, Absolute minimum in spring and autumn and average of the month of January, variables linked to the radiation budget instead push the points towards the lower left corner.
Figure 10.
The development stages of the Alpine spruce forest, the corresponding humus systems, arranged by an ACP in the space defined by climatic and pedological variables. Figure constructed from the work of Zampedri [66], published in [74]. Cutting the graph along the diagonal that starts from the upper left corner and reaches the lower right, the stations on carbonate rock (B, basic: limestone and dolomite) are positioned in the upper half on the right, while those on acidic rock (A, acidic: granites and porphyries) end in the lower half on the left. High pH, CEC%, thickness of the maA and meA horizons, average temperature for the month of May, soil humidity and number of days with temperatures below zero push the stations into the upper left box; High organic carbon, thickness of organic horizons OF and OH and thickness of organo-mineral horizon miA, average soil temperature at 10 cm depth of months 2, 3, 4, 5, 6, Absolute minimum in spring and autumn and average of the month of January, variables linked to the radiation budget instead push the points towards the lower left corner.
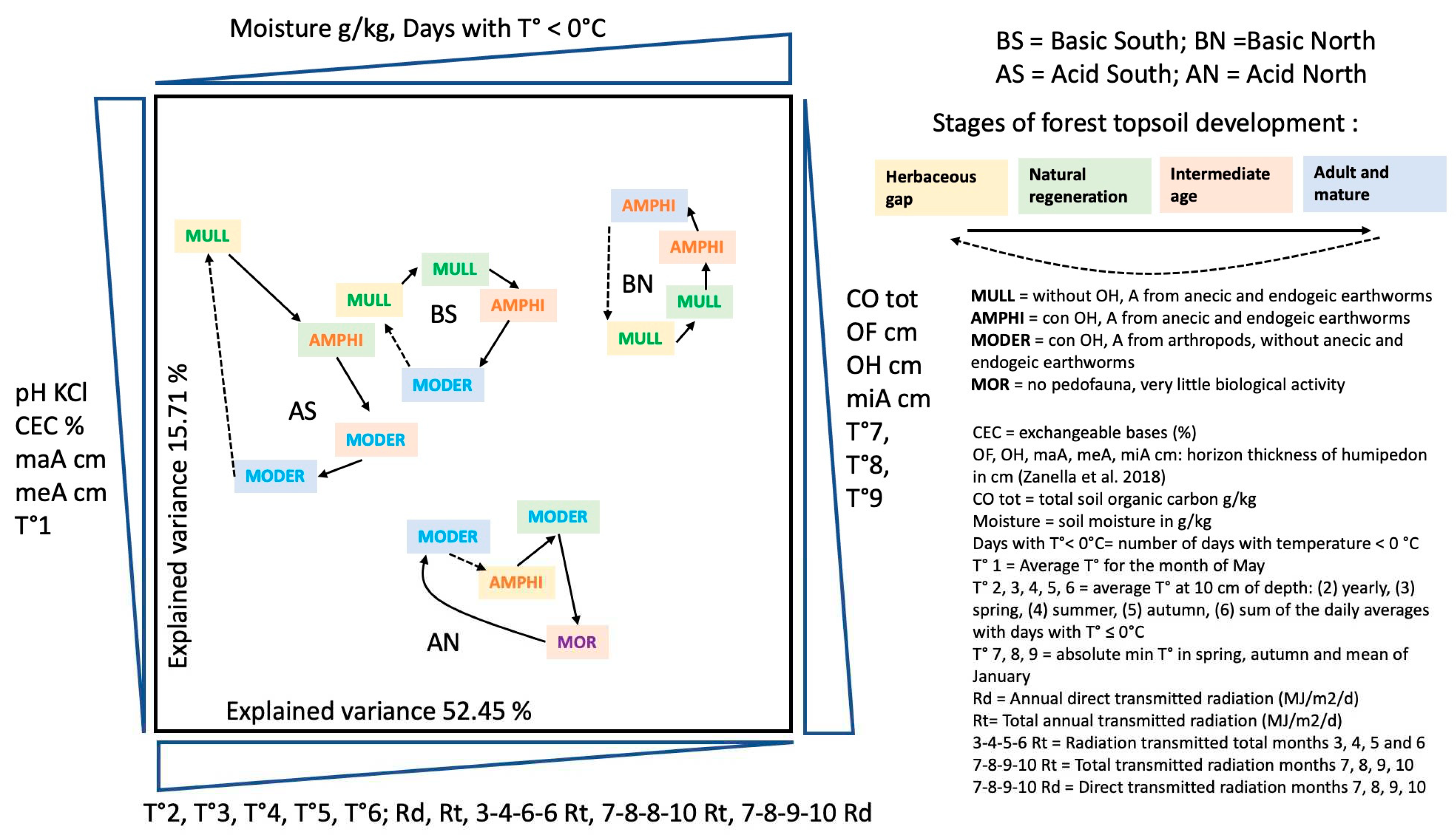
Figure 11.
A forest that turns and grows, like a wheel that recycles increasingly larger parts of the material from which it is built, to reinvest them in a new and more complex forest. Some of the growth is partially reinjected in the soil, which acts as an investment bank for the forest system.
Figure 11.
A forest that turns and grows, like a wheel that recycles increasingly larger parts of the material from which it is built, to reinvest them in a new and more complex forest. Some of the growth is partially reinjected in the soil, which acts as an investment bank for the forest system.
Figure 12.
The normalization cuts. In the figure the “norm” (theoretical) lines are shown in green, the crosses (+) indicate the real trees to be removed to normalize the distribution of the number of trees with age; the black points at the end of the cycle correspond to old trees, released to improve the biodiversity of the forest. The figure shows that two cycles of managed forest can be inserted in 300-400 years. It is as if, in order to increase the woody production of the forest, the forester had removed the forest from its “overripe” high forest stage and had forced the forest to remain young. This action determines the increase of the surface occupied by the central stages of the forestry cycle (2 and 3, young and mature high forest), to the detriment of that in which the regeneration and old age stages develop; the latter shorten and overlap (1-4). The middle stages with Moder humus systems become dominant.
Figure 12.
The normalization cuts. In the figure the “norm” (theoretical) lines are shown in green, the crosses (+) indicate the real trees to be removed to normalize the distribution of the number of trees with age; the black points at the end of the cycle correspond to old trees, released to improve the biodiversity of the forest. The figure shows that two cycles of managed forest can be inserted in 300-400 years. It is as if, in order to increase the woody production of the forest, the forester had removed the forest from its “overripe” high forest stage and had forced the forest to remain young. This action determines the increase of the surface occupied by the central stages of the forestry cycle (2 and 3, young and mature high forest), to the detriment of that in which the regeneration and old age stages develop; the latter shorten and overlap (1-4). The middle stages with Moder humus systems become dominant.
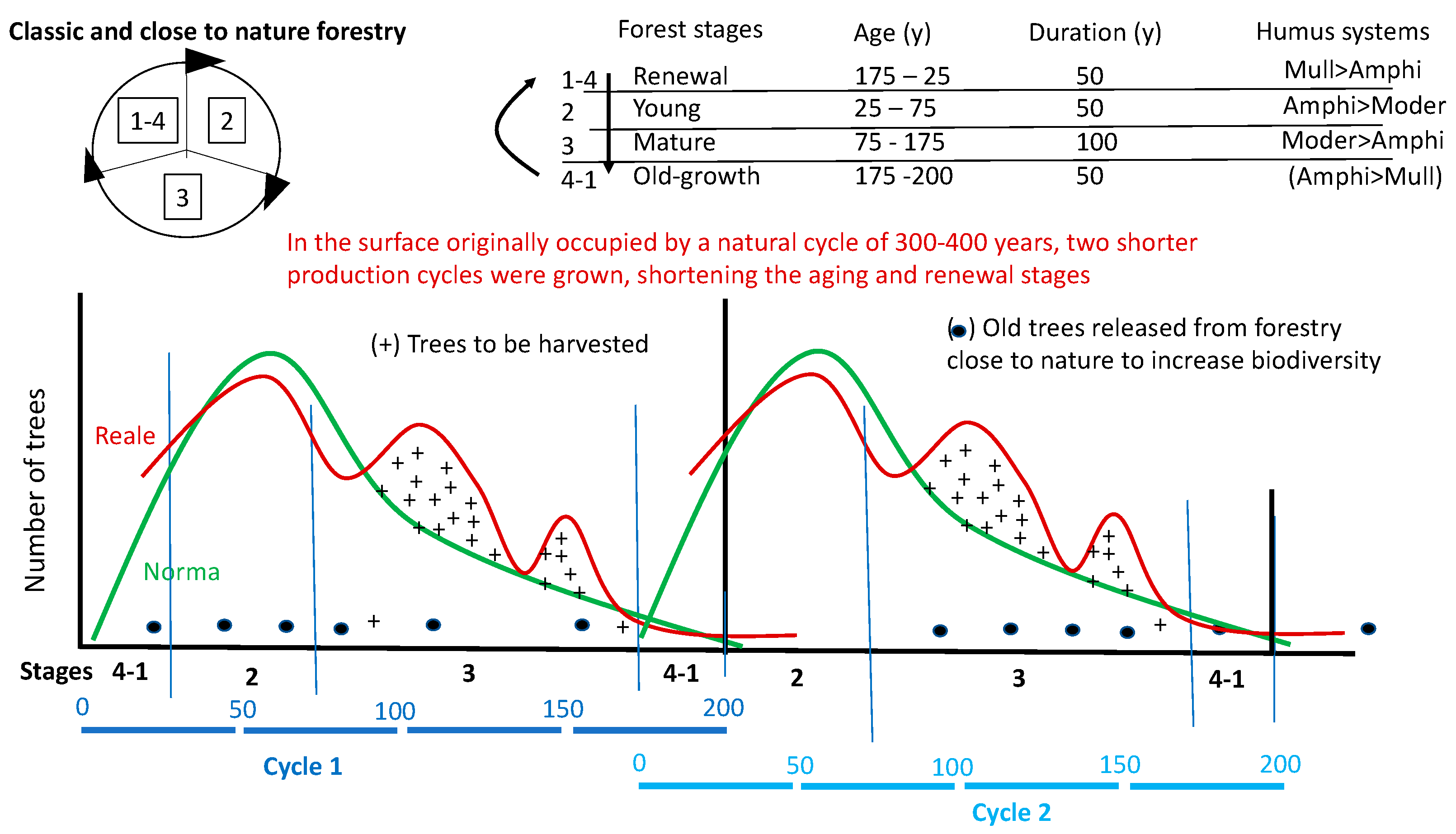
Figure 13.
Soil-conscious forestry involves: (1) promoting the aging of the population; (2) checking the presence of renewal in balance with the biological functionality of the soil; (3) careful sampling of the woody biomass in the humps of the distribution of the number of trees by age class; (4) doubting about the silvicultural action undertaken, and consequently, repeated monitoring of the forest’s response in the years following the felling. For the forest to continue to exist, it is necessary to respect the return to the soil of the nutrients that the trees have taken to grow. In the past, the length of the restitution phase was shortened because it was considered not essential for the functioning of the forest. The few trees of stage 4 correspond to rare individuals scattered among those of the three previous stages, released not for their action on the soil but to increase the biodiversity of the forest44,57,58. The space occupied by a long forestry cycle has been managed in such way as to generate two shorter cycles on the same surface, gaining in terms of volume produced and production times.
Figure 13.
Soil-conscious forestry involves: (1) promoting the aging of the population; (2) checking the presence of renewal in balance with the biological functionality of the soil; (3) careful sampling of the woody biomass in the humps of the distribution of the number of trees by age class; (4) doubting about the silvicultural action undertaken, and consequently, repeated monitoring of the forest’s response in the years following the felling. For the forest to continue to exist, it is necessary to respect the return to the soil of the nutrients that the trees have taken to grow. In the past, the length of the restitution phase was shortened because it was considered not essential for the functioning of the forest. The few trees of stage 4 correspond to rare individuals scattered among those of the three previous stages, released not for their action on the soil but to increase the biodiversity of the forest44,57,58. The space occupied by a long forestry cycle has been managed in such way as to generate two shorter cycles on the same surface, gaining in terms of volume produced and production times.
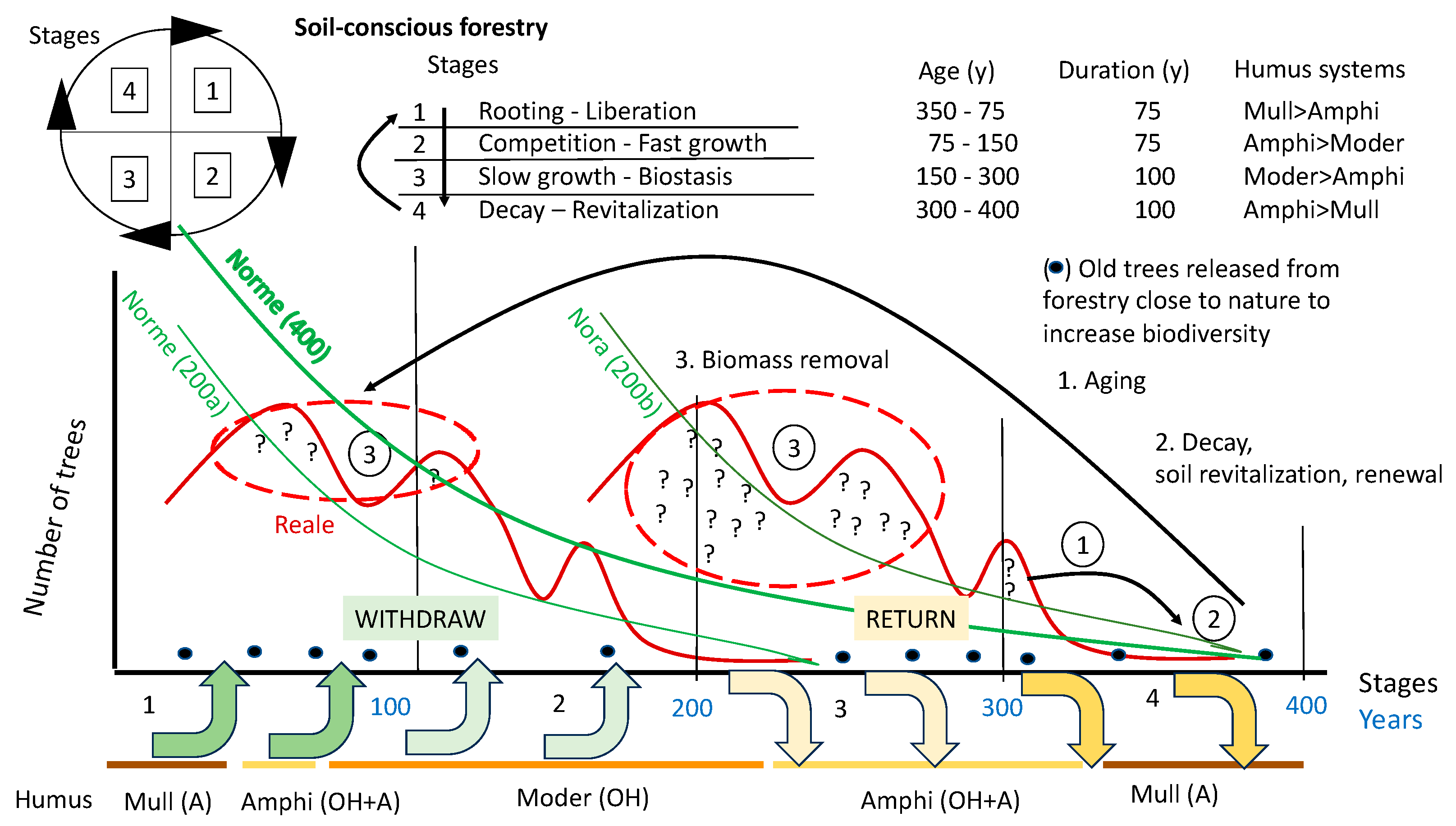
Figure 14.
Left: re-working of the figure on page 4 of PRO SILVA HEL-VETICA, Portrait de la forêt jardinée de l’Envers à Couvet / NE. Harmony that happens in the soil between utility and beauty (https://www.pro-silva-helvetica.ch/pdf/Portrait01_f.pdf ). The figure represents the number of stems per hectare on the ordinate for the list of 5 cm diameter categories on the abscissa. It originally showed the curves corresponding to four historical years (2018, 2002, 1946 and 1896) and also the norms calculated by renowned foresters (Liocourt and Schuetz). Two normal functions have been added to this graph in green (with beech dominant) and in blue (with fir dominant), defined starting from the same height (that of the tallest tree in Envers, 58 m), using two maximum diameters (Dmax) and two coefficients of passage from one 5 cm class to another (K). On the right: the cycle of withdrawal and return that occurs in the soil in harmony with the development of the chronological classes of the high forest, as illustrated in chapter 2.
Figure 14.
Left: re-working of the figure on page 4 of PRO SILVA HEL-VETICA, Portrait de la forêt jardinée de l’Envers à Couvet / NE. Harmony that happens in the soil between utility and beauty (https://www.pro-silva-helvetica.ch/pdf/Portrait01_f.pdf ). The figure represents the number of stems per hectare on the ordinate for the list of 5 cm diameter categories on the abscissa. It originally showed the curves corresponding to four historical years (2018, 2002, 1946 and 1896) and also the norms calculated by renowned foresters (Liocourt and Schuetz). Two normal functions have been added to this graph in green (with beech dominant) and in blue (with fir dominant), defined starting from the same height (that of the tallest tree in Envers, 58 m), using two maximum diameters (Dmax) and two coefficients of passage from one 5 cm class to another (K). On the right: the cycle of withdrawal and return that occurs in the soil in harmony with the development of the chronological classes of the high forest, as illustrated in chapter 2.
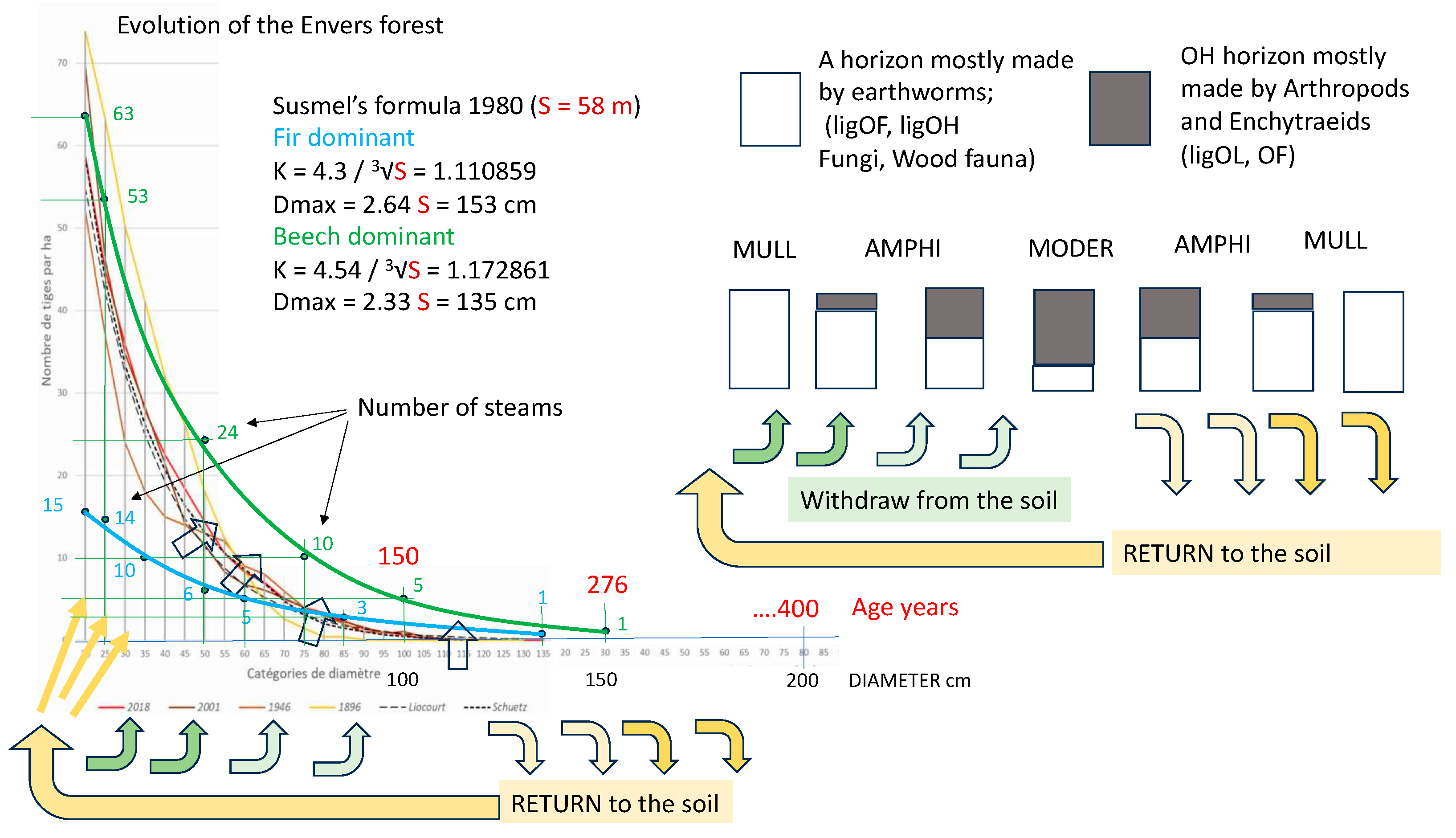
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



