
Submitted:
10 November 2023
Posted:
13 November 2023
You are already at the latest version
Abstract
CD147/Basigin, a transmembrane glycoprotein belonging to the immunoglobulin super family, is a multifunctional molecule with various binding partners. CD147 binds to monocarboxylate transporters (MCTs) and supports their expression on plasma membranes. MCTs play an important role in glycolysis, the enzymatic conversion of glucose to pyruvate and adenosine triphosphate (ATP). Pyruvate is further converted to lactic acid which is exported from the cytoplasm by MTC-1 and MCT-4 to maintain intracellular pH and a stable metabolic state. Under physiological conditions, cellular energy production is induced by mitochondrial oxidative phosphorylation. Glycolysis usually occurs under anaerobic conditions, whereas cancer cells depend on glycolysis under aerobic conditions. T cells also require glycolysis for differentiation, proliferation, and activation.
Human malignant melanoma cells expressed higher levels of MCT-1 and MCT-4, co-localized with CD147 on the plasma membrane and showed an increased glycolysis rate compared to normal human melanocytes. Silencing of CD147 by siRNA abrogated the membrane expression of MCT-1 and MCT-4 and disrupted glycolysis, inhibiting cancer cell activity.
The differentiation of CD4+ T cells into Th17 cells is a pivotal process in the development of various immune disorders, including psoriasis. Studies using CD147-deficient mice demonstrated the involvement of CD147 in psoriasis. MCT-1 was absent on CD4+ T cells in CD147-deficient mice. Naïve CD4+ T-cells from CD147-deficient mice show low potential for differentiation into Th17 cells. Imiquimod-induced skin inflammation was significantly milder in CD147-deficient mice than in wild-type mice. These findings indicate that CD147/Basigin is involved in the development of malignant tumors and T cell mediated immunological disorders via regulation of glycolysis.
Keywords:
CD147
; glycolysis
; malignant tumor
; psoriasis
; immune disorder
1. Introduction
CD147/Basigin, originally cloned as a carrier of the Lewis X carbohydrate antigen, is a transmembrane protein belonging to the immunoglobulin super family [1,2]. CD147 performs pleiotropic functions by binding to various molecules including monocarboxylate transporters (MCTs). MCTs, MCT-1 and MCT-4, play important roles in the regulation of glycolysis, the enzymatic conversion of glucose to pyruvate and adenosine triphosphate (ATP), to fuel the pathophysiological activities of cellular growth and proliferation [3]. CD147 is highly associated with MCT-1 and MCT-4 and facilitates their expression on the plasma membrane [4].
Malignant melanoma (MM) is highly malignant because of its rapid growth and metastatic properties. In MM, CD147 expression is enhanced on the cell membrane and CD147 plays an important role in tumor proliferation, production of matrix metalloproteinases (MMPs), angiogenesis, invasiveness, and metastatic activity in vitro and in vivo [5,6]. In human MM cells, CD147 co-localizes with MCT-1 and MCT-4 on the plasma membrane and promotes the proliferation, invasiveness, and metastasis of MM by regulating glycolysis. Silencing of CD147 by a small interfering RNA (siRNA) clearly abrogates the expression of MCT-1 and MCT-4 and their co-localization with CD147, dramatically decreases the glycolysis rate, extracellular pH, and the production of ATP. Thus, cell proliferation, invasiveness, and VEGF production are significantly decreased by siRNA. These previous observations strongly suggest that CD147 interacts with MCT-1 and MCT-4 to promote tumor cell glycolysis leading to the progression of MM [5,6,7].
T-cells also depend on glycolysis to generate energy for their differentiation, proliferation, and activation [8]. Psoriasis is a chronic inflammatory dermatosis characterized by recurrent episodes of sharply demarcated scaly erythematous plaques. Differentiation of naïve CD4+ T-cells into Th17 cells is important for the development of psoriasis [9,10]. We speculated that CD147 is involved in the pathogenesis of psoriasis through the regulation of glycolysis and demonstrated that CD147 is essential for the development of psoriasis via the induction of Th17 cell differentiation. The serum level of CD147 is increased in psoriasis patients and the expression of CD147 and MCT-1 is elevated on their dermal CD4+ RORγt+ Th17 cells. In vitro, the potential of naïve CD4+ T cells to differentiate into Th17 cells is abrogated in T-cells from CD147-deficient mice. Imiquimod (IMQ)-induced psoriasis-like dermatitis is significantly milder in CD147-deficient mice and bone marrow chimeric mice lacking CD147 in bone marrow hematopoietic cells. These findings demonstrate that CD147 is essential for the development of psoriasis via the induction of Th17 cell differentiation [11].
Here, we review the function of CD147 in glycolysis and its involvement in the development of malignant tumors and immune disorders.
2. CD147/Basigin
2.1. Discovery of CD147/Basigin
Cell surface glycoproteins regulate cellular activities, such as proliferation, differentiation, adhesion, migration, and transmembrane transportation [12,13]. CD147/Basigin was cloned as a carrier of Lewis X, a cell surface carbohydrate antigen expressed in embryonal carcinoma (EC) cells using the λgt11 expression library of F9 EC cells in our study of developmentally regulated cell surface markers [1]. CD147/Basigin is a transmembrane glycoprotein comprising two immunoglobulin (Ig)-like extracellular domains, a single transmembrane domain, and a short C-terminal cytoplasmic tail [1]. Extracellular Ig-like domains have strong homology with the Ig variable domain and the major histocompatibility complex class II β-chain. Based on the structural analysis, CD147/Basigin was identified as a new member of the Ig superfamily [1]. CD147/Basigin is expressed to a similar level in various adult organs, mouse embryos, and EC cells. Because of its broad distribution, we considered that this molecule has basic or fundamental functions and termed “Basigin (Bsg)” as an abbreviation of basic immunoglobulin [1]. It is located on chromosome 19 at p13.3 and consists of 10 exons spanning approximately 12 kb [14]. The molecular weight of this protein portion is 28 kDa. CD147 has three asparagine (ASN) glycosylation sites on its Ig-like domains and is highly glycosylated. The molecular weights of the glycosylated forms ranged from 43 to 66 kDa. Their diverse molecular weights are due to the different modes of glycosylation of molecules from various organs [2].
2.2. Discovery of related molecules
Identical molecules have been found independently from various perspectives in several laboratories and are given different names: M6, extracellular matrix metalloproteinase inducer (EMMPRIN), HT7, neurothelin, 5A11, gp42, OX-47, and CE9 [15,16,17,18,19,20,21,22]. Among these, M6 was cloned using peripheral granulocytes from patients with rheumatoid arthritis (RA) and was identified as a human leukocyte activation antigen [15]. EMMPRIN, previously known as tumor cell collagenase stimulatory factor (TCSF) [23], is implicated in the induction of matrix metalloproteinases (MMPs). Its expression is enhanced in various human carcinomas, including MM, and correlates with tumor progression and invasion by inducing the production of MMPs by stromal fibroblasts [24,25,26,27,28,29]. HT7 is a highly glycosylated protein localized in brain endothelial cells which is a receptor involved in cell surface recognition at the blood-brain barrier (BBB) [17]. Neurothelin is an inducible cell surface glycoprotein of BBB-specific endothelial cells and distinct neurons and regulates interaction between vascularization and neuronal differentiation [18]. Fadool and Linser reported that 5A11 is a functional molecule involved in neural-glial mutual recognition in the avian neural retina [19]. A murine fibroblast membrane glycoprotein, gp42, is a fibronectin-receptor-associated antigen involved in the neural cell adhesion [20]. OX-47 is a lymphocyte activation antigen. It presents on lymphocytes whose expression is markedly induced on activation with mitogens [21]. CE9 is a posterior-tail domain-specific integral plasma membrane glycoprotein cloned from a rat spermatozoon and plays an important role in spermatogenesis [22]. These studies suggest that CD147/Basigin is a multifunctional molecule involved in various physiological and pathological phenomena [30].
The gene and protein names given to these molecules are basigin both in humans (Locus Link) and mice (Mouse Genome Informatics) and the symbol provided by the Human Genome Organization is BSG in humans and Bsg in mice [31]. Because basigin is expressed on leukocytes, the Cluster of Differentiation Nomenclature “CD147” was given at the 5th International Workshop in 1993 [32].
3. Chaperone-like function of CD147/Basigin
Previous studies have revealed that CD147 has various binding partners such as cyclophilin A (CyPA) [33], integrins [34,35], P-glycoprotein [12,30,36], and MCTs [4]. The pleiotropic functions of CD147 are attributable to its binding partners. CD147 acts as a chaperone for their proper plasma membrane expression and catalytic activity and participates in many pathophysiological processes through these molecules [12,30].
CyPA, the major target of the immunosuppressive drug cyclosporin A, is a ubiquitously distributed intracellular protein. CyPA is secreted by cells in response to inflammatory stimuli and is a potent neutrophil and eosinophil chemoattractant. In the process of inflammation, CD147 acts as a cell surface receptor for CyPA and initiates signaling cascades leading to ERK activation [33]. CD147 binds to β1-integrin. β1-integrin induces aggregation of promonocyte line U937 cells via protein tyrosine phosphorylation. The antibody against CD147 inhibits the aggregation and the tyrosine phosphorylation by blocking the binding between β1-integrin and CD147 [34]. Multidrug resistance (MDR) of cancer cells is often associated with over expression of P-glycoprotein (P-gp), a transmembrane ATP-dependent transporter. Expression of P-gp and CD147 is up-regulated in Adriamycin (ADR)-resistant human mammary carcinoma cell line MCF7 (MCF7/Adr) compared to its non-ADR-resistant counterpart MCF7. Silencing of CD147 in MCF7/Adr by shRNA targeting CD147 resulted in the down-regulation of P-gp expression and the reduction of drug resistance [36].
The association between CD147 and MCTs yielded important clues for understanding the role of CD147. Cross-linking studies showed that CD147 forms a homo-oligomer [37]. Homodimerized CD147 binds to two MCT monomers and facilitates its proper folding and expression on the cell membrane (Figure 1). In MCT-transfected COS cells, expressed MCT proteins accumulate in the perinuclear compartment, whereas co-transfection with CD147 cDNA enables the expression of functional MCT-1 or MCT-4 on the plasma membrane [4]. As mentioned previously, CD147 consists of two Ig-like extracellular domains: a single transmembrane domain and a short C-terminal cytoplasmic tail [1]. MCT-1 directly binds to the transmembrane domain of CD147 [30,38]. The cryo-EM structure of the CD147/MCT complex has been determined [39,40] and Glu218 in the CD147 transmembrane domain is the binding site for MCT-1 [39,41]. Philp et al. demonstrated the role of CD147 in the retina, in association with MCTs [42]. CD147-deficient mice exhibit visual impairment. In CD147-deficient mice, morphology of the retina at the light microscopic level and the fundus and the fluorescein fundus angiography appeared to be normal until 8 weeks of age, whereas the amplitude of all components of both the photopic and scotopic electroretinograms was decreased indicating that both rod and cone functions were severely affected in CD147-deficient mice [43]. In these mice, cell membrane expression of MCT-1 and MCT-4 was greatly reduced in retinal photoreceptor cells and adjacent Müller cells. Müller cells are retinal glial cells, whose major role is to maintain the functional and structural stability of photoreceptor cells. Müller cells provide photoreceptor cells with lactate as fuel for normal functions [42]. Because of the absence of MCTs on the membrane of facing cells, the flux of lactate from Müller cells to photoreceptor cells is disrupted, and photoreceptor cell activity is lost owing to energy depletion [42]. The absence of CD147 results in the impaired expression of MCTs on the cell membrane leading to loss of vision.
Other binding partners of CD147 include glucose transporter-1 (GLUT1), CD44, the major hyaluronan receptor, CD43, CD98, γ-secretase, NOD2, γ-catenin, platelet glycoprotein VI (GPVI), and apolipoprotein D. Molecular interactions between CD147 and these binding partners have been previously documented [30].
4. CD147 and glycolysis
Elucidation of the mechanism of lactate flux by CD147 and MCTs in the retina prompted us to investigate the role of CD147 in glycolysis. Glycolysis is the enzymatic conversion of glucose to pyruvate to generate energy, which is stored in the form of ATP. Pyruvate is further converted to lactic acid, which is exported through the plasma membrane and is required for metabolism and intracellular pH regulation [38]. Lactic acid is transported by proton-linked/lactate co-transporters, MCTs, on the plasma membrane. The detailed metabolic pathway of glycolysis was initially studied in cancer cells. Under physiological conditions, cellular energy is provided by mitochondrial oxidative phosphorylation and glycolysis results from anaerobic enzymatic conversion. Warburg first reported that cancer cells depend on glycolysis for energy even in the presence of oxygen, i.e., under aerobic conditions [3]. They take up excess glucose through GLUT-1 or GLUT-3, which is then enzymatically converted into ATP to fuel the pathophysiological activities of cellular growth and proliferation [44]. During aerobic glycolysis in cancer cells, pyruvate is converted into lactic acid, which is excreted from the cytoplasm into the surrounding extracellular microenvironment through MCTs. Altered metabolism requires tumor cells to rapidly efflux lactate into the surrounding microenvironment to prevent self-poisoning. MCTs facilitate proton-linked monocarboxylate transport, leading to a decrease in the extracellular pH of tumors. The acidity of tumor microenvironment produces more aggressive phenotypes in cancer cells that exhibit increased proliferation, invasiveness, metastasis, and VEGF production [45,46,47,48].
Based on the observation in the retina, we investigated the involvement of CD147 in cancer cell glycolysis using MM cells. Human MM cells (A375) expressed higher levels of MCT-1, MCT-4, and CD147 and showed an increased glycolysis rate compared to normal human melanocytes. CD147 co-localized with MCT-1 and MCT-4 on the A375 cell membrane. CD147 silencing by siRNA abrogated MCT1 and MCT-4 expression and their co-localization with CD147. The glycolysis rate and ATP production were dramatically decreased and the extracellular pH increased. Subsequently, cell proliferation, invasiveness, and VEGF production were significantly inhibited [5]. Gallagher et al. documented similar findings in highly metastatic breast cancer cell line MDA-MB-231. In accordance with the findings in MM, lactate efflux was mediated by MCTs and the accessory subunit, CD147. CD147 was highly expressed on MDA-MB-231 cells and its expression was linked to MCT expression. MCT-4 mRNA and protein expression were increased in MDA-MB-231 cells compared to cells derived from normal mammary tissue. MCT-4 co-localized with CD147 in the plasma membrane. CD147 silencing resulted in the loss of MCT4 in the plasma membrane and the accumulation of the transporter in endo-lysosomes. On the other hand, silencing of MCT-4 impaired the maturation and trafficking of CD147 to the cell surface, resulting in accumulation of CD147 in the endoplasmic reticulum [49]. These findings strongly suggest that CD147 interacts with MCT-1 and MCT-4 to promote glycolysis in tumor cells, resulting in tumor progression (Figure 2).
5. T cell differentiation/proliferation and glycolysis
Glycolysis is also important for differentiation, proliferation, and activation of lymphocytes including T cells, B cells, and natural killer cells. Activated lymphocytes engage in robust growth and rapid proliferation. For these processes, lymphocytes adopt glycolysis [8]. Stimulated CD4+ T cells differentiate into effector T cells or inducible regulatory T cells. Differentiation of CD4+ T cells into distinct subsets, Th1, Th2, and Th17 cells, requires aerobic glycolysis. Th1, Th2, and Th17 cells express high surface levels of the GLUT-1 and are highly glycolytic [50]. Halestrap and Wilson demonstrated the importance of MCT-1 in T cell activation and proliferation. In T cells, energy metabolism is largely glycolytic even under aerobic conditions and lactic acid efflux from T cells is mediated by MCT-1. MCT-1 is important during the activation and proliferation of resting T cells, which is accompanied by a switch from aerobic to glycolytic metabolism and remarkable increase in lactate production and export [51]. In line with this, Murray et al. showed that a potent and specific MCT-1 inhibitor blocked T-cell proliferation and acted as an immunosuppressive agent [52]. T cell glycolysis is regulated by CD147 because MCT-1 requires CD147 as an ancillary protein in T cells [51]. Studies on the role of CD147 in T cell biology by Hahn et al. documented that CD147 is critical for energy metabolism and is involved in the development, activation, proliferation, migration, invasion, and adhesion of T-cells. The rapid proliferation and activation of T cells require glycolysis instead of oxidative phosphorylation to respond to their energy demands. Although glycolysis generates energy faster than oxidative phosphorylation, energy production is less efficient and leads to the intracellular accumulation of lactic acid. CD147 plays a role in alleviating lactate efflux for cell stability [53]. In patients with RA, CD147 mRNA expression was elevated in peripheral blood mononuclear cells. CD147 promotes the differentiation of CD4+ T cells into Th17 cells and induces production of Th17-secreting cytokine, interleukin (IL)-17, and Th17 differentiation-regulating cytokines, IL-6 and IL-1β [54]. These findings strongly suggest that CD147 is involved in the pathogenesis of T cell-mediated immune disorders.
6. Psoriasis
Psoriasis is a chronic inflammatory keratotic dermatosis characterized clinically by recurrent episodes of sharply demarcated scaly erythematous plaques and histologically by hyperkeratosis with parakeratosis, Munro microabscess, absence of granular layer, regular elongation of the rete ridges, marked dilatation of blood vessels in the papillary dermis, and dense clusters of inflammatory cells consisting of T cells and dendritic cells in the dermis. Our molecular and cellular understanding of the immunopathogenesis of psoriasis has progressed, and recent studies have revealed that Th17 cells and related signaling pathways play pivotal roles in the development of psoriasis [9,10,55]. First, tumor necrosis factor (TNF) and inducible nitric oxide synthase (iNOS)-producing dendritic cells (TIP-DC) which reside in the dermis are activated and produce TNF-α and IL-23. TNF-α is required for the maintenance of activatied state of TIP-DC and IL-23 promotes the differentiation of naïve CD4+ T cells into Th17 cells in human. Th17 cells produce the effector cytokines IL-17 and IL-22 which activate signal transducer and activator of transcription (STAT) 3 signaling in keratinocytes, leading to the production of chemokines, cytokines, and proinflammatory mediators such as IL-6, IL-8, GM-CSF, CXCL10, and CCL20 which are responsible for the proliferation of epidermal keratinocytes to develop psoriasis lesions [56]. Th17 cells are a distinct T cell subset derived from naïve CD4+ helper T cells and the mechanism underlying Th17 cell differentiation is known [57]. The combination of IL-6 and transforming growth factor-β (TGF-β) activates the retinoic acid receptor-related orphan nuclear receptor γt (RORγt) that is the key transcription factor directing the differentiation into Th17 cells [58]. Because the differentiation of naïve CD4+ helper T cells into Th17 cells is the fundamental process of psoriasis development [59,60] and requires glycolysis as an energy source, it is possible that CD147 participates in the pathogenesis of psoriasis.
CD147/Basigin and psoriasis
Previous studies showed that CD147 is highly expressed in peripheral blood neutrophils and neutrophils in the lesioned skin of patients with psoriasis [61]. The expression level was significantly correlated with disease severity as evaluated by the psoriasis area and severity index (PASI) [61]. In this study, inhibitory effect of CD147 on neutrophil chemotaxis was presented. An accepted model system for studying neutrophil chemotaxis using all-trans retinoic acid (ATRA)-induced differentiated HL-60 promyelocytic leukemia cells was employed. By treatment with ATRA, HL-60 cells were differentiated into neutrophils, which was confirmed by the expression of CD11b, cytoplasm to nucleus ratio, and nuclear segmentation accompanied by chromatin condensation. IL-8 and N-formyl-methyonylleucyl-phenylalanine (fMLP), well-known neutrophil chemo-attractants, induced the chemotaxis of ATRA-treated HL-60 cells. Silencing of CD147 significantly decreased the migration of these cells [61]. CD147 is cleaved and released into the peripheral blood as a 22-kDa fragment of the N-terminal extracellular domain [62]. Soluble CD147 has been detected in patients with malignant tumors such as MM and immune disorders including psoriasis and systemic lupus erythematosus (SLE) [63]. In patients with psoriasis, the serum level of soluble CD147 is significantly elevated compared to that in normal controls and correlates with PASI [64]. The effect of soluble CD147 on keratinocyte proliferation was examined. When HaCaT cells, an immortalized non-tumorigenic keratinocyte cell line, were treated with plasma from patients with psoriasis, proliferation was induced compared with that from normal subjects [64].
Regarding T cells in psoriasis, CD147 expression is increased in the CD3+ T cells in the dermis of psoriasis lesions. Its expression was significantly higher on CD4+ RORγt+ Th17 cells than on CD4+ RORγt- non-Th17 T cells. The expression level of MCT-1 on RORγt+ Th17 cells was upregulated and positively correlated with CD147 expression. These findings indicate that together with MCT-1, CD147 modulates CD4+ T-cell differentiation into Th17 cells [11].
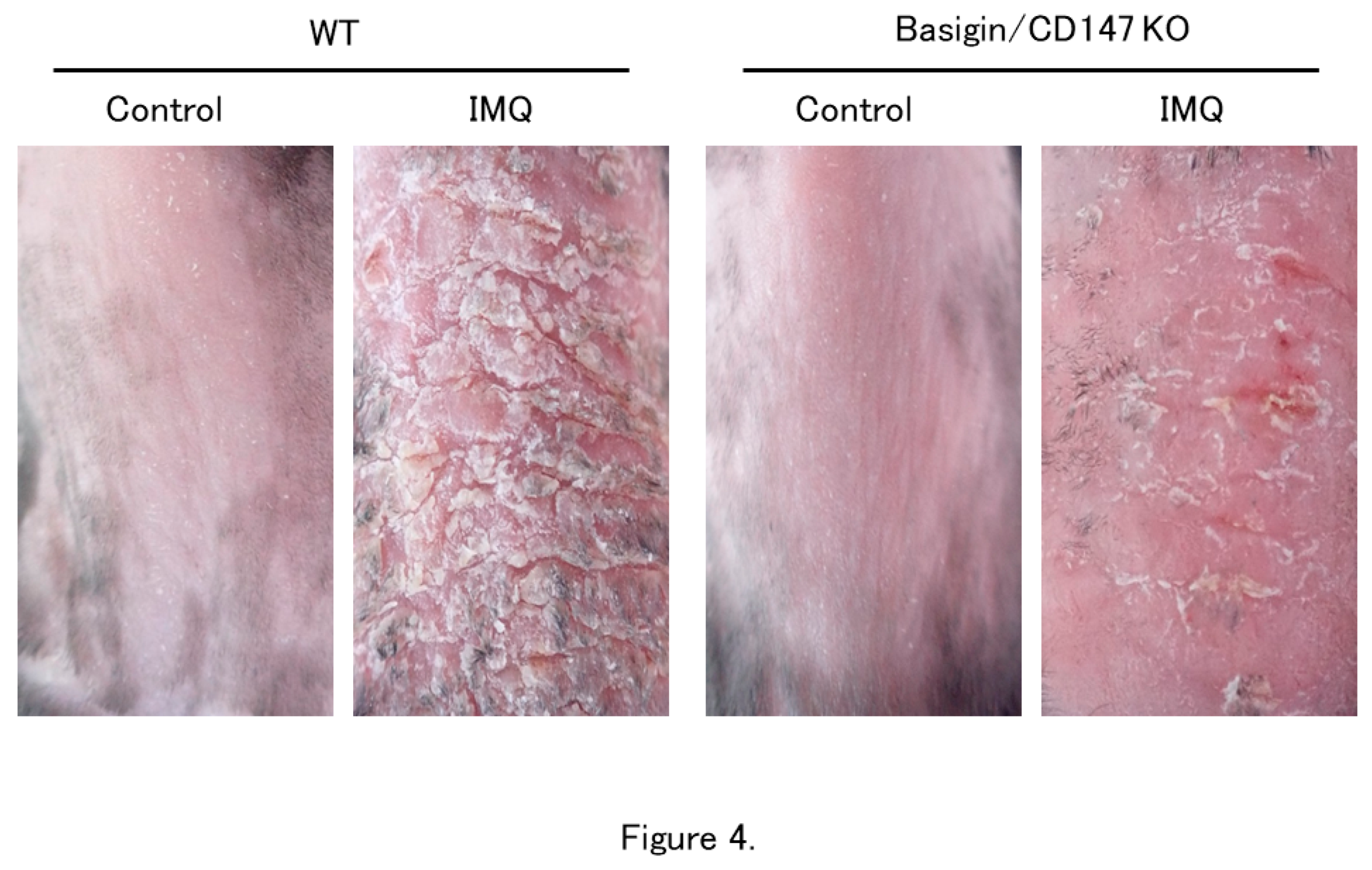
The involvement of CD147 in psoriasis development was investigated in wild-type (WT) and CD147-deficient mice. MCT-1 was absent on CD4+ T cells from CD147-deficient mice. Naïve CD4+ T-cells from spleen of CD147-deficient mice showed a low potential for differentiating into Th17 cells in response to stimulation with IL-6 and TGF-β, which are the differentiation stimuli into Th17 cells (Figure 3). Topical application of imiquimod (IMQ), a toll-like receptor-7 agonist, on the shaved skin induces psoriasis-like dermatitis. IMQ-induced dermatitis is an accepted psoriasis mouse model. After shaving the dorsal skin of CD147-deficient mice, 5% IMQ cream was applied for 7 consecutive days. The control WT mice were shaved but not treated with IMQ. The degree of skin inflammation was evaluated using the cumulative disease severity scoring system modified from the PASI score (modified PASI score). The degree of skin erythema, induration, and scaling was classified as 0 = none, 1 = mild, 2 = moderate, 3 = severe, and 4 = maximum. These scores were summed, resulting in a theoretical maximum score of 12. IMQ-induced skin inflammation was significantly milder in CD147-deficient mice than in WT mice (Figure 4). The modified PASI score in WT mice and CD147-deficient mice were 8.75 and 5.2 at day 6, and 8.0 and 4,6 at day 7, respectively. These studies on mice deficient in the CD147 gene yielded direct evidence that CD147 plays a major role in the development of psoriasis [11].
CD147 is ubiquitously expressed in various cells, including epidermal keratinocytes and lymphocytes. Epidermal CD147 plays a role in the pathogenesis of psoriasis via the production of IL-22 and subsequent STAT3 activation in the K5-promotor transgenic mice designed to overexpress CD147 in epidermal keratinocytes [65]. To examine the importance of CD147 in immune cells in psoriasis, bone marrow chimeric mice lacking CD147 in the bone marrow hematopoietic cells were produced. To produce bone marrow chimeric mice, Ly5.1 recipient mice were exposed to 10 Gy of total body irradiation. Bone marrow lumps were obtained from the femur of CD147-deficient and their wild type (WT) counterparts, and the lumps were transplanted to the recipient mice trans-venously. Leukocytes from the Ly5.1 recipients and the C57BL/6J donors expressed CD45.1 and CD45.2, respectively, which enables to distinguish the origin of the immune cells in the lesional skin. IMQ-induced skin lesions appeared significantly milder in the CD147-deficient chimeric mice than that in the WT chimeric mice. The modified PASI score in WT chimeric mice and CD147-deficient chimeric mice were 5.33 and 1.8 at day 6, and 5.67 and 1.4 at day 7, respectively. These results show that CD147 in immune cells is responsible for the development of psoriasis.
7. CD147/Basigin and other immune disorders
The involvement of CD147 in other immune disorders has been previously documented. As mentioned above, CD147 promotes Th17 cell differentiation in patients with RA. Expression of CD147 mRNA was approximately 3-fold higher in the peripheral blood mononuclear cells (PBMCs) from RA patients than those from normal subjects. Differentiation of CD4+ T cells into Th17 cells was promoted when CD4+ T cells were co-cultured with CD14+ monocytes in response to lipopolysaccharide (LPS) and anti-CD3 stimulation. The proportion of Th17 cells was significantly higher in RA patients than in normal subjects. Th17-secreting cytokine, IL-17, and Th17-regulating cytokines, IL-6 and IL-1β, were highly induced in the culture supernatants of the CD4+ /CD14+ co-culture. Anti-CD147 antibody reduced the proportion of Th17 cells and inhibited the production of IL-17, IL-6, and IL-1β [54].
Lung interstitial fibrosis is a chronic pulmonary disease characterized by excessive accumulation of extracellular matrix and often develops as a complication of various autoimmune diseases [66]. In a murine bleomycin-induced lung interstitial fibrosis model, CD147 neutralizing monoclonal antibodies (HAb18 mAbs) markedly reduced collagen accumulation and downregulated the proportion of M1 macrophages and Th17 cells. In vitro, M1 macrophages induced Th17 differentiation, which was significantly inhibited by treatment with HAb18 mAbs or by silencing CD147 by lentivirus interference in M1 macrophages. These observations indicate that CD147 promotes M1 macrophages and induces Th17 cell differentiation in lung interstitial fibrosis [67].
Multiple sclerosis (MS) is a demyelinating and neurodegenerative autoimmune disorder associated with the migration of activated lymphocytes and macrophages into central nervous system (CNS). These leukocytes enter the white matter by penetrating the BBB in areas of inflammation called perivascular cuffs and through the CNS-meningeal barrier. Accumulation of activated leukocytes in the lesion causes the loss BBB integrity leading to the transmigration of leukocytes into the brain parenchyma. Metabolic switch is seen in T cells and macrophages in the lesion; they heavily depend on glycolysis as energy source. In the experimental autoimmune encephalomyelitis (EAE) model of MS, macrophages within the perivascular cuffs of postcapillary venules were highly glycolytic as shown by strong expression of lactate dehydrogenase (LDHA) which converts pyruvate to lactate. These macrophages expressed prominent level of MCT-4, which engages the secretion of lactate from the cytoplasm of glycolytic cells. Silencing of LDHA or MCT-4 by each siRNA resulted in decreased lactate secretion and macrophage transmigration, indicating the functional relevance of glycolysis in the pathogenesis of EAE. In EAE, CD147 binds to MCT-4 and regulates its expression on the cell membrane of macrophages, which was confirmed by co-immunoprecipitation. Glycolysis and glycolytic capacity were significantly abrogated by CD147-knockdown by siRNA as measured by extracellular acidification rate (ECAR). These results demonstrate the importance of CD147, in association with MCT-4, in governing macrophage glycolysis and its migration in inflammatory perivascular cuffs of EAE. Relevance of MS was confirmed by the strong expression of CD147, MCT-4, and LDHA in perivascular macrophages in MS brains of human [68].
8. Conclusion and future directions
CD147 functions as a chaperone for various membrane proteins including CyPA, integrins, P-gp, MCTs, and so forth; and it supports their plasma membrane expression. In this review, we focused on MCTs. In association with MCT-1 and MCT-4, CD147 regulates glycolysis, an enzymatic metabolic system that generates energy that is stored as ATP. Glycolysis is required by cancer cells for their proliferation, invasiveness, and VEGF production and by T cells for their differentiation, proliferation, and activation. CD147 contributes to the development of malignant tumors and Th17 cell-mediated immune disorders including psoriasis. Therefore, CD147 is a promising therapeutic target for patients with these disorders.
References
- Miyauchi, T.; Kanekura, T.; Yamaoka, A.; Ozawa, M.; Miyazawa, S.; Muramatsu, T. Basigin, a new, broadly distributed member of the immunoglobulin superfamily, has strong homology with both the immunoglobulin V domain and the β-chain of major histocompatibility complex class II antigen. J. Biochem. 1990, 107, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Kanekura, T.; Miyauchi, T.; Tashiro, M.; Muramatsu, T. Basigin, a new member of the immunoglobulin super family: Genes in different mammalian species, glycosylation changes in the molecule from adult organs and possible variation in the N-terminal sequences. Cell Struct. Funct. 1991, 16, 23–30. [Google Scholar] [CrossRef]
- House, S.W.; Warburg, O.; Burk, D.; Schade, A.L. On respiratory impairment in cancer cells. Science 1956, 124, 269–270. [Google Scholar] [CrossRef]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef]
- Kanekura, T.; Chen, X. CD147/basigin promotes progression of malignant melanoma and other cancers. J. Dermatol. Sci. 2010, 57, 149–154. [Google Scholar] [CrossRef]
- Chen, X.; Lin, J.; Kanekura, T.; Su, J.; Lin, W.; Xie, H.; Wu, Y.; Li, J.; Chen, M.; Chang, J. A small interfering CD147-targeting RNA inhibited the proliferation, invasiveness, and metastatic activity of malignant melanoma. Cancer Res. 2006, 66, 11323–11330. [Google Scholar] [CrossRef]
- Su, J.; Chen, X.; Kanekura, T. A CD147-targeting siRNA inhibits the proliferation, invasiveness, and VEGF production of human malignant melanoma cells by down-regulating glycolysis. Cancer Lett. 2009, 273, 140–147. [Google Scholar] [CrossRef]
- Donnelly, R.P.; Finlay, D.K. Glucose, glycolysis and lymphocyte responses. Mol. Immunol. 2015, 68, 513–519. [Google Scholar] [CrossRef]
- Perera, G.K.; Di Meglio, P.D.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol. 2012, 7, 385–422. [Google Scholar] [CrossRef]
- Lynde, C.W.; Poulin, Y.; Vender, R.; Bourcier, M.; Khalil, S. Interleukin 17A: Toward a new understanding of psoriasis pathogenesis. J. Am. Acad. Dermatol. 2014, 71, 141–150. [Google Scholar] [CrossRef]
- Okubo, A.; Uchida, Y.; Higashi, Y.; Sato, T.; Ogawa, Y.; Ryuge, A.; Kadomatsu, K.; Kanekura, T. CD147 is essential for the development of psoriasis via the induction of Th17 cell differentiation. Int. J. Mol. Sci. 2021, 23, 177–191. [Google Scholar] [CrossRef]
- Muramatsu, T.; Miyauchi, T. Basigin (CD147): A multifunctional transmembrane protein involved in reproduction, neural function, inflammation and tumor invasion. Histol. Histopathol. 2003, 18, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Iacono, K.T.; Brown, A.L.; Greene, M.I.; Saouaf, S.J. CD147 immunoglobulin superfamily receptor function and role in pathology. Exp. Mol. Pathol. 2007, 83, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Kaname, T.; Miyauchi, T.; Kuwano, A.; Matsuda, Y.; Muramatsu, T.; Kajii, T. Mapping basigin (BSG), a member of the immunoglobulin superfamily, to 19p13.3. Cytogenet. Cell Genet. 1993, 64, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Kasinrerk, W.; Fiebiger, E.; Stefanová, I.; Baumruker, T.; Knapp, W.; Stockinger, H. Human leukocyte activation antigen M6, a member of the Ig superfamily, is the species homologue of rat OX-47, mouse basigin, and chicken HT7 molecule. J. Immunol. 1992, 149, 847–854. [Google Scholar] [CrossRef]
- Biswas, C.; Zhang, Y.; DeCastro, R.; Guo, H.; Nakamura, T.; Kataoka, H.; Nabeshima, K. The human tumor cell-derived collagenase stimulatory factor (renamed EMMPRIN) is a member of the immunoglobulin superfamily. Cancer Res. 1995, 55, 434–439. [Google Scholar]
- Seulberger, H.; Lottspeich, F.; Risau, W. The inducible blood-brain barrier specific molecule HT7 is a novel immunoglobulin-like cell surface glycoprotein. EMBO J. 1990, 9, 2151–2158. [Google Scholar] [CrossRef]
- Schlosshauer, B.; Herzog, K.H. Neurothelin: An inducible cell surface glycoprotein of blood-brain barrier-specific endothelial cells and distinct neurons. J. Cell. Biol. 1990, 110, 1261–1274. [Google Scholar] [CrossRef]
- Fadool, J.M.; Linser, P.J. 5A11 antigen is a cell recognition molecule which is involved in neuronal-glial interactions in avian neural retina. Dev. Dyn. 1993, 196, 252–262. [Google Scholar] [CrossRef]
- Altruda, F.; Cervella, P.; Gaeta, M.L.; Daniele, A.; Giancotti, F.; Tarone, G.; Stefanuto, G.; Silengo, L. Cloning of cDNA for a novel mouse membrane glycoprotein (gp42): Shared identity to histocompatibility antigens, immunoglobulins and neural-cell adhesion molecules. Gene 1989, 85, 445–451. [Google Scholar] [CrossRef]
- Fossum, S.; Mallett, S.; Barclay, A.N. The MRC OX-47 antigen is a member of the immunoglobulin superfamily with an unusual transmembrane sequence. Eur. J. Immunol. 1991, 21, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Nehme, C.L.; Cesario, M.M.; Myles, D.G.; Koppel, D.E.; Bartles, J.R. Breaching the diffusion barrier that compartmentalizes the transmembrane glycoprotein CE9 to the posterior-tail plasma membrane domain of the rat spermatozoon. J. Cell. Biol. 1993, 120, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; Troccoli, N.; Biswas, C. Coordinate increase in collagenase mRNA and enzyme levels in human fibroblasts treated with the tumor cell factor, TCSF. Biochem. Int. 1989, 19, 257–266. [Google Scholar] [PubMed]
- Bordador, L.C.; Li, X.; Toole, B.; Chen, B.; Regezi, J.; Zardi, L.; Hu, Y.; Ramos, D.M. Expression of EMMPRIN by oral squamous cell carcinoma. Int. J. Cancer. 2000, 85, 347–352. [Google Scholar] [CrossRef]
- Muraoka, K.; Nabeshima, K.; Murayama, T.; Biswas, C.; Koono, M. Enhanced expression of a tumor-cell-derived collagenase-stimulatory factor in urothelial carcinoma: Its usefulness as a tumor marker for bladder cancers. Int. J. Cancer. 1993, 55, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Polette, M.; Gilles, C.; Marchand, V.; Lorenzato, M.; Toole, B.; Tournier, J.M.; Zucker, S.; Birembaut, P. Tumor collagenase stimulatory factor (TCSF) expression and localization in human lung and breast cancers. J. Histochem. Cytochem. 1997, 45, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Sameshima, T.; Nabeshima, K.; Toole, B.P.; Yokogami, K.; Okada, Y.; Goya, T.; Koono, M.; Wakisaka, S. Glioma cell extracellular matrix metalloproteinase inducer (EMMPRIN) (CD147) stimulates production of membrane-type matrix metalloproteinases and activated gelatinase A in cocultures with brain-derived fibroblasts. Cancer Lett. 2000, 157, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Kanekura, T.; Chen, X.; Kanzaki, T. Basigin (CD147) is expressed on melanoma cells and induces tumor cell invasion by stimulating production of matrix metalloproteinases by fibroblasts. Int. J. Cancer 2002, 99, 520–528. [Google Scholar] [CrossRef]
- Kataoka, H.; DeCastro, R.; Zucker, S.; Biswas, C. Tumor cell-derived collagenase-stimulatory factor increases expression of interstitial collagenase, stromelysin, and 72-kDa gelatinase. Cancer Res. 1993, 53, 3154–3158. [Google Scholar]
- Muramatsu, T. Basigin (CD147), a multifunctional transmembrane glycoprotein with various binding partners. J. Biochem. 2016, 159, 481–490. [Google Scholar] [CrossRef]
- Simon-Chazottes, D.; Matsubara, S.; Miyauchi, T.; Muramatsu, T.; Guénet, J.L. Chromosomal localization of two cell surface-associated molecules of potential importance in development: Midkine (Mdk) and basigin (Bsg). Mamm. Genome. 1992, 2, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.N.; Marison, M.H.; Law, S.K.A.; McKnight, A.J.; Tomlinson, M.G.; van der Merwe, P.A. edt. The leucocyte antigen facts book. London: Academic Press Harcourt Brace & Company Publishers, 1997, p. 408–409.
- Yurchenko, V.; Zybarth, G.; O’Connor, M.; Dai, W.W.; Franchin, G.; Hao, T.; Guo, H.; Huang, H.-C.; Toole, B.; Gallay, P.; Sherry, B.; Bukrinsky, M. Active site residues of cyclophilin A are crucial for its signaling activity via CD147. J. Biol. Chem. 2002, 277, 22959–22965. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Fox, D.A.; Horejsi, V.; Sagawa, K.; Skubitz, K.M.; Katz, D.R.; Chain, B. The functional interaction s between CD98, β1-integrins, and CD147 in the induction of U937 homotypic aggregation. Blood 2001, 98, 374–832. [Google Scholar] [CrossRef]
- Nishibaba, R.; Higashi, Y.; Su, J.; Furukawa, T.; Kawai, K.; Kanekura, T. CD147-targeting siRNA inhibits cell-matrix adhesion of human malignant melanoma cells by phosphorylating focal adhesion kinase. J. Dermatol. 2012, 39, 63–67. [Google Scholar] [CrossRef]
- Li, Q.Q.; Wang, W.J.; Xu, J.D.; Cao, X.X.; Chen, Q.; Yang, J.M.; Xu, Z.D. Involvement of CD147 in regulation of multidrug resistance to P-gp substrate drugs and in vitro invasion in breast cancer cells. Cancer Sci. 2007, 98, 1064–1069. [Google Scholar] [CrossRef]
- Yoshida, S.; Shibata, M.; Yamamoto, S.; Hagihara, M.; Asai, N.; Takahashi, M.; Mizutani, S.; Muramatsu, T.; Kadomatsu, K. Homoligomer formation by basigin, an immunoglobulin superfamily member, via its N-terminal immunoglobulin domain. Eur. J. Biochem. 2000, 267, 4372–4380. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Meredith, D. The SLC16 gene family – From monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflugers Arch. 2004, 447, 619–628. [Google Scholar] [CrossRef]
- Köpnick, A.L.; Jansen, A.; Geistlinger, K.; Epalle, N.H.; Beitz, E. Basigin drives intracellular accumulation of L-lactate by harvesting protons and substrate anions. PLOS ONE. 2021, 16, e0249110. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Jiang, X.; Zhang, S.; Zhu, A.; Yuan, Y.; Xu, H.; Lei, J.; Yan, C. Structural basis of human monocarboxylate transporter 1 inhibition by anti-cancer drug candidates. Cell 2021, 184, 370–383.e13. [Google Scholar] [CrossRef]
- Manoharan, C.; Wilson, M.C.; Sessions, R.B.; Halestrap, A.P. The role of charged residues in the transmembrane helices of monocarboxylate transporter 1 and its ancillary protein basigin in determining plasma membrane expression and catalytic activity. Mol. Membr. Biol. 2006, 23, 486–498. [Google Scholar] [CrossRef]
- Philp, N.J.; Ochrietor, J.D.; Rudoy, C.; Muramatsu, T.; Linser, P.J. Loss of MCT1, MCT3, and MCT4 expression in the retinal pigment epithelium and neural retina of the 5A11/basigin-null mouse. Invest. Ophthalmol. Vis. Sci. 2003, 44, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Katayama, N.; Kachi, S.; Kondo, M.; Kadomatsu, K.; Usukura, J.; Muramatsu, T.; Mori, S.; Miyane, Y. Retinal dysfunction in basigin deficiency. Invest. Ophthalmol. Vis. Sci 2000, 41, 3128–3133. [Google Scholar]
- Weinhouse, S. Metabolism of respiratory fuels in cancer cells. Acta. Unio. Int. Contra. Cancrum. 1960, 16, 32–35. [Google Scholar] [PubMed]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Beckner, M.E.; Stracke, M.L.; Liotta, L.A.; Schiffmann, E. Glycolysis as primary energy source in tumor cell chemotaxis. J. Natl. Cancer Inst. 1990, 82, 1836–1840. [Google Scholar] [CrossRef] [PubMed]
- Rofstad, E.K.; Mathiesen, B.; Kindem, K.; Galappathi, K. Acidic extracellular pH promotes experimental metastasis of human melanoma cells in athymic nude mice. Cancer Res. 2006, 66, 6699–707. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, D.; Xu, L.; Chen, Y.; Gohongi, T.; Seed, B.; Jain, R.K. Hypoxia and acidosis independently up-regulate vascular endothelial growth factor transcription in brain tumors in vivo. Cancer Res. 2001, 61, 6020–6024. [Google Scholar] [PubMed]
- Gallagher, S.M.; Castorino, J.J.; Wang, D.; Philp, N.J. Monocarboxylate transporter 4 regulates maturation and trafficking of CD147 to the plasma membrane in the metastatic breast cancer cell line MDA-MB-231. Cancer Res. 2007, 67, 4182–4189. [Google Scholar] [CrossRef]
- Michalek, R.D.; Gerriets, V.A.; Jacobs, S.R.; Macintyre, A.N.; MacIver, N.J.; Mason, E.F.; Sullivan, S.A.; Nichols, A.G.; Rathmell, J.C. Cutting edge: Distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J. Immunol. 2011, 186, 3299–3303. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Wilson, M.C. The monocarboxylate transporter family - Role and regulation. IUBMB Life. 2012, 64, 109–119. [Google Scholar] [CrossRef]
- Murray, C.M.; Hutchinson, R.; Bantick, J.R.; Belfield, G.P.; Benjamin, A.D.; Brazma, D.; Bundick, R.V.; Cook, I.D.; Craggs, R.I.; Edwards, S.; Evans, L.R.; Harrison, R.; Holness, E.; Jackson, A.P.; Jackson, C.G.; Kingston, L.P.; Perry, M.W.D.; Ross, A.R.J.; Rugman, P.A.; Sidhu, S.S.; Sullivan, M.; Taylor-Fishwick, D.A.; Walker, P.C.; Whitehead, Y.M.; Wilkinson, D.J.; Wright, A.; Donald, D.K. Monocarboxylate transporter MCT1 is a target for immunosuppression. Nat. Chem. Biol. 2005, 1, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.N.; Kaushik, D.K.; Yong, V.W. The role of EMMPRIN in T cell biology and immunological diseases. J. Leuko.c Biol. 2015, 98, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, J.; Li, Y.; Yin, Z.J.; Lv, T.T.; Zhu, P.; Zhang, Y. CD147 modulates the differentiation of T-helper 17 cells in patients with rheumatoid arthritis. APMIS. 2017, 125, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Brembilla, N.C.; Boehncke, W.H. Revisiting the interleukin 17 family of cytokines in psoriasis: Pathogenesis and potential targets for innovative therapies. Front. Immunol. 2023, 14, 1186455. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.A.; Towne, J.E.; Kricorian, G.; Klekotka, P.; Gudjonsson, J.E.; Krueger, J.G.; Russell, C.B. The emerging role of IL-17 in the pathogenesis of psoriasis: Preclinical and clinical findings. J. Invest. Dermatol. 2013, 133, 17–26. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Cua, D.J. Th17 cell differentiation: The long and winding road. Immunity 2008, 28, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, B.; Ceribelli, A.; Massarotti, M.S.; Selmi, C. The Th17 axis in psoriatic disease: Pathogenetic and therapeutic implications. Auto. Immun. Highlights 2014, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Li, D.; Xiong, H.; Shi, D. Transcriptional regulation of effector T cells in the pathogenesis of psoriasis. Eur. J. Med. Res. 2023, 28, 182–189. [Google Scholar] [CrossRef]
- Lu, H.; Kuang, Y.H.; Su, J.; Chang, J.; Wu, L.S.; Kanekura, T.; Li, D.; Chen, M.-L.; Chen, X. CD147 is highly expressed on peripheral blood neutrophils from patients with psoriasis and induces neutrophil chemotaxis. J. Dermatol. 2010, 37, 1053–1056. [Google Scholar] [CrossRef]
- Egawa, N.; Koshikawa, N.; Tomari, T.; Nabeshima, K.; Isobe, T.; Seiki, M. Membrane type 1 matrix metalloproteinase (MT1-MMP/MMP-14) cleaves and releases a 22-kDa extracellular matrix metalloproteinase inducer (EMMPRIN) fragment from tumor cells. J. Biol. Chem. 2006, 281, 37576–37585. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Hori, M.; Kosugi, T.; Kojima, H.; Sato, W.; Inaba, S.; Maeda, K.; Nagaya, H.; Sato, Y.; Ishimoto, T.; Ozali, T.; Tsuboi, N.; Muro, Y.; Yuzawa, Y.; Imai, E.; Johnson, R.J.; Matsuo, S.; Kadomatsu, K.; Maruyama, S. Plasma CD147 reflects histological features in patients with lupus nephritis. Lupus. 2014, 23, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Liao, L.; Jia, X.; Kuang, Y.; Chen, X. Role of soluble CD147 in psoriatic patients: A preliminary study. J. Dermatol. 2018, 45, e266–267. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Zhang, S.; Lei, L.; Zhang, X.; Jia, X.; Luo, Z.; Huang, X.; Kuang, Y.; Zeng, W.; Su, J.; Chen, N. Epidermal CD147 expression plays a key role in IL-22-induced psoriatic dermatitis. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Wick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.F.; Feichtinger, J.; Zelger, B.; Sgonc, R.; Wolfram, D. The immunology of fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.J.; Zhang, K.; Chen, L.N.; Miao, J.L.; Yao, M.; Ren, Y.; Fu, Z.; Chen, Z.; Zhu, P. Enhancement of CD147 on M1 macrophages induces differentiation of Th17 cells in the lung interstitial fibrosis. Biochim. Biophys. Acta. 2014, 1842, 1770–1782. [Google Scholar] [CrossRef]
- Kaushik, D.K.; Bhattacharya, A.; Mirzaei, R.; Rawji, K.S.; Ahn, Y.; Rho, J.M.; Yong, V.W. Enhanced glycolytic metabolism supports transmigration of brain-infiltrating macrophages in multiple sclerosis. J. Clin. Invest. 2019, 129, 3277–3292. [Google Scholar] [CrossRef]
Figure 1.
CD147 associates with MCT-1 and MCT-4 and facilitates their expression in the plasma membrane. In the absence of CD147, MCTs accumulate in the cytosol and are not expressed on the plasma membrane (Modified from Reference 7).
Figure 1.
CD147 associates with MCT-1 and MCT-4 and facilitates their expression in the plasma membrane. In the absence of CD147, MCTs accumulate in the cytosol and are not expressed on the plasma membrane (Modified from Reference 7).
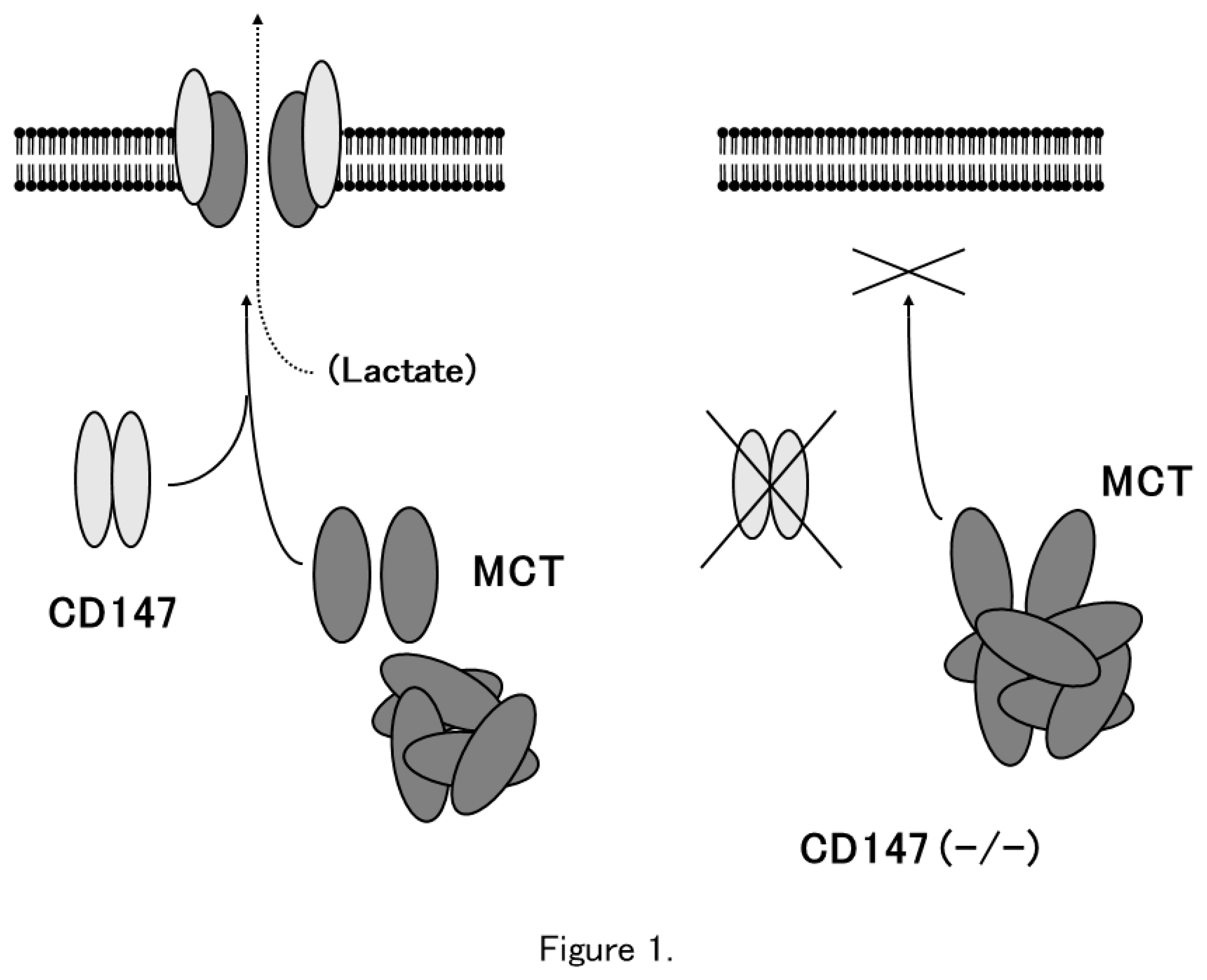
Figure 2.
Involvement of CD147 in tumor cell glycolysis. Glycolysis is the enzymatic conversion of glucose to pyruvate to produce ATP. Cancer cells depend on glycolysis for energy aerobic condition. Cancer cells take up excess glucose through GLUT-1 or GLUT-3 which is enzymatically converted to ATP to fuel the pathophysiological activities of cellular growth and proliferation. In aerobic glycolysis in cancer cells, pyruvate is further converted to lactic acid which is excreted from the cytoplasm into the surrounding extracellular microenvironment through MCT-1 or MCT-4. Homodimerized CD147 associates with two monomers of MCT1 or 4 and regulates lactate transport (Modified from Reference 7).
Figure 2.
Involvement of CD147 in tumor cell glycolysis. Glycolysis is the enzymatic conversion of glucose to pyruvate to produce ATP. Cancer cells depend on glycolysis for energy aerobic condition. Cancer cells take up excess glucose through GLUT-1 or GLUT-3 which is enzymatically converted to ATP to fuel the pathophysiological activities of cellular growth and proliferation. In aerobic glycolysis in cancer cells, pyruvate is further converted to lactic acid which is excreted from the cytoplasm into the surrounding extracellular microenvironment through MCT-1 or MCT-4. Homodimerized CD147 associates with two monomers of MCT1 or 4 and regulates lactate transport (Modified from Reference 7).
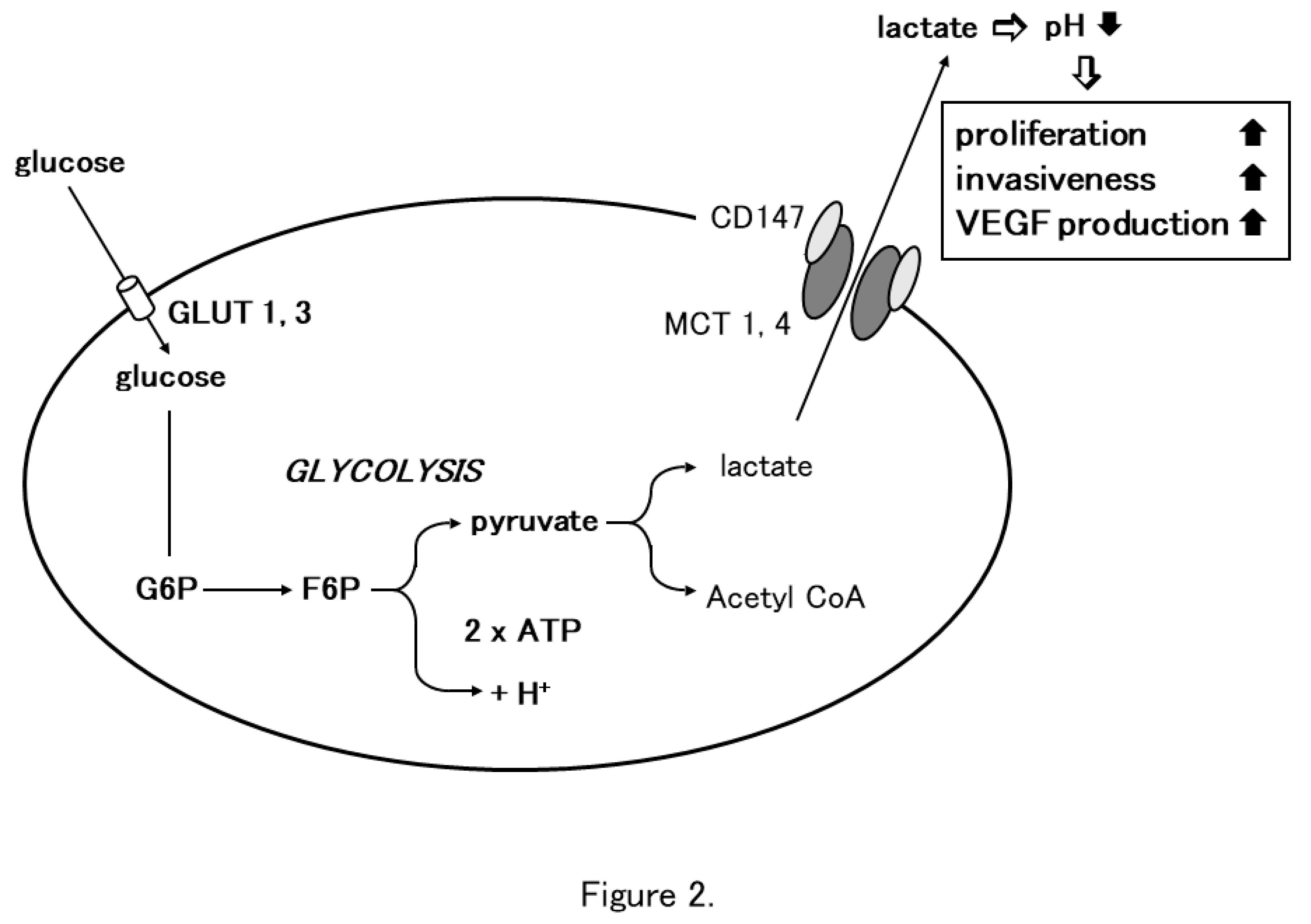
Figure 3.
In vitro effect of CD147 deficiency on the expression of MCT-1 and RORγt in CD4+ T cells. Naïve CD4+ T cells isolated from the spleens of CD147-/- mice and their wild-type (WT) counterparts were cultured on chamber slides and stimulated with IL-6 and TGF-β. RORγt expression is seen in WT CD4+ T cells, while it is significantly lower in the CD147-/- CD4+ T cells. MCT-1 is expressed on the plasma membrane of WT CD4+ but not on that of CD147-/- CD4+ T cells (Modified from Reference 11).
Figure 3.
In vitro effect of CD147 deficiency on the expression of MCT-1 and RORγt in CD4+ T cells. Naïve CD4+ T cells isolated from the spleens of CD147-/- mice and their wild-type (WT) counterparts were cultured on chamber slides and stimulated with IL-6 and TGF-β. RORγt expression is seen in WT CD4+ T cells, while it is significantly lower in the CD147-/- CD4+ T cells. MCT-1 is expressed on the plasma membrane of WT CD4+ but not on that of CD147-/- CD4+ T cells (Modified from Reference 11).
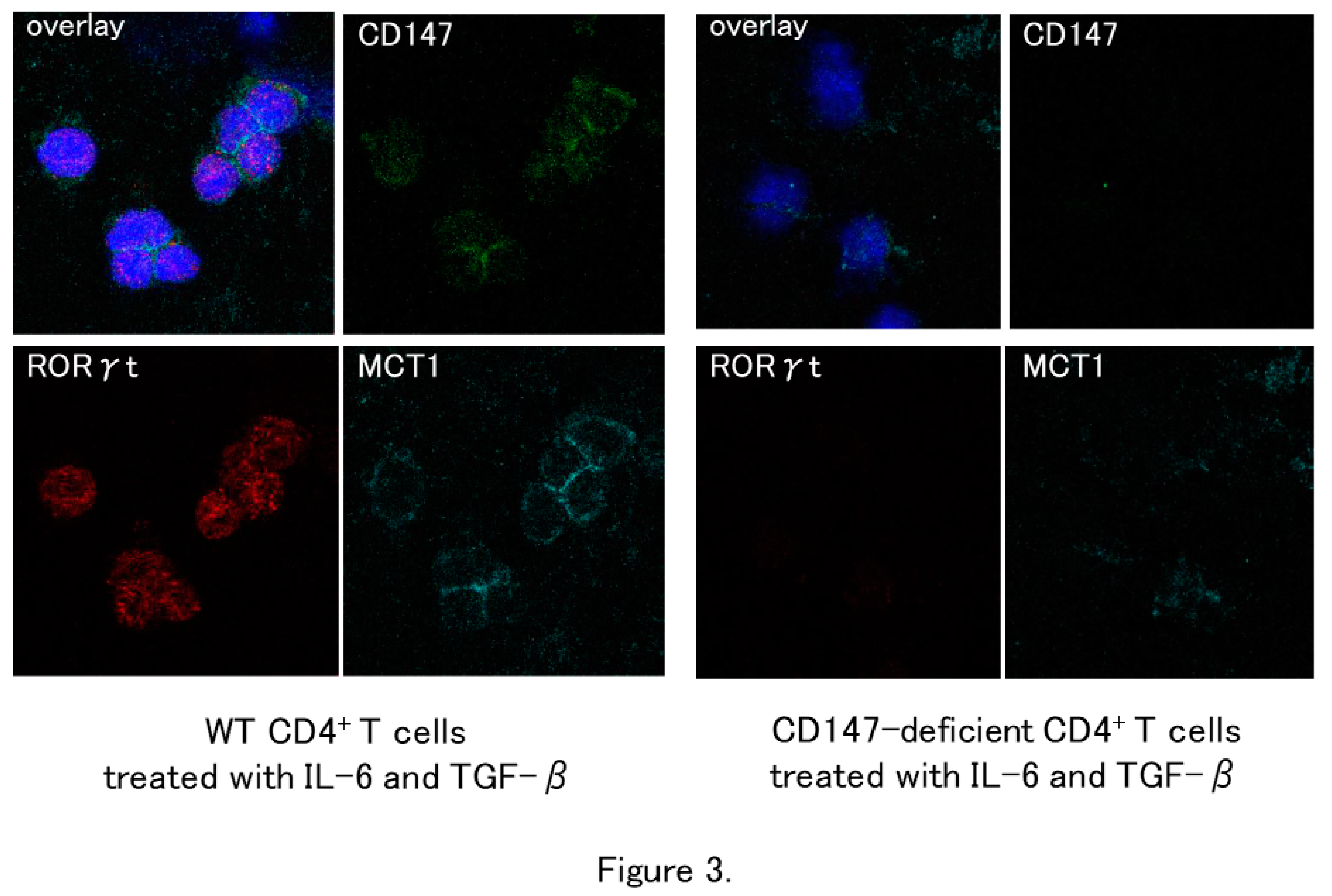
Figure 4.
Effect of CD147 deficiency on the development of IMQ-induced dermatitis. Imiquimod (IMQ) was applied on the dorsal skin of WT and CD147-deficient mice for seven consecutive days. Erythema, induration, and scaling are milder in CD147-deficient mice than in WT mice at day 7 (Modified from Reference 11).
Figure 4.
Effect of CD147 deficiency on the development of IMQ-induced dermatitis. Imiquimod (IMQ) was applied on the dorsal skin of WT and CD147-deficient mice for seven consecutive days. Erythema, induration, and scaling are milder in CD147-deficient mice than in WT mice at day 7 (Modified from Reference 11).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



