
Submitted:
09 November 2023
Posted:
10 November 2023
You are already at the latest version
Abstract
Objective: To investigate whether impaired plantar cutaneous vibration perception contributes to axial motor symptoms in Parkinson’s disease (PD) and whether anti-parkinsonian medication and subthalamic nucleus deep brain stimulation (STN-DBS), show different effects. Methods: Three groups were tested: PD patients on medication (PD-MED), PD patients on medication and additional STN-DBS (PD-MED–DBS), and healthy subjects (HS) as reference. Motor performance was analyzed using a pressure distribution platform. Plantar cutaneous vibration perception thresholds (VPT) were investigated using a customized vibration exciter at 30 Hz. Results: Motor performance of PD-MED and PD-MED–DBS was characterized by greater postural sway, smaller limits of stability ranges, and slower gait, due to shorter strides, fewer steps per minute, and broader stride widths compared to HS. Comparing patient groups, PD-MED–DBS showed better overall motor performance than PD-MED, particularly for the functional limits of stability and gait. VPTs were significantly higher for PD-MED compared to HS, which suggests impaired plantar cutaneous vibration perception in PD. However, PD-MED–DBS showed less impaired cutaneous vibration perception than PD-MED. Conclusion: PD patients suffer from poor motor performance compared to healthy subjects. Anti-parkinsonian medication in conjunction with STN-DBS seems to be superior for normalizing axial motor symptoms compared to medication alone. Plantar cutaneous vibration perception is impaired in PD patients, whereas anti-parkinsonian medication together with STN-DBS is superior for normalizing tactile cutaneous perception compared to medication alone. Consequently, based on our results and the findings of the literature impaired plantar cutaneous vibration perception might contribute to axial motor symptoms in PD.
Keywords:
Parkinson’s disease
; axial motor symptoms
; postural instability & gait difficulties
; functional limits of stability
; non-motor sensory symptoms
; sensorimotor integration
; somatosensory system
; plantar cutaneous vibration perception thresholds
; deep brain stimulation
; subthalamic nucleus
1. Introduction
Parkinson’s Disease (PD) is a widespread age-related and progressive neurodegenerative disorder, the primary cause of which is associated with the depletion of dopaminergic neurons within the substantia nigra pars compacta [1,2,3,4,5]. This lack of sufficient dopamine causes a hypo-dopaminergic state within the basal ganglia, leading to imbalanced excitatory and inhibitory communication within the nigro-striatal and thalami-cortical circuits, and therefore disrupts the network of brain structures that are responsible for selecting, planning, adapting, and executing motor functions [5,6,7,8]. Consequently, after losing a dramatic portion of about 80 % of dopamine-producing cells, clinical motor symptoms, including tremor, rigidity, bradykinesia, postural instability, and gait difficulties become noticeable [9,10,11]. Although treatments, such as dopamine replacement therapy or deep brain stimulation (DBS) of the subthalamic nucleus (STN) can relieve axial motor symptoms, their effects on postural and gait control are still controversial [8,12,13,14,15].
As those motor symptoms can be very prominent and cause a significant loss in patient quality of life, PD can furthermore be accompanied by numerous non-motor sensory symptoms [16,17,18,19]. Among those non-motor sensory symptoms, defective functionality of sensory systems, such as the visual [20,21,22,23], the vestibular [8,24,25,26,27,28], and the somatosensory systems [29,30,31,32,33,34,35,36,37,38], has been reported in PD patients. Since those sensory systems provide valuable information for controlling movements, impaired sensory system functionality might therefore contribute to PD patients’ motor symptoms. This might especially be true, as those sensory systems gather information about the surrounding environment, the position of the body in space, and the relative position of each joint with respect to other parts of the body, as well as information about the condition of the ground on which the person is standing or walking [24,39,40]. Given that only the soles of the feet are in direct contact with the ground during standing or walking, particularly afferent information from plantar cutaneous mechanoreceptors seems to be crucial for motor control. Plantar cutaneous mechanoreceptors gather information about the pressure distribution and loading shifts underneath the foot during movements, and therefore are involved in adapting muscle contraction tone and contraction patterns [41,42,43,44,45,46,47,48,49,50,51,52]. The sensorimotor integration of plantar mechanoreceptors has already been investigated in several studies with individuals without neurological diseases. Those studies have shown that decreased plantar cutaneous sensation achieved by anesthesia of the foot sole led to impaired control of static and dynamic balance abilities, as well as gait performance [42,46,49,50,51,52,53,54,55,56]. Conversely, several studies have demonstrated that sensory stimulation of the foot sole using various types of shoe insoles improved balance and gait performance in PD patients [41,43,44,45]. Accordingly, those findings suggest that PD patients might suffer from impaired plantar tactile cutaneous perception, while mechanical stimulation facilitates the sensory signal and therefore enhances sensorimotor integration.
This makes sense from a pathophysiological point of view, because the mechanisms of sensory symptoms in PD include findings of widespread deposits of α-synuclein, which is a fundamental pathological protein and a major component of malicious Lewy bodies in PD [16,57,58,59,60,61,62]. Impaired plantar tactile cutaneous perception in PD patients is therefore quite conceivable, since it has been shown that α-synuclein affects numerous sensory-related structures of the central nervous system, as well as the peripheral nervous system, including the skin [57,63,64,65,66,67,68]. This is supported by the study from Nolano et al., who investigated skin biopsies and found peripheral denervation in PD patients. More specifically, the authors reported demyelination of epidermal nerve fibers of the glabrous and the hairy skin, as well as a significantly reduced number of mechanoreceptors in PD patients compared to healthy subjects. In particular, Meissner corpuscles, which detect mechanical vibration stimuli, may be affected, whereas the loss of those mechanoreceptors correlated with the disease severity of the patients. The authors concluded that the peripheral deafferentation could play a major role in the pathogenesis of sensory dysfunction in PD [64]. Besides results showing impairments of the peripheral nervous system, there is also evidence for the defective central integration and processing of afferent information at a cerebral level in PD patients. Although the basal ganglia are considered well-established, primarily motor-related structures, they might also function as an active “sensory analyzer” for higher level central somatosensory processing [69,70,71,72,73]. This is plausible, since the basal ganglia have projections into the thalamus and the cortex and do not only receive input from motor areas, but also from cortical somatosensory areas, including the primary and the secondary somatosensory cortices [69,70,73,74]. Using 3D positron emission tomography, Boecker et al., showed that basal ganglia dysfunction in PD is characterized by abnormal sensory processing even for tasks devoid of any motor component [73]. Consequently, impaired tactile cutaneous perception in PD might be driven by both denervation of peripheral epidermal nerve fibers and mechanoreceptors and the defective central integration and processing of afferent information due to the diseased basal ganglia network [38,64,73,75,76,77,78,79,80,81,82,83,84,85,86].
Despite this clinical and experimental evidence of cutaneous denervation and impaired sensory processing in PD, investigations about tactile cutaneous perception are rare, especially for the foot sole, and the few existing studies show rather discordant results. While some studies found impaired plantar tactile cutaneous perception in PD patients [64,87,88], others failed to find PD affecting cutaneous thresholds for mechanical stimuli of the foot [48,89]. Lack of studies and contradicting results are also prominent for investigations about the effect of various therapies, such as anti-parkinsonian medication and STN-DBS for patients’ tactile cutaneous perception of the foot. According to the literature, anti-parkinsonian medication seems to have generally ´minor to no effects [48,64,87,89]. On the other hand, STN-DBS generally seems to be more promising for treating tactile cutaneous perception in PD [32,33,34,36,37,81,90,91,92]. However, the existing studies only included investigations of the upper body, such as the torso, arms, hands, or fingers, and to the best of our knowledge, no other studies have investigated the effect of STN-DBS on tactile cutaneous perception in PD patients’ feet.
Specifically investigating plantar tactile cutaneous perception in PD patients could be beneficial for several reasons. It has been reported that sensory symptoms may occur prior to the presence of motor symptoms, so they could be used for early diagnosis of PD [16,93,94,95]. Moreover, this may help to distinguish between PD and various other neurodegenerative diseases, such as multiple system atrophy, which has been demonstrated with a high sensitivity and specificity of approx. 80 % [87]. Consequently, testing plantar cutaneous sensation could serve as a potential surrogate marker. It may also add to our understanding of how PD affects the peripheral nervous system, as well as give insight into sensorimotor integration and potentially help to better understand the cause of axial motor symptoms in PD. Thinking ahead, using plantar tactile cutaneous perception measurements could help to develop and optimize low budget therapy devices, such as textured insoles, to stimulate plantar cutaneous mechanoreceptors and consequently enhance motor performance in PD patients [41,43,44,45]. As this is the first study investigating the effects of STN-DBS on plantar tactile cutaneous perception in PD patients, it therefore might provide valuable information for better understanding the neurophysiological processing of afferent sensory input, as well as for counseling patients on appropriate treatments, and assisting clinicians in tailoring and optimizing therapy strategies.
Therefore, we pursued two objectives with this study.
First, we investigated and characterized axial motor symptoms in PD patients, such as postural instability and gait difficulties. We analyzed whether patients treated with anti-parkinsonian medication in conjunction with STN-DBS have superior motor control compared to patients treated with medication alone. We also tested healthy elderly subjects as a reference. We hypothesized that patients’ motor performance is worse compared to healthy subjects, and that medication in conjunction with additional STN-DBS would be more advantageous in normalizing patients’ abnormal motor control compared to treatment with medication alone.
Second, we investigated somatosensory functionality by analyzing plantar cutaneous vibration perception thresholds (VPT) within the same study groups. Based on the pathophysiological mechanisms of PD and previous study findings, we hypothesized that patients’ plantar cutaneous vibration perception would be impaired compared to healthy subjects. Furthermore, we assumed that medication in conjunction with STN-DBS would show superior effects on normalizing patients’ impaired plantar cutaneous vibration perception compared to treatment with medication alone.
2. Methods
2.1. Subjects
Three different study groups were investigated: Patients with Parkinson’s disease (PD-MED), patients with Parkinson’s disease who had undergone previous deep brain stimulation surgery (PD-MED–DBS), and healthy subjects. Both patient groups comprised subjects who were at least 50 years old and who were diagnosed with idiopathic Parkinson's disease with a disease severity level between 2 and 3 on the Hoehn and Yahr scale, following the diagnostic criteria of the Movement Disorders Society [96]. Subjects assigned to group PD-MED–DBS had to have undergone bilateral high frequency (≥ 130 Hz) deep brain stimulation surgery of the STN at least one year prior to ensure optimized DBS settings and full efficiency [15,97]. The cut-off for the duration of DBS since surgery was defined as 5 years, since it has been shown that stimulation efficacy decreases gradually over time [98,99,100]. All enrolled DBS patients were positive responders to the surgery. Subjects of both patient groups were tested in the medication “on” state using regular anti-parkinsonian medication, including levodopa. Patients in the study group PD-MED–DBS were additionally in the DBS “on” stimulation state. The exclusion criteria for both patient groups comprised secondary pathologies affecting the motor and somatosensory systems, for instance atypical parkinsonism, severe camptocormia, normal pressure hydrocephalus, and diabetes mellitus with polyneuropathy. All patients had to be able to stand and walk without assistance. Patients with cognitive deficits (mini-mental state examination (MMSE) <24/30), psychiatric problems, or severe depression were excluded. All PD patients were recruited and tested during a patient consultation at Christian Oehlwein’s Neurological Outpatient Clinic for Parkinson’s Disease and Deep Brain Stimulation in Gera, Germany. Relevant clinical data from the most recent neurological examination was provided by the clinic for both patient groups (Table 1). The control group of healthy elderly subjects was tested in the laboratory of the Department of Human Locomotion (Chemnitz University of Technology, Germany). All healthy subjects were free of injuries or diseases, and took no medication that could have interfered with postural performance, cognition, or cutaneous perception.
2.2. Equipment and Testing Procedures
Prior to data acquisition, all subjects were informed about the purpose of this study and provided written informed consent. All procedures were conducted according to the recommendations of the Declaration of Helsinki and were approved by the ethics committee of the medical faculty of the University Leipzig (IRB number: 023/14-ff).
2.2.1. Motor Performance
Motor performance was quantified using a pressure distribution platform (Zebris FDM 1.5; Isny, Germany, sampling frequency 100 Hz). For subjects’ safety, the platform was integrated into a walkway so that it was level with the ground. All tests were carried out barefoot. Plantar skin temperature was monitored with an infrared thermometer (UNI-T UT302C, Augsburg, Germany), since alterations may affect tactile perception and consequently motor performance [56,101,102,103].
Quiescent Bipedal Stance
Subjects had to stand as still as possible in bipedal stance, with an upright posture, keeping the knees straightened but not locked, both arms hanging down loosely, and directing their gaze ahead. (Figure 1 A). Stance width could be chosen individually and was controlled measuring the heel-to-heel distance. Three consecutive trials lasting 20 s each were collected for each subject.
Functional Limits of Stability
To detect subjects’ limits of stability, they were instructed to stand as still as possible for approx. 10 s to define the neutral COP location. Thereafter, subjects had to lean forward three times and backward three times as far as they could, using the ankle joints as the pivot point. They were neither allowed to lift their heels or toes from the platform, nor were they allowed to bend their back or flex their knee and hip joints (Figure 1 B). When the subjects reached their maximum leaning posture, they were instructed to hold it for approx. 5 to 10 s [20,104,105,106,107]. The examiner stood nearby and carefully monitored the procedure and assisted in case of balance loss. If this happened, the trial was repeated.
Gait
Subjects were instructed to walk over the platform at least 8 times to generate sufficient data. Gait speed had to be quick but comfortable and safe, and could be chosen individually (Figure 1 C).
2.2.2. Plantar Vibration Perception
A customized vibration exciter (Tira Vib, TV51075, Schalkau, Germany) was used to quantify plantar cutaneous VPTs [47,108,109]. An oscillating brass probe (diameter 7.8 mm), protruding through a hole in an aluminum foot rest, applied sinusoidal vibration stimuli to the plantar foot (Figure 2). Vibration frequency was set at 30 Hz, a frequency which primarily innervates fast adapting (FA I) cutaneous afferents (Meissner corpuscles) [110,111], which are known to be impaired in PD [64]. Vibration frequency and amplitude were verified by an accelerometer (NXP Semiconductor, MMA2241KEG, Eindhoven, Netherlands), which was attached to the probe. Since changes in skin temperature affect tactile perception, the foot rest was equipped with a thermal element that was adjusted to 20 C° to assure comparability [101,112]. Prior to the examination, all subjects underwent an acclimatization period of approx. 10 min by placing their bare feet onto the aluminum plate, while comfortably sitting on a chair, with the hip and knee joints bent at approx. 90°. Subsequently, the first metatarsal head of the left or right foot was positioned precisely perpendicular to the probe in random order. Since alterations of the force the probe exerts against the skin affects tactile perception, the contact force was monitored throughout the measurement using a sensor (Hottinger, U9B, Darmstadt, Germany) [113,114]. A customized algorithm (written in Labview 2015, National Instruments, Austin, TX, USA), modified after Mildren et al. was implemented to automatically detect the VPT [115,116,117,118]. The algorithm started with a supra-threshold sinusoidal vibration burst (2 s duration), and patients indicated having perceived the vibration by pushing a handheld trigger. Consequently, the vibration amplitude was gradually reduced by 50 % until the subject no longer perceived the burst. Then, the average amplitude from the last perceived and the last unperceived burst was tested. The algorithm ended four bursts after the first undetected stimulus. To prevent bias from anticipation and habituation, the algorithm randomly varied the pause time between two consecutive vibration stimuli (ranging between 2-7 s). The recorded VPT was defined as the mean of the smallest perceived and largest unperceived vibration amplitude in µm. Each subject underwent one test trial and three main trials for both feet. To eliminate environmental noises, subjects were required to wear noise-canceling headphones (QuietComfort 25, Bose GmbH, Friedrichsdorf, Germany). Plantar temperatures of both feet were monitored using the same infrared thermometer used for the motor performance tests.
2.3. Data Processing
2.3.2. Motor Performance
All data were processed using a routine written in MATLAB R2022a (Math-WorksTM, Natick, MA, USA).
Quiescent Bipedal Stance
Raw COP displacement data were filtered using a recursive zero-phase-shift filter with a cut-off frequency of 35 Hz. Subsequently, the COP ranges, the COP maximum velocities (for both parameters in the anterior-posterior (AP) and medio-lateral (ML) directions), the 95 % COP confidence area, and the COP maximum velocity total were calculated. For each COP parameter, the mean over the three collected trials for each subject was calculated and used for further statistical analysis.
Functional Limits of Stability
Raw COP displacement data were filtered using a recursive zero-phase-shift filter with a cut-off frequency of 35 Hz. The COP signal was visually inspected and the neutral COP location and the three anterior and posterior maximal COP displacements were identified manually. The total COP range was calculated by adding together the anterior and the posterior COP displacements for each trial and subject. The neutral COP location, the maximal COP displacement, and the full range were normalized to the foot length of each subject [105,106,119]. Additionally, the motion times until maximal COP displacements were reached and the corresponding mean velocities for each trial were calculated (Figure 3). The mean over the three collected trials for each subject was calculated and used for further statistical analysis.
Gait
Spatial-temporal gait parameters, such as stride length, stride width, stride time, gait velocity, cadence, double support time, and COP length during the stance phase of gait were exported from the Software Zebris winFDM (version 0.1.11, Isny, Germany). The mean over all collected trials for each subject was calculated and used for further statistical analysis.
2.3.3. Plantar Cutanoues Vibration Perception
2.4. Statistical Analysis
Statistical analysis was performed using SPSS Statistics (IBM, version 29.0.0.0, Ehningen, Germany). The Shapiro-Wilk test (α = 0.05) was used to check for data distribution.
Differences between study groups were analyzed using the one-way analysis of variance for normally distributed data and the Kruskal–Wallis test for non-normally distributed data. To account for the study groups and testing conditions, the level of significance was Bonferroni-corrected individually. The effect sizes, r, were also calculated. To analyze the intra-group variability, an overall coefficient of variation for each test was calculated.
Intra-group comparisons between sex, body side, disease-dominant side, and AP vs. ML directions were performed using the t-test for paired or independent samples for normally distributed data. In the case of non-normally distributed data, the Wilcoxon–test was used.
3. Results
3.1. Demographic and Clinical Data
As shown in Table 1, there were considerably more male than female subjects in all study groups. On average, patient group PD-MED–DBS comprised younger subjects compared to both other study groups, with statistically significant differences compared to HS. Self-rated balance confidence and gait confidence were lower for both patient groups compared to the healthy subject group, HS. No differences were found for the clinical data MMSE, UPDRS III, UPDRS total, or Hoehn and Yahr ratings between PD-MED and PD-MED–DBS. On average, PD-MED–DBS suffered from Parkinson’s disease for more than twice as long as PD-MED. The disease-dominant side of the body was equally distributed for PD-MED. However, PD-MED–DBS comprised more patients with disease dominance on the right side of the body. In PD-MED, the time interval between the most recent clinical examination and the motor and vibration tests was longer and the variability was higher compared to PD-MED–DBS. The average duration of DBS since surgery was 33.1 ± 25.7 months, and patients’ self-rated satisfaction with DBS at the time of the tests was 80.1 ± 20.6 %.
3.2. Motor Performance
3.2.1. Quiescent Bipedal Stance
Inter-group Comparisons
Self-selected stance width did not differ between groups: PD-MED 13.1 cm ± 2.7 cm; PD-MED–DBS 13.7 cm ± 3.0 cm; HS 12.7 cm ± 2.0 cm. Statistically significant differences between groups were only found for spatial COP parameters (Figure 4, A-C), but not for COP velocities (Figure 4, E-F). In more detail, PD-MED and PD-MED–DBS showed significantly higher COP areas with higher COP ranges in the AP and ML directions, compared to HS. COP velocities only showed a trend towards higher values for PD-MED and PD-MED–DBS compared to HS. No statistically significant differences were found between PD-MED and PD-MED–DBS. The overall coefficient of variation showed higher variability for PD-MED (0.41 ± 0.12) and PD-MED–DBS (0.40 ± 0.11) compared to HS (0.34 ± 0.10).
Intra-group Comparisons
Within study groups, no significant differences were found between male and female subjects or between patients with left and right disease-dominant body sides. However, statistically significant intra-group differences (p<0.004) were found for each group between the AP and ML directions for COP ranges and COP max velocities. Consequently, all groups showed higher COP values in the AP direction compared to the ML direction.
3.2.3. Functional Limits of Stability
Inter-group Comparisons
The neutral COP location showed comparability between all groups: PD-MED 39.4 ± 5.8 %; PD-MED–DBS 38.0 ± 4.7 %, and HS 41.6 ± 4.3 % of the foot length measured from the heel. Various statistically significant inter-group differences were found for the COP displacement. Both PD-MED and PD-MED–DBS showed smaller COP displacements in the posterior direction and reduced total COP ranges compared to HS. Only PD-MED showed smaller values for the COP displacement in the anterior direction, which were statistically significant compared to HS. Comparing both patient groups, PD-MED–DBS revealed higher anterior COP displacements compared to PD-MED (Figure 5).
Both PD-MED and PD-MED–DBS showed longer motion times with slower velocities in the anterior and the posterior directions compared to HS (Table 2). No statistically significant differences were found comparing PD-MED and PD-MED–DBS. The overall coefficient of variation showed higher group variability for PD-MED (0.38 ± 0.16) compared to PD-MED–DBS (0.32 ± 0.14) and HS (0.30 ± 0.16).
Intra-group Comparisons
Within groups, no significant differences were found between male and female subjects or between patients with right-sided or left-sided disease-dominance. However, various statistically significant intra-group differences (p<0.004) between the anterior and the posterior directions were found for each group. Predominantly, COP displacements and COP mean motion velocities showed higher values for the anterior direction compared to the posterior direction.
3.2.2. Gait
Inter-group Comparisons
Statistically significant differences for gait parameters were mainly found between both patient groups and the group of healthy subjects. Patients in PD-MED and PD-MED–DBS walked significantly slower with less stride length and longer stride and double support times compared to HS. PD-MED took significantly fewer steps per minute with smaller COP lengths compared to HS. Both patient groups showed broader stride widths compared to HS, however, this was only statistically significant in PD-MED–DBS. No differences were found comparing PD-MED and PD-MED–DBS. (Table 3). The overall coefficient of variation for the gait parameters showed higher variability for PD-MED (0.20 ± 0.06) compared to PD-MED–DBS (0.15 ± 0.07) and HS (0.12 ± 0.05).
Intra-group Comparisons
Within groups, no significant differences were found between male and female subjects or between patients with left-sided and right-sided disease-dominance. There were also no differences for gait parameters between the left and right body sides.
3.3. Plantar Cutaneous Vibration Perception
Inter-group Comparisons
No considerable differences between groups were found for contact forces or plantar temperatures. As shown in Figure 6, only the left metatarsal head of PD-MED showed statistically significant differences for the VPTs compared to HS. Nevertheless, patient VPTs seemed to be higher in general compared to VPTs from HS. Although there were no statistically significant differences between patient groups, VPTs from PD-MED tended to be higher compared to the VPTs from PD-MED–DBS. The overall coefficient of variation for VPTs showed slightly higher variability for HS (0.25 ± 0.05) compared to PD-MED (0.18 ± 0.02) and PD-MED–DBS (0.19 ± 0.03).
Intra-group Comparisons
In all three groups, woman had lower VPTs than men, however these were only statistically significant for PD-MED. In PD-MED, the VPTs of left foot were significantly higher compared to the VPTs of the right foot. Moreover, an effect of the disease-dominant side was found, but was not consistent throughout both patient groups. For instance, patients in PD-MED with left-sided disease-dominance showed higher VPTs for the left foot (46.7 µm ± 24.4 µm) compared to the right foot (34.9 µm ± 20.5 µm). Nevertheless, this effect was not found for patients in PD-MED with right-sided disease-dominant or for patients in PD-MED–DBS.
4. Discussion
4.1. Motor Performance
Our first objective was to analyze patients’ axial motor symptoms, such as postural instability and gait difficulties, in comparison to healthy elderly subjects. We also analyzed whether patients treated with anti-parkinsonian medication in conjunction with STN-DBS have superior motor control compared to patients treated with medication alone. Therefore, we implemented different motor tests and used a pressure distribution platform to objectively assess subjects’ motor performance.
4.1.1. Quiescent Bipedal Stance:
Adequately functioning postural control is the foundation on which motor tasks are executed, such as quiescent upright bipedal stance [8,23,121]. As expected, the stance of both patient groups PD-MED and PD-MED–DBS was unstable and jerky compared to the healthy subject group HS. In more detail, patients showed higher COP sway displacements and higher COP sway velocities, which is consistent with several other studies and indicates that, even in situations with low balance requirements, PD patients have diminished postural control [12,13,20,21,23,26,44,104,105,106,122,123,124,125,126,127,128,129,130,131]. However, as shown by several other studies, patients’ postural instability during quiescent stance does not always manifest with increased COP values, but can also present as close to normal or even smaller values [12,28,132,133,134]. This might especially be the case for the akinetic-rigid PD subtype, which is characterized by increased background muscle activity, which leads to a higher degree of co-contraction of the leg muscles and higher joint stiffness [135,136,137]. Consequently, akinetic-rigid PD patients might resort to joint stiffness as a balance strategy, because it increases movement resistance and consequently diminishes sway amplitudes [135,136,137,138,139]. In our study, however, the patient groups comprised various symptom mixtures of the main PD subtypes akinetic-rigid, tremor-dominant, postural instability, and gait disturbance. The overall increased COP values we found for both patient groups might also be explained by the fact that even tremor can interfere with postural sway [140].
Despite their postural instability, the patients in this study did not choose wider stances than healthy subjects, which would have led to more stability, due to an increased base of support. One explanation for this might be that axial rigidity and bradykinesia can influence patients’ standing positions [27,135]. In this regard, it has been shown that PD patients have inadequate anticipatory postural adjustments prior to self-initiated movements and that they have difficulties to scale them up, as needed with wider stance. Therefore, it may become a subconscious habit for PD patients to stand with a narrow stance, similar to healthy subjects, even though this motor test did not require any movement initiation [26,27,135,141]. Another explanation why patients did not compensate for their unstable and jerky stance with a wider standing position might simply have to do with the low level of difficulty and little requirement for balance control necessary to execute bipedal quiescent stance. Even though postural instability is noticeable in early stages of the disease (Hoehn & Yahr Stage 2) and that our patients showed significantly lower self-rated balance confidence, bipedal quiescent stance, as a daily activity, still seems to be within the balance capacity of the PD patients tested in our study [12,23,142,143,144].
Among the investigated COP parameters, we found that spatial parameters, such as COP ranges and the COP 95 % confidence area, seem to be the most promising variables to discriminate between patients and healthy subjects. As supported by De la Casa-Fages et al., the COP 95 % confidence area showed a specificity of 70.6 % and a sensitivity of 100 % for differentiating between PD patients and healthy subjects [145]. In this respect, the COP velocity has also been mentioned as a valuable parameter [20,28].
On the other hand, there seems to be no consensus as to which body direction might be more affected by postural instability. While some authors argue that the ML direction is most impaired [20,131,146], others claim that patients are predominantly prone to falls in the AP direction [12,28,44,137,147]. In our study, we found that PD patients have pronounced postural instability in both the ML and AP directions compared to healthy subjects. However, comparing between both directions within the study groups, the AP directions seems to be more instable compared to the ML direction in general, even for the healthy subjects. There are basically two different postural strategies that control sway in the AP and ML directions; the hip strategy and the ankle strategy [21,135,148,149]. Hip adductor and abductor muscles are mostly involved in controlling postural sway in the ML direction, whereas postural sway in the AP direction involves the activity of plantarflexor and dorsiflexor muscles of the ankle joints [20,28,150]. The direction-specific postural instability in PD patients might be caused by neural constraints of the disease. Hence, patients’ poor muscular control and lower limb coordination caused by higher background activity and related muscle fatigue might be some factors for the jerky and instable balance performance in both directions, especially in the AP direction [12,27,107,151,152]. Nevertheless, both directions seem to be good discriminators between PD patients and healthy subjects [131].
The postural instability found in both of our patient groups might reflect the underlying pathophysiological mechanisms of the disease. Nevertheless, those mechanisms are complex and intricate, as they include numerous neurological structures and their functional interaction. However, the basal ganglia play a key role [4,153,154,155]. It is generally known that the depletion of dopaminergic neurons within the substantia nigra pars compacta causes a hypo-dopaminergic state within the basal ganglia. The lack of sufficient dopamine leads to imbalanced excitatory and inhibitory communication within the entire network of the nigro-striatal and thalami-cortical circuits [5,6,7,156]. This includes structures such as the basal ganglia, the thalamus, the primary and premotor cortices, the supplementary motor area, the visual area of the frontal lobes, the intermediate cingulate cortex, the brainstem, the reticulospinal system, and the cerebellar vermis of the cerebellum, to name a few [4,5,157,158]. Any abnormalities disrupting the interaction of those structures create obstacles for selecting, planning, adapting, and executing motor functions. While the lack of dopamine has been considered the main pathophysiological feature of the disease, there is also evidence that motor symptoms in PD are also intensified by the impairment of non-dopaminergic circuits, such as the cholinergic system [159,160,161,162]. Studies have shown that the reduced activity of structures that compose the mesencephalic locomotor region, such as the pedunculopontine nucleus and its thalamic projections, as well as the adjacent cuneiform nucleus, is associated with axial motor symptoms in PD [159,162,163,164,165,166]. The negative multisystem hypocholinergic effect on motor performance seems to be additionally intensified by the denervation of the basal forebrain [23,159,167].
Comparing PD-MED and PD-MED–DBS with each other, we could not find substantial differences for postural performance in the quiescent stance test. This seems surprising, since numerous studies have reported superior effects of DBS over medication for postural control in general [8,12,15,22,123,125,127,130,131,145,168,169,170,171,172,173,174]. However, there are also studies that report lagging beneficial effects of STN-DBS on postural control, especially in conjunction with medication, which may support our findings for this specific balance test [26,145,170,175,176,177,178]. For example, Yin et al. and De la Casa-Fages et al. concluded that STN-DBS only improved motor performance without medication [145,170]. McNeely and Earhart found no synergistic effect of medication and STN-DBS, and Rocchi et al. stated that DBS might have attenuated the negative effects on motor performance that were introduced by levodopa [178,179].
The heterogeneity between studies might be owed mainly to methodological differences, such as studies using more subjective clinical scales instead of computerized methods, or different parameters and testing conditions, as well as different group compositions [13,23,130,180]. In addition, the influence of varying disease severity and time-related fading of DBS-induced benefits might have had negative effects on our results [98,99,100,173,181,182]. Another explanation might have to do with the low level of difficulty of the quiescent bipedal stance test. As already suggested, this test might not have exploited patients’ balance capacities as needed to eventually differentiate between both patient groups. In this respect, Benatru et al., and Błaszczyk et al. concluded that quasi-static posturography in PD might have limited sensitivity, because it only assesses a single component of balance and therefore should be supplemented by additional, more challenging measures of the stability range, such as functional reach or maximal voluntary leaning tests [12,144].
4.1.2. Functional Limits of Stability
The functional limits of stability test has been suggested as a valuable application-oriented measure of dynamic stability to assess patients’ risk of falling [20,105,106,183]. Within a controlled and safe scenario, the test simulates challenging daily balance situations, such as the transition from sitting to standing, the initiation of gait, or leaning and reaching out to grab something [105,107,183,184,185]. Since PD patients are mostly confronted with self-generated internal balance perturbations, this test might be a sensitive and objective measure with potential to distinguish between PD patients and healthy subjects [104,106,107,150,184,185,186,187], specific PD subtypes [20,106,186,187], and therapy-induced effects on postural control [15,105,171,188,189]. In the current study, this test evaluated subjects’ ability to voluntarily lean as far as possible during standing without losing balance. Therefore, we analyzed the maximum COP displacements and the motion times and velocities during leaning in the anterior and posterior directions.
For the interpretation of maximum COP displacements, it is important to analyze the neutral COP location during upright stance, because any shift of the neutral COP location towards a specific direction influences the capabilities of voluntary maximum leaning. In this regard, studies have shown that concomitant parkinsonian symptoms can shift the neutral COP location during upright stance. For example, Schlenstedt et al. found that the COP location was situated more posteriorly in PD patients with distinct signs of freezing, which might have been a compensatory strategy to obtain a safe stance position and avoid falling forward [106]. The authors of another study reported that PD patients with pronounced camptocormia and stooped posture had a significant COP shift towards anterior compared to healthy subjects [26,144,190]. As our study groups did not comprise patients with severe freezing or camptocormia, the neutral COP position during upright standing was comparable between all three groups and was located at approx. 40 % of the foot length measured from the heel. Mancini et al. also found comparable COP locations between healthy subjects and PD patients during quiescent stance, which suggests better comparability between study groups for the functional limits of stability test [105].
As we found during forward and backward leaning, both of our patient groups showed significantly reduced total COP ranges due to smaller displacements in the anterior and posterior directions compared to our healthy subject group. In other words, PD patients had significantly diminished limits of stability in the sagittal plane and therefore had much less functional balance reserves, in which they can safely sway without falling or being forced to execute compensatory steps. [26,105,185]. Similar results have also been found by several other studies, however there is inconsistency regarding the leaning direction in which PD patients have greater balance restrictions [104,105,106,107,150,184,185,186,187,189]. For example, using force plates, Nikaido et al. and Dona et al. also found that PD patients’ functional balance reserve values in the sagittal plane were smaller compared to healthy subjects [104,150]. In an older study, Schiepatti et al. reported that during forward and backward leaning the total COP range in the anterior-posterior direction was approx. 40 % of the foot length for healthy subjects and approx. 30 % for PD patients. This, however, is much less compared to our study findings [185]. More comparable values were found in the study by Mancini et al., in which healthy subjects showed a total COP range in the sagittal plane of approx. 50 % of the foot length and PD patients showed approx. 40 %. However, in contrast to our study findings, the authors concluded that patients’ limits of stability were primarily a result of the reduction of the maximum body leaning forward, but not backward [105,106,124].
Besides their restricted limits of stability, our PD patients also required more time to reach their maximum leaning posture and consequently showed slower velocities compared to the healthy subjects. When comparing between forward and backward leaning, our patients required significantly longer to reach the maximum posterior COP displacement. These results are supported by Mancini et al., and Kelly et al.. Nevertheless, in Mancini’s study, patients only showed slower velocities during backward leaning [105,187].
There are several factors that might explain patients’ diminished limits of stability and slow movements during maximal voluntary leaning. Physiological factors, such as bradykinesia and rigidity, may have a strong contribution, since they lead to diminished joint mobility, axial inflexibility, and muscle weakness [144,186,191,192,193]. Due to the influence of axial rigidity, it has been reported that PD patients had less postural sway compared to healthy subjects while holding the critical inclined posture [28,185,194]. The authors concluded that the pronounced oscillating body sway in healthy subjects at the boundaries of stability might serve as a balance strategy, whereby motor control continuously changes between keeping balance (actively opposing gravity) and letting go (complying with gravity) [185]. Therefore, PD patients may not be able to reach and maintain higher COP displacements due to impaired temporal discrimination, which is important for executing those alternating minor sway movements, such as healthy subjects do [185,195,196]. Slowness of backward leaning may especially reflect patients’ weakness in the ankle extensors [105]. Another physiological factor includes somatosensory integration for regulating motor control. Parkinsonian motor deficits, such as rigidity and bradykinesia, are generally not isolated deficits, but strongly associated with impaired integration and processing of afferent sensory information from the visual [20,21,22,23], vestibular [8,24,25,26], and somatosensory [29,30,197] systems. Postural control is a closed-loop circuit, which integrates afferent inputs for controlling muscular agonists (activation) and antagonists (inhibition) through direct and indirect pathways of the basal ganglia [153,155,198,199,200]. In fact, the ability to use sensory information seems to depend on the stage of striatal dopamine loss, suggesting the role of dopaminergic pathways for central sensory integration [28,201]. Due to the inclined posture while performing the functional limits of stability test, balance control relies particularly on the vestibular and somatosensory systems. The vestibular system provides information about the position of the body in space and the relative position of each joint with respect to other parts of the body [24,39,40]. In PD patients, it has been reported that vestibular hypofunction and impaired signal processing in higher order sensory integration centers might cause impaired sense of trunk position and tilt misperceptions [8,24,25,27,28]. In addition, the functionality of somatosensory sensors, such as muscle spindles, Golgi tendon organs, joint receptors of the lower extremities, and plantar cutaneous mechanoreceptors, is important for leaning the body towards the limits of stability. These sensory systems provide valuable information about muscle contraction status and ligament and tendon tension [29,30,35,42,197,198,200]. Plantar cutaneous mechanoreceptors gather important information about the pressure distribution and loading shifts underneath the foot during movements and therefore are heavily involved in adapting muscle contraction tone and contraction patterns for reaching the maximal COP displacements [41,42,43,44,45,46,47,48,49,50,51,52]. In PD, all of these afferent sensors show deficits that could have prevented patients from reaching higher limits of stability. Note that the functionality of specific plantar cutaneous mechanoreceptors and therapy-induced effects in PD is another main focus of this study and will be discussed later in more detail. There are also psychological factors that might explain our results. The test is considered “functional” limits of stability, because patients’ postural performance also depends on subjective perception, internal postural control abilities, and fear of falling [20,105,188,202]. To minimize the weight of fear, our patients were assured that an examiner who was standing nearby during the test would keep them from falling. Nevertheless, as our patients also showed lower self-rated balance confidence values compared to the healthy subjects, fear of falling still might have played a major role preventing patients from leaning further. Slowness during leaning may also be related to cautiousness and fear of falling [202].
Regarding the effects of DBS and medication, PD-MED-DBS, which received both therapies simultaneously, performed overall better compared to PD-MED, which was treated with medication alone. More specifically, PD-MED–DBS showed larger limits of stability due to significantly higher COP displacements in the anterior direction. Furthermore, PD-MED–DBS showed a strong trend towards more normalized values for the COP motion times and velocities in both leaning directions compared to PD-MED. Even though the literature reports divergent results for the effects of each therapy on the functional limits of stability, better overall effects seem to be achieved with deep brain stimulation alone or in conjunction with medication, which supports our study findings [15,171,188,189]. For instance, Haito Li et al., showed that balance performance began to improve at 6 months post-surgery DBS, with significant improvements of patients’ limits of stability [171,188]. May et al. and St. George et al. showed improved abilities to reach and lean as far as possible after stimulation was turned on [15,189]. The authors suggest that STN-DBS may enhance the central integration of afferent sensor information by regulating the activity of neurons connected to STN. Hence, the normalized functionality of the entire cortical-striatum-pallid-thalamic-cortical system loop may improve sensory-motor interaction and enable patients to maintain balance, even during inclined posture [4,188,203,204,205,206].
The lack of efficacy for exclusive dopaminergic medication might be explained by the fact that motor dysfunction in PD is not solely generated by the depletion of the dopaminergic, but also by the cholinergic system. Hence, dopaminergic medication may relieve dopamine related dysfunction, but has no effect on motor dysfunction related to the cholinergic system [145,162,207]. Moreover, axial motor symptoms become increasingly refractory to medical treatment over time, and the adaptation to continually higher doses of medication might intensify additional balance related disabilities [98,163,205,208]. Nevertheless, the literature shows conflicting results for improvement [105,187] and no improvement [15,171] of the functional limits of stability after medication intake.
As in our study, the therapies DBS and levodopa medication are usually applied in combination. This common treated state offers positive synergistic effects, for example for PD patients’ postural performance. For instance, Bejjani et al. found that STN-DBS and levodopa in combination improved total motor ability by about 80 % compared to supra-threshold doses of levodopa alone [209]. Colnat-Coulbois et al. concluded that STN stimulation combined with levodopa treatment allowed the basal ganglia to function again, because the stimulation influences dopaminergic and non-dopaminergic pathways separately [174]. Lubik et al. also concluded that the combination of both therapeutic methods showed the best motor outcome [210]. The reduction of medication dosages in combination with DBS might be another major factor contributing to synergistic effects due to diminished levodopa-induced postural side effects [211,212]. Nevertheless, there are also contradictory findings that do not support the hypothesis of synergistic effects of STN-DBS and medication [145,170,176,177,178,179]. However, studies investigating synergistic effects on the limits of stability are rare and use rather subjective clinical scores. There are two studies showing positive effects using both therapies in tandem [15,189]. May et al. suggested that both treatment strategies improved participants’ abilities to reach and lean to the limits of stability. Furthermore, St. George et al. reported synergistic effects, suggesting that combining DBS and levodopa medication improved balance abilities beyond what either treatment provided alone [189]. In this regard, there are also studies that show that reducing medication-induced complications, such as dyskinesia and symptom fluctuations through DBS, might enhance patients’ balance performance [160,168,188,211,213].
The discrepancies between studies might mainly have methodological reasons, such as different independent group compositions with varying disease severities and stimulation durations. Moreover, varying executions of the functional limits of stability test and not normalizing the maximal COP displacement to length of the feet might have added a bias to the results.
4.1.3. Gait
The parkinsonian gait pattern is one of the most typical hallmark symptom of the disease, and generally manifests as a slow and unsteady gait, with shortened and shuffling strides. To characterize patient gait pattern more specifically and determine which parameters show the most clinical value in terms of differentiating between PD patients and healthy subjects, as well as therapy-induced differences between medication and medication in conjunction with DBS, we used a pressure distribution platform and analyzed various gait parameters.
As a spatial-temporal parameter, gait velocity depends on the stride length and the stride time, or rate at which a person walks [214,215]. As our study revealed, the fundamental disturbance of patients’ slowness of gait, seems to be primarily a result of significantly reduced stride length, rather than the reduction of cadence. Our results are therefore in agreement with most other study findings [104,210,214,215,216,217,218,219]. In a meta-analysis, including more than 50 studies, Zanardi et al. showed, that stride length in PD is on average of approx. 16 cm shorter than for healthy subjects, whereas cadence is on average of only approx. 1.75 steps higher than for healthy subjects [215]. Due to the reduction of the stride length, both of our patient groups only reached an average self-selected gait speed of approx. 0.8 m/s. According to Roper et al., this signals ‘‘limitations in community mobility’’ [216]. Patients’ reduced stride length might furthermore reflect the shortened lengths of the COP during the stance phase of gait. With shorter strides, the initial and terminal contact points of the foot with the ground are both shifted towards the middle of the foot, which consequently results in shorter COP lengths for PD patients during walking.
Another parameter indicating patients’ reduced quality of gait is the stride width. In contrast to our findings for the quiescent bipedal stance test, during which patients and healthy subjects had the same narrow foot positions, patients had a broader gait than our healthy subjects. As stride width generally enlarges the base of support, this could be interpreted as a compensation mechanism, through which patients try to counteract disease-related motor difficulties that might impair muscle coordination and stability [27,215,220,221,222]. During gait initiation, it has been shown that the first steps tend to be wider in PD [222]. Since gait velocity changes lower limb kinematics, patients’ broad gait might also have been an effect of slower walking with shortened strides. Due to patients‘ diminished forward walking progression, their limbs are aligned more broadly to compensate for medio-lateral instability [223]. Nevertheless, this is surprising, since most other studies, including a meta-analysis, found gait width in PD patients to be similar or even narrower than in healthy subjects [215,219,224]. Therefore, it has been suggested that patients’ compensatory gait broadness may be overcome by the interference of rigidity, which might impair the hip abductors and therefore affect the range of transversal hip rotation and pelvic tilt [220,221,225].
A good measure of instability during gait is the double support time of the gait cycle, which is defined as the period of time when both feet are in contact with the ground [210,215,226,227]. In our study, PD patients in both groups were in the double support phase significantly longer than the healthy subjects, which suggests reduced dynamic gait stability. This is supported by a meta-analysis by Zenardi et al., who found that PD patients’ double support times were approx. 1.79 % longer on average compared to healthy subjects [215]. Longer double support times can be attributed to an inability to adequately transfer weight to the standing foot, as well as to reduced ankle push off, which relates to rigidity and muscle weakness [228,229,230]. Since most gait parameters are closely related, the elongated double support time might also be a result of patients’ reduced walking speed, reduced stride length, and changes in the cadence [214,229].
Interestingly, those distinct gait difficulties that we found using the pressure distribution platform were poorly reflected by the values of the clinical rating scales. For example, our patients’ average disease severity was approx. 2 on the Hoehn and Yahr scale. This is a value that excludes impairments of balance or gait. However, the mismatch between our findings and the clinical ratings might be explained by the influence of subjectivity and the coarse scoring of most clinical rating scales [23,231]. Moreover, the tests in our study were carried out several months after the clinical examination, which could have emphasized this discrepancy due to disease progression [173,216].
The parkinsonian gait pattern with reduced velocity and shortened strides might be determined predominantly by rigidity and bradykinesia [172,175,210,214,232,233]. The presence of gait festination, which is the execution of quick and small strides to prevent patients from losing balance, could also contribute to the manifestation of this pattern [214,217]. Patients’ hypokinetic gait motion with reduced floor clearance relates to altered muscle activity with pronounced co-contraction and muscle weakness, which also reflects a higher metabolic effort for PD patients during walking [129,143,173,215,217,218,230]. Decreased arm swing amplitudes and chaotic arm swing timing during gait might also impede patients’ gait propulsion [12,137,147,234]. Since motor control is dependent on functional afferent sensory systems, disease-related deficits of the visual [20,21,22,23], vestibular [8,24,25], and somatosensory [29,30,42,197] systems may be another contributing factor to the parkinsonian gait pattern. Psychological factors contributing to patients’ slow gait might be predominantly fear of falling [151,235,236]. This is supported by the significantly lower self-rated gait confidence levels compared to our healthy subjects.
There is no easy answer as to what causes the underlying neurological pathophysiology of the parkinsonian gait pattern, mainly due to the intricate interaction between multiple structures of the central nervous system that are responsible for gait regulation [4,5,6,153,156,157,158,159,161,162,203]. Nevertheless, several mechanisms have been suggested in the literature. In addition to disturbances emanating from the basal ganglia, it has been proposed that degeneration of cholinergic neurons in the basal forebrain is a significant contributor to gait disturbance in PD [159,167,237]. Hence, PD patients’ decreased gait speed might be an early indicator of basal forebrain cholinergic deficits [159]. With advanced imaging technologies and microelectrode recordings, it has also been demonstrated that the mesencephalic locomotor region plays a crucial role in the pathophysiology of parkinsonian gait [238,239,240]. Since specific areas within the pedunculopontine nucleus comprise approx. 40 % of neurons responsible for motor control, their degeneration seems significant [159,162,164,165,166,240,241]. Hence, disrupted afferent connections between the pedunculopontine nucleus, the motor cortex, and the basal ganglia may lead to an inability to control stepping mechanisms [214,242,243,244]. Gait disturbance in PD might also be related to hyperactivation of the cerebellum, a structure known to be crucial for motor coordination [217,238,245]. Cerebellar hyperactivation is considered to be an adaptive response of the central nervous system to compensate for the impaired functionality of the basal ganglia and the brainstem [217,245]. Furthermore, patients’ disorganized movements and hypokinesia may be attributed to increased inhibition of thalamo-cortical fibers, caused by hyperactivity of the globus pallidus internus [129,246]. While interrupted supra-spinal communication between the phasic output from the basal ganglia to the supplementary motor area may explain impaired stride length regulation, defective mechanisms at the spinal level and the brainstem could play a crucial role in the dysregulation of cadence [214,218,243,247,248].
The multisystem degeneration of cholinergic structures, which are responsible for movement control, may explain the increasing dopamine non-responsiveness of gait difficulties in PD. DBS, which acts on the cholinergic systems, partially restoring the functionality of the dopaminergic system, and therefore improving dopamine-responsive symptoms, might help explain why DBS generally performs better than medication in treating parkinsonian gait disorders. Although the literature is controversial, there are various studies that report beneficial effects of DBS on parkinsonian gait [129,169,172,173,178,210,214,216,218,226,227,232,249,250,251,252,253]. In summary, most studies reported improved stride lengths, which was enhancing velocity, whereas cadence was mostly unaffected by DBS. As a general rule, the effect of DBS on gait parameters should be at least as effective as supra-threshold doses of levodopa, but is likely even more effective [210,214,216,218,253,254]. For example, using a pressure-sensitive treadmill, Navratilova et al. found increased step lengths and reduced double support times 3 months after starting neuro-stimulation. The authors concluded that DBS treatment has a superior impact on gait compared to medication alone [227]. This is supported by the results of a meta-analysis by Roper et al., who also reported improved gait speed after switching on DBS [216]. Faist et al. reported that patients’ stride length and mean walking velocity increased almost threefold with STN stimulation, whereas levodopa alone showed smaller effects [214]. Johnsen et al. and Lubik et al. found improved gait performance with increased velocity and stride length comparing the on vs. the off-stimulation states. The authors of both studies also reported improved balance during gait, since patients in the on stimulation state spent less time in the double support phase [210,226]. Lubik et al., however, reported that medication led to a more prominent increase of step length than DBS, whereby the increase in cadence was most pronounced with DBS [210]. Mera et al. and Hausdorff et al. reported reduced variability of gait parameters with DBS, which suggests enhanced gait coordination and stability [251,253,255].
As DBS is usually accompanied by medication, our question was whether both treatments in combination have a positive synergistic effect on gait performance in PD. According to the literature, reports about synergistic effects are inconsistent. For example, using clinical scales, such as the UPDRS and gait and balance scale, it has been reported that gait performance improved significantly more when patients were simultaneously in the on stimulation and on medication states [209,210,249,253]. Using computerized methods, Hausdorff et al., Lubik et al. and Faist et al. reported more specifically, that the combination of both therapeutic treatments showed the best results on gait parameters, including gait speed, stride length, and cadence, as well as stride-to-stride variability [210,214,253]. Stolze et al., reported that both treatments together further increased gait velocity by approx. 30 % compared to DBS alone [218]. However, such synergistic effects were questioned by the findings of various other studies, which failed to observe further improvements in spatial-temporal gait parameters when DBS was added to chronic daily dopaminergic therapy [177,178,179,256,257]. In our study, although the differences between both patient groups were subtle, the patients who received both treatments, medication and DBS, simultaneously, generally showed better gait performance compared to the patients treated with medication alone. Parameters that indicate better gait performance in PD-MED—DBS include stride length, cadence, double support times, and the COP length. Moreover, the overall coefficient of variation for gait parameters was smaller for PD-MED—DBS compared to PD-MED, which indicates better gait control. Therefore, our findings support those studies that reported beneficial synergistic effects.
Improvements of gait parameters using DBS are speculated to be secondary, mainly due to diminished rigidity, bradykinesia, and dyskinesia [172,173,251,253,254,258,259]. Therefore, enlarged stride length and higher gait velocity are the results of enhanced range of motion of the ankle, knee, and hip joints, as well as increased arm swing amplitudes [173,214,218,251,253,256,258,259,260,261]. In addition, improved vertical alignment of the trunk and shank with reduced trunk forward inclination and increased trunk mobility in terms of lateral bending and torsion range may be positive factors [122,127,190,254,258,262,263,264]. DBS-induced gait improvements might also be associated with enhanced lower limb muscle strength, the normalization of leg muscle contraction patterns, and inter-limb coordination [127,143,169,214,228,254,257,261,264,265,266]. As hypothesized in this study, the normalization of proprioceptive functionality and enhanced processing and sensory-motor integration of afferent information through DBS may have additionally lead to enhanced gait performance [19,32,33,34,36,74,90,91,267,268].
The exact mechanisms of DBS and its synergistic effects on gait in combination with medication are still not fully understood. However, thanks to modern functional imaging technologies, it has been proposed that DBS affects not only a single location, but has combined effects at multiple levels simultaneously, including the cortex, the brainstem, and circuits originating from the diencephalon [242,269]. Several studies suggest DBS inducing modulation effects, rather than an exclusive inhibition of neurons or axons in the stimulated area [204,226,270,271,272]. More precisely, DBS of the STN might improve gait by modulating cortical areas that are associated with the preparation and execution of movements. Moreover, STN-DBS may modulate pallido-nigrofugal projections to brainstem regions responsible for generating locomotor patterns [242,269]. Other studies report that the disrupted functional interaction between STN and the sensori-motor and fronto-parietal cortical regions in PD patients with freezing of gait might be improved directly through the stimulation effect of STN on the cortico-striato-pallido-thalamo-cortical system [172,273,274,275]. Investigations with positron-emission tomography of regional cerebral blood flow and metabolism indicate increased activity in the STN during stimulation and thus activity of the output nuclei globus pallidus interna and pars reticulata of substantia nigra [276,277]. Another concept includes DBS-induced normalization of the firing rate of the pedunculopontine nucleus [159,161,163,164,209,210,240]. Synergistic effects, in which the efficacy of levodopa was potentiated in the presence of DBS, may be related to different but additive effects of both treatments on the brain’s central pattern generators or on the pedunculopontine nucleus, which are structures that help regulate gait rhythmicity and stability [214,240,253,265,278,279]. Additive improvements in gait using STN-DBS and medication may also occur through improvements in abnormal cortical activity, including interactions between the supplementary motor area and the basal ganglia, which are important for movement preparation [178,257,280,281]. DBS-induced reductions of medication dosages, which in turn diminish medication-related side effects, such as troublesome dyskinesia, might be another major factor facilitating synergistic effects [211,212,232,282].
The subtle positive effects of DBS on gait found in our study could have several reasons. For example, we presume that disease progression was an issue. This is supported by our findings that patients in PD-MED–DBS were affected by the disease approx. 2.2 times longer, on average, compared to PD-MED. Due to the relatively long stimulation intervals of approx. 3 years since surgery, fading of DBS-induced benefits might also have had negative effects [98,99,100,173,181,182]. For example, Brozova et al. observed positive effects in patients who were treated within 1-2 years, and a significant decline in patients who were treated for 5 years or more. Analyzing results of 11 studies in their meta-analysis, St. George et al. found that gait performance progressively declined after as little as 2 years, and was even worse compared to the pre-surgery state [232]. As suggested, fading of stimulation-induced benefits may not only be associated with a natural progression of PD symptoms, but also with the progressive interference of non-dopaminergic mechanisms that are not responsive to DBS. [98,99,163,173,232,249,283]. Moreover, the lack of DBS efficacy can also be assigned to suboptimal stimulation settings [173]. Since some of our DBS patients were tested before their neurological consultations, they may not have benefited from optimized DBS settings, which could have biased our results towards lower efficacy.
4.2. Plantar Cutaneous Vibration Perception
Our second objective was to investigate tactile somatosensory functionality of the plantar skin in PD patients in comparison to healthy subjects. We also analyzed the therapeutic effects of whether anti-parkinsonian medication alone or medication in conjunction with DBS show different effects on tactile somatosensory functionality of the plantar skin in PD patients. Therefore, we implemented a customized vibration exciter to analyze subjects’ plantar cutaneous vibration perception thresholds.
We hypothesized that PD patients have higher plantar cutaneous vibration perception thresholds than healthy subjects, which was confirmed by our results. Both of our patient groups PD-MED and PD-MED–DBS showed generally higher thresholds, however, statistical significance was only found between PD-MED and HS. Although investigations about tactile cutaneous perception of vibration or pressure in PD are rare, especially for the foot sole, there are a few studies which help to explain our results. Compliance with our study findings can be found in three other studies [64,87,88], while two other studies failed to find PD affecting cutaneous thresholds for mechanical stimuli of the foot [48,89]. Testing cutaneous sensory functionality and analyzing skin biopsies of the foot, Nolano et al. showed that PD patients have significantly increased tactile thresholds, which are strongly associated with patients’ significant loss of epidermal nerve fibers and mechanoreceptors, such as Meissner corpuscles [64]. In another study, Prätorius et al. found significantly higher thresholds in PD patients when analyzing five sites of the plantar foot using a vibration-exciter at 30 Hz and Semmes-Weinstein Monofilaments for touch pressure perception. Their results showed that for each tested location (except the heel) the thresholds of the PD patients were at least twice as high as in the healthy control subjects [88]. Using electrical sinusoidal stimulation at 5, 250, and 2000 Hz at the external malleolus of the foot, Ikeda et al. also found significantly increased perception thresholds for PD patients compared to healthy controls [87]. However, McKeown et al. reported intact cutaneous functionality in PD. Using sophisticated methods, the authors investigated vibro-tactile thresholds at 30 and 250 Hz at the first metatarsal head of the foot and failed to find evidence of elevated plantar thresholds in PD patients [48]. Doty et al. also found no impaired tactile pressure sensitivity in PD patients at the medial sole of the foot and the plantar halluces using a forced-choice staircase threshold test paradigm [89]. The lack of agreement between different study results might primarily be attributed to varying methodological factors, such as the different severity of the disease between the study groups investigated. In this regard, Nolano et al. found that disease severity correlates with the loss of epidermal nerve fibers and Meissner corpuscles in PD. Hence, disease severity, which is associated with disease duration, might be a valuable factor influencing tactile perception [64]. However, when comparing the findings of different studies, there appears to be no consistent pattern for the influence of disease severity and duration on tactile cutaneous perception. In other words, even in studies in which patients had suffered from PD for a relatively long period or had a higher severity of the disease at the time of measurement, no differences in tactile perception were found between PD patients and healthy subjects [48,89]. As sensory perception underlies multifactorial influences, further factors, such as different measurement devices, testing other anatomical locations, and varying vibration frequencies, might also play a role. Furthermore, since only the current study and that by McKeown et al. controlled external factors, such as contact force, this could be another factor influencing tactile perception thresholds [48]. In addition, potential differences in patient skin temperature, which was only controlled in our study, might have had an unknown effect on tactile perception in other studies. Since our healthy subjects were slightly older compared to our patient groups, this could have biased their VPTs towards higher values as an effect of aging. Therefore, study groups with comparable age would have shown differences more dramatically. Finally, the lack of statistical power due to small sample sizes might have contributed to different results, as well. However, in addition to studies that investigated tactile perception of the foot, there are a number of studies that analyzed tactile perception at other anatomical locations in PD, including the arms, hands, fingers, and the torso [32,33,36,37,38,81,90,284]. Most of these studies found that PD increases tactile perception thresholds, which confirms the results of our study.
Given that only the soles of the feet are in direct contact with the ground while standing and walking, afferent information from plantar cutaneous mechanoreceptors is crucial input for motor control. Plantar cutaneous mechanoreceptors gather information about the pressure distribution and load shifts underneath the foot during movements, and therefore are involved in adapting muscle contraction tone and contraction patterns. Sensorimotor integration of plantar mechanoreceptors has already been investigated in several studies with individuals without neurological disease. In summary, decreased plantar cutaneous sensation achieved by anesthesia of the foot sole leads to impaired control of static and dynamic balance abilities [42,46,49,50,51,52,53,54,55,56]. Conversely, sensory stimulation of the foot sole has been shown to improve balance and gait performance. This has been demonstrated in several studies with PD patients using various types of shoe insoles. For instance, Phuenpathom et al. analyzed the effect of mechanical pressure stimulation of the foot sole during the initiation of gait in PD patients. They used insoles with thickened silicon pads and found that the pressure stimulation of the plantar foot skin reduced freezing of gait, which is a devastating motor symptom in PD [43]. In another study, Qiu et al. reported that textured insoles decreased postural sway and improved balance stability under challenging conditions in the PD group, due to facilitating afferent information from mechanoreceptors of the foot sole [44]. Another study analyzed predicting factors of falls in PD. Investigating muscle activity and spatial-temporal parameters during walking, Jenkins et al. found an improvement in the overall stability and safety of gait when using stimulating ribbed insoles. More specifically, the authors reported increased single-limb support time, which implies improved overall stability and normalized timing of the peak activation of the tibialis anterior muscles [45]. Novak and Novak used an elastic vibrating insole that delivered 70 Hz suprathreshold vibration bursts to the heel and the forefoot during the stance phase of gait. Their findings indicate that step-synchronized vibration stimulation of the plantar foot improves gait steadiness in PD with predominant balance impairment. The suprathreshold vibration stimulation improved gait performance by normalizing stride variability, walking speed, stride length, and cadence through enhanced sensory feedback [41]. In another study, the authors stated that the difference in touch thresholds they found between PD patients with and without a history of falling might be an association between reduced peripheral sensation and increased postural instability in the fallers group [285]. In summary, since the stimulation of the plantar foot enhances motor performance in PD patients, the link between reduced plantar sensation and motor symptoms in PD seems plausible. Hence, the reduced plantar cutaneous vibration perception found in our patient groups might be another factor contributing to motor symptoms in PD, such as postural instability or gait difficulties.
The causes of sensory symptoms in PD are multifactorial, however, there is a strong association with widespread deposits of α-synuclein, a fundamental pathological protein, which is also a major component of malicious Lewy bodies in PD patients [16,57,58,59,60,61,62]. Based on neuroanatomical models, the progression of α-synuclein might already begin in the prodromal stage of the disease, initiating in the lower brainstem, autonomic nervous system, and olfactory bulb, advancing in a caudal to rostral direction affecting the diencephalon, basal forebrain, medial temporal lobe structures, and finally neocortical areas [16,65,66,83,286,287,288]. While the progression of sensory symptoms in PD might not only be related to the extension of α-synuclein in specific dopaminergic structures, it also can be present in neurons, presynaptic terminals, and glial cells in the autonomic nervous system, the retina, the central and peripheral nervous systems, and therefore in epidermal nerves of the skin [57,63,64,65,67,68]. Studies have reported that sensory deficits are related to cutaneous denervation in PD, predominantly by α-synuclein. Investigating skin and cutaneous nerves from the abdominal wall in PD patients, Ikemura et al. found extensive Lewy body accumulation in up to 70 % of the investigated cases with Lewy stages II and III, which corresponds to preclinical and early stages of PD [63]. Using skin biopsy to assess peripheral denervation in PD patients, Nolano et al. found lower density of intrapapillary myelinated endings of the glabrous skin compared to healthy subjects. In both the glabrous and the hairy skin, the authors also observed axonal swelling and myelin abnormalities, such as paranodal and distal demyelination, profile segmentation and occasional internodal demyelination of the epidermal nerve fibers. This also includes myelinated axons of the fiber type Aβ, which are responsible for conducting afferent tactile information from mechanoreceptors, such as the Meissner corpuscles, to the central nervous system [289]. More specifically, the number of Meissner corpuscles that detect mechanical vibration stimuli was significantly reduced. Furthermore, Meissner corpuscles even presented a wide range of anomalies, which, according to the authors, suggests the coexistence of degenerative and regenerative processes. The loss of Meissner corpuscles also correlated with the disease severity of the patients. The authors concluded that peripheral deafferentation, including Meissner corpuscles in PD patients, could play a major role in the pathogenesis of sensory dysfunction and could account (at least partly) for the impairment in sensory function in PD [64]. Since we predominantly investigated the functionality of Meissner corpuscles, the findings from Nolano et al. might therefore be one reasonable explanation for why our PD patients also showed impaired plantar cutaneous vibration sensitivity. As those studies mainly show degeneration and deficits of the peripheral nervous system in PD patients, there is also evidence for defective central integration and processing of afferent information at a cerebral level in PD. Although the basal ganglia are considered well-established primarily motor-related structures, there is conjoining clinical and experimental evidence supporting basal ganglia as active “sensory analyzers” for higher-level central somatosensory processing [69,70,71,72,73]. This is plausible, since the basal ganglia are connected to the cortex and do not only receive input from motor areas, but also from cortical somatosensory areas [69,70]. Particularly the STN, one of the main input structures of the basal ganglia, receives projections from multiple cortical, predominantly sensorimotor, areas, whereas its disease-related hyperactivity might cause the loss of functional specificity and ultimately alter somatosensory and sensorimotor integration processing of tactile afferent information [36,290,291,292]. Boecker et al. investigated altered activity of various brain structures, including regional cerebral blood flow and metabolism, using 3D positron emission tomography while applying vibration stimuli to the skin of the index finger. Their results showed that sensory-evoked brain activation in PD patients was reduced in subcortical (basal ganglia) and cortical (parietal and frontal) areas compared to healthy control subjects. More specifically, PD patients showed decreased activation of the contralateral sensorimotor and lateral premotor cortex, the contralateral secondary somatosensory cortex, the contralateral posterior cingulate, the bilateral prefrontal cortex (Brodmann area 10), and the contralateral basal ganglia. In contrast, there was a relative enhanced activation of ipsilateral sensory cortical areas, notably caudal primary and secondary somatosensory cortices and the insular cortex, in PD patients compared to healthy subjects. The authors interpreted their findings as an indication of either altered central focusing and gating of afferent sensory impulses or enhanced compensatory recruitment of associative sensory areas in the presence of patients’ basal ganglia dysfunction. Hence, with their findings, Boecker et al. showed that basal ganglia dysfunction in PD is characterized by abnormal sensory processing, even for tasks devoid of any motor component [73]. In this context, other studies have also suggested that altered tactile perception, including impaired shape discrimination and tactile acuity, reduced roughness detection at the finger tips, altered two-point tactile discrimination thresholds, and abnormal weight perception thresholds, is the result of defective central processing due to diseased basal ganglia in PD [38,75,76,77,78,79,80,81,82]. It is assumed that so called “neural noise” in the somatosensory loops of the basal ganglia may also contribute to the increase of tactile detection thresholds [84,85,86,90]. Disease-related changes of the receptive fields for tactile inputs to the basal ganglia may introduce “noise” into sensory perception, resulting in increased thresholds and reduced discriminative capacities for different sensory modalities [84,85]. This might be emphasized by excessive pathological synchronous neural activity in the beta frequency band (8–35 Hz) throughout the cortico-basal ganglia network in PD patients. Accordingly, cortical oscillations in the beta-range would ‘contaminate’ the oscillatory activity of the basal ganglia and prevent their desynchronization, which is essential for movement control, but could possibly also play a role in sensory processing [86,293,294]. Besides impaired basal ganglia functionality, the dysfunction of extranigral pathways, including the brainstem nuclei, diencephalic and cortical areas, as well as extra-encephalic structures, such as the spinal cord and the autonomic enteric plexus, might be associated with sensory deficits in PD [61,90,295]. In summary, impaired tactile cutaneous perception in PD might be driven by denervation of peripheral epidermal nerve fibers and mechanoreceptors, as well as by defective central integration and processing of afferent information at a cerebral level. Therefore, those pathophysiological mechanisms might help to explain the increased plantar cutaneous vibration thresholds found in our patient groups.
As the pathophysiological mechanisms mentioned above can develop inconsistently and therefore dominate either the left or the right cerebral hemispheres and body sides, this can cause laterality of symptoms. Since laterality is common for motor symptoms, such as tremor, it might also apply to sensory symptoms. This might at least be partly true for our findings, since we found an effect of the disease-dominant side in PD-MED. Patients with a disease-dominant left side showed higher vibration perception thresholds of the left foot compared to the right foot. This is supported by other studies, which also report laterality of sensory symptoms [36,64,83,296]. Nevertheless, we did not find this effect for patients in group PD-MED with a disease-dominant right side or for patients in group PD-MED–DBS, which is consistent with various other studies [34,37,81,87,89,95].
When analyzing whether anti-parkinsonian medication alone or medication in conjunction with DBS results in differences for tactile somatosensory functionality of the plantar skin in PD patients, we generally found higher vibration perception thresholds for PD-MED compared to PD-MED–DBS. Although the comparison between PD-MED and PD-MED–DBS had no statistical significance, our results showed a strong trend towards more impaired tactile perception for patients treated with anti-parkinsonian medication alone. This trend is also supported by the fact that the vibration thresholds only differed significantly between PD-MED and HS, while there was no difference between PD-MED–DBS and HS. Although the effect of anti-parkinsonian medication on sensory deficits, including noxious and innoxious tactile thresholds, and thermal perception in PD is controversial, reports about general insufficiency seem to dominate the literature [19,48,83,87,89,297,298]. For example, investigating plantar vibration perception thresholds in PD patients on and off medication, McKeown et al. found no acute effects of ceasing levodopa intake on plantar sensitivity [48]. Doty et al. also reported that plantar point pressure sensitivity thresholds were not affected by levodopa [89]. Moreover, Gierthmühlen et al. reported that levodopa did not influence detection thresholds or pain sensitivity [37]. Investigating pain perception as a sensory symptom in PD patients, insufficiency of medical treatment was also reported by another study with a large number of patients with early to moderate PD. In this epidemiological study, approx. 80 % of PD patients reported no difference in pain between the on and off medication states [297]. It has even been reported that dopaminergic medication can worsen sensory symptoms in PD, such as proprioception, which might be related to medication-induced side effects due to heavy medication loads [19,89,267,299,300]. Although the processing of different sensory modalities, including proprioception, and noxious and innoxious tactile and thermal perception might not be the same, those studies show rather subtle effects of anti-parkinsonian medication on sensory symptoms and point towards little involvement of dopaminergic systems. Nevertheless, the contribution of dopaminergic systems on sensory symptoms in PD is still unclear.
On the other hand, several studies, including this current study, have shown that DBS of the STN is more promising than anti-parkinsonian medication alone for treating sensory symptoms in PD [19,32,33,36,37,83,90,91,92,95,284,301,302]. For example, Cury et al. stated that DBS has a clear effect on sensory thresholds and changes sensory abnormalities towards normal levels in PD patients [83]. Aman et al., who investigated haptic discrimination thresholds of the hand, also reported enhancements of more than 20 % with DBS compared to without DBS [36]. The authors concluded that improved haptic precision might indicate improved somatosensory functionality by STN-DBS. Their results support the findings from Maschke et al., who also showed a 20 % decrease in position sense threshold as a result of DBS [36,302]. In a more recent study, Sabourin et al. investigated specific settings of directional DBS electrodes on sensory symptoms using a quantitative sensory testing battery, including thermal, pressure, and vibration perception. Although the effects were subtle, their results demonstrated that DBS modulates thermal and mechanical cutaneous sensitivity. DBS pulse width modulated mechanical sensitivity, whereas the DBS total electrical energy modulated thermal sensitivity when using certain directional contacts of the electrodes [32]. Altering the stimulation frequency of DBS, Belasen et al. also analyzed its effects on sensory modalities, including cutaneous pressure and vibration perception. The authors reported that lower DBS frequencies resulted in changed detection thresholds for mechanical pressure and vibration to a greater extent than higher frequencies [33]. In another study, Cury et al. reported lower thermal and mechanical detection thresholds post DBS surgery compared to pre surgery. According to the authors, their data confirmed the existence of sensory abnormalities in PD and suggest that DBS mainly influences detection thresholds rather than painful sensations. In particular, DBS had a significant effect on mechanical and thermal detection thresholds, which were modified toward normal values after DBS surgery. Accordingly, DBS modulated both large and small fiber-dependent sensory input [90]. In contrast, the results of Ciampi de Andrade et al. showed that the detection of large fiber-mediated sensations, including vibration sensations at 100 Hz, did not change in PD patients between on-stim and off-stim conditions. However, PD patients had lower sensitivity to mechanical and thermal pain in the on-stim condition [284]. Dogru Huzmeli et al. also reported reduced thresholds of cutaneous two-point discrimination in PD patients after DBS, suggesting improved somatosensory processing [92]. Using questionnaires, such as the non-motor symptom scale, several other studies also found STN-DBS to improve sensory symptoms in PD patients [303,304,305]. Those study findings support our results, showing that DBS is more efficient in treating sensory symptoms and normalizing tactile cutaneous perception thresholds compared to anti-parkinsonian medication alone. This becomes even more interesting when we consider that our study group that received DBS was affected for twice as long as the group that received medication alone.
Since STN-DBS affects first and foremost the basal ganglia, changes in sensory perception are mainly associated with the modulation of somatosensory information at a cerebral level, while they probably have less effect on the peripheral nervous system per se [74,92,306,307,308,309,310]. The physiological mechanisms by which STN-DBS improves tactile cutaneous perception in PD patients remains unclear, but several hypotheses have been proposed. As STN-DBS acts on fibers and cells in close proximity to the implanted electrodes, an effect on specific somatosensory structures and pathways might be plausible, especially as the nearby thalamus plays a crucial role in processing sensory information [6,32,153,156,307]. In this context, it has been demonstrated that STN-DBS might modulate neural activity in the thalamus and other several cortical areas which are involved in processing tactile information [306,307,311]. Since the posterior parietal region receives information from prefrontal regions, the sensory cortex and multiple thalamic relay nuclei, STN-DBS may not only activate the frontal but also the parietal cortex, which suggests a contribution of the STN to sensory function [306,308]. The STN also has projections to the primary and secondary somatosensory cortices, which are responsible for processing tactile information, so that STN stimulation might affect sensory perception [32,74,306,308,309,312]. Using functional magnetic resonance imaging technology, DiMarzio et al. reported that the activity, especially of the primary somatosensory cortex, might be a promising indicator of whether sensory symptoms in PD patients will respond to STN-DBS [312]. Another study has also mentioned that STN-DBS may alter the activity of the secondary somatosensory cortex, but this still has to be proven [74]. 3D positron-emission tomography has also shown that DBS significantly increases the regional cerebral metabolic rate of glucose consumption in the frontal cortex, temporal cortex, parietal cortex, midbrain and basal ganglia, which may be associated with improved sensation [308,309]. Improved tactile perception through STN-DBS might therefore be the result of normalized inhibition-excitation communication of a comprehensive neuronal network, including numerous dopaminergic and nondopaminergic structures that are responsible for sensory processing [34,160,161,211,313]. Hence, DBS normalizes the disease-related hyperactivity of the STN, and consequently modulates the activation of the somatosensory cortex and enhances sensorimotor integration and processing of tactile afferent information [36,74,95,290,291,292,314]. It has been speculated that the high-frequency DBS signal overwrites the pathological activity of the STN, which causes dysfunction within the basal ganglia-thalamo-cortical circuit [90,310,315]. Given the connectivity between the subthalamus, pallidus, and thalamus and the ascending projections into the somatosensory cortices, DBS-induced regulation of neuronal firing bursts that improve somatosensory processing seems plausible [36,90,267,315,316]. Thus, this mechanism is believed to restore the ability of thalamo-cortical relay cells to respond to depolarizing inputs involved in sensorimotor integration [317,318]. Moreover, it might reduce nigrostriatal “noise” and enhance the signal-to-noise ratio for a better signal discrimination, which is needed for tactile perception [36,90,315,319,320].
Since we found rather subtle effects of STN-DBS on tactile cutaneous perception thresholds compared to various other studies, we questioned what could have caused a potential lack of DBS efficacy. Therefore, mainly methodological limitations, such as differences in surgical procedures, electrode localization in the STN, and different DBS settings, must be considered when interpreting our results. In this context, Pötter-Nerger and Volkmann reported that the lack of STN-DBS efficacy should be distinguished carefully, mainly by a “primary” failure ascribed to suboptimal DBS settings, and by “secondary” failure ascribed to the fading of stimulation-induced benefits due to disease progression [173]. Some of our STN-DBS patients were tested before and some were tested after the neurological consultation. Therefore, the patients who were tested before the consultation did not benefit from possible optimized DBS settings, which could have biased our results towards lower efficacy [182]. The fading of stimulation-induced benefits may also have been an issue, as our STN-DBS patients were affected by the disease for more than twice as long on average, and they also had relatively long STN stimulation intervals of approx. 3 years on average since surgery [98,99,100,173,181,182]. Further possible explanations include different study group compositions and the individual disease severity. Due to patients having high inter-individual symptom characteristics, a larger sample size might have helped to detect greater group differences. In addition, a longitudinal interventional study design, comparing tactile cutaneous perception pre vs. post DBS surgery with and without additional medication, instead of a cross-sectional study design, might have allowed a more precise investigation of which therapy conditions affect tactile cutaneous perception in PD patients.
5. Conclusion
This study investigated axial motor symptoms and somatosensory functionality in PD patients and analyzed whether anti-parkinsonian medication in conjunction with STN-DBS shows different effects compared to medication alone. Healthy subjects were included as a reference. With respect to axial motor symptoms, we hypothesized that patients’ motor performance would be worse compared to those of healthy subjects. Moreover, we assumed that medication in conjunction with additional STN-DBS would be more advantageous in normalizing patients’ impaired motor performance compared to treatment with medication alone.
We found that, regardless of the treatment conditions, PD patients in both study groups showed higher postural sway in the AP and ML directions during quiescent bipedal stance compared to the healthy subject group HS. No differences were found between treatment conditions. The functional limits of stability test revealed smaller base of support ranges, especially characterized by restricted limits of stability in the posterior direction for both patient groups PD-MED and PD-MED–DBS compared to the healthy subject group HS. Moreover, patients in both groups needed longer and moved slower during the test. Nevertheless, PD patients who received medication in conjunction with STN-DBS showed significantly larger limits of stability in the anterior direction, compared to patients who received anti-parkinsonian medication alone. Patient gait was mainly characterized by slowness due to shorter strides and fewer steps per minute, which also reflected longer strides and double support times compared to HS. PD patients also showed broader stride widths compared to HS. Only minor differences pointing towards better gait performance were found for PD-MED–DBS compared to PD-MED. To summarize those results, we found that PD patients suffer from impaired postural stability during quasi-static and dynamic balance situations, as well as from impaired gait performance compared to the healthy subject group, which therefore confirms our hypothesis. Anti-parkinsonian medication in conjunction with STN-DBS tended to be superior for improving patients’ overall motor performance compared to medication alone. This was especially true for the limits of stability and gait. Nevertheless, due to the lack of significant improvements in the majority of the investigated motor parameters, we reject our hypothesis that anti-parkinsonian medication in conjunction with STN-DBS is superior for treating axial motor symptoms compared to medication alone. The discrepancy between our findings and those of studies demonstrating clear DBS-induced positive effects on motor performance could be mainly been due to methodological reasons, as described above. Therefore, our results suggest that testing patients’ postural and gait performance should be challenging and quantify various different aspects of motor performance, in order to evaluate patients’ axial motor symptoms more comprehensively. As prominent clinical markers of patients’ motor disability, the limits of stability in the anterior and posterior directions and gait speed should be observed more carefully. Further studies investigating the effect of DBS and medication on axial motor symptoms should include varying stimulation parameters, as well as different combinations of anti-parkinsonian medication. Besides STN-DBS, the effects of stimulating other mesencephalic areas, such as the pedunculopontine nucleus and globus pallidus interna, on motor performance should also be analyzed. We also recommend additional investigations, including EMG analysis of several muscles controlling the ankle, knee, and the hip joints, as well as 3D motion analysis for investigating full body kinematics during motor performance.
The second objective of this study was to investigate somatosensory functionality by analyzing plantar cutaneous VPTs within the same study groups. Based on the pathophysiological mechanisms of PD and previous study findings, we hypothesized that patients’ plantar cutaneous vibration perception would be impaired compared to healthy subjects. Since our results showed that the plantar cutaneous vibration perception of PD patients who received only anti-parkinsonian medication were significantly higher compared to those of the healthy subject group, we can confirm our hypothesis. This suggests that the pathophysiological mechanisms of PD affect plantar tactile cutaneous perception. Moreover, our hypothesis is supported by the results of various other studies that found impaired tactile cutaneous perception in PD patients by analyzing numerous body regions, including the torso, arms, hands, and fingers, and the feet. Consistent with the argumentation above that plantar mechanoreceptor input contributes to motor control and the study findings that textured insoles improve motor performance in PD patients, our results also suggest that impaired plantar cutaneous vibration perception might therefore contribute to axial motor symptoms in PD patients. Hence, our results may be helpful for developing and implementing plantar tactile cutaneous perception-enhancing therapy strategies. Moreover, they can be used to design and optimize low-cost therapy devices, such as textured insoles, to stimulate plantar cutaneous mechanoreceptors and consequently enhance motor performance in PD patients. As this is the first study investigating the effects of STN-DBS on plantar tactile cutaneous perception in PD patients, we furthermore hypothesized that anti-parkinsonian medication in conjunction with STN-DBS might show superior effects on normalizing patients’ impaired plantar cutaneous vibration perception compared medication alone. Therefore, based on our results that STN-DBS improves plantar cutaneous vibration perception and on other study findings that also showed that STN-DBS improves tactile cutaneous perception, we confirm our hypothesis. Our results suggest that PD patients’ impaired plantar tactile cutaneous perception improves through STN-DBS, presumably by normalizing the integration and processing of afferent input on a higher order cerebral level. Further research investigating plantar tactile cutaneous perception in PD and the effects of anti-parkinsonian medication and DBS should include varying stimulation parameters, as well as different combinations of anti-parkinsonian medication. Besides STN-DBS, the effects of stimulating other mesencephalic areas, such as the pedunculopontine nucleus and globus pallidus interna, on tactile cutaneous perception should also be analyzed. Moreover, subsequent studies should focus on examining the functionality of various cutaneous mechanoreceptors that are sensitive to vibration stimuli at different frequencies, and to noxious and innoxious pressure and temperature stimuli.
Author Contributions
Conceptualization, T.H.; methodology, T.H.; software, T.H.; validation, T.H.; formal analysis, T.H.; investigation, T.H.; resources, T.H. and C.O.; data curation, T.H.; writing—original draft preparation, T.H.; writing—review and editing, T.H., T.L.M., C.O. and P.T.; visualization, T.H.; supervision, T.H., T.L.M; project administration, T.H.; funding acquisition, T.H. All authors have read and agreed to the published version of the manuscript.
Funding
The publication of this article was funded by Chemnitz University of Technology and by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) project number 491193532.
Institutional Review Board Statement
All procedures were conducted according to the recommendations of the Declaration of Helsinki and were approved by the ethics committee of the medical faculty of the University Leipzig (IRB number: 023/14-ff).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The dataset used and analyzed in this study is available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to thank all subjects of this study, and Christian Oehlwein for allowing to use his clinical premises. We would also like to extend special thanks to Renato Germano for supporting the data acquisition, to Ralph Dörfler and Philip Schulze for providing technical support, and to Lisa Peterson for proofreading the article as a native speaker.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weerasak, M.; Aju, M.; Hiroyuki, H.; David Seidel. A systematic review of the worldwide prevalence and incidence of Parkinson's disease. J. Med Assoc Thai 2011, 94, 19–55. [Google Scholar]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson's disease. J. Neural Transm. (Vienna) 2017, 124, 901–905. [Google Scholar] [CrossRef]
- Ou, Z.; Pan, J.; Tang, S.; Duan, D.; Yu, D.; Nong, H.; Wang, Z. Global Trends in the Incidence, Prevalence, and Years Lived With Disability of Parkinson's Disease in 204 Countries/Territories From 1990 to 2019. Front. Public Health 2021, 9, 776847. [Google Scholar] [CrossRef] [PubMed]
- Berardelli, A. Neurophysiology of basal ganglia diseases. Parkinson's Disease and Related Disorders, Part I; Elsevier, 2007; pp 67–75, ISBN 9780444519009.
- Agid, Y. Parkinson's disease: pathophysiology. Lancet 1991, 337, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Blandini, F.; Nappi, G.; Tassorelli, C.; Martignoni, E. Functional changes of the basal ganglia circuitry in Parkinson's disease. Prog. Neurobiol. 2000, 62, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.-E.; Lang, A.E. Parkinson disease. Nature Reviews Disease Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.D.; Allen, N.E.; Canning, C.G.; Fung, V.S.C. Postural instability in patients with Parkinson's disease. Epidemiology, pathophysiology and management. CNS Drugs 2013, 27, 97–112. [Google Scholar] [CrossRef]
- Sveinbjornsdottir, S. The clinical symptoms of Parkinson's disease. J. Neurochem. 2016, 139 (Suppl. S1), 318–324. [Google Scholar] [CrossRef]
- Schapira, A.H. Science, medicine, and the future: Parkinson's disease. BMJ 1999, 318, 311–314. [Google Scholar] [CrossRef]
- Shahed, J.; Jankovic, J. Motor symptoms in Parkinson's disease. Parkinson's Disease and Related Disorders, Part I; Elsevier, 2007; pp 329–342, ISBN 9780444519009.
- Benatru, I.; Vaugoyeau, M.; Azulay, J.-P. Postural disorders in Parkinson's disease. Neurophysiol. Clin. 2008, 38, 459–465. [Google Scholar] [CrossRef]
- Del Din, S.; Godfrey, A.; Coleman, S.; Galna, B.; Lord, S.; Rochester, L. Time-dependent changes in postural control in early Parkinson's disease: what are we missing? Med. Biol. Eng. Comput. 2016, 54, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.R.W.; Wieler, M. Treatment of Parkinson's disease. Can. J. Neurol. Sci. 2003, 30 (Suppl. S1), S27–33. [Google Scholar] [CrossRef] [PubMed]
- May, D.S.; van Dillen, L.R.; Earhart, G.M.; Rawson, K.S.; Perlmutter, J.S.; Duncan, R.P. Effects of Subthalamic Nucleus Deep Brain Stimulation and Levodopa on Balance in People with Parkinson's Disease: A Cross Sectional Study. Brain Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. Neuropathobiology of non-motor symptoms in Parkinson disease. J Neural Transm 2015, 122, 1429–1440. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Schapira, A.H.V. Non-motor symptoms of Parkinson's disease: dopaminergic pathophysiology and treatment. Lancet Neurol. 2009, 8, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, R.F.; Bodis-Wollner, I. Parkinsons Disease and Nonmotor Dysfunction 2013.
- Hogg, E.; Wertheimer, J.; Graner, S.; Tagliati, M. Deep Brain Stimulation and Nonmotor Symptoms. Int. Rev. Neurobiol. 2017, 134, 1045–1089. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, N.; Maas, K.C.; Shah, V.V.; Carlson-Kuhta, P.; Nutt, J.G.; Horak, F.B.; Asaka, T.; Mancini, M. Functional limits of stability and standing balance in people with Parkinson's disease with and without freezing of gait using wearable sensors. Gait Posture 2021, 87, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Nilsson, M.H.; Rehncrona, S.; Tjernström, F.; Magnusson, M.; Johansson, R.; Fransson, P.-A. Strategic alterations of posture are delayed in Parkinson's disease patients during deep brain stimulation. Sci. Rep. 2021, 11, 23550. [Google Scholar] [CrossRef]
- Patel, M.; Nilsson, M.H.; Rehncrona, S.; Tjernström, F.; Magnusson, M.; Johansson, R.; Fransson, P.-A. Effects of Deep Brain Stimulation on Postural Control in Parkinson's Disease. Comput. Biol. Med. 2020, 122, 103828. [Google Scholar] [CrossRef]
- Palakurthi, B.; Burugupally, S.P. Postural Instability in Parkinson's Disease: A Review. Brain Sci. 2019, 9. [Google Scholar] [CrossRef]
- Vitale, C.; Marcelli, V.; Furia, T.; Santangelo, G.; Cozzolino, A.; Longo, K.; Allocca, R.; Amboni, M.; Marciano, E.; Barone, P. Vestibular impairment and adaptive postural imbalance in parkinsonian patients with lateral trunk flexion. Mov. Disord. 2011, 26, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, G.; Wicki, A.; Baumann, C.R.; Straumann, D.; Palla, A. Impaired tilt perception in Parkinson's disease: a central vestibular integration failure. PLoS One 2015, 10, e0124253. [Google Scholar] [CrossRef]
- Park, J.-H.; Kang, Y.-J.; Horak, F.B. What Is Wrong with Balance in Parkinson's Disease? J. Mov. Disord. 2015, 8, 109–114. [Google Scholar] [CrossRef]
- Horak, F.B.; Dimitrova, D.; Nutt, J.G. Direction-specific postural instability in subjects with Parkinson's disease. Experimental Neurology 2005, 193, 504–521. [Google Scholar] [CrossRef] [PubMed]
- Termoz, N.; Halliday, S.E.; Winter, D.A.; Frank, J.S.; Patla, A.E.; Prince, F. The control of upright stance in young, elderly and persons with Parkinson's disease. Gait Posture 2008, 27, 463–470. [Google Scholar] [CrossRef]
- Carpenter, M.G.; Bloem, B.R. Postural control in Parkinson patients: a proprioceptive problem? Experimental Neurology 2011, 227, 26–30. [Google Scholar] [CrossRef]
- Ribeiro, L.; e Souza, T.M.; Bizarro, L.; Oliveira, A. Proprioceptive deficits in Parkinson's disease: From clinical data to animal experimentation. Psychology & Neuroscience 2011, 4, 235–244. [Google Scholar] [CrossRef]
- Vaugoyeau, M.; Viel, S.; Amblard, B.; Azulay, J.P.; Assaiante, C. Proprioceptive contribution of postural control as assessed from very slow oscillations of the support in healthy humans. Gait Posture 2008, 27, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Sabourin, S.; Khazen, O.; DiMarzio, M.; Staudt, M.D.; Williams, L.; Gillogly, M.; Durphy, J.; Hanspal, E.K.; Adam, O.R.; Pilitsis, J.G. Effect of Directional Deep Brain Stimulation on Sensory Thresholds in Parkinson's Disease. Frontiers in Human Neuroscience 2020, 14, 217. [Google Scholar] [CrossRef]
- Belasen, A.; Rizvi, K.; Gee, L.E.; Yeung, P.; Prusik, J.; Ramirez-Zamora, A.; Hanspal, E.; Paiva, P.; Durphy, J.; Argoff, C.E.; et al. Effect of low-frequency deep brain stimulation on sensory thresholds in Parkinson's disease. J. Neurosurg. 2017, 126, 397–403. [Google Scholar] [CrossRef]
- Ciampi de Andrade, D.; Lefaucheur, J.-P.; Galhardoni, R.; Ferreira, K.S.L.; Brandao Paiva, A.R.; Bor-Seng-Shu, E.; Alvarenga, L.; Myczkowski, M.L.; Marcolin, M.A.; de Siqueira, Silvia R D T; et al. Subthalamic deep brain stimulation modulates small fiber-dependent sensory thresholds in Parkinson's disease. Pain 2012, 153, 1107–1113. [CrossRef]
- Zia, S.; Cody, F.W.J.; O'Boyle, D.J. Identification of unilateral elbow-joint position is impaired by Parkinson's disease. Clin. Anat. 2002, 15, 23–31. [Google Scholar] [CrossRef]
- Aman, J.E.; Abosch, A.; Bebler, M.; Lu, C.-H.; Konczak, J. Subthalamic nucleus deep brain stimulation improves somatosensory function in Parkinson's disease. Mov. Disord. 2014, 29, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Gierthmühlen, J.; Arning, P.; Binder, A.; Herzog, J.; Deuschl, G.; Wasner, G.; Baron, R. Influence of deep brain stimulation and levodopa on sensory signs in Parkinson's disease. Mov. Disord. 2010, 25, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Maschke, M.; Tuite, P.J.; Krawczewski, K.; Pickett, K.; Konczak, J. Perception of heaviness in Parkinson's disease. Mov. Disord. 2006, 21, 1013–1018. [Google Scholar] [CrossRef]
- Shagufta Zia. Joint position sense is impaired by Parkinson's disease 2000.
- Day, B.L.; Fitzpatrick, R.C. The vestibular system. Curr. Biol. 2005, 15, R583-6. [Google Scholar] [CrossRef]
- Novak, P.; Novak, V. Effect of step-synchronized vibration stimulation of soles on gait in Parkinson's disease: a pilot study. J. Neuroeng. Rehabil. 2006, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Viseux, F.J. The sensory role of the sole of the foot: Review and update on clinical perspectives. Neurophysiologie Clinique 2020, 50, 55–68. [Google Scholar] [CrossRef]
- Phuenpathom, W.; Panyakaew, P.; Vateekul, P.; Surangsrirat, D.; Hiransuthikul, A.; Bhidayasiri, R. Vibratory and plantar pressure stimulation: Steps to improve freezing of gait in Parkinson's disease. Parkinsonism & Related Disorders 2022, 105, 43–51. [Google Scholar] [CrossRef]
- Qiu, F.; Cole, M.H.; Davids, K.W.; Hennig, E.M.; Silburn, P.A.; Netscher, H.; Kerr, G.K. Effects of textured insoles on balance in people with Parkinson's disease. PLoS One 2013, 8, e83309. [Google Scholar] [CrossRef]
- Jenkins, M.E.; Almeida, Q.J.; Spaulding, S.J.; van Oostveen, R.B.; Holmes, J.D.; Johnson, A.M.; Perry, S.D. Plantar cutaneous sensory stimulation improves single-limb support time, and EMG activation patterns among individuals with Parkinson's disease. Parkinsonism & Related Disorders 2009, 15, 697–702. [Google Scholar]
- Germano, A.M.C.; Heß, T.; Schmidt, D.; Milani, T.L. Effects of plantar hypothermia on quasi-static balance: Two different hypothermic procedures. Gait Posture 2018, 60, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Heß, T.; Milani, T.L.; Meixensberger, J.; Krause, M. Postural performance and plantar cutaneous vibration perception in patients with idiopathic normal pressure hydrocephalus. Heliyon 2021, 7, e05811. [Google Scholar] [CrossRef] [PubMed]
- McKeown, M.D.; Peters, R.M.; Pasman, E.P.; McKeown, M.J.; Carpenter, M.G.; Inglis, J.T. Plantar cutaneous function in Parkinson's disease patients ON and OFF L-dopa. Neuroscience Letters 2016, 629, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Meyer, P.F.; Oddsson, L.I.E.; Luca, C.J. de. The role of plantar cutaneous sensation in unperturbed stance. Exp. Brain Res. 2004, 156, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Nurse, M.A.; Nigg, B.M. The effect of changes in foot sensation on plantar pressure and muscle activity. Clinical Biomechanics 2001, 16, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.D.; Santos, L.C.; Patla, A.E. Contribution of vision and cutaneous sensation to the control of centre of mass (COM) during gait termination. Brain Research 2001, 913, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.D.; McIlroy, W.E.; Maki, B.E. The role of plantar cutaneous mechanoreceptors in the control of compensatory stepping reactions evoked by unpredictable, multi-directional perturbation. Brain Research 2000, 877, 401–406. [Google Scholar] [CrossRef]
- McKeon, P.O.; Hertel, J. Diminished Plantar Cutaneous Sensation and Postural Control. Percept Mot Skills 2007, 104, 56–66. [Google Scholar] [CrossRef]
- McKeon, P.O.; Hertel, J. Plantar hypoesthesia alters time-to-boundary measures of postural control. Somatosens. Mot. Res. 2007, 24, 171–177. [Google Scholar] [CrossRef]
- Song, K.; Kang, T.K.; Wikstrom, E.A.; Jun, H.-p.; Lee, S.Y. Effects of reduced plantar cutaneous sensation on static postural control in individuals with and without chronic ankle instability. J. Sci. Med. Sport 2017, 20, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Germano, A.M.C.; Schmidt, D.; Milani, T.L. Effects of hypothermically reduced plantar skin inputs on anticipatory and compensatory balance responses. BMC Neurosci. 2016, 17, 41. [Google Scholar] [CrossRef]
- Jellinger, K. The role of α-synuclein in neurodegeneration — An update. Translational Neuroscience 2012, 3, 1015. [Google Scholar] [CrossRef]
- Bennett, M.C. The role of α-synuclein in neurodegenerative diseases. Pharmacology & Therapeutics 2005, 105, 311–331. [Google Scholar] [CrossRef]
- Beach, T.G.; Adler, C.H.; Lue, L.; Sue, L.I.; Bachalakuri, J.; Henry-Watson, J.; Sasse, J.; Boyer, S.; Shirohi, S.; Brooks, R.; et al. Unified staging system for Lewy body disorders: correlation with nigrostriatal degeneration, cognitive impairment and motor dysfunction. Acta Neuropathol. 2009, 117, 613–634. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G.; Milenkovic, I.J.; Preusser, M.; Budka, H. Nigral burden of alpha-synuclein correlates with striatal dopamine deficit. Mov. Disord. 2008, 23, 1608–1612. [Google Scholar] [CrossRef] [PubMed]
- Gold, A.; Turkalp, Z.T.; Munoz, D.G. Enteric alpha-synuclein expression is increased in Parkinson's disease but not Alzheimer's disease. Mov. Disord. 2013, 28, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.K.; Zhang, Y.; Lim, K.L.; Tanaka, Y.; Huang, H.; Gao, J.; Ross, C.A.; Dawson, V.L.; Dawson, T.M. Parkin ubiquitinates the alpha-synuclein-interacting protein, synphilin-1: implications for Lewy-body formation in Parkinson disease. Nat. Med. 2001, 7, 1144–1150. [Google Scholar] [CrossRef]
- Ikemura, M.; Saito, Y.; Sengoku, R.; Sakiyama, Y.; Hatsuta, H.; Kanemaru, K.; Sawabe, M.; Arai, T.; Ito, G.; Iwatsubo, T.; et al. Lewy body pathology involves cutaneous nerves. J. Neuropathol. Exp. Neurol. 2008, 67, 945–953. [Google Scholar] [CrossRef]
- Nolano, M.; Provitera, V.; Estraneo, A.; Selim, M.M.; Caporaso, G.; Stancanelli, A.; Saltalamacchia, A.M.; Lanzillo, B.; Santoro, L. Sensory deficit in Parkinson's disease: Evidence of a cutaneous denervation. Brain 2008, 131, 1903–1911. [Google Scholar] [CrossRef]
- Braak, H.; Tredici, K.D. Nervous system pathology in sporadic Parkinson disease. Neurology 2008, 70, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Tredici, K.D.; Rüb, U.; Vos, R.A. de; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiology of Aging 2003, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Mori, F.; Tanji, K.; Orimo, S.; Takahashi, H. Involvement of the peripheral nervous system in synucleinopathies, tauopathies and other neurodegenerative proteinopathies of the brain. Acta Neuropathol. 2010, 120, 1–12. [Google Scholar] [CrossRef]
- Beach, T.G.; Adler, C.H.; Sue, L.I.; Vedders, L.; Lue, L.; White Iii, C.L.; Akiyama, H.; Caviness, J.N.; Shill, H.A.; Sabbagh, M.N.; et al. Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. 2010, 119, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Beudel, M.; Macerollo, A.; Brown, M.J.N.; Chen, R. Editorial: The Role of the Basal Ganglia in Somatosensory-Motor Interactions: Evidence From Neurophysiology and Behavior. Frontiers in Human Neuroscience 2019, 13, 451. [Google Scholar] [CrossRef]
- Nagy, A.; Eördegh, G.; Paróczy, Z.; Márkus, Z.; Benedek, G. Multisensory integration in the basal ganglia. Eur. J. Neurosci. 2006, 24, 917–924. [Google Scholar] [CrossRef]
- Colder, B. The basal ganglia select the expected sensory input used for predictive coding. Front. Comput. Neurosci. 2015, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Peller, M.; Zeuner, K.E.; Munchau, A.; Quartarone, A.; Weiss, M.; Knutzen, A.; Hallett, M.; Deuschl, G.; Siebner, H.R. The basal ganglia are hyperactive during the discrimination of tactile stimuli in writer's cramp. Brain 2006, 129, 2697–2708. [Google Scholar] [CrossRef]
- Boecker, H.; Ceballos-Baumann, A.; Bartenstein, P.; Weindl, A.; Siebner, H.R.; Fassbender, T.; Munz, F.; Schwaiger, M.; Conrad, B. Sensory processing in Parkinson's and Huntington's disease: investigations with 3D H(2)(15)O-PET. Brain 1999, 122 Pt 9, 1651–1665. [Google Scholar] [CrossRef]
- Korsun, O.; Renvall, H.; Nurminen, J.; Mäkelä, J.P.; Pekkonen, E. Modulation of sensory cortical activity by deep brain stimulation in advanced Parkinson's disease. Eur. J. Neurosci. 2022, 56, 3979–3990. [Google Scholar] [CrossRef]
- Schneider, J.S.; Morse, J.R.; Lidsky, T.I. Somatosensory properties of globus pallidus neurons in awake cats. Exp. Brain Res. 1982, 46, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Rothblat, D.S.; Schneider, J.S. Alterations in pallidal neuronal responses to peripheral sensory and striatal stimulation in symptomatic and recovered Parkinsonian cats. Brain Research 1995, 705, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rothblat, D.S.; Schneider, J.S. Response of caudate neurons to stimulation of intrinsic and peripheral afferents in normal, symptomatic, and recovered MPTP-treated cats. J. Neurosci. 1993, 13, 4372–4378. [Google Scholar] [CrossRef] [PubMed]
- Seitz, R.J.; Roland, P.E. Vibratory stimulation increases and decreases the regional cerebral blood flow and oxidative metabolism: a positron emission tomography (PET) study. Acta Neurologica Scandinavica 1992, 86. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S.; Diamond, S.G.; Markham, C.H. Parkinson's disease: sensory and motor problems in arms and hands. Neurology 1987, 37, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Sathian, K.; Zangaladze, A.; Green, J.; Vitek, J.L.; DeLong, M.R. Tactile spatial acuity and roughness discrimination: impairments due to aging and Parkinson's disease. Neurology 1997, 49, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Zia, S.; Cody, F.W.J.; O'Boyle, D.J. Discrimination of bilateral differences in the loci of tactile stimulation is impaired in subjects with Parkinson's disease. Clin. Anat. 2003, 16, 241–247. [Google Scholar] [CrossRef]
- Weder, B.J.; Leenders, K.L.; Vontobel, P.; Nienhusmeier, M.; Keel, A.; Zaunbauer, W.; Vonesch, T.; Ludin, H.-P. Impaired somatosensory discrimination of shape in Parkinson's disease: Association with caudate nucleus dopaminergic function. Hum. Brain Mapp. 1999, 8, 1–12. [Google Scholar] [CrossRef]
- Cury, R.G.; Galhardoni, R.; Fonoff, E.T.; Perez Lloret, S.; Dos Santos Ghilardi, M.G.; Barbosa, E.R.; Teixeira, M.J.; Ciampi de Andrade, D. Sensory abnormalities and pain in Parkinson disease and its modulation by treatment of motor symptoms. Eur. J. Pain 2016, 20, 151–165. [Google Scholar] [CrossRef]
- Conte, A.; Khan, N.; Defazio, G.; Rothwell, J.C.; Berardelli, A. Pathophysiology of somatosensory abnormalities in Parkinson disease. Nat. Rev. Neurol. 2013, 9, 687–697. [Google Scholar] [CrossRef]
- Conte, A.; Modugno, N.; Lena, F.; Dispenza, S.; Gandolfi, B.; Iezzi, E.; Fabbrini, G.; Berardelli, A. Subthalamic nucleus stimulation and somatosensory temporal discrimination in Parkinson's disease. Brain 2010, 133, 2656–2663. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.; Bergman, H.; Brown, P. Pathological synchronization in Parkinson's disease: networks, models and treatments. Trends in Neurosciences 2007, 30, 357–364. [Google Scholar] [CrossRef]
- Ikeda, K.; Deguchi, K.; Kume, K.; Kamada, M.; Touge, T.; Masaki, T. Assessment of sensory perception and processing using current perception threshold in Parkinson's disease. Neurol Clin Neurosci 2013, 1, 209–213. [Google Scholar] [CrossRef]
- Prätorius, B.; Kimmeskamp, S.; Milani, T. The sensitivity of the sole of the foot in patients with Morbus Parkinson. Neuroscience Letters 2003, 346, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Gandhi, S.S.; Osman, A.; Hurtig, H.I.; Pawasarat, I.; Beals, E.; Chung, I.; Dubroff, J.; Newberg, A.; Ying, G.-S.; et al. Point pressure sensitivity in early stage Parkinson's disease. Physiol. Behav. 2015, 138, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Cury, R.G.; Galhardoni, R.; Teixeira, M.J.; Dos Santos Ghilardi, M.G.; Silva, V.; Myczkowski, M.L.; Marcolin, M.A.; Barbosa, E.R.; Fonoff, E.T.; Ciampi de Andrade, D. Subthalamic deep brain stimulation modulates conscious perception of sensory function in Parkinson's disease, ----------; National Fire Protection Assoc: Quincy MA, 2016, ISBN ---------.
- Cury, R.G.; Galhardoni, R.; Fonoff, E.T.; Dos Santos Ghilardi, M.G.; Fonoff, F.; Arnaut, D.; Myczkowski, M.L.; Marcolin, M.A.; Bor-Seng-Shu, E.; Barbosa, E.R.; et al. Effects of deep brain stimulation on pain and other nonmotor symptoms in Parkinson disease. Neurology 2014, 83, 1403–1409. [Google Scholar] [CrossRef]
- Dogru Huzmeli, E.; Yilmaz, A.; Okuyucu, E. Analysis of the effects of subthalamic nucleus deep brain stimulation on somatosensation in Parkinson's disease patients. Neurol. Sci. 2020, 41, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-H.; Zhang, L.; Sperry, L.; Olichney, J.; Farias, S.T.; Shahlaie, K.; Chang, N.M.; Liu, Y.; Wang, S.-P.; Wang, C. Target Selection Recommendations Based on Impact of Deep Brain Stimulation Surgeries on Nonmotor Symptoms of Parkinson's Disease. Chin. Med. J. (Engl) 2015, 128, 3371–3380. [Google Scholar] [CrossRef]
- Djaldetti, R.; Melamed, E. Sensory symptoms in Parkinson's disease. Parkinson's Disease and Related Disorders, Part I; Elsevier, 2007; pp 377–384, ISBN 9780444519009.
- Maruo, T.; Saitoh, Y.; Hosomi, K.; Kishima, H.; Shimokawa, T.; Hirata, M.; Goto, T.; Morris, S.; Harada, Y.; Yanagisawa, T.; et al. Deep brain stimulation of the subthalamic nucleus improves temperature sensation in patients with Parkinson’s disease. Pain 2011, 152, 860–865. [Google Scholar] [CrossRef]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson's disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef]
- Beuter, A.; Modolo, J. Delayed and lasting effects of deep brain stimulation on locomotion in Parkinson's disease. Chaos 2009, 19, 26114. [Google Scholar] [CrossRef] [PubMed]
- Castrioto, A.; Lozano, A.M.; Poon, Y.-Y.; Lang, A.E.; Fallis, M.; Moro, E. Ten-year outcome of subthalamic stimulation in Parkinson disease: a blinded evaluation. Archives of Neurology 2011, 68, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Zibetti, M.; Merola, A.; Rizzi, L.; Ricchi, V.; Angrisano, S.; Azzaro, C.; Artusi, C.A.; Arduino, N.; Marchisio, A.; Lanotte, M.; et al. Beyond nine years of continuous subthalamic nucleus deep brain stimulation in Parkinson's disease. Mov. Disord. 2011, 26, 2327–2334. [Google Scholar] [CrossRef]
- Fasano, A.; Romito, L.M.; Daniele, A.; Piano, C.; Zinno, M.; Bentivoglio, A.R.; Albanese, A. Motor and cognitive outcome in patients with Parkinson's disease 8 years after subthalamic implants. Brain 2010, 133, 2664–2676. [Google Scholar] [CrossRef] [PubMed]
- Germano, A.M.C.; Heß, T.; Schmidt, D.; Milani, T.L. Effects of plantar hypothermia on quasi-static balance: Two different hypothermic procedures. Gait Posture 2018, 60, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Germano, A.M.C.; Schlee, G.; Milani, T.L. Effect of cooling foot sole skin receptors on achilles tendon reflex. Muscle Nerve 2016, 53, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Schlee, G.; Sterzing, T.; Milani, T.L. Foot sole skin temperature affects plantar foot sensitivity. Clin. Neurophysiol. 2009, 120, 1548–1551. [Google Scholar] [CrossRef]
- Nikaido, Y.; Akisue, T.; Kajimoto, Y.; Tucker, A.; Kawami, Y.; Urakami, H.; Iwai, Y.; Sato, H.; Nishiguchi, T.; Hinoshita, T.; et al. Postural instability differences between idiopathic normal pressure hydrocephalus and Parkinson's disease. Clin. Neurol. Neurosurg. 2018, 165, 103–107. [Google Scholar] [CrossRef]
- Mancini, M.; Rocchi, L.; Horak, F.B.; Chiari, L. Effects of Parkinson's disease and levodopa on functional limits of stability. Clin. Biomech. (Bristol, Avon) 2008, 23, 450–458. [Google Scholar] [CrossRef]
- Schlenstedt, C.; Muthuraman, M.; Witt, K.; Weisser, B.; Fasano, A.; Deuschl, G. Postural control and freezing of gait in Parkinson's disease. Parkinsonism & Related Disorders 2016, 24, 107–112. [Google Scholar] [CrossRef]
- Ganesan, M.; Pal, P.K.; Gupta, A.; Sathyaprabha, T.N. Dynamic posturography in evaluation of balance in patients of Parkinson’s disease with normal pull test: Concept of a diagonal pull test. Parkinsonism & Related Disorders 2010, 16, 595–599. [Google Scholar] [CrossRef]
- Müller, K.; Fröhlich, S.; Germano, A.M.C.; Kondragunta, J.; Agoitia Hurtado, M.F.D.C.; Rudisch, J.; Schmidt, D.; Hirtz, G.; Stollmann, P.; Voelcker-Rehage, C. Sensor-based systems for early detection of dementia (SENDA): a study protocol for a prospective cohort sequential study. BMC Neurol. 2020, 20, 84. [Google Scholar] [CrossRef]
- Schmidt, D.; Germano, A.M.; Milani, T.L.; Khaiyat, O. Subjective sensitivity data: Considerations to treat heteroscedasticity. Cogent Medicine 2019, 6, 1673086. [Google Scholar] [CrossRef]
- Cobo, R.; García-Piqueras, J.; Cobo, J.; Vega, J.A. The Human Cutaneous Sensory Corpuscles: An Update. J. Clin. Med. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- McGlone, F.; Reilly, D. The cutaneous sensory system. Neurosci. Biobehav. Rev. 2010, 34, 148–159. [Google Scholar] [CrossRef]
- Schlee, G.; Sterzing, T.; Milani, T.L. Foot sole skin temperature affects plantar foot sensitivity. Clin. Neurophysiol. 2009, 120, 1548–1551. [Google Scholar] [CrossRef]
- Hagander, L.G.; Midani, H.A.; Kuskowski, M.A.; Parry, G.J. Quantitative sensory testing: effect of site and pressure on vibration thresholds. Clin. Neurophysiol. 2000, 111, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Zippenfennig, C.; Wynands, B.; Milani, T.L. Vibration Perception Thresholds of Skin Mechanoreceptors Are Influenced by Different Contact Forces. J. Clin. Med. 2021, 10. [Google Scholar] [CrossRef]
- Mildren, R.L.; Strzalkowski, N.D.J.; Bent, L.R. Foot sole skin vibration perceptual thresholds are elevated in a standing posture compared to sitting. Gait Posture 2016, 43, 87–92. [Google Scholar] [CrossRef]
- Holowka, N.B.; Wynands, B.; Drechsel, T.J.; Yegian, A.K.; Tobolsky, V.A.; Okutoyi, P.; Mang'eni Ojiambo, R.; Haile, D.W.; Sigei, T.K.; Zippenfennig, C.; et al. Foot callus thickness does not trade off protection for tactile sensitivity during walking. Nature 2019, 571, 261–264. [Google Scholar] [CrossRef]
- Drechsel, T.J.; Monteiro, R.L.; Zippenfennig, C.; Ferreira, J.S.S.P.; Milani, T.L.; Sacco, I.C.N. Low and High Frequency Vibration Perception Thresholds Can Improve the Diagnosis of Diabetic Neuropathy. J. Clin. Med. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Claudio Zippenfennig; Tina J. Drechsel; Renan L. Monteiro; Isabel C. N. Sacco and Thomas L. Milani. The Mechanoreceptor’s Role in Plantar Skin Changes in Individuals with Diabetes Mellitus.
- Gomes Paiva, A.F.; Thoumie, P.; Missaoui, B. How far do stabilometric and clinical parameters correlate in peripheral neuropathies? Gait Posture 2017, 52, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Brehm, M.-A.; Scholtes, V.A.; Dallmeijer, A.J.; Twisk, J.W.; Harlaar, J. The importance of addressing heteroscedasticity in the reliability analysis of ratio-scaled variables: an example based on walking energy-cost measurements. Dev. Med. Child Neurol. 2012, 54, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.A. Human balance and posture control during standing and walking. Gait Posture 1995, 3, 193–214. [Google Scholar] [CrossRef]
- Fransson, P.-A.; Nilsson, M.H.; Rehncrona, S.; Tjernström, F.; Magnusson, M.; Johansson, R.; Patel, M. Deep brain stimulation in the subthalamic nuclei alters postural alignment and adaptation in Parkinson's disease. PLoS One 2021, 16, e0259862. [Google Scholar] [CrossRef] [PubMed]
- Schlenstedt, C.; Gavriliuc, O.; Boße, K.; Wolke, R.; Granert, O.; Deuschl, G.; Margraf, N.G. The Effect of Medication and Deep Brain Stimulation on Posture in Parkinson's Disease. Front. Neurol. 2019, 10, 1254. [Google Scholar] [CrossRef] [PubMed]
- Schlenstedt, C.; Mancini, M.; Horak, F.; Peterson, D. Anticipatory Postural Adjustment During Self-Initiated, Cued, and Compensatory Stepping in Healthy Older Adults and Patients With Parkinson Disease. Archives of Physical Medicine and Rehabilitation 2017, 98, 1316–1324.e1. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.A.; Polastri, P.F.; Baptista, A.M.; Lirani-Silva, E.; Simieli, L.; Orcioli-Silva, D.; Beretta, V.S.; Gobbi, L.T. Effects of disease severity and medication state on postural control asymmetry during challenging postural tasks in individuals with Parkinson’s disease. Human Movement Science 2016, 46, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Beretta, V.S.; Gobbi, L.T.B.; Lirani-Silva, E.; Simieli, L.; Orcioli-Silva, D.; Barbieri, F.A. Challenging Postural Tasks Increase Asymmetry in Patients with Parkinson's Disease. PLoS One 2015, 10, e0137722. [Google Scholar] [CrossRef]
- Collomb-Clerc, A.; Welter, M.-L. Effects of deep brain stimulation on balance and gait in patients with Parkinson's disease: A systematic neurophysiological review. Neurophysiol. Clin. 2015, 45, 371–388. [Google Scholar] [CrossRef]
- DiFrancisco-Donoghue, J.; Jung, M.-K.; Geisel, P.; Werner, W.G. Learning effects of the sensory organization test as a measure of postural control and balance in Parkinson's disease. Parkinsonism & Related Disorders 2015, 21, 858–861. [Google Scholar] [CrossRef]
- Erik Lisbjerg Johnsen. Gait and postural instability in Parkinson's disease treated with deep brain stimulation of the subthalamic nucleus 2011, 2011.
- Rocchi, L. Effects of deep brain stimulation and levodopa on postural sway in Parkinson's disease. Journal of Neurology, Neurosurgery & Psychiatry 2002, 73, 267–274. [Google Scholar] [CrossRef]
- Stylianou, A.P.; McVey, M.A.; Lyons, K.E.; Pahwa, R.; Luchies, C.W. Postural sway in patients with mild to moderate Parkinson's disease. Int. J. Neurosci. 2011, 121, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Diener, C.; Scholz, E.; Guschlbauer, B.; Dichgans, J. Increased shortening reaction in Parkinson's disease reflects a difficulty in modulating long loop reflexes. Mov. Disord. 1987, 2, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Schieppati, M.; Nardone, A. Free and supported stance in Parkinson's disease. The effect of posture and 'postural set' on leg muscle responses to perturbation, and its relation to the severity of the disease. Brain 1991, 114 Pt 3, 1227–1244. [Google Scholar] [CrossRef]
- Horak, F.B.; Nutt, J.G.; Nashner, L.M. Postural inflexibility in parkinsonian subjects. Journal of the Neurological Sciences 1992, 111, 46–58. [Google Scholar] [CrossRef]
- Dimitrova, D.; Horak, F.B.; Nutt, J.G. Postural muscle responses to multidirectional translations in patients with Parkinson's disease. J. Neurophysiol. 2004, 91, 489–501. [Google Scholar] [CrossRef]
- Beretta, V.S.; Carpenter, M.G.; Barbieri, F.A.; Santos, P.C.R.; Orcioli-Silva, D.; Pereira, M.P.; Gobbi, L.T.B. Does the impaired postural control in Parkinson's disease affect the habituation to non-sequential external perturbation trials? Clin. Biomech. (Bristol, Avon) 2021, 85, 105363. [Google Scholar] [CrossRef]
- Carpenter, M.G.; Allum, J.H.J.; Honegger, F.; Adkin, A.L.; Bloem, B.R. Postural abnormalities to multidirectional stance perturbations in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1245–1254. [Google Scholar] [CrossRef]
- Beretta, V.S.; Vitório, R.; Santos, P.C.R.D.; Orcioli-Silva, D.; Gobbi, L.T.B. Postural control after unexpected external perturbation: Effects of Parkinson's disease subtype. Human Movement Science 2019, 64, 12–18. [Google Scholar] [CrossRef]
- Winter, D.A.; Patla, A.E.; Prince, F.; Ishac, M.; Gielo-Perczak, K. Stiffness control of balance in quiet standing. J. Neurophysiol. 1998, 80, 1211–1221. [Google Scholar] [CrossRef]
- Néstor, G.-J. Parkinson’s Disease. Neurobiol. Dis. 2007, 51–67. [Google Scholar] [CrossRef]
- Rocchi, L.; Chiari, L.; Mancini, M.; Carlson-Kuhta, P.; Gross, A.; Horak, F.B. Step initiation in Parkinson’s disease: Influence of initial stance conditions. Neuroscience Letters 2006, 406, 128–132. [Google Scholar] [CrossRef]
- Song, J.; Sigward, S.; Fisher, B.; Salem, G.J. Altered Dynamic Postural Control during Step Turning in Persons with Early-Stage Parkinson's Disease. Parkinsons. Dis. 2012, 2012, 386962. [Google Scholar] [CrossRef] [PubMed]
- Heß, T.; Oehlwein, C.; Milani, T.L. Anticipatory Postural Adjustments and Compensatory Postural Responses to Multidirectional Perturbations—Effects of Medication and Subthalamic Nucleus Deep Brain Stimulation in Parkinson’s Disease. Brain Sci. 2023, 13, 454. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, J.W.; Orawiec, R.; Duda-Kłodowska, D.; Opala, G. Assessment of postural instability in patients with Parkinson's disease. Exp. Brain Res. 2007, 183, 107–114. [Google Scholar] [CrossRef]
- La Casa-Fages, B. de; Alonso-Frech, F.; Grandas, F. Effect of subthalamic nucleus deep brain stimulation on balance in Parkinson's disease: A static posturographic analysis. Gait Posture 2017, 52, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.L.; Collins, J.J.; Luca, C.J. de; Burrows, A.; Lipsitz, L.A. Open-loop and closed-loop postural control mechanisms in Parkinson's disease: increased mediolateral activity during quiet standing. Neuroscience Letters 1995, 197, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.E.; Oude Nijhuis, L.B.; Janssen, L.; Bastiaanse, C.M.; Borm, G.F.; Duysens, J.; Bloem, B.R. Dynamic posturography in Parkinson's disease: diagnostic utility of the "first trial effect". Neuroscience 2010, 168, 387–394. [Google Scholar] [CrossRef]
- Blenkinsop, G.M.; Pain, M.T.G.; Hiley, M.J. Balance control strategies during perturbed and unperturbed balance in standing and handstand. R. Soc. Open Sci. 2017, 4, 161018. [Google Scholar] [CrossRef]
- Horak, F.B.; Nashner, L.M.; Diener, H.C. Postural strategies associated with somatosensory and vestibular loss. Exp. Brain Res. 1990, 82, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Doná, F.; Aquino, C.C.; Gazzola, J.M.; Borges, V.; Silva, S.; Ganança, F.F.; Caovilla, H.H.; Ferraz, H.B. Changes in postural control in patients with Parkinson's disease: A posturographic study. Physiotherapy 2015. [CrossRef] [PubMed]
- Am Grimbergen, Y.; Munneke, M.; Bloem, B.R. Falls in Parkinson?: ?s disease. Curr. Opin. Neurol. 2004, 17, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Bosek, M.; Grzegorzewski, B.; Kowalczyk, A.; Lubiński, I. Degradation of postural control system as a consequence of Parkinson's disease and ageing. Neuroscience Letters 2005, 376, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Crenna, P.; Carpinella, I.; Lopiano, L.; Marzegan, A.; Rabuffetti, M.; Rizzone, M.; Lanotte, M.; Ferrarin, M. Influence of basal ganglia on upper limb locomotor synergies. Evidence from deep brain stimulation and L-DOPA treatment in Parkinson's disease. Brain 2008, 131, 3410–3420. [Google Scholar] [CrossRef] [PubMed]
- Shohamy, D.; Myers, C.E.; Onlaor, S.; Gluck, M.A. Role of the basal ganglia in category learning: how do patients with Parkinson's disease learn? Behav. Neurosci. 2004, 118, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Maschke, M.; Gomez, C.M.; Tuite, P.J.; Konczak, J. Dysfunction of the basal ganglia, but not the cerebellum, impairs kinaesthesia. Brain 2003, 126, 2312–2322. [Google Scholar] [CrossRef]
- Samadi, P.; Rouillard, C.; Bédard, P.J.; Di Paolo, T. Functional neurochemistry of the basal ganglia. Parkinson's Disease and Related Disorders, Part I; Elsevier, 2007; pp 19–66, ISBN 9780444519009.
- Takakusaki, K. Functional Neuroanatomy for Posture and Gait Control. J. Mov. Disord. 2017, 10, 1–17. [Google Scholar] [CrossRef]
- Takakusaki, K. Neurophysiology of gait: from the spinal cord to the frontal lobe. Mov. Disord. 2013, 28, 1483–1491. [Google Scholar] [CrossRef]
- Bohnen, N.I.; Frey, K.A.; Studenski, S.; Kotagal, V.; Koeppe, R.A.; Scott, P.J.H.; Albin, R.L.; Müller, M.L.T.M. Gait speed in Parkinson disease correlates with cholinergic degeneration. Neurology 2013, 81, 1611–1616. [Google Scholar] [CrossRef]
- Perez-Lloret, S.; Barrantes, F.J. Deficits in cholinergic neurotransmission and their clinical correlates in Parkinson's disease. NPJ Parkinsons. Dis. 2016, 2, 16001. [Google Scholar] [CrossRef] [PubMed]
- Rinne, J.O.; Ma, S.Y.; Lee, M.S.; Collan, Y.; Röyttä, M. Loss of cholinergic neurons in the pedunculopontine nucleus in Parkinson's disease is related to disability of the patients. Parkinsonism & Related Disorders 2008, 14, 553–557. [Google Scholar] [CrossRef]
- Karachi, C.; Grabli, D.; Bernard, F.A.; Tandé, D.; Wattiez, N.; Belaid, H.; Bardinet, E.; Prigent, A.; Nothacker, H.-P.; Hunot, S.; et al. Cholinergic mesencephalic neurons are involved in gait and postural disorders in Parkinson disease. J. Clin. Invest. 2010, 120, 2745–2754. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.-M.; Loria, Y.; Saint-Hilaire, M.-H.; Lhermitte, F.; Agid, Y. Does long-term aggravation of Parkinson\textquoterights disease result from nondopaminergic lesions? Neurology 1987, 37, 1539. [Google Scholar] [CrossRef] [PubMed]
- Pahapill, P.A.; Lozano, A.M. The pedunculopontine nucleus and Parkinson's disease. Brain 2000, 123 Pt 9, 1767–1783. [Google Scholar] [CrossRef]
- Zweig, R.M.; Jankel, W.R.; Hedreen, J.C.; Mayeux, R.; Price, D.L. The pedunculopontine nucleus in Parkinson's disease. Ann. Neurol. 1989, 26, 41–46. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Graybiel, A.M.; Duyckaerts, C.; Javoy-Agid, F. Neuronal loss in the pedunculopontine tegmental nucleus in Parkinson disease and in progressive supranuclear palsy. Proc. Natl. Acad. Sci. U. S. A. 1987, 84, 5976–5980. [Google Scholar] [CrossRef]
- Rochester, L.; Yarnall, A.J.; Baker, M.R.; David, R.V.; Lord, S.; Galna, B.; Burn, D.J. Cholinergic dysfunction contributes to gait disturbance in early Parkinson's disease. Brain 2012, 135, 2779–2788. [Google Scholar] [CrossRef]
- Oz, F.; Yucekeya, B.; Huzmeli, I.; Yilmaz, A. Does subthalamic nucleus deep brain stimulation affect the static balance at different frequencies? Neurocirugía 2023, 34, 60–66. [Google Scholar] [CrossRef]
- Sato, K.; Hokari, Y.; Kitahara, E.; Izawa, N.; Hatori, K.; Honaga, K.; Oyama, G.; Hatano, T.; Iwamuro, H.; Umemura, A.; et al. Short-Term Motor Outcomes in Parkinson's Disease after Subthalamic Nucleus Deep Brain Stimulation Combined with Post-Operative Rehabilitation: A Pre-Post Comparison Study. Parkinsons. Dis. 2022, 2022, 8448638. [Google Scholar] [CrossRef]
- Yin, Z.; Bai, Y.; Zou, L.; Zhang, X.; Wang, H.; Gao, D.; Qin, G.; Ma, R.; Zhang, K.; Meng, F.; et al. Balance response to levodopa predicts balance improvement after bilateral subthalamic nucleus deep brain stimulation in Parkinson's disease. NPJ Parkinsons. Dis. 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, S.; Yu, Y.; Wang, Y.; Cheng, Y.; Yang, H.; Tong, X. Effect of Subthalamic Nucleus Deep Brain Stimulation (STN-DBS) on balance performance in Parkinson's disease. PLoS One 2020, 15, e0238936. [Google Scholar] [CrossRef] [PubMed]
- Szlufik, S.; Kloda, M.; Friedman, A.; Potrzebowska, I.; Gregier, K.; Mandat, T.; Przybyszewski, A.; Dutkiewicz, J.; Figura, M.; Habela, P.; et al. The Neuromodulatory Impact of Subthalamic Nucleus Deep Brain Stimulation on Gait and Postural Instability in Parkinson's Disease Patients: A Prospective Case Controlled Study. Front. Neurol. 2018, 9, 906. [Google Scholar] [CrossRef] [PubMed]
- Pötter-Nerger, M.; Volkmann, J. Deep brain stimulation for gait and postural symptoms in Parkinson's disease. Mov. Disord. 2013, 28, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Colnat-Coulbois, S. Bilateral subthalamic nucleus stimulation improves balance control in Parkinson's disease. Journal of Neurology, Neurosurgery & Psychiatry 2005, 76, 780–787. [Google Scholar] [CrossRef]
- St George, R.J.; Carlson-Kuhta, P.; Burchiel, K.J.; Hogarth, P.; Frank, N.; Horak, F.B. The effects of subthalamic and pallidal deep brain stimulation on postural responses in patients with Parkinson disease. J. Neurosurg. 2012, 116, 1347–1356. [Google Scholar] [CrossRef]
- Benabid, A.-L.; Koudsié, A.; Benazzouz, A.; Fraix, V.; Ashraf, A.; Le Bas, J.F.; Chabardes, S.; Pollak, P. Subthalamic Stimulation for Parkinson’s Disease. Archives of Medical Research 2000, 31, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Maurer, C.; Mergner, T.; Xie, J.; Faist, M.; Pollak, P.; Lucking, C.H. Effect of chronic bilateral subthalamic nucleus (STN) stimulation on postural control in Parkinson's disease. Brain 2003, 126, 1146–1163. [Google Scholar] [CrossRef]
- McNeely, M.E.; Earhart, G.M. Medication and subthalamic nucleus deep brain stimulation similarly improve balance and complex gait in Parkinson disease. Parkinsonism & Related Disorders 2013, 19, 86–91. [Google Scholar] [CrossRef]
- Rocchi, L.; Carlson-Kuhta, P.; Chiari, L.; Burchiel, K.J.; Hogarth, P.; Horak, F.B. Effects of deep brain stimulation in the subthalamic nucleus or globus pallidus internus on step initiation in Parkinson disease: laboratory investigation. J. Neurosurg. 2012, 117, 1141–1149. [Google Scholar] [CrossRef]
- Niewboer. Walking ability after implantation of a pallidal stimulator:analysis of plantar force distribution in patients with Parkinson’s disease 1998.
- Nantel, J.; McDonald, J.C.; Bronte-Stewart, H. Effect of medication and STN-DBS on postural control in subjects with Parkinson’s disease. Parkinsonism & Related Disorders 2012, 18, 285–289. [Google Scholar] [CrossRef]
- Krishnamurthi, N.; Mulligan, S.; Mahant, P.; Samanta, J.; Abbas, J.J. Deep brain stimulation amplitude alters posture shift velocity in Parkinson's disease. Cogn. Neurodyn. 2012, 6, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Duncan, P.W.; Weiner, D.K.; Chandler, J.; Studenski, S. Functional reach: a new clinical measure of balance. J. Gerontol. 1990, 45, M192-7. [Google Scholar] [CrossRef] [PubMed]
- Stack, E.; Ashburn, A.; Jupp, K. Postural instability during reaching tasks in Parkinson's disease. Physiother. Res. Int. 2005, 10, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Schieppati, M.; Hugon, M.; Grasso, M.; Nardone, A.; Galante, M. The limits of equilibrium in young and elderly normal subjects and in parkinsonians. Electroencephalography and Clinical Neurophysiology/Evoked Potentials Section 1994, 93, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Behrman, A.L.; Light, K.E.; Flynn, S.M.; Thigpen, M.T. Is the functional reach test useful for identifying falls risk among individuals with Parkinson's disease? Archives of Physical Medicine and Rehabilitation 2002, 83, 538–542. [Google Scholar] [CrossRef]
- Kelly, V.E.; Hyngstrom, A.S.; Rundle, M.M.; Bastian, A.J. Interaction of levodopa and cues on voluntary reaching in Parkinson's disease. Mov. Disord. 2002, 17, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, S.; Yu, Y.; Wang, Y.; Cheng, Y.; Yang, H.; Tong, X. Clinical experience of comprehensive treatment on the balance function of Parkinson's disease. Medicine (Baltimore) 2020, 99, e20154. [Google Scholar] [CrossRef]
- St George, R.J.; Carlson-Kuhta, P.; Nutt, J.G.; Hogarth, P.; Burchiel, K.J.; Horak, F.B. The effect of deep brain stimulation randomized by site on balance in Parkinson's disease. Mov. Disord. 2014, 29, 949–953. [Google Scholar] [CrossRef]
- Schulz-Schaeffer, W.J.; Margraf, N.G.; Munser, S.; Wrede, A.; Buhmann, C.; Deuschl, G.; Oehlwein, C. Effect of neurostimulation on camptocormia in Parkinson's disease depends on symptom duration. Mov. Disord. 2015, 30, 368–372. [Google Scholar] [CrossRef]
- Gamborg, M.; Hvid, L.G.; Thrue, C.; Johansson, S.; Franzén, E.; Dalgas, U.; Langeskov-Christensen, M. Muscle Strength and Power in People With Parkinson Disease: A Systematic Review and Meta-analysis, United States, 2023 Jan 1.
- Wolke, R.; Kuhtz-Buschbeck, J.P.; Deuschl, G.; Margraf, N.G. Insufficiency of trunk extension and impaired control of muscle force in Parkinson’s disease with camptocormia. Clinical Neurophysiology 2020, 131, 2621–2629. [Google Scholar] [CrossRef]
- Santos, M.J.; Kanekar, N.; Aruin, A.S. The role of anticipatory postural adjustments in compensatory control of posture: 1. Electromyographic analysis. J. Electromyogr. Kinesiol. 2010, 20, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Rougier, P.; Burdet, C.; Farenc, I.; Berger, L. Backward and forward leaning postures modelled by an fBm framework. Neuroscience Research 2001, 41, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Artieda, J.; Pastor, M.A.; Lacruz, F.; Obeso, J.A. Temporal discrimination is abnormal in Parkinson's disease. Brain 1992, 115 Pt 1, 199–210. [Google Scholar] [CrossRef]
- Bernardinis, M.; Atashzar, S.F.; Jog, M.S.; Patel, R.V. Differential Temporal Perception Abilities in Parkinson's Disease Patients Based on Timing Magnitude. Sci. Rep. 2019, 9, 19638. [Google Scholar] [CrossRef]
- Vaugoyeau, M.; Hakam, H.; Azulay, J.-P. Proprioceptive impairment and postural orientation control in Parkinson's disease. Human Movement Science 2011, 30, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.G. Proprioceptive regulation of locomotion. Current Opinion in Neurobiology 1995, 5, 786–791. [Google Scholar] [CrossRef]
- Jacobs, J.V.; Horak, F.B. Abnormal proprioceptive-motor integration contributes to hypometric postural responses of subjects with Parkinson's disease. Neuroscience 2006, 141, 999–1009. [Google Scholar] [CrossRef]
- Konczak. Proprioception and Motor Control in Parkinson’s Disease 2009.
- Henderson, J.M.; Watson, S.; Halliday, G.M.; Heinemann, T.; Gerlach, M. Relationships between various behavioural abnormalities and nigrostriatal dopamine depletion in the unilateral 6-OHDA-lesioned rat. Behavioural Brain Research 2003, 139, 105–113. [Google Scholar] [CrossRef]
- Franchignoni, F.; Martignoni, E.; Ferriero, G.; Pasetti, C. Balance and fear of falling in Parkinson's disease. Parkinsonism & Related Disorders 2005, 11, 427–433. [Google Scholar] [CrossRef]
- Hariz, M.; Blomstedt, P. Deep brain stimulation for Parkinson's disease. J. Intern. Med. 2022, 292, 764–778. [Google Scholar] [CrossRef]
- Agnesi, F.; Johnson, M.D.; Vitek, J.L. Deep brain stimulation: how does it work? Handbook of Clinical Neurology 2013, 116, 39–54. [Google Scholar] [CrossRef]
- Groiss, S.J.; Wojtecki, L.; Südmeyer, M.; Schnitzler, A. Deep brain stimulation in Parkinson's disease. Ther. Adv. Neurol. Disord. 2009, 2, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, J.S.; Mink, J.W. Deep brain stimulation. Annu. Rev. Neurosci. 2006, 29, 229–257. [Google Scholar] [CrossRef]
- Bloem, B.R.; Beckley, D.J.; van Dijk, J.G.; Zwinderman, A.H.; Remler, M.P.; Roos, R.A. Influence of dopaminergic medication on automatic postural responses and balance impairment in Parkinson's disease. Mov. Disord. 1996, 11, 509–521. [Google Scholar] [CrossRef]
- Goetz, C.G.; Poewe, W.; Rascol, O.; Sampaio, C. Evidence-based medical review update: pharmacological and surgical treatments of Parkinson's disease: 2001 to 2004. Mov. Disord. 2005, 20, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Bejjani, B.P.; Gervais, D.; Arnulf, I.; Papadopoulos, S.; Demeret, S.; Bonnet, A.M.; Cornu, P.; Damier, P.; Agid, Y. Axial parkinsonian symptoms can be improved: the role of levodopa and bilateral subthalamic stimulation. J. Neurol. Neurosurg. Psychiatry 2000, 68, 595–600. [Google Scholar] [CrossRef]
- Lubik, S.; Fogel, W.; Tronnier, V.; Krause, M.; König, J.; Jost, W.H. Gait analysis in patients with advanced Parkinson disease: different or additive effects on gait induced by levodopa and chronic STN stimulation. J. Neural Transm. (Vienna) 2006, 113, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Goto, S.; Hamasaki, T.; Kuratsu, J.-I. Effect of bilateral subthalamic nucleus stimulation on levodopa-unresponsive axial symptoms in Parkinson's disease. Acta Neurochirurgica 2008, 150, 15–22; discussion 22. [Google Scholar] [CrossRef]
- Benabid, A.L. Deep brain stimulation for Parkinson’s disease. Current Opinion in Neurobiology 2003, 13, 696–706. [Google Scholar] [CrossRef]
- Krack, P.; Batir, A.; van Blercom, N.; Chabardes, S.; Fraix, V.; Ardouin, C.; Koudsie, A.; Limousin, P.D.; Benazzouz, A.; LeBas, J.F.; et al. Five-year follow-up of bilateral stimulation of the subthalamic nucleus in advanced Parkinson's disease. N. Engl. J. Med. 2003, 349, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Faist, M. Effect of bilateral subthalamic nucleus stimulation on gait in Parkinson's disease. Brain 2001, 124, 1590–1600. [Google Scholar] [CrossRef] [PubMed]
- Zanardi, A.P.J.; da Silva, E.S.; Costa, R.R.; Passos-Monteiro, E.; Dos Santos, I.O.; Kruel, L.F.M.; Peyré-Tartaruga, L.A. Gait parameters of Parkinson's disease compared with healthy controls: a systematic review and meta-analysis. Sci. Rep. 2021, 11, 752. [Google Scholar] [CrossRef]
- Roper, J.A.; Kang, N.; Ben, J.; Cauraugh, J.H.; Okun, M.S.; Hass, C.J. Deep brain stimulation improves gait velocity in Parkinson's disease: a systematic review and meta-analysis. J. Neurol. 2016, 263, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Grabli, D.; Karachi, C.; Welter, M.-L.; Lau, B.; Hirsch, E.C.; Vidailhet, M.; François, C. Normal and pathological gait: what we learn from Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 2012, 83, 979–985. [Google Scholar] [CrossRef]
- Stolze, H.; Klebe, S.; Poepping, M.; Lorenz, D.; Herzog, J.; Hamel, W.; Schrader, B.; Raethjen, J.; Wenzelburger, R.; Mehdorn, H.M.; et al. Effects of bilateral subthalamic nucleus stimulation on parkinsonian gait. Neurology 2001, 57, 144–146. [Google Scholar] [CrossRef]
- Stolze, H.; Kuhtz-Buschbeck, J.P.; Drücke, H.; Jöhnk, K.; Illert, M.; Deuschl, G. Comparative analysis of the gait disorder of normal pressure hydrocephalus and Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 2001, 70, 289–297. [Google Scholar] [CrossRef]
- Charlett, A.; Weller, C.; Purkiss, A.G.; Dobbs, S.M.; Dobbs, R.J. Breadth of base whilst walking: effect of ageing and parkinsonism. Age Ageing 1998, 27, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Kirollos, C.; Charlett, A.; O'Neill, C.J.; Kosik, R.; Mozol, K.; Purkiss, A.G.; Bowes, S.G.; Nicholson, P.W.; Hunt, W.B.; Weller, C.; et al. Objective measurement of activation of rigidity: diagnostic, pathogenetic and therapeutic implications in parkinsonism. Br. J. Clin. Pharmacol. 1996, 41, 557–564. [Google Scholar] [CrossRef]
- Bovonsunthonchai, S.; Vachalathiti, R.; Pisarnpong, A.; Khobhun, F.; Hiengkaew, V. Spatiotemporal gait parameters for patients with Parkinson's disease compared with normal individuals. Physiother. Res. Int. 2014, 19, 158–165. [Google Scholar] [CrossRef]
- Sarah A. Brinkerhoff; William M. Murrah; Jaimie A. Roper. All known cemeteries of Clayton County, Georgia; Ancestors Unlimited: Jonesboro Ga. (P.O. Box 1507 Jonesboro 30236), 1986, ISBN 0123456789.
- Bugalho, P.; Alves, L.; Miguel, R. Gait dysfunction in Parkinson's disease and normal pressure hydrocephalus: a comparative study. J. Neural Transm. (Vienna) 2013, 120, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Weller, C.; Humphrey, S.J.; Kirollos, C.; Bowes, S.G.; Charlett, A.; Dobbs, S.M.; Dobbs, R.J. Gait on a shoestring: falls and foot separation in parkinsonism. Age Ageing 1992, 21, 242–244. [Google Scholar] [CrossRef]
- Johnsen, E.L.; Sunde, N.; Mogensen, P.H.; Ostergaard, K. MRI verified STN stimulation site--gait improvement and clinical outcome. Eur. J. Neurol. 2010, 17, 746–753. [Google Scholar] [CrossRef]
- Navratilova, D.; Krobot, A.; Otruba, P.; Nevrly, M.; Krahulik, D.; Kolar, P.; Kolarova, B.; Kaiserova, M.; Mensikova, K.; Vastik, M.; et al. Deep Brain Stimulation Effects on Gait Pattern in Advanced Parkinson's Disease Patients. Front. Neurosci. 2020, 14, 814. [Google Scholar] [CrossRef]
- Johnsen, E.L.; Mogensen, P.H.; Sunde, N.A.; Ostergaard, K. Improved asymmetry of gait in Parkinson's disease with DBS: gait and postural instability in Parkinson's disease treated with bilateral deep brain stimulation in the subthalamic nucleus. Mov. Disord. 2009, 24, 590–597. [Google Scholar] [CrossRef]
- Peppe, A.; Chiavalon, C.; Pasqualetti, P.; Crovato, D.; Caltagirone, C. Does gait analysis quantify motor rehabilitation efficacy in Parkinson's disease patients? Gait Posture 2007, 26, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.E.; Canning, C.G.; Sherrington, C.; Fung, V.S.C. Bradykinesia, muscle weakness and reduced muscle power in Parkinson's disease. Mov. Disord. 2009, 24, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.R.; Grimbergen, Y.A.M.; Cramer, M.; Willemsen, M.; Zwinderman, A.H. Prospective assessment of falls in Parkinson's disease. J. Neurol. 2001, 248, 950–958. [Google Scholar] [CrossRef] [PubMed]
- St George, R.J.; Nutt, J.G.; Burchiel, K.J.; Horak, F.B. A meta-regression of the long-term effects of deep brain stimulation on balance and gait in PD. Neurology 2010, 75, 1292–1299. [Google Scholar] [CrossRef]
- van Nuenen, B.F.L.; Esselink, R.A.J.; Munneke, M.; Speelman, J.D.; van Laar, T.; Bloem, B.R. Postoperative gait deterioration after bilateral subthalamic nucleus stimulation in Parkinson's disease. Mov. Disord. 2008, 23, 2404–2406. [Google Scholar] [CrossRef]
- Lewek, M.D.; Poole, R.; Johnson, J.; Halawa, O.; Huang, X. Arm swing magnitude and asymmetry during gait in the early stages of Parkinson's disease. Gait Posture 2010, 31, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Adkin, A.L.; Frank, J.S.; Jog, M.S. Fear of falling and postural control in Parkinson's disease. Mov. Disord. 2003, 18, 496–502. [Google Scholar] [CrossRef]
- Cakit, B.D.; Saracoglu, M.; Genc, H.; Erdem, H.R.; Inan, L. The effects of incremental speed-dependent treadmill training on postural instability and fear of falling in Parkinson's disease. Clinical Rehabilitation 2007, 21, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Nicolaas I Bohnen; Martijn L T M M ller; Vikas Kotagal; Robert A Koeppe; Michael R Kilbourn; Sid Gilman; Roger L Albin; Kirk A Frey. Heterogeneity of cholinergic denervation in Parkinson's disease without dementia.
- Malouin, F.; Richards, C.L.; Jackson, P.L.; Dumas, F.; Doyon, J. Brain activations during motor imagery of locomotor-related tasks: a PET study. Hum. Brain Mapp. 2003, 19, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Jahn, K.; Deutschländer, A.; Stephan, T.; Strupp, M.; Wiesmann, M.; Brandt, T. Brain activation patterns during imagined stance and locomotion in functional magnetic resonance imaging. Neuroimage 2004, 22, 1722–1731. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, M.; Hamani, C.; Hutchison, W.D.; Moro, E.; Lozano, A.M.; Dostrovsky, J.O. Pedunculopontine nucleus microelectrode recordings in movement disorder patients. Exp. Brain Res. 2008, 188, 165–174. [Google Scholar] [CrossRef]
- Lozano, A.M.; Snyder, B.J. Deep brain stimulation for parkinsonian gait disorders. J. Neurol. 2008, 255 Suppl 4, 30–31. [Google Scholar] [CrossRef]
- Cossu, G.; Pau, M. Subthalamic nucleus stimulation and gait in Parkinson's Disease: a not always fruitful relationship. Gait Posture 2017, 52, 205–210. [Google Scholar] [CrossRef]
- Morris, M.E.; Iansek, R.; Matyas, T.A.; Summers, J.J. The pathogenesis of gait hypokinesia in Parkinson's disease. Brain 1994, 117 Pt 5, 1169–1181. [Google Scholar] [CrossRef]
- Morris, M.E.; Iansek, R.; Matyas, T.A.; Summers, J.J. Ability to modulate walking cadence remains intact in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 1994, 57, 1532–1534. [Google Scholar] [CrossRef]
- Wu, T.; Hallett, M. A functional MRI study of automatic movements in patients with Parkinson's disease. Brain 2005, 128, 2250–2259. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.R.; Grimbergen, Y.A.M.; van Dijk, J.G.; Munneke, M. The "posture second" strategy: a review of wrong priorities in Parkinson's disease. Journal of the Neurological Sciences 2006, 248, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.E.; Iansek, R.; Matyas, T.A.; Summers, J.J. Stride length regulation in Parkinson's disease. Normalization strategies and underlying mechanisms. Brain 1996, 119 Pt 2, 551–568. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.Z.; Davies, L.; Stein, J.; France, S. The role of descending basal ganglia connections to the brain stem in parkinsonian akinesia. Br. J. Neurosurg. 1998, 12, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Brozova, H.; Barnaure, I.; Ruzicka, E.; Stochl, J.; Alterman, R.; Tagliati, M. Short- and Long-Term Effects of DBS on Gait in Parkinson's Disease. Front. Neurol. 2021, 12, 688760. [Google Scholar] [CrossRef] [PubMed]
- Vallabhajosula, S.; Haq, I.U.; Hwynn, N.; Oyama, G.; Okun, M.; Tillman, M.D.; Hass, C.J. Low-frequency Versus High-frequency Subthalamic Nucleus Deep Brain Stimulation on Postural Control and Gait in Parkinson's Disease: A Quantitative Study. Brain Stimul. 2015, 8, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Mera, T.O.; Filipkowski, D.E.; Riley, D.E.; Whitney, C.M.; Walter, B.L.; Gunzler, S.A.; Giuffrida, J.P. Quantitative analysis of gait and balance response to deep brain stimulation in Parkinson's disease. Gait Posture 2013, 38, 109–114. [Google Scholar] [CrossRef]
- Brozova, H.; Barnaure, I.; Alterman, R.L.; Tagliati, M. STN-DBS frequency effects on freezing of gait in advanced Parkinson disease. Neurology 2009, 72, 770; author reply 770-1. [Google Scholar] [CrossRef] [PubMed]
- Hausdorff, J.M.; Gruendlinger, L.; Scollins, L.; O'Herron, S.; Tarsy, D. Deep brain stimulation effects on gait variability in Parkinson's disease. Mov. Disord. 2009, 24, 1688–1692. [Google Scholar] [CrossRef]
- Crenna, P.; Carpinella, I.; Rabuffetti, M.; Rizzone, M.; Lopiano, L.; Lanotte, M.; Ferrarin, M. Impact of subthalamic nucleus stimulation on the initiation of gait in Parkinson's disease. Exp. Brain Res. 2006, 172, 519–532. [Google Scholar] [CrossRef]
- Moore, S.T.; Dilda, V.; Hakim, B.; MacDougall, H.G. Validation of 24-hour ambulatory gait assessment in Parkinson's disease with simultaneous video observation. Biomed. Eng. Online 2011, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Peppe, A.; Pierantozzi, M.; Chiavalon, C.; Marchetti, F.; Caltagirone, C.; Musicco, M.; Stanzione, P.; Stefani, A. Deep brain stimulation of the pedunculopontine tegmentum and subthalamic nucleus: Effects on gait in Parkinson's disease. Gait Posture 2010, 32, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Krack, P.; Benabid, A.L.; Pollak, P. Effect of bilateral subthalamic nucleus stimulation on parkinsonian gait. J. Neurol. 2001, 248, 1068–1072. [Google Scholar] [CrossRef]
- Ferrarin, M.; Rizzone, M.; Lopiano, L.; Recalcati, M.; Pedotti, A. Effects of subthalamic nucleus stimulation and l-dopa in trunk kinematics of patients with Parkinson's disease. Gait Posture 2004, 19, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Ferrarin, M.; Rizzone, M.; Bergamasco, B.; Lanotte, M.; Recalcati, M.; Pedotti, A.; Lopiano, L. Effects of bilateral subthalamic stimulation on gait kinematics and kinetics in Parkinson?: S disease. Exp Brain Res 2005, 160, 517–527. [Google Scholar] [CrossRef]
- Bleuse, S.; Delval, A.; Blatt, J.L.; Derambure, P.; Destée, A.; Defebvre, L. Effect of bilateral subthalamic nucleus deep brain stimulation on postural adjustments during arm movement. Clin. Neurophysiol. 2011, 122, 2032–2035. [Google Scholar] [CrossRef]
- Carpinella, I.; Crenna, P.; Marzegan, A.; Rabuffetti, M.; Rizzone, M.; Lopiano, L.; Ferrarin, M. Effect of L-dopa and subthalamic nucleus stimulation on arm and leg swing during gait in Parkinson's Disease. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2007, 2007, 6665–6668. [Google Scholar] [CrossRef] [PubMed]
- St George, R.J.; Carlson-Kuhta, P.; King, L.A.; Burchiel, K.J.; Horak, F.B. Compensatory stepping in Parkinson's disease is still a problem after deep brain stimulation randomized to STN or GPi. J. Neurophysiol. 2015, 114, 1417–1423. [Google Scholar] [CrossRef]
- Weaver, F.M.; Follett, K.A.; Stern, M.; Luo, P.; Harris, C.L.; Hur, K.; Marks, W.J.; Rothlind, J.; Sagher, O.; Moy, C.; et al. Randomized trial of deep brain stimulation for Parkinson disease: thirty-six-month outcomes. Neurology 2012, 79, 55–65. [Google Scholar] [CrossRef]
- Rizzone, M.; Ferrarin, M.; Pedotti, A.; Bergamasco, B.; Bosticco, E.; Lanotte, M.; Perozzo, P.; Tavella, A.; Torre, E.; Recalcati, M.; et al. High-frequency electrical stimulation of the subthalamic nucleus in Parkinson's disease: kinetic and kinematic gait analysis. Neurol. Sci. 2002, 23 (Suppl. S2), S103–4. [Google Scholar] [CrossRef]
- Krystkowiak, P.; Blatt, J.-L.; Bourriez, J.-L.; Duhamel, A.; Perina, M.; Blond, S.; Guieu, J.-D.; Destée, A.; Defebvre, L. Effects of Subthalamic Nucleus Stimulation and Levodopa Treatment on Gait Abnormalities in Parkinson Disease. Arch Neurol 2003, 60, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.; Defebvre, L.; Destée, A.; Bleuse, S.; Clement, F.; Blatt, J.L.; Krystkowiak, P.; Devos, D. STN-DBS frequency effects on freezing of gait in advanced Parkinson disease. Neurology 2008, 71, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Wagle Shukla, A.; Moro, E.; Gunraj, C.; Lozano, A.; Hodaie, M.; Lang, A.; Chen, R. Long-term subthalamic nucleus stimulation improves sensorimotor integration and proprioception. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Sailer, A.; Molnar, G.F.; Paradiso, G.; Gunraj, C.A.; Lang, A.E.; Chen, R. Short and long latency afferent inhibition in Parkinson's disease. Brain 2003, 126, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- Knight, E.J.; Testini, P.; Min, H.-K.; Gibson, W.S.; Gorny, K.R.; Favazza, C.P.; Felmlee, J.P.; Kim, I.; Welker, K.M.; Clayton, D.A.; et al. Motor and Nonmotor Circuitry Activation Induced by Subthalamic Nucleus Deep Brain Stimulation in Patients With Parkinson Disease: Intraoperative Functional Magnetic Resonance Imaging for Deep Brain Stimulation. Mayo Clinic Proceedings 2015, 90, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Vitek, J.L. Deep brain stimulation: how does it work? Cleve. Clin. J. Med. 2008, 75 (Suppl. S2), S59–65. [Google Scholar] [CrossRef]
- Hamel, W.; Fietzek, U.; Morsnowski, A.; Schrader, B.; Herzog, J.; Weinert, D.; Pfister, G.; Müller, D.; Volkmann, J.; Deuschl, G.; et al. Deep brain stimulation of the subthalamic nucleus in Parkinson's disease: evaluation of active electrode contacts. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Vitek, J.L. Mechanisms of deep brain stimulation: Excitation or inhibition. Mov Disord. 2002, 17, S69–S72. [Google Scholar] [CrossRef] [PubMed]
- Santaniello, S.; McCarthy, M.M.; Montgomery, E.B.; Gale, J.T.; Kopell, N.; Sarma, S.V. Therapeutic mechanisms of high-frequency stimulation in Parkinson's disease and neural restoration via loop-based reinforcement. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, E586–95. [Google Scholar] [CrossRef]
- Shine, J.M.; Matar, E.; Ward, P.B.; Bolitho, S.J.; Gilat, M.; Pearson, M.; Naismith, S.L.; Lewis, S.J.G. Exploring the cortical and subcortical functional magnetic resonance imaging changes associated with freezing in Parkinson's disease. Brain 2013, 136, 1204–1215. [Google Scholar] [CrossRef]
- Deniau, J.-M.; Degos, B.; Bosch, C.; Maurice, N. Deep brain stimulation mechanisms: beyond the concept of local functional inhibition. Eur. J. Neurosci. 2010, 32, 1080–1091. [Google Scholar] [CrossRef]
- Arai, N.; Yokochi, F.; Ohnishi, T.; Momose, T.; Okiyama, R.; Taniguchi, M.; Takahashi, H.; Matsuda, H.; Ugawa, Y. Mechanisms of unilateral STN-DBS in patients with Parkinson's disease: A PET study. J. Neurol. 2008, 255, 1236–1243. [Google Scholar] [CrossRef]
- Geday, J.; Østergaard, K.; Johnsen, E.; Gjedde, A. STN-stimulation in Parkinson's disease restores striatal inhibition of thalamocortical projection. Hum. Brain Mapp. 2009, 30, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Stefani, A.; Lozano, A.M.; Peppe, A.; Stanzione, P.; Galati, S.; Tropepi, D.; Pierantozzi, M.; Brusa, L.; Scarnati, E.; Mazzone, P. Bilateral deep brain stimulation of the pedunculopontine and subthalamic nuclei in severe Parkinson's disease. Brain 2007, 130, 1596–1607. [Google Scholar] [CrossRef]
- Florio, T.; Scarnati, E.; Confalone, G.; Minchella, D.; Galati, S.; Stanzione, P.; Stefani, A.; Mazzone, P. High-frequency stimulation of the subthalamic nucleus modulates the activity of pedunculopontine neurons through direct activation of excitatory fibres as well as through indirect activation of inhibitory pallidal fibres in the rat. Eur. J. Neurosci. 2007, 25, 1174–1186. [Google Scholar] [CrossRef] [PubMed]
- Hanakawa, T.; Katsumi, Y.; Fukuyama, H.; Honda, M.; Hayashi, T.; Kimura, J.; Shibasaki, H. Mechanisms underlying gait disturbance in Parkinson's disease: a single photon emission computed tomography study. Brain 1999, 122 Pt 7, 1271–1282. [Google Scholar] [CrossRef]
- Limousin, P.; Greene, J.; Pollak, P.; Rothwell, J.; Benabid, A.L.; Frackowiak, R. Changes in cerebral activity pattern due to subthalamic nucleus or internal pallidum stimulation in Parkinson's disease. Ann. Neurol. 1997, 42, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, L.; Pecoraro, P.M.; Carbone, S.P.; Caminiti, M.L.; Di Lazzaro, V. Levodopa-Induced Dyskinesias in Parkinson’s Disease: An Overview on Pathophysiology, Clinical Manifestations, Therapy Management Strategies and Future Directions. JCM 2023, 12, 4427. [Google Scholar] [CrossRef] [PubMed]
- Rizzone, M.G.; Fasano, A.; Daniele, A.; Zibetti, M.; Merola, A.; Rizzi, L.; Piano, C.; Piccininni, C.; Romito, L.M.; Lopiano, L.; et al. Long-term outcome of subthalamic nucleus DBS in Parkinson's disease: from the advanced phase towards the late stage of the disease? Parkinsonism & Related Disorders 2014, 20, 376–381. [Google Scholar] [CrossRef]
- Ciampi de Andrade, D.; Lefaucheur, J.-P.; Galhardoni, R.; Ferreira, K.S.L.; Brandao Paiva, A.R.; Bor-Seng-Shu, E.; Alvarenga, L.; Myczkowski, M.L.; Marcolin, M.A.; de Siqueira, Silvia R D T; et al. Subthalamic deep brain stimulation modulates small fiber-dependent sensory thresholds in Parkinson's disease. Pain 2012, 153, 1107–1113. [CrossRef]
- Kerr, G.K.; Worringham, C.J.; Cole, M.H.; Lacherez, P.F.; Wood, J.M.; Silburn, P.A. Predictors of future falls in Parkinson disease. Neurology 2010, 75, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Dickson, D.W.; Uchikado, H.; Fujishiro, H.; Tsuboi, Y. Evidence in favor of Braak staging of Parkinson's disease. Mov. Disord. 2010, 25 (Suppl. S1), S78–82. [Google Scholar] [CrossRef]
- Kingsbury, A.E.; Bandopadhyay, R.; Silveira-Moriyama, L.; Ayling, H.; Kallis, C.; Sterlacci, W.; Maeir, H.; Poewe, W.; Lees, A.J. Brain stem pathology in Parkinson's disease: an evaluation of the Braak staging model. Mov. Disord. 2010, 25, 2508–2515. [Google Scholar] [CrossRef] [PubMed]
- Wolters, E.C.; Braak, H. Parkinson's disease: premotor clinico-pathological correlations. J. Neural Transm. Suppl. 2006, 309–319. [Google Scholar] [CrossRef]
- Nakanishi, T.; Tamaki, M.; Mizusawa, H.; Akatsuka, T.; Kinoshita, T. An experimental study for analyzing nerve conduction velocity. Electroencephalogr. Clin. Neurophysiol. 1986, 63, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Oroz, M.C.; Rodriguez, M.; Guridi, J.; Mewes, K.; Chockkman, V.; Vitek, J.; DeLong, M.R.; Obeso, J.A. The subthalamic nucleus in Parkinson's disease: somatotopic organization and physiological characteristics. Brain 2001, 124, 1777–1790. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, P.; Bronte-Stewart, H.; Heit, G.; Schaal, D.W.; Esposito, V. The functional organization of the sensorimotor region of the subthalamic nucleus. Stereotact. Funct. Neurosurg. 2004, 82, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Calvert, G.A.; Thesen, T. Multisensory integration: methodological approaches and emerging principles in the human brain. J. Physiol. Paris 2004, 98, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Kühn, A.A.; Kupsch, A.; Schneider, G.-H.; Brown, P. Reduction in subthalamic 8-35 Hz oscillatory activity correlates with clinical improvement in Parkinson's disease. Eur. J. Neurosci. 2006, 23, 1956–1960. [Google Scholar] [CrossRef]
- Kühn, A.A.; Williams, D.; Kupsch, A.; Limousin, P.; Hariz, M.; Schneider, G.-H.; Yarrow, K.; Brown, P. Event-related beta desynchronization in human subthalamic nucleus correlates with motor performance. Brain 2004, 127, 735–746. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Bratzke, H.; Hamm-Clement, J.; Sandmann-Keil, D.; Rüb, U. Staging of the intracerebral inclusion body pathology associated with idiopathic Parkinson's disease (preclinical and clinical stages). J. Neurol. 2002, 249 (Suppl. S3), II/1-5. [Google Scholar] [CrossRef]
- Riederer, P.; Sian-Hülsmann, J. The significance of neuronal lateralisation in Parkinson's disease. J. Neural Transm. (Vienna) 2012, 119, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Silverdale, M.A.; Kobylecki, C.; Kass-Iliyya, L.; Martinez-Martin, P.; Lawton, M.; Cotterill, S.; Chaudhuri, K.R.; Morris, H.; Baig, F.; Williams, N.; et al. A detailed clinical study of pain in 1957 participants with early/moderate Parkinson's disease. Parkinsonism & Related Disorders 2018, 56, 27–32. [Google Scholar] [CrossRef]
- K. Sathian; A. Zangaladze; J. Green; J. L. Vitek; M. R. DeLong. Tactile spatial acuity and roughness discrimination: Impairments due to aging and Parkinson's disease. Neurology 1997, 49, 168. [CrossRef]
- O'Suilleabhain, P.; Bullard, J.; Dewey, R.B. Proprioception in Parkinson's disease is acutely depressed by dopaminergic medications. J. Neurol. Neurosurg. Psychiatry 2001, 71, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Comi, C.; Oggioni, G.D.; Pacchetti, C.; Calandrella, D.; Moja, M.C.; Riboldazzi, G.; Tunesi, S.; Fante, M.D.; Manfredi, L.; et al. Prevalence and features of peripheral neuropathy in Parkinson’s disease patients under different therapeutic regimens. Parkinsonism & Related Disorders 2014, 20, 27–31. [Google Scholar] [CrossRef]
- Dellapina, E.; Ory-Magne, F.; Regragui, W.; Thalamas, C.; Lazorthes, Y.; Rascol, O.; Payoux, P.; Brefel-Courbon, C. Effect of subthalamic deep brain stimulation on pain in Parkinson's disease. Pain 2012, 153, 2267–2273. [Google Scholar] [CrossRef] [PubMed]
- Maschke, M.; Tuite, P.J.; Pickett, K.; Wächter, T.; Konczak, J. The effect of subthalamic nucleus stimulation on kinaesthesia in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 2005, 76, 569–571. [Google Scholar] [CrossRef]
- Ortega-Cubero, S.; Clavero, P.; Irurzun, C.; Gonzalez-Redondo, R.; Guridi, J.; Obeso, J.A.; Rodriguez-Oroz, M.C. Effect of deep brain stimulation of the subthalamic nucleus on non-motor fluctuations in Parkinson's disease: Two-year' follow-up. Parkinsonism & Related Disorders 2013, 19, 543–547. [Google Scholar] [CrossRef]
- Reich, M.M.; Ray Chaudhuri, K.; Ashkan, K.; Hulse, N.; Costello, A.; Moriarty, J.; Samuel, M. Changes in the non-motor symptom scale in Parkinson’s disease after deep brain stimulation. Basal Ganglia 2011, 1, 131–133. [Google Scholar] [CrossRef]
- Witjas, T.; Kaphan, E.; Régis, J.; Jouve, E.; Chérif, A.A.; Péragut, J.-C.; Azulay, J.P. Effects of chronic subthalamic stimulation on nonmotor fluctuations in Parkinson's disease. Mov. Disord. 2007, 22, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Taktakishvili, O.; Sivan-Loukianova, E.; Kultas-Ilinsky, K.; Ilinsky, I.A. Posterior parietal cortex projections to the ventral lateral and some association thalamic nuclei in Macaca mulatta. Brain Res. Bull. 2002, 59, 135–150. [Google Scholar] [CrossRef]
- Karen D., Davis; Chun L., Kwan; Adrian P., Crawley; and David, J. Mikulis. Functional MRI Study of Thalamic and Cortical Activations Evoked by Cutaneous Heat, Cold, and Tactile Stimuli.
- Hilker, R.; Voges, J.; Weisenbach, S.; Kalbe, E.; Burghaus, L.; Ghaemi, M.; Lehrke, R.; Koulousakis, A.; Herholz, K.; Sturm, V.; et al. Subthalamic nucleus stimulation restores glucose metabolism in associative and limbic cortices and in cerebellum: evidence from a FDG-PET study in advanced Parkinson's disease. J. Cereb. Blood Flow Metab. 2004, 24, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Trost, M.; Su, S.; Su, P.; Yen, R.-F.; Tseng, H.-M.; Barnes, A.; Ma, Y.; Eidelberg, D. Network modulation by the subthalamic nucleus in the treatment of Parkinson's disease. Neuroimage 2006, 31, 301–307. [Google Scholar] [CrossRef]
- Oswal, A.; Beudel, M.; Zrinzo, L.; Limousin, P.; Hariz, M.; Foltynie, T.; Litvak, V.; Brown, P. Deep brain stimulation modulates synchrony within spatially and spectrally distinct resting state networks in Parkinson's disease. Brain 2016, 139, 1482–1496. [Google Scholar] [CrossRef] [PubMed]
- Herzog, J.; Weiss, P.H.; Assmus, A.; Wefer, B.; Seif, C.; Braun, P.M.; Pinsker, M.O.; Herzog, H.; Volkmann, J.; Deuschl, G.; et al. Improved sensory gating of urinary bladder afferents in Parkinson's disease following subthalamic stimulation. Brain 2008, 131, 132–145. [Google Scholar] [CrossRef]
- DiMarzio, M.; Rashid, T.; Hancu, I.; Fiveland, E.; Prusik, J.; Gillogly, M.; Madhavan, R.; Joel, S.; Durphy, J.; Molho, E.; et al. Functional MRI Signature of Chronic Pain Relief From Deep Brain Stimulation in Parkinson Disease Patients. Neurosurgery 2019, 85, E1043–E1049. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, I.; St George, R.J.; Kalliolia, E.; Peters, A.L.; Limousin, P.; Day, B.L. Maintaining balance against force perturbations: impaired mechanisms unresponsive to levodopa in Parkinson's disease. J. Neurophysiol. 2016, 116, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.J.; Hirschmann, J.; Vesper, J.; Wojtecki, L.; Butz, M.; Schnitzler, A. Distinct cortical responses evoked by electrical stimulation of the thalamic ventral intermediate nucleus and of the subthalamic nucleus. Neuroimage Clin. 2018, 20, 1246–1254. [Google Scholar] [CrossRef]
- Garcia, L.; D'Alessandro, G.; Bioulac, B.; Hammond, C. High-frequency stimulation in Parkinson's disease: more or less? Trends in Neurosciences 2005, 28, 209–216. [Google Scholar] [CrossRef]
- Sailer, A.; Molnar, G.F.; Paradiso, G.; Gunraj, C.A.; Lang, A.E.; Chen, R. Short and long latency afferent inhibition in Parkinson's disease. Brain 2003, 126, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.E.; Terman, D. High frequency stimulation of the subthalamic nucleus eliminates pathological thalamic rhythmicity in a computational model. J. Comput. Neurosci. 2004, 16, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.; Oliviero, A.; Mazzone, P.; Insola, A.; Tonali, P.; Di Lazzaro, V. Dopamine dependency of oscillations between subthalamic nucleus and pallidum in Parkinson's disease. J. Neurosci. 2001, 21, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Rodríguez-Oroz, M.C.; Benitez-Temino, B.; Blesa, F.J.; Guridi, J.; Marin, C.; Rodriguez, M. Functional organization of the basal ganglia: therapeutic implications for Parkinson's disease. Mov. Disord. 2008, 23 Suppl 3, S548–59. [Google Scholar] [CrossRef]
- Sudhyadhom, A.; McGregor, K.; Okun, M.S.; Foote, K.D.; Trinastic, J.; Crosson, B.; Bova, F.J. Delineation of motor and somatosensory thalamic subregions utilizing probabilistic diffusion tractography and electrophysiology. J. Magn. Reson. Imaging 2013, 37, 600–609. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Testing subjects motor performance on the pressure distribution platform: A) Quiescent bipedal stance; B) Functional limits of stability - the red dot indicates the location of the center of pressure (COP); C) Gait.
Figure 1.
Testing subjects motor performance on the pressure distribution platform: A) Quiescent bipedal stance; B) Functional limits of stability - the red dot indicates the location of the center of pressure (COP); C) Gait.
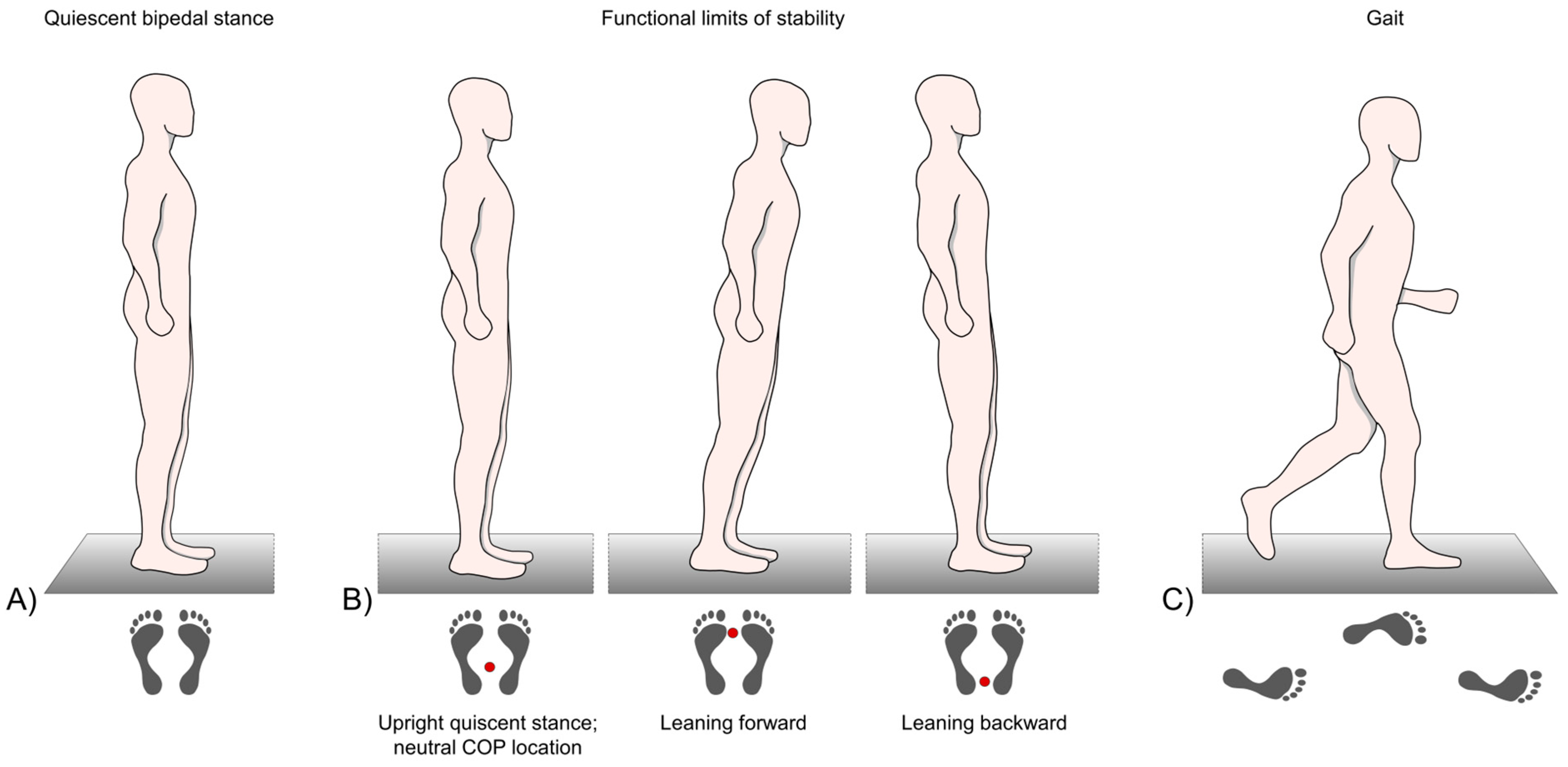
Figure 2.
Examination of the plantar cutaneous vibration perception threshold (VPT) at the first metatarsal head.
Figure 2.
Examination of the plantar cutaneous vibration perception threshold (VPT) at the first metatarsal head.
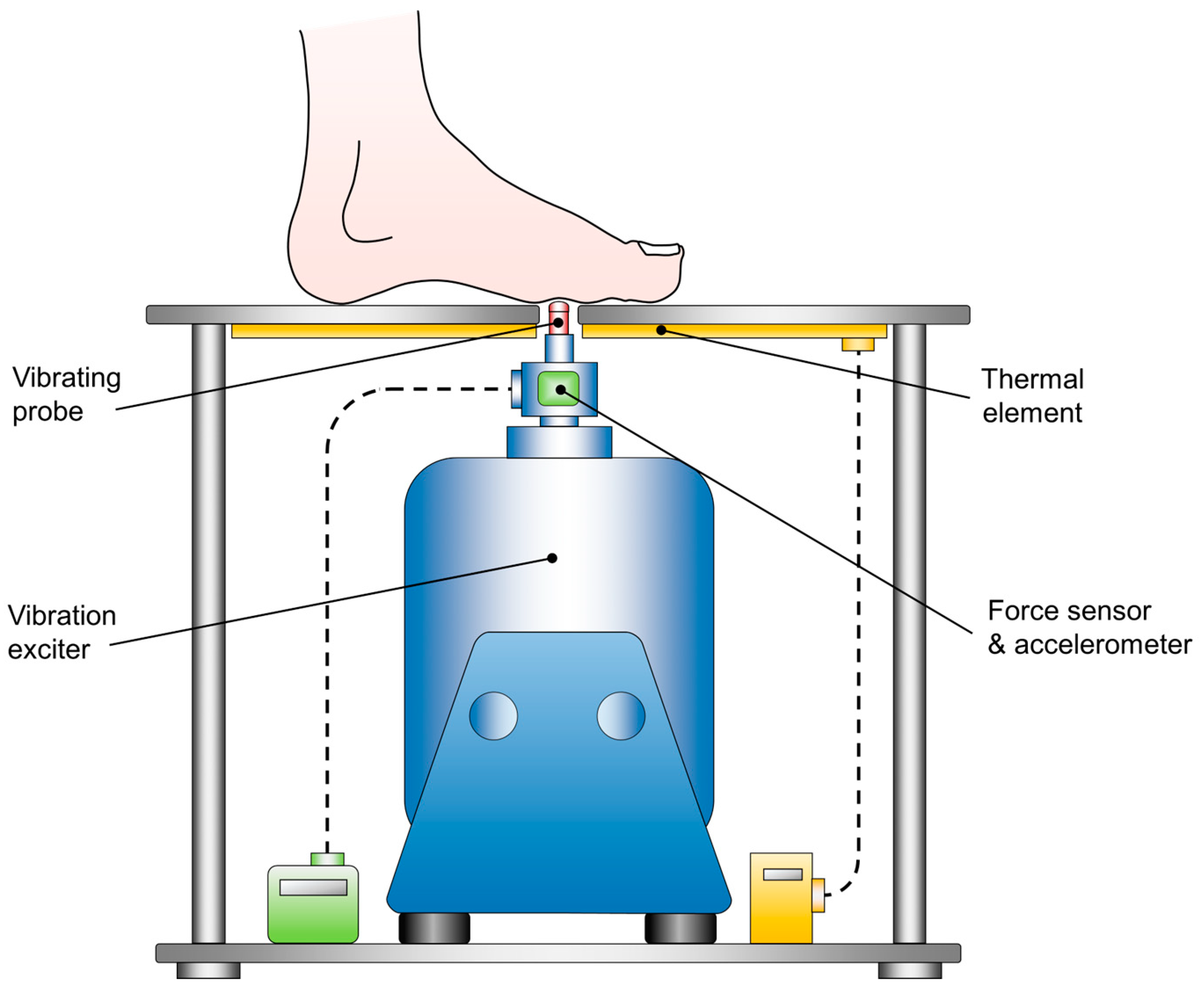
Figure 3.
Illustration of an actual COP displacement signal from a random patient in study group PD-MED–DBS, who performed the functional limits of stability test. The figure indicates the detection of the maximal COP displacement and the motion time in the anterior and posterior directions.
Figure 3.
Illustration of an actual COP displacement signal from a random patient in study group PD-MED–DBS, who performed the functional limits of stability test. The figure indicates the detection of the maximal COP displacement and the motion time in the anterior and posterior directions.
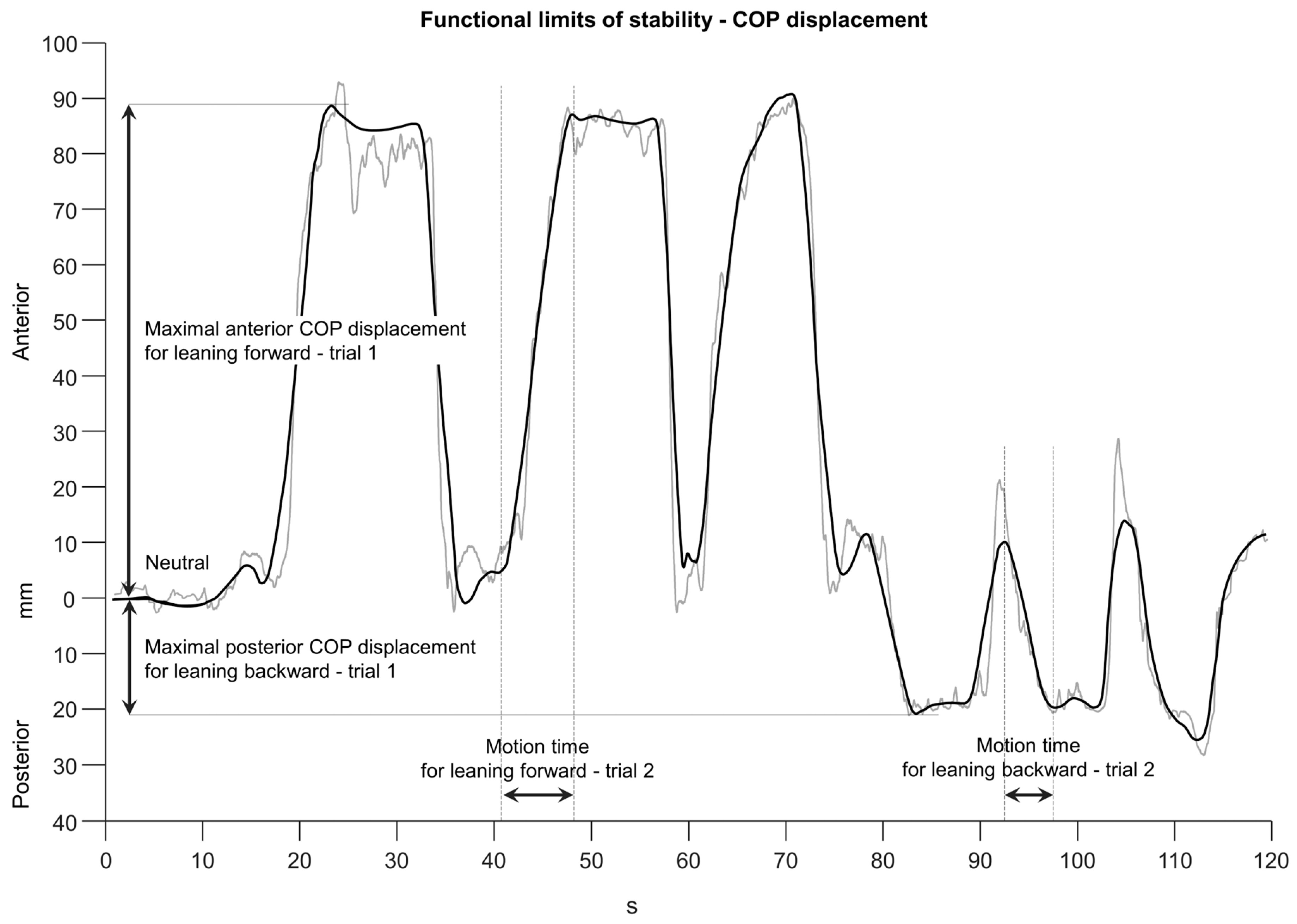
Figure 4.
Group comparisons for COP parameters: A) COP 95 % confidence area B) COP range AP; C) COP range ML; D) COP max velocity total; E) COP max velocity AP; F) COP max velocity ML. The cross within each box marks the mean value. Statistically significant differences between groups (p < 0.0028) are indicated, as well as effect sizes, r.
Figure 4.
Group comparisons for COP parameters: A) COP 95 % confidence area B) COP range AP; C) COP range ML; D) COP max velocity total; E) COP max velocity AP; F) COP max velocity ML. The cross within each box marks the mean value. Statistically significant differences between groups (p < 0.0028) are indicated, as well as effect sizes, r.
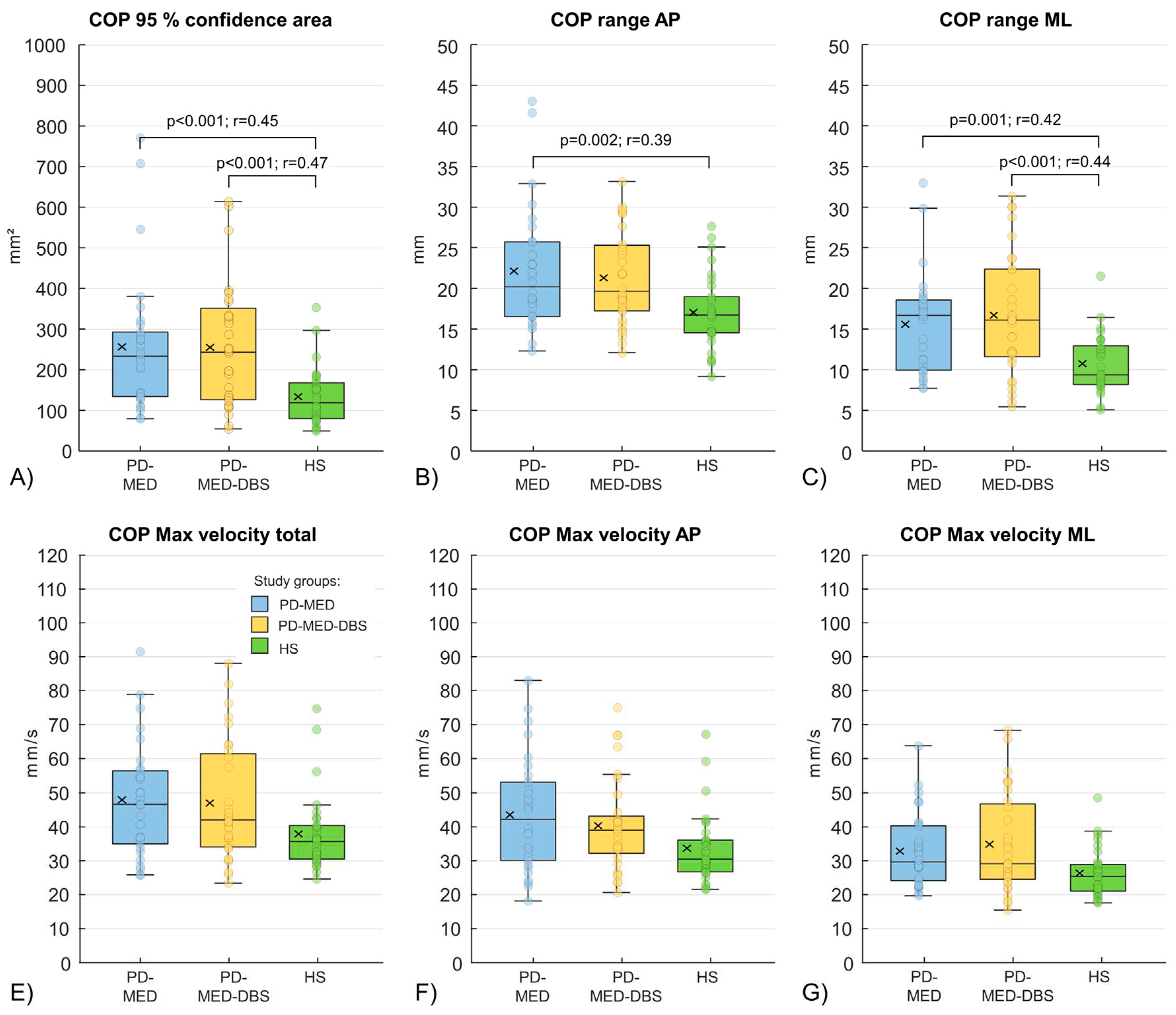
Figure 5.
Group comparisons for the maximal COP displacements in the anterior and posterior directions and the total COP range during the functional limits of stability test. Values are shown as percentage of the foot length (Mean + SD). Statistically significant differences (p < 0.0028) are shown, as well as effect sizes, r.
Figure 5.
Group comparisons for the maximal COP displacements in the anterior and posterior directions and the total COP range during the functional limits of stability test. Values are shown as percentage of the foot length (Mean + SD). Statistically significant differences (p < 0.0028) are shown, as well as effect sizes, r.
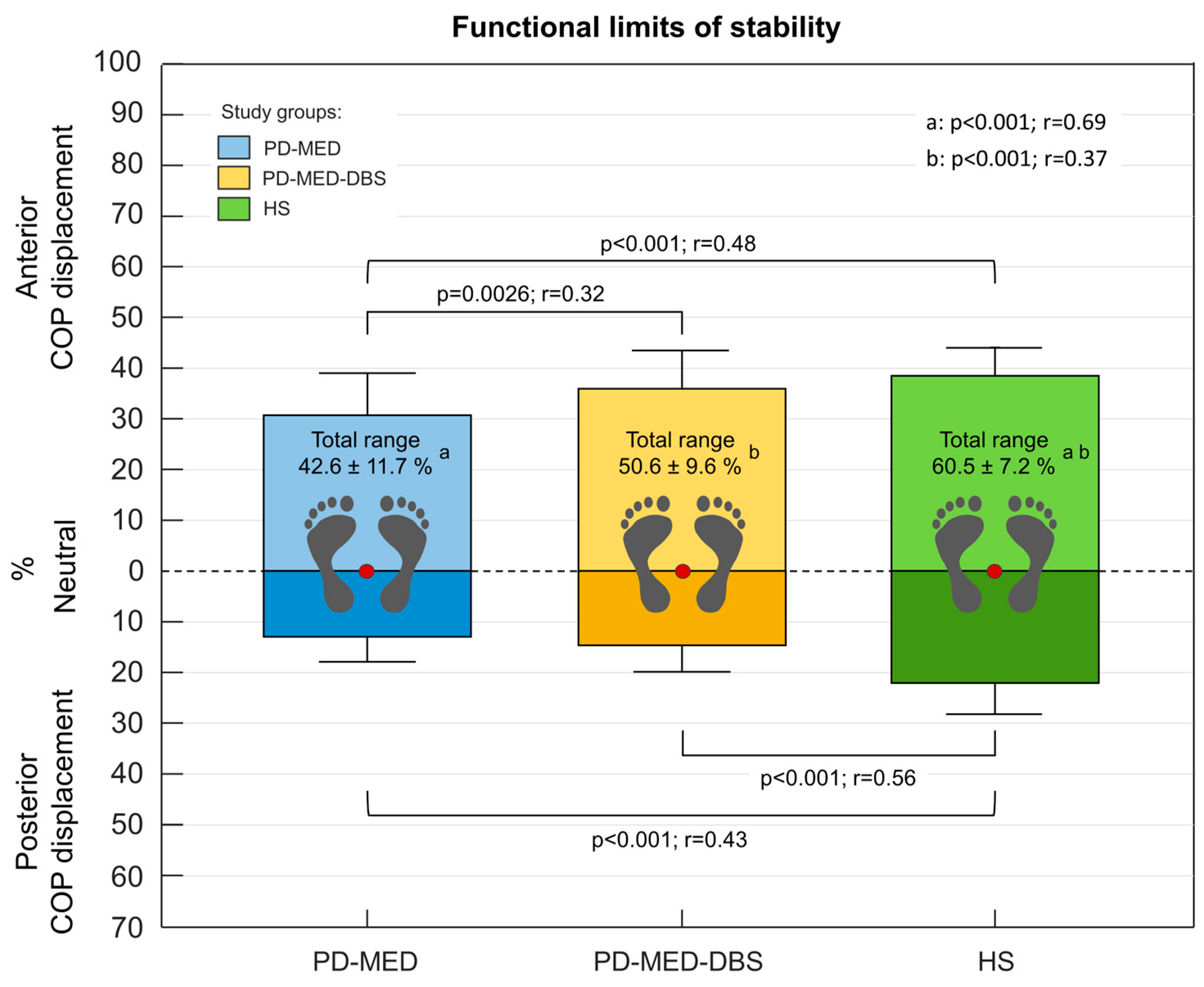
Figure 6.
Group comparisons of the VPTs from the left and right first metatarsal head of the plantar foot. Displayed are the untransformed absolute VPTs. The cross within each box marks the mean value. Statistically significant differences (p < 0.0083) are shown as well as effect sizes, r.
Figure 6.
Group comparisons of the VPTs from the left and right first metatarsal head of the plantar foot. Displayed are the untransformed absolute VPTs. The cross within each box marks the mean value. Statistically significant differences (p < 0.0083) are shown as well as effect sizes, r.
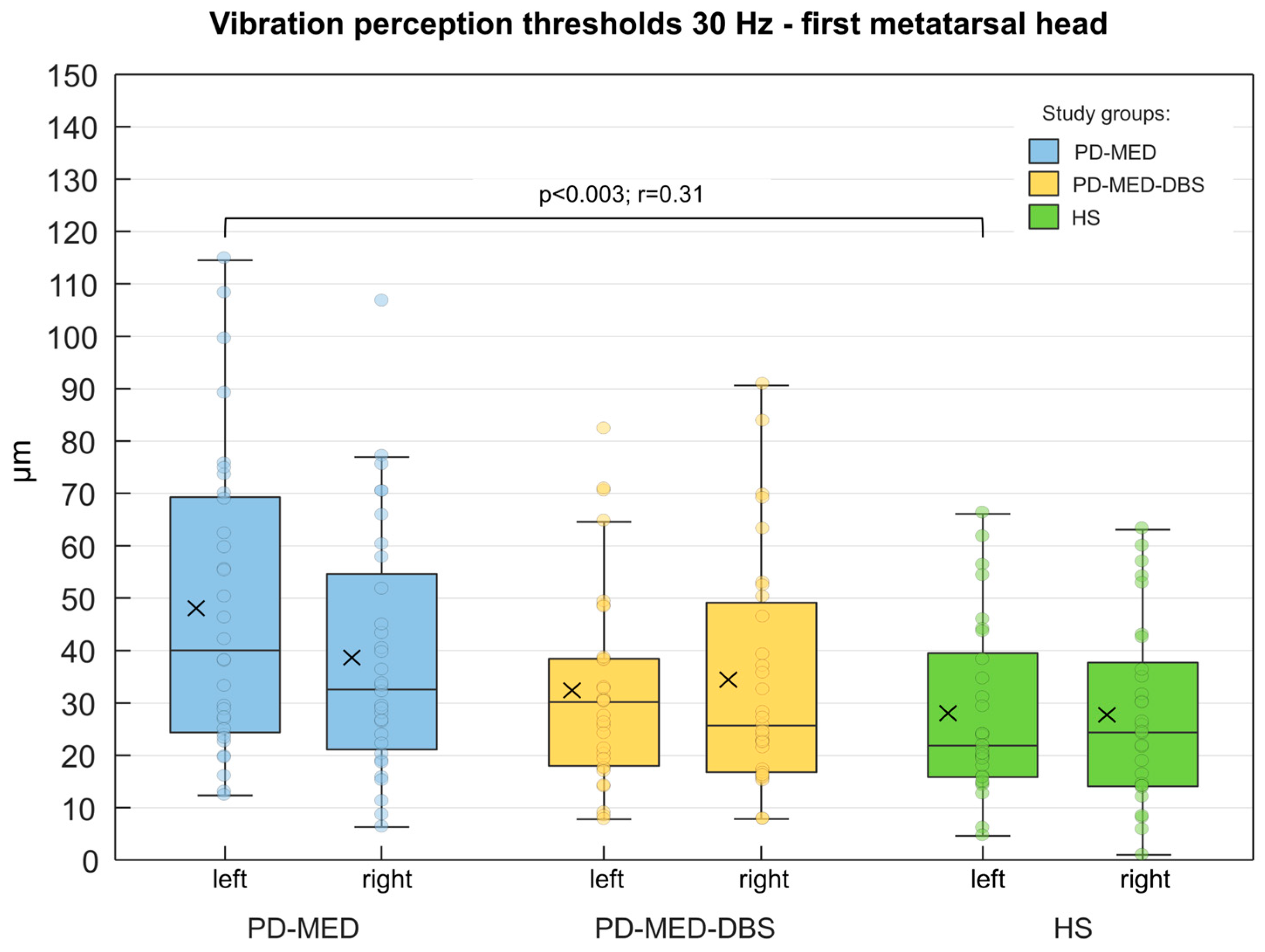

Table 1.
Demographic and clinical data (mean ± SD); statistically significant differences for PD-MED vs. PD-MED–DBS vs. HS, p < 0.0167 and PD-MED vs. PD-MED–DBS, p < 0.05 are marked with a and b; r-values indicate the effect sizes of statistically significant group differences.
Table 1.
Demographic and clinical data (mean ± SD); statistically significant differences for PD-MED vs. PD-MED–DBS vs. HS, p < 0.0167 and PD-MED vs. PD-MED–DBS, p < 0.05 are marked with a and b; r-values indicate the effect sizes of statistically significant group differences.
| PD-MED | PD-MED-DBS | HS | p-values; r | ||
|---|---|---|---|---|---|
| Demographic data: |
n/gender | 37 / ♂ 29 / ♀ 8 | 39 / ♂ 24 / ♀ 15 | 30 / ♂ 19 / ♀ 11 | – |
| age (years) | 68.5 ± 7.5 | 66.2 ± 6.5 b | 70.6 ± 5.7 b | b 0.023; 0.13 | |
| height (cm) | 169.6 ± 13.6 | 168.7 ± 8.1 | 171.3 ± 8.9 | – | |
| clinical data: |
self-rated balance confidence (0–100) (%) | 60.0 ± 20.2 a | 59.8 ± 20.1 b | 80.7 ± 9.4 a b |
a <0.001; 0.50 b <0.001; 0.43 |
| self-rated gait confidence (0–100) (%) | 64.0 ± 22.5 a | 63.3 ± 18.3 b | 89.1 ± 8.4 a b |
a <0.001; 0.55 b <0.001; 0.54 |
|
| MMSE (0–30) | 28.5 ± 1.7 | 28.4 ± 1.6 | – | – | |
| UPDRS III (0–108) | 16.9 ± 7.9 | 15.7 ± 6.6 | – | – | |
| UPDRS total (0–199) | 27.8 ± 13.2 | 29.2 ± 10.7 | – | – | |
| Hoehn and Yahr (0–5) | 2.1 ± 0.5 | 2.1 ± 0.4 | – | – | |
| disease duration since diagnosis (months) | 85.1 ± 65.7 | 189.2 ± 77.6 | – | – | |
| disease-dominant body side | left: 17; right: 20 | left: 15; right: 24 | – | – | |
| time btw. last neurological examination and testings (months) | 5.7 ± 15.2 a | 3.5 ± 3.9 a | – | a < 0.001; 0.66 | |
| DBS duration since surgery (months) | 33.1 ± 25.7 | – | – | ||
| self-rated satisfaction with DBS (%) | 80.1 ± 20.6 | – | – |
Table 2.
Group comparisons for the temporal and spatial-temporal COP parameters of the functional limits of stability test. Statistically significant differences (p < 0.0028) are shown (a, b) as well as effect sizes, r.
Table 2.
Group comparisons for the temporal and spatial-temporal COP parameters of the functional limits of stability test. Statistically significant differences (p < 0.0028) are shown (a, b) as well as effect sizes, r.
| PD-MED | PD-MED-DBS | HS | p-values; r | |
|---|---|---|---|---|
| COP motion time anterior (s) | 7.1 ± 2.6 a | 5.3 ± 2.1b | 3.0 ± 1.1 a b | a <0.001; 0.68; b 0.002; 0,43 |
| COP motion time posterior (s) | 6.3 ± 2.5a | 4.7 ± 1.7 | 3.2 ± 1.4a | a <0.001; 0.56 |
| COP mean motion velocity anterior (mm/s) | 12.9 ± 7.9a | 19.9 ± 10.9b | 37.3 ± 14.0a b | a <0.001; 0.71; b 0.002; 0,43 |
| COP mean motion velocity posterior (mm/s) | 6.2 ± 3.7a | 8.3 ± 3.2b | 21.4 ± 11.4a b | a <0.001; 0.72; b <0.001; 0,49 |
Table 3.
Group comparisons of the gait parameters. Statistically significant differences (p < 0.0023) are shown (a, b) as well as effect sizes, r.
Table 3.
Group comparisons of the gait parameters. Statistically significant differences (p < 0.0023) are shown (a, b) as well as effect sizes, r.
| PD-MED | PD-MED-DBS | HS | p-values; r | |
|---|---|---|---|---|
| Stride length (cm) | 91.1 ± 18.0 a | 94.3 ± 12.8 b | 118.3 ± 16.1 a b | a <0.001; 0.56; b <0.001; 0,52 |
| Stride width (cm) | 10.7 ± 3.1 | 11.1 ± 3.0b | 8.3 ± 1.8 b | – b <0.001; 0,29 |
| Stride time (s) | 1.2 ± 0.2 a | 1.2 ± 0.1 | 1.1 ± 0.1 a | a <0.001; 0.39; – |
| Gait velocity (m/s) | 0.8 ± 0.2 a | 0.8 ± 0.2b | 1.1 ± 0.2 a b | a <0.001; 0.60; b <0.001; 0,49 |
| Cadence (steps/minute) | 49.2 ± 6.0 a | 52.4 ± 5.3 | 55.4 ± 4.6a | a <0.001; 0.39; – |
| Double support time (%) | 35.3 ± 7.1 a | 32.7 ± 4.6b | 28.4 ± 3.7 a b | a <0.001; 0.47; b <0.001; 0,35 |
| COP length mean left right (%) | 84.5 ± 6.4 a | 85.8 ± 7.3 | 89.9 ± 6.2 a | a 0.002; 0.31; – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



