
Submitted:
07 November 2023
Posted:
08 November 2023
You are already at the latest version
Abstract
Preterm birth (PTB), a multi-causal syndrome, is one of the global epidemics. Maternal nutrition, but also neonatal and placental telomere length (TL), are among the factors affecting PTB risk. However, the exact relationship between these factors and the PTB outcome, remains obscure. The aim of this review, was to investigate the association between PTB, maternal nutrition and placental-infant TL. Observational studies were sought, with the keywords: maternal nutrition, placenta TL, newborn, TL, and PTB. No study was found that included all keywords simultaneously, and thus, the keywords, were searched in dyads, to reach assumptive conclusions. The findings show that maternal nutrition affects PTB risk, through its influence on maternal TL. On the other hand, maternal TL independently affects PTB risk, and at the same time PTB is a major determinant of offspring TL regulation. The strength of the associations and the extent of the influence from covariates, remains to be elucidated in future research. Furthermore, the question of whether maternal TL, is simply a biomarker of maternal nutritional status and PTB risk, or a causative factor of PTB, to date, remains to be answered.
Keywords:
maternal nutrition
; placenta telomeres
; newborn telomeres
; premature birth
; preterm infants
1. Introduction
Preterm birth (PTB), is defined as the birth whose onset occurs before the 37th week of pregnancy [1]. It commences automatically with the presence of one or more, of the following events: uterine contractions without rupture of membranes, premature rupture of membranes, induction of labor or the need of a caesarean section. Induction of preterm labor occurs in the presence of pregnancy-related-pathology, such as pre-eclampsia and intrauterine growth restriction (IUGR), where the fetuses’ delivery is necessary and/or urgent [2].
PTB is truly a global problem. An estimated 13.4 million infants were born prematurely in 2020, which translates to more than 1 in 10 babies, with the highest percentages, reported in southern Asia and sub-Saharan Africa [1]. Adding to the dim statistic, most countries show an increased rate of PTBs, over the last 20 years [3]. Prematurity is considered a multi-causal syndrome and one of the global epidemics [3]. Goldenberg et al. [2] identified prematurity as a syndrome, due to the contribution of multiple possible causes for its spontaneous onset, including: infection or inflammation, vascular diseases, uterine hypertension (e.g., twin pregnancy, polyhydramnios) etc.
The multiple risk factors implicated in premature labor, include, lifestyle, heredity, anthropometric characteristics, multiple pregnancy, and maternal age [4]. Other main causes of PTB receiving increased scientific interest recently include, placental premature cellular aging and/or a dysfunctional placenta [5]. Moreover, the cervixes’ short length in the second trimester of pregnancy (< 2.50 cm) and the increased concentration of fetal fibronectin in the cervical smear, are possible prognostic factors of spontaneous onset of PTB [4,6].
Even to this day, with the technological and research advances recorded, the causes, and mechanisms of the spontaneous onset of both preterm and full-term birth, remain poorly understood [1,7]. It is thought however, that the spontaneous onset of both, term- and preterm labor, share the same mechanism. The difference is that the onset of full term labor, is the result of a physiological maturation process, while the premature one, derives from a pathological process that activates one or more of the above stages of the mechanism [8].
The placenta which develops throughout pregnancy, has a key role in a variety of functions including labor outcome. It acts as an endocrine gland, an immunomodulatory organ, it is part of the fetal circulatory system, while it also participates in the nutrient / metabolite and oxygen transportation to and from the fetus [9]. Its anatomical formation separates the fetal from the maternal circulation, thus highlighting the importance of “filtering” both the macro- and micro-nutrient transportation [10], while the placenta’s fetomaternal nutrient transportation ability, depends on its size, weight and morphology [11,12,13,14]. However, the mechanism, in essence, is much more complex.
As the pregnancy progresses, the placenta undergoes aging-related-changes. During this process, its’ cells display reduced functionality, mitochondrial dysfunction and finally apoptosis. Placental aging, is a normal phenomenon, which was first described in the 70s, but only during the latest decade has the association between placenta aging and the various changes during pregnancy, been studied. Nevertheless, in some cases the placental cells’ aging occurs prematurely, disrupting its normal function [15,16]. Various factors may affect the aging pace of the placenta, proper fetal development and birth outcomes, including maternal dietary intake and nutritional status [17,18,19,20]. Not surprisingly, maternal nutritional status - among others, though its effect on the placenta - is associated with the onset of spontaneous PTB, low birth weight and irregular fetal growth [11,21,22,23,24,25,26,27,28,29,30]. In the majority of studies, beneficial dietary patterns that are often characterised by high consumption nutrient dense foods including vegetables, fruits, whole grains, fish and dairy products, appear to have a synergistic effect on reducing the risk of PTB. Unhealthy dietary patterns, on the other hand - such as the ones characterized by high intakes of refined grains, processed meat, and foods high in saturated fat or sugar - are associated with a higher risk of PTB [11]. However, the exact mechanism/s with which the characteristics of the diet (e.g. quality, quantity, intake timing and frequency) affect risk of PTB, is a complex research matter still under investigation.
Another factor, both linked to placental aging pace and PTB is telomere length (TL). Telomeres, used as potential indicators of the aging pace and disease burden, are repetitive, non-coding DNA sequences found at the ends of eukaryotic chromosomes that protect the integrity of DNA information throughout the cell cycle, preventing loss of DNA during cell division [31]. Telomeres are susceptible to oxidative stress damage, genetic/epigenetic and environmental factors including unfavourable nutritional habits, leading to premature length reduction and the subsequent development of chronic diseases and/or premature death [32,33].
Pregnancy is on its own characterized as a state of high oxidative stress. Indeed, as the metabolic demands of the developing foetus increase, the reactive oxygen’s species (ROS) production rises. However, a persistent, intense, and premature oxidative stress affects the placenta’s antioxidant capacity, followed by an accumulation of ROS and damage to lipids, proteins, and placenta’s DNA. Inductively, the placental cells, undergo aging and erosion of their TLs. Shorter TL has been associated with pregnancy complications, such as maternal and fetal mortality, preeclampsia, IUGR and PTB [34]. Additionally, placenta TL of PTBs, are reported to be shorter compared with the placentas of full-term pregnancies. This mechanism speculates, that the TL is associated with the premature placenta’s aging pace, as well as the adverse outcome of premature birth [35].
In recent years, TL and telomerase (i.e. the enzyme responsible for maintenance of the length of telomeres by addition of guanine-rich repetitive sequences), have been investigated as possible biomarkers of premature placental aging [15,16]. Physiologically, the cells’ division is accompanied by TL shortening that leads towards the aging trajectory. On the other hand, the enzyme telomerase is active in ensuring the preservation of placenta TL and subsequently its normal function, until the end of pregnancy. This balanced process, can be affected by various aggravating factors mentioned above (e.g. oxidative stress, inflammation and other genetic/epigenetic, immunological, physiological, lifestyle and environmental factors including nutrition), leading to an advanced aging pace and the consecutive dysfunction of the placenta [15,36]. As a result, complications during pregnancy, primarily gestational diabetes mellitus (GDM), preeclampsia, intrauterine growth restriction (IUGR), PTB, intrauterine death, arise [35,37]. Indeed, placentas of complicated pregnancies with preeclampsia, intrauterine growth retardation, are characterised by decreased activity of telomerase and generalised dysfunctionality of TL regulation. In complicated pregnancies, a short placental TL is recorded, but no similar findings are evident in the umbilical cord’s TLs. On the contrary, molar pregnancies have shown an increased telomerase activity, similar to the telomerase activity recorded in malignant cells and tissues [38].
The effect of the above influences is depicted in the offspring, and although the majority of prior telomere research has focused on adults, TL attrition in early life may be particularly important for lifelong health status, as evident from studies of childhood adversities and their effect on adult health trajectory [39].The importance of early life TL programming, is further evident, if one considers that TL in adulthood, is determined by both TL at birth (which serves as an “initial setting”) and by its subsequent attrition during development [40].
Since TL shortening is a biological phenomenon associated with many adverse health conditions, including various complications associated with pregnancy [41] and subsequent offspring health trajectory, it may serve as a reflection of the cumulative impact of stressors and/or as part of a fetal programming mechanism [42]. Indeed, newborn TL, seen as the primary setting of TL regulation, although highly variable among individuals [42], it carries important lifetime consequences for telomere dynamics and longevity [43,44]. Recent evidence regarding newborn TL, suggest plasticity exists in both the programming of telomere biology as well as, the initial setting of TL after birth [45]. The “in utero” environment appears as a significant contributor to this effect. Indeed, cord blood TL association studies, with specific fetal exposures to maternal nutrient status, e.g., folate [46] and vitamin D [47], smoking status [48] maternal educational profile [49], or maternal metabolic status [50,51,52] , and parental age [52,53,54,55,56], have provided insights into the mechanism of TL regulation and its trajectory following birth. Maternal stress [57], stress hormones [58], adverse early life events [39] and other related factors have been correlated with TL and point out the importance of in utero and early developmental exposures in the regulation of offspring TL in adult life. As shown by Gotlib et al. [59] chronic stress is a major determinant of telomere maintenance, both by direct exposure or through pathways of mother to fetus/infant transmission in early life.
Previous studies have shown that premature infants exhibit longer telomeres at birth [60,61], but shorter telomeres in young adulthood compared with term-born infants [62,63], suggesting that a more rapid attrition occurs in early life [64,65]. From this perspective, telomere regulation might be a key programming and compensatory mechanism, in premature infants. Furthermore, postnatal exposure to adversities associated with preterm birth, are a cause for enhanced TL attrition. In particular preterm infants, are characterized by an immature neurobehavioral profile at birth, even in absence of severe brain injuries and associated perinatal complications. Therefore, “by definition” they require long-lasting hospitalization in the Neonatal Intensive Care Unit (NICU). This increases the newborns’ survival, but at the same time entails a number of stressors, such as physical, pain related, and socio-emotional, thus representing an early adverse experience, linked to detrimental consequences for neurological, neuro-endocrinal, sensorial, behavioural, emotional, and socio-emotional development, as well as to increased chronic disease risk, later in life [60,66]. Indeed, elevated oxidative stress and inflammation, both of which contribute to telomere shortening, has been recorded in premature hospitalized preterm infants, while a measurable decline in TL during hospitalization, may convey information about NICU exposures that carry both immediate and long-term health risks [67,68,69,70]. It is therefore not surprising that researchers assume preterm infants display an “aged” phenotype [61], further increasing the importance of TL regulation and monitoring. Nevertheless, whether infant and placental TL are biomarkers of PTB, or the actual cause of this adverse outcome, remains to be discovered.
The aim of this literature review is to investigate the possible associations between maternal nutrition, placental - newborn TL and PTB.
2. Materials and Methods
Description of research strategy
This is a review study of the international scientific literature. Articles published in scientific journals were sought, from the online databases PubMed and EBSCO host, from 2013 to 2023 in an effort to reflect current knowledge and well established research trends. The search took place from March to September 2023 and the terms used, were:
Maternal (nutrition OR diet), Pregnancy (nutrition OR diet), Fetal (nutrition OR nutrients)
AND
Placenta (telomeres OR telomere length) OR (Newborn OR Infant OR Neonatal OR Offspring)
AND
(Preterm OR Premature) (birth OR labor) (infant OR newborn OR offspring)
AND
(Observational studies OR Cohort studies OR Case control studies OR Cross sectional studies).
The searches, were made independently by two reviewers, namely NL and IT. Initially, the abstracts were read. In this way, research selection was directly related to the subject. Citation chaining also took place, in order to avoid omitting relevant studies. With the completion of the article searches, the studies were classified into thematic categories, based on the exposure and the outcome (PTB, maternal nutrition, placental and neonatal TL).
Study inclusion criteria:
- Studies published from 2013 to 2023.
- Observational studies (i.e. cohort, case-control, cross-sectional studies).
- Direct relation to the subject and inclusion of the specified keywords.
- Conceptual connection of keywords with the title and summary.
- Inclusion of studies with positive, negative or neutral effect on the association of maternal nutrition with placental / newborn TL and PTB.
- Description of the search strategy and eligibility of studies according to the four-phase flow chart of PRISMA guidelines [71].
3. Results
The search returned several studies for each keyword separately. However, when all keywords were combined together, no study was found associating maternal nutrition with placenta and/or newborn TL and PTB.
For this reason, the keywords A: Maternal nutrition, B: PTB, and C: Placental / newborn TL, were searched in dyads as follows: concept “A” relation to concept “B”, “C” to “A”, and “C” to “B”. It is however impossible, to find chronological sequence of the exposure and the outcome, but evidence of associations between these factors, would imply a potential relationship (either causal or correlational) [72].
3.1. Studies eligibility
As previously mentioned, from the thorough preliminary search of relevant literature in PubMed and EBSCO host electronic databases, no studies from the combination of keywords, were found. The process of selecting and excluding the studies was carried out according to the PRISMA guidelines, as shown in Figure 1 [71].
Thus, the literature search was performed linking concepts A to B, B to C and to A to C. In total 40 observational studies containing the key words in the combinations described above, were included.
The selected studies were evaluated for their methodological quality, on the basis of the checklist of the 22 thematic modules, examined by the STROBE scale, and were all considered to be of high quality.
3.2. Maternal nutrition and PTB
Of the above 40 included studies, 21 were found to associate maternal nutrition with premature birth. Publication years were from 2014 to 2023. Five of the studies took place in China, 1 in Indonesia, 1 in Norway, 2 in Singapore, 3 in the United States of America, 1 in Canada, 1 in Malawi, 1 in Pakistan, 1 in Mexico, 1 in Brazil, 1 in Switzerland, 1 in Portugal, 1 in Bangladesh and 1 in Italy.
Regarding the methodology, 12 of the studies concerned cohort studies, 4 studies were cross-sectional and 5 studies case-control. Overall, the studies’ population, were pregnant women, with a singleton pregnancy and the absence of any pathology or pregnancy complications.
Dietary intake was assessed with validated questionnaires, and/or measurements of anthropometric characteristics of the pregnant woman (BMI, arm circumference etc.), as well as the newborn (sex, weight, length) were recorded. Also, delivery characteristics such as gestational age (GA) at delivery, were documented.
The results showed both positive and negative associations, between maternal nutrition (either as a dietary pattern adherence or individual food items, or supplement/s intake) and PTB.
Adherence to a healthy diet, as recorded by a maternal nutritional score and GA at birth, were positively correlated. In particular, a significant increase in the risk of PTB, was associated with low nutritional scores from a questionnaire developed and based on the International Federation of Gynecology and Obstetrics (FIGO) recommendations on adolescent, preconception, and maternal nutrition. Furthermore, a significant increase in preterm deliveries, with a relative risk of 1.44, was recorded for women with a 1st trimester nutritional score lower than five. However, the single food items of the score calculation, were not associated with both early placental markers and complex pregnancy outcomes [73].
The Norwegian Fit for Delivery diet score either pre-pregnancy or in early pregnancy, was protectively associated with excessive GWG and risk of PTB. The protective association with high birthweight was confined to pre-pregnancy diet and with preeclampsia to early pregnancy diet. The study recorded no association between pre-pregnancy diet score and pre-eclampsia [74].
Dietary patterns related to reduced food intake or fasting, in the 2nd trimester of pregnancy (Ramadan period of the Muslim religion), were positively associated with increased risk of very PTB between 28 and 31 weeks of gestation. At 15-21 weeks the risk increased by 1.33 times and at 22-27 weeks by 1.53 times [75].
Yogurt consumption during pregnancy appeared to have a negative (protective) association with PTB, especially when combined with a normal BMI [76]. However, in the study by Lu et al. [77] higher dairy consumption compared with vegetable consumption, showed a strong correlation with PTB.
In very PTB, dietary maternal consumption during pregnancy of Portuguese women giving birth very prematurely, was assessed. Consumption of certain foods did not comply with the recommendations for pregnant women by the Portuguese Directorate General of Health. In particular, consumption was below the recommended levels for dairy products (1 vs 3 portions), vegetables (2 vs 3 portions) and fruits (1 vs 4 portions). The study also recorded a very low cereal intake (average of one portion per day ingested). Furthermore, PTB associated pregnancy-induced hypertension was associated with increased consumption of pastry products, fast food, bread, pasta, rice, and potatoes. However, only bread consumption had a weak but statistically significant association with pregnancy-induced hypertension in a multivariate analysis model [78].
A fruit/vegetable/rice-based dietary pattern during pregnancy appeared protective against PTB and small for gestational age (SGA) infants, compared with two other dietary patterns; namely, one based on consumption of seafood and noodles and the second on consumption of processed meat, cheese, pasta. Only the fruit/vegetable/rice pattern contributed positively to the development of long for gestational age (LGA) neonate [79].
In the prospective cohort of Martin et al. [80], the diet quality during pregnancy was associated with PTB. Specifically, greater adherence to a healthy dietary pattern, such as the DASH diet, reduced the odds of PTB, especially during the 2nd trimester. On the contrary a greater adherence to a dietary pattern of poorer quality, such as pre-procced or fast foods, high fat foods and confectionary, increased the odds of PTB.
The use of folic acid supplements of 400μg or more, showed a negative correlation with PTB. Folic acid use, appeared to reduce PTB by 14%. Strong association with spontaneous PTB and non-significant for iatrogenic PTB or prolonged spontaneous rupture of membranes was also recorded [81]. Furthermore, the large cohort study of Wu et al. [82] showed similar results with folic acid periconceptional supplementation associated with a lower risk of PTB. In particular, the earlier women started taking their folic acid supplements prior to pregnancy (i.e. at least 3 months before their last menstrual period), the more likely to reduce the risk of PTB, compared with women who started taking folic acid later (i.e. 1–2 months before their last menstrual period) .
Multivitamin use in the 3rd trimester of pregnancy, had a negative correlation with preterm and very PTB, especially in pregnant women of African non-Hispanic descent. However, the dose response effect was not investigated [83]. Similar results by Olapeju et al. [84] support that multivitamin supplement intake of at least 3 times/week throughout pregnancy, is significantly associated with a reduction in the odds of PTB. Use during the third trimester, was especially associated with a greater reduction in PTB odds than use in the first trimester, but no significant association between preconceptional supplement intake and PTB. Also, higher plasma folate levels were associated with lower risk of PTB.
Ren et al. [85] in a case-control study, they examined hair levels of trace elements, including endocrine disrupting metal(loid)s (EDMs) such as lead, mercury (Hg), arsenic, and cadmium, and nutritional trace metal(loid)s (NTMs) such as zinc (Zn), iron (Fe), copper, and selenium, in 415 control pregnant women with birth at term, and 82 pregnant women with PTB. Negative (protective) correlation between increased levels of NTMs, especially Fe and Zn and PTB occurrence, was recorded. Also, a potential protective effect of Hg was seen against PTB, while for EDMs, only maternal hair Mercury (Hg), was negatively associated with PTB risk.
PTB was associated with higher serum concentration of heavy metals (such as mercury and lead, lower maternal serum concentration of AtRA) and all micronutrients, and with lower placental concentration of manganese, iron, copper, zinc, selenium, AtRA, 25(OH)D, and higher placental concentration of mercury and lead. Compared with PTB group, term birth group had higher concentration of copper and AtRA in cord blood [86]. Copper in maternal serum concentrations has been recorded above the upper normal limit in both term and PTB groups of a nested case control study in Malawi. At the same time, PTB was associated with higher maternal serum concentrations of copper and zinc [87].
In the cohort study of Perveen and Soomro [88] iron deficiency anemia (Hb<11mg/dl) appeared positively associated with PTB, low birth weight, fetal mortality and low Apgar score at the 1st and 5th minutes of birth. Additionally, increased levels of folic acid, at 3rd trimester were associated with reduced risk of PTB and longer duration of gestation of PTB. Little or no correlation was found between increased levels of B6 & B12 and PTB or SGA [89].
In the cross sectional study of Christoph et al. [90] the lower the maternal serum vitamin D level, the higher the GA at birth, but no association was observed between vit D levels and PTB. In addition, the nested case – control study in Bangladesh, that vitamin D deficiency, common in Bangladeshi pregnant women, is associated with an increased risk of PTB [91].
Inverse relationship between maternal total protein levels (via the diet) and PTB during the 3rd trimester of pregnancy, especially in female fetuses was recorded in a large Chinese cohort study [92]. The odds of adverse outcomes were higher in non-white obese women with high protein consumption, in the nested case control study by Miele et al. [93] The anthropometric classification of obesity had a greater impact on PE and GDM, in contrast to PTB and SGA. In total, obesity had a small effect on PTB.
3.3. Maternal nutrition, placental and newborn telomeres
In total, 13 studies were retrieved and included in this review investigating the relationship between maternal nutrition, placental and newborn telomeres.
The studies publication years were from 2015 to 2022. Studies were conducted in various geographic locations, namely China (n=1), Argentina (n=2), Italy (n=1), USA (n=4), Belgium (n=1), Seoul (n=1), Seychelles (n=1), Rwanda (n=1), and Singapore (n=1).
In terms of methodology, 12 were cohort studies and 1 was cross sectional. The study populations primarily consisted of pregnant women, while others mother-newborn dyads. The basic characteristics of the majority of the studies populations, concerned pregnant women, in the 1st trimester of pregnancy, with singleton pregnancy and the absence of pathology.
Blood samples of pregnant women, questionnaires monitoring their eating habits, supplement intake and/or measurements of anthropometric characteristics of the pregnant woman (BMI, arm circumference), of the newborn (gender, weight, length) and delivery characteristics (GA at delivery), were recorded. In addition, observations were made in regards to the characteristics of the placentas, measurements of their TLs as well as the leucocyte TL (LTL) of the newborns from samples of umbilical cord blood at delivery.
The results showed both positive and negative associations between maternal diet and placental and neonatal TL.
In terms of dietary lipids, one study examined the effect of omega-3 intake and their effect on umbilical cord, placenta and infant TL. Keeping higher levels of maternal n-3 polyunsaturated fatty acids (PUFAs) during pregnancy, may help maintain TL in the offspring, which is beneficial to long-term offspring health [94].
No clear associations were recorded, for prenatal or postnatal PUFAs status and methylmercury exposure with offspring TL. Higher prenatal n–6:n–3 PUFA ratio was however, associated with longer maternal TL [95]. On the contrary, a maternal high-fat dietary consumption pattern during pregnancy, was associated with shortened TL among fetuses, after accounting for the effects of potential covariates [96].
One study, correlated placental and umbilical cord blood TL with maternal nutritional profile. In specific, a positive association between plasma vitamin 25(OH)D3 and placental TL was observed, while an inverse association was observed between BMI, body fat percentage, vitamin B12 and placental TL. Furthermore, a negative correlation was found between the above and the length of umbilical cord blood telomeres [97].
Furthermore, vitamin D levels during pregnancy and especially in the 1st trimester, were positively correlated with neonatal leukocyte TL at delivery [47,98]. Positive correlation was also seen between neonatal leukocyte TL and maternal leukocyte TL, maternal vitamin D concentration, maternal energy intake and newborn weight [47].
A positive correlation was also seen between folic acid and neonatal TL. Each 10ng/dl increase in pregnant women's folic acid intake, was associated with a 5.8% increase in mean TL. The average TL of newborns of mothers with low folate serum concentration, was shorter compared with those whose mothers belonged to the group with increased levels [46].
Furthermore, maternal folic acid supplementation after the first trimester and throughout pregnancy was associated with longer newborn TL [99]. However, in the same cohort, no significant association was found between maternal folic acid supplementation in the first trimester and newborn TL. One more study of the effect of folate on newborn TL, showed a positive association between umbilical cord RBC folate and fetal TL at birth [100].
TL, but only in female newborns and not male, was more susceptible to variation from maternal vitamin B12 levels, as well as maternal TL, and mental health. In total, maternal TL was strongly associated with antenatal factors, especially metabolic health and nutrient status [65].
The increased exposure of pregnant women to toxic metals (antimony, lithium, arsenic) was positively correlated to placental TL and newborn sex. Antioxidants (zinc, selenium, folate, Vit D3), did not contribute to the modification of the above process. The TL of the placenta, decreases as the age of the pregnant woman increases, approximately by 1% per year. Lithium appears to increase the mother's TL. Lead (in umbilical cord blood) showed an inversely proportional correlation with infant TL, particularly in male neonates [101].
Magnesium deficiency was negatively associated with maternal RTL after adjusting for covariates. A positive association between maternal intake of magnesium and TL of cfDNA from amniotic fluid, while results on other micronutrients (i.e., vitamin B1 and iron) were marginally significant [102]. Also, Myers et al. [103] examined the relationship of vitamin C intake and cord blood TL. The sample similarly to the Louis-Jacques, et al. [100] study was drawn as a part of an ongoing prospective cohort study conducted by the University of South Florida (USF) Morsani College of Medicine. A positive association between maternal vitamin C intake and fetal TL.
3.4. PTB, placental and newborn telomeres
As for the association of PTB with both the placental and the newborn telomeres, 6 studies published from 2015-2023, were found. The study countries were Brazil (n=1), United Kingdom (n=1), Indonesia (n=1), Israel (n=1), India (n=1) and France (n=1). In terms of methodology, 2 were cohort, 1 case-control and 3 were cross sectional studies.
The population of the studies was heterogenous and included pregnant women in normal labor, mother-infant dyads, or 2- and 7-9-year-old children, who were born either full term or prematurely. Data analysis included: anthropometric measurements, dietary intake questionnaires, observations regarding pregnancy course, DNA methylation of umbilical cord blood and placental tissue, umbilical cord blood and placental tissue LTL.
A study indicating no effect of PTB on TL, showed similar placenta TL in PTB and term labor placentas. In this cross sectional study, early telomere shortening in PTB, was observed, that mimics the term placenta. Markers 8-OHdG and HMGB1, did not correlate with placental telomere ratio, while HMGB1 from the placenta of both PTB and term labor showed no significant difference. Equal relative amount of telomeric DNA (T) to the beta-globin single copy gene (S), calibrated to a plate reference genomic DNA sample (T/S ratio) of the placenta from PTB and term labor was recorded [104].
In contrast, telomere shortening in foetal membranes, was suggestive of senescence associated with triggering of labor at term. As such, foetal membranes from the term labor group showed TL reduction compared with those from the others. However, telomerase activity did not change in foetal membranes, irrespective of pregnancy outcome [105].
In the case-control study by Vasu et al. [61] preterm infants showed longer TL than full-term infants. A positive correlation was observed between maternal age and the telomere/shelterin (protein) ratio, while as the age of the mother increased, the longer were the telomeres of the newborn (p=0.011). Accordingly, maternal blood and placental samples from spontaneous PTB, presented shorter telomeres and increased Gal-3 expression, compared with spontaneous term pregnancies group [106].
Another longitudinal cohort study, tracked premature infants into adulthood by studying TL in saliva, as well as lung function. A positive correlation was recorded between TL and abnormal lung airflow in the adult population who were born prematurely. There was no apparent association with perinatal causes of PTB. Nevertheless, no apparent association with perinatal events and TL was noted [70].
Increased LTL attrition was observed in those born before 37 weeks of GA, as well as, in those who gained weight as adults in a cohort study in India. GA was positively associated with offspring RTL, although no significant association of offspring RTL with body size at birth including birthweight, birth length, and birth BMI. Conditional BMI gain at 2 and 11 years not associated with RTL. BMI gain at 29 year was negatively associated with RTL. Born SGA was not associated with RTL in adulthood [107].
3.5. Figures & Tables
Figure 2.
Schematic representation of aggregated key findings. PTB: preterm birth, TL: telomere length, VFR: vegetables/ fruits/rice, Zn: zinc, Fe: iron, Hg: Mercury, AtRA: All-trans retinoic acid, Mn: manganese, Cu: copper, Se: selenium, Pb: lead, Li: Lithium, PUFAs: Polyunsaturated fatty acids, Mg: magnesium, GA: gestational age.
Figure 2.
Schematic representation of aggregated key findings. PTB: preterm birth, TL: telomere length, VFR: vegetables/ fruits/rice, Zn: zinc, Fe: iron, Hg: Mercury, AtRA: All-trans retinoic acid, Mn: manganese, Cu: copper, Se: selenium, Pb: lead, Li: Lithium, PUFAs: Polyunsaturated fatty acids, Mg: magnesium, GA: gestational age.

4. Discussion
In the current study, the systematic review traditional methodology was rejected since the combination of the search keywords “Maternal nutrition”, “Preterm birth” and “Placental and/or newborn telomeres”, returned zero results. For this reason, the keywords A: Maternal nutrition, B: PTB, and C: Placental and/or newborn TL, were searched in dyads as follows: is concept “A” related to concept “B”, “C” to “A”, and “C” to “B”; thus leading to the assumptive inference of A being related to B and C. It is however, impossible to find chronological sequence of the exposure and the outcome, but evidence of associations between these factors, would imply either a causal or a correlational relationship [72].
The data from the relevant literature search, led to the final selection, of 40 observational studies. The overall results indicate a relationship between maternal nutrition and PTB, as well as, maternal nutrition and newborn and placenta TL. However, inconclusive were the evidence for the relationship between PTB and TL.
Maternal nutrition and PTB
In terms of maternal nutrition and PTB, all 21 studies included in this review, indicated that there are nutrients (either through dietary intake or supplementation) and eating patterns, associated with increased risk of PTB, while others play a beneficial and/or protective role against PTB.
Nutritional status, as assessed by selected biomarkers, is associated with higher risk of PTB and GA at birth. In particular, protein level in maternal blood plasma, is inversely associated with risk of PTB and positively associated with gestational duration, in the 3rd trimester of pregnancy, particularly in the female fetus [92]. Dietary protein is a macronutrient well known for its relationship with foetal health in the past decades [108,109,110,111,112,113]. However, few studies have investigated the role of protein on PTB risk, and the results are inconclusive especially when examining protein supplementation and PTB incidence [114]. By using selected valid and reproducible biomarkers of maternal protein plasma levels, additionally to dietary intake assessments - which present numerous challenges to obtaining accurate dietary intake and nutritional status - significant confounders are omitted (i.e. quantification of intake, self-report bias, pathological causes of reduced absorption etc.) [92].
Another nutrient biomarker examined for its role in PTB, is folate. Higher maternal serum folate concentration at approximately the start of the 3rd trimester, was associated with longer duration of gestation and lower risk of PTB, while little or no correlation was seen between serum B6 & B12 and PTB [89]. Supportive of the above findings regarding folate, one more study, showed that higher plasma levels of the nutrient, are associated with lower risk of PTB [84]. The relationship between folate levels and PTB, is further supported in a recent meta-analysis of folic acid and risk of PTB. The meta-analysis evidence indicates that early pre-conceptually high maternal folate levels, are significantly associated with a lower risk of PTB. Moreover, protective action against spontaneous PTB was seen, with the daily use of 400μg folate periconceptionally [115]. However, folate supplementation does not appear to protect against iatrogenic PTB or prolonged spontaneous rupture of fetal membranes [81]. The large cohort study of Wu et al. [82] showed similar results with folic acid periconceptional supplementation being associated with lower risk of PTB. Specifically, the earlier women started taking their folic acid supplements prior to pregnancy (i.e. at least 3 months before their last menstrual period), the more likely to reduce the risk of PTB, compared with women who started taking folate later (i.e. 1–2 months before their last menstrual period) [82]. An earlier systematic review suggested that supplementation, is associated with a significant reduction in the risk of PTB, but only when being initiated after conception [23]. Although further RCTs are warranted, to establish the relationship between folate and PTB, the beneficial effect of folate on the reduction of birth defects, ascertain it’s necessity in adequate amounts during the peri-conceptional period.
Vitamin D is another micronutrient that was studied for its effects on PTB risk, in three studies included in this review. Firstly, the cross sectional study of Christoph et al. [90] showed that the lower the vitamin D maternal level, the higher the GA at birth. The 2nd cross sectional study in Indonesia, comparing the PTB group with the term birth group, found that the latter had higher placental concentration of 25(OH)D [86]. In addition, in the 3rd nested case – control study in Bangladesh, vitamin D deficiency in pregnant women, was associated with an increased risk of PTB [91]. An earlier meta-analysis, had already shown evidence of the association between maternal circulating 25-OHD deficiency (rather than insufficiency), and an increased risk of PTB. To enhance the argument, the meta-analysis concludes that vitamin D supplementation alone during pregnancy, may reduce PTB risk [116]. However, RCTs to date, have unanimously failed to ascertain the prophylactic effect of vitamin D against PTB, with the emergence of conflicting evidence [117]. It is suggested that low levels of vitamin D, may reflect poor general maternal health status. Thus, priority should be given to the attainment of general health, rather than vitamin D supplementation per se [118].
Maternal iron deficiency anaemia was positively associated with PTB [88]. In particular, hemoglobin (Hb) levels of less than 11mg/dl were directly associated with PTB, low birth weight, fetal mortality and a low Apgar score in the 1st and 5th minute of birth [88]. In a recent systematic review and meta-analysis, iron deficiency anaemia was indeed found to be a contributing factor towards PTB during the first trimester, but, not in the second and third trimester [119]. When accounting for iron during pregnancy, it is imperative to consider its critical role for embryonic development and fetal growth when transported through the placenta from the mother to the fetus. Furthermore, since iron cannot be synthesized by the body, sufficient iron absorption from dietary sources is very important for both mother and fetus [120]. Still, little is known about the iron states in the mother, the placenta, and the fetus, and what mechanisms responsible for iron transport, contribute to PTBs. A recent study attempting to characterize maternal and fetal iron metabolism in pregnant women with PTB, found a dysregulated iron homeostasis in both sides and a disordered placental iron equilibrium, which were presumed to account for the compromised fetal iron supply [121]. Prevention or treatment with either the intravenous iron supplementation or oral medication, showed no significant differences in maternal and neonatal outcomes, thus directing the interest towards nutritional correction of iron deficient states during pregnancy [122].
The effect of the availability of other micronutrients on PTB risk, is more complex than the previously discussed folate, vit. D and iron. Lower concentrations of all-trans retinoic acid (AtRA), in maternal serum, were inversely associated with PTB, while PTB was also positively associated with higher serum concentrations of heavy metals such as mercury and lead, in the Irwinda et al. [86] cross sectional study. Also, compared with the PTB group, term birth group had higher maternal serum concentration of higher placental concentration of manganese, iron, copper, zinc, selenium, AtRA, lower placental concentration of mercury and lead, and in cord blood a higher concentration of copper and AtRA [86]. A current scoping review, also documented a higher incidence of PTB with lead and cadmium exposures. The findings however for mercury and arsenic exposures, were inconclusive. The most common pathways through which heavy metals and metalloids lead to increased risk of PTB are, placental oxidative stress, epigenetic modifications, inflammation, and endocrine disruptions [123].
In hair samples, nutritional trace metal(loid)s concentrations especially Fe and Zn are negatively associated with PTB and surprisingly the endocrine disrupting metal(loid) Hg, was also negatively associated with PTB [85]. Conversely, PTB was associated with higher maternal serum concentrations of copper and zinc in a nested case control study in Malawi, indicating perhaps the difference between type of sampling source of the biomarker (e.g. blood serum and plasma, cellular fractions, tissue etc.) [87]. Due to the conflicting results, and the low-to-moderate-certainty evidence, it is suggested that zinc supplementation may reduce PTB risk in women with low zinc intake and level or poor nutrition [114].
Any type of multivitamin supplementation during the 3rd trimester of pregnancy (but not earlier) was associated with a significant reduction in PTB among non-Hispanic and African women in a large USA cohort study [83]. In agreement is the case control study by Olapeju et al. [84] where multivitamin supplement intake of at least 3 times/week throughout pregnancy, was significantly associated with a reduction in the odds of PTB, while the use during the third trimester was also associated with a greater reduction in PTB odds, than use in the first trimester. In both studies described above no significant association was recorded between preconceptional multivitamin intake and risk of PTB. On the contrary, in a systematic review and meta-analysis, the use of multivitamins and adverse birth outcomes in high income countries, did not change the risk of PTB [124]. However, the authors stressed the need for additional data on multivitamin intake in pregnancy from randomized controlled trials or large cohort studies controlling for multiple confounders.
In terms of dietary patterns, religious fasting (such as the Ramadan period of the Muslim religion) in the 2nd trimester of pregnancy, increased the risk of very PTB between the 28th and 31st week of gestation, while at 15-21 weeks the risk seemed to increase by 1.33 times and at 27 weeks by 1.53 times [75]. This cohort had a large population of 78,109 births making the evidence robust. On the contrary, in a 2012 smaller cohort study, the practice of Ramadan fasting did not increase baseline risk of PTB in pregnant women, regardless of the GA during which this practice is adhered. The recent review and meta-analysis that examined the effect of Ramadan fasting during pregnancy on perinatal outcomes of 5600 births, showed no effect on PTB [125]. Studies indeed have shown conflicting results on the effect of fasting on PTB, while the timing of exposure to maternal fasting may affect birth outcome including birth weight [125]. More well-designed studies with large samples, investigating all types of fasting during all trimesters of pregnancy, are required to shed light on its impact on maternal and fetal health [126].
A dietary pattern during pregnancy based on fruits, vegetables and rice, protected women against PTB and SGA infants, compared with dietary patterns based on seafood and noodles or based on processed meat, cheese, pasta. Nevertheless, the fruit/vegetables/rice pattern contributed towards the development of LGA infant [79]. Supportive of this evidence, is the study by Teixeira et al. [78] in which the dietary habits of women giving birth prematurely were assessed. In the group of women giving birth prematurely, the consumption of certain foods (i.e. dairy products, cereals, vegetables and fruits) did not comply with the recommendations for pregnant women by the Portuguese Directorate General of Health, providing evidence for their important role in PTB risk. Furthermore, in the very PTB group, pregnancy-induced hypertension was associated with increased consumption of pastry products, fast food, bread, pasta, rice, and potatoes, but only bread consumption had a weak but statistically significant association, with pregnancy-induced hypertension.
Furthermore, dairy products seem to have a controversial effect. It is argued that an increased consumption of dairy products has a protective effect against PTB when combined with a maternal normal BMI [76]. Otherwise, the increased consumption of dairy products, especially milk, as well as Cereals, eggs, and Cantonese soups’ and ‘Fruits, nuts, and Cantonese desserts’ groups, compared with vegetable consumption, had a strong correlation with PTB [77]. When examining the effect of dietary patterns on PTB severity levels, a pattern rich in rice and nuts lowered odds of very/moderately PTB compared with late PTB or term birth, while a high dietary consumption of starch foods dietary was associated with the most severe level of PTB outcome incidence [127].
On the same note, in the prospective cohort of Martin et al. [80] maternal diet quality was associated with PTB, where a greater adherence to a healthy dietary pattern, such as the DASH diet, reduced the odds of PTB, especially during the 2nd trimester. On the contrary a greater adherence to a dietary pattern of poorer quality, such as pre-procced or fast foods, high saturated fat foods and confectionary, increased the odds of PTB.
Comparing five different food patterns, namely “Obesogenic” “Traditional” “Intermediate” “Vegetarian” and “Protein”, Miele et al. [93] performed a nested case control study in Brazil, in order to investigate their association with unfavourable pregnancy outcomes and PTB. They report a diet rich in protein increases the probability of developing preeclampsia and PTB. However, the anthropometric classification of obesity had a greater impact on preeclampsia (PE) and gestational diabetes mellitus (GDM), in contrast to PTB and SGA outcomes, suggesting that the effect of the dietary pattern on PTB is dependent upon other anthropometric characteristics. In a recent review, which included 40 observational studies, the dietary patterns during pregnancy associated with a lower risk of PTB, were also characterised by high consumption of vegetables, fruits, as well as whole grains, fish and dairy products [11].
In general, the evidence indicates that a Western-type diet, high in meat and fats and low in fruits and vegetables, is associated with increased odds of induced PTB [128]. On the contrary adherence to a Mediterranean and /or healthy diet pattern during pregnancy, appears associated with reduced risk of PTB [129,130]. It may also be that the protective effect increasing the intake of foods associated with a Mediterranean or healthy dietary pattern is more important than totally excluding highly processed food, fast food, confectionary, and snacks [131].
Similar results were noted with nutritional scoring systems. In the cohort of Parisi et al. [73] a positive association between healthy maternal nutritional score and GA at birth was reported, while a significant increase in the risk of PTB was associated with low nutritional scores. However, the single food items score calculations, were not associated with both early placental markers and complex pregnancy outcomes. On the same note, a high Norwegian Fit for Delivery diet score, either pre-pregnancy or in early pregnancy, was inversely associated with gestational weight gain (GWG) and PTB risk [74].
Examining dietary patterns as a whole, independent of the way of assessment, has emerged as a holistic approach for capturing the complex interactions between nutrients and foods [132]. In total, the evidence shows that maternal nutrition during pregnancy, as assessed through dietary patterns, is a major determinant for birth outcomes and, consequently, offspring health outcomes in later life [133].
Surprisingly, no recent studies were found that examine exclusively maternal BMI and the risk of PTB. Previous evidence showed that either a high or low BMI is associated with increased risk of PTB. However, when limited to developing countries low BMI was not significantly associated with PTB [118]. BMI is an obscure measure of nutritional status and health, especially since it does not take into account body composition (e.g. muscle mass, bone density etc.) and racial and sex differences [134], and thus is not considered as a valid factor accounting for PTB risk.
As a whole, the above evidence indicate that dietary patterns linked to enhanced anti-inflammatory and anti-oxidant properties, containing a variety nutrient dense, non-processed foods that reduce the risk of developing nutritional deficiencies, are the most appropriate for the ascertaining a healthy period of pregnancy and fetal development as well as the avoidance of adverse birth outcomes.
Maternal nutrition, placental and newborn telomeres
In total 13 studies were included in this review, exploring the association between maternal nutrition and placental and infant TL. The findings conclude that diet has an important role in whether or not the placental TL will be maintained. Specifically, the level of maternal plasma vitamin D, seems to be positively correlated with placental TL, thus contributing towards TL maintenance or reduction of TL attrition. Conversely, an inverse effect was observed for BMI, body fat percentage, vitamin B12 and placental TL, but no associations were apparent for cord’s blood TL [97].
Further studies support the protective role of maternal plasma vitamin D in maintaining the newborn’s TL. In two studies included in this review, the concentration of serum vitamin D during pregnancy, was positively correlated with the neonate’s LTL at birth [47,98]. Furthermore, a positive correlation was observed between the newborn’s LTL with maternal LTL, vitamin D levels in maternal plasma, maternal energy intake and the newborn’s weight [47]. The pleiotropic effects of vitamin D in the organism and especially the immunomodulatory effects of vitamin D, may be one mechanism by which it is protective against telomere attrition [135]. More recently, these evidences were strengthened by a cohort in Hong Kong where 25(OH)D, D3 and D3 epimer both in utero and at birth, impacted childhood LTLs [136]. Furthermore, insufficient maternal vitamin D (25(OH)D) has been associated with increased offspring risk for many diseases and later life adverse outcomes [137].
On the contrary, Herlin et al. found no association between antioxidant maternal serum levels of zinc, selenium, folate, and Vit D3 and maternal or newborn TL [101]. Increased exposure to toxic metals (antimony, lithium, arsenic) during pregnancy was positively associated with a decrease in placental TL. Lithium had a positive effect on maternal TL, while lead (in cord blood) showed an inverse association with TL, particularly in male infants. In addition, placental TL decreased as the pregnant woman’s age increased about 1 % per year (p = 0.0009).
The contradicting results, of maternal vitamin D levels and their effect on offspring TL, may be due to the differences in the timing of vitamin D level assessments during pregnancy. For example, Daneels et al. showed that maternal nutrition during early pregnancy and in particular the first trimester, was associated with TL at birth [98]. In support of the above, another study in Switzerland showed that the effects of vitamin D are more pronounced during the earlier gestational period [138].
Three studies included in this review, investigated the effect of folate on fetal and placental TL [46,99,100]. A positive association was observed between maternal folic acid levels early in pregnancy and newborn cord blood TL. Indeed, each increase of 10 ng/dl was associated with a 5.8% increase in median newborn TL, in the cohort study of Entringer et al. [46]. The average neonatal TL in the group of mothers with reduced concentration of folate, was shorter compared with those whose mothers belonged to the high folate concentration. The authors supported that fetal telomeres exhibit developmental plasticity and show that maternal nutrition can affect or even "program" this system.
Louis-Jacques, et al. study [100], also showed a positive association between umbilical cord red blood cell folate levels and fetal TL at birth, while a possible association between maternal folic acid supplementation during pregnancy and longer newborn TL was suggested in the cohort study by Fan et al. [99]. Folate’s role in DNA methylation and oxidative stress are proposed mechanisms, through which it influences TL regulation in the offspring [139]. On the contrary in the study by Kim et al. dietary intake of dietary folate equivalents, assessed by 24-hour recalls, was not associated with fetal TL [47]. A recent systematic review of the effect of maternal diet and offspring TL, also concluded that higher circulating maternal folate and 25-hydroxyvitamin D3 concentrations were associated with longer offspring TL adding to the equation a protective effect for higher maternal dietary caffeine intakes [140]. As of yet, contradictory remain the data regarding higher dietary intake of carbohydrate, folate, n-3 polyunsaturated fatty acids, vitamin C, or sodium in regards to offspring TL regulation [103].
Another study examining maternal micronutrient status and TL in maternal serum, cord blood, amniotic fluid and placenta, showed that only magnesium deficiency negatively associated with maternal RTL. Furthermore, a positive association between maternal intake of magnesium and TL of cfDNA from amniotic fluid was seen, while results on other micronutrients (i.e., vitamin B1 and iron) were marginally significant [141]. Moreover, Nsereko et al. concluded that lower ferritin, soluble transferrin receptor levels, and retinol-binding protein levels are associated with longer maternal TL [142].
The effects of dietary fat and its biomarkers, has also been investigated. Shortened TL among fetuses exposed to maternal high fat consumption during pregnancy, after accounting for the effects of potential covariates, was recorded in the Salihu, et al. [96], but with no association recorded for a high carbohydrate diet. However, according to Liu et al. study concentrations of total n3- PUFAs and docosahexaenoic acid (DHA) in maternal erythrocytes, are closely correlated to infant TL and the telomerase reverse transcriptase (TERT) promoter methylation [94]. Maternal n-3 PUFAs (a-linolenic, eicosapentaenoic and docosahexaenoic acids) in cord blood and the placenta correlated to infant TL and TERT promoter methylation, differently in cord blood and the placenta. Maintaining higher levels of maternal n-3 PUFAs during pregnancy may help to conserve the offspring TL, accompanied by potential benefits to long-term health effects.
Contradictory evidence for the maternal PUFA status and its relation to offspring TL was recorded in the cohort study by Yeates et al. [95]. There were no clear associations for both prenatal or postnatal PUFA status and methylmercury exposure with offspring TL, recorded. However, the higher prenatal n–6:n–3 PUFA ratio was associated with longer TL in mothers.
In terms of maternal dietary patterns and offspring TL regulation later in life, the - recent first of its kind- systematic review examining the impact of dietary intake factors on TL in childhood and adolescence, suggests that a higher consumption of fish, nuts and seeds, fruits and vegetables, green leafy and cruciferous vegetables, olives, legumes, polyunsaturated fatty acids, and an antioxidant-rich diet might positively affect TL. Conversely, a high intake of dairy products, simple sugar, sugar-sweetened beverages, cereals, especially white bread, and a diet high in glycemic load were associated with enhanced TL shortening in the offspring during childhood and adulthood [143].
Offspring sex, also appears to affect the associations between maternal nutritional intake and status and TL. Indeed, TL in female newborn was shown to be more susceptible to variation from maternal TL and vitamin B12 levels, while variation in newborn male TL was more explained by parental age, maternal education, plasma fasting glucose, DGLA%, and IGFBP3 levels [65]. Thus, demographic factors such as offspring sex and maternal ethnicity seem to also affect the relationship between maternal nutrition and offspring TL, and thus should be treated as confounding factors in all relevant investigations [103].
It is relatively well established in both animal and human studies that nutrition plays a profound role on DNA integrity, epigenetic mechanisms, and TL regulation [32,144,145,146]. Indeed, various nutrients influence TL through mechanisms depicting their role in cellular functions including inflammation, oxidative stress, DNA integrity and methylation, as well as telomerase activity [144]. However, the evidence from human studies, examining the impact of maternal nutrition on newborn TL remains scarce, not allowing the reach of definite conclusions. Since TL at birth represents an individual’s initial setting of TL and predicts later life TL [147], the effects of maternal diet on offspring TL, should be thoroughly investigated. The need for longitudinal studies, assessing nutritional intake and/or status - with various indices including nutrient biomarkers - and exploring possible associations with offspring and placenta TL regulation, is warranted. Upon ascertaining the potential nutritional determinants of offspring TL regulation, TL status and attrition rates could become valuable as markers of future chronic disease risk.
PTB, placental and newborn telomeres
As for the association of PTB with both the placental and the newborn TL, only 6 studies were found. TL appears to be highly variable in newborn infants. In particular, PTB infants, were found to have longer TL than full-term, while TL was significantly negatively correlated with GA and birth weight. A positive correlation was also observed between maternal age and the telomere/ beta-globin single copy gene (T/S) ratio, indicating that the older the mother, the longer the newborn’s telomeres, possibly indicating a compensatory mechanism. In the same study, longitudinal assessment of preterm infants who had TL measurements available at the age of 5 years, suggests that TL attrition rate is negatively correlated with increasing GA [61]. In accordance are the findings of the systematic review and meta-analysis of Niu et al. [62]. In this study, IUGR was associated with shorter placental TL, while PTB with longer birth TL, but only as measured by qPCR and not by the method of telomere restriction fragment (TRF). This, may explain the lack of correlation of TL measurements, when assessed by different laboratory methods, which may explain controversies in study results. The TRF method has been proposed as a method of high accuracy, as it results in lower variation than the PCR method [148]. However, possibly due to its time consuming, labor intensive, costly and expertise-demanding characteristics, TRF is used in very few studies compared with PCR.
Furthermore, in the Niu et al. [62] review and meta-analysis, IUGR was associated with shorter birth TL, only when birth TL was measured in placenta, but not in newborn blood. TL is sensitive to the types of tissues used for measurement, as previous studies reported the placental TL relatively longer than cord blood TL [149]. Nevertheless, irrespective of the studied biological matrix (i.e. cord leukocytes or placenta), newborn TL measurements remain predictive of TL in leukocytes at the age of 4 years [43]. Although, the choice of sample type is a complex matter, involving numerous issues such as, attainability, availability and ethics, it must be made clear that when comparing studies using different biological samples, this could lead to discordant conclusions.
Foetal membranes from term labors also showed TL reduction compared with those from the PTB group. Telomerase activity did not change in foetal membranes irrespective of pregnancy outcome. The authors suggested, that telomere shortening in foetal membranes is indicative of senescence associated with the triggering of labor at term [105]. The longer TL recorded in PTB offspring, is a surprising finding since PTB has been associated with early aging phenotypes [150], but may be explained by the fewer cellular replications and reduced DNA turnover in the final several weeks of gestation, otherwise missed when born preterm [151]. Indeed, in a twin study, placental TL gradually decreased with GA, indicating that enhanced TL attrition is more prominent with advancing GA [152]. Adjustments for GA, should be made when measuring birth TL in all studies between groups of different maternal nutritional exposures.
On the other hand, the study by Saroyo et al. showed that telomere T/S ratio of the placenta did not differ between PTB and term labor despite difference in GA, suggesting perhaps similar TLs in the timing of sampling between the two groups, due to early telomere shortening in PTB that mimics the term placenta [104]. The timing of sampling is indeed of extreme importance when assessed in PTB infants, especially if one considers the effects of trauma in the NICU, and the previously reported enhanced TL shortening of preterm infants experiencing these adversities after birth [68].
On the contrary, GA has also been positively associated with offspring RTL. Longitudinal data, indicate an increased LTL attrition in those born before 37 weeks of GA, as well as in those who gained weight as adults (29 years) but conditional BMI gain at 2 and 11 years, was not associated with RTL [107]. Previous studies have shown however that birth TL differs according to body size and GA of the newborn [153,154]. Maternal anthropometric characteristics may perhaps explain to an extent, why studies reach inconclusive outcomes when comparing TLs from spontaneous PTB offspring, compared with term born infants [106].
Another longitudinal cohort, followed premature newborns into their adulthood by studying the TL in their saliva and assessing their lung function. A positive correlation between TL and abnormal lung airflow in an adult population born prematurely, was observed, but there was no apparent association with perinatal causes. It is speculated that there is continuous oxidative stress of the airways, which leads to persistent inflammation, lung function alteration and increased sensitivity for chronic obstructive pulmonary disease [70]. Furthermore, TL attrition rates are associated with diseases characterised by increased inflammation and oxidative stress [155], thus affecting TL regulation in the offspring in an independent manner.
Evidence to date have confirmed that newborn TL strongly predicts child- and adulthood TL [43,148], while it is the strongest predictor of TL change over time [156,157]. Thus, TL measured at any point in life is jointly determined by TL at birth and subsequent TL attrition in later life, with a rapid attrition rate before the age of 5 years old and then remains relatively stable for the remaining of life course [158]. TL at birth indeed has been reported to vary as much as 3000-5000 bp inter-individually, a proportion that is well above the overall TL attrition occurring in the whole lifespan [159]. Therefore, birth TL is extremely important for the lifetime health trajectory and aging pace rhythm determines the interindividual variation of adult TL, which in turn has been proposed as a biomarker and potential contributor to the development of aging-related chronic diseases [160,161].
Maternal nutrition, placental-newborn telomeres and PTB
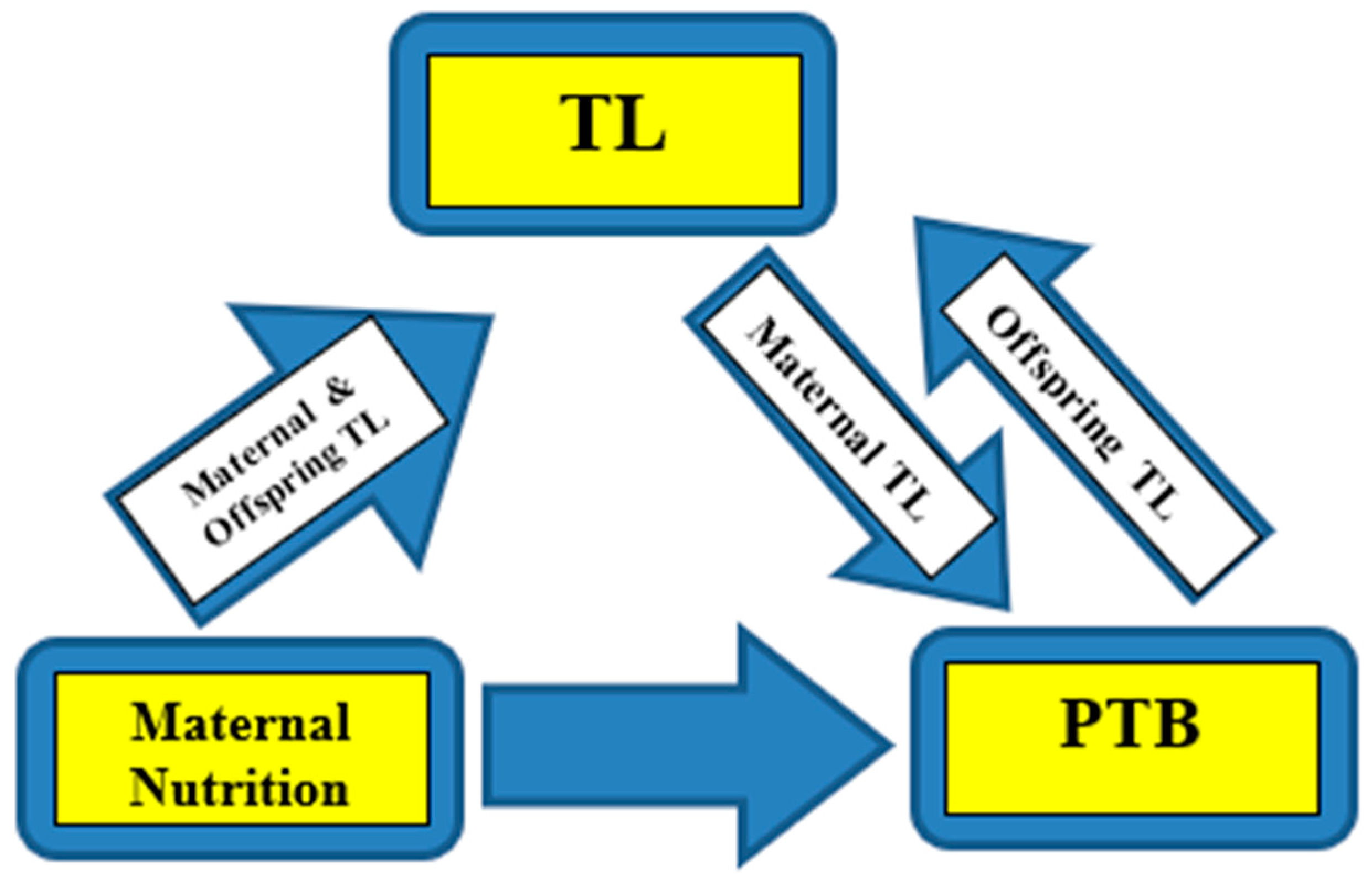
From the above evidence, it appears that maternal nutrition affects PTB risk, partly through its influence on maternal TL. Indeed, the telomere-regulated-clock mechanism determines the length of gestation, leading to the onset of labor (parturition), and at the same time PTB is a major determinant of offspring TL regulation. The strength of the associations and the extent of the influence from covariates, remains to be elucidated in future research. Furthermore, the question of whether maternal TL, is simply a biomarker of maternal nutritional status and in effect PTB risk, or a causative factor of PTB through nutritional habits and status, to date remains unanswered.
Following telomere expansion at the beginning of pregnancy, TL in gestational tissues (i.e., the placenta and fetal membranes) gradually shortens throughout the remainder of pregnancy. The rate of telomere shortening can be affected by nutritional practices, through their inflammatory/anti-inflammatory and oxidative/antioxidant properties [41,162], while indirectly influencing other aspects of health also known to affect TL, such as mental health (MH). Indeed, nutrition is associated with MH outcomes, such as a higher risk for developing depression and anxiety [163,164] while, on the other hand, MH may affect nutritional status (e.g. through appetite regulation) [165]. Additionally, evidence shows a link between various types of MH disorders and TL, including stress and anxiety [166,167,168], bipolar disorder [169,170], and psychotic disorders [171,172]. Thus, it appears that MH may independently affect maternal and offspring TL, through its effects on maternal dietary habits. To our knowledge, no studies to date however, have examined the bi-directional relationship between maternal nutrition and MH, assessing simultaneously TL regulation in the premature offspring.
As previously mentioned, due to the critical role of birth TL in determining TL in later life [154,173,174,175], it is intriguing to explore the potential mediation role of TL biology, underlying the relationship between intrauterine exposures and aging-related diseases by first comparing TL at birth between PTB and term birth outcomes. The current findings, thus emphasize the necessity for prospective, longitudinal studies that investigate the relationship between placenta / newborn TL and maternal nutrition - particularly during conception and pregnancy - in PTB risk. With the use of machine learning, algorithms and big data analysis, insights on the exact relationship of the aforementioned factors may be provided, also taking into account personal characteristics, genetics profiles, MH, way of life and general wellbeing.
Figure 3.
Representation of the associations. Maternal nutrition affects PTB risk, partly through its influence on maternal TL. On the other hand, maternal TL independently affects PTB risk, and at the same time PTB is a major determinant of offspring TL regulation. PTB: Preterm birth, TL: Telomere length.
Figure 3.
Representation of the associations. Maternal nutrition affects PTB risk, partly through its influence on maternal TL. On the other hand, maternal TL independently affects PTB risk, and at the same time PTB is a major determinant of offspring TL regulation. PTB: Preterm birth, TL: Telomere length.
Limitations of the study
This study has several limitations. First, studies included in this review used different research methodologies and assessment methods, variable set outcomes and exposures and geographical, demographic and anthropometric population characteristics. Population sizes are furthermore extremely variant, with many studies having small population sizes and thus power of study. Secondly, many studies did not adjust for sex and race/ethnicity. Thirdly, exposures especially those with key roles in the outcomes of studies, for example MH status (i.e. subclinical depression, traumatic experiences, eating disorders etc.) remained unexplored or unadjusted for. Fourthly, the well documented difficulty leading to compromised validity and reliability of nutritional intake and status assessments, was not adequately addressed in the majority of studies.
Also, we searched for relevant publications only in 2 electronic databases and did not include any unpublished studies or articles such as meeting abstracts and dissertations, which might have introduced publication biases. Finally, we included solely English written articles, which might also have caused the omission of articles published in other languages.
Future studies using individual participant data with adjustments for sex, race/ethnicity, anthropometry and other potential confounders, are imperative for valid comparisons. In addition, cross-sectional studies do not provide information of causality, because of the unclear temporality between the exposure and outcome and this should be considered in review and meta-analysis studies.
5. Conclusions
There is a relationship between maternal nutrition, placental-newborn TL and PTB. Specifically, maternal nutrition influences both positively and negatively PTB risk, to an extent through its influence on maternal TL. Furthermore, maternal TL independently affects PTB risk, while at the same time PTB appears as a major determinant of offspring TL regulation. The strength of the associations and the extent of the influence from various covariates, remains largely unexplored. The question also remains of whether maternal TL, is simply a biomarker of maternal nutritional status and in effect PTB risk, or a causative factor of PTB, through nutritional practices and status.
From a nutritional point of view, a diet high in carbohydrates and low in protein in the 3rd trimester and fat in the 1st trimester of pregnancy, was associated with PTB and SGA infants. Micronutrients and their adequate intake, may also play an important role in PTB risk. In particular, iron, zinc, B vitamins (folic acid, B6, B12) seem to affect PTB prevention and folic acid, vitamin D, B12 and n-3 PUFAs are factors that aid in maintaining both the placental and infant TL. However, the strength of the evidence is not adequate enough to reach definite conclusions and in effect the formulation of clinical guidelines.
In addition, placental telomeres play an important role in PTB and act as a biomarker of both the mother’s and the newborn’s nutritional status. Indeed, placental and maternal TL in PTBs is reported shorter compared with full-term pregnancies, thus TL may be considered as a biomarker of prognostic value for the adverse outcome of premature labor. Maternal leukocytes’ TL on the other hand, at the beginning of pregnancy, constitute a prognostic indicator of maternal biological aging and further, indirectly, a prognostic indicator of the offspring’s health from fetal life to the early post-natal period. Since premature infants exhibit a more rapid TL attrition in early life, while a measurable decline in TL during NICU hospitalization has been reported, the rate of TL change in early life, may convey information about prematurity and NICU exposures, that carry both immediate and long-term health risks.
The findings of this review, signify the urgent need for further research that will assess the relationship of the abovementioned factors simultaneously, especially in longitudinal observational studies, following populations from pregnancy to offspring’s’ adult life. Preventative medicine and future research may use this information regarding TL (maternal and placental) as well as, maternal nutritional status, for screening of high-risk preterm delivery or pregnancies, quantification of premature offspring adversity burden in early life and recording of their long-term health consequences in offspring adult life.
Author Contributions
It should be noted that each author has made substantial contributions to the completion of the current article. All authors have approved the submitted version. Furthermore, all authors agree to be personally accountable for their own contributions and ensuring that questions related to the accuracy or integrity of any part of the work, even ones in which they were not personally involved, are appropriately investigated, resolved, and documented in the literature. Nikoletta Lis and Dr Irene Tzanetakou contributed to the aspects of conceptualization, data curation, methodology design, investigation, resources, project administration, and visualization. Supervision, Dr Irene Tzanetakou. The writing part, both the original draft and review editing conducted by all authors, Nikoletta Lis, Demetris Lamnisos, Aikaterini Bograkou-Tzanetakou , Elena Hadjimbei and Irene Tzanetakou.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Ethical issues
The studies used in this literature review comply with a series of conditions such as the validity of the content from accredited databases, acceptance by the National Bioethics Commission and meeting the rules of ethics and research, without the existence of conflicts of interest.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Preterm birth, Fact sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/preterm-birth#:~:text=Across%20countries%2C%20the%20rate%20of,of%20babies%20born%20in%202020 (accessed on 9 October 2023).
- Goldenberg, R.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of PTB. The Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Wagura, P.; Wasunna, A.; Laving, A.; Wamalwa, D.; Ng’ang’a, P. Prevelance and factors associated with PTB at Kenyatta national hospital. BMC Pregnancy and Childbirth 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Suman, V.; Luther, E. Preterm Labor. 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK536939/ (accessed on 10 August 2023).
- Morgan, T. Placental insufficiency is a leading cause of preterm labor. NeoReviews 2014, 15, e518–e525. [Google Scholar] [CrossRef]
- Glover, A.; Manuck, T. Screening for spontaneous PTB and resultant therapies to reduce neonatal morbidity and mortality: A review. Seminars in Fetal & Neonatal Medicine 2017, 23, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.B.; Buhimschi, C.S.; Norwitz, E.R. Normal labor: mechanism and duration. Obstetrics and Gynecology Clinics 2005, 32, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Gibb, W.; Challis, J.R.G. Mechanism of term and PTB. Journal of Obstetrics and Gynaecology Canada 2020, 23, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Mousiolis, A. The role of leptin and syncytin upon the placenta function. (Publication No.28617) [Doctoral thesis, University of Thessaly, Greece] National Documentation Centre 2011. [CrossRef]
- Wang, Y.; Zhao, S. Vascular Biology of the Placenta; Integrated Systems Physiology: From Molecules to Function to Disease. San Rafael (CA) 2010. [CrossRef] [PubMed]
- Gete, D.; Waller, M.; Mishra, G. Effects of maternal diets on PTB and low birth weight: a systematic review. British Journal of Nutrition 2020, 12, 446–461. [Google Scholar] [CrossRef] [PubMed]
- McAninch, D.; Bianco-Miotto, T.; Gatford, K.L.; Leemaqz, S.Y.; Andraweera, P.H.; Garrett, A.; Plummer, M.D.; Dekker, G.A.; Roberts, C.T.; Smithers, L.G.; et al. The metabolic syndrome in pregnancy and its association with child telomere length. Diabetologia 2020, 63, 2140–2149. [Google Scholar] [CrossRef]
- Mizgier, M.; Jarzabek-Bielecka, G.; Mruczyk, K. Maternal diet and gestational diabetes mellitus development. The Journal of Maternal-Fetal & Neonatal Medicine 2021, 34, 77–86. [Google Scholar] [CrossRef]
- Brett, E.; Ferraro, Z.M.; Yockell-Lelievre, J.; Gruslin, A.; Adamo, K.B. Maternal-Fetal nutrient transport in pregnancy pathologies :The role of the placenta. International Journal of Molecular Sciences 2014, 15, 16153–16185. [Google Scholar] [CrossRef]
- Manna, S.; McCarthy, C.; McCarthy, F.P. Placental ageing in adverse pregnancy outcomes: Telomere shortening, cell senescence, and mitochondrial dysfunction. Oxidative medicine and cellular longevity 2019, 2019(3095383), 11. [Google Scholar] [CrossRef] [PubMed]
- Jirkovska, M.; Korabecna, M.; Lassakova, S. Telomeres and telomerase activity in the human placenta. In: Telomerase and non-Telomerase mechanisms of telomere maintenance. Morrish, T.A. Ed.; IntechOpen 2019; pp 1-20. [CrossRef]
- Connor, K.; Kibscull, M.; Matysiak-Zablocki, E.; Ngoc Nguyen, T.; Matthews, S.G.; Lye, S.; Bloise, E. Maternal malnutrition impacts placental morphology and transport. An origin for poor offspring growth and vulnerability to disease. The Journal of Biochemistry 2019, 78. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Phillips, J.A.; Bianco-Miotto, T.; McAninch, D.; Goh, Z.; Anderson, P.H.; Roberts, C.T. Reduced dietary calcium and vitamin D results in PTB and altered placental morphogenesis in mice during pregnancy. Reproductive Sciences 2020, 27, 1330–1339. [Google Scholar] [CrossRef]
- Dunlop, A.; Taylor, R.N.; Tangpricha, V.; Fortunato, S.; Menon, R. Maternal micronutrient status and preterm versus term birth for black and white US women. Reproductive Sciences 2012, 19, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.; Robinson, S.; Barker, D.J.; Osmond, C.; Cox, V. Maternal nutrition in early pregnancy in relation to placental and fetal growth. BMJ 1996, 312, 410–4. [Google Scholar] [CrossRef]
- Oh, C.; Keats, E.C.; Bhutta, Z.A. Vitamin and mineral supplementation during pregnancy on maternal birth, child health and development outcomes in low- and middle- income countries: A systematic review and meta-analysis. Nutrients 2019, 419, 1–30. [Google Scholar] [CrossRef]
- Emond, J.; Karagas, M.R.; Baker, E.R.; Gilbert-Diamond, D. Better diet quality during pregnancy is associated with a reduced likelihood of an infant born small for gestational age: an analysis of the prospective New Hampshire birth cohort study. J Nutrients 2018, 148, 22–30. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, H.; Perkins, A.; Wang, Y.; Sun, J. Maternal dietary nutrient intake and its association with PTB: A case-control study in Beijing, China. Nutrients 2017, 9, 221. [Google Scholar] [CrossRef]
- Heppe, D.; Steegers, E.A.; Timmermans, S.; den Breeijen, H.; Tiemeier, H.; Hofman, A.; Jaddoe, V.W. Maternal fish consumption, fetal growth and the risks of neonatal complications: the Generation R Study. British Journal Nutrition 2011, 105, 938–94. [Google Scholar] [CrossRef]
- Heppe, D.; van Dam, R.M.; Willemsen, S.P.; den Breeijen, H.; Raat, H.; Hofman, A.; Steegers, E.A.; Jaddoe, V.W. Maternal milk consumption, fetal growth, and the risks of neonatal complications: the Generation R Study. American Journal of Clinical Nutrition 2011, 94, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, T.; Osterdal, M.L.; Knudsen, V.K.; Haugen, M.; Meltzer, H.M.; Bakketeig, L.; Olsen, S.F. Association between a Mediterranean-type diet and risk of PTB among Danish women: a prospective cohort study. Acta Obstetricia et Gynecologica Scandinavica 2008, 87, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Myhre, R.; Brantsæter, A.L.; Myking, S.; Gjessing, H.K.; Sengpiel, V.; Meltzer, H.M.; Haugen, M.; Jacobsson, B. Intake of probiotic food and risk of spontaneous preterm delivery. The American Journal of Clinical Nutrition 2011, 93, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Moore, V.; Worsley, A.; Robinson, J.S. Dietary composition of pregnanct women is related to size of the baby at birth. The journal of nutrition 2004, 134, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Haugen, M.; Meltzer, H.M.; Brantsaeter, A.L.; Mikkelsen, T.; Osterdal, M.L.; Alexander, J.; Olsen, S.F.; Bakketeig, L. Mediterranean-type diet and risk of PTB among women in the Norwegian Mother and Child Cohort Study (MoBa): a prospective cohort study. Acta Obstetricia et Gynecologica Scandinavica 2008, 87, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, U.; Manjrekar, R.; Rivera, J.; Gonzalez-Cossio, T.; Martorell, R. Micronutrients and pregnancy outcome : A review of the literature. Nutrition Research 1999, 9, 103–159. [Google Scholar] [CrossRef]
- Shammas, M. Telomeres, lifestyle, cancer and aging. Current Opinion in Clinical Nutrition and Metabolic Care 2011, 14, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Galiè, S.; Canudas, S.; Muralidharan, J.; García-Gavilán, J.; Bulló, M.; Salas-Salvadó, J. Impact of nutrition on telomere health: systematic review of observational cohort studies and randomized clinical trials. Advances in Nutrition 2020, 11, 576–601. [Google Scholar] [CrossRef]
- Ballan, E.; Decottignies, A.; Deldicque, L. Physical activity and nutrition: Two promising strategies for telomere maintenance? Nutrients 2018, 10, 1942. [Google Scholar] [CrossRef] [PubMed]
- Polettini, J.; da Silva, M. Telomere-Related disorders in fetal membranes associated with birth and adverse pregnancy outcomes. Frontiers in Physiology 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Ferrari, F.; Facchinetti, F.; Saade, G.; Menon, R. Placental telomere shortening in stillbirth: a sign of premature senescence? Journal of Maternal-Fetal & Neonatal Medicine 2016, 29, 1283–8. [Google Scholar] [CrossRef]
- Menon, R.; Boldogh, I.; Hawkins, H.K.; Woodson, M.; Polettini, J.; Ali Syed, T.; Fortunato, S.J.; Saade, G.R.; Papaconstantinou, J.; Taylor, R.N. Histological evidence of oxidative stress and premature senescence in preterm premature rupture of the human fetal membranes recapitulated in vitro. The American Journal of Pathology 2014, 184, 1740–1751. [Google Scholar] [CrossRef] [PubMed]
- Sultana, Z.; Maiti, K.; Dedman, L.; Smith, R. Is there a role for placental senescence in the genesis of obstetric complications and fetal growth restriction? American Journal of Obstetrics & Gynecology 2018, 218, S762–S773. [Google Scholar] [CrossRef]
- Hallows, S.; Regnault, T.R.H.; Betts, D.H. The long and the short of it: The role of telomeres in fetal origins of adult disease. Journal of pregnancy 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.; Levandowski, M.; Ridout, S.J.; Gantz, L.; Goonan, K.; Palermo, D.; Price, L.H.; Tyrka, A.R. Early life adversity and telomere length: a meta-analysis. Molecular Psychiatry 2018, 23, 858–71. [Google Scholar] [CrossRef]
- Factor-Litvak, P.; Susser, E. The importance of early life studies of telomere attrition. Paediatric and perinatal epidemiology 2015, 29, 144. [Google Scholar] [CrossRef]
- Sarkar, S.; Godara, K.; Sarkar, S.; Ghosh, S.; Maity, A.; Team, G.; Sindhu, B.; Bhatnagar, S.; Wadhwa, N.; Maitra, A.; et al. Preterm Birth is Associated with Reduction of Maternal Telomere Length During Pregnancy. Research Square 2021. [Google Scholar] [CrossRef]
- Panelli, D.; Bianco, K. Cellular aging and telomere dynamics in pregnancy. Current Opinion in Obstetrics and Gynecology 2022, 34, 57–61. [Google Scholar] [CrossRef]
- Martens, D.S.; van der Stukken, C.; Derom, C.; Thiery, E.; Bijnens, E.M.; Nawrot, T.S. Newborn telomere length predicts later life telomere length: Tracking telomere length from birth to child-and adulthood. EBioMedicine 2021, 103164, 63. [Google Scholar] [CrossRef] [PubMed]
- Benetos, A.; Verhulst, S.; Labat, C.; Lai, T.; Gireld, N.; Toupance, S.; Zannad, F.; Rossignol, P.; Aviv, A. Telomere length tracking in children and their parents: implications for adult onset diseases. FASEB J 2019, 33, 14248–53. [Google Scholar] [CrossRef]
- Entringer, S.; de Punder, K.; Buss, C.; Wadhwa, P.D. The fetal programming of telomere biology hypothesis: an update. Philosophical Transactions of the Royal Society of London. Biological sciences 2018, 373. [Google Scholar] [CrossRef]
- Entringer, S.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Buss, C.; Shahbaba, B.; Gillen, D.L.; Venkataramanan, R.; Simhan, H.N.; Wadhwa, P.D. Maternal folate concentration in early pregnancy and newborn telomere length. Annals of Nutrition and Metabolism 2015, 66, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, G.J.; Lee, D.; Ko, J.H.; Lim, I.; Bang, H.; Koes, B.W.; Seong, B.; Lee, D.C. Higher maternal vitamin D concentrations are associated with longer leukocyte telomeres in newborns. Maternal & child nutrition 2018, 14, e12475. [Google Scholar] [CrossRef]
- Mirzakhani, H.; De Vivo, L.; Leeder, J.S.; Gaedigk, R.; Vyhlidal, C.A.; Weiss, S.T.; Tantisira, K. Early pregnancy intrauterine fetal exposure to maternal smoking and impact on fetal telomere length. European Journal of Obstetrics & Gynecology and Reproductive Biology 2017, 218, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Wojcicki, J.; Olveda, R.; Heyman, M.B.; Elwan, D.; Lin, J.; Blackburn, E.; Epel, E. Cord blood telomere length in Latino infants: relation with maternal education and infant sex. Journal of Perinatology 2016, 36, 235–41. [Google Scholar] [CrossRef]
- Tellechea, M.; Fernandez-Gianotti, T.; Alvarinas, J.; Gonzalez, C.D.; Sookoian, S.; Pirola, C.J. Telomere length in the two extremes of abnormal fetal growth and the programming effect of maternal arterial hypertension. Scientific Reports 2015, 5, 7869. [Google Scholar] [CrossRef]
- Martens, D.; Plusquin, M.; Gyselaers, W.; de Vivo, I.; Nawrot, T. Maternal pre-pregnancy body mass index and newborn telomere length. BMC Med 2016, 14, 148. [Google Scholar] [CrossRef]
- Xu, J.; Ye, J.; Wu, Y.; Zhang, H.; Luo, Q.; Han, C.; Ye, X.; Wang, H.; He, J.; Huang, H.; et al. Reduced fetal telomere length in gestational diabetes. Plos one 2014, 9, e8616. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; Du, M.; Wong, J.Y.Y.; Han, J.; de Vivo, I. Paternal age at birth is associated with offspring leukocyte telomere length in the nurses’ health study. Human Reproduction 2012, 27, 3622–3. [Google Scholar] [CrossRef]
- Broer, L.; Codd, V.; Nyholt, R.D.; Deelen, J.; Mangino, M.; Willemsen, G.; Albrecht, E.; Amin, N.; Beekman, M.; de Geus, J.C.E.; et al. Meta-analysis of telomere length in 19,713 subjects reveals high heritability, stronger maternal inheritance and a paternal age effect. European Journal of Human Genetics 2013, 21, 1163–8. [Google Scholar] [CrossRef]
- Kimura, M.; Cherkas, L., F.; Kato, B., S.; Demissie, S.; Hjelmborg, J.B.; Brimacombe, M.; Cupples, A.; Hunkin, J.L.; Gardner, J.P.; Lu, X.; et al. Offspring’s leukocyte telomere length, paternal age, and telomere elongation in sperm. Plos Genetics 2008, 4, e37. [Google Scholar] [CrossRef] [PubMed]
- Nordfjall, K.; Larefalk, A.; Lindgren, P.; Holmberg, D.; Roos, G. Telomere length and heredity: indications of paternal inheritance. Proceedings of the National Academy of Sciences. U S A 2005, 102, 16374–8. [Google Scholar] [CrossRef] [PubMed]
- Send, T.; Gilles, M.; Codd, V.; Wolf, I.; Bardtke, S.; Streit, F.; Strohmaier, J.; Frank, J.; Schendel, D.; Sütterlin, M.W.; et al. Telomere length in newborns is related to maternal stress during pregnancy. Neuropsychopharmacology 2017, 42, 2407–13. [Google Scholar] [CrossRef]
- Entringer, S.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Buss, C.; Simhan, H.N.; Wadhwa, P.D. Maternal estriol concentrations in early gestation predict infant telomere length. The Journal of Clinical Endocrinology & Metabolism 2015, 100, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Gotlib, I.H.; LeMoult, J.; Colich, N.L.; Foland-Ross, L.C.; Hallmayer, J.; Joormann, J.; Lin, J.; Wolkowitz, O.M. Telomere length and cortisol reactivity in children of depressed mothers. Molecular psychiatry 2015, 20, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.; Hansen, N.I.; Bell, E.F.; Shankaran, S.; Laptook, A.R.; Walsh, M.C.; Hale, E.C.; Newman, N.S.; Schibler, K.; Carlo, W.A.; et al. Neonatal outcomes of extremely preterm infants from the NICHD neonatal research network. Pediatrics 2010, 126, 443–56. [Google Scholar] [CrossRef] [PubMed]
- Vasu, V.; Turner, K.J.; George, S.; Greenall, J.; Slijepcevic, P.; Griffin, D.K. Preterm infants have significantly longer telomeres than their term born counterparts. Plos one 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Li, K.; Xie, C.; Wen, X. Adverse birth outcomes and birth telomere length: a systematic review and meta-analysis. The Journal of Pediatrics 2019, 215, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Smeets, C.; Codd, V.; Samani, N.J.; Hokken-Koelega, A.C.S. Leucocyte telomere length in young adults born preterm: Support for accelerated biological ageing. Plos one 2015, 10, e0143951. [Google Scholar] [CrossRef]
- Provenzi, L.; Giorda, R.; Fumagalli, M.; Brambilla, M.; Mosca, F.; Borgatti, R.; Montirosso, R. Telomere length and salivary cortisol stress reactivity in very preterm infants. Early Human Development 2019, 129, 1–4. [Google Scholar] [CrossRef]
- Chen, L.; Tan, K.M.L.; Gong, M.; Chong, M.F.; Tan, K.H.; Chong, Y.S.; Tan, K.H.; Chong, Y.S.; Meaney, M.J.; Gluckman, P.D.; et al. Variability in newborn telomere length is explained by inheritance and intrauterine environment. BMC medicine 2022, 20, 20. [Google Scholar] [CrossRef] [PubMed]
- Saigal, S.; Day, K.L.; van Lieshout, R.J.; Schmidt, L.A.; Morrison, K.M.; Boyle, M.H. Health, wealth, social integration, and sexuality of extremely low-birth-weight prematurely born adults in the fourth decade of life. JAMA Pediatrics 2016, 170, 678. [Google Scholar] [CrossRef] [PubMed]
- Belfort, M.; Qureshi, F.; Litt, J.; Bosquet Enlow, M.; De Vivo, I.; Gregory, K.; Tiemeier, H. Telomere length shortening in hospitalized preterm infants: A pilot study. Plos one 2021, 16, e0243468. [Google Scholar] [CrossRef] [PubMed]
- Provenzi, L.; Scotto di Minico, G.; Giorda, R.; Montirosso, R. Telomere length in preterm infants: a promising biomarker of early adversity and care in the neonatal intensive care unit? . Frontiers in endocrinology 2017, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.; Vasu, V.; Greenall, J.; Griffin, D.K. Telomere length analysis and preterm infant health: the importance of assay design in the search for novel biomarkers. Biomarkers in Medicine 2014, 8, 485–98. [Google Scholar] [CrossRef] [PubMed]
- Hadchouel, A.; Marchand-Martin, L.; Franco-Montoya, M.L.; Peaudecerf, L.; Ancel, P.Y.; Delacourt, C. Salivary Telomere Length and Lung Function in Adolescents Born Very Preterm: A Prospective Multicenter Study. Plos one 2015, 10, e0136123. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Journal of Clinical Epidemiology 2009, 62, 1006–1. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.L.; Arocha, J.F.; Zhang, J. Thinking and reasoning in medicine. The Cambridge handbook of thinking and reasoning 2005, 14, 727–750. [Google Scholar]
- Parisi, F.; Savasi, V.M.; di Bartolo, I.; Mandia, L.; Cetin, I. Associations between first trimester maternal nutritional score, early markers of placental function, and pregnancy outcome. Nutrients 2020, 12, 1799. [Google Scholar] [CrossRef]
- Hillesund, E.R.; Bere, E.; Sagedal, L.R.; Vistad, I.; Seiler, H.L.; Torstveit, M.K.; Øverby, N.C. Pre-pregnancy and early pregnancy dietary behavior in relation to maternal and newborn health in the Norwegian Fit for Delivery study–a post hoc observational analysis. Food & nutrition research, 2018; 62. [Google Scholar] [CrossRef]
- Tith, R.M.; Bilodeau-Bertrand, M.; Lee, G.E.; Healy-Profitós, J.; Auger, N. Fasting during Ramadan increases risk of very PTB among Arabic-speaking women. The Journal of Nutrition 2019, 149, 1826–1832. [Google Scholar] [CrossRef]
- Kriss, J.L.; Ramakrishnan, U.; Beauregard, J.L.; Phadke, V.K.; Stein, A.D.; Rivera, J.A.; Omer, S.B. Yogurt consumption during pregnancy and preterm delivery in Mexican women: A prospective analysis of interaction with maternal overweight status. Maternal & child nutrition 2018, 14, e12522. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.S.; He, J.R.; Chen, Q.; Lu, J.; Wei, X.; Zhou, Q.; Chan, F.; Zhang, L.; Chen, N.; Qiu, L.; et al. Maternal dietary patterns during pregnancy and preterm delivery: a large prospective cohort study in China. Nutrition journal 2018, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Cardoso, M.; Dias, C.C.; Pereira-da-Silva, L.; de Silva, D. Eating Habits During Pregnancy of Women Giving Birth Very Prematurely: An Exploratory Analysis. Acta Médica Portuguesa 2023, 36, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Chia, A.R.; De Seymour, J.V.; Colega, M.; Chen, L.W.; Chan, Y.H.; Aris, I.M.; Tint, M.T.; Quah, P.L.; Godfrey, K.M.; Yap, F.; et al. A vegetable, fruit, and white rice dietary pattern during pregnancy is associated with a lower risk of PTB and larger birth size in a multiethnic Asian cohort: the Growing Up in Singapore Towards healthy Outcomes (GUSTO) cohort study. The American journal of clinical nutrition 2016, 104, 1416–1423. [Google Scholar] [CrossRef]
- Martin, C.L.; Sotres-Alvarez, D.; Siega-Riz, A.M. Maternal dietary patterns during the second trimester are associated with preterm birth. The Journal of nutrition 2015, 145, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ye, R.; Zhang, L.; Li, H.; Liu, J.; Ren, A. Periconceptional folic acid supplementation and the risk of preterm births in China: a large prospective cohort study. International Journal of Epidemiology 2014, 43, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yuan, Y.; Kong, C.; Ma, Q.; Ye, H.; Jing, W.; Liu, J.; Liu, M. The association between periconceptional folic acid supplementation and the risk of preterm birth: a population-based retrospective cohort study of 200,000 women in China. European Journal of Nutrition 2021, 60, 2181–2192. [Google Scholar] [CrossRef]
- Johnston, E.O.; Sharma, A.J.; Abe, K. Association between maternal multivitamin use and PTB in 24 states, pregnancy risk assessment monitoring system, 2009–2010. Maternal and child health journal 2016, 20, 1825–1834. [Google Scholar] [CrossRef]
- Olapeju, B.; Saifuddin, A.; Wang, G.; Ji, Y.; Hong, X.; Raghavan, R.; Summers, A.; Keiser, A.; Ji, H.; Zuckerman, B.; et al. Maternal postpartum plasma folate status and PTB in a high-risk US population. Public health nutrition 2019, 22, 1281–1291. [Google Scholar] [CrossRef]
- Ren, M.; Zhao, J.; Wang, B.; An, H.; Li, Y.; Jia, X.; Wang, J.; Wang, S.; Yan, L.; Liu, X.; et al. Associations between hair levels of trace elements and the risk of PTB among pregnant women: A prospective nested case-control study in Beijing Birth Cohort (BBC), China. Environment International 2022, 158, 106965. [Google Scholar] [CrossRef]
- Irwinda, R.; Wibowo, N.; Putri, A.S. The concentration of micronutrients and heavy metals in maternal serum, placenta, and cord blood: a cross-sectional study in PTB. Journal of pregnancy 2019, 2019, 5062365. [Google Scholar] [CrossRef] [PubMed]
- Chiudzu, G.; Choko, A.T.; Maluwa, A.; Huber, S.; Odland, J. Maternal serum concentrations of selenium, copper, and zinc during pregnancy are associated with risk of spontaneous PTB: a case-control study from Malawi. Journal of Pregnancy 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Soomro, T.K. Sideropaenic anaemia: Impact on perinatal outcome at tertiary care hospital. The Journal of the Pakistan Medical Association 2016, 66, 952–956. [Google Scholar] [PubMed]
- Chen, L.W.; Lim, A.L.; Colega, M.; Tint, M.T.; Aris, I.M.; Tan, C.S.; Chong, Y.S.; Gluckman, P.D.; Godfrey, K.M.; Kwek, K.; van Dam, R.M. Maternal folate status, but not that of vitamins B-12 or B-6, is associated with gestational age and PTB risk in a multiethnic Asian population. The Journal of nutrition 2015, 145, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Christoph, P.; Challande, P.; Raio, L.; Surbek, D. High prevalence of severe vitamin D deficiency during the first trimester in pregnant women in Switzerland and its potential contributions to adverse outcomes in the pregnancy. Swiss medical weekly 2020, 150. [Google Scholar] [CrossRef]
- Tahsin, T.; Khanam, R.; Chowdhury, N.H.; Hasan, A.T.; Hosen, M.B.; Rahman, S.; Roy, A.K.; Ahmed, S.; Raqib, R.; Baqui, A.H. Vitamin D deficiency in pregnancy and the risk of preterm birth: a nested case–control study. BMC pregnancy and childbirth 2023, 23, 322. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Wu, Y.; Huang, L.; Chen, X.; Zhang, Y.; Zhong, C.; Gao, Q.; Hong, M.; Hu, X.; Yang, X.; et al. Association between the maternal protein nutrition status during pregnancy and the risk of PTB. Maternal & Child Nutrition 2021, 17, e13043. [Google Scholar] [CrossRef] [PubMed]
- Miele, M.J.; Souza, R.T.; Calderon, I.M.; Feitosa, F.E.; Leite, D.F.; Rocha Filho, E.A.; Vettorazzi, J.; Mayrink, J.; Fernandes, K.G.; Vieira, M.; et al. Maternal nutrition status associated with pregnancy-related adverse outcomes. Nutrients 2021, 13, 2398. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, Q.; Fan, X.; Chen, H.; Chen, N.; Zhao, Y.; Qi, K. Associations of maternal polyunsaturated fatty acids with telomere length in the cord blood and placenta in Chinese population. Frontiers in Nutrition 2022, 8, 779306. [Google Scholar] [CrossRef]
- Yeates, A.J.; Thurston, S.W.; Li, H.; Mulhern, M.S.; McSorley, E.M.; Watson, G.E.; Shamlaye, C.F.; Strain, J.J.; Myers, G.J.; Davidson, P.W.; Broberg, K. PUFA status and methylmercury exposure are not associated with leukocyte telomere length in mothers or their children in the Seychelles Child Development Study. The Journal of Nutrition 2017, 147, 2018–2024. [Google Scholar] [CrossRef]
- Salihu, H.M.; Adegoke, K.K.; King, L.M.; Daas, R.; Paothong, A.; Pradhan, A.; Aliyu, M.H.; Whiteman, V.E. Effects of maternal carbohydrate and fat intake on fetal telomere length. Southern Medical Journal 2018, 111, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Vahter, M.; Broberg, K.; Harari, F. Placental and cord blood telomere length in relation to maternal nutritional status. The Journal of nutrition 2020, 150, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Daneels, L.; Martens, D.S.; Arredouani, S.; Billen, J.; Koppen, G.; Devlieger, R.; Nawrot, T.S.; Ghosh, M.; Godderis, L.; Pauwels, S. Maternal vitamin D and newborn telomere length. Nutrients 2021, 13, 2012. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Song, L.; Liu, Q.; Wu, M.; Bi, J.; Xu, L.; Xiong, C.; Cao, Z.; Xu, S.; Wang, Y. Association of maternal folic acid supplementation during pregnancy with newborn telomere length. Reproductive Toxicology 2022, 114, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Louis-Jacques, A.F.; Salihu, H.M.; King, L.M.; Paothong, A.; Sinkey, R.G.; Pradhan, A.; Riggs, B.M.; Siegel, E.M.; Salemi, J.L.; Whiteman, V.E. A positive association between umbilical cord RBC folate and fetal TL at birth supports a potential for fetal reprogramming. Nutrition Research 2016, 36, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Herlin, M.; Broberg, K.; Igra, A.M.; Li, H.; Harari, F.; Vahter, M. Exploring telomere length in mother–newborn pairs in relation to exposure to multiple toxic metals and potential modifying effects by nutritional factors. BMC medicine 2019, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Magnano San Lio, R.; Barchitta, M.; Maugeri, A.; La Rosa, M.C.; Caruso, M.; Giunta, G.C.; Panella, M.; Cianci, A.; Agodi, A. The effect of nutrients on telomere length of fetal DNA: findings from the Mamma & Bambino cohort. European Journal of Public Health 2021, 31 (Supplement_3), ckab164-074. [Google Scholar] [CrossRef]
- Myers, K.O.; Ibrahimou, B.; Yusuf, K.K.; Mauck, D.E.; Salihu, H.M. The effect of maternal vitamin C intake on fetal telomere length. The Journal of Maternal-Fetal & Neonatal Medicine 2021, 34, 1143–1148. [Google Scholar] [CrossRef]
- Saroyo, Y.B.; Wibowo, N.; Irwinda, R.; Prijanti, A.R.; Yunihastuti, E.; Bardosono, S.; Krisnadi, S.R.; Permata, P.I.; Wijaya, S.; Santawi, V.P.A. Oxidative stress induced damage and early senescence in preterm placenta. Journal of Pregnancy 2021, 9923761. [Google Scholar] [CrossRef]
- Colatto, B.N.; de Souza, I.F.; Schinke, L.A.A.; Noda-Nicolau, N.M.; da Silva, M.G.; Morceli, G.; Menon, R.; Polettini, J. Telomere length and telomerase activity in foetal membranes from term and spontaneous PTBs. Reproductive Sciences 2020, 27, 411–417. [Google Scholar] [CrossRef]
- Farladansky-Gershnabel, S.; Dekel, N.; Biron-Shental, T.; Shechter-Maor, G.; Amiel, A.; Weisz, A.; Benchetrit, S.; Zitman-Gal, T. Spontaneous Preterm Birth: Elevated Galectin-3 and Telomere Shortening May Reflect a Common Pathway of Enhanced Inflammation and Senescence. Reproductive Sciences 2023, 30, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Tarik, M.; Ramakrishnan, L.; Sinha, S.; Sachdev, H.P.S.; Tandon, N.; Roy, A.; Bhargava, S.K. Association of birth outcomes and postnatal growth with adult leukocyte telomere length: Data from New Delhi Birth Cohort. Maternal & Child Nutrition 2019, 15, e12857. [Google Scholar] [CrossRef] [PubMed]
- Blumfield, M.L.; Collins, C.E. High-protein diets during pregnancy: Healthful or harmful for offspring? The American Journal of Clinical Nutrition 2014, 100, 993–995. [Google Scholar] [CrossRef] [PubMed]
- Blumfield, M.L.; Hure, A.J.; Macdonald-Wicks, L.; Smith, R.; Collins, C.E. Systematic review and meta-analysis of energy and macronutrient intakes during pregnancy in developed countries. Nutrition Reviews 2012, 70, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Morisaki, N.; Nagata, C.; Yasuo, S.; Morokuma, S.; Kato, K.; Sanefuji, M.; Shibata, E.; Tsuji, M.; Senju, A.; Kawamoto, T.; et al. Japan Environment and Children's Study Group . Optimal protein intake during pregnancy for reducing the risk of fetal growth restriction: The Japan Environment and Children's Study. British Journal of Nutrition 2018, 120, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Ota, E.; Mori, R.; Middleton, P.; Tobe-Gai, R.; Mahomed, K.; Miyazaki, C.; Bhutta, Z.A. Zinc supplementation for improving pregnancy and infant outcome. Cochrane Database of Systematic Reviews 2015, 2015, CD000230. [Google Scholar] [CrossRef] [PubMed]
- Switkowski, K.M.; Jacques, P.F.; Must, A.; Kleinman, K.P.; Gillman, M.W.; Oken, E. Maternal protein intake during pregnancy and linear growth in the offspring. American Journal of Clinical Nutrition 2016, 104, 1128–1136. [Google Scholar] [CrossRef]
- Awasthi, S.; Chauhan, M.; Pandey, M.; Singh, S.; Singh, U. Energy and protein intake during pregnancy in relation to preterm birth: A case control study. Indian Pediatrics 2015, 52, 489–492. [Google Scholar] [CrossRef]
- Ota, E.; Hori, H.; Mori, R.; Tobe-Gai, R.; Farrar, D. Antenatal dietary education and supplementation to increase energy and protein intake. Cochrane Database of Systematic Reviews 2015, 2, CD000032. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Peng, X.; Zhang, S.; Wang, X.; Zhu, C. Folic acid and risk of preterm birth: a meta-analysis. Frontiers in Neuroscience 2019, 13, 1284. [Google Scholar] [CrossRef]
- Zhou, S.S.; Tao, Y.H.; Huang, K.; Zhu, B.B.; Tao, F.B. Vitamin D and risk of preterm birth: Up-to-date meta-analysis of randomized controlled trials and observational studies. Journal of Obstetrics and Gynaecology Research 2017, 43, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Bialy, L.; Fenton, T.; Shulhan-Kilroy, J.; Johnson, D.W.; McNeil, D.A.; Hartling, L. Vitamin D supplementation to improve pregnancy and perinatal outcomes: an overview of 42 systematic reviews. BMJ Open 2020, 10, e032626. [CrossRef] [PubMed]
- Gillespie, B. Nutritional Status and the Risk of Preterm Birth. In Evidence Based Global Health Manual for Preterm Birth Risk Assessment; Anumba, D.O., Jayasooriya, S.M., Eds.; Springer: Cham, 2022; pp. 41–51. [Google Scholar] [CrossRef]
- Rahmati, S.; Azami, M.; Badfar, G.; Parizad, N.; Sayehmiri, K. The relationship between maternal anemia during pregnancy with preterm birth: a systematic review and meta-analysis. The Journal of Maternal-Fetal & Neonatal Medicine 2020, 33, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wu, Y.; Zhang, N.; Liu, S.; Zhou, L. Disordered Maternal and Fetal Iron Metabolism Occurs in Preterm Births in Human. Disease Markers 2022, 2022, 1664474. [Google Scholar] [CrossRef] [PubMed]
- Kemppinen, L.; Mattila, M.; Ekholm, E.; Pallasmaa, N.; Törmä, A.; Varakas, L.; Mäkikallio, K. Gestational iron deficiency anemia is associated with preterm birth, fetal growth restriction, and postpartum infections. Journal of Perinatal Medicine 2021, 49, 431–438. [Google Scholar] [CrossRef]
- Khanam, R.; Kumar, I.; Oladapo-Shittu, O.; Twose, C.; Islam, A.A.; Biswal, S.S.; Raqib, R.; Baqui, A.H. Prenatal environmental metal exposure and preterm birth: a scoping review. International journal of environmental research and public health 2021, 18, 573. [Google Scholar] [CrossRef] [PubMed]
- Wolf, H.T.; Hegaard, H.K.; Huusom, L.D.; Pinborg, A.B. Multivitamin use and adverse birth outcomes in high-income countries: a systematic review and meta-analysis. American journal of obstetrics and gynecology 2017, 217, 404-e1. [Google Scholar] [CrossRef]
- Glazier, J.D.; Hayes, D.J.; Hussain, S.; D’Souza, S.W.; Whitcombe, J.; Heazell, A.E.; Ashton, N. The effect of Ramadan fasting during pregnancy on perinatal outcomes: a systematic review and meta-analysis. BMC pregnancy and childbirth 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Susser, E.; Ananth, C.V. Invited commentary: is prenatal fasting during Ramadan related to adult health outcomes? A novel and important question for epidemiology. American journal of epidemiology 2013, 177, 737–740. [Google Scholar] [CrossRef]
- Mitku, A.A.; Zewotir, T.; North, D.; Jeena, P.; Naidoo, R.N. Modeling differential effects of maternal dietary patterns across severity levels of preterm birth using a partial proportional odds model. Scientific reports 2020, 10, 5491. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.A.; Maslova, E.; Halldorsson, T.I.; Olsen, S.F. Characterization of dietary patterns in the Danish national birth cohort in relation to preterm birth. Plos one 2014, 9, e93644. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, S.; Soltani, S.; de Souza, R.J.; Forbes, S.C.; Toupchian, O.; Salehi-Abargouei, A. Associations between maternal dietary patterns and perinatal outcomes: a systematic review and meta-analysis of cohort studies. Advances in nutrition 2021, 12, 1332–1352. [Google Scholar] [CrossRef] [PubMed]
- Chia, A.R.; Chen, L.W.; Lai, J.S.; Wong, C.H.; Neelakantan, N.; van Dam, R.M.; Chong, M.F.F. Maternal dietary patterns and birth outcomes: a systematic review and meta-analysis. Advances in Nutrition 2019, 10, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Englund-Ögge, L.; Brantsæter, A.L.; Sengpiel, V.; Haugen, M.; Birgisdottir, B.E.; Myhre, R.; Meltzer, H.M.; Jacobsson, B. Maternal dietary patterns and preterm delivery: results from large prospective cohort study. BMJ 2014, 348, g1446. [Google Scholar] [CrossRef] [PubMed]
- McGuire, S. Scientific report of the 2015 dietary guidelines advisory committee. Washington, DC: US Departments of Agriculture and Health and Human Services, 2015. Advances in nutrition 2016, 7, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Abu-Saad, K.; Fraser, D. Maternal nutrition and birth outcomes. Epidemiologic reviews 2010, 32, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, F.Q. Body mass index: Obesity, BMI, and health: a critical review. Nutrition Today 2015, 50, 117–28. [Google Scholar] [CrossRef] [PubMed]
- Mulhern, M.S.; Broberg, K.; McAfee, A.; Laird, E.; Li, H.; Wallace, J.M.W.; McSorley, E.M.; Myers, G.J.; Davidson, P.W.; Thurston, S.W.; et al. Vitamin D status is a predictor of telomere length during pregnancy. Proceedings of the Nutrition Society 2012, 71(OCE2), E124. [Google Scholar] [CrossRef]
- Wong, K.K.; Cheng, F.; Mao, D.; Lim, C.K.; Wang, R.; Tam, C.H.; Joglekar, M.A.; Hardikar, A.A.; Jenkins, A.; Metzger, B.E.; et al. 275-OR: Maternal Vitamin D Levels during Pregnancy Is Associated with Childhood Telomere Length: Analysis from a Longitudinal Mother-Child Cohort. Diabetes, 2021; 70. [Google Scholar] [CrossRef]
- Reichetzeder, C.; Chen, H.; Föller, M.; Slowinski, T.; Li, J.; Chen, Y.P.; Lang, F.; Hocher, B. Maternal vitamin D deficiency and fetal programming-lessons learned from humans and mice. Kidney and Blood Pressure Research 2014, 39, 315–329. [Google Scholar] [CrossRef]
- Cabaset, S.; Krieger, J.P.; Richard, A.; Elgizouli, M.; Nieters, A.; Rohrmann, S.; Quack Lötscher, K.C. Vitamin D status and its determinants in healthy pregnant women living in Switzerland in the first trimester of pregnancy. BMC pregnancy and childbirth 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moores, C.J.; Fenech, M.; O’Callaghan, N.J. Telomere dynamics: the influence of folate and DNA methylation. Annals of the New York Academy of Sciences 2011, 1229(1), 76–88. [Google Scholar] [CrossRef] [PubMed]
- Habibi, N.; Bianco-Miotto, T.; Phoi, Y.Y.; Jankovic-Karasoulos, T.; Roberts, C.T.; Grieger, J.A. Maternal diet and offspring telomere length: a systematic review. Nutrition reviews 2021, 79, 148–159. [Google Scholar] [CrossRef]
- Magnano San Lio, R.; Maugeri, A.; La Rosa, M.C.; Giunta, G.; Panella, M.; Cianci, A.; Caruso, M.A.T.; Agodi, A.; Barchitta, M. Nutrients intakes and telomere length of cell-free circulating DNA from amniotic fluid: findings from the Mamma& Bambino cohort. Sci Rep. 2022, 12, 11671. [Google Scholar] [CrossRef]
- Nsereko, E.; Uwase, A.; Muvunyi, C.M.; Rulisa, S.; Ntirushwa, D.; Moreland, P.; Corwin, E.J.; Santos, N.; Lin, J.; Chen, J.L.; et al. Association between micronutrients and maternal leukocyte telomere length in early pregnancy in Rwanda. BMC Pregnancy and Childbirth 2020, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Valera-Gran, D.; Prieto-Botella, D.; Hurtado-Pomares, M.; Baladia, E.; Petermann-Rocha, F.; Sánchez-Pérez, A.; Navarrete-Muñoz, E.M. The Impact of Foods, Nutrients, or Dietary Patterns on Telomere Length in Childhood and Adolescence: A Systematic Review. Nutrients 2020, 14, 3885. [Google Scholar] [CrossRef] [PubMed]
- Paul, L. Diet, nutrition and telomere length. The Journal of nutritional biochemistry 2011, 22, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.F. Nutriomes and personalised nutrition for DNA damage prevention, telomere integrity maintenance and cancer growth control. Advances in nutrition and cancer 2014, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Burton, M.A.; Godfrey, K.M.; Lillycrop, K.A. Linking nutrition to long-term health: Epigenetic mechanisms. In Early Nutrition and Long-Term Health; Woodhead Publishing, 2022; pp. 257–277. [Google Scholar] [CrossRef]
- Okuda, K.; Bardeguez, A.; Gardner, J.P.; Rodriguez, P.; Ganesh, V.; Kimura, M.; Skurnick, J.; Awad, G.; Aviv, A. Telomere length in the newborn. Pediatric research 2002, 52, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Aviv, A.; Chen, W.; Gardner, J.P.; Kimura, M.; Brimacombe, M.; Cao, X.; Srinisavan, R.S.; Berenson, G.S. Leukocyte telomere dynamics: longitudinal findings among young adults in the Bogalusa Heart Study. American journal of epidemiology 2009, 169, 323–329. [Google Scholar] [CrossRef]
- Davy, P.; Nagata, M.; Bullard, P.; Fogelson, N.S.; Allsopp, R. Fetal growth restriction is associated with accelerated telomere shortening and increased expression of cell senescence markers in the placenta. Placenta 2009, 30, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Elbers, C.C.; Garcia, M.E.; Kimura, M.; Cummings, S.R.; Nalls, M.A.; Newman, A.B.; Park, V.; Sanders, J.L.; Tranah, G.J.; Tishkoff, S.A.; et al. Comparison between southern blots and qPCR analysis of leukocyte telomere length in the health ABC study. Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences 2014, 69, 527–531. [Google Scholar] [CrossRef]
- Friedrich, U.; Schwab, M.; Griese, E.U.; Fritz, P.; Klotz, U. Telomeres in neonates: new insights in fetal hematopoiesis. Pediatric research 2021, 49, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Gielen, M.; Hageman, G.; Pachen, D.M.F.A.; Derom, C.; Vlietinck, R.; Zeegers, M.P. Placental telomere length decreases with gestational age and is influenced by parity: a study of third trimester live-born twins. Placenta 2014, 35, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Demerath, E.W.; Cameron, N.; Gillman, M.W.; Towne, B.; Siervogel, R.M. Telomeres and telomerase in the fetal origins of cardiovascular disease: a review. Human biology 2004, 76, 127. [Google Scholar] [CrossRef]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Telomere length and oxidative stress and its relation with metabolic syndrome components in the aging. Biology 2021, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Zeichner, S.L.; Palumbo, P.; Feng, Y.; Xiao, X.; Gee, D.; Sleasman, J.; Goodenow, M.; Biggar, R.; Dimitrov, D. Rapid telomere shortening in children. Blood. The Journal of the American Society of Hematology 1999, 93, 2824–2830. [Google Scholar] [CrossRef]
- Frenck Jr, R.W.; Blackburn, E.H.; Shannon, K.M. The rate of telomere sequence loss in human leukocytes varies with age. Proceedings of the National Academy of Sciences 1998, 95, 5607–5610. [Google Scholar] [CrossRef] [PubMed]
- Daniali, L.; Benetos, A.; Susser, E.; Kark, J.D.; Labat, C.; Kimura, M.; Desai, K.K.; Granick, M.; Aviv, A. Telomeres shorten at equivalent rates in somatic tissues of adults. Nature communications 2013, 4, 1597. [Google Scholar] [CrossRef]
- Akkad, A.; Hastings, R.; Konje, J.C.; Bell, S.C.; Thurston, H.; Williams, B. Telomere length in small-for-gestational-age babies. BJOG: An International Journal of Obstetrics & Gynaecology 2006, 113, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, S.; Verhulst, S.; Guirguis, G.; Kark, J.D.; Labat, C.; Roche, N.E.; Martimucci, K.; Patel, K.; Heller, D.S.; Kimura, M.; et al. Telomere length dynamics in early life: the blood-and-muscle model. The FASEB journal 2018, 32, 529. [Google Scholar] [CrossRef]
- Factor-Litvak, P.; Susser, E.; Kezios, K.; McKeague, I.; Kark, J.D.; Hoffman, M.; Kimura, M.; Wapner, R.; Aviv, A. Leukocyte Telomere Length in Newborns: Implications for the Role of Telomeres in Human Disease. Pediatrics 2016, 137. [Google Scholar] [CrossRef] [PubMed]
- Phillippe, M. Telomeres, oxidative stress, and timing for spontaneous term and preterm labor. American Journal of Obstetrics and Gynecology 2022, 227, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Pinilla, F. Brain foods: the effects of nutrients on brain function. Nature reviews neuroscience 2008, 9, 568–578. [Google Scholar] [CrossRef]
- Firth, J.; Gangwisch, J.E.; Borsini, A.; Wootton, R.E.; Mayer, E.A. Food and mood: how do diet and nutrition affect mental wellbeing? BMJ 2020, 369, m2382. [Google Scholar] [CrossRef]
- Rao, T.S.; Asha, M.R.; Ramesh, B.N.; Rao, K.J. Understanding nutrition, depression and mental illnesses. Indian journal of psychiatry 2008, 50, 77. [Google Scholar] [CrossRef]
- Malouff, J.M.; Schutte, N.S. A meta-analysis of the relationship between anxiety and telomere length. Anxiety, Stress, & Coping 2017, 30, 264–272. [Google Scholar] [CrossRef]
- Mathur, M.B.; Epel, E.; Kind, S.; Desai, M.; Parks, C.G.; Sandler, D.P.; Khazeni, N. Perceived stress and telomere length: A systematic review, meta-analysis, and methodologic considerations for advancing the field. Brain, behavior, and immunity 2016, 54, 158–169. [Google Scholar] [CrossRef]
- Needham, B.L.; Mezuk, B.; Bareis, N.; Lin, J.; Blackburn, E.H.; Epel, E.S. Depression, anxiety and telomere length in young adults: evidence from the National Health and Nutrition Examination Survey. Molecular psychiatry 2015, 20, 520–528. [Google Scholar] [CrossRef]
- Martinez, D.; Lavebratt, C.; Millischer, V.; de Jesus, R.; de Paula, V.; Pires, T.; Michelon, L.; Camilo, C.; Esteban, N.; Pereira, A.; Schalling, M.; Vallada, H. Shorter telomere length and suicidal ideation in familial bipolar disorder. Plos one 2022, 17, e0275999. [Google Scholar] [CrossRef]
- Colpo, G.D.; Leffa, D.D.; Koehler, C.A.; Kapczinski, F.; Quevedo, J.; Carvalho, A.F. Is bipolar disorder associated with accelerating aging? A meta-analysis of telomere length studies. Journal of affective disorders 2015, 186, 241–248. [Google Scholar] [CrossRef]
- Ayora, M.; Fraguas, D.; Abregu-Crespo, R.; Recio, S.; Blasco, M.A.; Moises, A.; Derevyanko, A.; Arango, C.; Diaz-Caneja, C.M. Leukocyte telomere length in patients with schizophrenia and related disorders: a meta-analysis of case-control studies. Molecular Psychiatry 2022, 27, 2968–2975. [Google Scholar] [CrossRef]
- Russo, P.; Prinzi, G.; Proietti, S.; Lamonaca, P.; Frustaci, A.; Boccia, S.; Amore, R.; Lorenzi, M.; Onder, G.; Marzetti, E.; Bonassi, S. Shorter telomere length in schizophrenia: evidence from a real-world population and meta-analysis of most recent literature. Schizophrenia research 2018, 202, 37–45. [Google Scholar] [CrossRef]
- Benetos, A.; Kark, J.D.; Susser, E.; Kimura, M.; Sinnreich, R.; Chen, W.; Steenstrup, T.; Christensen, K.; Herbig, U.; von Bornemann Hjelmborg, J.; Aviv, A. Tracking and fixed ranking of leukocyte telomere length across the adult life course. Aging cell 2013, 12, 615–621. [Google Scholar] [CrossRef]
- Entringer, S.; Epel, E.S.; Lin, J.; Buss, C.; Blackburn, E.H.; Simhan, H.N.; Wadhwa, P.D. Prenatal programming of newborn and infant telomere length. European Journal of psychotraumatology 2012, 3. [Google Scholar] [CrossRef]
- Rewak, M.; Buka, S.; Prescott, J.; de Vivo, I.; Loucks, E.B.; Kawachi, I.; Non, A.L.; Kubzansky, L.D. Race-related health disparities and biological aging: does rate of telomere shortening differ across blacks and whites? Biological psychology 2014, 99, 92–99. [Google Scholar] [CrossRef]
Figure 1.
PRISMA flow diagram 2009 [71]. Methodology used for studies selection.
Figure 1.
PRISMA flow diagram 2009 [71]. Methodology used for studies selection.
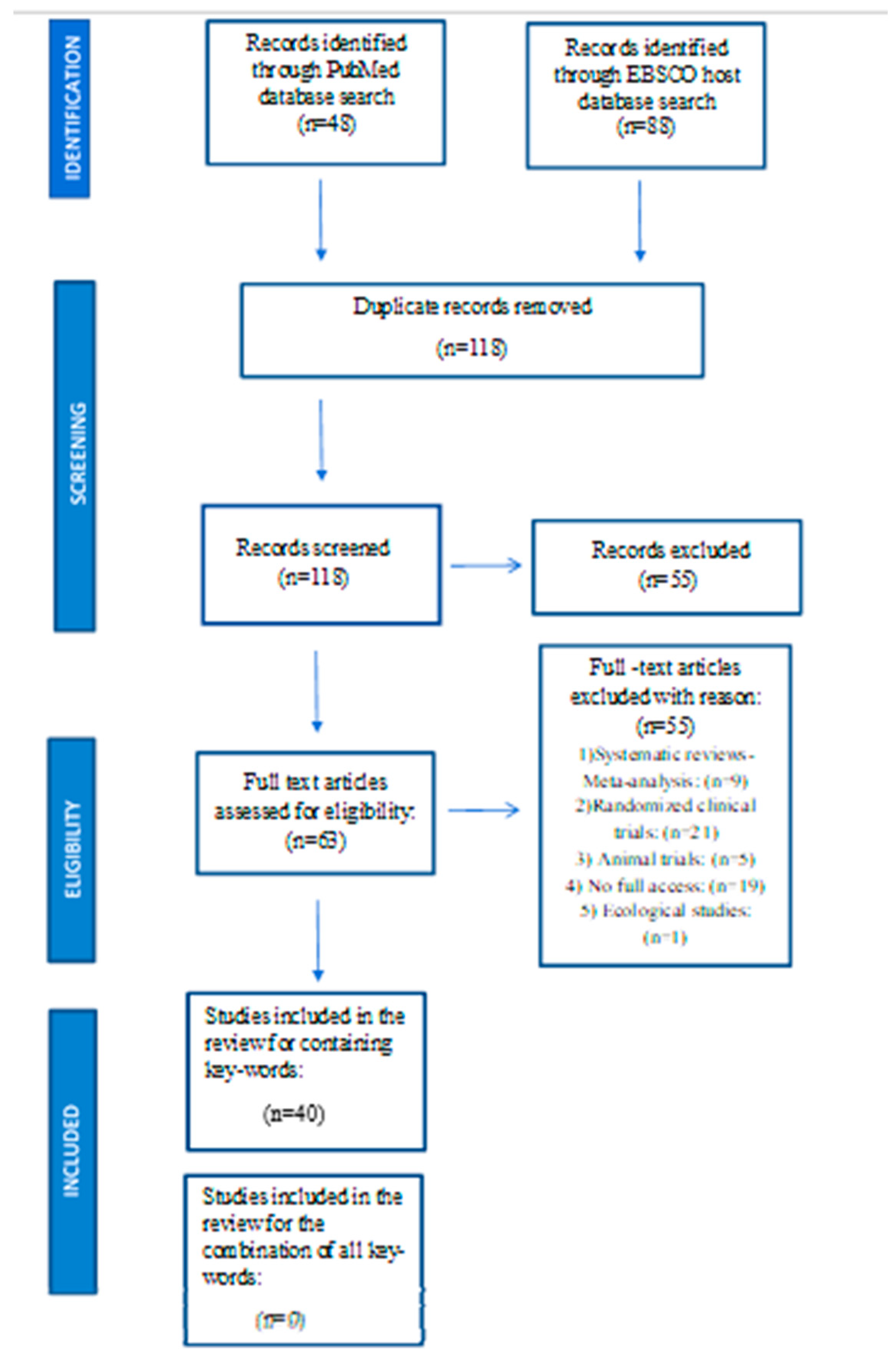

Table 1.
Studies associating maternal nutrition and preterm birth (n=21).
| Research |
Type of study | Country | Population/ Subject characteristics |
Outcome |
|---|---|---|---|---|
| 1. Xiong et al, 2021 [92] | Cohort | China | 3,382 mother- newborn second-trimester pairs and 3,478 mother- third-trimester newborn pairs Absence of pathology |
3rd trimester MTP level, inversely associated with PTB risk and positively associated with gestational duration. The effects of the 3rd trimester MTP level on PTB risk and gestational duration, were stronger in women carrying female offspring than those carrying male. |
| 2. Kriss et al, 2018 [76] | Cohort | Mexico | 965 pregnant women 18-22 weeks gestation - follow up 2 years post-birth 18 -39 years old, Absence of pathology |
No overall association between prenatal yogurt consumption and PTB. In non-overweight women, higher prenatal yogurt consumption was associated with reduced PTB risk. |
| 3. Tith et al, 2019 [75] | Cross-sectional | Canada | 78.019 births of Arabic speaking women 3 trimesters of pregnancy during Ramadan period Absence of pathology |
Ramadan fasting during the 2nd trimester of pregnancy was associated with increased risk of very PTB (28-31 weeks gestation). |
| 4. Johnston et al, 2016 [83] | Cohort | USA | 62.443 pregnant women ≥18 weeks gestation Singleton pregnancy Absence of pathology |
Any multivitamin use in the last 3 months of pregnancy is associated with a significant reduction in PTB among non-Hispanic and African women. |
| 5. Chia et al, 2016 [79] | Cohort | Singapore | 923 pregnant women Absence of pathology |
The VFR pattern is associated with a lower incidence of PTB. The VFR pattern was also associated with a higher birth weight, higher ponderal index, and increased risk of LGA deliveries. |
| 6. Li et al, 2014 [81] | Cohort | China | 207 936 singleton live births Delivered at GAs of 20–42 weeks Absence of pathology |
Daily consumption of 400 μg folic acid alone during the periconceptional period is related to a reduced risk of spontaneous PTB. These reduced risks were greater for early-age spontaneous PTBs. |
| 7. Chen et al, 2015 [89] | Cohort | Singapore | 999 pregnant and <14 weeks gestation & post-partum women, aged 18-50 years old. Only parents of the same ethnicity Absence of pathology |
Higher maternal folate concentrations at approximately the start of the 3RD trimester significantly associated with longer duration of gestation and lower risk of PTB. Little or no correlation between ↑folic acid and SGA. Little or no correlation between ↑B6 &B12 vitamins and PTB or SGA. |
| 8. Perveen & Soomro, 2016 [88] | Cohort | Pakistan | 234 pregnant women aged 20-35 and at 35-42 weeks gestation Singleton pregnancy Absence of pathology |
Fe deficiency anemia positively associated with PTB , low newborn birth weight, fetal mortality and ↓ Apgar score at the 1st and 5th minutes of birth. |
| 9. Lu et al, 2018 [77] | Cohort | China | 7352 pregnant women <20 weeks gestation Singleton pregnancy. Absence of pathology |
Women in the high consumption of ‘Milk’ group, had greater odds of PTB, spontaneous PTB and late PTB, than those in the ‘Vegetables’ group. Compared with women in the ‘Vegetables’ group, those in the ‘Cereals, eggs, and Cantonese soups’ and ‘Fruits, nuts, and Cantonese desserts’ groups had increased odds of late PTB. Maternal pregnancy diet with frequent consumption of milk and less frequent consumption of vegetables is found to be associated with increased odds of PTB among Chinese women. |
| 10. Ren et al, 2022 [85] | Case-control study | China | 509 pregnant women: Controls: 415 pregnant women with birth at term and Cases:82 pregnant women with PTB Singleton pregnancy Absence of pathology |
Negative correlation between levels of NTMs,and PTB Especially Fe and Zn may protect against PTB occurrence. Potential protective effect of Hg. |
| 11. Parisi et al, 2020 [73] | Cohort | Italy | 112 pregnant women (11+-13+6 weeks of gestation) Absence of pathology Singleton pregnancy Natural conception Healthy neonates |
Positive association between maternal nutritional score and GA at birth. Significant increase in the risk of PTB associated with low nutritional scores. The single food items of the score calculation were not associated with both early placental markers and complex pregnancy outcomes. |
| 12. Teixeira et al, 2023 [78] | Cross-sectional | Portugal | 60 Portuguese women with a diagnosis of high-risk pregnancy Birth before 33 weeks GA |
In very PTB pregnancy-induced hypertension was associated with increased consumption of pastry products, fast food, bread, pasta, rice, and potatoes. Only bread consumption had a weak but statistically significant association with pregnancy-induced hypertension in a multivariate analysis. |
| 13. Hillesund et al, 2018 [74] | Cohort | Norway | 591 nulliparous pregnant women 18 years or older 20 weeks pregnant or less at inclusion Singleton pregnancy Absence of pathology, disabilities and substance abuse |
A higher diet score either pre-pregnancy or in early pregnancy was protectively associated with excessive GWG and PTB risk. The protective association with high birthweight was confined to pre-pregnancy diet and with preeclampsia to early pregnancy diet. No association between pre-pregnancy diet score and preeclampsia. |
| 14. Chiudzu et al, 2020 [87] | Nested case-control | Malawi | 181 pregnant women: 90/181 (49.7%) term and 91/181 (50.3%) PTBs. Singleton pregnancy Presenting in spontaneous labor with intact membranes Absence of pathology |
Copper maternal serum concentrations were above the upper normal limit in both term and PTB groups. PTB was associated with higher maternal serum concentrations of copper and zinc. |
| 15. Olapeju et al, 2019 [84] | Case - control | USA | 7675 mother–infant dyads Cases: mother–infant dyads, singleton, live, low-birth-weight (<2500 g) or preterm infants (<37 weeks of gestation) regardless of birth weight Controls: mother–infant dyads with singleton, live, term infants with birth weight of ≥2500 g or more |
Multivitamin supplement intake of at least 3 times/week throughout pregnancy was significantly associated with a reduction in the odds of PTB. Use during the third trimester associated with a greater reduction in PTB odds than use in the first trimester. No significant association between preconceptional supplement intake and PTB. Higher plasma folate levels were associated with lower risk of PTB. |
| 16. Christoph et al, 2020 [90] | Cross-sectional | Switzerland | 1382 pregnant women Supplemented with 600 IU/d orally throughout the pregnancy, and at least 1000 IU/d in cases of deficiency |
No association between the 25(OH)D serum level and, PTB, preeclampsia, postdate pregnancy, miscarriage, intrauterine growth restriction, bacterial vaginosis, mode of delivery, or neonatal birth weight and length. The lower the vitamin D maternal level, the higher the GA at birth. |
| 17. Miele et al, 2021 [93] | Nested case-control | Brazil | 1165 nulliparous pregnant women between 19 and 21 weeks of gestation Singleton pregnancies Absence of pathology Absence of medication or supplementation |
The odds of adverse outcomes were higher in non-white (p < 0.05) obese women and with high protein consumption. The anthropometric classification of obesity had a greater impact on PE and GDM, in contrast to PTB and SGA. |
| 18. Irwinda et al, 2019 [86] | Cross sectional | Indonesia | 51 pregnant women undergoing birth (term group: 25, PTB group: 26) Singleton pregnancy Absence of pathology |
PTB associated with lower concentrations of micronutrients in maternal serum. PTB associated with higher concentration of heavy metals such as Hg and Pb. Compared with PTB group, term birth group had higher maternal serum concentration of AtRA serum. Compared with PTB group, term birth group had higher placental concentration of Mn, Fe, Cu, Zn, Se, AtRA, 25(OH)D, and lower placental concentration of mercury and Pb. Compared with PTB group, term birth group had higher concentration of Cu and AtRA in cord blood. |
| 19. Wu et al, 2021 [82] | Cohort | China | 201,477 pregnant women aged 18–49 Singleton livebirth |
Periconceptional supplementation with FA was associated with a lower risk of PTB. Women who started taking FA at least 3 months before their last menstrual period were more likely to reduce the risk of PTB. |
| 20. Tahsin, et al, 2023 [91] | Nested case–control | Bangladesh | 930 pregnant women at 8–19 weeks of gestation 262 PTB and 668 term births |
Vitamin D deficiency is common in Bangladeshi pregnant women and is associated with an increased risk of PTB. |
| 21. Martin et al, 2015 [80] | Cohort | USA | 3143 pregnancies at 26–29 week of gestation | Greater adherence to a healthy dietary pattern, such as the DASH diet, reduced the odds of PTB. Greater adherence to a dietary pattern of poorer quality, such as factors 2 (i.e. high intakes of beans, corn, French fries, hamburgers or cheeseburgers, white potatoes, fried chicken, spaghetti dishes, cheese dishes, cornbread or hushpuppies, processed meats, biscuits, and ice cream) and factors 3 (i.e. high consumption of collard greens, coleslaw or cabbage, red processed meats, fried chicken, fried fish, cornbread or hushpuppies, eggs or egg biscuits, gravy, whole milk, and vitamin C-rich drinks), increased the odds of PTB. |
[n]: Reference number. PTB: Preterm birth, SPTB: Spontaneous Preterm Birth, SGA: Small for gestational age, LGA: Large for gestational age, KTR: kynurenine to tryptophan ratio, PE: preeclampsia, MTP: Maternal plasma total protein, VFR: vegetables/ fruits/rice, SfN: seafood & noodles, PCP: processed meat, cheese, pasta, EDMs: Endocrine disrupting metal(loid)s, Mn: manganese, Cu: copper, Se: selenium, Pb, lead, Hg: Mercury, Se: Selenium, Pb: Lead, NTMs: Nutritional trace metal(loid)s, Zn: Zinc, Fe: Iron (Fe), FA: Fatty acids, Cu: Copper, GWG: Gestational weight gain, UAC: upper arm circumference, PE: preeclampsia and GDM: gestational diabetes mellitus.
Table 2.
Studies associating maternal nutrition with placental and newborn telomeres (n=13).
| Research |
Type of study | Country | Population/ Subject characteristic |
Outcome |
|---|---|---|---|---|
| 1.Vahter et al, 2020 [97] | Cohort | Argentina | 99 Pregnant women in 1st trimester, Singleton pregnancy |
Maternal BMI, BFP, and vitamin B-12 were inversely associated. 25(OH)D3 was positively associated, with placental TL. No association between the above factors was observed with cord blood TL. |
| 2. Herlin et al, 2019 [101] | Cohort | Argentina | 169 pregnant women in 1st trimester, Singleton pregnancy |
More associations with relative TL in maternal blood leukocytes during pregnancy (inverse associations with boron and antimony, positive association with lithium), than in the placenta (positive with arsenic) and cord blood (inverse with lead). Nutritional antioxidants (Zn, Se, folate, Vit D3) did not affect the associations. |
| 3. Daneels et al, 2021 [98] | Cohort | Belgium | 108 pregnant women in 1st trimester or women who are trying to conceive Absence of pathology Singleton pregnancy |
Positive association of maternal vitamin D concentration (diet and/or supplement) in the 1st trimester with TL of the newborn at birth. |
| 4. Kim et al, 2018 [47] | Cross-sectional | Seoul | 106 mother-newborn dyads. Pregnant women in 3rd trimester Absence of maternal and fetal pathology |
Positive correlation between the concentration of vitamin D of the pregnant woman and the neonatal leukocyte TL (β = .33, p < .01). |
| 5.Entringer et al, 2015 [46] | Cohort | USA | 119 mother-newborn dyads enrolled at 9.5 weeks gestation Absence of pathology or fetal abnormality Use of folic acid supplements |
Maternal total folate concentration in early pregnancy was significantly and positively associated with newborn cord blood TL. |
| 6. Yeates et al, 2017 [95] | Cohort | Seychelles | 229 mothers enrolled at 28 week gestation and followed through delivery Their children (at 5 years of age) |
No clear associations of both prenatal or postnatal PUFA status and methylmercury exposure with child TL. Higher prenatal n–6:n–3 PUFA ratio was associated with longer TL in mothers. |
| 7. Liu et al, 2022 [94] | Cohort | China | 274 mother newborn dyads. Pregnant women enrolled during 3rd trimester. Absence of pathology |
Positive association of the concentrations of DHA and total n-3 PUFAs in maternal erythrocytes with DNA methylation of the TERT promoter in the cord blood instead of the placenta. Maternal PUFAs closely correlated to infant TL and TERT promoter methylation, differently affected by maternal n-3 PUFAs between the cord blood and the placenta. High levels of maternal n-3 PUFAs during pregnancy maintain offspring TL. |
| 8. Chen et al, 2022 [65] | Cohort | Singapore | 950 mother-offspring dyads In their first trimester of pregnancy Participants with homogeneous parental ethnic backgrounds. |
TL in female newborn more susceptible to variation from maternal TL, mental health, and vitamin B12 levels. Variation in newborn male TL was more explained by their parental age, maternal education, plasma fasting glucose, DGLA%, and IGFBP3 levels. Overall, maternal TL strongly associated with antenatal factors, especially metabolic health and nutrient status. |
| 9. Magnano San Lio et al, 2022 [102] | Cohort | Italy | 174 pregnant women, at 4th–20th gestational week Singleton pregnancyAbsence of pathology and pregnancy complications Planned follow-up of their children at delivery and up to 2 years of age |
Magnesium deficiency negatively associated with maternal RTL after adjusting for the same covariates Positive association between maternal intake of magnesium and TL of cfDNA from amniotic fluid, while results on other micronutrients (i.e., vitamin B1 and Fe) were marginally significant. |
| 10. Salihu, et al, 2018 [96] | Cohort | USA | 62 women upon admission for delivery Singleton pregnancyNo indication of congenital malformations |
Shortened TL among fetuses exposed to maternal high fat consumption during pregnancy, after accounting for the effects of potential covariates. |
| 11. Fan et al, 2022 [99] | Cohort | China | 746 mother-newborn pairs < 16 weeks of pregnancy Singleton gestation |
Possible association between maternal FA supplementation during pregnancy with longer newborn TL was suggested. |
| 12. Louis-Jacques, et al, 2016 [100] | Cohort | USA | 96 maternal-fetal dyads Singleton gestation No indication of congenital malformations |
Positive association between umbilical cord RBC folate and fetal TL at birth. |
| 13. Myers et al, 2021 [103] | Cohort | USA | 96 maternal–fetal dyads Singleton gestation No indication of congenital malformations |
Positive association between maternal vitamin C intake and fetal TL. |
[n]: Reference number. STI: Sexually Transmitted Infection, sTfR: soluble Transferrin Receptor, RBP: Retinol Binding Protein, PUFA: Polyunsaturated fatty acids, cfDNA: cell-free DNA, IGFBP3: Insulin-like growth factor binding protein 3, CRP:C-reactive protein, Fe: Iron, Zn: Zinc, Se: Selenium, DGLA: Dihomo-γ-linolenic acid, AGP: Ambulatory glucose profile. PTB: Preterm birth, BFM: body fat mass, BFP: body fat percentage, RTL: Relative telomere length, TL: Telomere length, GA: Gestational age, RBC: Red blood cell. TERT: Telomerase reverse transcriptase, DHA: Docosahexaenoic acid.
Table 3.
Studies associating preterm birth with placental and newborn telomeres (n=6).
| Research |
Type of study | Country | Population/ Subject characteristics |
Outcome |
|---|---|---|---|---|
| 1. Vasu et al, 2017 [61] | Case-control | United Kingdom |
78 newborn (47 premature <32gestational weeks and 31 term newborns). Absence of antenatal or postnatal severe congenital malformation or unlikely to survive. |
RTL significantly negatively correlated with GA and birth weight in preterm infants. RTL highly variable in newborn infants. Preterm infants at term equivalent age, have significantly longer TLs than term born infants. Positive correlation between maternal age and T/S ratio. Longitudinal assessment in preterm infants with TL measurements available at birth and term age (n = 5) suggests that TL attrition rate is negatively correlated with increasing GA. RTL was significantly shortest in the term born control group compared with both PTB groups and longest in the PTB at birth group. In addition, TL was not significantly different between preterm infants sampled at birth and those sampled at term equivalent age. |
| 2. Hadchouel et al, 2015 [70] | Cohort | France | 274 adults Group 1:236 adults born prematurely Group 2:38 adults born full-term |
No apparent association with perinatal events. Positive association between TL and abnormal lung airflow in the population born prematurely. |
| 3. Farladansky-Gershnabel, et al, 2023 [106] | Cross sectional | Israel | 19 women with spontaneous term pregnancies and 11 with SPTB Absence of preterm premature rupture of membranes or induced preterm labor, Absence of pregnancy pathology or pregnancy complications, Nulliparous, Absence of fetal malformations |
Maternal blood and placental samples from SPTB had shorter telomeres and increased Gal-3 expression. compared with spontaneous term pregnancies group. |
| 4. Saroyo et al, 2021 [104] | Cross - sectional | Indonesia | 67 placentas (34 placentas from preterm and 33 placentas from term birth were included). Singleton pregnancy, Absence of pathology and pregnancy complications |
Similar TL due to early telomere shortening in PTB that mimics the term placenta. 8-OHdG and HMGB1, do not correlate with placental telomere ratio. HMGB1 from the placenta of both PTB and term labor has no significant difference. Equal telomere T/S ratio of the placenta from PTB and term labor. |
| 5 Colatto et al, 2020 [105] | Cross sectional | Brazil | 80 Singleton pregnancies Foetal membrane samples collected from (1) pregnant women with pPROM, (2) PTB with intact membranes, (3) term labor and (4) term not in labor (TNL). Absence of pathology and pregnancy complications |
Foetal membranes from the term group showed TL reduction compared with those from the other groups. Telomerase activity did not change in foetal membranes irrespective of pregnancy outcome. Telomere shortening in foetal membranes is suggestive of senescence associated with triggering of labor at term. |
| 6. Tarik et al, 2019 [107] | Cohort | India | 1309 offspring, Mean maternal age 39.08y (±3.29), BMI gain at 2, 11, and 29 years |
GA positively associated with offspring RTL No significant association of offspring RTL with body size at birth including birthweight, birth length, and birth BMI. Conditional BMI gain at 2 and 11 years not associated with RTL. BMI gain at 29 year was negatively associated with RTL. Born SGA was not associated with RTL in adulthood. Increased LTL attrition was observed in those born before 37 weeks of GA, as well as in those who gained weight as adults. |
[n]: Reference number. LTL: Leucocyte telomere length, RTL: Relative telomere length, pPROM: preterm premature rupture of membranes, TL: Telomere length, GA: gestational age, PTB: PTB, BMI: body mass index, T/S ratio: telomere/single copy gene ratio, OR: Odds ratio, GA: Gestational age, SGA: Small for gestational age, EVT: extravillous trophoblast, SPTB: spontaneous preterm birth, Gal-3: Galanine, TNL: term not in labor, pPROM: preterm premature rupture of membranes, PTL spontaneous preterm labor with intact membranes, BMI: Body mass index, HMGB1: High mobility group box 1, 8-OHdG: 8-hydroxy-2'-deoxyguanosine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



