
Submitted:
09 November 2023
Posted:
10 November 2023
You are already at the latest version
Abstract
In Poland, the mountainous and hilly regions until recently were territories with unbalanced and poor diet quality, especially in early spring. Wild edible plants were or still are influential in supplementing a human diet. One of the earliest wild edible plants to appear in the growing season, characteristic of a broad swath of Europe from Ukraine to Germany, remains wild (bear) garlic (Allium ursinum L.). This paper assesses the impact of the vegetation site and preservation methods on selected parameters of the chemical composition of bear garlic. In addition, the connection between its genome pattern and chemical composition was assessed. The plants were collected in three locations in the Carpathian foothills, and the reference sample was plants collected in the lowlands. The site from which the raw material was harvested has a statistically significant effect on the chemical composition of the preserved leaves of bear garlic. The profiles of the analysed components differed in the material from a different site. Bear garlic leaves from mountainous areas had more analyzed components than those obtained from the lowland sites. Moreover, the material from the lowlands had a different genetic pattern from the leaves from the mountainous areas..

Keywords:
wild garlic
; freezing
; air-drying
; freeze-drying
; genome pattern
; polyphenol
; antioxidant activity
1. Introduction
Plant-based foods play an increasingly important role in modern consumer live styles due to changing behaviour and eating habits. The reasons for these changes vary; for example, changes in worldview that preclude the consumption of any or some animal products or beliefs related to the perception that raising animals for food causes more significant environmental damage than growing plants. Many consumers are also open to current nutritional recommendations that call for increasing the proportion of plant products in everyday food, mainly fruits and vegetables. It may be easier to change eating habits by expanding the range of available plant products with unknown or forgotten plant species. These sometimes include species that are difficult to cultivate or are unprofitable in large-scale farming.
The food group in question may include plants growing in the natural environment, i.e. in forest areas, wastelands, or abandoned/neglected human settlements. According to FAO [1], this group of plants is referred to as wild edible plants. Such plants are usually very well known to local communities, especially in low-income areas, as they often supplement people's daily food. Such a situation was common in the past and now can be observed in poorly developed agricultural mountainous regions [2,3]. In Europe, an example is the Carpathian Mountains and their foothills. Therefore, the experiences of the people living in these areas can be used to promote plant foods by popularising forgotten edible plant species.
One of the most characteristic plants for the areas mentioned above is wild (bear) garlic (Allium ursinum L.). Wild garlic belongs to the economically important genus and subgenus Allium. Almost all of whose species are present in temperate Europe, the Mediterranean region, the Near and Middle East, the Caucasus, and northern Asia, with only a few species being found outside this area [4,5]. In the wild, it grows on the floor of wet, usually mountainous forests [6,7].
The characteristic and dominant flavour of Allium and its possible nutritional effects and medical applications have attracted food and medical researchers [8]. Wild garlic's properties are similar to those of common cultivated garlic. Primarily, the plant's large, green leaves are gathered. There has been a growing interest in using wild garlic as a functional food and dietary supplement. Plants of the genus Allium, including wild garlic, contain active biological substances, phenolic and sulphur compounds, that are potentially useful for humans and animals [9,10].
Wild garlic is an interesting food product due to its early vegetation (March/April). It is one of the earliest spring plants in forests and wild areas near forests. It is also increasingly grown in home gardens and is used in the local cuisine. European people consume wild garlic as a fresh or processed vegetable or condiment. The leaves are added mainly for salads, soups, and as a component of pie fillings [11]. In addition to its culinary uses, wild garlic is also increasingly used as an additive to processed food products. In Poland, Turkey, and Czechia, the leaves are added to local rennet and camembert cheeses [12,13].
This study aimed to compare the genome, and chemical composition of the wild garlic (Allium ursinum L.) leaves collected in four locations in southern Poland.
2. Materials and Methods
2.1. Material
The research material consisted of whole leaves (the above-ground part) of wild (bear) garlic. Fresh material was collected from land belonging to private owners located in four sites in Poland, ranging from 200 to 600 meters above sea level: Krakow (K) – an old orchard at 200 m; Limanowa (L) – a privately-owned forest at 400 m; Sucha Beskidzka (S) – a private woodland at 500 m, and Ropa (R) – a private forest at 600 m. The last three places are situated in the West Beskid Mountains, a part of the West Outer Carpathians. Before analysis, the material was shock-frozen and dried using convection and freeze-drying. It was processed into frozen and dried products immediately after harvesting. The products and chemical determinations were made at the Department of Plant Product Technology and Nutrition Hygiene at the University of Agriculture in Krakow. The DNA analysis was done at the Department of Genetics and Plant Breeding, Faculty of Agrobiology and Food Resources, Slovak University of Agriculture in Nitra.
The leaves were harvested before the plant flowered in late March and early April. After picking, the fresh material was packed into a cooled box and transferred to the laboratory within 60–120 min. The raw material was processed on the same day. As part of preparing the raw material for freezing and drying, the leaves were washed in cold tap water. Surface water was removed by gentle centrifugation in a leafy-vegetable centrifuge and ambient air blowing from a fan. Any leaves that were mechanically damaged or diseased during these actions were removed. They were frozen by a convective shock freezer, a jet of cold air (-40 °C). Before freezing, the leaves were packed in polyethylene bags in batches of about 300 g and pressed to form a layer 1–1.5 cm thick. The bags were frozen for about 1.5 hours until the temperature reached -25 °C inside the package.
Convection drying was carried out in a ProfiLine-type chamber dryer with airflow parallel to the sieves (Hendi, Netherlands). The charge of the material was 2 kg per 1 m2 of the screen. The drying temperature was 40±1 °C, and the time was about 40 hours until the humidity reached 10%. Freeze-drying was performed in a Gamma 1-16 LSC freeze dryer (Christ, Germany). Before placing the garlic leaves into the freeze dryer, they were air-cooled in the freeze dryer trays in a shock freezer (as described above) to speed up the drying. They were lyophilised with the following parameters: an initial product temperature of -30 °C, a condenser temperature of -52 °C, a shelf temperature during actual drying of 20 °C, a shelf temperature while drying of 30 °C, and a drying time of 6 h. Drying was carried out for 24 hours until the moisture content was about 5%.
2.2. Methods
The gravimetric method determined the dry matter (DM) content according to the AOAC standard [14]. The analysed material was ground with double-distilled water (1:1). The samples were dried in an air dryer to a constant weight at a temperature of 98 °C.
Vitamin C was measured by high-performance liquid chromatography. The analysed material was prepared using the PN-EN 14130 [15] standard. Vitamin C was determined as the sum of L-ascorbic acid and dehydroascorbic acid, using L-cysteine to reduce dehydroascorbic acid. Analysis was performed on a La-Chrome Merck-Hitachi liquid chromatograph (Tokyo, Japan). The set was equipped with an L-7612 degasser, programmable L-7250 autosampler, L-7100 pump, L-7360 column oven thermostat, L-7420 UV-VIS detector, D-7000 interface with D-7000 HPLC System Manager software and a Bionacom Velocity C18 PLMX (4.6 × 250 mm, 5 µm) Bionacom Ltd. (UK) column with a pre-column. Isocratic elution was run with a flow of 0.6 ml/min of 0.1% metaphosphoric acid solution as a mobile phase. The measurement was done at a wavelength of 254 nm. An external standard was used for the identification and quantitative analysis of vitamin C: L-ascorbic acid dissolved in 20% metaphosphoric acid at concentrations of 1.0–10.0 µg/ml.
The colourimetric method determined the content of chlorophyll a and b and β-carotene [16]. A sample of the material was homogenised with a mixture of acetone and hexane (4:6) for 2 min. The entire homogenate was transferred to a 25-mL volumetric flask with the same solvents up the mark and centrifuged at 4 °C, 5000 rpm, for 10 min. After centrifugation, the absorbance (A) was measured at four wavelengths: 453, 505, 645, and 663 nm. The content of individual compounds was calculated from the following equations:
Chlorophyll a = 0.999A663 – 0.0989A645 [mg/100 ml],
Chlorophyll b = -0.328A663 + 1.77A645 [mg/100 ml],
β-carotene = 0.216A663 – 0.304A505 + 0.452A453 [mg/100 ml].
In order to determine total polyphenols (TP) and antioxidant activity, sample extracts were prepared using 80% ethanol. Polyphenols were determined by the Folin-Ciocalteu method [17], according to which the Folin–Ciocalteu reagent and 25% sodium carbonate were added to the extract, diluted with deionised water. After 60 min, the absorbance was measured at 765 nm. The content of polyphenols was calculated from the standard curve prepared for gallic acid expressed as gallic acid equivalent (GAE) g/100 g DM.
The antioxidant activity was determined using ABTS, DPPH, and FRAP assays. The evaluation of this parameter against the ABTS radical (2,2'-azino-bis- [3-ethylbenzothiazoline-6-sulfonic acid) was carried out according to Re et al. [18]. The determinations were carried out in 80% ethanol extracts at 734 nm after 10 min. The results are given in μmol of Trolox (2-carboxyl-6-hydroxy-2,5,7,8-tetramethylchroman, a water-soluble analogue of α-tocopherol) in 1 g of the sample. In turn, in the second determination, the ability to reduce the persistent free radical DPPH' (1,1-diphenyl-2-picrylhydrasyl) of an 80% ethanol solution was analysed (Pekkarinen et al. 1999) [19]. The absorbance was measured at a wavelength of 516 nm. The activity level was determined in µmol of Trolox (2-carboxy-6-hydroxy-2,5,7,8-tetramethylchroman) in 1 g. The antioxidant activity against the FRAP (ferric reducing antioxidant power) was defined as the ability to reduce iron, and it was measured in µmol of Fe2+ in 100 g of DM [20]. TPTZ reagent (iron-2,4,6-tripyridyl-S-triazine complex) was added to the extracts and incubated for 10 min in a water bath at 37 °C. After this time, the absorbance was measured at 595 nm.
2.3. DNA Analysis
2.3.1. DNA Extraction
Total genomic DNA was isolated from the lyophilised samples of wild garlic leaves (Allium ursinum) with the GeneAll Exgene™ isolation kit (Cambio) following the manufacturer's instructions (Plant SV). One sample originated from an urban area of Krakow (sample K), and the others were grown in different mountainous regions of Poland (samples R, L, and S). The quantity and quality of the extracted DNA were measured with a P-Class NanoPhotometer® (Implen).
2.3.2. CDDP and PBA Marker Assay
DNA markers based on conserved regions of DNA or gene families were used in the study. PBA primer combinations (CYPA1F+CYPA1R; CYP2BF+CYP2BR; CYP2CF+CYP2CR) were used as designed by Inui et al. [21]. CDDP primer combinations (WRKY F/R1; WRKY F/R2; WRKY F/R2b; WRKY F/R3; WRKY F/R3b) were used as designed by Xie et al. [22]. Polymerase chain reactions (PCRs) were performed with MyTaq 2x master mix (Bioline) with 100 ng of DNA and 320 nmol of each primer. Amplification reactions were performed in a C1000 thermocycler (BioRad) with the following temperature and time profile: 95 °C for 5 min (95 °C for 60 s; 50 °C for 60 s; 72 °C for 120 s) 45 times, plus 72 °C for 10 min for the final elongation.
2.3.3. Microsatellite Markers Assay
The extracted DNA was diluted to a concentration of 10 ng/µl. The microsatellite loci were amplified by two combinations of primers, amplifying the (AG)9 and (AG)16 repeat motifs [23]. The PCRs were performed in 25 µl, containing 2.5 µl of 10× Dream Taq reaction buffer (plus 50 mM of MgCl2) (Thermo Scientific), 0.5 µl of a mix of all four dNTPs (10 mM each), 0.5 µl forward and reverse primer (10 µM), 0.1 µl
Dream Taq DNA polymerase (5 U/µl ), 1.0 µl of bovine serum albumin (10 mg/ml), 1.0 µl of 10 ng/µl genomic DNA, and sterilised water up to a final volume of 25 µl.
PCR reactions were performed using a C1000 Thermal Cycler (Biorad, USA) under the following conditions: initial denaturation at 94 °C for 3 min; 40 cycles of 94 °C for 30 s, 55 °C for 45 s, and 72 °C for 1 min; 8 cycles of 94 °C for 30 s, 53 °C for 45 s, and 72 °C for 1 min; and a final extension step of 72 °C for 5 min. The resulting PCR products were run on 1.5% agarose gel together with the size standard 100 bp Ready-to-use DNA Ladder (Bioron). The fragments were scored by GeneTool software (Syngene).
2.3.4. Direct Sequencing
Standard barcoding primers were used for the amplification [24]. The PCR amplification of the ITS regions was carried out in a SureCycler 8800 thermal cycler (Agilent). The PCRs were performed with Phusion High-Fidelity PCR Master Mix (Thermo Scientific) under the following conditions: 98 °C for 30 s (98 °C for 10 s, 55 °C for 40 s, and 72 °C for 40 s) 35 times, then 72 °C for 10 min final elongation. After the control display of the specificity of amplicons on 1.5% agarose gel, the product was purified with an ExpinTM Combo Kit (GeneAll) according to the manufacturer's instructions. GenSeq sequenced the ITS amplicons.
2.3.5. Data Analysis
The amplified fragments were separated in 6% PAGE stained by GelRed (Biotium); they were analysed online by GelAnalyser software. The amplified loci were scored by 0–1 matrices and compared for amplification patterns. ITS raw sequences were trimmed and analysed by BioEdit software. Individual sequences were BLASTed against Allium sp. taxa in the NCBI database.
3. Results and Discussion
The frozen leaves of wild garlic after the harvest had a dry matter (DM) content ranging from 9.8–12.6 g/100 g of fresh matter (FM) (Table 1). The leaves obtained in Krakow and Sucha Beskidzka had significantly higher DM content than those from Limanowa and Ropa. The last two had amounts similar to the value reported by Jędrszczyk et al. [25] and Leahu [26], 9.79–10.77 g/100 g and 9.75 g/100 g, respectively; these levels were higher than that found by Gîtin et al. (2012). However, according to Błażewicz-Woźniak [27], the DM content in wild garlic leaves (6.69–15.7 g/100 g FM) largely depended on the ecotype and the year of cultivation. After drying to a constant weight, the air-dried leaves contained 85.5–92.7 g of DM, and the freeze-dried leaves had 94.3–96.9 g of DM per 100 g of FM. Tomšik et al. [28] were able to dry wild garlic leaves up to 91.19 g and 92.03 g of DM per 100 g of FM using air at 40 °C and 60 °C, respectively.
Plant pigments, based on their chemical structure, can be classed into five families: tetrapyrroles (e.g. chlorophyll), carotenoids (e.g. β-carotene), phenolic compounds (e.g. theaflavins), and N-heterocyclic compounds (e.g. betalains) [29]. Chlorophyll a and chlorophyll b are essential pigments of plant photosystems. Its concentration in leaves is a crucial parameter of the plants' photosynthetic mechanism and metabolism. Chlorophyll production mainly depends on the intensity of sunlight and the nitrogen content [30]. Chlorophylls and carotenoids are responsible for the colour of plants and are even used as parameters of maturity, quality, and freshness of food crops. The amount of chlorophyll in the leaves is influenced by many environmental factors [31]. The degradation of chlorophyll pigments during processing causes colour changes in the product; a change from green to yellow or red indicates that the carotenoids were uncovered by the destruction of chlorophyll [32]. Chlorophyll is used as a photosensitiser for cancer therapy. The pigment and its derivatives have potent antioxidant and radioprotective effects. They inhibit lipid peroxidation, protein oxidation, DNA damage, and membrane damage [33].
In the frozen and dried leaves of wild garlic, the sum of chlorophyll a and b was 638–1532 mg/100 g DM, depending on the location and processing methods. The fraction of chlorophyll a ranged from 68% to 74% (Table 1), and the chlorophyll a:b ratio was 2–3:1, which is consistent with a study by Srichaikul et al. [34]. For comparison, Luta et al. [35] reported 174 mg of chlorophyll/100 g FM in fresh wild garlic leaves. Regardless of the processing method, the highest level of total chlorophyll was in the frozen material collected in the Limanowa region (a mountainous area), an average of 19%–89% more; the lowest content was in the leaves collected near Sucha Beskidzka (also mountainous). Among the preservation methods, freezing turned out to be the best and air-drying the worst way to protect the chlorophylls in garlic leaves. On average, the frozen product contained 15% more chlorophylls than the air-dried product. The decomposition of these pigments likely occurred due to the thin wild garlic leaves' tissue aeration and exposure to high temperatures (40 °C) for tens of hours. According to Rocha et al. [36], a short steam blanching (15 s) can reduce the degradation of chlorophyll in basil leaves dried at high temperatures (60 °C). Still, if the samples were not blanched before drying, drying at low temperatures was the best method to prevent the degradation of chlorophylls.
In wild garlic leaves, 155–221 mg of β-carotene per 100 g of DM was measured (Table 1). The most considerable amount of this discriminant was found in the leaves collected near Limanowa (a mountainous area). At the same time, in the case of other frozen products, the location did not affect the amount of β-carotene. The preservation method usually had little effect on the level, with the best process being freeze-drying. For comparison, Luta et al. [35] found only 5.3 mg of total carotenoids in 100 g of fresh wild garlic leaves, using 80% acetone for the extraction. In turn, Stajner et al. [37] showed that cultivated leaves of Allium vienale contain over two times more chlorophyll a and chlorophyll b in 100 g of FM than wild growing. Carotenoids are essential not only for the colouration of food but also for their acknowledged roles in health. They are vitamin A sources and a critical factor for human vision. In addition, they have an essential role in preventing various diseases associated with oxidative stress, such as cancer, cardiovascular diseases, and other chronic diseases [38,39].
Vitamin C is one of the most critical water-soluble vitamins with different essential biological functions. It is one of the most active antioxidants, which helps the body neutralise free radicals. It takes part in the biosynthesis of collagen, carnitine, hormones, immune response, and iron absorption. It is vulnerable to light, air, and high temperature and decomposes completely at approximately 190 °C [40]. The content of vitamin C in fruits, vegetables, and herbs is highly variable and dependent on many factors: species, variety, climate, a season of the year, method of storage after harvest, and country of origin [41]. Stajner et al. [37] noted that cultivated leaves of Allium vienale contain over three times as much vitamin C (18 mg/100 g FM) as wild-growing varieties. In turn, Jędrszczyk et al. [25] showed that the amount of vitamin C in wild garlic depends on the part of the plant because the leaves and stems contain 7–15 and 4–6 mg of vitamin C/100 g FM, respectively. In the wild garlic leaves analysed in this study, 20–1673 mg of vitamin C/100 g DM were measured, depending on the harvest site and the method of preservation (Table 2). For comparison, in fresh wild garlic leaves, Luta et al. [35] found only 80 mg of vitamin C per 100 g of FM. They determined the amount of vitamin C using titration, not chromatography.
The most vitamin C was found in the frozen material collected in the vicinity of Sucha Beskidzka (a mountainous area), whilst the lowest was in the material from Krakow (an urban area). Among the drying methods used, air-drying was the least favourable, usually resulting in almost complete decomposition of vitamin C. Vitamin C's low resistance to high temperatures might be related to its chemical structure [41]. Wolska et al. [42] noted that the brewing temperature significantly impacted the vitamin C content in infusions of nettle leaves. Brewing temperatures of 25–50 ℃ were more favourable than 70–80 ℃, and the optimal dewatering time was 10 min. According to Lee and Kader [43], depending on the species, L-ascorbic acid (AA) accounted for 52%–93% and 73%–100% of the vitamin C in fruits and vegetables, respectively. In the product prepared from wild garlic leaves, the vitamin C contained up to 45%–50%, 25%–75%, and 41%–54% L-ascorbic acid in frozen, air-dried, and freeze-dried leaves, respectively; the rest was L-dehydroascorbic acid (DHAA). As a rule, a minor proportion of AA was recorded in leaves collected in Krakow (an urban area).
The total polyphenol (TP) content in frozen wild garlic leaves obtained from the four sites was in the range of 1.64–1.95 g gallic acid equivalent (GAE)/100 g DM (Table 3). However, no difference was found between the frozen product gathered in Krakow and Sucha Beskidzka, whilst the product from Limanowa and Ropa contained significantly more of these compounds. The observed TP content was higher than that previously found in fresh leaves harvested in Poland – 1.48 g/100 g DM [44] – but they were in the range of 1.31–2.11 g/100 g reported in garlic leaves from Montenegro and Bosnia and Herzegovina [45]. These values, on the other hand, were lower than those of the leaves of wild garlic harvested in Serbia [46,47,48] – 0.92–6.05 g/100 g DM, depending on the solvent used for extraction – and of garlic cultivated in Poland [25] – 2.78–3.83 g/100 g DM. According to Gîtin et al. [49] and Pop et al. [50], there was even 24.6–27.0 g TP/100 g DM in garlic leaves.
The TP content in dried wild garlic leaves was 1.41–2.14 g/100 g DM. Compared to frozen leaves, air drying resulted in a slight increase in TP content in the leaves from Ropa and a decrease in leaves from Sucha Beskidzka. In contrast, the freeze-dried leaves of plants showed a reduction in TP content regardless of the place of harvest. The decline was not statistically significant in the case of dried products from raw materials obtained in Krakow. According to Tomšik et al. [28], the TP content in frozen and dried leaves was much higher (9.18–10.10 /100 g DM) than in this study.
The antioxidant activity of wild garlic depends on the harvest date. Jędrszczyk et al. [25] found that the antioxidant activity against DPPH differed almost twice between the harvest years. Similarly, Lachowicz et al. [51] found higher antioxidant activity against ABTS and DPPH but lower using the FRAP method in leaves obtained in June than in March. Kovarovič et al. [52] found that the antioxidant activity of wild garlic leaves against the DPPH radical is higher than that of white and yellow onion, leeks, and common garlic but half that of red onion. Krivokapič et al. [48] demonstrated that the antioxidant activity (IC50) of wild garlic leaves against DPPH was in the range of 111–391 μg/mL of the extract, depending on the solvent used for extraction. Similar conclusions were reached by Pop et al. [50]. According to Tomšik et al. [28], wild garlic leaves dried at 40 °C, and 60 °C did not differ in their antioxidant activity against DPPH.
The antioxidant activity measured against the cation radical ABTS in the frozen leaves was at 347–511 µmol Trolox/g DM; the lowest level was observed in the leaves harvested in Krakow, whilst the highest was in those from Limanowa. The drying contributed to a significant increase in the antioxidant activity – 2.8 to 5.4 times more – compared to frozen leaves. These results were significantly higher in air-dried leaves than in freeze-dried leaves.
The antioxidant activity measured against the free radical DPPH in frozen leaves was 11–27 µmol Trolox/g DM. The dried leaves had a higher level of this index (36–131 µmol Trolox/g DM), and, as in the case of the measurements against ABTS, the freeze-dried leaves had higher antioxidant activity than air-dried ones.
The ferric-reducing antioxidant power (FRAP) of frozen wild garlic leaves was 707–798 µmol Fe2+/g DM. After drying, a significant increase in this indicator was found, in comparison to frozen: 47%–102% more in air-dried and 46%–81% more in freeze-dried. The advantage of air-drying was confirmed only for leaves from Limanowa and Ropa, while in the case of freeze-drying in leaves from Sucha Beskidzka. The drying method did not affect the antioxidant activity only in the leaves harvested in Krakow.
CDDP and PBA marker profiling
Different marker systems were developed within the trend toward gene-targeted functional markers rather than markers of non-coding regions. The conserved DNA-derived polymorphism technique (CDDP) and P450-based analogue markers (PBA) target candidate plant genes [21,53]. Similar to RAPD and ISSR techniques, the method is based on the single primer amplified region principle, as a single primer is used as a forward and reverse primer.
Different CDDP and PBA fingerprint profiles were obtained in accessions from the Krakow control sample and samples from the mountainous areas (Table 4). Monomorphic profiles with no fingerprint changes were only for two primer combinations of CDDP analyse.
Barcoding analysis of ITS region
Primers for barcoding analyse were selected for their putative amplification of the DNA sequences that codify for small- (18S, SSU) and large-subunit (28S, LSU) ribosomal RNA (rRNA). The primers were selected for their previous use; they are considered standard primers for amplifying these regions in various species of fungi and higher plants [24,54]. According to many authors, thanks to their variability, ITS regions yielded sufficient molecular information, which can be directly used as identifiers or molecular markers in the phylogenetic analysis [54,55,56]. In our case, the ITS product was variable in some nucleotide positions when compared to the ITS sequence of Allium ursinum stored in the NCBI database (Figure 1). For comparison purposes, registered NCBI database sequences were used, and their sequences were compared with the BLAST algorithm.
Microsatellite markers assay
Based on Kumar et al. [57], it was expected that clonal propagation would have little effect on the level of variation in the population structure. In our study, there were differences in the DNA fingerprinting of the garlic samples originating from different environmental conditions (Figure 2). Our results are consistent with the variability in Indian garlic germplasm consisting of accessions obtained from public research organisations and farmers' markets (natural landraces). Similarly, cluster analysis based on microsatellite markers grouped garlic accessions according to their flowering behaviour, botanical variety, and ecophysiological characteristics [58]. The potential of SSR markers in plant germplasm evaluation is also supported by the study of da Cunha et al. [59], where hierarchical clustering (UPGMA dendrogram) analysis reflected the classification of garlic accessions according to maturity time (growth cycle).
4. Conclusions
- Regardless of the processing method, the highest levels of chlorophylls, β-carotene, and vitamin C were found in the frozen product from raw material collected in Limanowa and Sucha Beskidzka mountainous areas.
- The preservation method significantly impacted chlorophyll and vitamin C content and less on β-carotene content. The best way for preserving chlorophyll content was freezing; freeze-drying was best for β-carotene. In the case of vitamin C, air drying resulted in almost complete decomposition. In contrast, freezing and freeze-drying leaves only resulted in significant differences in the raw material collected in the Sucha Beskidzka (mountainous area), where freezing became the better choice.
- There was no difference in the total polyphenol content in the frozen leaves between those obtained in Krakow and Sucha Beskidzka. In contrast, the frozen leaves harvested from Limanowa and Ropa contained significantly more of these compounds. Convection drying of the leaves resulted in a slight increase in TP content in the leaves from Ropa and a decrease in the leaves from Sucha Beskidzka, compared to the frozen leaves. In contrast, lower TP content was found in the freeze-dried leaves regardless of the collection site. Drying contributed to a significant increase in antioxidant activity in comparison to freezing. The results were significantly higher for the air-dried leaves than the freeze-dried ones.
- Marker analysis of coding region-based CDDP and PBA techniques showed sequence variability in the samples from the mountainous areas – Ropa, Limanowa, and Sucha Beskidzka – compared to the sample from Krakow. One of the reasons for this variability might be environmental conditions.
- Taking into account the level of total polyphenols and antioxidant activity, the best preservation method was usually air drying; in the case of vitamin C and chlorophylls, it was freezing; and in the case of β-carotene, freeze-drying was best.
- The leaves collected in the Limanowa area were characterised by the highest level of total polyphenols, chlorophylls, β-carotene, and antioxidant activity (FRAP), and those picked in the Sucha Beskidzka area had the highest levels of vitamin C.
Author Contributions
Conceptualization and methodology, P.G., E.B. and J.S.; validation, E.B., J.S., K.R, and J.Ž.; material preparation, P.G., E.B. and J.S.; formal analysis and investigation, P.G., E.B., J.S., K.R, and J.Ž.; data curation, E.B., J.S., K.R, and J.Ž.; writing - original draft preparation, visualization, review and editing, P.G., E.B., J.S., K.R, and J.Ž. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Polish National Agency for Academic Exchange in the project PPI/APM/2018/1/00010/U/001.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maurer, M.; Schueckler, A. Eds. Use and potential of wild plants in farm households, FAO, Rome, 1999. Available online: http://www.fao.org/3/W8801E/w8801e00.htm#toc_00.
- Bvenura, C.; Sivakumar, D. The role of wild fruits and vegetables in delivering a balanced and healthy diet. Food Res Int 2017, 99, 15. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, S.; Ahmad, M.; Haroon, M. Edible wild plants: A solution to overcome food insecurity. Springer, Cham, Switzerland, 2017. [CrossRef]
- Odeny, D.A.; Narina, S.S. Allium. In: Wild crop relatives: Genomic and breeding resources; Kole C. Ed.; Springer, Berlin, Heidelberg, Germany, 2011. pp. 1-10. [CrossRef]
- Khassanov, F.O. Taxonomical and ethnobotanical aspects of Allium species from Middle Asia with particular reference to subgenus Allium. In The Allium genomes. Compendium of plant genomes; Shigyo, M.; Khar, A.; Abdelrahman, M. Eds.; Springer, Cham, 2018, 11-21. [CrossRef]
- Rola, K. Taxonomy and distribution of Allium ursinum (Liliaceae) in Poland and adjacent countries. Biol 2012, 67, 1080–1087. [Google Scholar] [CrossRef]
- Peinado, M.; Aguirre, J.L.; Aparicio, A. The Iberian ranges and highlands. In: The vegetation of the Iberian peninsula. Plant and vegetation, vol. 12; Loidi, J. Ed.; Springer, Cham, Switzerland, 2017; pp. 439-512. [CrossRef]
- Bisen, P.S.; Emerald, M. Nutritional and therapeutic potential of garlic and onion (Allium sp.). Curr Nutr Food Sci 2016, 12, 1–10. [Google Scholar] [CrossRef]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Bursać Kovačević, D. An overview of organosulfur compounds from Allium spp.: from processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem 2019, 276, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Skrovankova, S.; Snopel, L.; Mlcek, J.; Planetova, T. Polyphenols and antioxidant capacity in different types of garlic. Potravinarstvo Slov J Food Sci 2018, 12, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Barla, G.; Poroch-Seritan, M.; Sanduleac (Tudosi), E.; Ciornei (Stefaroi), S.E. Antioxidant activity and total phenolic content in Allium ursinum and Ranunculus ficaria. Food Env Saf 2014, 13, 349. [Google Scholar]
- Apak, R. Current issues in antioxidant measurement. J Agric Food Chem 2019, 67, 9187. [Google Scholar] [CrossRef] [PubMed]
- MRiRW. Produkty regionalne i tradycyjne. (Polish Ministry of Agriculture and Rural Development, Local and traditional products database). Available online: https://www.gov.pl/web/rolnictwo/produkty-regionalne-i-tradycyjne (accessed on 10 May 2023).
- AOAC. Official methods of analysis, 14th ed.; Association of Official Chemists, Arlington, VA, USA, 1984.
- PN EN 14130:2004. Foodstuffs - determination of vitamin C by HPLC. Polish Commite of Normalisation, Warsaw, Poland.
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shok Kog Gakk 1999, 39, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Bio Med 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pekkarinen, S.S.; Heinonen, I.M.; Hopia, A.I. Flavonoids, quercetin, myrcetin, kaempferol and (+) catechin and antioxidants in methyl linoleate. J Sci Food Agr 1999, 79, 499–506. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ”Antioxidant Power”: The FRAP assay. Anal Biochem 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Inui, H.; Komada, T.; Ohkawa, Y.; Ohkawa, H. Herbicide metabolism and cross-tolerance in transgenic potato plants co-expressing human CYP1A1, CYP2B6 and CYP2C19. Pestic Biochem Physiol 2000, 66, 116–129. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, Z.L.; Zou, X.; Huang, J.; Ruas, P.; Thompson, D.; Shen, Q.J. Annotations and functional analyses of the rice WRKY gene superfamily reveals positive and negative regulators of abscisic acid signaling in aleurone cells. Plant Physiol 2005, 137, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Mansour, H.; Sliwinska, E. Genetic diversity and inbreeding level of Cotoneaster orbicularis Schltdl. in the Sinai Mountains revealed by microsatellite markers and flow cytometry. Egypt J Bot 2017, 57, 351–361. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR Protocols: a guide to methods and applications Innis, M.A.; Gelfand, D.H.; Sninsky, J.J.; White, T.J. Eds.; Academic Press, New York, USA, 1999, pp. 315–322.
- Jędrszczyk, E.; Kopeć, A.; Bucki, P.; Ambroszczyk, A.M.; Skowera, B. The enhancing effect of plants growth biostimulants in garlic cultivation on the chemical composition and level of bioactive compounds in the garlic leaves, stems and bulbs. Not Bot Horti Agrobot 2019, 47(1), 81. [Google Scholar] [CrossRef]
- Leahu, A.; Damian, C.; Oroian, M.; Juravle, L.; Ropciuc, S. Physico-chemical and antioxidant properties of two medicinal wild plants grown in Moldova region. Sci Pap Anim Sci Biotech 2015, 48(1), 382–388. [Google Scholar]
- Błażewicz-Woźniak, M.; Michowska, A. The growth, flowering and chemical composition of leaves of three ecotypes of Allium ursinum L. Acta Agrobot 2011, 64(4), 171–180. [Google Scholar] [CrossRef]
- Tomšik, A.; Radojčin, M.; Stamenković, Z.; Kevrešan, Ž.; Mastilović, J.; Pavkov, I.; Vidović, S. Convective drying and preservation of functional ingredients of wild garlic (Allium ursinum L.) in dependence of drying temperature. In: Preceding’s of the III International Congress on Food Technology, Quality and Safety, 25-27 October 2016, University of Novi Sad, Institute of Food Technology, Novi Sad, Serbia, pp. 646-650.
- Schoefs, B. Chlorophyll and carotenoid analysis in food products. Properties of the pigments and methods of analysis. Trends Food Sci Tech 2002, 13, 361–371. [Google Scholar] [CrossRef]
- Padilla, F.M.; Peña-Fleitas, M.T.; Gallardo, M.; Giménez, C. Derivation of sufficiency values of a chlorophyll meter to estimate cucumber nitrogen status and yield. Comput Electron Agr 2017, 141, 54–64. [Google Scholar] [CrossRef]
- Çetİn, M. Changes in the amount of chlorophyll in some plants of landscape studies. Kastamonu Üniv J Forest Fac 2016, 16(1), 239-245.
- Marty-Audouin, C.; Leber, A.; Rocha-Mier, T. Influence of drying on the colour of plant products. In: Developments in drying: Food dehydration (Vol. 1); Mujumdar, A.S.; Sirikayala, S. Eds.; Kasetsart University Press, Bangkok, Thailand, 1999, pp. 207-233.
- Mishra, V.K.; Bacheti, R.K.; Husen, A. Medicinal uses of chlorophyll: a critical overview. In: Chlorophyll: Structure, function and medicinal uses. Le, H.; Salcedo, E. Eds.; Nova Science Publishers, Inc., Hauppauge, NY, USA, 2011, pp. 177-196.
- Srichaikul, B.; Bunsang, R.; Samappito, S.; Butkhup, S.; Bakker, G. Comparative study of chlorophyll content in leaves of Thai Morus alba Linn. species. Plant Sci Res 2011, 3, 17–20. [Google Scholar] [CrossRef]
- Luta, G.; Gherghina, E.; Balan, D.; Israel-Roming, F. Bioactive compounds and antioxidant properties of some wild plants with potential culinary uses. Rev Chim 2020, 71(2), 179–184. [Google Scholar] [CrossRef]
- Rocha, T.; Marty-Audouin, C.; Lebert, A. Effect of drying temperature and blanching on the degradation of chlorophyll a and b in mint (Mentha spicata Huds.) and basil (Ocimum basilicum): analysis by high performance liquid chromatography with photodiode array detection. Chromatographia 1993, 36(1), 152-156. [CrossRef]
- Štajner, D.; Milić, N.; Mimica-Dukić, N.; Lazić, B.; Igić, R. Antioxidant abilities of cultivated and wild species of garlic. Phytother Res 1998, 12(S1), S13–S14. [Google Scholar] [CrossRef]
- Sangeetha, R.K.; Baskaran, V. Carotenoid composition and retinol equivalent in plants of nutritional and medicinal importance. Efficacy of β-carotene from Chenopodium album in retinol-deficient rats. Food Chem 2010, 119, 1584–1590. [Google Scholar] [CrossRef]
- Žnidarčič, D.; Ban, D.; Šircelj, H. Carotenoid and chlorophyll composition of commonly consumed leafy vegetables in Mediterranean countries. Food Chem 2011, 129(3), 1164–1168. [Google Scholar] [CrossRef]
- Davey, M.W.; Montagu, M.V.; Inzé, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J.P. Plant L-ascorbic acid: chemistry, function, metabolism, bioavailability and effects of processing. J Sci Food Agr 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Janda, K.; Kasprzak, M.; Wolska, J. Witamina C – budowa, właściwości, funkcje i występowanie (Vitamin C – structure, properties, occurrence and functions). Pom J Life Sci 2015, 61(4), 419. [Google Scholar]
- Wolska, J.; Czop, M.; Jakubczyk, K.; Janda, K. Influence of temperature and brewing time of nettle (Urtica dioica L.) infusions on vitamin C content. Rocz Panstw Zakl Hig.
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol Tec 2000, 20(3), 207–220. [Google Scholar] [CrossRef]
- Lachowicz, S.; Kolniak-Ostek, J.; Oszmiański, J.; Wiśniewski, R. Comparison of phenolic content and antioxidant capacity of bear garlic (Allium ursinum L.) in different maturity stages. J Food Process Pres 2017, 41, e13089. [Google Scholar] [CrossRef]
- Pejatović, T.; Samardžić, D.; Krivokapić, S. Antioxidative properties of a traditional tincture and several leaf extracts of Allium ursinum L. (collected in Montenegro and Bosnia and Hercegovina). J Mater Environ Sci, 2017; 8, 1929–1934. [Google Scholar]
- Tomšik, A.; Pavlić, B.; Vladić, J.; Ramić, M.; Brindza, J.; Vidović, S. Optimization of ultrasound-assisted extraction of bioactive compounds from wild garlic (Allium ursinum L.). Ultrason Sonochem 2016, 29, 502–511. [Google Scholar] [CrossRef]
- Pavlović, D.R.; Veljković, M.; Stojanović, N.M.; Gočmanac-Ignjatović, M.; Mihailov-Krstev, T.; Branković, S.; Sokolović, D.; Marčetić, M.; Radulović, N.; Radenković, M. Influence of different wild-garlic (Allium ursinum) extracts on the gastrointestinal system: spasmolytic, antimicrobial and antioxidant properties. J Pharm Pharmacol 2017, 69(9), 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Krivokapic, M.Z.; Jakovljevic, V.L.; Sovrlic, M.M.; Bradic, J.V.; Petkovic, A.M.; Radojevic, I.D.; Brankovic, S.R.; Comic, L.R.; Andjic, M.M.; Kocovic, A.G.; Tomovic, M.T. Biological activities of different extracts from Allium ursinum leaves. Acta Pol Pharm Drug Res 2020, 77(1), 121–129. [Google Scholar]
- Gîtin, L.; Dinicâ, R.; Parnavel, R. The influence of extraction method on the apparent content of bioactive compounds in Romanian Allium spp. leaves. Not Bot Horti Agrobot 2012, 40(1), 93–97. [Google Scholar] [CrossRef]
- Pop, R.M.; Bocsan, I.C.; Buzoianu, A.D.; Chedea, V.S.; Socaci, S.A.; Pecoraro, M.; Popolo, A. Evaluation of the antioxidant activity of Nigella sativa L. and Allium ursinum extracts in a cellular model of doxorubicin-induced cardiotoxicity. Molecules. [CrossRef]
- Lachowicz, S.; Kolniak-Ostek, J.; . Oszmiański, J.; Wiśniewski, R. Comparison of phenolic content and antioxidant capacity of bear garlic (Allium ursinum L.) in different maturity stages. J Food Process Pres 2017, 41, e12921. [Google Scholar] [CrossRef]
- Kovarovič, J.; Bystrická, J.; Fehér, A.; Lenková, M. Evaluation and comparison of bioactive substances in selected species of the genus Allium. Potravinarstvo Slovak J Food Sci 2017, 11(1), 702–708. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.; Mackill, D.J. Conserved DNA-derived polymorphism (CDDP): a simple and novel method for generating DNA markers in plants. Plant Mol Biol Rep 2009, 27(4), 558–562. [Google Scholar] [CrossRef]
- Smolik, M.; Malinowska, K.; Smolik, B.; Pacewicz, K. Polymorphism in Random amplified and nuclear rDNA sequences assessed in certain apple (Malus x domestica Borkh.) cultivars. Not Bot Horti Agrobot, 2011; 39, 264–270. [Google Scholar]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol Phylogenet Evol 2003, 29, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Nalini, E.; Bhagwat, S.G.; Jawali, N. Identification and characterization of some ITS variants from hexaploid wheat (Triticum aestivum L.). Plant Sci 2007, 173, 262–268. [Google Scholar] [CrossRef]
- Kumar, M.; Rakesh Sharma, V.; Kumar, V.; Sirohi, U.; Chaudhary, V.; Sharma, S.; Saripalli, G.; Naresh, K R.; Yadav, K.H.; Sharma, S. Genetic diversity and population structure analysis of Indian garlic (Allium sativum L.) collection using SSR markers. Physiol Mol Biol Plants 2019, 25, 377–386. [Google Scholar] [CrossRef]
- Barboza, K.; Beretta, V.; Kozub, P.C.; Salinas, C.; Morgenfeld, M.M.; Galmarini, C.R.; Cavagnaro, P.F. Microsatellite analysis and marker development in garlic: distribution in EST sequence, genetic diversity analysis, and marker transferability across Alliaceae. Mol Genet Genomics 2018, 293, 1091–1106. [Google Scholar] [CrossRef]
- da Cunha, CP,; Resende, FV.; Zucchi, MI.; Pinheiro, JB. SSR-based genetic diversity and structure of garlic accessions from Brazil. Genetica 2014, 142(5), 419-431. [CrossRef]
Figure 1.
Polymorphism analysis of Allium ursinum samples by SSR marker. Highlighted are differences in DNA profile between control and mountainous samples of wild garlic. K - Krakow, control sample; R - Ropa, L - Limanowa, S - Sucha Beskidzka – samples from mountainous areas.
Figure 1.
Polymorphism analysis of Allium ursinum samples by SSR marker. Highlighted are differences in DNA profile between control and mountainous samples of wild garlic. K - Krakow, control sample; R - Ropa, L - Limanowa, S - Sucha Beskidzka – samples from mountainous areas.

Figure 2.
Polymorphism analysis of Allium ursinum samples by SSR marker. Highlighted are differences in DNA profile between control and mountainous samples of wild garlic. K - Krakow, control sample; R, L, S – samples from the mountainous areas.
Figure 2.
Polymorphism analysis of Allium ursinum samples by SSR marker. Highlighted are differences in DNA profile between control and mountainous samples of wild garlic. K - Krakow, control sample; R, L, S – samples from the mountainous areas.
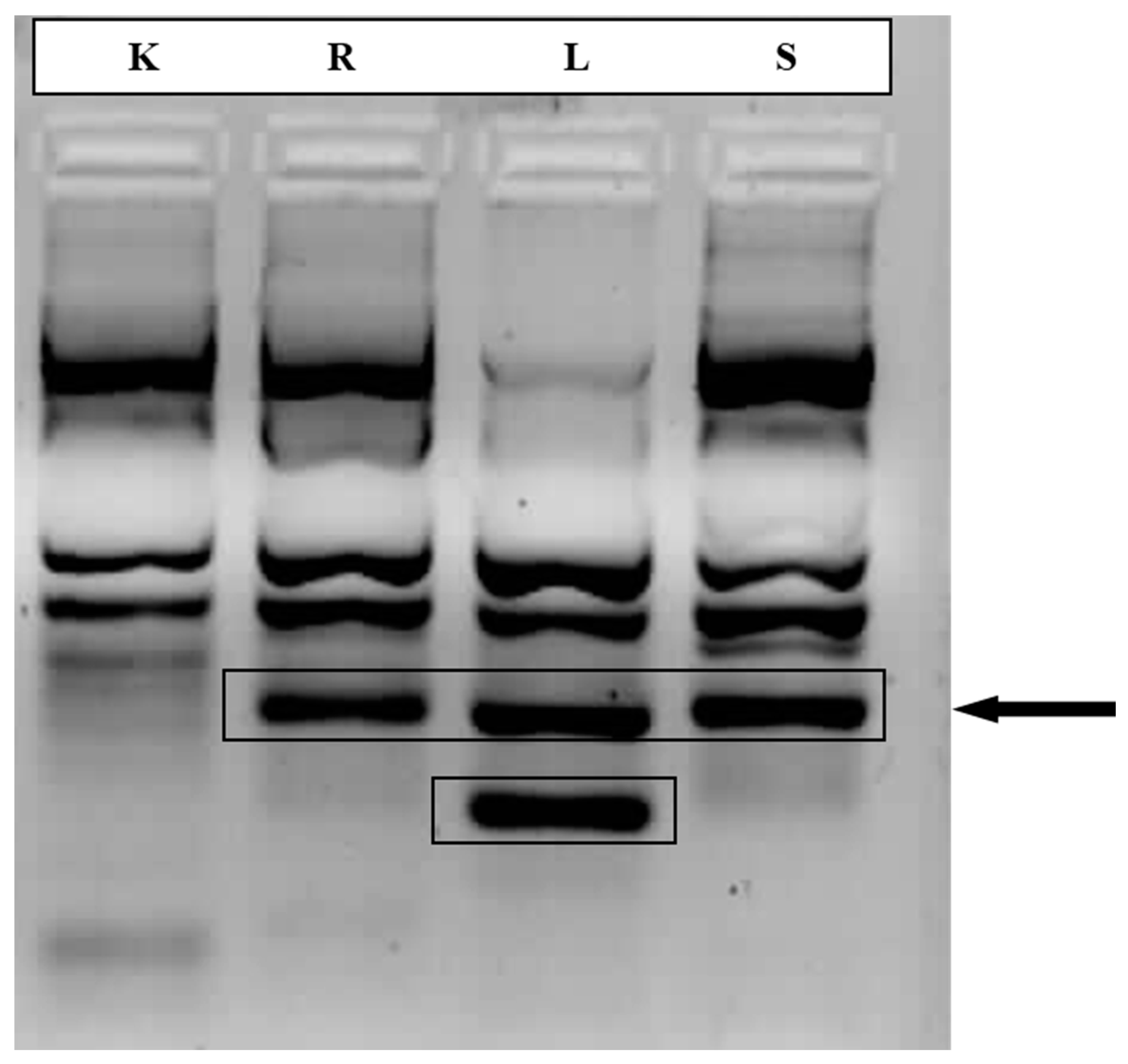

Table 1.
The content of dry matter, chlorophylls, and β-carotene in frozen, air-dried, and freeze-dried wild garlic leaf*.
Table 1.
The content of dry matter, chlorophylls, and β-carotene in frozen, air-dried, and freeze-dried wild garlic leaf*.
| Component | Product | Location | ||||
|---|---|---|---|---|---|---|
| Kraków | Limanowa | Ropa | Sucha Beskidzka |
mean | ||
|
Dry matter g/100 g FM |
frozen | 12.0b | 10.7a | 9.8a | 12.6b | 11.3A |
| air-dried | 91.3e | 87.5d | 92.7f | 85.5c | 89.2B | |
| freeze-dried | 94.5g | 94.3g | 96.9h | 95.4g | 95.3C | |
| mean | 65.9B | 64.2A | 66.4B | 64.5A | ||
|
Chlorophyll a mg/100 g DM |
frozen | 914e | 1081f | 936e | 641b | 893C |
| air-dried | 711c | 1078f | 830d | 450a | 767A | |
| freeze-dried | 817d | 1047f | 920e | 617b | 850B | |
| mean | 814B | 1069D | 895C | 569A | ||
|
Chlorophyll b mg/100 g DM |
frozen | 379fg | 430hi | 414h | 255c | 370B |
| air-dried | 342de | 454i | 324d | 188a | 327A | |
| freeze-dried | 329de | 401gh | 358ef | 220b | 327A | |
| mean | 350B | 428C | 365B | 221A | ||
|
Total chlorophyll (a+b) mg/100 g DM |
frozen | 1293e | 1511f | 1350e | 896b | 1263C |
| air-dried | 1053c | 1532f | 1154d | 638a | 1094A | |
| freeze-dried | 1146d | 1448f | 1278e | 837b | 1177B | |
| mean | 1164B | 1497D | 1261C | 790A | ||
|
β
-carotene mg/100 g DM |
frozen | 180bc | 213e | 155a | 187bcd | 184A |
| air-dried | 188bcd | 221e | 185bcd | 171ab | 191AB | |
| freeze-dried | 178bc | 220e | 204de | 193cd | 199B | |
| mean | 182A | 218B | 181A | 184A | ||
* FM – fresh matter, DM - dry matter; different small letter for each object within individual component represents significant differences between means at P < 0.05; different capital letters in column or row represent significant differences between means for the type of product from a different location and different product from a specific location, respectively, at P < 0.05.
Table 2.
The content of vitamin C in frozen, dried, and freeze-dried wild garlic leaves [mg/100 g DM]*.
Table 2.
The content of vitamin C in frozen, dried, and freeze-dried wild garlic leaves [mg/100 g DM]*.
| Component | Product | Location | ||||
|---|---|---|---|---|---|---|
| Kraków | Limanowa | Ropa | Sucha Beskidzka | mean | ||
| L-ascorbic acid (AA) | frozen | 379cd | 473d | 441cd | 840f | 533C |
| air-dried | 15a | 50ab | 130b | 14a | 52A | |
| freeze-dried | 365c | 455cd | 445cd | 619e | 471B | |
| mean | 253A | 326B | 339B | 491C | ||
| L-dehydroascorbic acid (DHAA) | frozen | 462e | 497g | 542h | 833i | 584C |
| air-dried | 5a | 41b | 170c | 40b | 64A | |
| freeze-dried | 516g | 481f | 521g | 537h | 514B | |
| mean | 328A | 339B | 411C | 470D | ||
| Vitamin C | frozen | 841c | 969de | 983e | 1673g | 584C |
| air-dried | 20a | 91a | 300b | 54a | 64A | |
| freeze-dried | 881cd | 936de | 966de | 1156f | 514B | |
| mean | 328A | 339B | 411C | 470D | ||
* DM - dry matter; different small letter for each object within individual component represents significant differences between means at P < 0.05; different capital letters in column or row represent significant differences between means for the type of product from a different location and a different product from a specific location, respectively, at P < 0.05.
Table 3.
Total polyphenol content and antioxidant activity in frozen, air-dried, and freeze-dried wild garlic leaf*.
Table 3.
Total polyphenol content and antioxidant activity in frozen, air-dried, and freeze-dried wild garlic leaf*.
| Component | Product | Location | ||||
|---|---|---|---|---|---|---|
| Kraków | Limanowa | Ropa | Sucha Beskidzka |
mean | ||
|
Total polyphenols g/100 g DM |
frozen | 1.64cd | 1.95e | 1.91e | 1.68d | 1.80B |
| air-dried | 1.73d | 2.01ef | 2.14f | 1.47ab | 1.84C | |
| freeze-dried | 1.59bcd | 1.51abc | 1.41a | 1.45ab | 1.49A | |
| mean | 1.65B | 1.83C | 1.82C | 1.53A | ||
|
ABTS ABTS µmol Trolox/g DM |
frozen | 347a | 511b | 418ab | 397ab | 418A |
| air-dried | 1860e | 1955e | 1889e | 1919e | 1905C | |
| freeze-dried | 1636d | 1434c | 1523c | 1725d | 1580B | |
| mean | 1281A | 1300AB | 1276A | 1347B | ||
|
DPPH ABTS µmol Trolox/g DM |
frozen | 20ab | 12a | 11a | 27bc | 18A |
| air-dried | 124f | 99e | 110e | 131f | 116C | |
| freeze-dried | 36cd | 48d | 48d | 42d | 43B | |
| mean | 60AB | 53A | 56A | 67B | ||
|
FRAP µmol Fe2+/g DM |
frozen | 707a | 798b | 717a | 716a | 734A |
| air-dried | 1295e | 1609g | 1379f | 1050c | 1333C | |
| freeze-dried | 1283e | 1205d | 1046c | 1248de | 1195B | |
| mean | 1095C | 1204D | 1047B | 1004A | ||
* DM - dry matter; different small letter for each object within individual component represents significant differences between means at P < 0.05; different capital letters in column or row represent significant differences between means for the type of product from a different location and a different product from a specific location, respectively, at P < 0.05.
Table 4.
Marker analysis of coding regions based techniques CDDP and PBA.
| CDDP analyse | ||
| Primer combination | Results of differentiation | Type of profile change |
| F/R1 | - | Monomorphic profile |
| F/R2 | + | Locus insertion in sample R |
| F/R2b | - | Monomorphic profile |
| F/R3 | + | Locus deletion in sample R |
| F/R3b | + | Locus deletion in sample R Locus insertion in sample L Locus insertion in sample S |
| PBA analyse | ||
| Primer combination | Results of differentiation | Type of profile change |
| CYP A1 F+R | + | Locus deletion in sample R Locus deletion in sample L Locus deletion in sample S |
| CYP 2B F+R | + | Locus deletion in sample R Locus deletion in sample L Locus deletion in sample S Locus insertion in sample R Locus insertion in sample L Locus insertion in sample S |
| CYP 2C F+R | + | Locus deletion in sample R Locus deletion in sample L Locus deletion in sample S Locus insertion in sample R Locus insertion in sample L Locus insertion in sample S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



