
Submitted:
10 November 2023
Posted:
13 November 2023
You are already at the latest version
Abstract
Plant pathogenic fungi are the most important cause of losses in agriculture. In the face of the overuse of synthetic fungicides, natural products are an encouraging alternative to control these plant pathogenic fungi. In this study, we tested the antifungal capacity of metabolites produced by strains of the genus Streptomyces against the plant pathogenic fungi Alternaria alternata, Botrytis cinerea and Fusarium oxysporum. In addition, we investigated the feasibility of applying these metabolites in a postharvest setting for the control of B. cinerea on cherry tomato fruits. The results revealed the ability of the strains, especially Streptomyces netropsis, to inhibit the growth of plant pathogenic fungi and, in some cases, the sporulation of these fungi. In addition, we observed an interesting phenomenon in which Streptomyces strains physically encapsulated the mycelium of the fungi, which contributed to their inhibition. Finally, the protective activity of S. netropsis extract on Cherry tomato fruits infected with B. cinerea was demonstrated. In conclusion, this research opens the door to studies on the understanding of physical microbial interactions in natural environments and holds promise in agriculture, especially in the protection of agricultural products during the post-harvest stage as a safer and more sustainable alternative.
Keywords:
Actinomycetes
; Streptomyces
; Alternaria alternata
; Botrytis cinerea
; Fusarium oxysporum
; Antifungal activity
; postharvest
1. Introduction
Humanity’s most significant challenge today is securing an adequate food supply for an ever-growing world population. This overpopulation, as well as the industrialization of the food sector, has driven an unprecedented intensification of crop cultivation, which puts significant pressure on natural resources and available arable land [1]. A direct consequence is soil quality has been observed. This degradation, coupled with the deterioration together with the loss of natural resources, is a problem that threatens agriculture’s ability to keep pace with the growing demand for food [2].
In addition, plant diseases cause significant losses in crop production worldwide, with their consequent economic impact [3]. Among the causal agents of these diseases, plant pathogenic fungi are the main cause of agricultural losses, responsible for infections affecting a wide range of cultivated plants. Among them, Alternaria alternata, Botrytis cinerea, and Fusarium oxysporum stand out as widely distributed pathogens that can cause considerable damage to crops of economic interest, causing root, fruit, and flower rot, vascular wilt and, ultimately, plant death [4].
Synthetic chemical fungicides are the most widely used to combat plant pathogenic fungi. However, the excessive use of these synthetic fungicides has caused a significant impact on human health due to the presence of residues in food and the environment. In this context, natural products or extracts are an encouraging alternative to synthetic chemical fungicides currently used to control plant pathogenic fungi [5]. Much of the losses due to these plant pathogenic fungi occur during the post-harvest stage, a critical stage in the supply chain of agricultural products [6], where quality and safety of products are key factors. Therefore, the direct application of bacteria poses significant challenges, as they could cause plant infections or raise food safety concerns for consumers [7]. Thus, the ideal is to adopt approaches that allow the antimicrobial compounds produced by bacteria to be harnessed without the need to apply the bacterial cells themselves to the cultures.
In this context, searching for effective and sustainable methods to control these diseases has prompted research in microbiology and biotechnology. One of the promising strategies in this field is the use of beneficial microorganisms such as actinomycetes. These Gram-positive and filamentous bacteria have the genus Streptomyces as their primary proponent due to their known ability to produce a wide variety of bioactive compounds with potential pharmaceutical, biotechnological, and agricultural applications, including inhibition of plant pathogenic fungi [8,9]. There impressive diversity of secondary metabolites of interest, including antibiotics, anti-tumor, and immunosuppressive agents, which have been the subject of study and use for decades [10]. Due to their relevance in the search for new Streptomyces strains capable of producing new therapeutic agents and bioproducts for various uses is ongoing. Whole genome sequences have shown that actinomycetes, including Streptomyces spp., have larger potential for biosynthesis of new compounds than that shown in axenic culture. Genomics has shown that a single species has more than 20 gene clusters potentially encoding new compounds. For example, more than 20 clusters for the production of secondary metabolites are detected in the genome of Streptomyces coelicolor (8.66 Mb) [11], and 25 clusters are included in the genome of Streptomyces avermitilis (9.02 Mb) [12]. However, a significant proportion of these genes remain unexpressed under axenic culture. In this task for finding new active compounds, apart from more classical studies analyzing compounds produced by Streptomyces when growing in axenic culture, the co-culture of microorganisms has emerged as a valuable approach. Co-culture better simulates ecological habitat interactions and extends the exploration of the impact of microbial interactions on the biosynthesis of new secondary metabolite in Streptomyces. Some representative examples from other studies are the co-culturing of Streptomyces with Myxococcus xanthus, which triggered the production of secondary metabolites of interest [13,14].
The present study is focused on evaluating the extracts in vitro antifungal capacity produced by four strains from different species of the genus Streptomyces against the plant pathogenic fungi A. alternata, B. cinerea, and F. oxysporum. In addition, the viability of these extracts was studied in a post-harvest setting for the fungal control of B. cinerea on cherry tomato fruits.
2. Materials and Methods
2.1. Strains, culture growth and co-cultures
The plant pathogenic reference fungi used in this trial were A. alternata strain Aa100, B. cinerea strain B05.10, and F. oxysporum strain 2715 belong to strain collection in University La Laguna. The actinomycetes strains used in the present study were isolated in a previous study from soils of Tenerife (Canary Islands, Spain) carried out in 2016, following the methodology described previously [9], classified according to their sequences in their closest species and deposited in the Nertalab S.L. strain collection. These strains were: Streptomyces ‘ardesiacus’ P3, Streptomyces ‘mauvecolor’ P46M, both from pine forest environment; Streptomyces ‘netropsis’ A52M, coastal environment, and Streptomyces ‘pratensis’ i3E, high mountain environment. Actinomycetes and fungi were cultured on TSA and PDA agar plates. Bacteria were grown for five days at 23°C, forming a homogeneous lawn using a sterile swab. Similarly, fungi strains were grown on PDA agar plates under the same conditions.
Actinomycetes and fungi co-culture was carried out in 250 ml Erlenmeyer flasks containing 50 ml TSB. Two discs of 8 mm diameter of the chosen actinomycete strain and one disc of the same size of one of the phytopathogenic fungi were added. Controls were inoculated with bacterial or fungi strains alone. Each treatment was carried out in triplicate, resulting in three replicates of the extracts for each actinomycete with each fungus. Erlenmeyer flasks were incubated at the same temperature under agitation, and 2 ml aliquots were taken at 24 hours and then every 48 hours for seven days. They were examined under the light microscope by making a fresh smear, without staining, to observe actinomycete and fungal growth in all treatments. Subsequently, the samples were subjected to two successive centrifugations of 10 minutes at 13,000 rpm to remove the microorganisms and to obtain a supernatant used for the fungal in vitro inhibition assay.
2.2. Antifungal activity of the media
The antifungal activity of the co-culture media was tested in PDA agar plates. All plates had the same volume (20 ml) and thickness to ensure uniform product diffusion across all plates. Each plate was surface inoculated with 50 µl of a spore suspension at a concentration of 2 · 104 spores/µl of B. cinerea and F. oxysporum. The spore suspension was prepared from plates of the different fungi, grown in the dark at 23°C for 5 to 7 days. Approximately 5 ml of sterile water was poured onto each plate, and the surface was scraped. The resulting suspension was recovered and filtered through sterile cotton wool into a falcon tube. Subsequently, an aliquot of the suspension was taken and quantified in a hemocytometer, allowing normalization of the spore concentration [15]. In the case of A. alternata, as it was impossible to obtain a spore suspension, a plate of 5-7 days of growth was taken and sown on grass using a sterile swab.
The agar diffusion method was used to test the antifungal capacity against the indicated fungi [16]. Briefly, 8 mm diameter circumferences were removed with a sterile punch, creating wells in the agar, three per plate. Then, 100 μl of each supernatant from each actinomycete with each fungal was added to each well, using TSB as a negative control. This was done in triplicate with each replicate of the co-cultures. The plates were incubated at 26°C in the dark, and inhibition halos were measured after five days. Compared to the paper disc diffusion method [17], this method allows more product volume to be added for evaluation and more uniform diffusion through the agar. The measurements taken were separated according to the type of inhibition observed. Thus, total growth inhibition, partial or affected inhibition, and inhibition of spore production were evaluated.
2.3. Medium solvent extraction and concentration
The co-culture medium was harvested and centrifuged in 50 ml Falcon tubes at 4800 rpm for 15 minutes to remove cells, hyphae, and debris. The centrifuged media was mixed with petroleum ether, hexane, diethyl ether, dichloromethane, ethyl acetate, and isobutyl alcohol in the ratio of (1:1) (v/v) and shaken vigorously for 10 min in a solvent extraction funnel. The solvent phase was separated from the aqueous phase, and this process was repeated three times. The solvent phase was evaporated to dryness in a water bath at 80-90°C and resuspended in sterile water (30-35 ml). The antifungal activity of the extract was analyzed following the agar diffusion method again in triplicate [16].
2.4. Extract antifungal activity on post-harvest cherry tomatoes
The extract with the highest antifungal activity in vitro was used for fruit post-harvest treatment against B. cinerea infection. Cherry tomato fruits (Solanum lycopersicum var. cerasiforme) were washed for 15 minutes in water with Tween 80 and bleach at 0.05% and 0.15%, respectively [18]. Finally, tomatoes were rinsed thrice (10 minutes each) in distilled water.
Skin fruits were wounded with a sterile 0.9x25 mm hypodermic needle. Tomatoes were inoculated with 5 µl of a B. cinerea spore suspension at 2·104 spores/µl. After draining the spore suspension, the infected fruits were sprayed with the chosen extract. Two treatments were carried out in parallel, one with an application of the product at the time of inoculation (A1) and the other with the same application and a booster after 24 hours (A2). Controls were sprayed with water, and the same protocol was carried out: one control with a single water application (C1) and a second application after 24 hours (C2).
Ten fruits were infected for each treatment and incubated for seven days in darkness in closed trays with moistened paper, maintaining a high relative humidity. They were observed every day, and after a week, the disease incidence was measured as the percentage of infected wounds [4]. In addition, it was also assessed whether the infection was only superficial or internal and the turgidity to the touch of the fruit using tweezers, discriminating between the two results. This trial was carried out 3 times.
2.5. Statistical analysis
Statistical analyses were performed with SPSS Statistics software (IBM, United States). Homoscedasticity was assessed by Levene’s test, while normality was corroborated by the Shapiro-Wilk test, since the sample size was less than 50. Thus, the different treatments were compared with the control using the Mann-Whitney U-test.
3. Results
3.1. Actinomycetes co-culture growth inhibition and antifungal activity in vitro
Fungal growth inhibition was observed from the beginning of the co-culture (24 hours), except for F. oxysporum. Actinomycetes growth was prominent, while fungi hyphae and spores were covered by actinomycete mycelium with structures like cysts (Figure 1). Moreover, in the case of the S. netropsis strain, the fungal mycelium and spores covered by the bacterial mycelium acquired a blue coloration, even though no dye was used (Figure 1a).
Screening of media aliquots taken every 24 hours from each co-culture allowed the selection of the optimal culture time to obtain the highest activity in vitro antifungal tests. All the Actinomycetes species exhibited similar antifungal kinetics in the antifungal activity tests. None of the strains showed activity before 48 hours of culture. In the case of S. mauvecolor in co-culture with A. alternaria, B. cinerea and F. oxysporum (Figure 2) the highest activity was shown after 48h, decreasing progressively after the fifth day. At seven days, the antifungal activity of the aliquots decreased except against Fusarium oxysporum. Furthermore, the S. mauvecolor co-culture extracts against A. alternaria showed complete growth inhibition (Figure 3a), while the same extracts tested with B. cinerea and F. oxysporum only exhibited partial inhibition (Figure 3b,c). In extracts from cocultures of the other actinomycete species, the decrease in activity on day 7 was drastic.
3.2. Actinomycetes species antifungal activity in vitro
The antifungal activity was analyzed for the rest of the Actinomycetes species, using media collected from five days of co-culture. The media with lower activity was found in S. pratensis co-culture. Although this actinomycete was able to encapsulate the mycelium of A. alternata inhibiting this fungus in co-culture, in vitro its extract did not inhibit its growth nor F. oxysporum. However, it successfully significantly inhibited the growth of B. cinerea as well as its ability to sporulates resulting on the plates in two distinctive concentric halos, an inner halo without fungal growth and an outer halo where sporulation is inhibited (Figure 3a). On its part, S. ardesiacus, significantly inhibited the mycelial growth of A. alternata and the sporulation of both B. cinerea and F. oxysporum (Figure 3b–d). Nevertheless, the best antifungal activity was observed for media extracts of S. netropsis, which significantly inhibited the sporulation of F. oxysporum and the growth of A. alternata and B. cinerea, showing large halos (Figure 3e–g). Finally, S. mauvecolor significantly inhibited the growth of A. alternata, although with smaller halo sizes (Figure 3h), while its effect on B. cinerea were like those of S. ardesiacus, showing a potent inhibition of fungal sporulation but not of mycelial growth (Figure 3i), making its use against this fungus less attractive than S. netropsis. In addition, S. netropsis was the only species that inhibited the sporulation and mycelial growth of F. oxysporum (Figure 3j).
3.3. Organic solvent extraction and antifungal activity
The use of less polar solvents (petroleum ether, hexane, diethyl ether, dichloromethane) for extraction of actinomycetal compounds exhibited no antifungal activity in vitro. However, the water phase retained similar activity ratios on fungi than above shown. Consequently, increasing the solvent polarity (ethyl acetate) was necessary to detect the antifungal activity. However, with ethyl acetate, the highest antifungal activity stayed in the aqueous phase, with a slight reduction in mycelia growth (Figure 4a,b). The highest antifungal activity was obtained using isobutyl alcohol, the most polar solvent immiscible with water. The solvent phase showed almost the same antifungal activity as the crude supernatant media, and hardly any growth inhibition halos were created with the aqueous phase extracted with isobutyl alcohol tests (Figure 4a,c).
3.4. Botrytis growth inhibition on cherry tomatoes
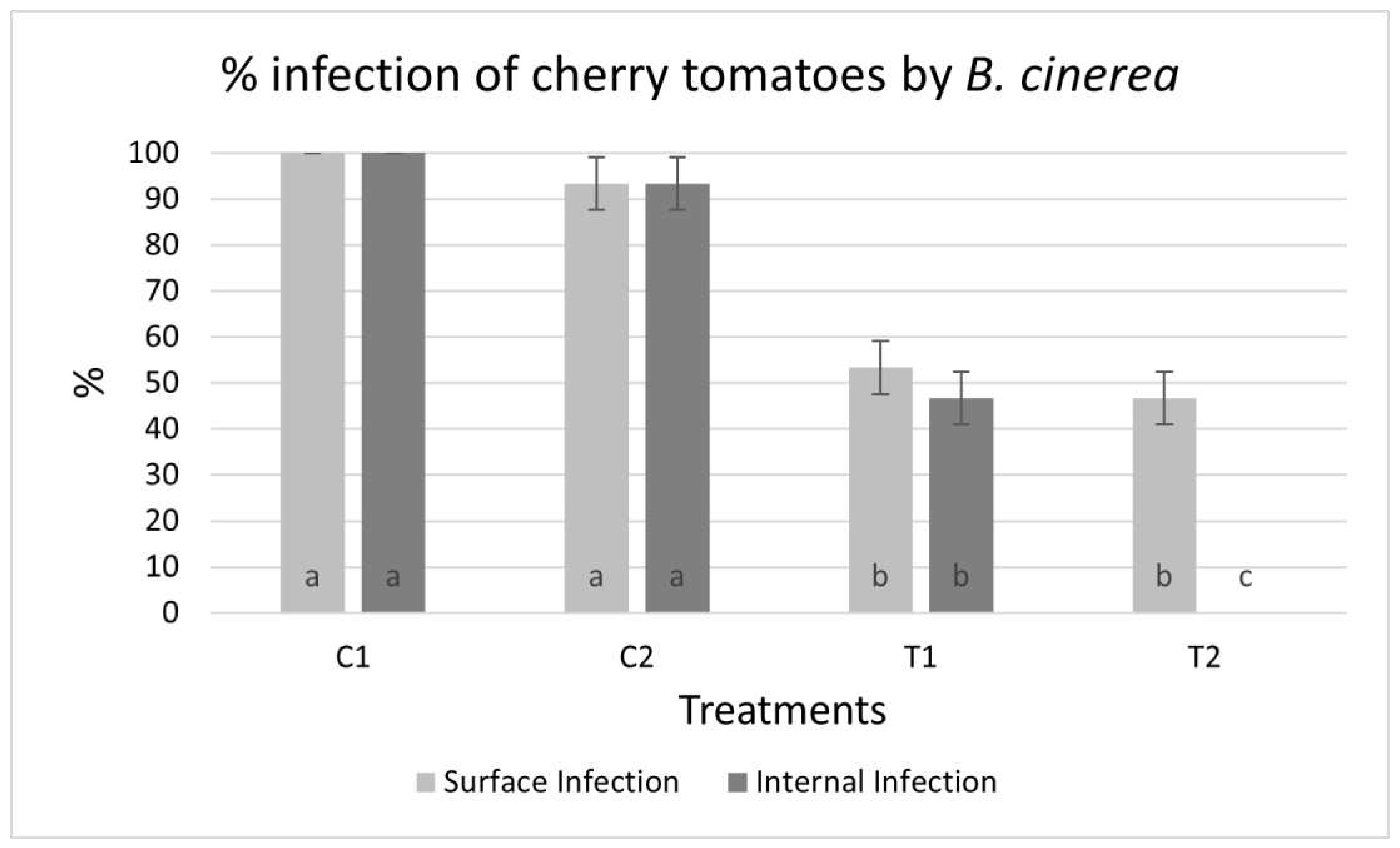
The isobutyl alcohol extracts were dissolved in water and tested on cherry tomatoes infected with B. cinerea. Seven days later, 100% of the control fruits of a water application (C1) and 90% of those subjected to reinforcement (C2) showed visible infection with aerial mycelium of the fungus. The area around the infection injury was sunken, and an internal rot was highly developed, resulting in the absence of turgor in all infected fruit (Figure 5a,b). However, fruits sprayed with S. netropsis isobutyl alcohol extract showed much lower infection rates. In those subjected to a single application (A1), only 50% of the fruits showed infection by B. cinerea, with low internal damage causing loss of turgor (Figure 5c). Finally, the cherry tomatoes subjected to the reinforcement treatment with the extract (A2) also showed a surface 50% infection. However, none of the fruits showed internal rotting, 100% of them retaining their turgor (Figure 5d). For both treatments, the fruits did not show infection but did show scarring tissue, which acquired a dark color. Thus, the treatments resulted in a significant reduction in the percentage of infected fruit (Figure 6).
4. Discussion
Previous works described the antifungal activity of many Actinomycetes strains against numerous plant pathogenic fungi [19]. In this line, in the present study we also show the good activity in vitro of compounds present in culture extracts of different Streptomyces spp. to inhibit the growth and sporulation of several plant pathogenic fungi. Further work is required to identify these compounds and to investigate the effect of inoculation of these Streptomyces strains in field cultures. In addition, we show the existence of a physical control mechanism consisting of the “trapping” of the fungus inside the mycelium of the actinomycete. These physical mechanisms, which can be easily observed by a color change on a simple smear of a co-culture sample, open new research areas, which could have implications for understanding how these microorganisms interact in natural environments as well as how these interactions could be used to design new strategies for agricultural or biotechnological applications. Exploring the nature of these “cysts” and its relationship to antifungal activity could also lead to a better understanding of the microbial defense mechanisms in the interaction between these filamentous bacteria and fungi when coexisting in a particular habitat. Although the production of bioactive secondary metabolites by the genus Streptomyces is well known [20,21], no similar data have been reported on the encapsulation of fungi by Streptomyces mycelium, nor on the compounds related to the color change or staining of fungal mycelium when growing in co-cultures, making these data highly novel. Related results reported are the induction by several Streptomyces species of the formation of mounds of M. xanthus [22]. The coloring could be due to compounds produced by the actinomycete or by the fungus itself. For example, there are studies in which co-culture induces the production of metabolites by the actinomycete, as in the case of S. coelicolor when co-cultured with M. xanthus, which produces actinorhodin, curiously blue colored [13]. In contrast, in other studies, such as with S. peucetius, co-culture has induced the synthesis of DK-Xanthene by M. xanthus, which is yellow in color [22]. Therefore, the coloring of the fungal mycelium could be due to this fungal stimulation, as secondary metabolites of fungi with blue coloring, such as azulenes or atrovenetin produced by Penicillium herquei or Lactarius sp. have also been reported [23]. The literature seems to indicate that one of the crucial aspects of this interaction is the competition for iron, which induces the production of siderophores triggering the activation of biosynthetic pathways related to the synthesis of secondary metabolites by the actinomycete [14], while small molecules of less than 3 kDa have been identified in the stimulation of M. xanthus [22]. In our case, further studies are needed to determine the nature and origin of this staining.
Regarding the kinetics of production of active compounds, we revealed the antifungal activity of Streptomyces spp. was different depending on the phytopathogenic species, the best antifungal activity probably being for S. netropsis, negatively affecting F. oxysporum, A. alternata and B. cinerea. Moreover, we found that 3-5 days of co-culture are optimal to reach the highest activity before collecting the media, which generally decreased later at 7 days. These results may seem surprising because secondary metabolism usually happens late when the culture has passed its exponential growth phase [24]. However, several of these metabolites may arise as part of their survival strategy in their natural habitat and produced earlier, as actinomycete show limited competitiveness in growth, with faster-growing bacteria due to their slow growth [25]. Elucidating the production kinetics of these antifungal compounds would require a more exhaustive study of the extracts, with precise identification of the compounds and a detailed analysis of the metabolism involved in their generation.
In terms of the importance of these plant pathogenic fungi, A. Alternaria and B. cinerea are responsible for causing significant losses in producing and marketing fresh fruits, vegetables, and ornamentals worldwide. Therefore, postharvest treatment is a crucial aspect of the supply chain of agricultural products, as it directly influences the quality, shelf life, and food safety of fruits [5,26]. Postharvest fungus control using synthetic fungicides allows the control of losses due to plant rot. However, their adverse effects on environmental and human health make it necessary to search for safer and more environmentally friendly alternatives [27]. Numerous research fronts are currently open in this field, and they constantly rely on bioactive compounds produced by bacteria for fungal biocontrol [28,29,30]. Thus, the excellent inhibition results of S. netropsis and its lack of pathogenicity in humans and plants make this bacterium an excellent and promising alternative in the fight against fungal biocontrol.
F. oxysporum is not a problem in post-harvest, but in the field, where it causes Panama disease on banana trees. It’s infection takes place via the root, by penetration through the epidermis into the xylem vessels and establishment throughout the vascular system [31], while that of A. alternata and B. cinerea occurs directly in leaves, young shoots, and fruits [32,33]. This result is exciting in a region like the Canary Islands since more than 9,000 hectares are used in banana crops. At the beginning of the century, this crop accounted for 25% of the gross value of agricultural production, representing 6% of the employment of the active population in the Canary Islands [34]. This percentage has decreased over these two decades. The highest production is found on the islands of Tenerife (43%) and La Palma (35%). However, it is on the second island where the relative importance of the crop concerning other economic sectors is more prominent. This is due to the large surface area this crop covers and the fact that in past decades, there has been no other crop with such a high demand to cover production costs [35]. This is why S. netropsis extract is the most interesting for postharvest application. Thus, once the optimal culture time had been established and the activity ratio for this strain had been verified, the supernatant was extracted with organic solvents to eliminate the remains of the culture medium and other substances that were not of interest.
As for the organic solvent extraction of actinomycetes co-culture media showed the polar nature of the molecule(s) responsible for the antifungal activity. Therefore, isobutyl alcohol was used to extract the active molecules from the co-culture medium, leaving behind other unwanted substances; the extract was considered suitable for evaluating its effectiveness in postharvest treatments.
Finally, with respect to the B. cinerea growth inhibition on cherry tomatoes assay, the protective activity of S. netropsis extract was demonstrated. As previously mentioned, this pathogen can infect fruits and leaves, generating considerable losses in agricultural yield before and after harvest [26]. The fact that tomato is one of the most consumed vegetables in the world, with more than 5 million hectares cultivated [36], makes controlling biotic stress a priority nowadays. Mainly, cherry tomato presents a compact size, making it an ideal candidate to be used as a model in laboratory trials, as stated by other authors such as Fernandez-San Millan et al. [4]. The results obtained with S. netropsis increase the interest in this species for future research in the fight against phytopathogens, both pre and postharvest. The most suitable dosage, mode of application, molecule/s identification, and safety for human consumption of treated foods still need to be established with detailed studies.
5. Conclusions
In conclusion, the positive outcomes of antifungal activities obtained by using crude extracts of Streptomyces co-cultures with pathogenic fungi could have a significant impact on improving agricultural production. This can be achieved by controlling fungal infections during crops or post-harvest by inoculating with these Streptomyces strains or spraying their purified active products. This strategy would reduce the dependence on chemical fungicides, promoting more environmentally friendly practices and food safety. Specifically, the extract of a single strain of S. netropsis greatly inhibited the sporulation of F. oxysporum and the growth of A. alternata and B. cinerea. Furthermore, it showed promising results for postharvest application on tomato fruits against B. cinerea, as it halved the number of infected fruits. Other Streptomyces strains showed more specific effects on the pathogenic fungi, with extract of S. mauvecolor only able to completely inhibit F. oxysporum and extracts of S. pratensis and S. ardesiacus, although with less-than-optimal results, also able to inhibit at least totally or partially one of the three phytopathogenic fungi. Remarkable was the observation of the formation of actinomycetal “cysts” surrounding and trapping the fungal mycelium, an easily distinguishable physical encapsulation by the specific acquired blue coloration of the fungal mycelium, and that resulted in the inhibition of fungal growth, F. oxysporum. These novel results are of interest for new lines of research, which could significantly impact the understanding of interactions between microorganisms in natural environments. Finally, the extracts of these new strains of Streptomyces could be a new source of environmentally friendly antifungal control.
Author Contributions
Conceptualization, D.T.M.F., E.H.B., M.L.B and J.C.L.J.; methodology, D.T.M.F., E.H.B., E.H.A. and S.D.; software, D.T.M.F. and J.C.L.J.; validation, D.T.M.F., E.H.B. and J.C.L.J.; formal analysis, D.T.M.F. and E.H.B.; investigation, D.T.M.F., E.H.B., E.H.A. and S.D.G.; resources, S.D.G., M.L.B., N.A.C. and J.C.L.J.; data curation, D.T.M.F. and E.H.B.; writing—original draft preparation, D.T.M.F.; writing—review and editing, D.T.M.F, M.L.B and J.C.L.J.; visualization, D.T.M.F. and J.C.L.J.; supervision, J.C.L.J.; project administration, M.L.B., N.A.C. and J.C.L.J.; funding acquisition, J.C.L.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the aid program of the Consejería de Economía, Industria, Comercio y Conocimiento for training research staff in the Canary Islands, co-financed by the Fondo Social Europeo (grant numbers: TESIS2020010011, TESIS2017010078, TESIS2021010033). We also declare funding from Nertalab S.L.
Data Availability Statement
The data used to support the findings of this study are included within the article.
Acknowledgments
We thank Ana Portero Álvarez for her collaboration in the laboratory and Concepción González Martín for her recommendations and for allowing us to use her equipment when necessary. We would also like to thank Dr. Cristina Giménez Mariño for providing the strains of the plant pathogenic fungi.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bahadur, I. The novel potassic bio-fertilizers: a promising approach for evergreen agriculture. Int. J. Microbiol. Res. 2015, ISSN, 0975-5276.
- Bahadur, I.; Maurya, R.; Roy, P.; Kumar, A. Potassium-solubilizing bacteria (KSB): a microbial tool for K-solubility, cycling, and availability to plants. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability: From Theory to Practices; Kumar, A., Meena, V., Eds.; Springer: Singapore; pp. 257–265. [CrossRef]
- Bandara, A.Y.; Weerasooriya, D.K.; Bradley, C.A.; Allen, T.W.; Esker, P.D. Dissecting the economic impact of soybean diseases in the United States over two decades. PLOS one. 2020, 15, e0231141. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-San Millan, A.; Larraya, L.; Farran, I.; Ancin, M.; Veramendi, J. Successful biocontrol of major postharvest and soil-borne plant pathogenic fungi by antagonistic yeasts. Biol. Control. 2021, 160, 104683. [Google Scholar] [CrossRef]
- Parra-Amin, J.E.; Cuca, L.E.; González-Coloma, A. Antifungal and phytotoxic activity of benzoic acid derivatives from inflorescences of Piper cumanense. Nat. Prod. Res. 2021, 35(16), 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Mederos-Torres, Y.; Bernabé-Galloway, P.; Ramírez-Arrebato, M.A. Películas basadas en polisacáridos como recubrimientos biodegradables y su empleo en la postcosecha de los frutos. Cultivos Tropicales. 2020, 41(3). [Google Scholar]
- Truchado, P.; Allende, A. La implicación de las frutas y hortalizas en las toxiinfecciones alimentarias y la relevancia del estado fisiológico de las bacterias. arbor. 2020, 196(795), a541–a541. [Google Scholar] [CrossRef]
- Salwan, R.; Sharma, V. Bioactive compounds of Streptomyces: biosynthesis to applications. Stud. Nat. Prod. Chem. 2020, 64, 467–491. [Google Scholar] [CrossRef]
- Hernández-Bolaños, E.; Montesdeoca-Flores, D.T; Abreu-Yanes, E.; Barrios, M.; Abreu-Acosta, N. Evaluating different methodologies for bioprospecting actinomycetes in Canary Islands soils. Curr. Microbiol. 2020, 77, 2510–2522. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, M.H.; Rasmey, A.H.M.; El-Sayed, E.S.A.; El-Kady, I.A.; Yassin, I.M. Biosynthesis of anti-inflammatory immunosuppressive metabolite by Streptomyces variabilis ASU319. Eur. J. Biol. Res. 2016, 6(3). [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; ... Hopwood, D.A. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3 (2). Nature 2002, 417, 141–147. [CrossRef]
- Ōmura, S.; Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Takahashi, C.; Shinose, M.; ... Hattori, M. Genome sequence of an industrial microorganism Streptomyces avermitilis: deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 12215–12220. [CrossRef] [PubMed]
- Pérez, J.; Muñoz-Dorado, J.; Braña, A.F.; Shimkets, L.J.; Sevillano, L.; Santamaría, R.I. Myxococcus xanthus induces actinorhodin overproduction and aerial mycelium formation by Streptomyces coelicolor. Microb. Biotechnol. 2011, 4(2), 175–183. [Google Scholar] [CrossRef]
- Lee, N.; Kim, W.; Chung, J.; Lee, Y.; Cho, S.; Jang, K.S.; ... Cho, B.K. Iron competition triggers antibiotic biosynthesis in Streptomyces coelicolor during coculture with Myxococcus xanthus. ISME J. 2020, 14(5), 1111–1124. [CrossRef]
- Chen, H.; Xiao, X.; Wang, J.; Wu, L.; Zheng, Z.; Yu, Z. Antagonistic effects of volatiles generated by Bacillus subtilis on spore germination and hyphal growth of the plant pathogen, Botrytis cinerea. Biotechnol. Lett. 2008, 30, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Boorn, K.L.; Khor, Y.Y.; Sweetman, E.; Tan, F.; Heard, T.A.; Hammer, K.A. Antimicrobial activity of honey from the stingless bee Trigona carbonaria determined by agar diffusion, agar dilution, broth microdilution and time-kill methodology. J. Appl. Microbiol. 2010, 108(5), 1534–1543. [Google Scholar] [CrossRef]
- Esteban, A.; Abarca, M.L.; Cabañes, F.J. Comparison of disk diffusion method and broth microdilution method for antifungal susceptibility testing of dermatophytes. Med. Mycol. 2005, 43(1), 61–66. [Google Scholar] [CrossRef]
- Garmendia, G.; Vero, S. Métodos para la desinfección de frutas y hortalizas. Hortic. 2006, 197, 18–27. [Google Scholar]
- Abdelmoteleb, A.; González-Mendoza, D. A novel Streptomyces rhizobacteria from desert soil with diverse anti-fungal properties. Rhizosphere. 2020, 16, 100243. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, H.; Deng, Y.; Tian, W.; Fan, G.; Sun, X. Antifungal Activity of Streptomyces hygroscopicus JY-22 against Alternaria alternata and Its Potential Application as a Biopesticide to Control Tobacco Brown Spot. Agronomy 2023, 13(7), 1944. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, D.; Yu, Z. New Lactones Produced by Streptomyces sp. SN5431 and Their Antifungal Activity against Bipolaris maydis. Microorganisms. 2023, 11(3), 616. [Google Scholar] [CrossRef]
- Santamaría, R.I.; Martínez-Carrasco, A.; Tormo, J.R.; Martín, J.; Genilloud, O.; Reyes, F.; Díaz, M. Interactions of Different Streptomyces Species and Myxococcus xanthus Affect Myxococcus Development and Induce the Production of DK-Xanthenes. Int. J. Mol. Sci. 2023, 24(21), 15659. [Google Scholar] [CrossRef]
- Afroz-Toma, M.; Rahman, M.H.; Rahman, M.S.; Arif, M.; Nazir, K.N.H.; Dufossé, L. Fungal Pigments: Carotenoids, Riboflavin, and Polyketides with Diverse Applications. J. Fungi. 2023, 9(4), 454. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, A. Metabolitos secundarios de actinomicetos. In Impacto de la biología molecular y las nuevas tecnologías en el conocimiento de la función celular y sus aplicaciones, 1st ed.; Fierro, F.F., Onofre, M.V. Universidad Autónoma Metropolitana, México, 2011; pp. 27-38.
- Evangelista-Martínez, Z.; Moreno-Enríquez, A. Metabolitos secundarios de importancia farmacéutica producidos por actinomicetos. Rev. Biotecnol. 2007, 11, 37–50. [Google Scholar]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; ... Foster, G.D. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13(4), 414–430. [CrossRef] [PubMed]
- Tsalidis, G.A. Human health and ecosystem quality benefits with life cycle assessment due to fungicides elimination in agriculture. Sustainability 2022, 14(2), 846. [Google Scholar] [CrossRef]
- Chacon-Lopez, A.; Guardado-Valdivia, L.; Banuelos-Gonzalez, M.; Lopez-Garcia, U.; Montalvo-González, E.; Arvizu-Gomez, J.; ... Aguilera, S. Effect of metabolites produced by Bacillus atrophaeus and Brevibacterium frigoritolerans strains on postharvest biocontrol of Alternaria alternata in tomato (Solanum lycopersicum L.). Biocontrol Sci. 2021, 26(2), 67–74. [CrossRef] [PubMed]
- Chaouachi, M.; Marzouk, T.; Jallouli, S.; Elkahoui, S.; Gentzbittel, L.; Ben, C.; Djébali, N. Activity assessment of tomato endophytic bacteria bioactive compounds for the postharvest biocontrol of Botrytis cinerea. Postharvest Biol. Technol. 2021, 172, 111389. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63(1), 34–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, J.H.; Liu, G.; Yao, X.F.; Li, P.F.; Yang, X.P. Characterization of the watermelon seedling infection process by Fusarium oxysporum f. sp. niveum. Plant Pathol. 2015, 64(5), 1076–1084. [Google Scholar] [CrossRef]
- Prins, T.W.; Tudzynski, P.; von Tiedemann, A.; Tudzynski, B.; Ten Have, A.; Hansen, M.E.; ... van Kan, J.A. Infection strategies of Botrytis cinerea and related necrotrophic pathogens. Fungal Pathol. 2000, 33–64. [CrossRef]
- Zhu, L.; Ni, W.; Liu, S.; Cai, B.; Xing, H.; Wang, S. Transcriptomics analysis of apple leaves in response to Alternaria alternata apple pathotype infection. Front. Plant Sci. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Carpena, R.; Ritter, A.; Socorro, A.R; Perez, N. Nitrogen evolution and fate in a Canary Islands (Spain) sprinkler fertigated banana plot. Agric. Water Manag. 2002, 52(2), 93–117. [Google Scholar] [CrossRef]
- de Cossío, Á.G. Nueva ayuda al sector de producción de plátanos de Canarias. Hacienda Canaria. 2008, (24), 129–152. [Google Scholar]
- Costa, J.M.; Heuvelink, E.P. The global tomato industry. In Tomatoes; Heuvelink, E.P., Ed.; Wallingford, United Kingdom, 2018; pp. 1–26. [Google Scholar]
Figure 1.
Light microscopy photomicrographs of actinomycetes smears. (a) Mycelium of F. oxysporum encapsulated by S. netropsis. Note the blue staining on the encapsulated fungal hyphae but not on the free hyphae. (b) Spores of B. cinerea encapsulated by S. ardesiacus. (c) Mycelium of A. alternata encapsulated by S. pratensis. (d) Hypha of B. cinerea encapsulated by S. mauvecolor.
Figure 1.
Light microscopy photomicrographs of actinomycetes smears. (a) Mycelium of F. oxysporum encapsulated by S. netropsis. Note the blue staining on the encapsulated fungal hyphae but not on the free hyphae. (b) Spores of B. cinerea encapsulated by S. ardesiacus. (c) Mycelium of A. alternata encapsulated by S. pratensis. (d) Hypha of B. cinerea encapsulated by S. mauvecolor.
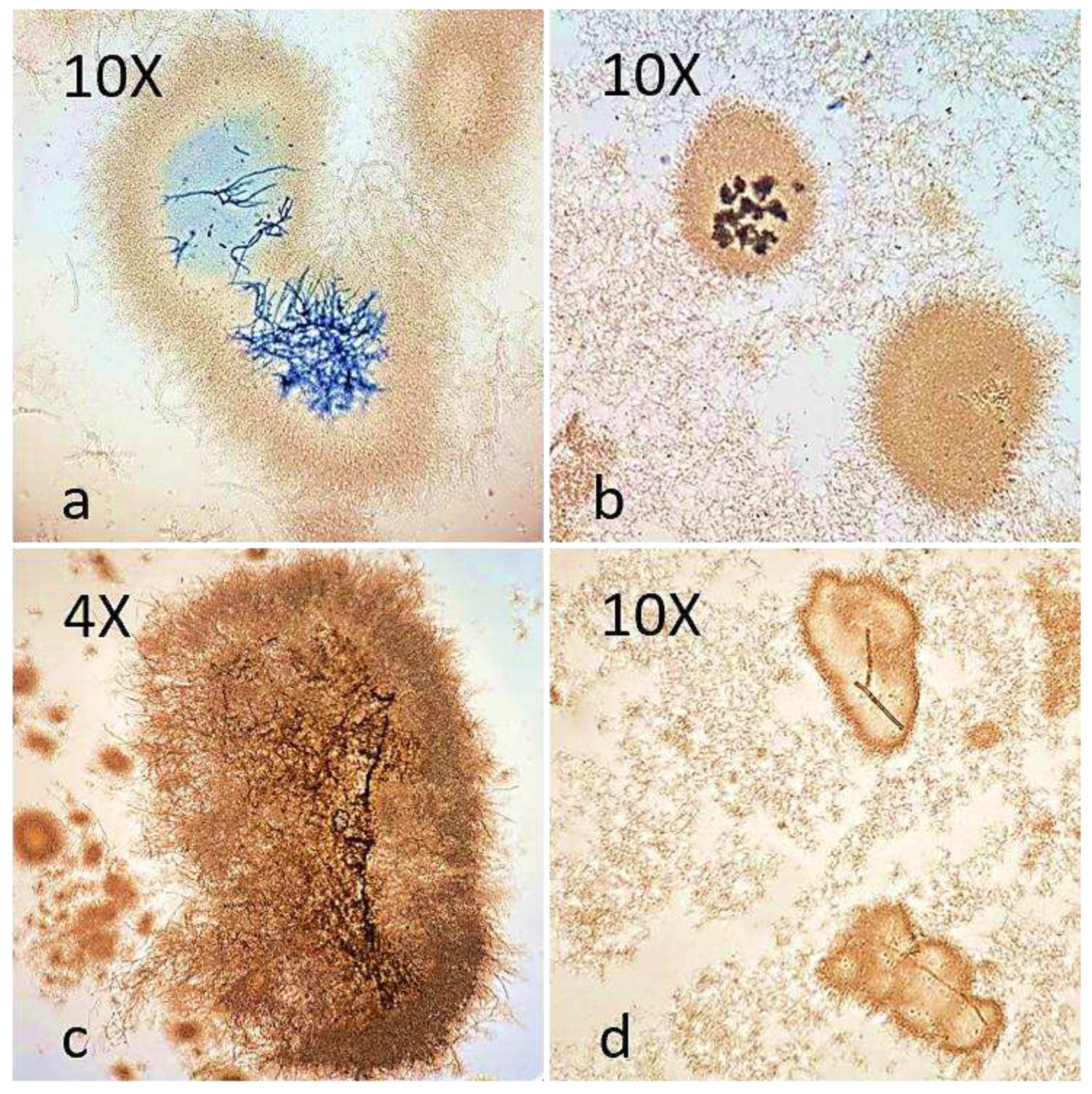
Figure 2.
Kinetics of production of compound(s) in supernatants of S. mauvecolor with activity in co-cultive with A. alternaria, B. cinerea and F. oxysporum against the three phytopathogenic fungi. (a) Kinetics of activity against A. alternata and plate at 5 days. (b) Kinetics of activity against B. cinerea and plate at 5 days (c) Kinetics and plate growth of the activity against F. oxysporum at 5 days of co-culture. Note that b and c are partial affection activity.
Figure 2.
Kinetics of production of compound(s) in supernatants of S. mauvecolor with activity in co-cultive with A. alternaria, B. cinerea and F. oxysporum against the three phytopathogenic fungi. (a) Kinetics of activity against A. alternata and plate at 5 days. (b) Kinetics of activity against B. cinerea and plate at 5 days (c) Kinetics and plate growth of the activity against F. oxysporum at 5 days of co-culture. Note that b and c are partial affection activity.
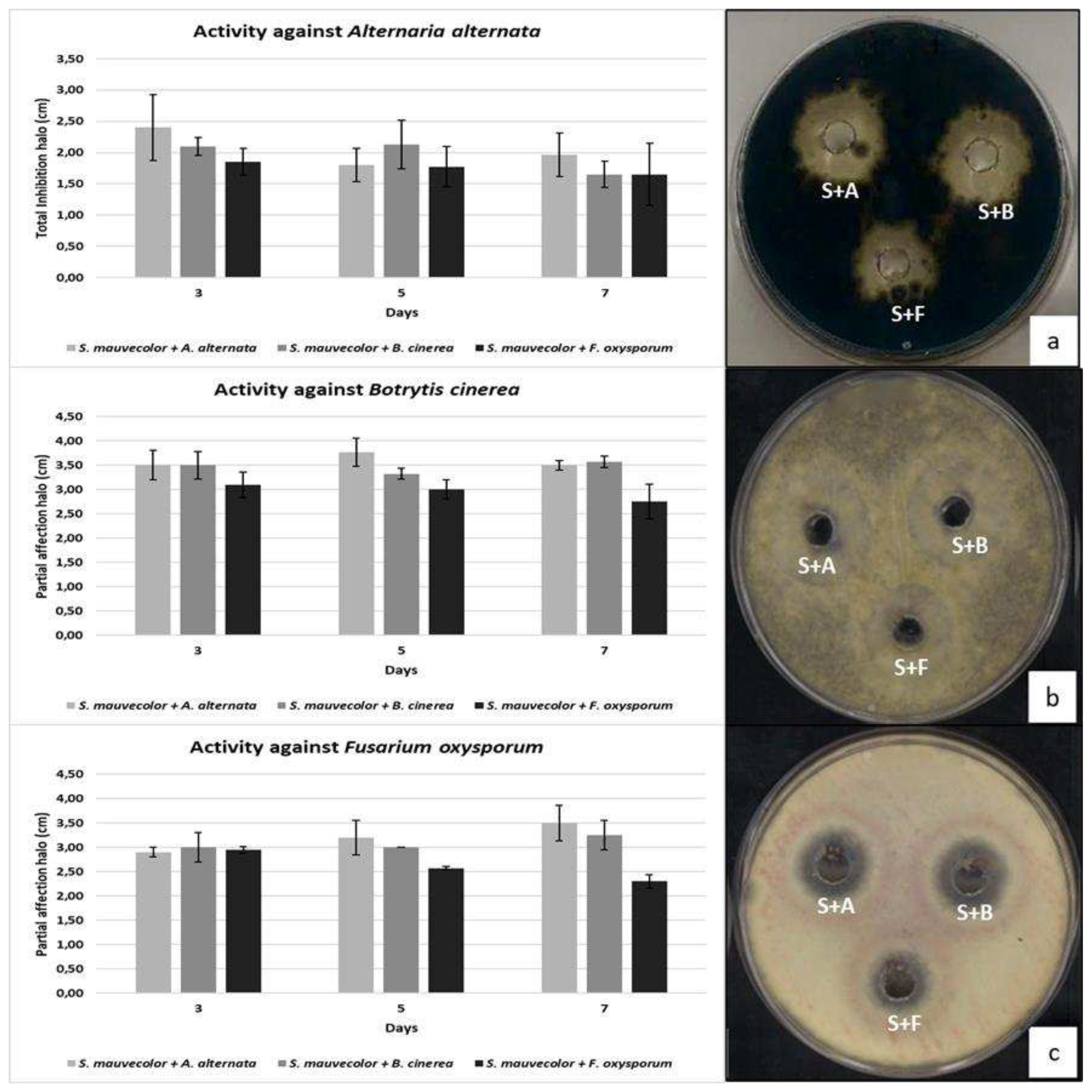
Figure 3.
Effect of different actinomycetal extracts against three phytopathogenic fungi. (a) Plate of B. cinerea with S. pratensis supernatant. Note the double halo, an inner halo without fungal growth and an outer halo where sporulation is inhibited. (b-d) Phytopathogens with S. ardesiacus supernatant. (b) Plate of A. alternata. Note the low mycelial density around the wells. (c) Plate of B. cinerea. Note the halo with affected fungal growth, without conidia formation. (d) Plate of F. oxysporum. Note the greatly reduced fungal growth around the well and the outer halo with increased mycelial density but no sporulation. (e-g) Phytopathogens with S. netropsis supernatant. (e) F. oxysporum plate. Note the low mycelial density around the wells, with no spore formation. (f) Plate of A. alternata. Note the large fungal growth inhibition halo. (g) Plate of B. cinerea. Note the large fungal growth inhibition halo. (h-j) Phytopathogens with supernatants of S. mauvecolor. (h) Plate of A. alternata. Note the mycelial growth inhibition halo. (i) Plate of B. cinerea. Note the halo with affected fungal growth, without conidia formation. (j) Plate of F. oxysporum. Note the total inhibition of fungal growth around the well and the external halo with mycelium development but without sporulation.
Figure 3.
Effect of different actinomycetal extracts against three phytopathogenic fungi. (a) Plate of B. cinerea with S. pratensis supernatant. Note the double halo, an inner halo without fungal growth and an outer halo where sporulation is inhibited. (b-d) Phytopathogens with S. ardesiacus supernatant. (b) Plate of A. alternata. Note the low mycelial density around the wells. (c) Plate of B. cinerea. Note the halo with affected fungal growth, without conidia formation. (d) Plate of F. oxysporum. Note the greatly reduced fungal growth around the well and the outer halo with increased mycelial density but no sporulation. (e-g) Phytopathogens with S. netropsis supernatant. (e) F. oxysporum plate. Note the low mycelial density around the wells, with no spore formation. (f) Plate of A. alternata. Note the large fungal growth inhibition halo. (g) Plate of B. cinerea. Note the large fungal growth inhibition halo. (h-j) Phytopathogens with supernatants of S. mauvecolor. (h) Plate of A. alternata. Note the mycelial growth inhibition halo. (i) Plate of B. cinerea. Note the halo with affected fungal growth, without conidia formation. (j) Plate of F. oxysporum. Note the total inhibition of fungal growth around the well and the external halo with mycelium development but without sporulation.
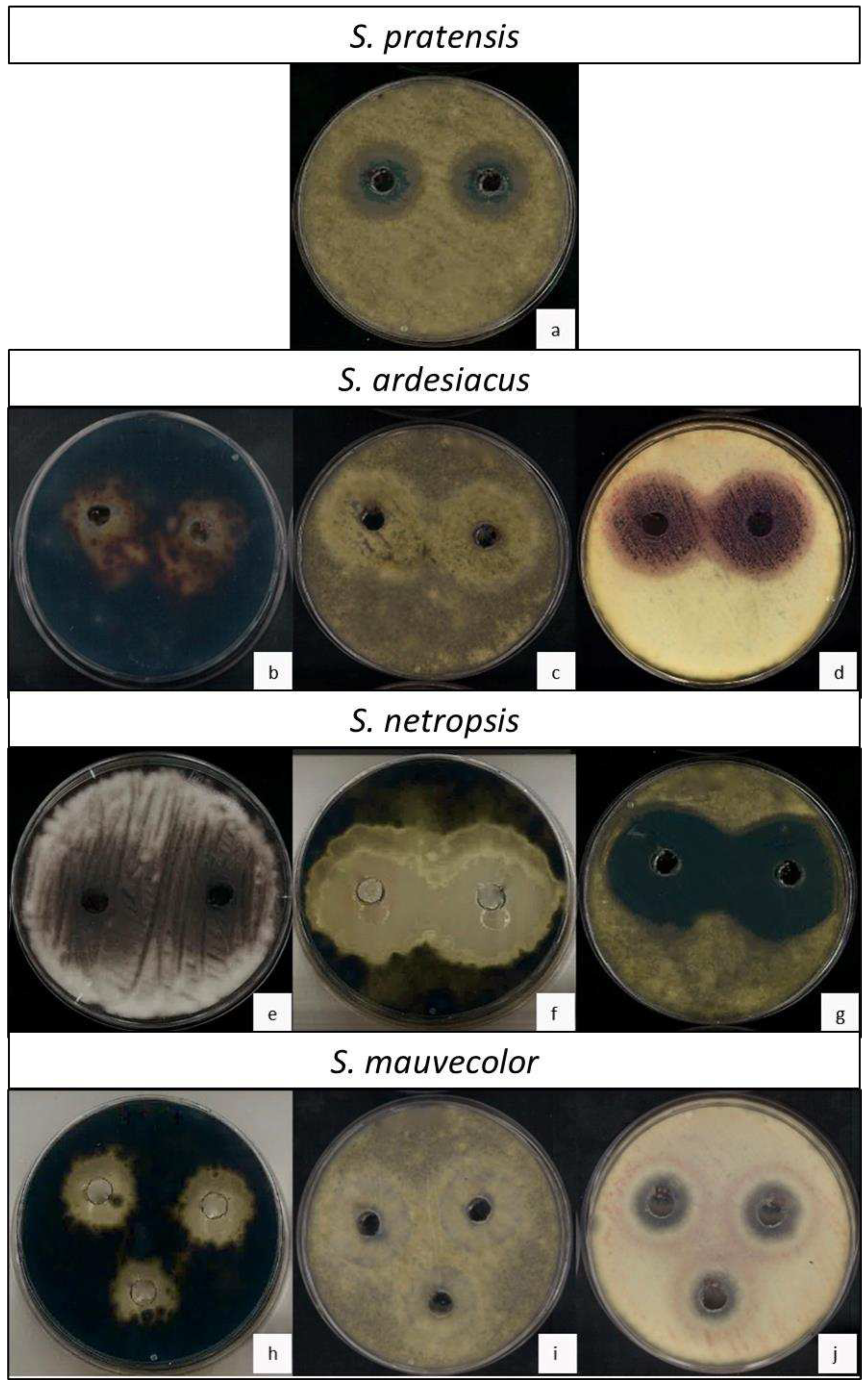
Figure 4.
B. cinerea behavior against S. netropsis extracts. (a) Crude extract. (b) Extract in ethyl acetate. The halo on the right shows the organic phase (EA-org.f), while the halo on the left shows the aqueous phase (EA-aq.f). (c) Extract in isobutyl alcohol. The halo on the right shows the organic phase (iB-org.f), while the halo on the left shows the aqueous phase (iB-aq.f).
Figure 4.
B. cinerea behavior against S. netropsis extracts. (a) Crude extract. (b) Extract in ethyl acetate. The halo on the right shows the organic phase (EA-org.f), while the halo on the left shows the aqueous phase (EA-aq.f). (c) Extract in isobutyl alcohol. The halo on the right shows the organic phase (iB-org.f), while the halo on the left shows the aqueous phase (iB-aq.f).
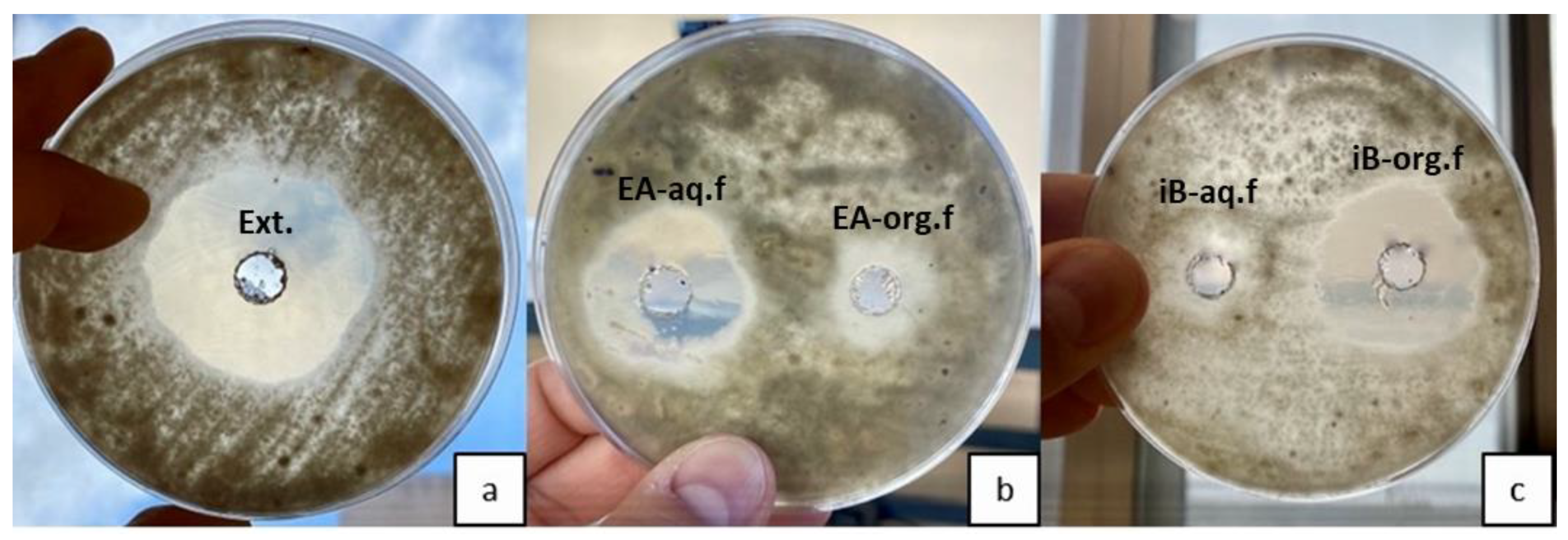
Figure 5.
Cherry tomato fruits infected by B. cinerea. (a) Control (C1) - 100% infection. (b) Control booster (C2) - 90% infection. (c) Treatment with S. netropsis extract with one application (A1) - 50% infection. (d) Treatment with S. netropsis extract with booster (A2) - 50% infection.
Figure 5.
Cherry tomato fruits infected by B. cinerea. (a) Control (C1) - 100% infection. (b) Control booster (C2) - 90% infection. (c) Treatment with S. netropsis extract with one application (A1) - 50% infection. (d) Treatment with S. netropsis extract with booster (A2) - 50% infection.

Figure 6.
Surface and internal infection of cherry tomatoes by B. cinerea in %. Values with the same letter do not differ significantly (p ≤ 0.05%).
Figure 6.
Surface and internal infection of cherry tomatoes by B. cinerea in %. Values with the same letter do not differ significantly (p ≤ 0.05%).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



