
Submitted:
07 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
The CDPK family genes play crucial roles in signal transduction pathways during plant develop-ment and stress response. In this study, we comprehensively analyzed the CDPK family genes in mustard (Brassica juncea L.), resulting in the identification of 101 genes (BjuCDPK1–101) located on chromosomes AA_Chr01 to BB_Chr08. RNA-seq analysis showed that most BjuCDPK genes were predominantly expressed in the root and flower bud, suggesting their organ-specific expression patterns. Furthermore, the expression levels of most BjuCDPK genes were significantly altered by cold stress. The promoter regions of BjuCDPK genes had abundant cis-acting elements related to hormone and stress. The interaction network analysis predicted that BjuCDPK proteins respond to abiotic stress probably via interaction with ABF1, ABF4, and/or DI19. Our findings provide valu-able information for further investigation of the cold stress adaption of mustard via the CDPK signaling pathway.
Keywords:
mustard (Brassica juncea L.)
; CDPK gene family
; gene expression
; cold stress
1. Introduction
Calcium signature refers to the stimulus-specific increase in cytosolic calcium ion (Ca2+), which serves as a second messenger in various signal transduction pathways and environmental stimulus responses of plants [1,2]. The intracellular modulation and cytosolic concentration of Ca2+ are highly responsive to different abiotic stresses, including drought, high salinity, ultraviolet light, heavy metals, waterlogging, extreme temperature, and wounding [3]. Three classes of calcium sensors, including calmodulins (CaMs) and calmodulin-like proteins (CaMLs), calcineurin B-like proteins (CBLs), and calcium-dependent protein kinases (CDPKs), modulate intracellular Ca2+ transductions in plant signals [4,5,6]. These proteins can be divided into two modes according to the mechanisms of their activation by Ca2+: (1) sensor relay proteins, including CaMs, CaMLs, and CBLs, which have no intrinsic enzymatic activity and interact with sensor responder proteins to transduce signals after binding to Ca2+; (2) sensor responder proteins, unique in CDPKs, which have additional kinase activities besides all the functions of sensor relay proteins [7,8].
The structure of CDPK proteins consists of four domains, including a variable N-terminal domain, a catalytic domain of serine/threonine kinase, a junction domain containing an autoinhibitory region, and a calmodulin-like regulatory domain (CaM-LD) [9]. The N-terminal of CDPKs is highly variable in length and sequence, and sometimes undergoes post-translational modifications, such as myristoylation and palmitoylation, to anchor CDPKs to the membrane of plant cells [10]. In Arabidopsis thaliana, 97.05% (33/34) of CDPKs have a variable N-terminal domain with a length of 25–180 amino acids (aa) that may play an essential role in substrate recognition [11]. The autoinhibitory junction domain acts as an autoinhibitor and contains a pseudosubstrate sequence interacting with the active site and then inhibiting the activity of CDPKs [12]. The catalytic domain, including highly conserved regions such as the ATP-binding site, Ser/Thr protein kinase active site, and 11 subdomains (I-XI subdomains), plays a central role in the function of CDPKs [13]. The C-terminal CaM-LD of CDPKs contains four elongation factor (EF) hands, a helix-loop-helix structure (about 29 aa) that senses calcium signals, and can be directly activated by free Ca2+ binding [14].
CDPK genes were found to distribute in various plant species, including early lower plants (such as green algae), and they were function as transcriptional regulators and play critical roles in regulating of signal transduction and stress responses [15]. For example, overexpression of GmCDPKSK5 enhances the tolerance to high temperature and humidity stress and improves the vitality of soybean seeds [16]. Additionally, AtCPK6 overexpression can improve drought and salt tolerance, but the mutant of AtCPK6 showed no significant difference from the wild type, indicating redundancy of functions among AtCPK genes [17]. Specially, cold stress can trigger the influx of calcium ions into plant cells, leading to physical changes in cellular structure, such as membrane fluidity and cytoskeletal reorganization [18,19]. Previous studies have revealed the positive role of CDPKs in response to cold or freezing tolerance. For instance, cold-induced calcium signals could be rapidly decoded by AtCPK28 and phosphorylated NIN-LIKE PROTEIN 7 (NLP7), which controls the expression of a set of downstream COR genes, and thus enhance the cold tolerance of Arabidopsis plants [20]. In rice, OsCDPK13 is required for signal transduction in response to cold stress and gibberellin (GA) in rice seedlings [21]. OsCPK17 plays an important role in cold tolerance by affecting the activity of membrane channels and sugar and nitrogen metabolism [22]. OsCPK24 is another positive regulator of response to cold stress in rice by participating in the calcium signaling pathway and phosphorylation of OsGrx10 to maintain a higher glutathione level [23]. VaCPK20 from a wild grapevine species (Vitis amurensis Rupr.) was reported to be involved in cold and drought stress response pathways, as overexpression of VaCPK20 in Arabidopsis could increase the expression of several stress-responsive genes [24]. Overexpression of Populus euphratica CPK10 in Arabidopsis could also enhance the freezing tolerance of transgenic lines [25]. In some other cases, CDPKs can play negative roles in regulating of the cold stress response as well. For instance, ZmCPK1 acts as a negative regulator and calcium-independent kinase in response to cold stress, and can suppress the expression of the cold-induced marker gene Zmerf3 in maize [26].
In recent years, more than a dozen of CDPK gene family have been comprehensively identified in various plant species. For instance, genome-wide analysis has identified 40 CDPK genes in maize (Zea mays) [27], 34 in Arabidopsis [28], 31 in pepper (Capsicum annuum) [29], 30 in each of grass (Brachypodium distachyon) and poplar (Populus trichocarpa) [30,31], 29 in each of rice (Oryza sativa) and foxtail millet (Setaria italica) [32,33], 25 in each of canola (Brassica napus) and legume (Medicago truncatula) [34,35], 19 in cucumber (Cucumis sativus) [36], 17 in peach (Prunus persica) [37], and 11 in strawberry (Fragaria x ananassa) [38]. Wild tomato (Solanum habrochaites), which is closely related to cultivated tomato and has strong resistance to cold stress, has been found to possess 33 CDPK genes, whereas the cultivated variety S. lycopersicum only comprised 29 CDPK genes [39,40]. To date, the genomic information and expression patterns of CDPK genes in mustard (Brassica juncea) remain unknown. In this study, we identified the CDPK family genes in mustard, and investigated their phylogeny, gene structure, conserved motifs, chromosomal localization, gene duplication, proteins interaction, and expression levels in different organs and under cold stress. The findings provide a foundation for future research on the roles of CDPK genes in the response of mustard to stresses.
2. Materials and Methods
2.1. Identification of BjuCDPK genes in the genome of Brassica juncea
For the identification of the members in the CDPK gene family, the genome of Brassica juncea cv. tumida T84-66 V2.0 was obtained from the Brassicaceae Database (BRAD V3.0) [41]. A Hidden Markov Model (HMM) search was conducted via the HMMER 3.0 software (E value < 1 × 10-5) with the Pkinase domain (PF00069) and EF-hand 7 domain (PF13499), which were retrieved from the Pfam database (http://pfam.xfam.org/ accessed on 12 December 2022). After that, all putative CDPKs were retested through the NCBI conserved domain database [42]. The ProtParam module in Biopython [43] was used to predict the basic physicochemical characteristics of the BjuCDPK proteins, including protein length (aa), molecular weight (MW), grand average of hydropathy (GRAVY), instability index, and theoretical isoelectric point (pI). The DeepLoc 2.0 online tool [44] was employed to characterize the subcellular location of each BjuCDPK protein.
2.2. Gene structure, chromosomal mapping, and gene duplication analysis
The gene information, including location of exons, introns, and untranslated region of each identified BjuCDPK genes, and their predicted positions mapped onto the Brassica juncea chromosomes was downloaded from BRAD, and visualized by TBtools [45]. The duplication events across the BjuCDPK genes were detected by the WGDI (v0.6.1) software [46], and visualized by shinyCircos [47]. Each of the duplicated BjuCDPK gene pairs was aligned by the ParaAT 2.0 [48] with MUSCLE (v3.8.31) [49] as multiple sequence aligner, and non-synonymous (Ka) and synonymous substitution rates (Ks) were estimated by KaKs_Calculator 3.0 [50] using the MA (model averaging) method.
2.3. Phylogenetic analysis, conserved motif recognition, and promoter analysis
The CDPK protein sequences of Arabidopsis and rice, which were obtained from the Phytozome (https://phytozome.jgi.doe.gov/ accessed on 12 December 2022, reference genome TAIR10) and Rice Genome Annotation Project (http://rice.plantbiology.msu.edu/ accessed on 12 December 2022, reference genome MSU7), respectively, were employed to construct phylogenetic trees with sequences of mustard. CDPK protein sequences from three species (mustard, Arabidopsis, and rice) were aligned with Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/ accessed on 12 December 2022) and then submitted for phylogenetic analysis by MEGA 11 (v11.011). A maximum-likelihood (ML) phylogenetic tree was constructed with 1000 bootstrap replicates. The BjuCDPK protein sequences were submitted to MEME software (https://meme-suite.org/meme/tools/meme accessed on 12 December 2022) for recognition of conserved motifs by setting the number of motifs as 10. To analyze the hormone-responsive and stress-responsive cis-elements in the promoters, the promoter sequences (2 kb upstream of the translation start site) of the BjuCDPK genes were obtained and submitted to the PlantCARE database [51].
2.4. Protein-protein interaction network prediction
Protein-protein interaction network analysis for all 101 BjuCDPK protein sequences was performed by the STRING online tool (https://string-db.org/ accessed on 12 December 2022). The orthologs of Arabidopsis were selected as the reference.
2.5. Plant materials, cold stress treatments, and transcriptome analysis
Mustard seeds were germinated on wet filter paper at 26 °C. After that, the germinated seedlings were transplanted into growth chambers at 14 h/10 h (day/night) cycle at 26 °C. The 21-day old seedlings were exposed to cold stress at 14 h/10 h (day/night) cycle at 4 °C. Leaf samples of seedlings were harvested at 0 h before cold treatment (unstressed seedlings), and 1 h, 3 h, 6 h, 10 h and 24 h after cold stress treatment. Each time point involved three biological replicates. About three micrograms of total RNA isolated by TRIzol Reagent (Invitrogen, Waltham, MA, USA) from each sample was used to generate sequencing libraries by NEBNext® UltraTM RNA Library Prep Kit (New England Biolabs, Ipswich, MA, USA). The sizes of complementary DNA (cDNA) fragments ranged from 250 to 300 bp. Universal PCR primers, Phusion High-Fidelity DNA polymerase, and Index (X) Primer (New England Biolabs, Ipswich, MA, USA) were employed for PCR amplification. Clustering of the index-coded samples was conducted on AcBot Cluster Generation System with TruSeq PE Cluster Kit v3-cBot-HS (Illumina, San Diego, CA, USA). Library sequencing was performed on an Illumina NovaSeq instrument with 150 bp pair-end reads. NGS QC Toolkit (v2.3.3) [52] was used for quality control. The clean reads with high quality were mapped to reference genome Braju_tum_V2.0 using HISAT2 (v2.2.1) [53], and relative expression levels in transcripts per million (TPM) for each gene of each sample were calculated by StringTie (v2.2.0) [54] with default parameters.
2.6. qRT-PCR for expression analysis
Two micrograms of total RNA for each leaf sample of seedlings were reverse transcribed using HiScript® III Reverse Transcriptase (Vazyme Biotech Co., Ltd.). Real-time PCR was conducted on a CFX Connect™ real-time PCR detection system (Bio-Rad Laboratories, Inc.) using ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd.). TIP41 (tonoplastic intrinsic protein 41) was used as the reference gene, and each sample was assessed in triplicate of technical replication. All primers used are listed in Table S1.
3. Results
3.1. Identification and characterization of CDPK family genes in Brassica juncea
In this study, the Hidden Markov Model (HMM) profiles of the Pkinase domain (PF00069) and EF-hand 7 domain (PF13499) were used to characterize putative CDPK proteins. A total of 101 candidate CDPK genes were identified in the Brassica juncea genome (Braju_tum_V2.0). The physicochemical properties of their encoded proteins were analyzed based on their putative amino acid sequences (Table S2). These CDPK genes were designated as BjuCDPK1 to BjuCDPK101 based on their chromosome localization (Figure 1 and Table S2). The Gene Location Visualize of TBtools was used to construct a chromosomal location map. The results showed that the 101 BjuCDPK genes were unevenly distributed on all chromosomes except for AA_Chr08 (Figure 1). Among them, 12 BjuCDPK genes were detected on BB_Chr02, 10 on AA_Chr03, eight each on of BB_Chr05 and BB_Chr08, seven on AA_Chr09, six each on of AA_Chr02, AA_Chr10, and BB_Chr03, five on each of AA_Chr06, AA_Chr07, and BB_Chr07, four each on of AA_Chr01, AA_Chr05, BB_Chr01, BB_Chr04, and BB_Chr06, and three on AA_Chr04 (Figure 1).
The length of the 101 BjuCDPK proteins ranged from 419 aa (BjuCDPK74) to 915 aa (BjuCDPK31), and the MWs ranged from 47.234 kDa (BjuCDPK74) to 104.460 kDa (BjuCDPK31). The pI varied from 4.83 (BjuCDPK8) to 9.34 (BjuCDPK29), with 91 BjuCDPKs being acidic proteins and the remaining being basic proteins. The predicted grand average of hydropathy (GRAVY) values of all BjuCDPK proteins were negative, suggesting that all these proteins are hydrophilic. Instability index analysis revealed that 51.49% (52/101) of BjuCDPK proteins were unstable, while the rest were stable proteins (Table S2). Subcellular localization analysis indicated that 83.17% (84/101) of BjuCDPK proteins were located at the cell membrane, while 16 proteins were located in the cytoplasm, and only one protein (BjuCDPK31) was localized in the nucleus. All BjuCDPK proteins contained EF-hand domains. Most BjuCDPK proteins (86/101) contained four EF-hand domains, while six had three EF-hand domains and nine had only two EF-hand domains. In terms of lipid modification sites, 81 BjuCDPK proteins might have myristoylation sites in their N-terminus and 82 BjuCDPK proteins might contain S-palmitoylation sites (Table S2).
3.2. Phylogenetic relationship of CDPKs in Arabidopsis, rice, and mustard
To investigate the evolutionary relationships of CDPK proteins in different plant species, a phylogenetic tree was constructed for 164 CDPKs, including 29 OsCPKs, 34 AtCPKs, and 101 BjuCDPKs (Figure 2). Based on the structural characteristics of protein sequences and previous classification of CDPK proteins in Arabidopsis, the 101 BjuCDPKs were clustered into four distinct groups, namely Group I to IV [26]. The largest group (Group I) contained 34 BjuCDPKs; Group II comprised 26 BjuCDPKs; Group III included 32 BjuCDPKs; while Group IV had only nine BjuCDPKs. An analysis of the evolutionary relationships indicated that the CDPK proteins in mustard have a closer relationship with those in Arabidopsis than with those in rice (Figure 2).
3.3. Conserved motif and gene structural analysis of BjuCDPKs
To investigate the structural characteristics of the mustard CDPKs, a motif analysis was performed by using the MEME server (Figure S1). A total of 10 different motifs were identified, including six protein kinase and four EF-hand domains. Most BjuCDPK proteins contained all these ten motifs, and most members in the same group contained uniform motif arrangements (Figure 3A, 3B). The exceptions included BjuCDPK74 and BjuCDPK52 in Group I, which were lack of two protein kinase domains, BjuCDPK41 and BjuCDPK99 in Group II, which were lack of three protein kinase domains, BjuCDPK83, which was lack of two protein kinase domains, as well as BjuCDPK18 and BjuCDPK100 in Group III, which were lack of one protein kinase domain. In addition, all members of Group IV contained six protein kinase domains. The number of EF-hand domains in each BjuCDPK protein was presented in Table S2.
The gene structure varied among the 101 BjuCDPKs. The number of exons in the coding sequence (CDS) ranged from five (BjuCDPK5) to 14 (BjuCDPK41). Group III and Group IV members exhibited a similar exon/intron structure in their encoding genes. BjuCDPK genes in Group III harbored seven to eight exons, and those in Group IV had 11 to 13 exons. However, there were considerable differences in the number of exons between Group I (6–11 exons) and Group II (5–14 exons) members (Figure 3A, 3C).
3.4. Gene duplication of BjuCDPKs
The obviously larger number of BjuCDPK genes indicated that the BjuCDPK gene family may undergone significant expansion compared with the family of other plants. To investigate the underlying mechanism, we analyzed the gene duplication patterns of BjuCDPK genes, including tandem and segmental duplications. As shown in Figure 4, a total of 188 segmental duplication events were observed, but no tandem duplication event was detected.
To further understand the selective pressure and evolutionary fates of the CDPK gene pairs resulting from duplication, the nonsynonymous/synonymous substitution ratio (Ka/Ks) was calculated for the full-length gene regions. Generally, a Ka/Ks value greater than 1 indicates positive selection, and a value of 1 indicates neutral selection, while a value lower than 1 indicates purifying or negative selection [55]. In this study, the Ka/Ks values of all 188 BjuCDPK gene pairs resulting from segmental duplication were lower than 0.6 (ranging from 0.028 to 0.476, with an average of 0.123) (Table S3). These results suggested that the BjuCDPK genes have undergone strong purifying selection and slow evolution at the protein level.
3.5. Expression analysis of BjuCDPK genes in various organs of mustard
To reveal the potential biological functions of BjuCDPKs during the growth and development of mustard, the RNA-seq data from different mustard organs, including the leaf, stem, root, flower, pod, pod shell, and seed, were obtained (Figure 5). A total of 83 BjuCDPK genes showed TPM values higher than 1 in at least one of the tested organs. The BjuCDPK genes from different groups exhibited inconsistent expression patterns in different organs, but those closely related BjuCDPKs showed similar expression patterns. The majority of BjuCDPKs exhibited organ-specific expression. The BjuCDPK genes were predominantly expressed in the root and flower bud, suggesting that these genes possibly have important roles in the development of these two organs. In vegetative organs, 33.66% (34/101) of BjuCDPKs showed a root-specific expression pattern, while 12 and five BjuCDPKs were preferentially expressed in the stem and leaf, respectively. In reproductive organs, 16 BjuCDPKs were only expressed in the flower bud, among which six belonged to Group II, and eight and two were clustered in Group III and IV, respectively. At different developmental stages of the pod, seven BjuCDPKs were preferentially expressed at 7 days after pollination (DAP); only two BjuCDPKs were preferentially expressed at 15 DAP; while eight BjuCDPKs showed preferential expression in the pod shell of 20 DAP. No BjuCDPKs showed preferential expression in the seed. These results suggested that the BjuCDPK family genes may be involved in the development of multiple mustard organs.
3.6. Expression analysis of BjuCDPK genes under cold stress
To assess the potential function of BjuCDPK genes in response to cold stress, their transcript accumulation was assessed using RNA-seq data from mustard leaves at 0, 1, 3, 6, 10, and 12 h after exposure to cold stress at 4 °C (Table S4). As a result, 62 BjuCDPKs averagely presented TPM values higher than 1 in at least one sample. These BjuCDPKs were analyzed and clustered into three groups, with 23 BjuCDPKs in Group a, 20 BjuCDPKs in Group b, and 19 BjuCDPKs in Group c (Figure 6A). In Group a, most BjuCDPKs showed up-regulated expression at 6 h after cold treatment, while a few exhibited down-regulated expression. In contrast, most members in Group b exhibited the highest expression levels before cold treatment, while only six members showed increases in expression levels at 6 h after cold treatment. In addition, BjuCDPK genes from both Group a and Group b displayed lower expression levels at 1, 3, and 10 h after cold treatment. In Group c, over half of the members were up-regulated at 10 h, while the remaining members were highly expressed at 0, 1, 3, and 6 h after cold treatment (Figure 6A).
qRT-PCR was performed to validate the expression changes of ten randomly selected BjuCDPK genes under cold stress treatment (Figure 6B). BjuCDPK39, BjuCDPK68, BjuCDPK75, and BjuCDPK90 exhibited similar trends with the highest expression at 6 h, while the expression of BjuCDPK10 and BjuCDPK67 peaked at 10 h. BjuCDPK13, BjuCDPK71, BjuCDPK76, and BjuCDPK96 showed the highest expression level at 24 h.
3.7. Analysis of cis-acting elements of CDPK family genes in mustard
To illustrate how the promoter structure influences the gene expression levels, we extracted and analyzed 2000 bp upstream sequences of 101 BjuCDPKs. A total of 31 types of cis-elements were identified, including 12 hormone-responsive elements and 19 stress-responsive elements (Figure 7). Notably, all BjuCDPK genes contained at least one type of hormone-responsive and stress-responsive elements. For instance, the ABRE element is one of the ABA-responsive elements with a highly conserved ACGTG motif, and 84 BjuCDPK genes contained at least one ABRE element, indicating that they might play crucial roles in ABA signaling response. Two MeJA-responsive elements, CGTCA and TGACG motifs, were present in the promoter regions of 68.32% (69/101) of the BjuCDPK genes. All BjuCDPK genes had the MYB element and almost all BjuCDPKs (except for BjuCDPK84) had the MYC element, suggesting that the BjuCDPK genes may be regulated by MYB and MYC transcription factors. Furthermore, 94 BjuCDPK genes had the anaerobic induction element ARE, and 42 BjuCDPK genes contained at least one low-temperature responsive element LTR. These results indicated that BjuCDPK genes might play vital roles in the growth, development, and environmental stress response of mustard.
3.8. Protein-protein interaction network analysis of BjuCDPKs
To explore the potential interaction and regulatory network between BjuCDPKs and other proteins, we predicted the interaction partners of key BjuCDPK proteins using the STRING software based on the orthologs in Arabidopsis. Overall, three pairs of protein-protein interactions were predicted within BjuCDPKs. BjuCDPK34, BjuCDPK68, and BjuCDPK75 might interact with BjuCDPK101, BjuCDPK96, and BjuCDPK76, respectively. In addition, 53 pairs of protein-protein interactions were predicted between BjuCDPKs and other proteins, including SLAH3 (slow anion channel 1 homolog 3, 12 pairs), ABF4 (ABA-responsive element binding factor 4, 12 pairs), ABF1 (ABA-responsive element binding factor 1, 11 pairs), OZS1 (ozone-sensitive, 11 pairs), and DI19 (dehydration-induced 19, seven pairs) (Figure 8 and Table S5).
4. Discussion
CDPKs are serine/threonine protein kinases playing crucial regulatory roles in the development and stress response of plants. Various numbers of CDPK genes have been identified in different species through bioinformatics analysis of their genomes, such as maize (40 CDPKs), Arabidopsis (34 CDPKs), rice (29 CDPKs), peach (17 CDPKs), and strawberry (11 CDPKs) [27,28,37,38]. In this study, 101 CDPK genes were identified in mustard genome, which were unevenly distributed in different chromosomes, indicating possible genetic variation and species evolution. Gene duplication analysis revealed that 99 out of the 101 BjuCDPK genes (98.02%) are associated with segmental duplication, suggesting that large fragment duplication is the main force driving the expansion of CDPK gene family in mustard. Gene structure analysis revealed that great variations were present in the number of introns and exons among the 101 BjuCDPK genes, indicating that the CDPK family genes might have differential biological functions in mustard.
Previous studies showed that plant CDPK genes have distinct structural features. Based on the evolutionary distance, the 101 BjuCDPKs were clustered into four groups (Group I to IV), all of which were more closely related to AtCPKs than to OsCPKs. The N-terminal of 101 BjuCDPKs varied in length and sequence, and they were predicted to be located at the cell membrane (84 BjuCDPKs), cytoplasm (16 BjuCDPKs), and nucleus (one BjuCDPK). The cytoplasm-located BjuCDPKs consisted of 41 members in Group I and two members in Group II, which had no myristoylation site or S-palmitoylation site in their N-terminus. The only protein located in the nucleus was BjuCDPK31, which had the highest protein length, particularly the length of the N-terminus, and it was lack of myristoylation sites. Typically, the N-terminal domains of CDPKs are characterized by potential N-myristoylation and N-palmitoylation sites, which determine the membrane targeting of plant CDPKs [56]. For example, AtCPK5 is myristoylated and membrane-targeted in Arabidopsis, while the mutation in the myristoylation (G2A) site resulted in a defect in its membrane localization [57]. The catalytic kinase region, including the typical Ser/Thr protein kinase, and the regulatory region containing 1-4 EF-hand domains, were present in all BjuCDPK proteins, which is consistent with the previous speculation that the CDPK genes may have evolved from the fusion of an ancestral CaM kinase gene with a calmodulin gene [58,59].
CDPKs have been reported to coordinate developmental processes and potentially interact with the phytohormone network. In Arabidopsis, AtCPK28 regulates the transition from vegetative growth to reproductive growth and stem elongation during plant development by altering the expression of NAC transcriptional and gibberellic acid homeostasis regulators [60]. OsCPK13 is a callus- and leaf sheath-specific expressed protein with important functions in rice leaf sheath elongation, and its transcription levels and protein accumulation are induced by gibberellins (GA) but suppressed by abscisic acid (ABA) and brassinolide (BL) [61]. In this study, 12 hormone-responsive elements were found in the promoter regions of BjuCDPK genes. The differential gene expression patterns of CDPKs in different organs may suggest their differential roles in plant growth and development. In eight different organs of mustard (such as leaf, stem, root, flower bud, pod at 7 DAP, pod at 15 DAP, pod shell at 20 DAP, and seed at 20 DAP), 83 BjuCDPK genes were expressed in at least one organ as indicated by RNA-seq data analysis and the TPM values were more than 1. Notably, most BjuCDPK genes were preferentially expressed in the roots and flower buds of mustard (Figure 5), indicating that these genes may play important roles in regulating the development of these two organs of mustard.
Cis-element analysis revealed the presence of various hormone- and stress-responsive elements in the promoter regions of BjuCDPK genes, suggesting that these genes may play important roles in response to stress, which is consistent with previous findings in other species [62,63,64,65]. In particular, 42 out of 101 BjuCDPK genes contained at least one LTR element (responding to cold stress) in their promoter regions (Figure 7), and a large number of BjuCDPK genes displayed altered expression under cold stress via RNA-seq data (Figure 6A), indicating their possible roles in response to cold stress. In Solanum habrochaites, 11 ShCDPK family members were found to have LTR element, and the expression of a set of ShCDPK genes changed to a certain extent under cold stress [66]. Similar results were also found in other plants, such as Gossypium barbadense [67], Medicago truncatula [68]. Moreover, the expression of ten seleted BjuCDPK genes were observably induced by cold stress based on qRT-PCR analysis (Figure 6B), further supported the RNA-seq data. And the expression profiles of CDPK orthologous genes in different plants were found to regulated by cold stress, which indicated that the role of CDPK genes in cold stress response is conserved across different plants. As a key Ca2+ sensor group of protein kinase, CDPK proteins play important roles in Ca2+-mediated signaling pathways under cold stress, and the studies of CDPK roles in cold tolerance were also reported in other species, including VaCPK20 in Vitis amurensis [69], MdCPK1a in Malus domestica [70], SpCPK33 in Solanum pennellii [71], as well as in other reports [21,22,23,72,73].
The interaction network prediction by STRING revealed three pairs of protein interactions between BjuCDPK proteins, including BjuCDPK34 and BjuCDPK101, BjuCDPK68 and BjuCDPK96, and BjuCDPK75 and BjuCDPK76. Furthermore, BjuCDPKs were predicted to interact with several other proteins, including SLAH3, ABF4, ABF1, OZS1, and DI19. In the pollen tube tip, phosphorylation by AtCPK2 and AtCPK20 activates the S-type anion channel SLAH3, which regulates pollen tube growth through Ca2+ and cytosolic anion gradients [74]. AtCPKs have been reported to participate in response to stress by interacting with ABF1, ABF4, and DI19 in Arabidopsis. AtCPK32 can be induced by high-salt stress, and its overexpression lines are sensitive to ABA and NaCl during seed germination. AtCPK32 regulates gene expression in response to ABA by phosphorylating ABF4 [75]. Under salt stress, AtCPK4 and AtCPK11 participate in ABA signal transduction by phosphorylating the ABA-responsive transcription factors ABF1 and ABF4 [76]. AtDi19, a stress-induced protein, can also be phosphorylated by AtCPK11 to regulate plant stress response via ABA signal transduction pathway [77]. In this study, five BjuCDPKs (BjuCDPK68, BjuCDPK76, BjuCDPK91, BjuCDPK98, and BjuCDPK101) were predicted to interact with ABF1, ABF4 and/or DI19, while six other BjuCDPKs (BjuCDPK58, BjuCDPK59, BjuCDPK69, BjuCDPK75, BjuCDPK78, and BjuCDPK84) might interact with ABF1 and/or ABF4. Based on the qRT-PCR result that BjuCDPK68, BjuCDPK75, and BjuCDPK76 were up-regulated after cold treatment, it can be speculated that these proteins play crucial roles in response to cold stress as interaction partners of ABF1, ABF4 and/or DI19.
5. Conclusions
In this study, 101 BjuCDPK genes were systematically identified and characterized in mustard. According to the diversity of conserved domains, BjuCDPK proteins could be divided into four groups, in which proteins with close phylogenetic relationships exhibited strong conservation in gene structure and motif organization. Additionally, the BjuCDPK genes harbor many cis-elements responsive to various hormones and stresses. RNA-seq analysis revealed that most BjuCDPK genes were upregulated upon cold stress treatment. Moreover, proteins predicted to interact with BjuCDPKs are associated with abiotic stress responses in plants. Overall, this study provides vital information for further biological functional characterization of BjuCDPK genes and a theoretical basis for improvement of cold stress tolerance of mustard.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Motif analysis of BjuCDPKs using MEME; Table S1: Information of the primers used in this study; Table S2: Identification and characterization of CDPK family genes in Brassica juncea; Table S3: Ka/Ks values of duplicated BjuCDPK gene pairs; Table S4: TPM values of 101 BjuCDPK genes under cold treatment; Table S5: Protein-protein interaction network prediction of BjuCDPKs.
Author Contributions
Data curation, Haibo Li, Hao Wu, Jiaxin Deng, Jiaxian Liu, Weifeng Huang, Chuanhong Li, Cui Mao, Yang Zhang, Yukun Wang, and Jie Zheng; Formal analysis, Yukun Wang and Jie Zheng; Funding acquisition, Haibo Li, Hao Wu, and Jie Zheng; Investigation, Haibo Li, Hao Wu, Jiaxin Deng, Jiaxian Liu, Weifeng Huang, and Chuanhong Li; Methodology, Hao Wu, Jiaxin Deng, Jiaxian Liu, Weifeng Huang, Chuanhong Li, Cui Mao, Yang Zhang, Yukun Wang, and Jie Zheng; Resources, Haibo Li, Weifeng Huang, Chuanhong Li, Cui Mao, and Yang Zhang; Software, Haibo Li, Hao Wu, Yukun Wang, and Jie Zheng; Validation, Jiaxin Deng, Jiaxian Liu; Writing – original draft, Haibo Li, Hao Wu, Yukun Wang, and Jie Zheng; Writing – review & editing, Haibo Li, Hao Wu, Yukun Wang, and Jie Zheng.
Funding
This study is supported by Guangdong Provincial Science and Technology Special Fund Project (210903164532210), Key Projects of Shaoguan University (SZ2019ZK03, SZ2019ZK10), and Doctoral Scientific Research Startup Foundation of Shaoguan University (99000632, 99000631).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.A.R.; McAinsh, M.R.; Taylor, J.E.; Hetherington, A.M. Calcium ions as intracellular second messengers in higher plants. In Advances in Botanical Research. Elsevier Academic Press Inc, 1996; pp. 45-96. [CrossRef]
- Li, Y.; Liu, Y.; Jin, L.; Peng, R. Crosstalk between Ca2+ and other regulators assists plants in responding to abiotic stress. Plants 2022, 11, 1351. [Google Scholar] [CrossRef]
- Romeis, T.; Herde, M. From local to global: CDPKs in systemic defense signaling upon microbial and herbivore attack. Curr. Opin. Plant Biol. 2014, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B-like proteins: calcium sensors for specific signal response coupling in plants. Plant Cell 2002, 14, S389–S400. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; An, L.; Li, W. The CBL-CIPK network mediates different signaling pathways in plants. Plant Cell Rep. 2014, 33, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.F.; Breton, G.; Harmon, A. Decoding Ca2+ signals through plant protein kinases. Annu. Rev. Plant Biol. 2004, 55, 263–288. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ito, T. Structure and function of CDPK: a sensor responder of calcium. In Coding and decoding of calcium signals in plants, S. Luan, Editor., Springer, Berlin, Heidelberg, 2011; pp. 129-146. [CrossRef]
- Breviario, D.; Morello, L.; Giani, S. Molecular cloning of two novel rice cDNA sequences encoding putative calcium-dependent protein kinases. Plant Mol. Biol. 1995, 27, 953–967. [Google Scholar] [CrossRef]
- Ito, T.; Nakata, M.; Fukazawa, J.; Ishida, S.; Takahashi, Y. Alteration of substrate specificity: the variable N-terminal domain of tobacco Ca2+-dependent protein kinase is important for substrate recognition. Plant Cell 2010, 22, 1592–1604. [Google Scholar] [CrossRef]
- Klimecka, M.; Muszynska, G. Structure and functions of plant calcium-dependent protein kinases. Acta. Biochim. Pol. 2007, 54, 219–233. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Q.; Chen, X.; Tian, C.; Wang, X.; Xing, T.; Li, Y.; Wang, Y. Cloning and biochemical properties of CDPK gene OsCDPK14 from rice. J. Plant Physiol. 2005, 162, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.C. Calcium-regulated protein kinases of plants. Gravit. Space Biol. Bull. 2003, 16, 83–90. [Google Scholar] [PubMed]
- Li, M.; Chen, X.; Huang, W.; Li, Y.; Liu, Q.; Yan, W.; Guo, C.; Shu, Y. Genome-wide analysis of the CDPK gene family and their important roles response to cold stress in white clover. Plant Signal Behav. 2023, 18, 2213924. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Hu, H.; Shen, Y.; Zhu, Y.; Liu, X.; Wei, J.; Yu, X.; Liu, S.; Ma, H. GmCDPKSK5 interacting with GmFAD2-1B participates in regulation of seed development in soybean under high temperature and humidity stress. Plant Mol. Biol. Rep. 2022, 40, 402–417. [Google Scholar] [CrossRef]
- Xu, J.; Tian, Y.S.; Peng, R.H.; Xiong, A.S.; Zhu, B.; Jin, X.F.; Gao, F.; Fu, X.Y.; Hou, X.L.; Yao, Q.H. AtCPK6, a functionally redundant and positive regulator involved in salt/drought stress tolerance in Arabidopsis. Planta 2010, 231, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Orvar, B.L.; Sangwan, V.; Omann, F.; Dhindsa, R.S. Early steps in cold sensing by plant cells: the role of actin cytoskeleton and membrane fluidity. Plant J. 2000, 23, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.; Foulds, I.; Singh, J.; Dhindsa, R.S. Cold-activation of Brassica napus BN115 promoter is mediated by structural changes in membranes and cytoskeleton, and requires Ca2+ influx. Plant J. 2001, 27, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Zhang, Y.; Zhong, M.; Hussian, J.; Tang, Y.; Liu, S.; Qi, G. Calcium signaling-mediated transcriptional reprogramming during abiotic stress response in plants. " Theor. Appl. Genet. 2023, 136, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Shen, S.; Yang, S.; Komatsu, S. OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced in response to cold and gibberellin. Plant Physiol. Bioch. 2003, 41, 369–374. [Google Scholar] [CrossRef]
- Almadanim, M.C.; Alexandre, B.M.; Rosa, M.T.G.; Sapeta, H.; Leitao, A.E.; Ramalho, J.C.; Lam, T.T.; Negrao, S.; Abreu, I.A.; Oliveira, M.M. Rice calcium-dependent protein kinase OsCPK17 targets plasma membrane intrinsic protein and sucrose-phosphate synthase and is required for a proper cold stress response. Plant Cell Environ. 2017, 40, 1197–1213. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, C.; Zhu, Y.; Zhang, L.; Chen, T.; Zhou, F.; Chen, H.; Lin, Y. The calcium-dependent kinase OsCPK24 functions in cold stress responses in rice. J. Integr. Plant Biol. 2018, 60, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xue, B.; Xia, X.; Yin, W. A novel calcium-dependent protein kinase gene from Populus euphratica, confers both drought and cold stress tolerance. Biochem. Biophys. Res. Commun. 2013, 441, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, P.; Ehlert, B.; Romeis, T. ZmCPK1, a calcium-independent kinase member of the Zea mays CDPK gene family, functions as a negative regulator in cold stress signalling. Plant Cell Environ. 2015, 38, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Lv, W.; Jiang, S.; Zhang, D.; Cai, G.; Pan, J.; Li, D. Genome-wide identification and expression analysis of calci-um-dependent protein kinase in maize. BMC Genomics 2013, 14, 433. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Willmann, M.R.; Chen, H.C.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef]
- Cai, H.; Cheng, J.; Yan, Y.; Xiao, Z.; Li, J.; Mou, S.; Qiu, A.; Lai, Y.; Guan, D.; He, S. Genome-wide identification and ex-pression analysis of calcium-dependent protein kinase and its closely related kinase genes in Capsicum annuum. Front. Plant Sci. 2015, 6, 737. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Ye, F.; Xiao, Z.; Liao, L.; Li, T.; Jia, M.; Liu, X.; Wu, X. Genome-wide survey and expression analysis of calcium-dependent protein kinase (CDPK) in grass Brachypodium distachyon. BMC Genomics 2020, 21, 53. [Google Scholar] [CrossRef] [PubMed]
- Zuo, R.; Hu, R.; Chai, G.; Xu, M.; Qi, G.; Kong, Y.; Zhou, G. Genome-wide identification, classification, and expression analysis of CDPK and its closely related gene families in poplar (Populus trichocarpa). Mol. Biol. Rep. 2013, 40, 2645–62. [Google Scholar] [CrossRef]
- Yu, T.F.; Zhao, W.Y.; Fu, J.D.; Liu, Y.W.; Chen, M.; Zhou, Y.B.; Ma, Y.Z.; Xu, Z.S.; Xi, Y.J. Genome-wide analysis of CDPK family in foxtail millet and determination of SiCDPK24 functions in drought stress. Front. Plant Sci. 2018, 9, 651. [Google Scholar] [CrossRef]
- Asano, T.; Tanaka, N.; Yang, G.; Hayashi, N.; Komatsu, S. Genome-wide identification of the rice calcium-dependent protein kinase and its closely related kinase gene families: comprehensive analysis of the CDPKs gene family in rice. Plant Cell Physiol. 2005, 46, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhang, Y.; Li, F.; He, X.; Zhang, X.; Yi, Y. Bioinformatic analysis of calcium-dependent protein kinase family of Medicago truncatula. Mol. Plant Breeding 2018, 9, 26–35. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, W.Z.; Zhang, Y.; Deng, M.; Niu, F.; Yang, B.; Wang, X.; Wang, B.; Liang, W.; Deyholos, M.K.; Jiang, Y.Q. Identification, expression and interaction analyses of calcium-dependent protein kinase (CPK) genes in canola (Brassica napus L.). BMC Genomics 2014, 15, 211. [Google Scholar] [CrossRef]
- Xu, X.; Liu, M.; Lu, L.; He, M.; Qu, W.; Xu, Q.; Qi, X.; Chen, X. Genome-wide analysis and expression of the calcium-dependent protein kinase gene family in cucumber. Mol. Genet. Genomics 2015, 290, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xie, B.; Hou, Y.; Zhao, Y.; Zheng, Y.; Jin, P. Genome-wide identification of the CDPK gene family reveals the CDPK-RBOH pathway potential involved in improving chilling tolerance in peach fruit. Plant Physiol. Biochem. 2022, 191, 10–19. [Google Scholar] [CrossRef]
- Crizel, R.L.; Perin, E.C.; Vighi, I.L.; Woloski, R.; Seixas, A.; da Silva Pinto, L.; Rombaldi, C.V.; Galli, V. Genome-wide iden-tification, and characterization of the CDPK gene family reveal their involvement in abiotic stress response in Fragaria x ananassa. Sci. Rep. 2020, 10, 11040. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Liang, S.; Chen, X.; Liu, J.; Zhang, Y.; Wang, A. Identification of CDPK gene family in Solanum habrochaites and its function analysis under stress. Int. J. Mol. Sci. 2022, 23, 4227. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Xu, Y.P.; Munyampundu, J.P.; Liu, T.Y.; Cai, X.Z. Calcium-dependent protein kinase (CDPK) and CDPK-related kinase (CRK) gene families in tomato: genome-wide identification and functional analyses in disease resistance. Mol. Genet. Genomics 2016, 291, 661–76. [Google Scholar] [CrossRef]
- Chen, H.; Wang, T.; He, X.; Cai, X.; Lin, R.; Liang, J.; Wu, J.; King, G.; Wang, X. BRAD V3.0: an upgraded Brassicaceae da-tabase. Nucleic Acids Res. 2022, 50, D1432–D1441. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; Yang, M.; Zhang, D.; Zheng, C.; Lanczycki, C.J.; Marchler-Bauer, A. CDD/SPARCLE: the conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Cock, P.J.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; de Hoon, M.J. Biopython: freely available Python tools for computational molecular biology and bioinformatics. Bioin-formatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Sonderby, C.K.; Sonderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: prediction of protein sub-cellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: an integrative toolkit developed for in-teractive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Jiao, B.; Yang, Y.; Shan, L.; Li, T.; Li, X.; Xi, Z.; Wang, X.; Liu, J. WGDI: A user-friendly toolkit for evolutionary analyses of whole-genome duplications and ancestral karyotypes. Mol. Plant 2022, 15, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ouyang, Y.; Yao, W. shinyCircos: an R/Shiny application for interactive creation of Circos plot. Bioinformatics 2018, 34, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, J.; Wu, J.; Zhang, H.; Liu, G.; Wang, X.; Dai, L. ParaAT: a parallel tool for constructing multiple protein-coding DNA alignments. Biochem. Bioph. Res. Co. 2012, 419, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z. KaKs_Calculator 3.0: calculating selective pressure on coding and non-coding sequences. Genom. Proteom. Bioinf. 2022, 20, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic. Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Liu, Z.; Patel, R.K.; Jain, M. NGS QC Toolkit: a toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Long, M.; Fan, C. gKaKs: the pipeline for genome-level Ka/Ks calculation. Bioinformatics 2013, 29, 645–646. [Google Scholar] [CrossRef]
- Singh, A.; Sagar, S.; Biswas, D.K. Calcium dependent protein kinase, a versatile player in plant stress management and development. Crit. Rev. Plant Sci. 2018, 36, 336–352. [Google Scholar] [CrossRef]
- Lu, S.X.; Hrabak, E.M. The myristoylated amino-terminus of an Arabidopsis calcium-dependent protein kinase mediates plasma membrane localization. Plant Mol. Biol. 2013, 82, 267–278. [Google Scholar] [CrossRef]
- Harmon, A.C.; Gribskov, M.; Gubrium, E.; Harper, J.F. The CDPK superfamily of protein kinases. New Phytol. 2001, 151, 175–183. [Google Scholar] [CrossRef]
- Suen, K.L.; Choi, J.H. Isolation and sequence analysis of a cDNA clone for a carrot calcium-dependent protein kinase: homology to calcium/calmodulin-dependent protein kinases and to calmodulin. Plant Mol. Biol. 1991, 17, 581–590. [Google Scholar] [CrossRef]
- Matschi, S.; Werner, S.; Schulze, W.X.; Legen, J.; Hilger, H.H.; Romeis, T. Function of calcium-dependent protein kinase CPK28 of Arabidopsis thaliana in plant stem elongation and vascular development. Plant J. 2013, 73, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Onodera, H.; Toki, S.; Tanaka, H.; Komatsu, S. OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced by cold and gibberellin in rice leaf sheath. Plant Mol. Biol. 2004, 55, 541–552. [Google Scholar] [CrossRef]
- Chen, F.; Fasoli, M.; Tornielli, G.B.; Dal Santo, S.; Pezzotti, M.; Zhang, L.; Cai, B.; Cheng, Z.M. The evolutionary history and diverse physiological roles of the grapevine calcium-dependent protein kinase gene family. PLoS One 2013, 8, e80818. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, D.; Wang, L.; Pan, J.; Liu, Y.; Kong, X.; Zhou, Y.; Li, D. A maize calcium-dependent protein kinase gene, ZmCPK4, positively regulated abscisic acid signaling and enhanced drought stress tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2013, 71, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Vivek, P.J.; Tuteja, N.; Soniya, E.V. CDPK1 from ginger promotes salinity and drought stress tolerance without yield penalty by improving growth and photosynthesis in Nicotiana tabacum. PLoS One 2013, 8, e76392. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Baldrich, P.; Messeguer, J.; Lalanne, E.; Coca, M.; San Segundo, B. Overexpression of a calcium-dependent protein kinase confers salt and drought tolerance in rice by preventing membrane lipid peroxidation. Plant Physiol. 2014, 165, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Liang, S.; Chen, X.; Liu, J.; Zhang, Y.; Wang, A. Identification of CDPK gene family in Solanum habrochaites and its function analysis under stress. Int. J. Mol. Sci. 2022, 23, 4227. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Zhu, X. Genome-wide identification and functional characterization of CDPK gene family reveal their involvement in response to drought stress in Gossypium barbadense. PeerJ 2022, 10, e12883. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Liu, Y.; Kong, W.; Ji, J.; Cai, T.; Guo, Z. Genome-wide identification and characterization of calcium-dependent protein kinase (CDPK) and CDPK-related kinase (CRK) gene families in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 1044. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef]
- Dong, H.; Wu, C.; Luo, C.; Wei, M.; Qu, S.; Wang, S. Overexpression of MdCPK1a gene, a calcium dependent protein kinase in apple, increase tobacco cold tolerance via scavenging ROS accumulation. PLoS One 2020, 15, e0242139. [Google Scholar] [CrossRef]
- Hu, J.; Wang, B.; Yang, T.; Li, N.; Yang, H.; Yu. Q.; Wang J. A calcium-dependent protein kinase gene SpCPK33 from Solanum pennellii associated with increased cold tolerance in tomato. J. Plant Physiol. 2022, 279, 153834. [Google Scholar] [CrossRef]
- Saijo, Y.; Kinoshita, N.; Ishiyama, K.; Hata, S.; Kyozuka, J.; Hayakawa, T.; Nakamura, T.; Shimamoto, K.; Yamaya, T.; Izui, K. A Ca2+-dependent protein kinase that endows rice plants with cold- and salt-stress tolerance functions in vascular bundles. Plant Cell Physiol. 2001, 42, 1228–1233. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, H.; Wu, S.; Fu, D.; Li, M.; Gong, Z.; Yang, S. CPK28-NLP7 module integrates cold-induced Ca2+ signal and transcriptional reprogramming in Arabidopsis. Sci. Adv. 2022, 8, eabn7901. [Google Scholar] [CrossRef] [PubMed]
- Gutermuth, T.; Lassig, R.; Portes, M.T.; Maierhofer, T.; Romeis, T.; Borst, J.W.; Hedrich, R.; Feijo, J.A.; Konrad, K.R. Pollen tube growth regulation by free anions depends on the interaction between the anion channel SLAH3 and calcium-dependent protein kinases CPK2 and CPK20. Plant Cell 2013, 25, 4525–4543. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.I.; Park, H.J.; Park, J.H.; Kim, S.; Im, M.Y.; Seo, H.H.; Kim, Y.W.; Hwang, I.; Kim, S.Y. Arabidopsis calcium-dependent protein kinase AtCPK32 interacts with ABF4, a transcriptional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol. 2005, 139, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Y.; Yu, X.C.; Wang, X.J.; Zhao, R.; Li, Y.; Fan, R.C.; Shang, Y.; Du, S.Y.; Wang, X.F.; Wu, F.Q.; Xu, Y.H.; Zhang, X.Y.; Zhang, D.P. Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Ar-abidopsis. Plant Cell 2007, 19, 3019–3036. [Google Scholar] [CrossRef]
- Rodriguez Milla, M.A.; Uno, Y.; Chang, I.F.; Townsend, J.; Maher, E.A.; Quilici, D.; Cushman, J.C. A novel yeast two-hybrid approach to identify CDPK substrates: characterization of the interaction between AtCPK11 and AtDi19, a nuclear zinc finger protein. FEBS Lett. 2006, 580, 904–911. [Google Scholar] [CrossRef]
Figure 1.
Distribution of 101 BjuCDPK genes in mustard genome. Chromosome numbers are listed above and the chromosome sizes are indicated on the left of the figure. The length of each chromosome was estimated in mega base (Mb).
Figure 1.
Distribution of 101 BjuCDPK genes in mustard genome. Chromosome numbers are listed above and the chromosome sizes are indicated on the left of the figure. The length of each chromosome was estimated in mega base (Mb).
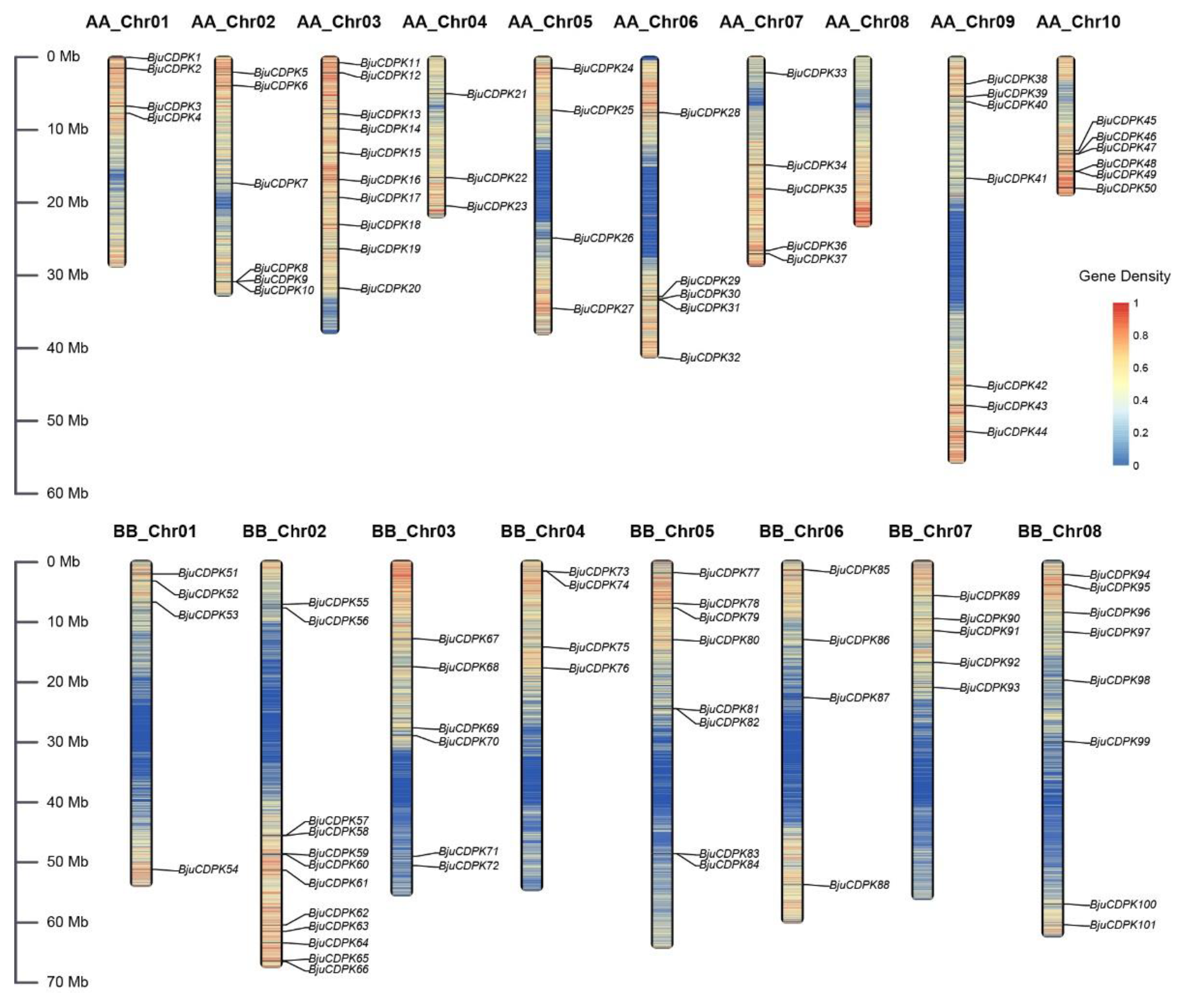
Figure 2.
Phylogenetic analysis of the CDPKs in Arabidopsis thaliana, rice (Oryza sativa), and mustard (Brassica juncea). A total of 165 CDPKs were classified into four distinct groups (Group I to IV) based on their protein sequence structural characteristics. The red triangles, blue cycles, and green squares represent AtCPKs in Arabidopsis, OsCPKs in rice, and BjuCDPKs in mustard, respectively.
Figure 2.
Phylogenetic analysis of the CDPKs in Arabidopsis thaliana, rice (Oryza sativa), and mustard (Brassica juncea). A total of 165 CDPKs were classified into four distinct groups (Group I to IV) based on their protein sequence structural characteristics. The red triangles, blue cycles, and green squares represent AtCPKs in Arabidopsis, OsCPKs in rice, and BjuCDPKs in mustard, respectively.
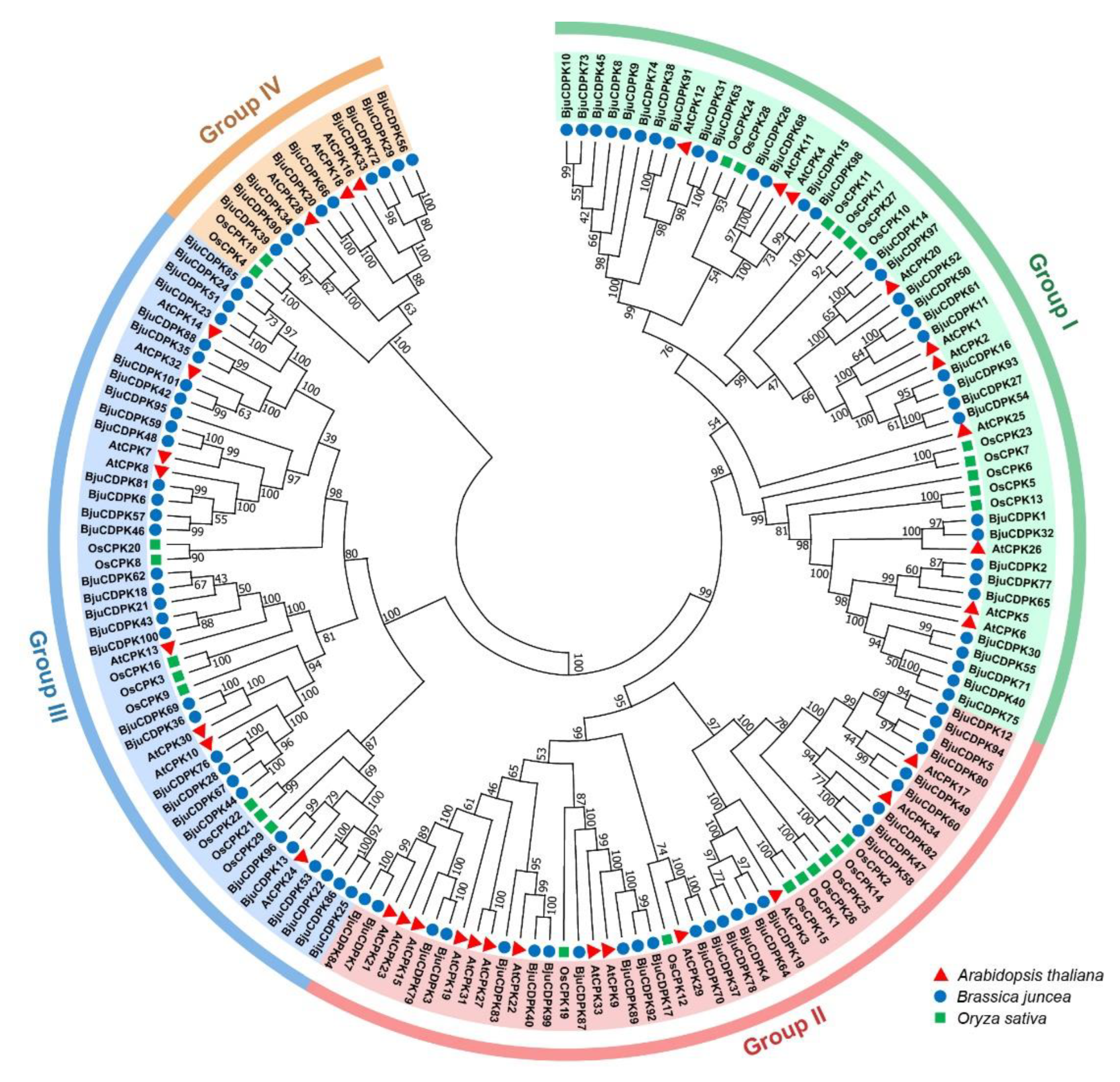
Figure 3.
Phylogenetic analysis, conserved motifs, and structure of encoding genes of BjuCDPKs. (A) Motif composition and distribution of 101 BjuCDPK proteins based on the phylogenetic relationship analysis. (B)Ten motifs are indicated by different colors. The lengths of the boxes and lines are proportional to protein lengths. (C) Structure analysis of BjuCDPKs genes. UTRs and exons are indicted by green and yellow boxes, respectively, and introns are shown by black lines. The lengths of the boxes and lines are proportional to gene lengths.
Figure 3.
Phylogenetic analysis, conserved motifs, and structure of encoding genes of BjuCDPKs. (A) Motif composition and distribution of 101 BjuCDPK proteins based on the phylogenetic relationship analysis. (B)Ten motifs are indicated by different colors. The lengths of the boxes and lines are proportional to protein lengths. (C) Structure analysis of BjuCDPKs genes. UTRs and exons are indicted by green and yellow boxes, respectively, and introns are shown by black lines. The lengths of the boxes and lines are proportional to gene lengths.

Figure 4.
Synteny analysis of interchromosomal relationships of BjuCDPK genes. BjuCDPK gene pairs in the B. juncea genome are indicated by black lines.
Figure 4.
Synteny analysis of interchromosomal relationships of BjuCDPK genes. BjuCDPK gene pairs in the B. juncea genome are indicated by black lines.
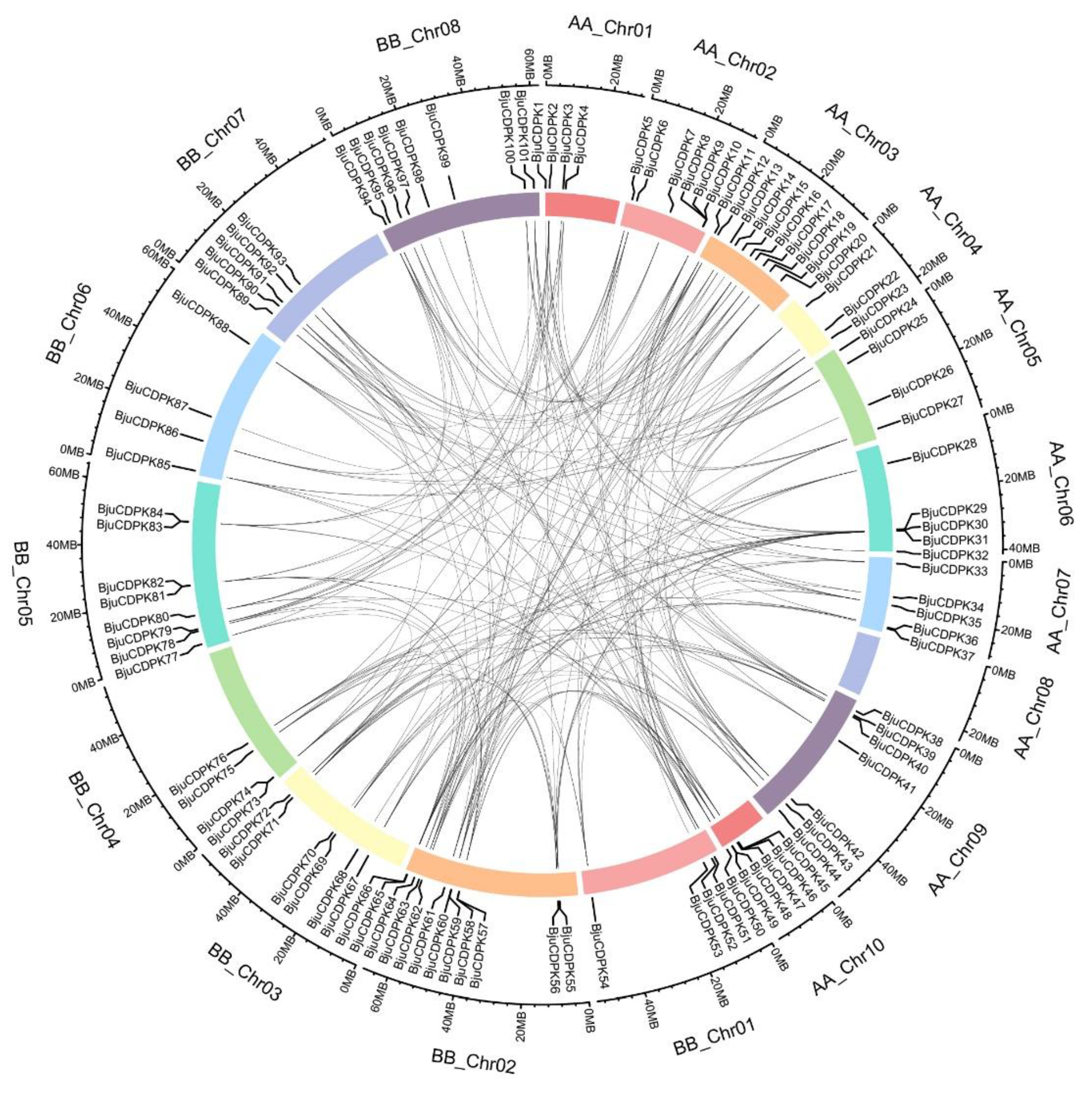
Figure 5.
Expression patterns of 101 BjuCDPK genes in the leaf, stem, root, flower bud, and pod at different developmental stages. The relative expression levels were determined by RNA-seq data and normalized to 0–1.
Figure 5.
Expression patterns of 101 BjuCDPK genes in the leaf, stem, root, flower bud, and pod at different developmental stages. The relative expression levels were determined by RNA-seq data and normalized to 0–1.
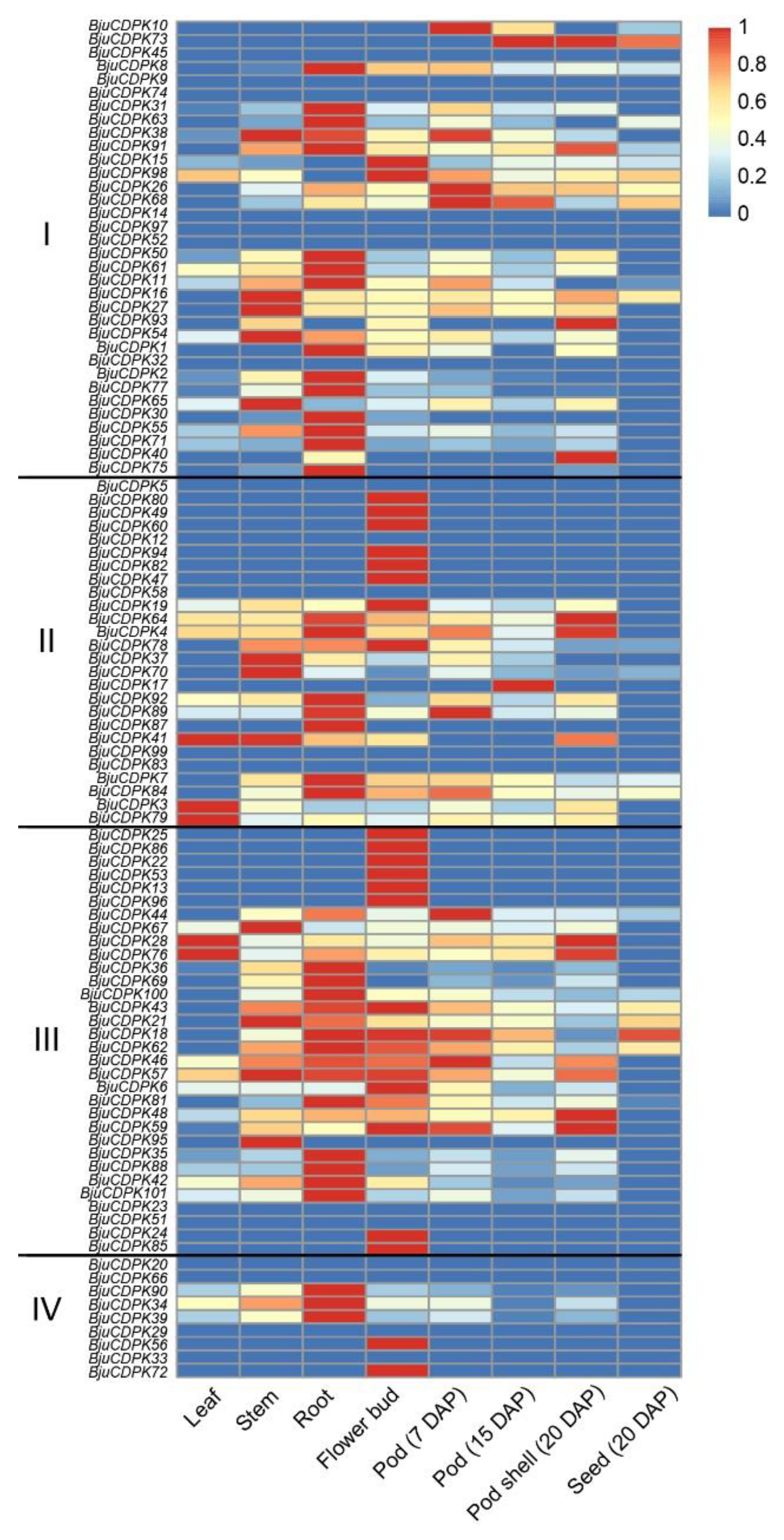
Figure 6.
Expression patterns of BjuCDPK genes under cold stress. (A). Heatmap of expression profiles of BjuCDPK genes in seedlings under cold (4 °C) treatment, and the TPM value for each gene was normalized to 0–1. (B). qRT-PCR validation of ten selected BjuCDPK genes under cold stress.
Figure 6.
Expression patterns of BjuCDPK genes under cold stress. (A). Heatmap of expression profiles of BjuCDPK genes in seedlings under cold (4 °C) treatment, and the TPM value for each gene was normalized to 0–1. (B). qRT-PCR validation of ten selected BjuCDPK genes under cold stress.
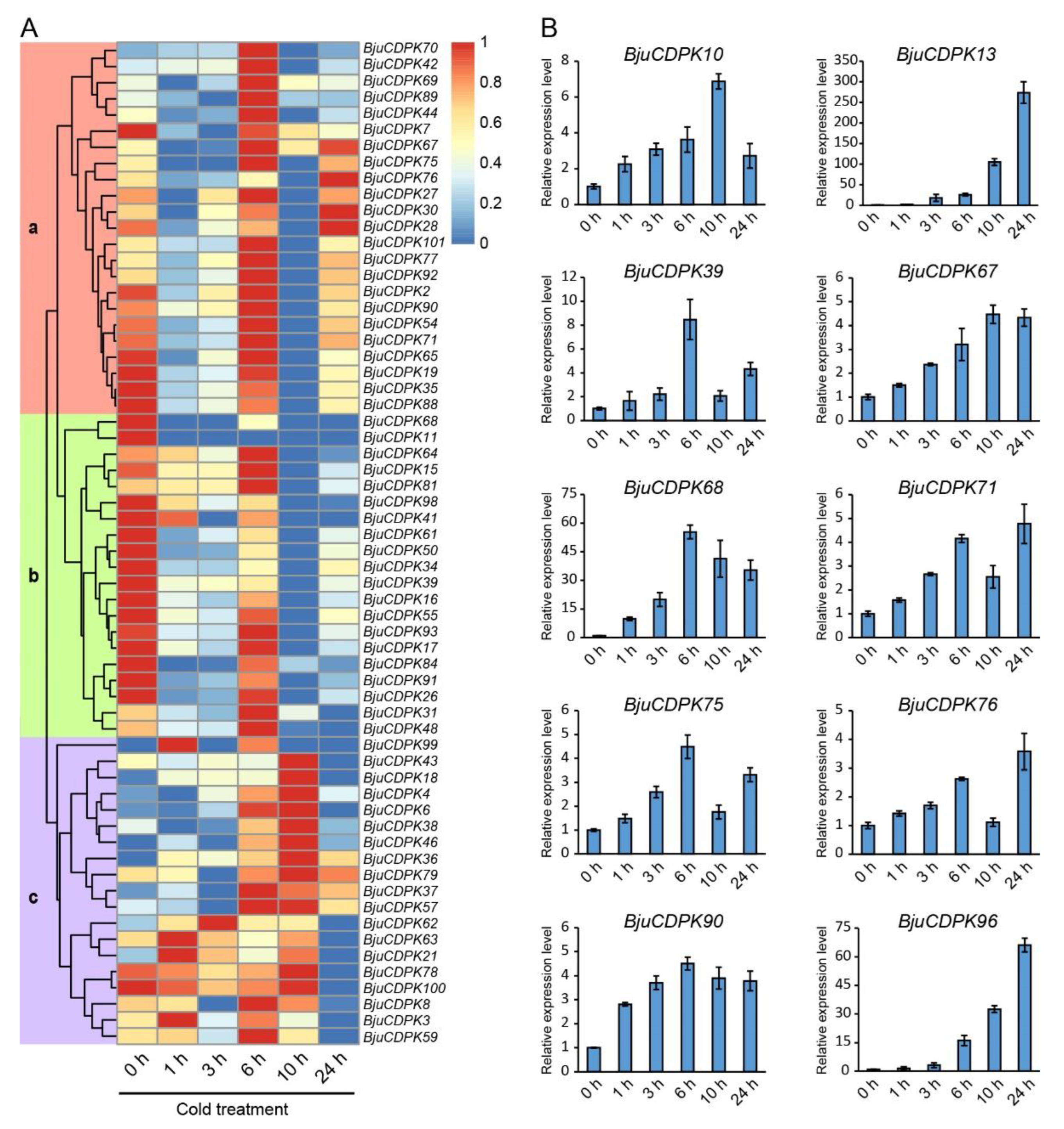
Figure 7.
Numbers of 12 types of hormone-responsive elements and 19 types of stress-responsive elements in BjuCDPK gene promoter regions.
Figure 7.
Numbers of 12 types of hormone-responsive elements and 19 types of stress-responsive elements in BjuCDPK gene promoter regions.
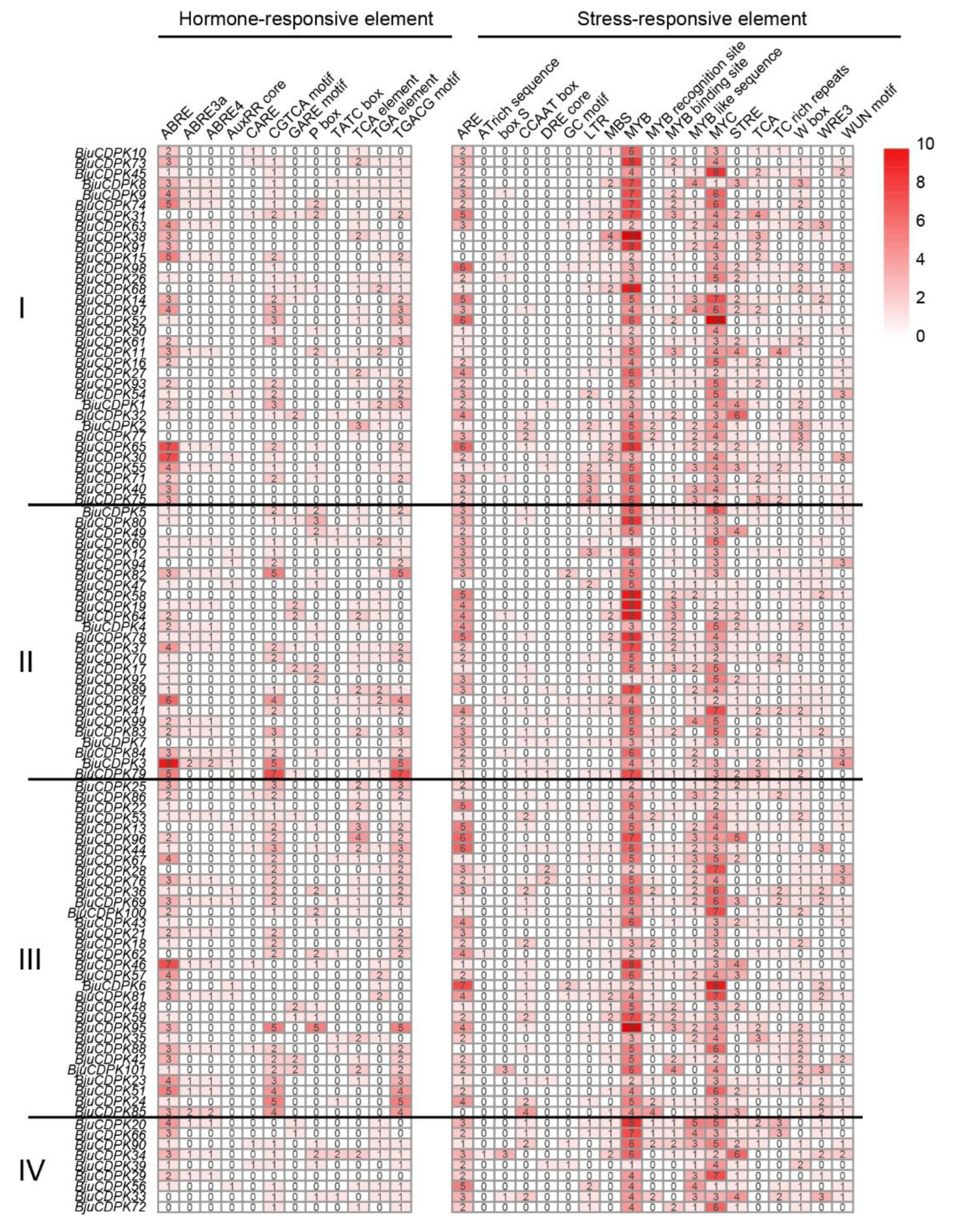
Figure 8.
Protein-protein interaction network of BjuCDPKs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



