Submitted:
13 November 2023
Posted:
13 November 2023
You are already at the latest version
Abstract
High-resolution forecasting of vegetation type shifts may prove essential for anticipating and mitigating the impacts of future climate change on bird populations. Here, we used the US Forest Service Ecological Response Unit (ERU) classification to develop and assess vegetation-based breeding habitat profiles for eight owl species occurring in foothills and mountains of the southwestern US. Shifts in mapped habitat were forecast using an ecosystem vulnerability model based on the pre-1990 climate envelopes of ERUs and the A1B moderate IPCC emission scenario of future climate. For five of the eight owl species, regional breeding habitat extent was projected to decline by at least 60% by 2090. Three species, the boreal owl (Aegolius funereus; at the trailing edge of its distribution), flammulated owl (Psiloscops flammeolus), and northern pygmy-owl (Glaucidium gnoma) were projected to experience the steepest habitat-loss rates, or 85%, 85%, and 76%, respectively. Projected vegetation shifts overlaid with well-documented flammulated owl breeding populations showed complete or near complete loss of habitat by 2090 in areas of montane forest currently supporting dense aggregations of owl territories. Generalist or lower-elevation owl species were predicted to be less impacted, while for the whiskered screech-owl (Megascops trichopsis), the contraction of current habitat was nearly offset by a projected northward expansion. In general, the results of this study suggest high exposure to climate change impacts for upper-elevation forest owls of semi-arid southwestern North America. Long distance migration and low natal philopatry may prove important to some montane owl populations for adapting to regional loss of habitat.
Keywords:
Aegolius acadicus
; Aegolius funereus
; Asio otus
; boreal owl
; Bubo virginianus
; climate change
; ecosystem vulnerability model
; flammulated owl
; Glaucidium gnoma
; habitat model forecasts
; long-eared owl
; Megascops kennicotti
; Megascops trichopsis
; northern pygmy-owl
; northern saw-whet owl
; great horned owl
; Psiloscops flammeolus
; western screech-owl
; whiskered screech-owl
1. Introduction
If left unchecked, anthropogenic climate change could soon result in the extinction of one in six species on Earth [1]. To minimize global (and regional) biodiversity losses, there is an urgent need to identify both the species at risk and the extent of their vulnerability [2,3]. A useful framework for assessing vulnerability to climate change relies on the three concepts of exposure, sensitivity, and adaptive capacity [3]. Exposure is defined as the degree to which a species may experience change in its surrounding environment, whether from temperature or precipitation shifts, altered vegetation cover, or the disruption of trophic interactions. Sensitivity relates to the species’ dependence on specific environmental or ecological conditions that are projected to change. Adaptive capacity is the ability of a species to persist under changing conditions through dispersal, migration, or other adaptation mechanisms [4,5].
Several approaches have been proposed to assess climate change risks and vulnerabilities in bird populations. Already measurable and/or predicted climate change impacts on owls in particular have been studied using demographic models based on simulated weather variables and their effect on population vital rates, either directly or through their interplay with trophic interactions e.g., [6,7]. In high-Arctic Greenland, snowy owl (Bubo scandiacus) reproductive success and population densities were predicted to decline due to ongoing demographic shifts (i.e., collapsing of the high-amplitude population cycle) in a main prey, the collared lemming (Dicrostonyx groenlandicus) [8,9]. Similarly, stochastic demographic modelling showed that already observable dampening vole prey cycles may be driving a tawny owl (Strix aluco) population from northern England toward extinction [6]. At much lower latitudes, in Switzerland, Jenouvrier [10] projected a dramatic barn owl (Tyto alba) population decline under a scenario of non-linear relationships between climate change and population vital rates. During past, extreme harsh winters, recorded only twice in a 58-year monitoring period, adult and juvenile barn owl survivorship plummeted due to extended snow cover duration reducing small mammal prey availability [11]; under the scenario of non-linear climate change, the incidence of extreme harsh winters would increase with a direct impact on long-term barn owl stochastic growth rate [10]. In the southwestern U.S., negative associations between warm, dry conditions and spotted owl (Strix occidentalis) fecundity led to predictions of rapid population declines and a much higher probability of extinction under three emission scenarios compared to current climate conditions [7]. Compared to other birds, spotted owls have relatively narrow neutral thermal zones as they are seemingly less heat-tolerant [12,13].
Here, we provide a new approach to predicting impacts on owl populations at a local and regional scale based on the Intergovernmental Panel on Climate Change’s (IPCC) A1B moderate IPCC emission scenario of future climate [14] and an ecosystem-based vulnerability assessment developed for the southwestern U.S. (i.e., the two states of Arizona and New Mexico) [15]. The vulnerability assessment is represented by the level of departure for late 21st century climate from the characteristic pre-1990 climate envelope of the ecosystem type at each given location, resulting in a probability surface of climate impacts for the two-state area [15]. This emission scenario is considered a plausible balance of expected technological and energy development and a standard representative for mid-range climate forcing and climate research [14,15].
Land cover in southwestern North America has changed dramatically since the last ice age, when climate conditions were significantly cooler and more humid and the vegetation consisted largely of coniferous forests and woodlands, all as recently as 12,000 years ago [16,17]. Xerophytic vegetation types would have been limited in extent and concentrated in southernmost areas of the region. The abundance and location of ecosystem types found today in southwestern North America reflect a warmer and drier climate than that 12,000 years ago, the result of natural change over time [18]. High topographic relief has facilitated a conspicuous elevational zonation of the vegetation with xerophytic vegetation (e.g., grassland, shrubland, and desert scrub) at lower elevations grading into more mesic plant communities (woodlands and forests) at upper elevations (Figure 1). A number of owl species breed in southwestern North America in association with those woodlands and forests, and increasingly warmer and drier conditions brought about by ongoing and projected climate change raise the possibility of large declines in the extent of their breeding habitat as woodlands and forests retreat farther up the elevational gradient. The model based on the climate envelopes of individual ecosystem types and the magnitude of departure from those envelopes allows quantitative projections of vegetation-based habitat by the late 21st century [19]. This type of analysis has implications for arid southwestern North America and similar environments elsewhere in the World.
2. Materials and Methods
Habitat profiles and spatial rendering
Spatial models were developed to represent the approximate extent and distribution of breeding habitat for eight species of owls in montane woodlands and forests of the southwestern US: flammulated owl (Psiloscops flammeolus)., western screech-owl (Megascops kennicottii), whiskered screech-owl (M. trichopsis), great horned owl (Bubo virginianus), northern pygmy-owl (Glaucidium gnoma), long-eared owl (Asio otus), boreal owl (Aegolius funereus), and northern saw-whet owl (A. acadicus).
As a first step, breeding habitat profiles were created for each of those species based on a literature review and expert opinion and using the 2017 version of Ecological Response Units [20], each Ecological Response Unit (ERU) being similar in vegetation potential and plant dominants, historical fire regime, plant succession, and key ecological processes (Table 1). Vegetation provides the basic structure and the primary functions for ecosystems and species habitat [21]. Vegetation is a major environmental factor influencing habitat selection in bird species [22,23], while high-resolution mapping of vegetation cover is also essential to model the distribution of populations and improve the management of breeding habitats [24]. Not included in our analysis was the Mexican spotted owl (Strix occidentalis lucida), a threatened subspecies of the spotted owl occurring in the southwestern U.S. Although the Mexican spotted owl is a montane owl, extensive research on that subspecies shows that it selects its habitat based on topography and/or canopy closure rather than vegetation type [25,26].
Of the eight owl species, five have their distribution in the southwestern U.S. restricted to wooded foothills and mountains during the breeding season e.g., [27,28], and the habitat profiles used for this study approximated the totality of their regional breeding habitat distribution in 2017. To varying extents, western screech-owls, great horned owls, and long-eared owls also breed in floodplain riparian woodlands and in other low-elevation vegetation types not found in foothills and mountains [27,28]. The montane habitat profiles of those three species therefore represented only partial representations of their total habitat distributions in the southwestern U.S. We excluded pinyon-juniper woodland and Madrean pinyon-oak woodland from the habitat profiles of the flammulated owl and the northern saw-whet owl as the relatively small number of breeding records from those vegetation types tended to be restricted to the ecotone with ponderosa pine forest, with denser and taller pinyon pines (Pinus spp.) and junipers (Juniperus spp.) and scattered ponderosa pines (Pinus ponderosa) [29,30].
Spatial rendering of each profile was performed in GIS using state boundaries to focus on Arizona and New Mexico, a geodataset of all Ecological Response Units and their climate vulnerability forecasts [15], and a layer of Ecological Subregions [31] to constrain species biogeography. Figure 2 shows the model zones used to geographically limit habitat profiles for some species, each zone composed of one or more Subregions.
Ecosystem vulnerability model and projections of future habitat distribution
Projections of future habitat distribution were based on measured shifts in climate conditions, from past to future, and expected changes in vegetation pattern based on suitable climate conditions among ERUs and their known climate envelopes [15]. Climate envelopes were built with five climate variables (e.g., Julian date that the sum of degree-days >5°C reaches 100degree-days >5°C based on mean monthly temperature) found to optimize class separation of ERUs and selected from over 20 variables including several indices corresponding to combinations of temperature and precipitation measurements [15]. The following equation was used to assign the vulnerability of an ERU at a given location.
where VS is the vulnerability score for a polygon, is the mean of one pre-1990 climate variable, Valseg is the Year 2090 value for the variable and polygon segment, and s is the inter-annual, pre-1990 climate standard deviation for the ERU [15].
The same climate variables used to develop the underlying vulnerability surface were applied to determine the most likely outcomes in the future distribution of ERUs, according to forecast climate exposure relative to conditions represented by each envelope. Multiple studies from the region and the western US corroborate directional change in vegetation patterns anticipated with warmer and drier conditions, including elevational shifts [32,33,34,35] and associated changes in plant composition [15,36,37]. For instance, tree species recruitment in ponderosa pine communities with the greatest climate departure projections are showing increased probability in the regeneration of pinyon and juniper species, components of warmer-dryer downslope affinities [15]. Vegetation type projections sometimes indicate more than one possible outcome due to overlap among some climate envelopes. In these cases, the interpretation of future distribution erred towards inclusion for any given habitat profile. For upland shrub types (not selected in any breeding habitat profile, but present in the woodland and montane zones), a specific rule was applied to narrow potential transitions of shrub types to other shrub vegetation types, given the importance of site factors (especially slope, aspect, edaphic properties), shrub recruitment strategies, and natural fire regimes in perpetuating shrub dominance [38,39]. Thus, no shrub types were projected to convert to woodlands and forests under climate model projections. But woodlands, at their lower ecotone, could be projected to grassland or shrubland types (e.g., Colorado Plateau/Great Basin Grassland, Intermountain Salt Scrub).
Our analyses were restricted to mid-to-upper elevation ecosystem types and did not include lowlands including desert areas, for which some uncertainty remains regarding climate envelopes. Samples for the four desert ERUs (Chihuahuan Desert Scrub, Chihuahuan Salt Desert Scrub, Mojave-Sonoran Desert Scrub, and Sonora-Mojave Mixed Salt Desert Scrub) were available only from the northern extents of the Chihuahuan and Sonoran provinces, in turn possibly skewing climate envelopes and overpredicting vulnerability (15). All four of these systems are especially resistant to stress and drought extremes, as well as to variability of environmental conditions across temporal scales [40,41,42].
For all species, current and future habitat models were parameterized conservatively, erring to commission, by the inclusion of vegetation types and model zones. In the case of the whiskered screech owl, a Central American species whose range extends just into the southern borderlands of Arizona and Mexico, an allowance was made for the future projection and potential northward expansion of the species by adding two model zones.
Habitat model validation datasets
Habitat model validation was performed using a total of 834 locational data points corresponding to geo-referenced museum specimen records, i-Naturalist research-grade verified observations (non-duplicate locations only), and owl or owl territory detections in survey or monitoring datasets. All records with a precision > 1 km were excluded. Other than western and whiskered screech-owls, known to remain on their territories year-round, museum specimen records were restricted to the months of May–July to avoid inclusion of data points during migration. I-Naturalist records used for habitat model validation consisted of those during the same period of May–July and/or based on breeding pair, nest, or fledgling photos or recorded singing. Owl surveys were conducted during the May–July period and detections typically involved hearing territorial calls. Our occurrence data do not represent a statistical sample; however, these records provide a sense of data quality of habitat mapping and the degree of model omission and commission. A habitat buffer of 1 km was used to account for uncertainty in both habitat mapping and occurrence records. Owl survey datasets were also used to investigate projected shifts in vegetation conditions in areas with documented owl populations or inference of higher-than-average owl densities. Though typically limited in terms of their spatiotemporal coverage, survey datasets consist of presence-absence data found to perform better in predictive models than opportunistic records and other presence only data [43]. At a local scale at least, owl survey datasets likely improved the accuracy of our assessment of projected climate-induced impacts on the extent of habitat for some owl species.
Boreal owl surveys (New Mexico)
Boreal Owls were recorded during standardized, conspecific playback surveys in northern New Mexico during 1989–1993 [44]. Documented or suspected breeding locales were all revisited in 2012, with additional sites also surveyed [45]. All detections occurred in just three mountain ranges [44,45]. The high rate of detection success at revisited sites in 2012 suggested long-persisting populations [45]. For this study, all documented boreal owl territories were overlaid on maps showing the current and predicted late-21st century distribution of vegetation types generalized from ERUs. The percentage of territories included within our habitat model under current and projected conditions was calculated in ArcGIS.
Flammulated owl surveys (New Mexico)
From 1996 through 1998, systematic flammulated owl surveys were conducted between 2000 and 2700 m in the Jemez, Zuni, Magdalena, and San Mateo mountains, and in the Black Range, all in western New Mexico [30]. Additional surveys were completed through 2006 and in 2011 for the Oso Ridge flammulated owl population in the Zuni Mountains (D. Arsenault, unpubl. data). All surveys involved the search for flammulated owl territories during the nesting season, and nest and territorial male locations were determined with an accuracy of ≤ 3 and 10 m, respectively. The surrounding vegetation consisted primarily of ponderosa pine and Gambel oak (Quercus gambelii) forests with herbaceous understories and scattered shrubs; remnant groves of narrowleaf cottonwood (Populus angustifolia) in some riparian areas; and ponderosa pine mixed with quaking aspen (Populus tremuloides) and Douglas-fir (Pseudotsuga menziesii) at upper elevations and with pinyon pine (Pinus edulis) at mid-elevations [30]. From 1996 through 1998, a total of 86 occupied territories were documented and georeferenced along 149 km of transects [30]. The total number of occupied nesting territories reached 148 with the later surveys in the Zuni Mountains. Owls were detected in montane forest at elevations ranging from 2260 m (2070 m in the Magdalena Mountains) to 2680 m. All documented flammulated owl territories were overlaid on maps showing the current and predicted late 21st century distribution of vegetation types generalized from ERUs. The percentage of territories included within our habitat model under current and projected conditions was again calculated in ArcGIS.
Quadrat owl detections (Arizona and New Mexico)
Location data on various species of owls were collected opportunistically in 1999 during nocturnal, systemic broadcast Mexican spotted owl surveys conducted in 25 study areas or quadrats of varying size scattered across central Arizona and into New Mexico [46]. Surveys were solely aimed at detecting Mexican spotted owls. However, non-target owls are often heard or seen during nocturnal surveys, with some species even known to respond to heterospecific call-broadcast [47,48]. A few of the locations were visual sightings, but most were auditory detections. Surveys were conducted from georeferenced call points. If an owl was very close to that point, observers recorded the call point location as the owl location. For more distant owls, observers estimated distance and bearing to the calling owl from the call point and then recorded spatial coordinates from an estimated map location based on that distance and bearing. A total of 303 separate detections were recorded, mostly of great horned owls and flammulated owls, but also including smaller numbers of some of the other owls included in this study. The locations all represented calling owls, not nests. Sampling effort was similar in that call points were generated in each quadrat to ensure complete survey coverage, and multiple surveys were completed at all call points. The total number of complete surveys varied from 3–6 per quadrat, reflecting the degree of difficulty of completing surveys (access was more difficult in roadless quadrats). Although heterospecific call-broadcast has variable effects on detectability across owl species [48], it likely would not produce any bias in detectability among habitats within species.
A total of 262 detections were recorded within the 25 quadrats. Within a quadrat, multiple locations may have corresponded to a smaller number of unique individuals or territories, constraining strength of inference regarding habitat associations. In one quadrat with multiple northern saw-whet owl detections, we created 250 m buffers around each detection point in ArcMAP to limit overlap with adjacent points and generate what we then considered separate samples of habitat use. In the Sierra Nevada, Groce and Morrison [49] used the same approach to obtain a general sense of habitat associations at a scale greater than a nest or roost site.
Great horned owl detections (New Mexico)
Great horned owls are known to readily respond to heterospecific call-broadcast [47]. During spotted owl surveys in south-central New Mexico’s Sacramento Mountains from 2003–2006 [50], great horned owl detections were opportunistically also recorded (J. Ganey, unpubl. data). The study area covered approximately 50,000 ha at elevations ranging from 2,000 m to 2,800 m. The terrain consisted primarily of heavily forested montane slopes and minor drainages, with interspersed meadows in the larger valley bottoms. The predominant forest type was mixed-conifer, singularly or co-dominated by white fir (Abies concolor) and Douglas-fir, with more limited extents of ponderosa pine forest. All recorded locations (n = 45) represented calling owls, and most detections were auditory, a few visual observations.
Whiskered and western screech-owl nest box study (Arizona)
A nest box study was conducted during the breeding seasons of 2015–2022 in Miller Creek Canyon in the Peloncillo Mountains in southeastern Arizona [51]. Elevation ranged from 1,553 to 1,610 m. The vegetation within the study area was dominated by an oak overstory and a mixed understory of alligator juniper (Juniperus deppeana), pointleaf manzanita (Arctostaphylos pungens), and catclaw acacia (Senegalia greggii). A total of 15 nest boxes installed in the study area were monitored for occupancy and nesting success [51].
Spatial model validation
Validation is the most challenging task in the habitat modelling process, with no single test or approach accepted as the gold standard for assessing model performance [52,53]. Compositional procedures have been recommended for the validation of habitat models in the absence of ad hoc, presence and absence datasets [53]. We used Arc GIS Pro and a spatial randomized sampling procedure to test the performance of our species habitat profiles and habitat maps under current conditions. Within an area corresponding to all woodland and forest types (woodland, montane, and subalpine zones; Table 1), we randomly generated sample points equal in number to the occurrence records for a given species in our validation dataset. To exclude areas where species are known to be absent, the sampling procedure was constrained geographically for the whiskered screech-owl (Sky Island -Bootheel ecoregion; Figure 2), boreal owl (San Juan-Sangre de Cristo and Jemez mountains ecoregions), and northern saw-whet owl (all ecoregions except for the Sonoran ecoregion). Using the Python function arcpy.analysis.Intersect(), we then calculated the number of random points intersecting with polygons representing the predicted habitat of the species. For every species, the randomization procedure was repeated for a total of 500 iterations before applying one-tailed z-tests to test the null hypothesis, “the proportion of occurrence records captured by the predicted habitat model is not significantly greater than that of a randomly generated sample”. Z-tests were conducted in R 4.3.1 (R Core Team 2022) using proportions of records falling within predicted mapped habitat with and without the 1-km buffer that was used to account for spatial error in mapping and occurrence records (e.g., species recorded in grassland within 1 km of a woodland or forest polygon).
Assessment of climate change impacts
For each of the eight species, we used R 4.3.1 and McNemar tests for paired, binary response data to test the null hypothesis that the proportion of records or detection points within vs. outside 2017 mapped modeled habitat does not differ between current and projected climate-induced conditions. McNemar tests are preferred over Chi-squared tests to determine whether there is a significant change in nominal data before and after an event (in this case climate change), based on repeated observations of the same individuals, with data presented in a 2x2 table showing the numbers of concordant vs. discordant cells instead of the Chi-squared test’s contingency table [54]. Tests were repeated with and without the 1-km buffer around mapped habitat. McNemar’s continuity correction was applied whenever any of the cell counts in the 2x2 matrix were less than 5.
3. Results
3.1. Spatial Model Validation
In terms of omission errors, our current (2017) habitat model performed well for five species, with 100% of geo-referenced records captured within the buffer for the boreal owl and whiskered screech-owl, 98.6% for the northern pygmy-owl, 95.6% for the flammulated owl, and 93.2% for the great-horned owl (Table 2). Performance was lower for the northern saw-whet owl (82.4% captures within the buffer), long-eared owl (78.3%), and western screech-owl (53.3%). For all 8 species, use of the 1-km buffer improved the performance of our habitat model. Some records fell just outside mapped habitat, as was the case for two of the three boreal owl records not directly captured by the model but within less than 100 m of mapped extents (and thus captured inside the buffer). Without applying the buffer, as many as 88 (34%) of all 256 records outside mapped habitat fell in riparian areas nestled within woodlands and forests, another 48 (19%) in montane/subalpine and other grasslands. For the whiskered screech-owls, 23 riparian records represented the majority (56%) of all whiskered screech-owl records falling outside mapped habitat. Of the total number of northern pygmy-owl records outside mapped habitat, 16 (73%) also occurred in riparian areas. Smaller numbers of records mapped in upland shrub including 8 flammulated owl records falling in Mojave-Sonoran Desert Scrub evidently represented mapping errors, which were limitations of the validation dataset.
Without applying the 1-km buffer, habitat models significantly outperformed randomized samples (generated from woodlands and forests) in the case of the flammulated owl, northern pygmy-owl, long-eared owl, boreal owl, and northern saw-whet owl; results for the western screech-owl were marginally significant (Table 2). With the 1-km buffer included, the western screech-owl and whiskered screech-owl habitat models also significantly outperformed random sampling procedures. Everything else being equal, it becomes increasingly more difficult for a habitat model to outperform randomization procedures as the number of woodland forest ERUs in the habitat profile increases. Because all woodland and forest types were included in the great horned owl habitat profile, all random points (from woodland and forest types) necessarily intersected with that profile. Lack of significant results for the great horned owl thus does not mean that its habitat model is flawed. Nonetheless, greater caution is needed in interpreting model projections for that species, in addition to the western and whiskered screech-owls, given the likelihood for greater uncertainty in the results.
3.2. Climate Change Projections
Under current conditions, approximately 88% of all owl survey records were captured by the habitat profile mapping when allowing for a 1-km buffer to account for spatial accuracy in the data. When intersected with late 21st century ERU projections, only about 43% of owl records or detection points fell within modeled habitat (with the 1-km buffer), with percent capture values ranging between 5% for the whiskered screech-owl and nearly 90% for the great horned owl but overall much lower than under current conditions for nearly every species (Figure 3).
When using the 1-km buffer, McNemar’s test results showed that the proportion of owl record locations falling outside our spatial model was indeed significantly higher under projected future conditions compared to current conditions for all but one species (Table 3). Results for the great horned owl were only marginally significant. Without applying the 1-km buffer, the proportion of owl records falling outside our mapped habitat was significantly higher under projected conditions for all eight owl species.
Comparison of the current and projected distribution of modeled owl habitat indicated range contractions within the region for all eight owl species with mid to upper-elevation habitat declines varying from -5% to -86%, and with the boreal and flammulated owls predicted to have the greatest habitat loss (Figure 4, Table 4). The habitat range of one species, the whiskered screech-owl, was projected to expand northward, nearly offsetting losses in habitat extent over the long term, with an estimated decrease of only 5%. Wherever habitat gains were projected (mostly observed for the whiskered and western screech-owls), new mapped habitat was typically not adjacent to existing mapped habitat. Current mapped habitat for the boreal owl (all in norther New Mexico) consisted of 899 dissolved polygons (i.e., separate patches) ranging in size from 0.12 to 80,179 ha (x̄ = 331.39 ha ± 3146.82). Projected 2090 mapped habitat was reduced to 311 separate patches averaging 133.59 ha ± 615.22 with a maximum size of just under 6,128 ha. The 8 boreal owl records still within mapped habitat under projected future conditions were located within 5 patches totaling 6,401 ha (x̄ =1,280 ha, range= 11.3-3,563 ha), compared to 6 patches totaling 149,285 ha (x̄ =24,881 ha, range= 4,162- 80,179 ha) under current conditions.
3.2. Flammulated owl breeding populations
Projected flammulated owl habitat shifts affecting 148 documented breeding territories across five mountain ranges of New Mexico (32; D. Arsenault, unpubl. data) are summarized in Table 5 and shown in Figure 7. About 92% of territories were found to currently fall within the expected habitat profile of flammulated owls, with a strong preference for dry forests (ponderosa pine, pine oak, and frequent-fire mixed conifer forests). Three territories occurred in pinyon-juniper stands within 600 m or less of dry forest and leaving open the possibility of spatial uncertainty in vegetation type mapping or territory records. Nine documented territories occurred in nearby riparian corridors or shrub types, all within 200 meters of dry forest. Late 21st century habitat projections suggested that climate conditions in over 80% of territories currently suitable for owl habitat will favor the development of pinyon-juniper and Madrean woodland types, with the possibility of some territories falling into upland shrublands and less than 10% of current owl territories remaining within the present concept of flammulated owl habitat.
Of the five flammulated owl populations analyzed, four were projected to face vegetation shifts placing all or nearly all current nesting territories outside the species’ habitat profile, whether pinyon-juniper woodland, Madrean woodland, or upland shrub (Figure 5). The only exception was the Jemez Mountains flammulated owl population, which would still find suitable vegetation where nesting territories currently exist.
3.3. Northern saw-whet detection points
As the best approximation of habitat use by northern saw-whet owls in any given area (based on our dataset), or even of the existence of a breeding population, a total of 13 independent detection points (i.e., detection points with non-overlapping, 250-m-diameter buffers around them) for that species occurred in one sampling quadrat in Arizona. Of the 13 independent detection points, 12 (92%) were mapped within ponderosa pine forest, which covered 93% of the quadrat’s area (Figure 6). Under 2090 projections, however, all of those detection points were mapped outside the species’ habitat profile.
4. Discussion
Differences between current and future habitat distribution reflect a 21st-century departure from historical climate envelopes for significant underlying features of vegetation types at any given location. Modeled habitat distributions suggest substantial change in familiar ecosystem conditions and, in particular, significant loss in montane woodland and forest habitat for most owl species evaluated with this study. This forecast does not factor disturbance processes such as wildfires, broadly considered catalysts for rapid and permanent type conversion of vegetation under this century’s climate trends [55].
The modeling results show that climate-driven range contractions can be expected in the southwestern U.S. for several owl species. Of those species assessed, the boreal owl is arguably the most sensitive to warming trends as trailing-edge populations (near the low-latitude edge of the distribution) are generally predicted to face the highest risk of climate-induced extinction [56]. The southernmost part of the boreal owl’s range narrowly extends into three mountain ranges in northern New Mexico [44,45], and 2090 projected conditions include the complete loss of historical habitat in one of those mountain ranges, in addition to severe habitat losses in the other two ranges (Figure 4). Particularly concerning are our projections of important reductions in the size of future, individual mapped habitat areas for the boreal owl in New Mexico. Summer home range areas in Idaho were found to average 1,182 ± 335 ha for 15 owls, with an observed minimum of 229 ha and an observed maximum of 2,386 ha [57]. Although in Idaho boreal owl were not restricted to spruce-fir forest (they nested in mixed-conifer forest where they presumably found more suitable nesting cavities but roosted and foraged in spruce fir forest), they also selected cool roosting sites during hot weather periods, when they often exhibited signs of heat stress [57]. Our projections of only 4 future habitat area patches exceeding the maximum home range size documented in Idaho (and only 8 and 26 greater in size than the observed average and minimum home range, respectively) raise the possibility that under future conditions in New Mexico, the boreal owl will become extirpated from the state. To persist in New Mexico, any remaining boreal owls would likely need to maintain smaller home ranges or increase their relative use of lower-elevation vegetation despite an increased likelihood of heat stress. Under one possible scenario, northern New Mexico would function as sink habitat. Some patches of spruce-fir forest would continue to be occupied, but only with an influx of immigrants from Colorado’s high mountainous terrain to the north.
Warming trends and increased aridity are not only likely to reduce the amount of owl habitat in the region but also favor population shifts northward and upward in elevation within the region. Indeed, model outputs for boreal, western screech, and whiskered screech-owls (figures 4a-h) corroborate a theorized northward shift in habitat conditions for some species. Outputs for boreal, flammulated, long-eared, and northern pygmy-owls support expectations that habitat will shift upward in elevation. Any shift north in habitat for the whiskered screech-owl may represent a range expansion, though the species will nonetheless likely experience a net loss of habitat with ongoing climate trends (Table 4).
Model uncertainty
There is always uncertainty in model outputs as with results reported here on habitat distribution and decline. The model validation showed that the habitat profile tested was spatially inclusive of all survey data when a buffer was applied, and that model parameterization may reflect a reasonable balance of omission and commission error. Nevertheless, greater confidence may be warranted for results for percent change in habitat conditions (current to future) versus specific area estimates in Table 4. Uncertainty in the underlying climate vulnerability projections tends to be greater in mountainous terrain and lower in basins and plains of the region [15] with the implication that habitat models associated with upper life zones may have added uncertainty – i.e., for boreal, northern saw-whet, and flammulated owls. Although not included in our modeling, nest locations for the long-eared, western screech, and great horned owls can also occur in low elevation riparian areas separate from suitable upland forest and woodland habitats. The ability to include riparian habitats in future habitat models could improve the predictive ability and usefulness of model outputs. Riparian and canyon habitats represented in some survey records for the western screech-owl, habitat features not expressed in its model profile, helps to explain lower validation results for this species. These valley bottom settings usually possess upland vegetation including that of cool-moister life zones of higher elevations, given how riparian and canyon bottoms are protected from sun exposure and how they concentrate moisture. In addition to the great horned owl, the western screech-owl is known to breed in low-elevation areas such as the Sonoran Desert in Arizona [27,28]. Ongoing trends and 21st-century forecasts for temperature and aridity in the region [58,59] suggest that desert regions may increase in extent [60].
Vegetation vs. resource habitat models
Resource-based habitat models have been shown to perform better than vegetation-based habitat models for some species [61,62]. Although our spatial model functioned well for most of the owl species included in this study, they likely did not fully capture spatial occurrence, or lack thereof, when scaled over space and time. The northern saw-whet owl tends to occur sporadically throughout its distribution, being locally common in some years and absent in others, perhaps tracking irruptive populations of small mammals, an important, temporal habitat component [63]. High prey densities are also believed to be key for the long-eared owl, often referred to as a nomadic breeder [64]. Nest cavities may represent another important limiting resource in the flammulated owl, western and whiskered screech owls, northern pygmy-owl, boreal owl, and northern saw-whet owl, all secondary cavity nesters. In New Mexico, for example, areas with suitable nest cavities were found to be saturated with flammulated owl territories [30]. Cavities large enough for flammulated owls in New Mexico are those excavated by the acorn woodpecker (Melanerpes formicivorus) and the northern flicker (Colaptes auratus), but the distribution of the latter species is largely limited by the availability of soft wood in the form of ponderosa pine snags and large quaking aspens [30,65].
Fire
There is increasing evidence of shrinking forest-woodland extent in the West and shifting vegetation types [32,66] with upward changes in elevation at an average rate of 15m or more per decade [33,36,37]. Concerns about shifts in vegetation types and related habitat conditions should be heightened by the potential for abrupt and lasting change with severe wildfire [67]. Of particular concern are the combined effects of warming, drought, and wildfire on forests and woodlands [68,69] and compounding effects of fire exclusion and unnaturally high tree densities in fire-adapted forests and woodlands [70]. From 2011 to 2013, two wildfires in the Jemez Mountains (one of the three mountain ranges harboring the totality of New Mexico’s boreal owl population) totaling > 700 km2 reduced the extent of spruce–fir forest around the southern and eastern portions of the Valles Caldera National Preserve by 34 km2 [45]. During those same three years, three fires in the southern Sangre de Cristo Mountains burned through 54 km2 of spruce–fir forest. In 2013, one of these fires burned through one of the boreal owl’s historical locations [45]. The 2022 Calf Canyon/Hermits Peak Fire became the largest fire in the state of New Mexico’s history after burning through 135,934 hectares in the Sangre de Cristo Mountains, of which 29,604 hectares (22%) corresponded to spruce-fir forest. As no boreal owl surveys were ever conducted in spruce-fir forest stands affected by the 2022 Calf Canyon/Hermits Peak Fire (D. W. Stahlecker, unpubl. data), we cannot rule out the loss of more boreal owl historical habitat estimated at a maximum of only 306,000 ha in New Mexico (Table 4). Another species that may stand to lose from an increased incidence of severe fires and lack of forest regeneration is the northern saw-whet owl, which showed near complete avoidance of burned areas in the Pacific Northwest [71].
Adaptation
Some mechanisms for coping with climate change have already been documented in owls and other bird species. Research conducted in Finland used 28 years of past climate data to also link milder winter conditions with the disappearance of selective predation pressure against the tawny owl’s brown morph (determined largely through genetic inheritance and considered generally more cryptic when there is less snow), an example of adaptive capacity through microevolution [72]. The adaptive capacity of bird populations to warming trends is also revealed by uphill movement to cooler-moisture and more familiar climate conditions, with short-lived species holding an advantage in relocating more quickly [73]. If this evidence applies to owls of the Southwest, the northern saw-whet, flammulated, and northern pygmy-owls may possess some advantage in coping with rapid warming trends relative to longer-lived species such as the long-eared owl. Throughout much of New Mexico, however, flammulated owl populations breeding in ponderosa pine and/or mixed conifer forest may already find themselves on the highest slopes within the same mountain range (e.g., Zuni Mountains). At the same time, flammulated owls likely have good potential for adaptability because they are long-distance migrants with low natal philopatry making them able to colonize isolated habitats [74].
Though a long-lived species and projected by us to lose 55% of its mapped upper elevation habitat (due to conversion to non-woodland types), the great horned owl may nonetheless be uniquely suited to adapt to projected future climate change. It is known to occupy, and breed in, a very broad range of vegetation types from desert to subalpine or boreal forest [75]. It exhibits much size and other phenotypic variation [76], can switch between being a prey specialist and a generalist [77], and is capable of long-distance movements following habitat quality reductions [78]. Though much remains to be learned about natal dispersal in the western and whiskered screech-owl, these two species may be characterized by lower adaptive capacity as adults typically remain within the same breeding territories year-round in the southwestern U.S. Our study shows that gains in habitat will likely not be enough to offset habitat losses, not to mention that these gains will also not happen in areas neighboring existing habitat. Western and whiskered screech-owl may still be able to shift their distribution through natal dispersal, which remains poorly known. In southwestern Idaho, juvenile western screech-owls were found to disperse an average of 10.6 ± 1.8 km from natal to overwintering sites, with possible further dispersal later to first breeding sites [79].
Mitigation
The directional change in elevation and latitude that is anticipated for wildlife species and their habitat can help set priorities for land management aimed at climate adaptation. Repeat inventory of historical vegetation samples in the Southwest corroborate expectations for rapid warming trends and habitat responses [33,36,37]. Wildlife specialists and natural resource managers can evaluate the likely shifts among vegetation types under increased warming and aridity, assess the sensitivity and adaptive capacity of plant dominants that make up habitat structure, and evaluate ongoing owl population trends. Legacy mapping of vegetation and wildlife habitat should not be viewed as static and will challenge practitioners and applied scientists to adapt conventional roles for the interpretation, communication, and guidance to others on the use of spatial information.
Broader-scale implications
Most of the World’s owl species now appear to be experiencing global population declines [80]. High exposure and high sensitivity to ongoing and future climate change have been shown in high-latitude owls in particular [6,8,9,81]. Our research points to what may represent important loss of habitat not just in the southwestern U.S. but perhaps also in many mountainous regions of arid southwestern North America and arid zones of the Subtropics. With climate having a powerful influence on the distribution of biota around the world [82,83], tropical mountain regions have long been known as biodiversity hotspots [84], but there is also increasing recognition of the ecological distinctiveness of subtropical, arid-zone mountain ranges [85,86,87]. Unlike at more temperate latitudes, arid-zone mountains tend to harbor biota distinct from neighboring lowlands, reflecting their role as refugia during past periods of warming temperatures and drying conditions [85,86]. These refugia are now threatened by anthropogenic climate change, with the taxa they harbor facing range contractions [87] and increased isolation [88]. Added to projected loss of refugia habitat in arid-zone subtropical regions are findings linking likely future population declines and more extreme temperature and/or reduced precipitation, not just for the Mexican spotted owl in southwestern North America [7] but also for the tawny owl in Israel [89].
5. Conclusions
We project severe, region-wide breeding habitat loss for most montane owls in the southwestern U.S. by 2090. Near complete loss of current breeding habitat is projected in particular for known flammulated owl populations in western New Mexico, while the projected boreal owl habitat distribution will only consist of isolated areas perhaps too small to sustain the regional persistence of the species. Three more species, the northern pygmy-owl, long-eared owl, and northern saw-whet owl, are projected to lose at least 60% of their current breeding habitat. The whiskered screech-owl is predicted to lose all its current habitat and will only persist in the region if it can track areas that become suitable to the north of its current range. More uncertainty exists for two species with distributions that extend down to lower elevations.
Author Contributions
Conceptualization, J.E.C., F.J.T.; methodology, J.E.C., F.J.T.; validation, J.E.C., F.J.T., K.C.C.; formal analysis, J.E.C., F.J.T.; investigation, J.E.C., F.J.T., D.W.S., D.P.A., J.L.G., C.D.H., H.K.T., M.C.C.; resources, J.E.C., F.J.T., H.K.T., M.C.C.; data curation, J.E.C., F.J.T., D.W.S., D.P.A., J.L.G., C.D.H., H.K.T., M.C.C., K.C.C.; writing—original draft preparation, J.E.C., F.J.T.; writing—review and editing, J.E.C., F.J.T., D.W.S., D.P.A., J.L.G., K.C.C.; visualization, J.E.C., F.J.T.; supervision, J.E.C.; project administration, J.E.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Museum specimen records from New Mexico and Arizona can be publicly queried at https://arctos.database.museum/search.cfm; i-Naturalist records are publicly available at https://www.inaturalist.org/observations. Our validation dataset is available from the authors upon request.
Acknowledgments
The authors wish to acknowledge the foundational work of Esteban Muldavin (Natural Heritage New Mexico and University of New Mexico Department of Biology) and Max Wahlberg (USDA Forest Service, Pacific Northwest Region), who contributed to the development of the ecosystem vulnerability assessment underlying the present study. The lead author also benefitted from discussions with Andrew Johnson (Museum of Southwestern Biology) on the use of museum records in our validation dataset.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Stanton, J.C., Shoemaker, K.T., Pearson, R.G., Akçakaya, H.R. Warning times for species extinctions due to climate change. Global Change Biol. 2015, 21, 1066–1077. [CrossRef]
- Williams, S.E., Shoo, L.P., Isaac, J.L., Hoffmann, A.A., Langham, G. Towards an Integrated Framework for Assessing the Vulnerability of Species to Climate Change. PLOS Biol. 2008, 6, 2621–2626. [CrossRef]
- Dawson, T.P., Jackson, S.T., House, J.I., Prentice, I.C., Mace, G.M. Beyond Predictions: Biodiversity Conservation in a Changing Climate. Science 2011, 332, 53–58. [CrossRef]
- Glick, P., Stein, B.A., Edelson, N. (eds.). Scanning the conservation horizon: a guide to climate change vulnerability assessment. National Wildlife Federation: Washington, D.C., 2011. Available from: www.nwf.org/vulnerabilityguide.conservation.
- Millon, A., Petty, S.J., Little, B., Gimenez, O., Cornulier, T., Lambin, X. Dampening prey cycle overrides the impact of climate change on predator population dynamics: a long-term demographic study on tawny owls. Global Change Biol. 2014, 20, 1770–1781. [CrossRef]
- Peery, M.Z., Guttiérez, R.J., Kirby, R., Ledee, O.E., Layahe, W. Climate change and spotted owls: potentially contrasting responses in the Southwestern United States. Global Change Biol. 2011. [CrossRef]
- Gilg, O., Sittler, B., Hanski, I. Climate change and cyclic predator–prey population dynamics in the high Arctic. Global Change Biol. 2009, 15, 2634–2652. [CrossRef]
- Schmidt, N.M., Ims, R.A., Høye, T.T., Gilg, O., Hansen, L.H., Hansen, J., Lund, M., Fuglei, E., Forchhammer, M.C., Sittler, B. Response of an arctic predator guild to collapsing lemming cycles. Proc Biol Sci. 2012, 279, 4417–22. [CrossRef]
- Jenouvrier, S. Impacts of climate change on avian populations. Global Change Biol. 2013. [Google Scholar] [CrossRef]
- Altwegg R., Roulin, A., Kestenholz, M., Jenni, L. Demographic effects of extreme winter weather in the barn owl. Oecologia 2006, 149, 44–51. [CrossRef] [PubMed]
- Ganey, J.L., Balda, R.P., King, R.M. 1993. Metabolic rate and evaporative water loss of Mexican spotted and great horned owls. Wilson Bull. 1993, 105, 645–656.
- Weathers, W.W., Hodum, P.J., Blakesley, J.A. Thermal ecology and ecological energetics of California spotted owls. Condor 2001, 103, 678–690. [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change). Climate change 2007: the physical science basis. In S. Solomon, editor. Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press: Cambridge, UK, 2007.
- Triepke, F.J., Muldavin, E.H., Wahlberg, M.M. Using climate projections to assess ecosystem vulnerability at scales relevant to managers. Ecosphere 2019, 10, e02854. [CrossRef]
- Dick-Peddie, W.A. New Mexico Vegetation Past, Present and Future; University of New Mexico Press: Albuquerque, New Mexico, USA, 1993. [Google Scholar]
- Allen, C.D., Betancourt, J.L., Swetnam, T.W. 1998. Landscape changes in the southwestern United States: techniques, long-term data sets, and trends. In Perspectives on the land use history of North America: a context for understanding our changing environment; Sisk, T., Ed. U.S. Geological Survey, Biological Science Report USGS/BRD/BSR-1998-0003; pp. 71–84.
- Triepke, J.F., Lowrey, T.K. 2023. Vegetation, fire ecology, and climate of New Mexico’s ecosystems. In Wild Carnivores of New Mexico; Cartron, J.-L.E., Frey, J.K., Eds.; University of New Mexico Press: Albuquerque, USA, 2023; pp. 21–49.
- Cartron, J.-L.E., Triepke, F.J., Gutzler, D.S., Steckbeck, K.M., Calhoun, K.C. Future climate-driven impacts and the conservation of carnivores in New Mexico. In Wild Carnivores of New Mexico; Cartron, J.-L.E., Frey, J.K., Eds.; University of New Mexico Press: Albuquerque, USA, 2023; pp.91–127.
- Moreland, J.C., Robbie, W.A., Triepke, F.J., Muldavin, E.H., Malusa, J.R. Ecological Response Units: ecosystem mapping system for the Southwest US. In Proceedings of the sixth natural history of the Gila symposium, 25-27 February 2016, Silver City, New Mexico, USA, 25-27 February 2016.
- Box, E.O., Fujiwara, K. 2005. Vegetation types and their broad–scale distribution. In Vegetation Ecology; van der Maarel, E., Ed.; Blackwell Publishing: Malden, Massachussets, USA, 2005; pp. 106–128.
- MacArthur, R., MacArthur, J.W. On Bird Species Diversity. Ecology 1961, 42, 594–598. [CrossRef]
- Cody, M.L. Habitat Selection in Birds: The Roles of Vegetation Structure, Competitors, and Productivity. BioScience 1981, 31, 107–113. [CrossRef]
- Besnard, A.G., Davranche, A., Maugenest, S., Bouzillé, J.B., Vian, A., Secondi, J. (2015). Vegetation maps based on remote sensing are informative predictors of habitat selection of grassland birds across a wetness gradient. Ecological Indicators 2015, 58, 47–54. [CrossRef]
- Timm, B.C., McGarigal, K., Cushman, S.A., Ganey, J.L. Multi-scale Mexican spotted owl (Strix occidentalis lucida) nest/roost habitat selection in Arizona and a comparison with single-scale modeling results. Landscape Ecol 2016, 31, 1209–1225. [CrossRef]
- Wan, H.Y., McGarigal, K., Ganey, J.L., Lauret, V., Timm, B.C., Cushman, S.A. Meta-replication reveals nonstationarity in multi-scale habitat selection of Mexican Spotted Owl. Condor 2017, 119, 641–658. [CrossRef]
- Corman, T. E., Wise-Gervais, C. Eds. Arizona Breeding Bird Atlas; University of New Mexico Press: Albuquerque, New Mexico, USA, 2005.
- Cartron, J.-L.E. Ed. 2010. Raptors of New Mexico; University of New Mexico Press: Albuquerque, New Mexico, USA, 2010.
- Wise-Gervais, C. Flammulated owl. In Arizona Breeding Bird Atlas; Corman, T.E., Wise-Gervais, C. Eds.; University of New Mexico Press: Albuquerque, New Mexico, USA, pp. 208–209.
- Arsenault, D.P. Distribution and density of flammulated owls in western New Mexico. New Mexico Ornithol. Bull. 2007, 35, 41–53. [Google Scholar]
- Cleland, D.T., Freeouf, J.A., Nowacki, G.J., Carpenter, C., Keys, J.E., McNab, W.H. Ecological subregions: Sections and subsections of the conterminous United States. USDA Forest Service General Technical Report WO-76; Washington Office, Washington, DC, USA, 2007. https://databasin.org/datasets/.
- Bell, D.M., Bradford, J.B., Lauenroth, W.K. Early indicators of change: Divergent climate envelopes between tree life stages imply range shifts in the western United States. Glob. Ecol. Biogeogr. 2014, 23, 168–180. [CrossRef]
- Kelly, A.E., Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proceedings of the National Academy of Sciences 2008, 105, 11823–11826. [CrossRef]
- Monleon, V.J., Lintz, H.E. Evidence of tree species’ range shifts in a complex landscape. PLoS ONE 2015, 10, e0118069. [CrossRef]
- Hill, A.P., Field, C.B. Forest fires and climate-induced tree range shifts in the western US. Nat. Commun. 2021, 12, 1–10. [CrossRef]
- Brusca, R.C., Wiens, J.F., Meyer, W.M., Eble, J., Franklin, K., Overpeck, J.T., Moore, W. Dramatic response to climate change in the Southwest: Robert Whittaker’s 1963 Arizona Mountain plant transect revisited. Ecol. Evol. 2013, 3, 3307–3319. [CrossRef] [PubMed]
- Guida, R.J., Abella, S.R., Smith, W.J., Stephen, H., Roberts, C.L. Climatic change and desert vegetation distribution: Assessing thirty years of change in southern Nevada’s Mojave Desert. Prof. Geogr. 2014, 66, 311–322. [CrossRef]
- Vogl, R.J. Chaparral succession. In Conrad, C.E., Oechel, W.C., Eds., Proceedings of the symposium on dynamics and management of Mediterranean-type ecosystems. USDA Forest Service Gen. Tech. Rep. PSW-GTR-58; Pacific Southwest Forest and Range Experiment Station, Berkeley, CA, USA, 1982. Pp. 81–85.
- Kitchen, S.G., Meyer, S.E., Carlson, S.L. 2015. Mechanisms for maintenance of dominance in a nonclonal desert shrub. Ecosphere 2015, 6, 1–15. [CrossRef]
- Pockman, W.T., Speery, J.S. Vulnerability to xylem cavitation and the distribution of Sonoran Desert vegetation. Am. J. Bot. 2000, 87, 1287–1299. [CrossRef]
- Enright, N., Miller, B. Livestock grazing impacts on desert vegetation, Khirthar National Park, Pakistan. Rangel. Ecol. Manag. 2007, 60, 680–684. [CrossRef]
- Bhattachan, A., D'Odorico, P., Dintwe, K., Okin, G.S., Collins, S.L. Resilience and recovery potential of duneland vegetation in the southern Kalahari. Ecosphere 2014, 5, 2. [CrossRef]
- Brotons, L., Thuiller, W., Arauijo, M. B. and Hirzel, A. H. Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 2004, 27, 437–44. [CrossRef]
- Stahlecker, D.W., Duncan, R.B. 1996. The boreal owl at the southern terminus of the Rocky Mountains: Undocumented longtime resident or recent arrival? Condor 1996, 98, 153–160. [CrossRef]
- Stahlecker, D.W., MacKerrow, E.P., Walker, H.A., Batkin, J.P., Foy, B.R. Persistence of the boreal owl in New Mexico: 1987-2012. Western Birds 2014, 45, 166–175.
- Ganey, J.L., Benoit, M.A. Using terrestrial ecosystem survey data to identify potential habitat for the Mexican spotted owl on National Forest System lands: a pilot study. Gen. Tech. Rep. RMRS-GTR-86, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO, 2002.
- Bosakowski, T., Smith, D.G. Response of a forest raptor community to broadcasts of heterospecific and conspecific calls during the breeding season. Can. Field-Nat. 1998, 112, 198–203. [CrossRef]
- Lima, K.A., Call, E.M., Hodgman, T.P., Potter, D.S., Gallo, S., Blomberg, E.J. Environmental conditions and call-broadcast influence detection of eastern forest owls during standardized surveys. Condor 2020, 122, duaa016. [CrossRef]
- Groce, J.E. Groce, J.E., Morrison, M.L. Habitat Use by Saw-Whet Owls in the Sierra Nevada. J. Wildl. Manag. 2010, 74, 1523–1532. [Google Scholar] [CrossRef]
- Ganey, J.L., White, G.C., Ward, J.P., Jr., Kyle, S.C., Apprill, D.L., Rawlison, T.A., Jonnes, R.S. Demography of Mexican spotted owls in the Sacramento Mountains, New Mexico. J. Wildl. Manag. 2014, 78, 42–49. [CrossRef]
- Hathcock, C.D. Use of nest boxes, reproductive success, and sex ratio in two screech-owls in the Peloncillo Mountains of southeastern Arizona. Wilson J. Ornith. 2023, 135, 283–287. [Google Scholar] [CrossRef]
- Fielding, A.H. What are appropriate characteristics of an accuracy measure? In Predicting Species Occurrences: Issues of Accuracy and Scale; Scott, J.M., Heglund, P.J., Morrison, M.L., Haufler, J.B., Raphael, M.G., Wall, W.A., Samson, F.B., Eds.; Island Press: Washington, USA, 2002; pp. 271–280. [Google Scholar]
- Ottaviani, D., Lasinio, G.J., Boitani, L. Two statistical methods to validate habitat suitability models using presence-only data. Ecol. Model. 2004, 179, 417–443. [CrossRef]
- Pembury Smith, M.Q.R., Ruxton, G.D. Effective use of the McNemar test. Behav Ecol Sociobiol 2020, 74, 133. [CrossRef]
- Keyser, A.R., Krofcheck, D.J., Remy, C.C., Allen, C.D., and Hurteau, M.D. Simulated increases in fire activity reinforce shrub conversion in a southwestern US forest. Ecosystems 2020, 23, 1702–1713. [CrossRef]
- Merker, S.A., Chandler, R.B. Identifying global hotspots of avian trailing-edge population diversity. Global Ecology and Conservation 2020, 22, e00915. [CrossRef]
- Hayward, G.D., Hayward, P.H., Garton, E.O. Ecology of Boreal Owls in the Northern Rocky Mountains, USA. Wildl. Monogr. 1993, 124.
- Seager, R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G., Huang, H.-P., Harnik, N., Leetmaa, A., Lau, N.-C., Li, C., Velez, J., Naik, N. Model projections of an imminent transition to a more arid climate in southwestern North America. Science 2007, 316, 1181–4. [CrossRef]
- Gutzler, D.S., Robbins, T.O. Climate variability and projected change in the western United States: Regional downscaling and drought statistics. Climate Dyn. 2010, 37, 835–849. [CrossRef]
- Rehfeldt, G.E., Crookston, N.L., Sáenz-Romero, C., Campbell, E.M. North American vegetation model for land-use planning in a changing climate: A solution to large classification problems. Ecological Applications 2012, 22, 119–141. [CrossRef]
- Dennis, R.L.H., Shreeve, T.G., Van Dyck, H. Habitats and Resources: The Need for a Resource-based Definition to Conserve Butterflies. Biodivers Conserv 2006, 15, 1943–1966. [CrossRef]
- Turlure, C., Schtickzelle N., Dubois Q., Baguette M., Dennis R.L.H., Van Dyck, H. Suitability and transferability of the resource-based habitat concept: a test with an assemblage of butterflies. Front. Ecol. Evol. 2019, 7, 127. [CrossRef]
- Wise-Gervais, C. Northern saw-whet owl. In Arizona Breeding Bird Atlas; Corman, T.E., Wise-Gervais, C. Eds.; University of New Mexico Press: Albuquerque, New Mexico, USA, 2005, pp. 228–229.
- Friederici, P. Long-eared owl. In Arizona Breeding Bird Atlas; Corman, T.E., Wise-Gervais, C. Eds.; University of New Mexico Press: Albuquerque, New Mexico, USA, 2005, pp. 226–227.
- Arsenault, D.P. Differentiating nest sites of primary and secondary cavity-nesting birds in New Mexico," Journal of Field Ornithology 2004, 75, 257–265, (1 July 2004). [CrossRef]
- Hill, A.P., Field, C.B. Forest fires and climate-induced tree range shifts in the western US. Nat. Commun. 2021, 12, 1–10. [CrossRef]
- Guiterman, C.H., Gregg, R.M., Marshall, L.A., Beckmann, J.J., van Mantgem, P.J., Falk,D.A., Keeley, J.E., Caprio, A.C., Coop, J.D., Fornwalt, P.J., Haffey, C. Vegetation type conversion in the US Southwest: frontline observations and management responses. Fire Ecol. 2022, 18, 1–16. [CrossRef]
- Breshears, D.D., Myers, O.B. Myers, Meyer, C.W., Barnes, F.J., Zou, C.B., Allen, C.D., McDowell, M.G., Pockman, W.T. Tree die-off in response to global change-type drought: mortality insights from a decade of plant water potential measurements. Front. Ecol. Environ. 2009, 7, 185–189. [CrossRef]
- Allen, C.D., Macalady, A.K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Kitzberger, T., Rigling, A., Breshears, D.D., Hogg, E.T., Gonzalez, P. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Manage 2010, 259, 660–684. [CrossRef]
- Westerling, A.L., Hidalgo, H.G., Cayan, D.R., Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [CrossRef] [PubMed]
- Duchac, L.S., Lesmeister, D.B., Dugger, K.M., Davis, R.J. Differential landscape use by forest owls two years after a mixed-severity wildfire. Ecosphere 2021, 12, e03770. [CrossRef]
- Karell, P., Ahola, K., Karstinen, T., Valkama, J., Brommer, J.E. Climate change drives microevolution in a wild bird. Nature Commun 2011, 2, 208. [CrossRef]
- Couet, J., Marjakangas, E., Santangeli, A., Kålås, J.A., Lindström, Å., Lehikoinen, A. Short-lived species move uphill faster under climate change. Oecologia 2022, 198, 877–888. [CrossRef]
- Arsenault, D.P., Stacey, P.B., Hoelzer, G.A. Mark–recapture and DNA fingerprinting data reveal high breeding-site fidelity, low natal philopatry, and low levels of genetic population differentiation in flammulatedowls (Otus Flammeolus). Auk 2005, 122, 329–337. [CrossRef]
- Burton, J. A. Owls of the World; William Collins: Glasgow; 1984.
- Ostrow, E.N., DeCicco, L.H., Moyle, R.G. Range-wide phylogenomics of the Great Horned Owl (Bubo virginianus) reveals deep north-south divergence in northern Peru. PeerJ 2023, 11, e15787. [CrossRef]
- Tyson, R., Lutscher, F. Seasonally Varying predation behavior and climate shifts are predicted to affect predator-prey cycles. Am. Nat. 2016, 188, 539–553. [CrossRef]
- Adamcik, R.S. Keith, L.B. Regional movements and mortality of great horned owls in relation to snowshoe hare fluctuations. Can. Field-Nat. 1978, 92, 228–234. [CrossRef]
- Ellsworth, E., Belthoff, J.R. Sex-biased dispersal of young Western Screech-owls (Otus kennicottii) in southwestern Idaho. In: Duncan, J.R.; Johnson, D.H.; Nicholls, T.H., eds. Biology and conservation of owls of the Northern Hemisphere: 2nd International symposium. Gen. Tech. Rep. NC-190. St. Paul, MN: U.S. Dept. of Agriculture, Forest Service, North Central Forest Experiment Station. 1997, 155–159.
- IUCN. 2022. The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org. Accessed on 3 November 2023.
- Orlando, G., Passarotto, A., Morosinotto, C., Ahola, K., Karstinen, T., Brommer, J.E., Koskenpato, K., Karell, P. Changes in over-winter prey availability, rather than winter climate, are associated with a long-term decline in a northern Tawny Owl population. J Ornithol, 2023. [CrossRef]
- Woodward, F.I. Climate and plant distribution. Cambridge University Press, Cambridge, United Kingdom. 1987, 174 pages.
- Cox, C.B., Moore, P.D., Ladle, R.J. Biogeography: An ecological and evolutionary approach. Wiley-Blackwell, Hoboken, NJ, USA. 2016, Ninth Edition, 512 pages.
- Sonne, J., Dalsgaard, B., Borregaard, M.K., Kennedy, J., Fjeldså, J., Rahbek, C. Biodiversity cradles and museums segregating within hotspots of endemism. Proc. R. Soc. B 2022, 289, 20221102. [CrossRef]
- Byrne, M., Yeates, D.K., Joseph, L., Kearney, M., Bowler, J., Williams, M.A.J., Cooper, S., Donnellan, S.C., Keogh, J.S., Leys, R., Melville, J., Murphy, D.J., Porch, N., Wyroll, K.-H. Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota. Molecular Ecology 2008, 17, 4398–4417. [CrossRef]
- Rahbek, C., Borregaard, M.K., Colwell, R.K., Dalsgaard, B., Holt, B.G., Morueta-Holme, N., Nogues-Bravo, D., Whittaker, R.J., Fjeldså, J. Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science 2019, 365(6458), 1108–1113. [CrossRef]
- McDonald, P.J. McDonald, P.J., Jobson, P., Köhler, F., Nano, C.E.M., Oliver, P.M. The living heart: climate gradients predict desert mountain endemism. Ecol. Evol. 2021, 11, 4366–4378. [Google Scholar] [CrossRef] [PubMed]
- Haire, S.L. Haire, S.L., Villarreal, M.L., Cortés-Montaño, C., Flesch, A.D., Iniguez, J.M., Romo-Leon, J.R., Sanderlin, J.S. Climate refugia for Pinus spp. in topographic and bioclimatic environments of the Madrean sky islands of México and the United States. Plant Ecol. 2022, 223, 577–598. [Google Scholar] [CrossRef]
- Comay, O., Ezov, E., Yom-Tov, Y., Dayan, T. In its southern edge of distribution, the tawny owl (Strix aluco) is more sensitive to extreme temperatures than to rural development. Animals 2022, 12, 641. [CrossRef]
Figure 1.
Elevational zonation of vegetation types in the southwestern U.S. showing desert and grassland at the lower elevations and woodlands and forests at mid to upper elevations.
Figure 1.
Elevational zonation of vegetation types in the southwestern U.S. showing desert and grassland at the lower elevations and woodlands and forests at mid to upper elevations.
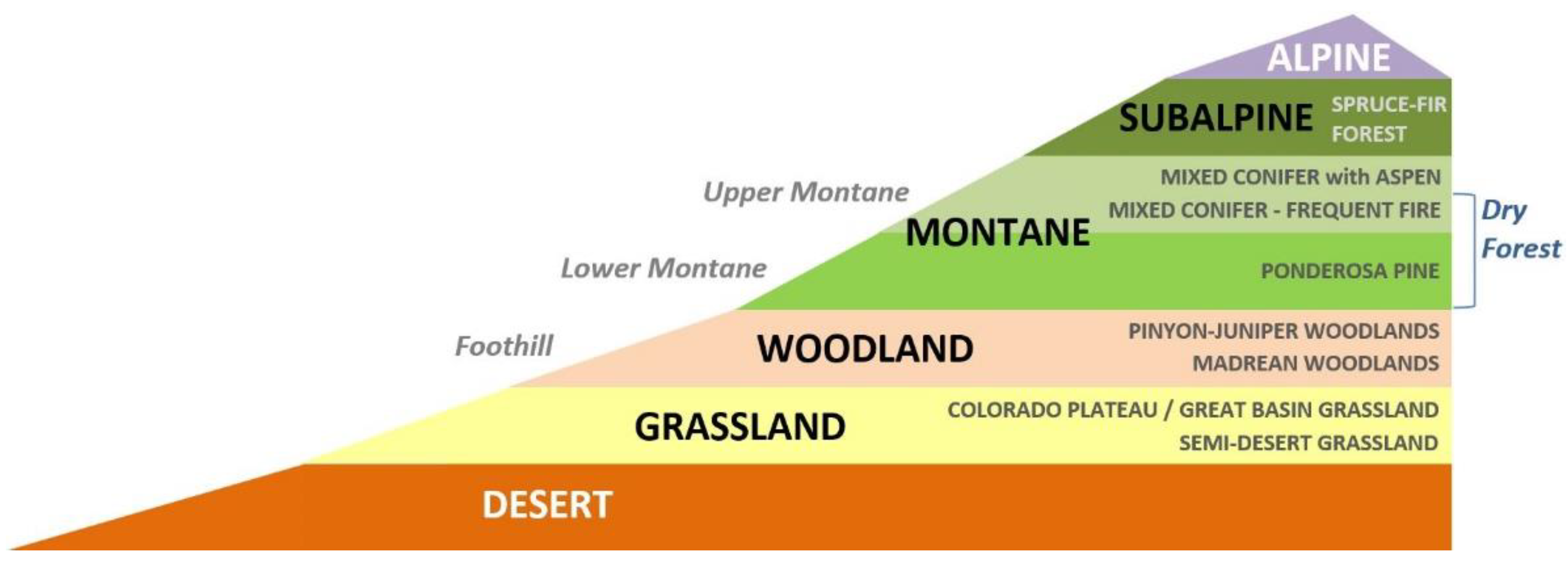
Figure 2.
Model zones used for owl habitat profiles and alignment with reported range distributions.
Figure 2.
Model zones used for owl habitat profiles and alignment with reported range distributions.
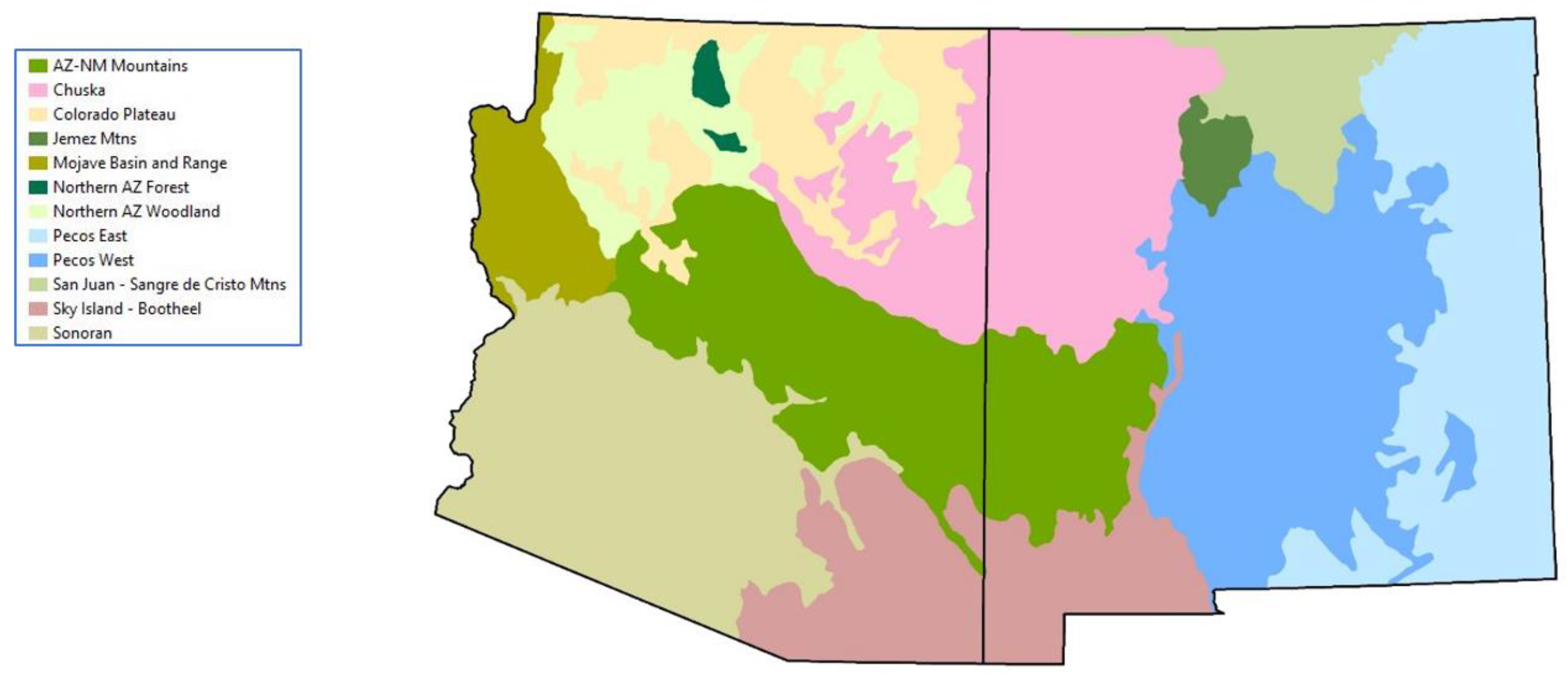
Figure 3.
Model validation results and difference in the proportion of records within vs. outside mapped habitat under current and projected conditions.
Figure 3.
Model validation results and difference in the proportion of records within vs. outside mapped habitat under current and projected conditions.
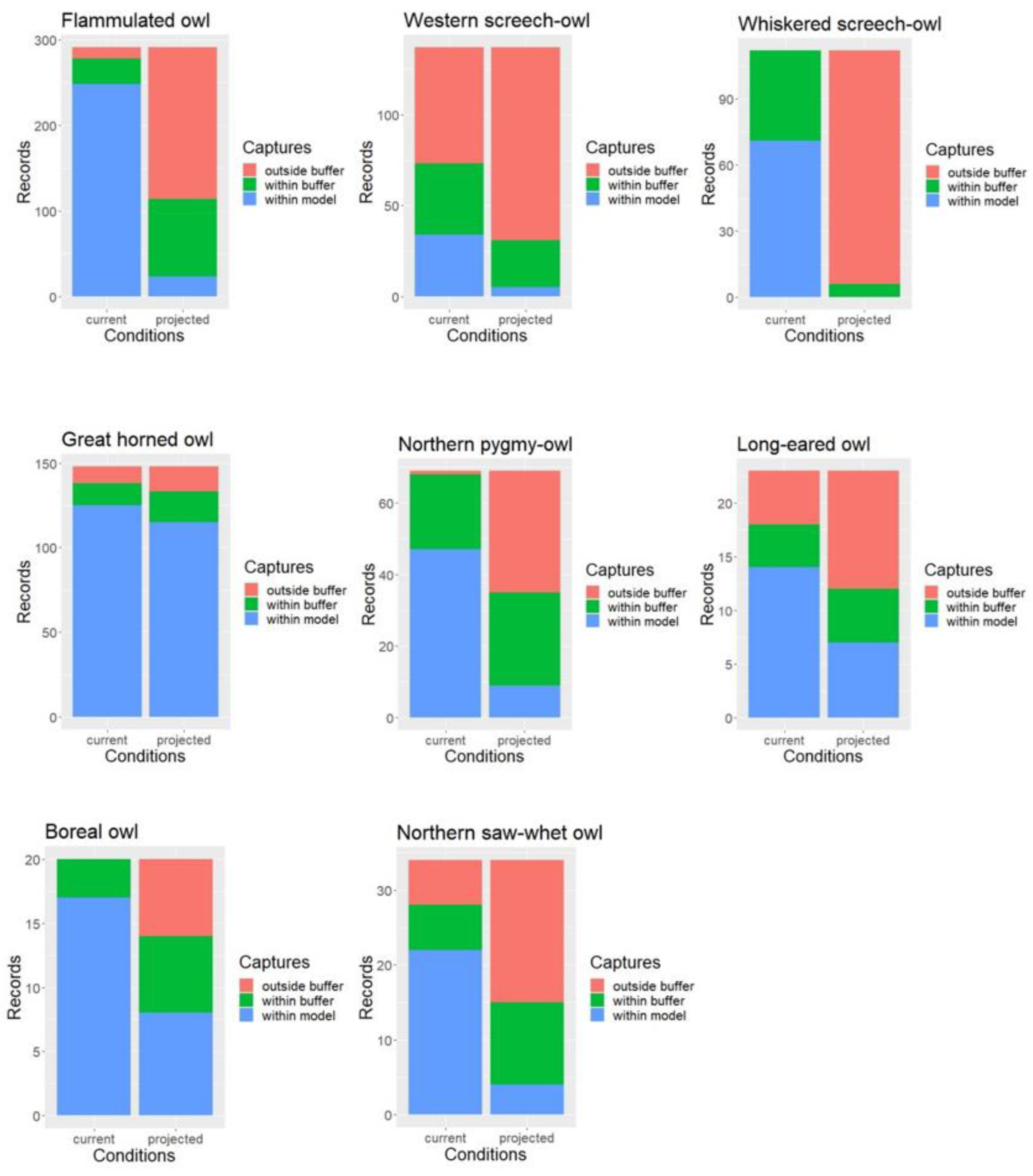
Figure 4.
Distribution of modeled habitat for eight owl species under current vs. 2090 projected conditions, southwestern U.S. .
Figure 4.
Distribution of modeled habitat for eight owl species under current vs. 2090 projected conditions, southwestern U.S. .
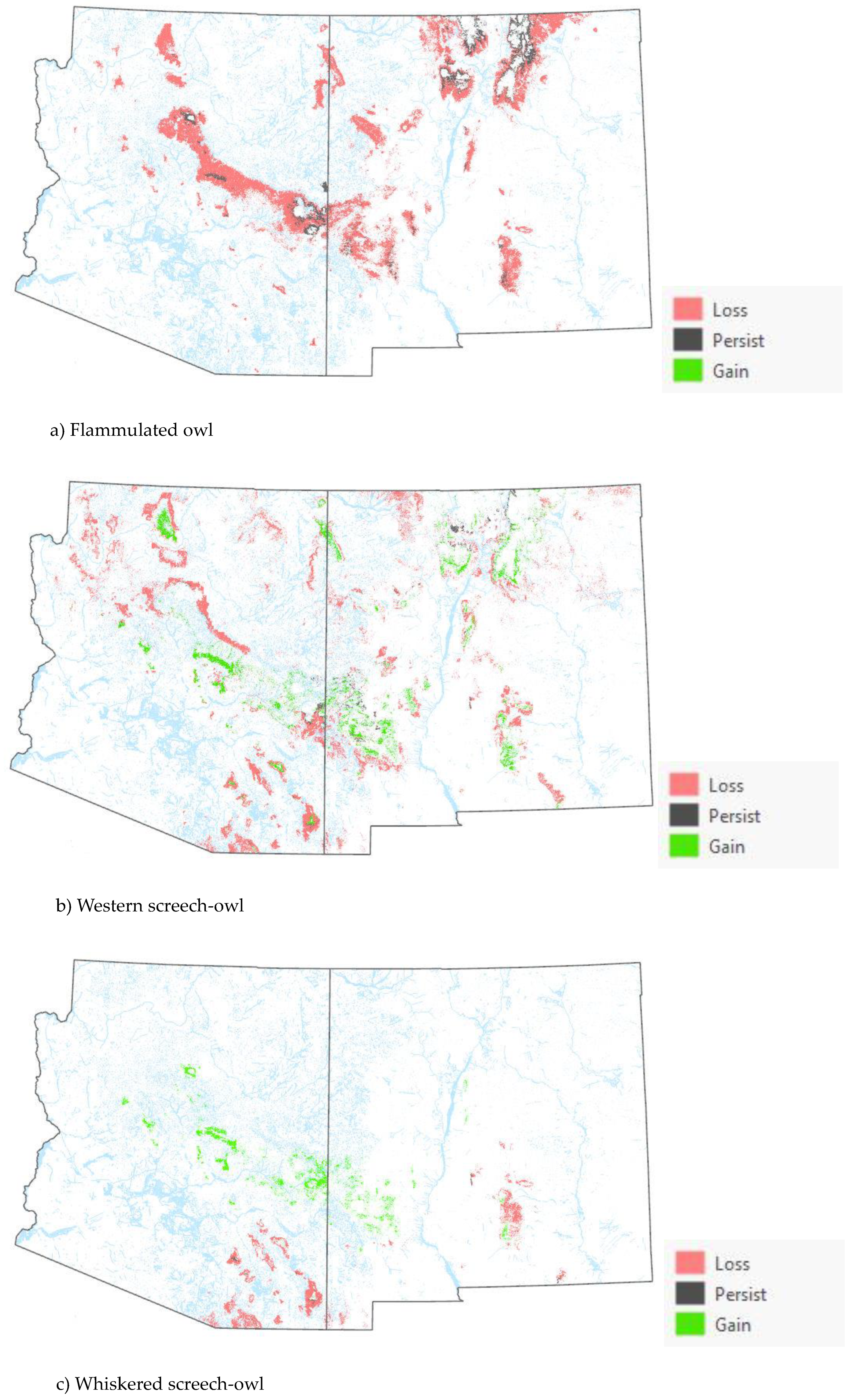
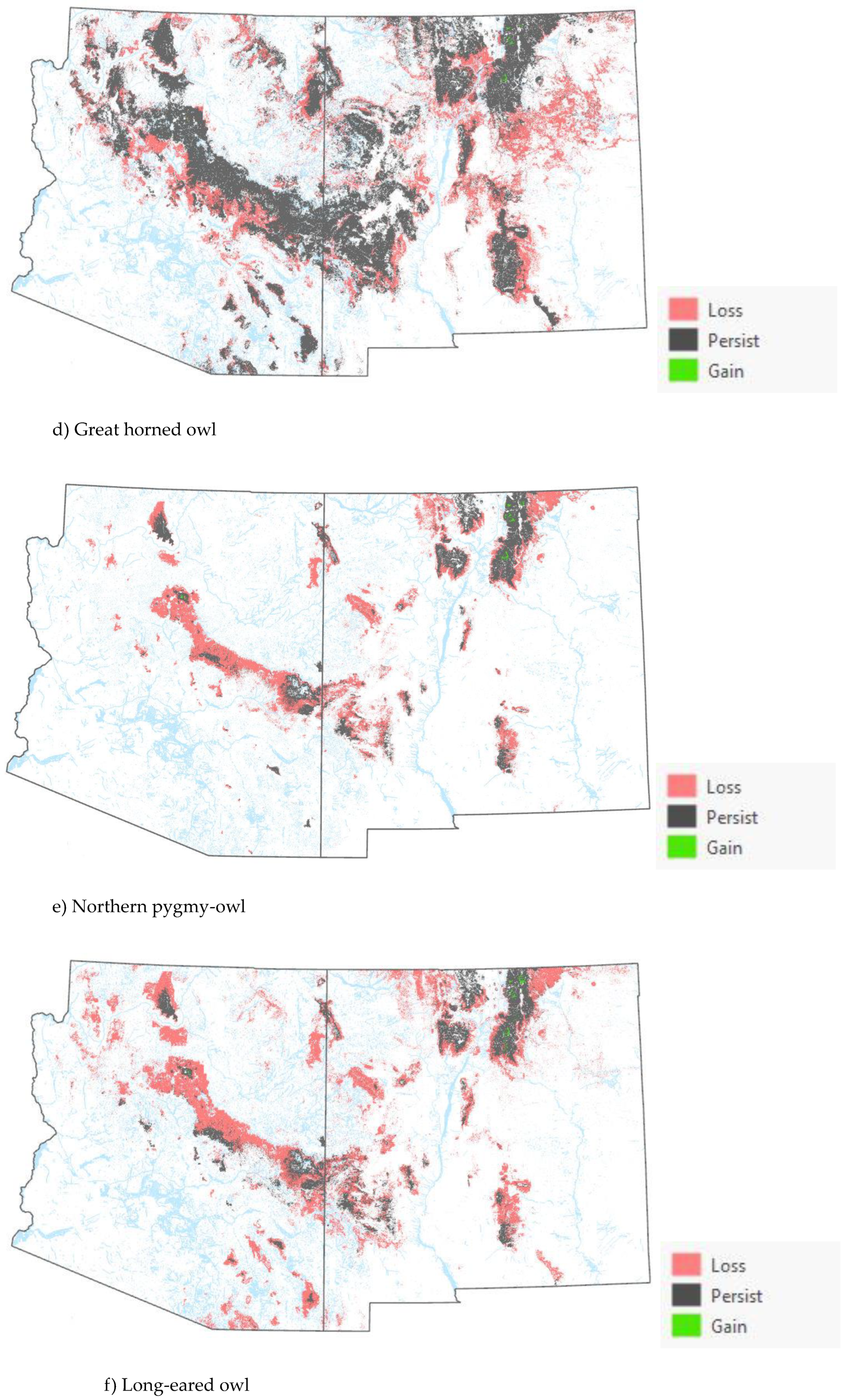
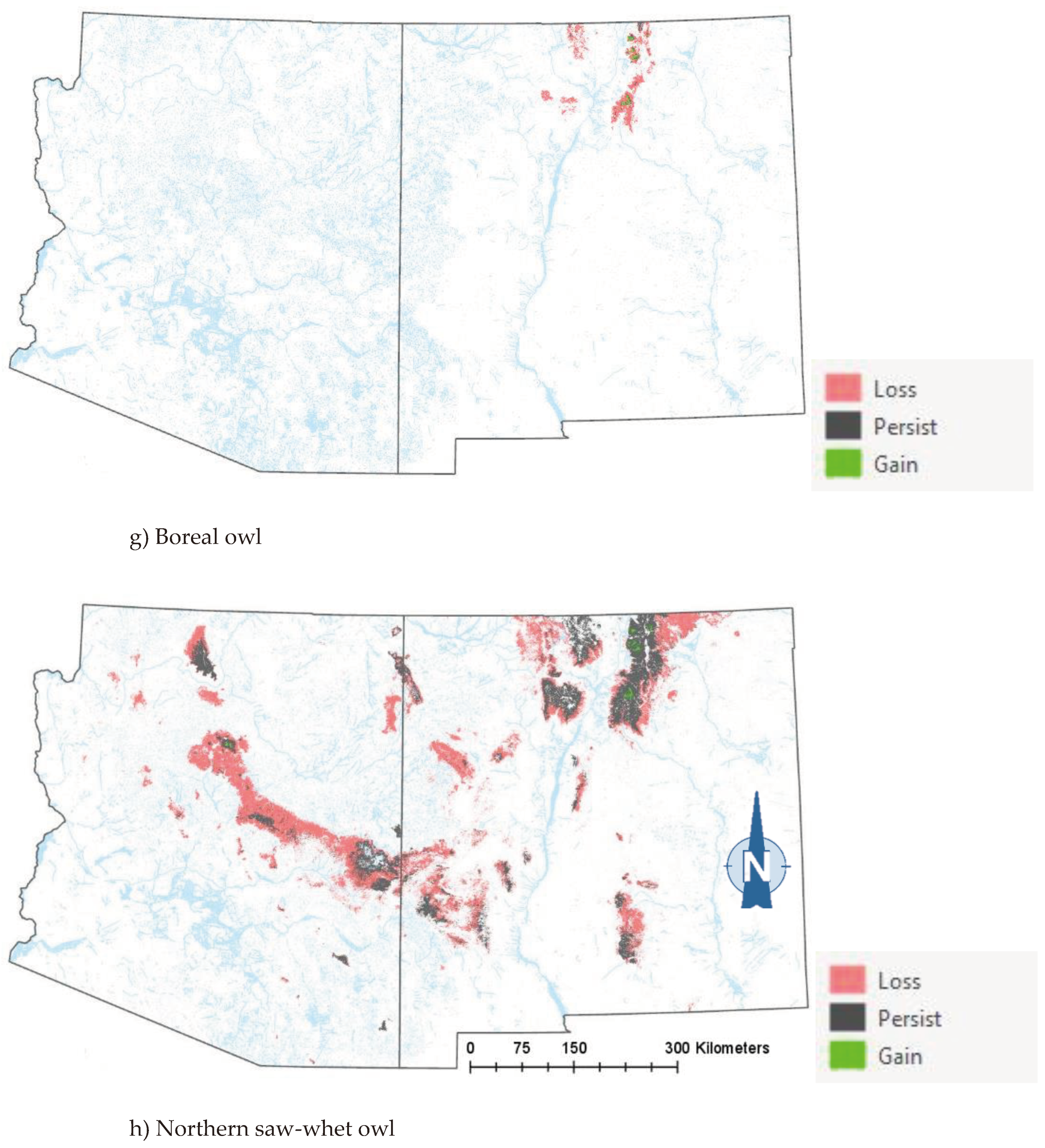
Figure 5.
Distribution of flammulated owl territories (red triangles) and nests (black triangles) across current and 2090 projected general vegetation types in the Black Range and in the Jemez, Magdalena, San Mateo, and Zuni Mountains of New Mexico. In the Zuni Mountains, all 72 recorded flammulated owl territories and nests were mapped in dry forest (ponderosa pine, pine-oak, and frequent-fire mixed-conifer forest) under current conditions; vegetation shifts projected by 2090 placed all 72 territories and nests in pinyon-juniper instead, outside the species’ habitat profile.
Figure 5.
Distribution of flammulated owl territories (red triangles) and nests (black triangles) across current and 2090 projected general vegetation types in the Black Range and in the Jemez, Magdalena, San Mateo, and Zuni Mountains of New Mexico. In the Zuni Mountains, all 72 recorded flammulated owl territories and nests were mapped in dry forest (ponderosa pine, pine-oak, and frequent-fire mixed-conifer forest) under current conditions; vegetation shifts projected by 2090 placed all 72 territories and nests in pinyon-juniper instead, outside the species’ habitat profile.
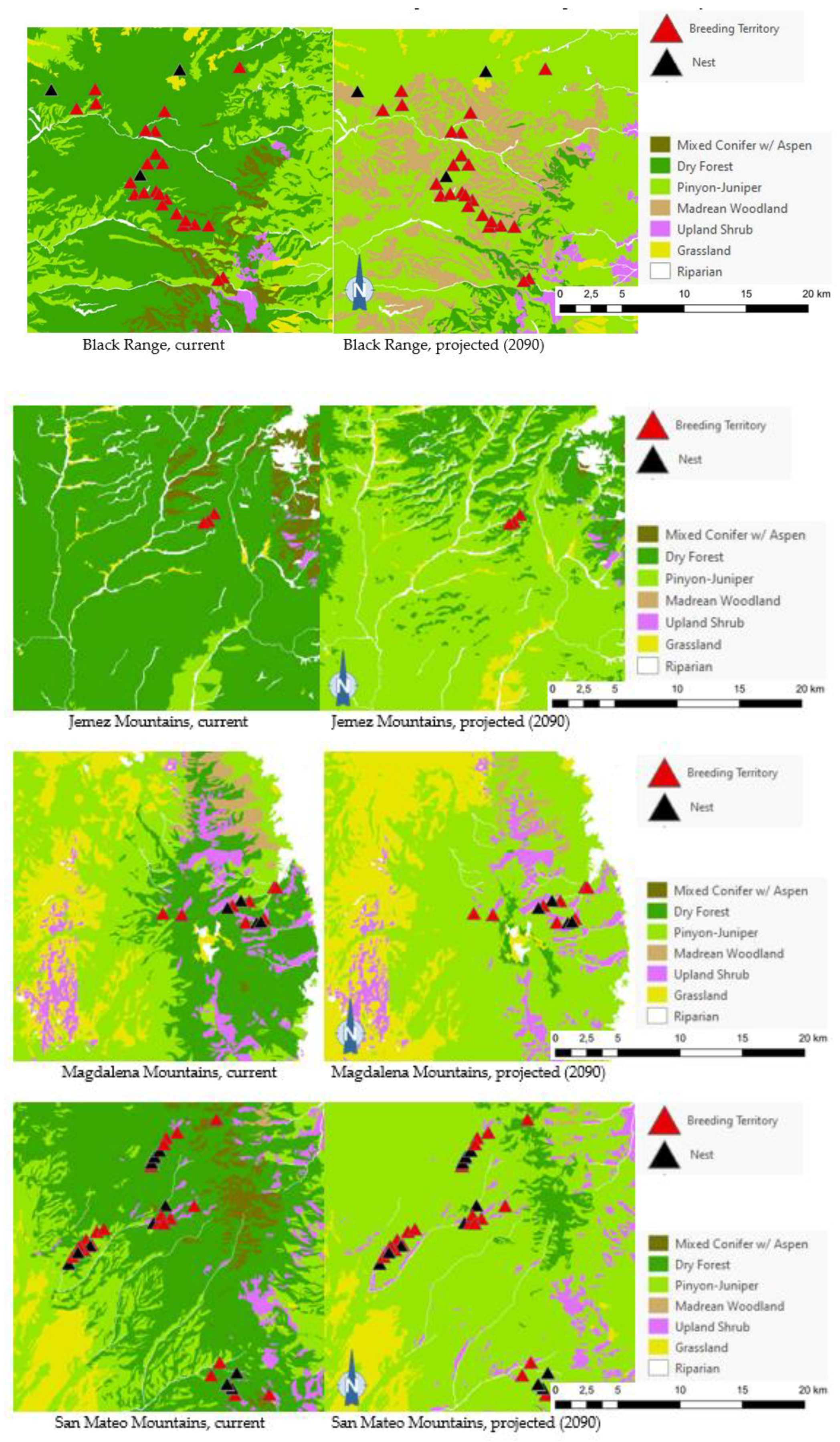
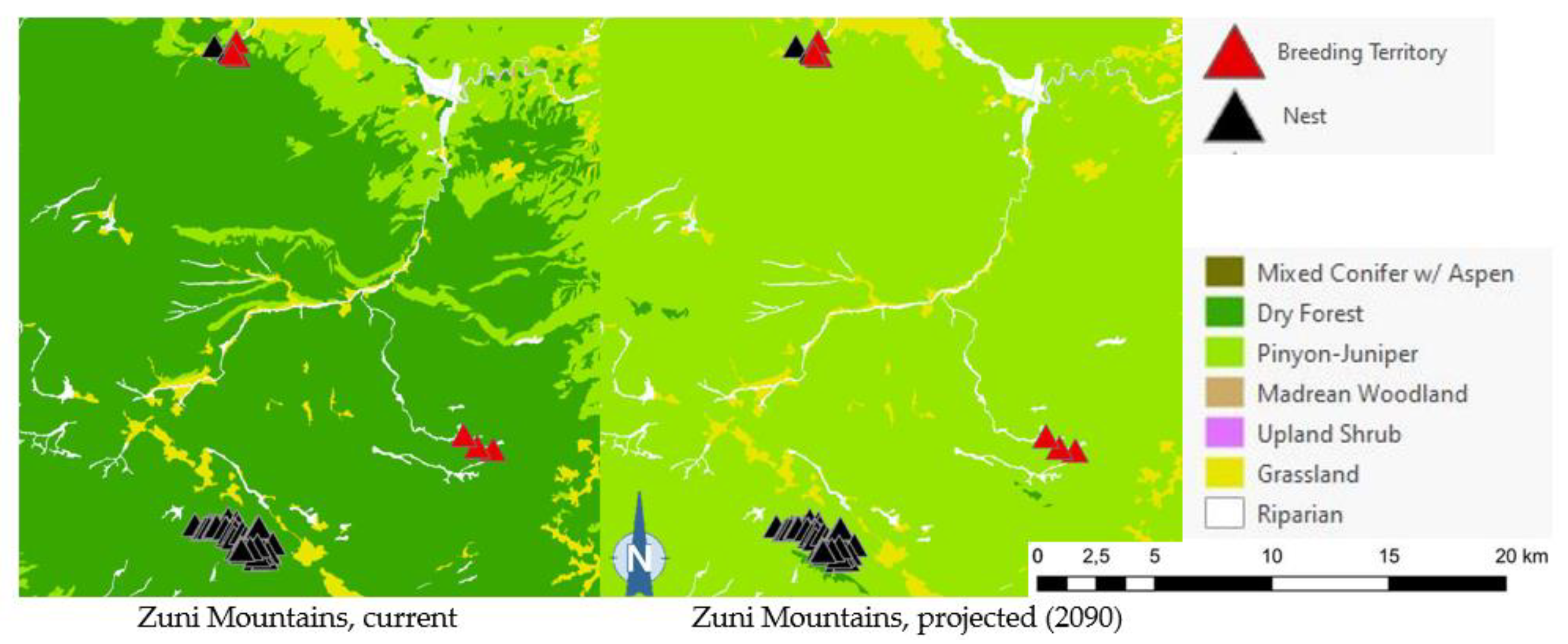
Figure 6.
Distribution of northern saw-whet owl detection points (red squares) among current and 2090 projected general vegetation types in the White Horse Sampling Quadrat on Arizona’s Coconino Plateau. All 13 northern saw-whet owl 1999 detection points were mapped in ponderosa pine forest under current conditions; vegetation shifts projected by 2090 placed those same detection points in pinyon-juniper instead, outside the species’ habitat profile.
Figure 6.
Distribution of northern saw-whet owl detection points (red squares) among current and 2090 projected general vegetation types in the White Horse Sampling Quadrat on Arizona’s Coconino Plateau. All 13 northern saw-whet owl 1999 detection points were mapped in ponderosa pine forest under current conditions; vegetation shifts projected by 2090 placed those same detection points in pinyon-juniper instead, outside the species’ habitat profile.
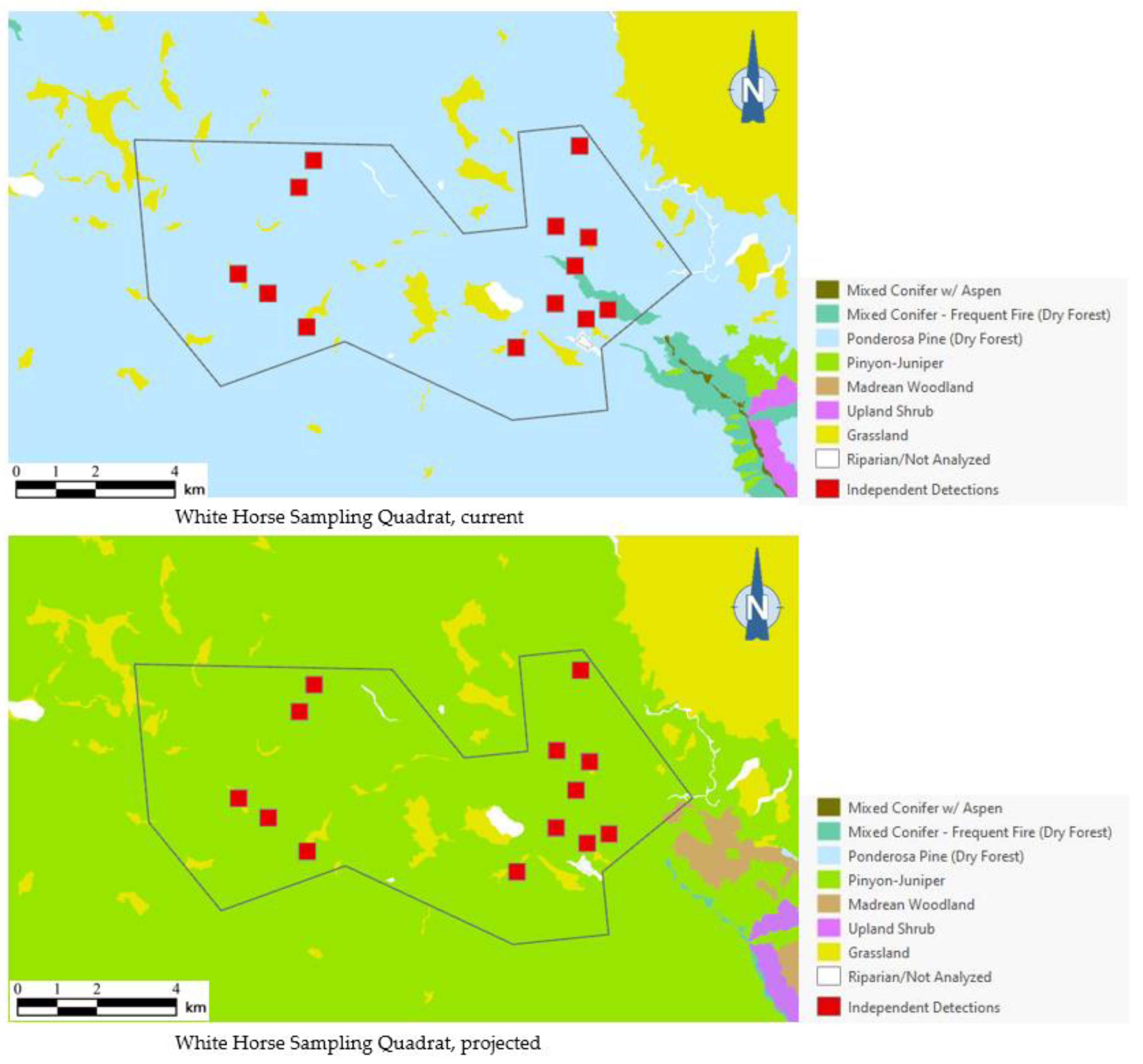

Table 1.
Ecological Response Units present in foothills and mountains of the southwestern U.S. (New Mexico and Arizona) and used for species habitat models1.
Table 1.
Ecological Response Units present in foothills and mountains of the southwestern U.S. (New Mexico and Arizona) and used for species habitat models1.
| Vegetation Zone | Ecological Response Unit |
|---|---|
| Subalpine Zone | Spruce-fir forest |
| Bristlecone pine forest | |
| Montane Zone | Mixed conifer with aspen |
| Mixed conifer-frequent fire | |
| Ponderosa pine forest | |
| Ponderosa pine-evergreen oak forest | |
| Woodland Zone | Pinyon-juniper woodland |
| Madrean pinyon-oak woodland | |
| Madrean encinal woodland | |
| Pinyon-juniper sagebrush | |
| Pinyon-juniper evergreen shrub | |
| Pinyon-juniper deciduous shrub | |
| Pinyon-juniper grass | |
| Juniper grass |
1 Though they represent additional Ecological Response Units, riparian systems were excluded from species habitat models as they could not be modeled under projected future (2090) conditions.
Table 2.
Habitat model performance and results of spatial randomized sampling procedures and 1-tailed z-tests to accept or reject the null hypothesis that predicted habitat captured a proportion of occurrence records that is not significantly greater than that of a randomly generated sample from within woodlands and forests1, 2.
Table 2.
Habitat model performance and results of spatial randomized sampling procedures and 1-tailed z-tests to accept or reject the null hypothesis that predicted habitat captured a proportion of occurrence records that is not significantly greater than that of a randomly generated sample from within woodlands and forests1, 2.
| Species | No. records | No. simulations equaling or outperforming habitat model | ||||
|---|---|---|---|---|---|---|
| Withinmodel | Within buffer | Outside buffer | Total | Without the 1-km buffer | With the 1-km buffer | |
| Flammulated owl | 248 | 30 | 13 | 291 | 0**** | 0**** |
| Western screech-owl | 34 | 39 | 60 | 137 | 34(*) | 0**** |
| Whiskered screech-owl | 71 | 41 | 0 | 112 | 307 | 0**** |
| Great horned owl | 125 | 13 | 10 | 148 | 500 | 500 |
| Northern pygmy-owl | 47 | 21 | 1 | 69 | 0**** | 0**** |
| Long-eared owl | 14 | 4 | 5 | 23 | 17* | 0**** |
| Boreal owl | 17 | 3 | 0 | 20 | 0**** | 0**** |
| Northern saw-whet owl | 22 | 6 | 6 | 34 | 0**** | 0**** |
1 Randomized sampling results were based on 500 iterations for every species. 2 Significant P–values are indicated in bold; **** P < 0.0001, *** P < 0.001, ** P < 0.01, P < 0.05, (*) P < 0.1.
Table 3.
Results of McNemar tests (Chi-square values) to accept or reject the null hypothesis that the proportion of records or detection points within vs. outside 2017 mapped modeled habitat did not differ between current and projected climate-induced conditions1.
Table 3.
Results of McNemar tests (Chi-square values) to accept or reject the null hypothesis that the proportion of records or detection points within vs. outside 2017 mapped modeled habitat did not differ between current and projected climate-induced conditions1.
| Species | No. observations | Mapped habitat | |||
|---|---|---|---|---|---|
| With the 1-km buffer | Without the 1-km buffer | ||||
| χ2 | P-value | χ2 | P-value | ||
| Flammulated owl | 291 | 162.01 | <0.0001**** | 223 | <0.0001**** |
| Western screech-owl | 137 | 40.02 | <0.0001**** | 27.03 | <0.0001**** |
| Whiskered screech-owl | 112 | 104.01 | <0.0001**** | 69.01 | <0.0001**** |
| Great horned owl | 148 | 3.2 | 0.07(*) | 8.1 | 0.004** |
| Northern pygmy-owl | 69 | 31.03 | <0.0001**** | 36.03 | <0.0001**** |
| Long-eared owl | 23 | 4.17 | 0.04* | 5.14 | 0.02* |
| Boreal owl | 20 | 4.17 | 0.04* | 7.11 | 0.008** |
| Northern saw-whet owl | 34 | 11.08 | 0.0008*** | 16.06 | <0.0001**** |
1 Significant P–values are indicated in bold; **** P < 0.0001,*** P < 0.001, ** P < 0.01, P < 0.05, (*) P < 0.1.
Table 4.
Current and projected habitat area for each owl species analyzed, along with percent change in extent.
Table 4.
Current and projected habitat area for each owl species analyzed, along with percent change in extent.
| Species | Vegetation Types Representing Preferred or Primary Habitat | Owl Habitat Model Zones(Figure 1) | Approx. Current 2017 Extent (ha) | Approx. Projected 2090 Extent (ha) | Projected 2090 % Change |
|---|---|---|---|---|---|
| Flammulated owl | Montane forests | Regionwide | 4,684,000 | 714,000 | -85% |
| Western screech-owl | Pinyon-juniper and Madrean woodlands | Regionwide | 3,186,000 | 1,511,000 | -55% |
| Whiskered screech-owl | Ponderosa and ponderosa pine-evergreen oak forest, and Madrean woodlands | Sky Island – Bootheel, Pecos East (Guadalupe Mtns only), AZ-NM Mtns (2090 only), Pecos West (2090 only) | 714,000 | 677,000 | -5% |
| Great horned owl | All forest and woodland types | Regionwide | 16,984,000 | 11,273,000 | -35% |
| Northern pygmy-owl | Subalpine and montane forests, pinyon-juniper and Madrean woodlands | Regionwide | 8,267,000 | 2,021,000 | -75% |
| Long-eared owl | Subalpine and montane forests, pinyon-juniper and Madrean woodlands | Regionwide | 8,263,000 | 2,623,000 | -70% |
| Boreal owl | Subalpine forests | Sangre de Cristo, San Juan, and Jemez mountains | 306,000 | 42,000 | -85% |
| Northern saw-whet owl | Subalpine and montane forests | Regionwide except Sonoran zone | 5,080,000 | 2,008,000 | -60% |
Table 5.
Number of flammulated owl sample territories among current and projected vegetation types in the Black Range and the Jemez, Magdalena, San Mateo, and Zuni Mountains in western New Mexico.
Table 5.
Number of flammulated owl sample territories among current and projected vegetation types in the Black Range and the Jemez, Magdalena, San Mateo, and Zuni Mountains in western New Mexico.
| Projected Vegetation Type (to) | ||||||
|---|---|---|---|---|---|---|
| Current Vegetation Type (from) | Dry Forest | Madrean Woodland | Pinyon-Juniper | Riparian | Gambel Oak Shrubland | Total |
| Dry Forest1 | 1 | 9 | 122 | 132 | ||
| Mixed Conifer w/ Aspen | 4 | 4 | ||||
| Pinyon-Juniper | 2 | 1 | 3 | |||
| Riparian | 7 | 7 | ||||
| Upland Shrub | 2 | 2 | ||||
| Total | 5 | 9 | 124 | 7 | 3 | 148 |
1 Dry forest consists of Ponderosa Pine Forest, Ponderosa Pine–Evergreen Oak Forest, and Mixed Conifer / Frequent Fire combined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



