
Submitted:
08 November 2023
Posted:
09 November 2023
You are already at the latest version
Abstract
Zika Virus (ZIKV) is a pathogenic member of the flavivirus family, with several unique charac-teristics. Unlike any other arbovirus, ZIKV can be transmitted sexually and maternally, and thus produce congenital syndromes (CZS) due to its neurotropism. This challenges the search for safe active molecules that can protect pregnant women and their fetuses. In this context, and in the ab-sence of any existing treatment, it seemed worthwhile to test whether the known cytoprotective properties of adiponectin and its pharmacological analog, AdipoRon, could influence the outcome of ZIKV infection. We showed that both AdipoRon and adiponectin could significantly reduce in vitro infection of A549 epithelial cells, a well-known cell model for flaviviruses infection studies. This effect was particularly observed when a pre-treatment was carried out. Conversely, ZIKV revealed an ability to down-regulate adiponectin receptor expression and thereby limit adi-ponectin signaling.
Keywords:
Zika virus
; antiviral
; AdipoRon
; adiponectin agonist
; drug repositioning
; metabolism
1. Introduction
Zika virus (ZIKV) is a mosquito-borne virus which, on the eve of the global crisis caused by SARS-CoV-2, had already alerted the international scientific community. The awareness was due to an unanticipated emergence, an ability to spread rapidly worldwide and, above all, a capacity to cause severe and previously unknown neurological diseases in newborns[1,2]. ZIKV at the time of its re-emergence and successive epidemics, firstly in Micronesia in 2007, French Polynesia in 2013, followed by Brazil in 2015, was not totally unknown[3]. This flavivirus is a member of the Flaviviridae family that was first isolated in a rhesus monkey from the Zika rainforest in Uganda in 1947, indicating the existence of an African and an Asian lineage[4,5]. Both are mainly transmitted by mosquitoes of the genus Aedes. While the historical African strain of ZIKV seemed to cause only discrete and sporadic cases, the Asian strain has suddenly emerged as a threat. This urged the World Health Organization (WHO) to declare this virus as a major health issue in 2015. Among flaviviruses, several are of medical concern, including Dengue Virus (DENV), Japanese Encephalitis Virus (JEV), and West Nile Virus (WNV). As with the latter, ZIKV infection often goes asymptomatic in approximately 80% of cases and induces only mild symptoms in most patients. Unfortunately, contemporary epidemics revealed that ZIKV could also induce severe neurological diseases like Guillain-Barré syndrome in adults and congenital ZIKA syndromes (CZS) in infants[6,7]. Conversely to other flaviviruses, ZIKV has also shown the ability to bypass vector-based transmission through sexual and maternal-fetal transmissions, providing an additional mode of dissemination for this virus[8]. These specific features accentuate the risks taken by pregnant women or those planning to give birth in ZIKA epidemic and endemic zones. To date, there is no treatment for ZIKV infection and no way to prevent the neurological damage that could occur in infected fetuses during pregnancy. Microcephaly, the most striking manifestation of fetal infection leading to CZS, is a phenomenon characterized by a decreased brain size. It has been shown that ZIKV can deregulate genes involved in cell cycle, neurogenesis, and apoptosis in cortical progenitor cells and related neural progenitor cells. For key cells in brain development, disruption of cell cycle progression, reduced proliferation, premature differentiation, or increased mortality are highly detrimental[9,10]. In this context, the search for a molecule with a neuroprotective and antiviral role adapted to this particular situation of pregnancy becomes a priority.
Adiponectin is a cytokine produced by adipose tissue that is attracting growing interest due to its multiple physiological properties[11]. It was first identified for its anti-inflammatory effects and its ability to increase insulin sensitivity. Adiponectin is thus implied in a variety of metabolic diseases such as diabetes, atherosclerosis and dyslipidemia. Adiponectin signaling depends on two main types of adiponectin receptors, AdipoR1 and AdipoR2, which are ubiquitously expressed in a wide range of human tissues[12]. The adiponectin can be found in monomeric and multimeric forms. Multimers include trimers, hexamers and higher molecular weight oligomers (of 12-18 mers, >300 kDa)[13]. The different forms, including high-molecular-weight complexes, circulate in plasma and the trimeric and hexameric forms are found in human cerebrospinal fluid (CSF). Adiponectin in CSF would benefit glial and neuronal cells in oxidative stress-, inflammation- and apoptosis-dependent pathological processes[14,15,16]. Adiponectin signaling in the central nervous system (CNS) has been shown to be involved in the regulation of brain homeostasis, neurogenesis, synaptic plasticity, and memory improvement[17,18,19]. It is therefore considered neuroprotective and of therapeutic interest for various CNS disorders such as depression[20,21], stroke[22,23] and neurodevelopmental diseases[24]. AdipoR expression in neurons of the hypothalamus, brainstem, cortex and the brain blood vessels supports a role for adiponectin signaling in the brain[25]. Moreover, adiponectin was shown to promote neurogenesis and cell protection, mainly via AdipoR1 signaling[26].
Due to its pleiotropic protective properties, adiponectin has been proposed as a pharmacological approach for the treatment of several pathologies. However, the high molecular weight of the most active form, its short half-life and its stability have compromised its potential for clinical use[27], and directed research toward easier-to-use agonist molecules. AdipoRon is a synthetic small molecule and a potent adiponectin receptor agonist that can cross the blood-brain barrier. Several studies have shown its beneficial effects on brain functions[28]. A low dose of AdipoRon promoted hippocampal cell proliferation and increased serum adiponectin levels in mice[29]. In addition, AdipoRon has been shown to compensate for the depletion of neural stem cells in the hippocampus by activating the AdipoR1/AMP-activated protein kinase (AMPK) pathway[20].
Based on the deleterious impact of ZIKV on neural progenitor cell proliferation and differentiation, we hypothesized that treatment with AdipoRon might provide a beneficial neuroprotective effect in the context of ZIKV infection. To conduct an in vitro study of the effect of AdipoRon treatment on ZIKV infection, we used the A549 epithelial cells, an already well-characterized ZIKV-permissive cell model[30]. We first verified that these cells expressed the adiponectin receptors. We then studied whether commercial purified adiponectin and its pharmacological analog, AdipoRon, could modify the outcome of in vitro infection. We also showed that AdipoRon treatment, at a non-cytotoxic dose, was able to promote the expression of adiponectin receptors in A549 cells. Surprisingly, this ability was reduced by ZIKV in infected cells. However, a pre or co-treatment with AdipoRon revealed promising antiviral effects in A549 epithelial cells infected with ZIKV.
2. Materials and Methods
1. Virus, Cell Culture, and Reagents.
Human epithelial A549Dual cells (ATCC, CCL-185, Manassas, VA, USA) were cultured in DMEM (Dulbecco's Modified Eagle Medium from Gibco/Invitrogen, Carlsbad, CA, USA) that contains 10% heat-inactivated Fetal Bovine Serum (FBS, Good purchased from Invitrogen), 1 mmoL·L-1 sodium pyruvate, 2 mmoL·L-1 L-Glutamine, 0.1 mg·mL-1 of streptomycin, 100 U·mL-1 of penicillin and 0.5 µg·mL-1 of fungizone (PAN Biotech, Aidenbach, Germany). Cells were harvested at 37 °C and under a 5% CO2 atmosphere. For ZIKV, a clinical isolate of the asian lineage, isolated during the french polynesian outbreak, PF-25013-18 (ZIKVPF13) was used and has been previously described[31]. AdipoRon (SML0998) and Adiponectin (1065-AP-050) were purchased from Sigma-Aldrich (Saint-Louis, MO, USA) and RnD Systems (Minneapolis, USA) respectively.
2. Lactate dehydrogenase assay (LDH)
A549Dual cells were plated in a 96-well plate with a density of seeding corresponding to 10,000 cells per well. Cells were then treated with increasing doses of AdipoRon (ranging from 0 µg·mL-1 to 100 µg·mL-1). DMEM – dimethyl sulfoxide (DMSO) 10% was used as a positive death inductor. 24 and 48 hours later, the supernatant was harvested for LDH assay and cells underwent lactate dehydrogenase assay.
LDH assay was achieved using CytoTox 96® Non-Radioactive Cytotoxicity assay from Promega following manufacturer recommendations. Briefly, 50 µL of LDH was added to 50 µL of LDH substrate and incubated at 37°C for 15 minutes. The absorbance was then read at 490 nm using a FLUOstar® Omega (BMG LABTECH, Offenburg, Germany).

3. Neutral Red assay
A549Dual cells were plated in a 96-well plate with a density of seeding corresponding to 10,000 cells per well. Cells were then treated with increasing doses of AdipoRon (ranging from 0 µg·mL-1 to 100 µg·mL-1). DMEM – DMSO 10% was used as a positive death inductor. 24 hours later, the cells undergo neutral red assay.
Cell viability was assessed with a neutral red uptake assay, as described by Repetto et al. (10.1038/nprot.2008.75). Briefly, 100 µL of medium containing 40 µg·mL-1 of neutral red was added to each well after removal of the cell culture supernatant. Cells were then incubated for 2 hours at 37°C. Following incubation, the medium was removed and cells were washed with PBS. Finally, 150 µL of neutral red destain (50% ethanol, 49% H2O, 1% glacial acetic acid) was added per well. Absorbance was read at 540 nm using a FLUOstar® Omega microplate reader (BMG LABTECH, Offenburg, Germany).
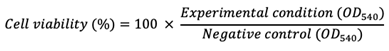
4. Cell infection and treatments with AdipoRon or Adiponectin
The potential effect of AdipoRon treatment on the outcome of ZIKV infection was assessed on A549 cells. Briefly, after seeding, cells were infected for 24 and 48 hours with ZIKVPF13 at a given multiplicity of infection (MOI 1, 2, or 5). A condition of pre-treatment was performed, with AdipoRon added to the cells 16 hours before infection (AdipoRon pre ZIKV). A co-treatment was realized with AdipoRon added simultaneously with the virus inoculum (AdipoRon + ZIKV). A post-infection treatment was realized by adding AdipoRon 24 hours post-infection, for 24 hours. Another treatment was performed only during the last 8 hours of infection (ZIKV 40h pre adipoRon). Infection was monitored by flow cytometry following ZIKV-E-positive cells, using the 4G2 antibody. Cell culture supernatants were harvested and stored at -80°C, while cells were lysed using RLT Lysis Buffer (Qiagen, Hilden, Germany) for subsequent RNA extraction.
5. Flow cytometry and antibody
For flow cytometry assay, cells were gently harvested by trypsinization, fixed with 3.7% FA in PBS for 10 minutes and 4G2 positive cells were analyzed with a Cytoflex flow cytometer (Beckman Coulter, Villepinte, France). For viral detection, a primary mouse 4G2 antibody (anti-flavivirus E protein) from RD biotech (Besançon, France) was used at 1:1000 in PBS-BSA 1%. Donkey anti-mouse Alexa Fluor 488 IgG was used as the secondary antibody at 1:1000 in PBS-BSA 1%.
6. RNA extraction and qRT-PCR
Total RNA has been extracted from cell lysates using the RNeasy Plus Mini Kit (cat. 74136, Qiagen, Hilden, Germany). The total cDNA was obtained by reverse transcription using random primers from Invitrogen (ref. 58875, ThermoFisher, Waltham, MA, USA) and M-MLV reverse transcriptase enzyme (ref. M1708, Promega, Madison, WI, USA) at 42 °C for 60 minutes. Next, cDNAs were subjected to a quantitative polymerase chain reaction, using a CFX96 Connect™ Real-Time Detection System (Bio-Rad, Hercule, CA, USA). For amplification, ABsolute™ 2X qPCR MasterMix (ref. AB1163A, Thermofischer, Waltham, MA, USA) and specific primers were used to assess gene transcripts expression for the following targets: RNAPII, GAPDH and RPLP0 for housekeeping genes and ADIPOR1, ADIPOR2, ZIKV-E (ZIKV Envelope), CPT1 for targets of interest. A threshold cycle (Ct) was calculated for each single sample amplification reaction in the exponential phase of amplification, using Bio-Rad CFX Manager 3.1 (Bio-Rad, Hercule, CA, USA).

| Gene | Forward primer | Reverse primer |
|---|---|---|
| GAPDH | 5’-CAAATTCCATGGCACCGTCA-3’ | 5’-GGAGTGGGTGTCGCTGTTGA-3’ |
| RNAPII | 5'-GAGAGCGTTGAGTTCCAGAACC-3' | 5'-TGGATGTGTGCGTTGCTCAGCA-3' |
| RPLP0 | 5'-AGATGCAGCAGATCCGCAT-3' | 5’-GGATGGCCTTGCGCA-3' |
| ADIPOR1 | 5’-AGCCTGCGGCTTAATTTGAC-3’ | 5’-CAACTAAGAACGGCCATGCA-3’ |
| ADIPOR2 | 5’-GCAGCCAAGTTTTACCGAAG-3’ | 5’-CACCTCAAATGTGGGCTTTT-3’ |
| CPT1a | 5'-GATCCTGGACAATACCTCGGAG-3' | 5'-CTCCACAGCATCAAGAGACTGC-3' |
| ZIKV-E | 5’-CTGGTCACCTGGGGAAACTA-3’ | 5’- GAGCCTTCTCAAAGCACACC-3’ |
7. Statistical analyses
Statistical analyses were performed by one-way ANOVA with Dunnett's correction using Graph-Pad Prism software version 9. All values are expressed as mean ± SD of three independent experiments. Values of p<0.05 were considered statistically significant. The degrees of significance are indicated as follows in the figure legends: ns = not significant, * p < 0.05; ** p < 0.001; *** p < 0.0002; **** p < 0.0001.
3. Results
3.1. A549 cells express both adiponectin receptor genes, ADIPOR1 and ADIPOR2
Although AdipoR1 and AdipoR2, the main receptors for adiponectin, are known to be ubiquitously expressed[32,33,34], we first ensured that it was the case in our A549 cell model of ZIKV infection. We showed that both ADIPOR1 and ADIPOR2 are expressed, with ADIPOR1 expression 6-fold higher than that of ADIPOR2 (Figure 1).
3.2. Assessment of the non-toxic concentration of AdipoRon in A549 cells
Given the concentration-dependent dual properties of AdipoRon on cell survival, and in order to test its pharmacological use, we treated cells with increasing concentrations of AdipoRon for 24 hours. We estimated cell cytotoxicity assays with cell leakage measurement (LDH activity in cell supernatants) and neutral red uptake which measures the integrity of the endocytosis and lysosome pathways. We covered a spectrum of concentrations, from 3 to 100 µg·mL-1. 25 µg·mL-1 of AdipoRon was found to be the highest non-cytotoxic concentration (Figure 2). When tested for 48 hours of treatment, this concentration did not result in excess mortality (Figure 3D), indicating that it can be used over 24 and 48 hours of infection.
3.3. AdipoRon displays antiviral and cytoprotective effects
We therefore infected A549 cells with ZIKVPF13, i) pre-treating them with adipoRon (25 µg·mL-1) 16 hours before infection, ii) co-treating them all along the 24 and 48 hours of infection, iii) post-treating them for 24 hours after 24 hours of ZIKV infection. iiii) post-treating them for 8 hours, 40 hours after the onset of infection (Figure 3A).
At both 24 and 48 hours post-infection, a treatment with AdipoRon for 16 hours before infection with ZIKVPF13 resulted in a significant two-thirds reduction in the percentage of infected cells, immuno-detected for the envelope protein (ZIKV-E) (Figure 3B and 3C). In addition, 48 hours of co-treatment with AdipoRon led to an approximately one-third reduction in the percentage of infected cells (Figure 3C). A similar reduction was observed when the AdipoRon treatment took place 24 hours after infection. However, a treatment applied 40 hours after ZIKV infection for 8 hours had no significant impact on the number of infected cells (Figure 3C).
To confirm the inhibitory effect of AdipoRon on ZIKV replication, we quantified the viral RNA in infected and treated cell extracts by qRT-PCR. We observed a significant decrease in ZIKV RNA levels when cells were pre-treated, co-treated, or treated with AdipoRon 24 hours after incubation with ZIKV (Figure 3D), which is consistent with the reduction in the percentage of infected cells assessed by cytometry. AdipoRon cell treatment prior to infection led to the lowest ZIKV infection as observed in Figure 3C and 3D. Cytotoxicity was analyzed by measuring the amount of LDH released from cells, indicating that only a pre-treatment with AdipoRon for 16 hours prior infection led to significant cell protection (Figure 3E).
3.4. AdipoRon and adiponectin provide anti-ZIKV properties
To test whether the observed antiviral effect of AdipoRon is a property shared with adiponectin, we tested the effect of commercially purified adiponectin upon A549 cells infection with ZIKVPF13 at MOI 2. Based on known concentrations of adiponectin circulating in body fluids[25] and after verification of its innocuity on cultivated A549 cells (Figure 4A), we incubated cells with adiponectin at 5 µg·mL-1. As with AdipoRon, adiponectin showed an ability to reduce cell infection when a 16h treatment was applied to cells prior to incubation with ZIKV. In contrast, co-treatment or the addition of adiponectin 24 hours post-infection had no effect on the percentage of ZIKV-E positive cells detected 48 hours post-infection (Figure 4B).
3.5. ZIKV downregulates ADIPOR1 and CPT1 gene expression
To investigate the involvement of the adiponectin signaling in limiting the infectious process, we analyzed the expression levels of genes in the AdipoR1/AMPK pathway under different conditions of ZIKV infection and AdipoRon treatment. In this axis, AMPK leads to activation of p38 mitogen-activated protein kinase (MAPK) which in turn activates the peroxisome proliferator-activated receptor (PPARα). PPARα is a transcription factor that up-regulates expression, among others, of the carnitine palmitoyltransferase 1 (CPT1) gene[35] (Figure 5A). Increased transport of fatty acids into mitochondria will result in their increased aerobic catabolism in response to adiponectin. The expression levels were analyzed by qRT-PCR for ADIPOR1 and CPT1, the signaling target known to be upregulated upon adiponectin transduction and stimulation of the AdipoR1/AMPK/PPARα pathway.
Unexpectedly, while treatment with AdipoRon led to the upregulation of both ADIPOR1 and CPT1 expression, we observed that transcriptional expression of both genes was halved under a 48 h ZIKV infection condition (Figure 5B). Upregulation of ADIPOR1 was maintained under the AdipoRon pre-treatment conditions, which may account for the protective effect obtained on cells subsequently infected with ZIKV. AdipoRon when added at the same time as ZIKV or after 24 h of infection seemed to attenuate the inhibitory effect of ZIKV on the expression of both genes. To ascertain ZIKV's ability to inhibit adiponectin receptor expression, we infected A549 cells with ZIKVPF13 at different MOI and monitored the effect on both ADIPOR1 and ADIPOR2 expressions after 24 hours of infection. We confirmed that ADIPOR1 expression was downregulated by half as early as 24 hours post-infection, independently of the MOI used (Figure 5B). Furthermore, we found that the down-regulation also affected ADIPOR2 to the same extent.
4. Discussion
Following worldwide Zika epidemics in the 2006-2016 period, and given the explosion of reported cases of adverse outcomes of pregnancies, congenital syndromes and birth defects (including about 2000 cases of ZIKV-linked microcephaly during the Brazilian outbreak[36], ZIKV must be considered a dangerous teratogenic pathogen[37]. Under these conditions, the search for antiviral drugs that can be administered to pregnant women living in ZIKV-endemic regions became a challenge. This search for safe, bio-active molecules capable of protecting the developing CNS of the fetus is indeed a necessity, especially since vertical transmission from mother to child is possible and deleterious at various stages of pregnancy[38], including the possibility of an early contamination via the father's semen[39]. This research is also needed to anticipate the possibility of the reemergence of the ZIKV Asian strains that still circulate silently in several world spots[40,41,42,43] but also in response to the threat of emergence of contemporary African strains whose teratogenic capacities are under the spotlight[44]. Unfortunately, and even if the literature on the discovery of antivirals selected from high-throughput screening of compound libraries, from observation of traditional pharmacopoeia usages, notably based on plant and marine biodiversity[45,46], or from de novo design, is plethoric, it has to be admitted that, to date, few therapeutic strategies have resulted in conclusive clinical phases[47]. Among the huge efforts to find suitable prophylactics and treatments, drug repositioning offers several undeniable advantages. One specificity in the case of ZIKV is that the profile of a good drug candidate will be based on its ability to cross the placental and blood-brain barriers to reach and act on the fetus while being completely safe for its development. There are several possible strategies. A first approach is to test molecules with proven antiviral activity, already validated and already used for other viral infections. A potentially pan-viral curative treatment regimen combining drugs targeting several proteins (the viral replicase and proteases) of the hepatitis C virus (HCV), a Flaviviridae member related to ZIKV has been developed with success[48,49]. Another approach would turn to molecules with cytoprotective effects used in CNS pathologies not necessarily of infectious origin. This is why we considered adiponectin, and more specifically its pharmacological analog AdipoRon, as a potential candidate of interest. These molecules, already known to exert neuroprotective activity fit the essential criteria set out above[14,50]. Hypothesizing that the cytoprotective action of AdipoRon could favorably influence the outcome of ZIKV infection, we found in our in vitro infection model of A549 cells expressing adiponectin receptors (mainly ADIPOR1 and to a lesser extent ADIPOR2, Figure 1) that AdipoRon was indeed able to limit infection. Pre-treatment of the cells at a concentration of 25 µg·mL-1, validated as non-cytotoxic (2) reduced by 2/3 the number of replicating ZIKV cells, and the associated cytopathy (Figure 3). While treatment of cells with AdipoRon leads to overexpression of the gene encoding AdipoR1 and that of CPT1, a transcriptional target of PPARα activated during adiponectin signaling, we revealed that ZIKV was able to interfere with this signaling pathway (Figure 5). We demonstrated ZIKV's ability to halve ADIPOR1 and ADIPOR2 expression as early as 24h post-infection. This suggests that the adiponectin signaling pathway and its amplification loop are detrimental to ZIKV.
To our knowledge, we have shown for the first time that the adiponectin signaling pathway has antiviral potential, making it a target that ZIKV subverts to limit its effect. In our study, we highlighted that the ability of AdipoRon to limit infection when administered in pretreatment is a feature shared by adiponectin. However, considering the lesser effect observed with adiponectin compared to its pharmacological analog, we speculate that AdipoRon may mimic the most active form of adiponectin in its anti-ZIKV capacity. This form, yet to be identified, would be weakly represented in the population of various multimers possibly present in the commercial suspension of adiponectin we used. Adiponectin doses may need to be increased to provide efficacy similar to that of its analog. Nevertheless, we do not exclude that AdipoRon may have activities independent of its ability to mimic adiponectin. However, the literature does not mention any related effects of this molecule, currently characterized exclusively for its “adiponectin receptor agonist” effects.
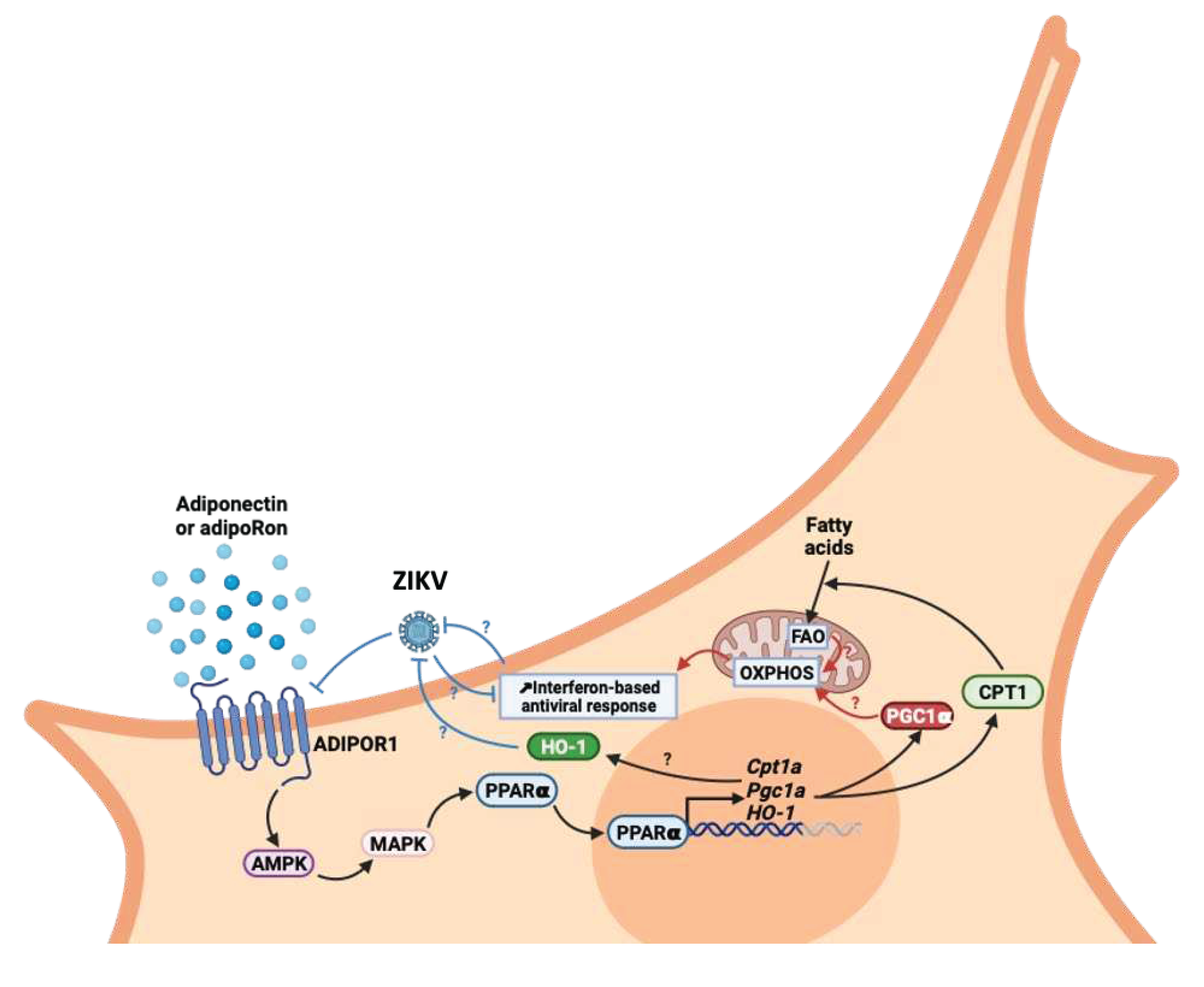
Finally, we still need to understand the cellular and molecular mechanisms by which adiponectin signaling exerts an inhibitory effect on ZIKV multiplication. Several hypotheses can be formulated to investigate the links between the identified cytoprotective capacities of adiponectin signaling and cellular activities that might impair viral multiplication. Among the possibilities to be explored, metabolic reprogramming is one of the most exciting. On one hand, AdipoR1/AMPK signaling has been shown to be responsible for a cellular shift towards increased oxidative metabolism. Data available in the literature repeatedly mention that AdipoR signaling favors mitochondrial biogenesis and activity, oxidative phosphorylation (OXPHOS) and ATP synthesis through the Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) activation and its downstream targets[51,52,53,54]. On the other hand, the demonstration was made that viruses benefit from glycolytic cellular metabolism[55]. Conversely, aerobic metabolism and high OXPHOS are associated with a better interferon response and antiviral properties[56,57]. It therefore appears that viruses have strategies to favor glycolytic use of glucose instead of OXPHOS, to increase viral production and limit antiviral response. In this regard, ZIKV and few other flaviviruses are known examples of viruses that promotes metabolic reprogramming to their benefit[58,59,60,61,62]. Indeed, AMPK system is a sensor of cellular energy status that responds primarily to changes in AMP:ATP ratio and activated OXPHOS. This diaphonic relationship between reprogramming of cellular metabolism and infection could therefore be disrupted by agonists of the AdipoR1/AMPK/PPARα axis. Potentiation of the antiviral response is not the only lever on which AdipoR1 agonists could act. Potentiation of cytoprotective mechanisms could also explain the beneficial effects of AdipoR1 agonists. Of note, some work describes how adiponectin-mediated Heme oxygenase-1 (HO-1) induction could contribute to the cytoprotective effects attributed to adiponectin signaling[63]. HO-1 has been qualified as antiviral against several viruses[64], including ZIKV but with underlying mechanisms that are not known. Interestingly, in a previous work we demonstrated that HO-1 was a cellular antiviral player that ZIKV was able to subvert[65]. Setting these data in perspective suggests that adiponectin signaling and HO-1 antiviral activity may be linked. The cyto-protection associated with signaling by adiponectin and the AdipoR1/AMPK pathway is also attributed to a capacity to inhibit inflammation and oxidative stress via inhibition of the activity of the transcription factor nuclear factor kappa B (NF-κB)[11]. AMPK inactivates the P65 subunit of NF-κB by stimulating the expression of the gene encoding the “silent information regulator sirtuin 1” (SIRT1), which deacetylates P65. This SIRT1 expression is mediated by the transcription factor PGC-1α, activated by the AdipoR1-AMPK axis[66,67]. Another mechanism of regulation by AMPK is inhibition of p65 phosphorylation by targeting IKKb, leading to the absence of nuclear translocation of p65[68].
We highlighted in our study that the effect of AdipoRon or adiponectin is limited once infection has set in. This limitation may be due to the mechanism of action of these agonists and/or to the virus' ability to downregulate the expression of the Adiponectin receptors as we stated (Figure 6). With the hypothesis that AdipoR1/R2 agonists enhance OXPHOS, it's worth remembering that infection, as discussed above, promotes aerobic glycolysis. AdipoR agonists and the virus therefore act in opposition for metabolic reprogramming. Indeed, it has been shown that M1 macrophages that display low OXPHOS have a lower expression of ADIPOR1/R2 than those of the M2 phenotype exhibiting high OXPHOS[69,70]. Then, one explanation for the decrease in ADIPOR1/R2 expression following ZIKV infection could be the virus-induced metabolic reprogramming towards aerobic glycolysis. These metabolic redirection patterns, which would explain the crosstalk relationship between ZIKV infection and the AdipoR-AMPK axis, must therefore be evaluated in future research. It should be added that a direct action of ZIKV in inhibiting ADIPOR1/2 expression cannot be ruled out. Indeed, ZIKV's ability to reduce the expression of certain genes involved in metabolic control was not entirely surprising, as shown by numerous transcriptome analyses[71,72]. Here again, the mechanisms by which ZIKV directs targeted gene silencing remain to be explored.
Whatever, taken together, these observations point to a potential beneficial effect of the AdipoR1/AMPK pathway stimulation during ZIKV infection (Figure 6). This can be driven by adiponectin or its pharmacological analog AdipoRon with benefits at different levels. Like in a recent study on new therapeutic opportunities to fight West Nile virus[62], our study demonstrates the value of antiviral molecules targeting the potentiation of responses and metabolic orientation of the host cells rather than targeting the virus particle or viral factors involved in the viral cycle. Our results suggest that preventive rather than therapeutic use of adiponectin receptor agonists could be proposed until effective vaccines against ZIKV become available. This drug repositioning approach of agonists of the adiponectin signaling pathways seems to us to hold out hope for people wishing to procreate in ZIKA-endemic areas. At the end, this work is a first step towards preclinical model to study beneficial effect of adiponectin receptor agonists on viral load, but also on neurogenesis under ZIKV infection conditions in vivo in animal models or in ex-vivo models of developing brain, like cerebral organoids from human embryonic stem cells.
Author Contributions
Conceptualization, P.K-T. and W.V.; methodology and experiments D.E-S., G.L., J.T.; contribution to new reagents/analytic tools C.L.H., N.D., formal analysis, D.E-S., G.L., J.T., W.V. and P.K-T.; resources, C.L.H., N.D., W.V..; writing—original draft preparation, D.E-S, G.L, W.V. and P.K-T.; writing—review and editing, P.K-T.; supervision, W.V. and P.K-T.; funding acquisition, C.L.H and P.K-T.
Funding
This work was supported by the Federation BioST from Reunion Island University (ZIKAPONE project). D.E.S., and G.L have Ph.D. degree scholarships from Réunion University (Ecole doctorale STS) funded by DIRED/2021-1115, DIRED/2021-0161 from Région Réunion Council, respectively.
Acknowledgments
Illustration were created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Song, B.-H.; Yun, S.-I.; Woolley, M.; Lee, Y.-M. Zika Virus: History, Epidemiology, Transmission, and Clinical Presentation. Journal of Neuroimmunology 2017, 308, 50–64. [CrossRef]
- Gubler, D.J.; Vasilakis, N.; Musso, D. History and Emergence of Zika Virus. The Journal of Infectious Diseases 2017, 216, S860–S867. [CrossRef]
- White, M.K.; Wollebo, H.S.; David Beckham, J.; Tyler, K.L.; Khalili, K. Zika Virus: An Emergent Neuropathological Agent. Annals of Neurology 2016, 80, 479–489. [CrossRef]
- Yun, S.-I.; Lee, Y.-M. Zika Virus: An Emerging Flavivirus. J Microbiol. 2017, 55, 204–219. [CrossRef]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 1640. [CrossRef]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An Update on Zika Virus Infection. The Lancet 2017, 390, 2099–2109. [CrossRef]
- Marbán-Castro, E.; Goncé, A.; Fumadó, V.; Romero-Acevedo, L.; Bardají, A. Zika Virus Infection in Pregnant Women and Their Children: A Review. European Journal of Obstetrics & Gynecology and Reproductive Biology 2021, 265, 162–168. [CrossRef]
- Runge-Ranzinger, S.; Morrison, A.C.; Manrique-Saide, P.; Horstick, O. Zika Transmission Patterns: A Meta-review. Tropical Med Int Health 2019, 24, 523–529. [CrossRef]
- Wen, Z.; Song, H.; Ming, G. How Does Zika Virus Cause Microcephaly? Genes Dev. 2017, 31, 849–861. [CrossRef]
- Christian, K.M.; Song, H.; Ming, G. Pathophysiology and Mechanisms of Zika Virus Infection in the Nervous System. Annu. Rev. Neurosci. 2019, 42, 249–269. [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. IJMS 2020, 21, 1219. [CrossRef]
- Heiker, J.T.; Kosel, D.; Beck-Sickinger, A.G. Molecular Mechanisms of Signal Transduction via Adiponectin and Adiponectin Receptors. Biological Chemistry 2010, 391. [CrossRef]
- Tsao, T.-S. Assembly of Adiponectin Oligomers. Rev Endocr Metab Disord 2014, 15, 125–136. [CrossRef]
- Miao, W.; Jiang, L.; Xu, F.; Lyu, J.; Jiang, X.; He, M.; Liu, Y.; Yang, T.; Leak, R.K.; Stetler, R.A.; et al. Adiponectin Ameliorates Hypoperfusive Cognitive Deficits by Boosting a Neuroprotective Microglial Response. Progress in Neurobiology 2021, 205, 102125. [CrossRef]
- Chan, K.-H.; Lam, K.S.-L.; Cheng, O.-Y.; Kwan, J.S.-C.; Ho, P.W.-L.; Cheng, K.K.-Y.; Chung, S.K.; Ho, J.W.-M.; Guo, V.Y.; Xu, A. Adiponectin Is Protective against Oxidative Stress Induced Cytotoxicity in Amyloid-Beta Neurotoxicity. PLoS ONE 2012, 7, e52354. [CrossRef]
- Qiu, G.; Wan, R.; Hu, J.; Mattson, M.P.; Spangler, E.; Liu, S.; Yau, S.-Y.; Lee, T.M.C.; Gleichmann, M.; Ingram, D.K.; et al. Adiponectin Protects Rat Hippocampal Neurons against Excitotoxicity. AGE 2011, 33, 155–165. [CrossRef]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plasticity 2018, 2018, 1–15. [CrossRef]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. IJMS 2020, 21, 2010. [CrossRef]
- Thundyil, J.; Pavlovski, D.; Sobey, C.G.; Arumugam, T.V. Adiponectin Receptor Signalling in the Brain. British J Pharmacology 2012, 165, 313–327. [CrossRef]
- Liu, J.; Guo, M.; Zhang, D.; Cheng, S.-Y.; Liu, M.; Ding, J.; Scherer, P.E.; Liu, F.; Lu, X.-Y. Adiponectin Is Critical in Determining Susceptibility to Depressive Behaviors and Has Antidepressant-like Activity. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, 12248–12253. [CrossRef]
- Wędrychowicz, A. Peptides from Adipose Tissue in Mental Disorders. WJP 2014, 4, 103. [CrossRef]
- Song, W.; Huo, T.; Guo, F.; Wang, H.; Wei, H.; Yang, Q.; Dong, H.; Wang, Q.; Xiong, L. Globular Adiponectin Elicits Neuroprotection by Inhibiting NADPH Oxidase-Mediated Oxidative Damage in Ischemic Stroke. Neuroscience 2013, 248, 136–144. [CrossRef]
- Bai, H.; Zhao, L.; Liu, H.; Guo, H.; Guo, W.; Zheng, L.; Liu, X.; Wu, X.; Luo, J.; Li, X.; et al. Adiponectin Confers Neuroprotection against Cerebral Ischemia-Reperfusion Injury through Activating the CAMP/PKA-CREB-BDNF Signaling. Brain Research Bulletin 2018, 143, 145–154. [CrossRef]
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: Adiponectin. Nat Metab 2019, 1, 334–339. [CrossRef]
- Clain, J.; Couret, D.; Planesse, C.; Krejbich-Trotot, P.; Meilhac, O.; Lefebvre d’Hellencourt, C.; Viranaicken, W.; Diotel, N. Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke. Brain Sciences 2022, 12, 680. [CrossRef]
- Song, J.; Kang, S.M.; Kim, E.; Kim, C.-H.; Song, H.-T.; Lee, J.E. Adiponectin Receptor-Mediated Signaling Ameliorates Cerebral Cell Damage and Regulates the Neurogenesis of Neural Stem Cells at High Glucose Concentrations: An in Vivo and in Vitro Study. Cell Death Dis 2015, 6, e1844–e1844. [CrossRef]
- Nigro, E.; Daniele, A.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. AdipoRon and Other Adiponectin Receptor Agonists as Potential Candidates in Cancer Treatments. IJMS 2021, 22, 5569. [CrossRef]
- Formolo, D.A.; Lee, T.H.; Yu, J.; Lin, K.; Chen, G.; Kranz, G.S.; Yau, S.-Y. Increasing Adiponectin Signaling by Sub-Chronic AdipoRon Treatment Elicits Antidepressant- and Anxiolytic-Like Effects Independent of Changes in Hippocampal Plasticity. Biomedicines 2023, 11, 249. [CrossRef]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.-F.; Xu, A.; So, K.-F.; Yau, S. AdipoRon Treatment Induces a Dose-Dependent Response in Adult Hippocampal Neurogenesis. IJMS 2021, 22, 2068. [CrossRef]
- Frumence, E.; Roche, M.; Krejbich-Trotot, P.; El-Kalamouni, C.; Nativel, B.; Rondeau, P.; Missé, D.; Gadea, G.; Viranaicken, W.; Desprès, P. The South Pacific Epidemic Strain of Zika Virus Replicates Efficiently in Human Epithelial A549 Cells Leading to IFN-β Production and Apoptosis Induction. Virology 2016, 493, 217–226. [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J Virol 2015, 89, 8880–8896. [CrossRef]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of Adiponectin Receptors That Mediate Antidiabetic Metabolic Effects. Nature 2003, 423, 762–769. [CrossRef]
- Miller, M.; Cho, J.Y.; Pham, A.; Ramsdell, J.; Broide, D.H. Adiponectin and Functional Adiponectin Receptor 1 Are Expressed by Airway Epithelial Cells in Chronic Obstructive Pulmonary Disease. The Journal of Immunology 2009, 182, 684–691. [CrossRef]
- Chen, H.-M.; Yang, C.-M.; Chang, J.-F.; Wu, C.-S.; Sia, K.-C.; Lin, W.-N. AdipoR-Increased Intracellular ROS Promotes CPLA 2 and COX-2 Expressions via Activation of PKC and P300 in Adiponectin-Stimulated Human Alveolar Type II Cells. American Journal of Physiology-Lung Cellular and Molecular Physiology 2016, 311, L255–L269. [CrossRef]
- Yoon, M.J.; Lee, G.Y.; Chung, J.-J.; Ahn, Y.H.; Hong, S.H.; Kim, J.B. Adiponectin Increases Fatty Acid Oxidation in Skeletal Muscle Cells by Sequential Activation of AMP-Activated Protein Kinase, P38 Mitogen-Activated Protein Kinase, and Peroxisome Proliferator–Activated Receptor α. Diabetes 2006, 55, 2562–2570. [CrossRef]
- de Araújo, T.V.B.; Ximenes, R.A. de A.; Miranda-Filho, D. de B.; Souza, W.V.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; de Albuquerque, M. de F.P.M.; Braga, C.; Filho, S.P.B.; et al. Association between Microcephaly, Zika Virus Infection, and Other Risk Factors in Brazil: Final Report of a Case-Control Study. The Lancet Infectious Diseases 2018, 18, 328–336. [CrossRef]
- Gilbert, R.K.; Petersen, L.R.; Honein, M.A.; Moore, C.A.; Rasmussen, S.A. Zika Virus as a Cause of Birth Defects: Were the Teratogenic Effects of Zika Virus Missed for Decades? Birth Defects Research 2023, 115, 265–274. [CrossRef]
- Zorrilla, C.D.; García García, I.; García Fragoso, L.; De La Vega, A. Zika Virus Infection in Pregnancy: Maternal, Fetal, and Neonatal Considerations. The Journal of Infectious Diseases 2017, 216, S891–S896. [CrossRef]
- Watts, J.L.; Ralston, A. The Fetal Lineage Is Susceptible to Zika Virus Infection within Days of Fertilization. Development 2022, 149, dev200501. [CrossRef]
- Bonifay, T.; Le Turnier, P.; Epelboin, Y.; Carvalho, L.; De Thoisy, B.; Djossou, F.; Duchemin, J.-B.; Dussart, P.; Enfissi, A.; Lavergne, A.; et al. Review on Main Arboviruses Circulating on French Guiana, An Ultra-Peripheric European Region in South America. Viruses 2023, 15, 1268. [CrossRef]
- Noisumdaeng, P.; Dangsagul, W.; Sangsiriwut, K.; Prasertsopon, J.; Changsom, D.; Yoksan, S.; Ajawatanawong, P.; Buathong, R.; Puthavathana, P. Molecular Characterization and Geographical Distribution of Zika Virus Worldwide from 1947 to 2022. International Journal of Infectious Diseases 2023, 136, 5–10. [CrossRef]
- Marquine, S.; Durand, G.A.; Modenesi, G.; Khouadhria, S.; Piorkowski, G.; Badaut, C.; Canivez, T.; De Lamballerie, X.; Grard, G.; Klitting, R. Sequence Data From a Travel-Associated Case of Microcephaly Highlight a Persisting Risk Due to Zika Virus Circulation in Thailand. The Journal of Infectious Diseases 2023, jiad322. [CrossRef]
- Braga, C.; Martelli, C.M.T.; Souza, W.V.; Luna, C.F.; Albuquerque, M. de F.P.M.; Mariz, C.A.; Morais, C.N.L.; Brito, C.A.A.; Melo, C.F.C.A.; Lins, R.D.; et al. Seroprevalence of Dengue, Chikungunya and Zika at the Epicenter of the Congenital Microcephaly Epidemic in Northeast Brazil: A Population-Based Survey. PLoS Negl Trop Dis 2023, 17, e0011270. [CrossRef]
- Aubry, F.; Jacobs, S.; Darmuzey, M.; Lequime, S.; Delang, L.; Fontaine, A.; Jupatanakul, N.; Miot, E.F.; Dabo, S.; Manet, C.; et al. Recent African Strains of Zika Virus Display Higher Transmissibility and Fetal Pathogenicity than Asian Strains. Nat Commun 2021, 12, 916. [CrossRef]
- Pereira, R.S.; Santos, F.C.P.; Campana, P.R.V.; Costa, V.V.; de Pádua, R.M.; Souza, D.G.; Teixeira, M.M.; Braga, F.C. Natural Products and Derivatives as Potential Zika Virus Inhibitors: A Comprehensive Review. Viruses 2023, 15, 1211. [CrossRef]
- Salampe, M.; Mamada, S.S.; Evary, Y.M.; Mitra, S.; Bin Emran, T.; Harapan, H.; Nainu, F.; Simal-Gandara, J. Promising Marine Natural Products for Tackling Viral Outbreaks:A Focus on Possible Targets and Structure-Activity Relationship. CTMC 2023, 23, 1352–1379. [CrossRef]
- Bernatchez, J.A.; Tran, L.T.; Li, J.; Luan, Y.; Siqueira-Neto, J.L.; Li, R. Drugs for the Treatment of Zika Virus Infection. J. Med. Chem. 2020, 63, 470–489. [CrossRef]
- Martinello, M.; Naggie, S.; Rockstroh, J.K.; Matthews, G.V. Direct-Acting Antiviral Therapy for Treatment of Acute and Recent Hepatitis C Virus Infection: A Narrative Review. Clinical Infectious Diseases 2023, 77, S238–S244. [CrossRef]
- Mayberry, J.; Lee, W.M. The Revolution in Treatment of Hepatitis C. Medical Clinics of North America 2019, 103, 43–55. [CrossRef]
- Ng, R.C.-L.; Jian, M.; Ma, O.K.-F.; Bunting, M.; Kwan, J.S.-C.; Zhou, G.-J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.-K.; Li, M.; et al. Chronic Oral Administration of AdipoRon Reverses Cognitive Impairments and Ameliorates Neuropathology in an Alzheimer’s Disease Mouse Model. Mol Psychiatry 2021, 26, 5669–5689. [CrossRef]
- Zhang, S.; Wu, X.; Wang, J.; Shi, Y.; Hu, Q.; Cui, W.; Bai, H.; Zhou, J.; Du, Y.; Han, L.; et al. Adiponectin/AdiopR1 Signaling Prevents Mitochondrial Dysfunction and Oxidative Injury after Traumatic Brain Injury in a SIRT3 Dependent Manner. Redox Biology 2022, 54, 102390. [CrossRef]
- Qiao, L.; Kinney, B.; Yoo, H. sun; Lee, B.; Schaack, J.; Shao, J. Adiponectin Increases Skeletal Muscle Mitochondrial Biogenesis by Suppressing Mitogen-Activated Protein Kinase Phosphatase-1. Diabetes 2012, 61, 1463–1470. [CrossRef]
- Majeed, Y.; Halabi, N.; Madani, A.Y.; Engelke, R.; Bhagwat, A.M.; Abdesselem, H.; Agha, M.V.; Vakayil, M.; Courjaret, R.; Goswami, N.; et al. SIRT1 Promotes Lipid Metabolism and Mitochondrial Biogenesis in Adipocytes and Coordinates Adipogenesis by Targeting Key Enzymatic Pathways. Sci Rep 2021, 11, 8177. [CrossRef]
- Pal, S.; Singh, M.; Porwal, K.; Rajak, S.; Das, N.; Rajput, S.; Trivedi, A.K.; Maurya, R.; Sinha, R.A.; Siddiqi, M.I.; et al. Adiponectin Receptors by Increasing Mitochondrial Biogenesis and Respiration Promote Osteoblast Differentiation: Discovery of Isovitexin as a New Class of Small Molecule Adiponectin Receptor Modulator with Potential Osteoanabolic Function. European Journal of Pharmacology 2021, 913, 174634. [CrossRef]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral Hijacking of Cellular Metabolism. BMC Biol 2019, 17, 59. [CrossRef]
- Lebeau, G.; Paulo-Ramos, A.; Hoarau, M.; El Safadi, D.; Meilhac, O.; Krejbich-Trotot, P.; Roche, M.; Viranaicken, W. Metabolic Dependency Shapes Bivalent Antiviral Response in Host Cells: The Role of Glutamine; Immunology, 2023;
- Lebeau, G.; El Safadi, D.; Paulo-Ramos, A.; Hoareau, M.; Desprès, P.; Krejbich-Trotot, P.; Chouchou, F.; Roche, M.; Viranaicken, W. The Efficient Antiviral Response of A549 Cells Is Enhanced When Mitochondrial Respiration Is Promoted. Pathogens 2022, 11, 1168. [CrossRef]
- Chen, Q.; Gouilly, J.; Ferrat, Y.J.; Espino, A.; Glaziou, Q.; Cartron, G.; El Costa, H.; Al-Daccak, R.; Jabrane-Ferrat, N. Metabolic Reprogramming by Zika Virus Provokes Inflammation in Human Placenta. Nat Commun 2020, 11, 2967. [CrossRef]
- Gilbert-Jaramillo, J.; Purnama, U.; Molnár, Z.; James, W.S. Zika Virus-Induces Metabolic Alterations in Fetal Neuronal Progenitors That Could Influence in Neurodevelopment during Early Pregnancy. Biology Open 2023, 12, bio059889. [CrossRef]
- Fontaine, K.A.; Sanchez, E.L.; Camarda, R.; Lagunoff, M. Dengue Virus Induces and Requires Glycolysis for Optimal Replication. J Virol 2015, 89, 2358–2366. [CrossRef]
- Jordan, T.X.; Randall, G. Flavivirus Modulation of Cellular Metabolism. Current Opinion in Virology 2016, 19, 7–10. [CrossRef]
- Mingo-Casas, P.; Blázquez, A.-B.; Gómez de Cedrón, M.; San-Félix, A.; Molina, S.; Escribano-Romero, E.; Calvo-Pinilla, E.; Jiménez de Oya, N.; Ramírez de Molina, A.; Saiz, J.-C.; et al. Glycolytic Shift during West Nile Virus Infection Provides New Therapeutic Opportunities. J Neuroinflammation 2023, 20, 217. [CrossRef]
- Lin, H.; Yu, C.-H.; Jen, C.-Y.; Cheng, C.-F.; Chou, Y.; Chang, C.-C.; Juan, S.-H. Adiponectin-Mediated Heme Oxygenase-1 Induction Protects Against Iron-Induced Liver Injury via a PPARα-Dependent Mechanism. The American Journal of Pathology 2010, 177, 1697–1709. [CrossRef]
- Espinoza, J.A.; León, M.A.; Céspedes, P.F.; Gómez, R.S.; Canedo-Marroquín, G.; Riquelme, S.A.; Salazar-Echegarai, F.J.; Blancou, P.; Simon, T.; Anegon, I.; et al. Heme Oxygenase-1 Modulates Human Respiratory Syncytial Virus Replication and Lung Pathogenesis during Infection. The Journal of Immunology 2017, 199, 212–223. [CrossRef]
- El Kalamouni, C.; Frumence, E.; Bos, S.; Turpin, J.; Nativel, B.; Harrabi, W.; Wilkinson, D.; Meilhac, O.; Gadea, G.; Desprès, P.; et al. Subversion of the Heme Oxygenase-1 Antiviral Activity by Zika Virus. Viruses 2018, 11, 2. [CrossRef]
- Barroso, E.; Eyre, E.; Palomer, X.; Vázquez-Carrera, M. The Peroxisome Proliferator-Activated Receptor β/δ (PPARβ/δ) Agonist GW501516 Prevents TNF-α-Induced NF-ΚB Activation in Human HaCaT Cells by Reducing P65 Acetylation through AMPK and SIRT1. Biochemical Pharmacology 2011, 81, 534–543. [CrossRef]
- Salminen, A.; Hyttinen, J.M.T.; Kaarniranta, K. AMP-Activated Protein Kinase Inhibits NF-ΚB Signaling and Inflammation: Impact on Healthspan and Lifespan. J Mol Med 2011, 89, 667–676. [CrossRef]
- Antonia, R.J.; Baldwin, A.S. IKK Promotes Cytokine-Induced and Cancer-Associated AMPK Activity and Attenuates Phenformin-Induced Cell Death in LKB1-Deficient Cells. Sci. Signal. 2018, 11, eaan5850. [CrossRef]
- Stijn, C.M.W.; Kim, J.; Lusis, A.J.; Barish, G.D.; Tangirala, R.K. Macrophage Polarization Phenotype Regulates Adiponectin Receptor Expression and Adiponectin Anti-inflammatory Response. FASEB j. 2015, 29, 636–649. [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [CrossRef]
- Chen, Z.; Yin, Z.; Qiu, T.; Chen, J.; Liu, J.; Zhang, X.; Xu, J. Revealing the Characteristics of ZIKV Infection through Tissue-Specific Transcriptome Sequencing Analysis. BMC Genomics 2022, 23, 697. [CrossRef]
- Tiwari, S.K.; Dang, J.; Qin, Y.; Lichinchi, G.; Bansal, V.; Rana, T.M. Zika Virus Infection Reprograms Global Transcription of Host Cells to Allow Sustained Infection. Emerging Microbes & Infections 2017, 6, 1–10. [CrossRef]
Figure 1.
Adiponectin receptor ADIPOR1 and ADIPOR2 expression in A549 cell model. The basal expression level of ADIPOR1 and ADIPOR2 mRNA was measured by qRT-PCR in A549 cells under normal growing conditions. Expression levels were normalized to the housekeeping gene (RPLP0). ADIPOR1 is almost 6-fold more expressed than ADIPOR2 in A549 cells. The error bars represent the standard deviations of three independent experiments.
Figure 1.
Adiponectin receptor ADIPOR1 and ADIPOR2 expression in A549 cell model. The basal expression level of ADIPOR1 and ADIPOR2 mRNA was measured by qRT-PCR in A549 cells under normal growing conditions. Expression levels were normalized to the housekeeping gene (RPLP0). ADIPOR1 is almost 6-fold more expressed than ADIPOR2 in A549 cells. The error bars represent the standard deviations of three independent experiments.
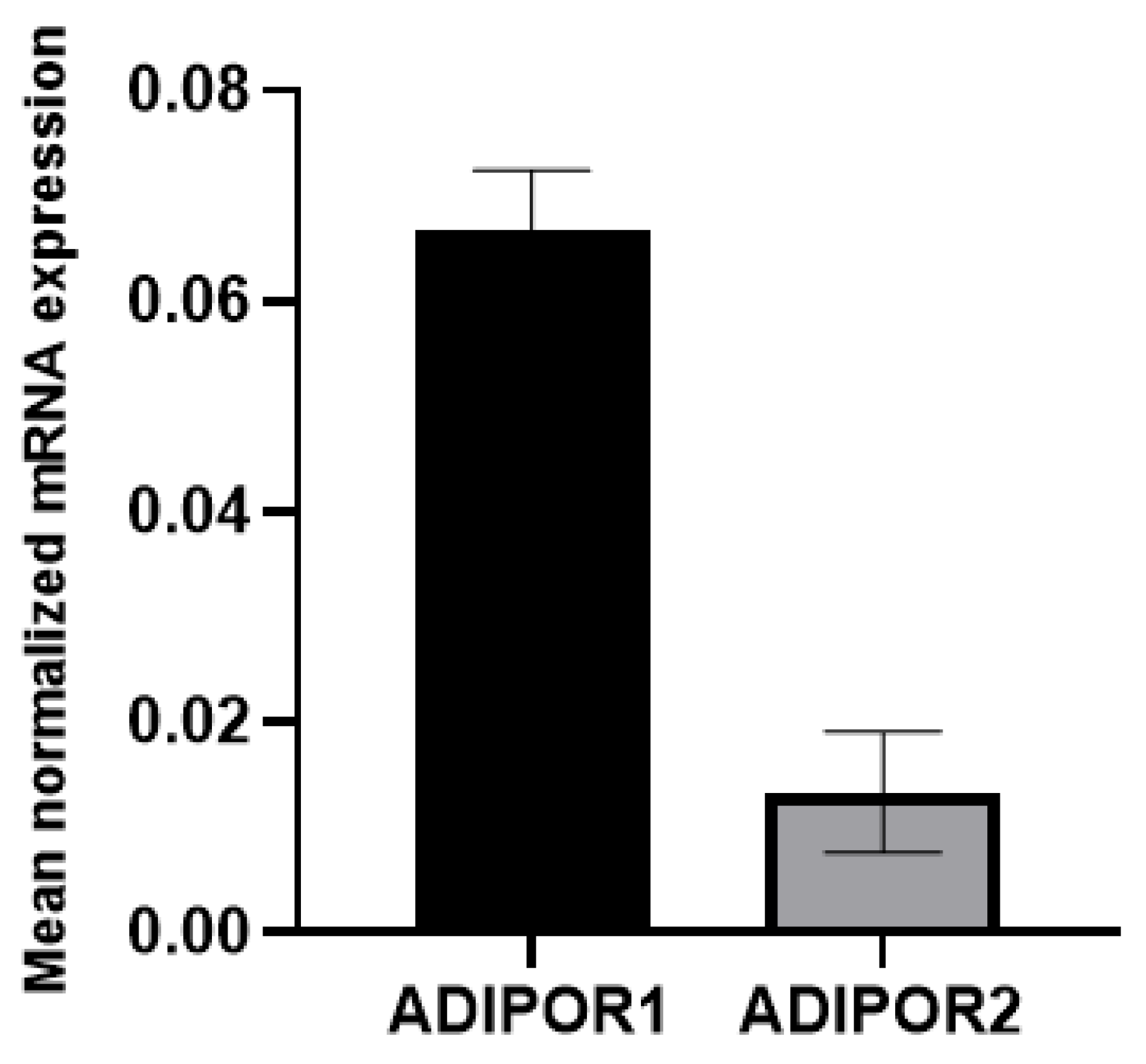
Figure 2.
Dose-dependent cytotoxicity of AdipoRon on A549 cells. AdipoRon-related cytotoxicity was assessed by (a) LDH, and (b) neutral red assays. Cells were plated and incubated for 24h with different concentrations of AdipoRon (3.125, 6.25, 12.5, 25, 50 and 100 µg·mL-1) as indicated in the figures. Values represent the mean and the standard deviations of three independent experiments, ** p < 0.001; *** p < 0.0002; **** p < 0.0001. .
Figure 2.
Dose-dependent cytotoxicity of AdipoRon on A549 cells. AdipoRon-related cytotoxicity was assessed by (a) LDH, and (b) neutral red assays. Cells were plated and incubated for 24h with different concentrations of AdipoRon (3.125, 6.25, 12.5, 25, 50 and 100 µg·mL-1) as indicated in the figures. Values represent the mean and the standard deviations of three independent experiments, ** p < 0.001; *** p < 0.0002; **** p < 0.0001. .
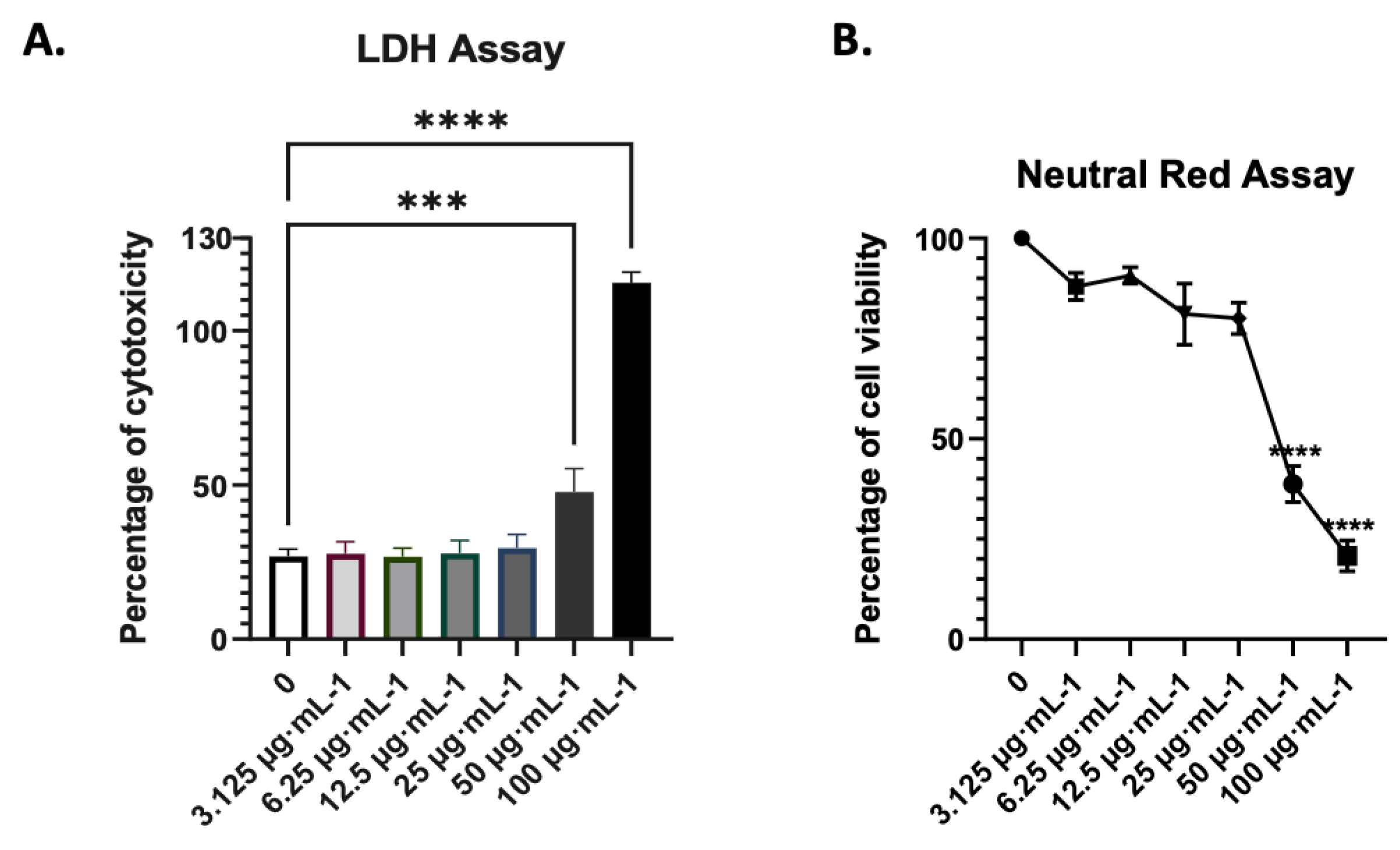
Figure 3.
AdipoRon treatment of A549 cells protects against ZIKV infection and cytopathy. (a) Schematic illustration of the experimental design of AdipoRon treatments and ZIKV infection. (b) Percentage of ZIKV-E positive cells stained by the 4G2 antibody, 24 hours post-infection with ZIKVPF13 at MOI 2. (c) Percentage of ZIKV-E positive cells stained by the 4G2 antibody, 48 hours post-infection with ZIKVPF13 at MOI 2. (d) ZIKV replication was determined by quantification of the viral RNA levels (ZIKV-E) by qRT-PCR. Expression levels were normalized to the housekeeping gene (RNAPII) and fold changes are related to viral RNA quantification at 48h post infection. (e) Percentage of cytotoxicity measured by LDH release in cell culture supernatants of treated and infected cells. Values represent the mean and the standard deviations of three independent experiments, ns: non-significant; **** p < 0.0001. .
Figure 3.
AdipoRon treatment of A549 cells protects against ZIKV infection and cytopathy. (a) Schematic illustration of the experimental design of AdipoRon treatments and ZIKV infection. (b) Percentage of ZIKV-E positive cells stained by the 4G2 antibody, 24 hours post-infection with ZIKVPF13 at MOI 2. (c) Percentage of ZIKV-E positive cells stained by the 4G2 antibody, 48 hours post-infection with ZIKVPF13 at MOI 2. (d) ZIKV replication was determined by quantification of the viral RNA levels (ZIKV-E) by qRT-PCR. Expression levels were normalized to the housekeeping gene (RNAPII) and fold changes are related to viral RNA quantification at 48h post infection. (e) Percentage of cytotoxicity measured by LDH release in cell culture supernatants of treated and infected cells. Values represent the mean and the standard deviations of three independent experiments, ns: non-significant; **** p < 0.0001. .
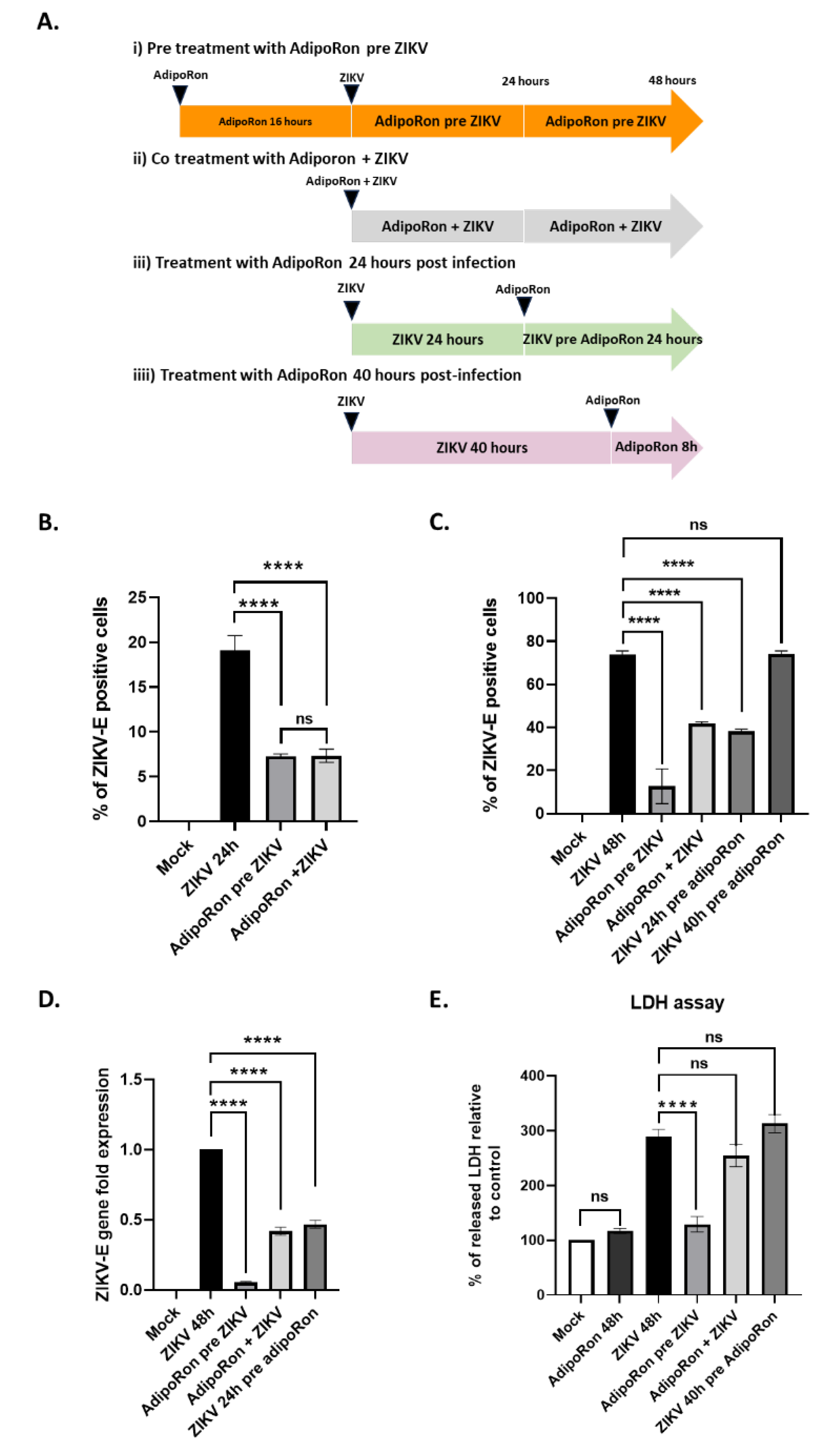
Figure 4.
A549 cell treatment with Adiponectin and ZIKV infection. (a) Effect of 5 µg·mL-1 of adiponectin treatment on cytotoxicity, related to non-treated cells (measured by LDH release in cell culture supernatants); (b) Effects of adiponectin treatment (5 µg·mL-1) on the percentage of ZIKV-E positive cells stained by the 4G2 antibody, 48 hours post infection. Values represent the mean and the standard deviations of three independent experiments. ns = not significant, * p < 0.05; *** p < 0.0002; **** p < 0.0001.
Figure 4.
A549 cell treatment with Adiponectin and ZIKV infection. (a) Effect of 5 µg·mL-1 of adiponectin treatment on cytotoxicity, related to non-treated cells (measured by LDH release in cell culture supernatants); (b) Effects of adiponectin treatment (5 µg·mL-1) on the percentage of ZIKV-E positive cells stained by the 4G2 antibody, 48 hours post infection. Values represent the mean and the standard deviations of three independent experiments. ns = not significant, * p < 0.05; *** p < 0.0002; **** p < 0.0001.
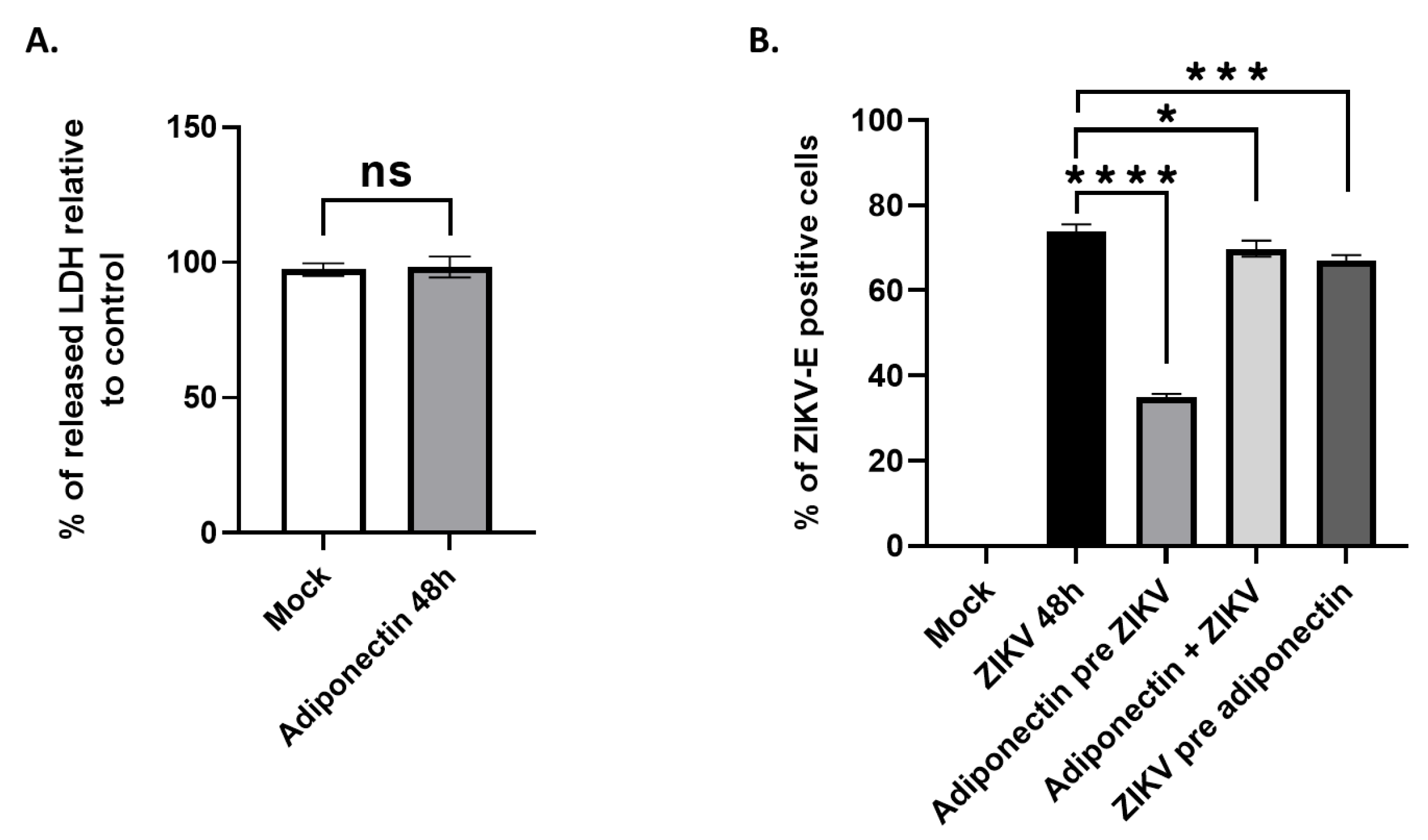
Figure 5.
ZIKV infection and AdipoRon treatment of A549 cells modify gene expression in the Adiponectin signaling pathway. (a) Schematic representation of the PPARα-CPT1 axis of the AdipoR1/AMPK signaling pathway. (b) The ADIPOR1 gene and the CPT1 gene, a gene under the control of the adiponectin/AdipoR1 pathway, are down-regulated after 48 hours of ZIKV infection. (c) ZIKV controls adiponectin receptor expression. Expression of the ADIPOR1 and ADIPOR2 genes was normalized to the RNAPII housekeeping gene. Values represent the mean and the standard deviations of three independent experiments. ns = not significant, * p < 0.05; ** p < 0.001; *** p < 0.0002; **** p < 0.0001.
Figure 5.
ZIKV infection and AdipoRon treatment of A549 cells modify gene expression in the Adiponectin signaling pathway. (a) Schematic representation of the PPARα-CPT1 axis of the AdipoR1/AMPK signaling pathway. (b) The ADIPOR1 gene and the CPT1 gene, a gene under the control of the adiponectin/AdipoR1 pathway, are down-regulated after 48 hours of ZIKV infection. (c) ZIKV controls adiponectin receptor expression. Expression of the ADIPOR1 and ADIPOR2 genes was normalized to the RNAPII housekeeping gene. Values represent the mean and the standard deviations of three independent experiments. ns = not significant, * p < 0.05; ** p < 0.001; *** p < 0.0002; **** p < 0.0001.
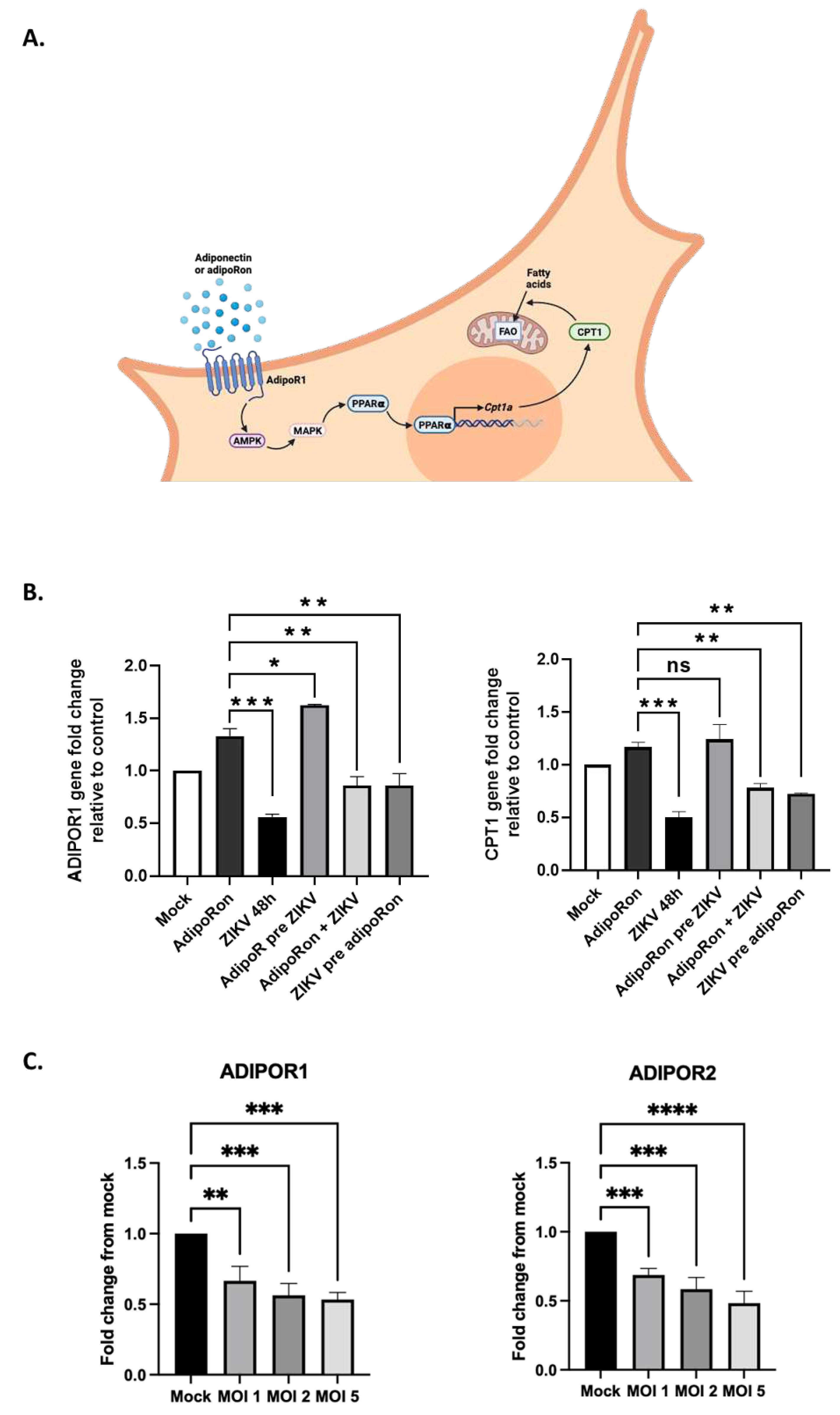
Figure 6.
Hypotheses for antiviral modes of action of AdipoR1/AMPK signaling and its PGC-1α and CPT1 axis on infection and ZIKV's ability to interfere with these pathways.
Figure 6.
Hypotheses for antiviral modes of action of AdipoR1/AMPK signaling and its PGC-1α and CPT1 axis on infection and ZIKV's ability to interfere with these pathways.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



