
Submitted:
12 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
The blood–brain barrier (BBB) plays pivotal roles in synaptic and neuronal functioning by sealing the space between adjacent microvascular endothelial cells. BBB breakdown is present in patients with mild cognitive impairment (MCI) or Alzheimer disease (AD). Claudin-5 (CLDN-5) is a tetra-spanning protein essential for sealing the intercellular space between adjacent endothelial cells in the BBB. In this study, we developed a blood-based assay for CLDN-5 and investigated its diagnostic utility by using 100 cognitively normal (control) subjects, 100 patients with MCI, and 100 patients with AD. Plasma CLDN-5 levels were increased in patients with AD (3.08 ng/mL) compared with controls (2.77 ng/mL). Plasma levels of phosphorylated tau (pTau181), a biomarker of pathological tau, were elevated in patients with MCI or AD (2.86 and 4.20 pg/mL, respectively) compared with control subjects (1.81 pg/mL). In patients with MCI or AD, plasma levels of CLDN-5—but not pTau181—decreased with age, suggesting some age-dependent BBB changes in MCI and AD. These findings suggest that plasma CLDN-5 may a potential biochemical marker for the diagnosis of MCI and AD.
Keywords:
blood-brain barrier
; claudin-5
; tight junction
; biomarker
; dementia
; Alzheimer disease
1. Introduction
Currently more than 50 million people worldwide have some form of dementia, and this population is anticipated to grow to 78 million by 2030 [1]. Alzheimer disease (AD) is the most common cause of dementia. AD is a progressive disease, the preclinical stage of which includes mild cognitive impairment (MCI). Current therapeutic strategies for AD aim to reduce cognitive decline and involve various cognitive enhancers (e.g., donepezil, memantine) and disease modifiers (e.g., aducanumab, lecanemab) [2,3]. To maximize therapeutic efficacy, identifying people at risk for AD and initiating treatment during preclinical or early AD is crucial.
The clinical symptoms of AD reflect the associated progressive brain atrophy and subsequent cognitive decline. However, various molecular pathologies in the brain—including the accumulation of amyloid protein, phosphorylation of tau protein, and breakdown of the blood–brain barrier (BBB)—typically precede clinical symptoms [4,5]. Diagnosis of AD rests on measuring the accumulation of amyloid or tau proteins in the brain or their spillover into the cerebrospinal fluid (CSF) or blood. However, the collection of CSF typically involves lumbar puncture, which is an invasive procedure. An alternative method is positron emission tomography (PET) using amyloid and tau probes, which detects deposits of these proteins in the brain. Although PET imaging is minimally invasive, it is not widely used because of its expense [6]. Blood-based tests have recently gained attention as alternative diagnostic options because they are minimally invasive and cost effective. Blood tests for amyloid and phosphorylated tau proteins have been developed and are used in people who have symptoms of MCI or AD [7,8].
The BBB functions as a selective gate for the uptake of essential molecules from blood into the brain and the excretion of harmful molecules from the brain into blood via transporters and receptors on cellular membranes [9]. In addition, the BBB prevents the influx of blood-borne neurotoxins, cells, and pathogens into the brain because of the formation of tight junctions (TJs) in the intercellular space between adjacent macrovascular endothelial cells. Loss of BBB integrity has been observed in neuroinflammatory disorders [5], and patients with early AD demonstrate BBB leakage [10]. In addition, patients with early cognitive dysfunction show BBB breakdown in the hippocampus, which occurs independently of brain accumulation of amyloid and tau [11]. These several findings indicate that BBB TJ-sealing components might be impaired in MCI- and AD-related pathology.
Claudin is a tetra-transmembrane protein and an essential component of the TJ seal [12]. In mammals, claudins comprise a protein family of 27 members, and claudin-5 (CLDN-5) is a pivotal TJ-sealing protein in the BBB [12,13]. Blood CLDN-5 levels are elevated in patients with multiple sclerosis [14], and loss of CLDN-5 was accompanied by BBB breakdown in a mouse model of autoimmune encephalomyelitis [15]. These findings suggested to us that blood CLDN-5 levels might be altered in patients with MCI and AD compared with their cognitively normal individuals. To investigate this idea, we first used a monoclonal antibody (mAb) that recognizes the extracellular loops of human CLDN-5 to develop a highly sensitive single-molecular array (Simoa) assay, which is more sensitive than conventional immunoassays. We then used this assay to qualitatively investigate plasma CLDN-5 levels in cognitively normal adults and patients with MCI or AD and considered the clinical implications of plasma CLDN-5 levels in AD.
2. Results
2.1. Establishment of an immunoassay system using Simoa
To investigate whether plasma CLDN-5 protein levels are changed in patients with MCI and AD, we established a sensitive immunoassay system using Simoa technology and an anti-CLDN-5 mAb against the second extracellular loop domain of human CLDN-5 [16].
To this end, we chemically coated magnetic beads with antibody recognizing the extracellular loop domain of CLDN-5. Samples were incubated with anti-CLDN-5–coated beads; the bound CLDN-5 was sandwiched with a biotinylated anti-CLDN-5 antibody against the intracellular domain of CLDN-5; and the complexes were incubated with streptavidin β-galactosidase, followed by addition of chemiluminescent substrate (Figure 1A). The generated chemiluminescence signal was measured through digital scanning. The Simoa immunoassay for CLDN-5 had a dynamic range of 0.01 to 3 ng/mL, with a limit of detection of 0.001 ng/mL; a regression line with good fit (R2 = 0.992); low coefficient of variability; high accuracy; and high CLDN-5 specificity (Figure 1B,C, Table 1).
Spearman rank-correlation analyses of plasma CLDN-5 concentration and age are shown for (A) cognitively normal controls, (B) patients with MCI, and (C) patients with AD; Those of plasma pTau-181 and age are shown for (D) cognitively normal controls, (E) patients with MCI, and (F) patients with AD; and those of plasma CLDN-5 and pTau-181 concentrations are shown in (G) cognitively normal controls, (H) patients with MCI, and (I) patients with AD.
2.2. Association of plasma CLDN-5 levels with the stages of cognitive decline
We obtained plasma samples from cognitively healthy (control) adults and patients with MCI or AD patients from Japan’s National Center for Geriatrics and Gerontology Biobank (Table 2). The median plasma CLDN-5 level increased with cognitive impairment: 2.77, 2.91, and 3.08 ng/mL in the control, MCI, and AD groups, respectively (Figure 2A). Plasma CLDN-5 levels were significantly higher in AD patients than in cognitively healthy adults (p < 0.05). As previously reported, plasma pTau-181 levels showed a positive relationship with the stages of cognitive decline (Figure 2B) [7]. Whereas cognitively normal subjects did not show an age-dependent change in CLDN-5 (Figure 3A), plasma CLDN-5 levels were higher in younger than in older patients in both the MCI (p < 0.001) and AD (p < 0.01) groups (Figure 3B,C). However, neither MCI nor AD patients showed an age-related change in plasma pTau-181 levels, a biomarker of tau phosphorylation (Figure 3D–F). CLDN-5 levels were not associated with pTau-181 levels in any of the groups, indicating that these are independent biomarkers (Figure 3G–I).
3. Discussion
A body of evidence supports that microvascular degeneration is associated with the pathophysiology of AD [5,17]. In the current study, we investigated associations of the plasma concentration of CLDN-5, a pivotal TJ-sealing protein in the microvascular endothelial cells of the BBB, with MCI and AD. We found that plasma CLDN-5 levels were higher in patients with AD than in cognitively healthy adults and plasma CLDN-5 levels were higher in younger than in older patients in both the MCI and AD groups.
Breakdown of the BBB, which is associated with CNS diseases, is accompanied by the invasion of leukocytes and activation of astrocytes [5]. The matrix metalloproteinases (MMPs) secreted by these invading leukocytes led to the degradation of CLDN-5 in the BBB of mice [18]. In a rat ischemic model, MMPs secreted from astrocytes likewise degraded CLDN-5 in the BBB [19]. In addition, the number of pericytes in the BBB was greater in patients with AD compared with cognitively healthy peers, perhaps reflecting a response to endothelial breakdown [20]. Similar to our findings for patients with MCI or AD, circulating CLDN-5 levels are elevated in other CNS diseases, including ischemic stroke, bipolar disorder, and obsessive–compulsive disorder [21,22,23]. The CLDN-5 circulating in blood might be derived from the endothelial cells in the BBB.
Interestingly, we found significant negative association of plasma CLDN-5 level with age in MCI and AD. Ultrastructural analysis of TJ seals in the BBB did not reveal normal age-associated changes [24]. In contrast, magnetic resonance imaging showed that people with no cognitive impairment had an age-dependent progressive loss of BBB integrity in the hippocampus, which plays a pivotal role in learning and memory and which is damaged in early AD [25]. In another study, adults with early cognitive dysfunction developed brain capillary damage and BBB breakdown in the hippocampus [11]. Furthermore, the number of pericytes in the BBB was increased in patients with AD compared with age-matched controls without dementia [20]. In the current study, CLDN-5 levels were higher in younger than in older patients in both the MCI and AD groups. This is consistent with the analysis of autopsied brains, which reported that CLDN-5 level decreases with AD progression [26]. Because reactive astrocytes and endothelial cells in the BBB in AD produced MMPs [27], prolonged activation of MMPs might lead to the degradation of CLDN-5 and thus the lower plasma CLDN-5 levels in older compared with younger patients with cognitive deficits.
We acknowledge several limitations of this study. First, the source of the circulating CLDN-5 remains unknown. CLDN-5 is expressed in endothelial cells throughout the body [28] and at greater levels by CNS endothelial cells than by those outside of the CNS [29]. Endothelial cell-specific CLDN-5+/– mice, whose CLDN-5 protein level was 50% lower than that of CLDN-5+/+ mice, had learning and memory deficits [30,31]. Downregulation of CLDN-5 in the prefrontal cortex promoted anxiety-like and depression-like behaviors in mice [32]. Suppression of CLDN-5 in the hippocampus impaired spatial recognition memory in mice [31]. Together, these data suggest that local suppression of CLDN-5 levels may affect the onset of AD. Second, we did not investigate comorbidities of AD. Patients with chronic kidney diseases have higher risks of dementia and MCI than do members of the general population [33]. In addition, heart failure is a risk factor for AD [34,35], and epidemiologic studies indicate that AD is seen more frequently in people with asthma [36]. The influences of these complications need to be defined to clarify the clinical significance of an increased circulating CLDN-5 level. Third, we did not address whether the detected CLDN-5 protein is the post-translationally modified form. CLDN-5 has multiple glycosylation, phosphorylation, and ubiquitination sites, which may be associated with enhancing the permeability of the BBB [37,38]. Further investigation into the clinical meaning and utility of circulating CLDN-5 will be useful for the diagnosis of MCI and AD. In conclusion, CLDN-5 may be a potential biochemical marker of MCI and AD.
4. Materials and Methods
4.1. Clinical samples
Samples of human plasma were obtained from the national biobank of the National Center for Geriatrics and Gerontology (Aichi, Japan) [39]. To explore the diagnostic utility of a plasma CLDN-5 assay, we obtained plasma samples from cognitively normal subjects and those with MCI or AD from the National Center for Geriatrics and Gerontology Biobank (Table 2). Basic demographic data including age, gender, diagnosis, and Mini-Mental State Examination score were obtained from the biobank. To avoid analytical bias, the analysis of plasma CLDN-5 was carried out in a blinded manner, and the sample identities were only revealed from the biobank after the analysis. This study was approved by the ethics committees of Osaka University (protocol no. Yakuhito2020-1-4), and the National Center for Geriatrics and Gerontology (20TB6). All methods were performed in accordance with relevant guidelines and regulations.
4.2. Purification of anti-CLDN-5 mAbs
Purified mAbs (clone R9) were prepared as described previously [40]. Briefly, hybridoma cells were cultured in Hybridoma SFM medium (Thermo Fisher Scientific, Waltham, MA) containing 10% BM Condimed H1 (Roche, Mannheim, Germany). Rat anti-CLDN-5 mAb was purified from the culture media by using Protein G Sepharose 4 Fast Flow columns (Cytiva, Marlborough, MA, USA). Buffer exchange was performed by using phosphate-buffered saline (PBS, pH 7.4) and centrifugal filter tubes with a molecular weight cutoff of 100 kDa (Amicon Ultra-100K, Merck Millipore, Burlington, MA, USA). Purified mAb was sterilized by filtering it through a 0.22-µm filter and was stored at −30°C. The concentration of mAb was quantified by using a BCA Protein Assay kit (Nacalai Tesque, Kyoto, Japan), with bovine serum albumin as the standard.
4.3. Preparation of CLDNs
Cell-free synthesis of CLDN recombinant proteins was performed with the bilayer-dialysis method as described previously with minor modifications [41,42]. Briefly, in vitro transcription was performed with SP6 RNA polymerase (CellFree Sciences, Matsuyama, Japan). The translation reaction mixture (500 µL) containing mRNA (25%), WEPRO 7240 wheat germ extract (25%; CellFree Sciences), creatine kinase (40 μg/mL; Roche), and asolectin liposomes (10 mg lipids/mL) was overlaid with 2 mL of SUB-AMIX SGC dialysis solution (CellFree Sciences) in a 10-K MWCO Slide-A-Lyzer MINI dialysis device (Thermo Fisher Scientific); the cup was then immersed in 3.5 mL of SUB-AMIX SGC solution and incubated at 16 °C for 24 h. Cell-free synthesized proteoliposomes were collected by centrifugation at 20,000 × g for 10 min at 4 °C, and the resultant pellet was suspended in HBS buffer (20 mM Hepes-NaOH, pH 7.2, 150 mM NaCl). Proteoliposomes were washed three times with HBS, finally being resuspended in 125 μL of HBS buffer. To solubilize the proteoliposomes, 125 μL of a solution (20 mM Hepes-NaOH, pH 7.2, 150 mM NaCl, 1% n-dodecyl-β-D-maltoside (Dojindo, Kumamoto, Japan), 10% glycerol (Nacalai Tesque), 1 mM DTT) was mixed with the 125 µL proteoliposomes suspension. The mixture was rotated gently at 4 ˚C for 1 h, sonicated for 3 min at 4 ˚C using an ultrasonic disruptor (SONIFIER model 450 Advanced, BRANSON, CT, USA) then centrifuged at 17,800 × g and 4 ˚C for 15 min. The supernatant was aliquoted in small portions, frozen in liquid nitrogen, and stored at –80 ˚C.
4.4. Simoa assay for CLDN-5
Plasma CLDN-5 concentrations were determined with Simoa technology and a Homebrew assay kit (Quanterix, Billerica, MA, USA) in accordance with the manufacturer’s instructions. In the first step of the assay, carboxylated paramagnetic beads were activated by using 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride (Thermo Fisher Scientific), followed by incubation with rat anti-CLDN-5 mAb (clone R9) for 2 h at 4 ºC with rotation. After blocking of the antibody-coupled beads with Bead Blocking Buffer (Quanterix), the beads were resuspended with Bead Diluent (Quanterix) and stored at 4 ºC until further use. For biotinylation of the detection antibody, bovine serum albumin and azide-free rabbit anti-CLDN-5 mAb (Abcam, Cambridge, United Kingdom) were mixed with NHS-PEG4-Biotin (Thermo Fisher Scientific) and reacted at room temperature for 30 min. The biotinylated antibody was buffer-exchanged into PBS by using an Amicon centrifugal filter with a cutoff value of 50 kDa (Merck Millipore) and stored at –20ºC until use.
Singleplex Simoa assays were performed by using the HD-X Analyzer (Quanterix, Billerica, MA, USA). Human plasma samples and standard human CLDN-5 proteins were incubated with the antibody-coated capture beads (250,000 beads per test) for 15 min. After washes, the beads were incubated with biotinylated detector antibody for 5.25 min. After a series of washes, streptavidin–β-galactosidase (Quanterix) solution was added and incubated for 5.25 min. The beads were washed again, mixed with the enzyme substrate resorufin β-D-galactopyranoside (Quanterix), and loaded onto the disc microarray. The array was sealed with oil, and the average enzyme per bead values were calculated by the software in the HD-X Analyzer [43].
Plasma pTau-181 concentrations were determined by using Simoa technology and the pTau-181 Advantage V2 kit (Quanterix) on the HD-X Analyzer in accordance with the manufacturer’s instructions.
4.5. Data analysis
Data are expressed as mean ± standard deviation. Standard curves for determining unknown sample concentrations were fitted by using Prism (version 9, GraphPad Software, Boston, MA, USA) four-parameter logistic fit. Statistical analyses were performed by using the Dunn multiple-comparison test (Prism, GraphPad Software), and correlations were calculated by using Spearman rank correlation analysis (Prism, GraphPad Software).
Author Contributions
KT: RH, KH, NS, TK, HT, and MK wrote the manuscript; KT, RH, KH, NS, TK, and MK designed the research project; KT performed research experiments; KT, RH, KH, NS, TK, HT, and MK analyzed the data; and KH, NS, TK, and HT contributed new reagents or analytical tools.
Acknowledgments
This study was supported by the Japan Agency for Medical Research and Development (AMED) [grant JP20hm0102084] and JSPS KAKENHI [grant numbers: 21H03354, 22H03953, 22K19936]. We are grateful to Drs. Toshiyuki Sato and Kenta Noda (Sysmex) for advice during the development of the Simoa system for CLDN-5, and Mrs. Yuko Fujieda for excellent technical support.
References
- World-Alzheimer-Report 2022.
- Hameed, S.; Fuh, J.L.; Senanarong, V.; Ebenezer, E.G.M.; Looi, I.; Dominguez, J.C.; Park, K.W.; Karanam, A.K.; Simon, O. Role of fluid biomarkers and PET imaging in early diagnosis and its clinical implication in the management of Alzheimer’s disease. J. Alzheimers Dis. Rep. 2020, 4, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Lee, G.; Nahed, P.; Kambar, M.E.Z.N.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2022. Alzheimers Dement. (N Y) 2022, 8, e12295. [Google Scholar] [CrossRef]
- Chen, H.; Xu, J.; Xu, H.; Luo, T.; Li, Y.; Jiang, K.; Shentu, Y.; Tong, Z. New insights into Alzheimer’s disease: Novel pathogenesis, drug target and delivery. Pharmaceutics 2023, 15, 1133. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Rabinovici, G.D.; Gatsonis, C.; Apgar, C.; Chaudhary, K.; Gareen, I.; Hanna, L.; Hendrix, J.; Hillner, B.E.; Olson, C.; Lesman-Segev, O.H.; et al. Association of Amyloid Positron Emission Tomography With Subsequent Change in Clinical Management Among Medicare Beneficiaries With Mild Cognitive Impairment or Dementia. JAMA 2019, 321, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, E.H. La Joie, R.; Wolf, A.; Strom, A.; Wang, P.; Iaccarino, L.; Bourakova, V.; Cobigo, Y.; Heuer, H.; Spina, A.; et al. Diagnostic value of plasma phosphorylated tau181 in Alzheimer’s disease and frontotemporal lobar degeneration. Nat. Med. 2020, 26, 387–397. [CrossRef] [PubMed]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.X.; Martins, R.; Rowe, C.; et al. High performance plasma amyloid-β biomarkers for Alzheimer’s disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Nelson, A.R.; Betsholtz, C.; Zlokovic, B.V. Establishment and dysfunction of the blood-brain barrier. Cell 2015, 163, 1064–1078. [Google Scholar] [CrossRef]
- van de Haar, H.J.; Burgmans, S.; Jansen, J.F.; van Osch, M.J.; van Buchem, M.A.; Muller, M.; Hofman, P.A.M.; Verhey, F.R.J.; Backes, W.H. Blood-brain barrier leakage in patients with early Alzheimer disease. Radiology 2016, 281, 527–535. [Google Scholar] [CrossRef]
- Nation D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A,R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 2019, 25, 270–276. [CrossRef]
- Tsukita, S.; Tanaka, H.; Tamura, A. The claudins: From tight junctions to biological systems. Trends Biochem. Sci. 2019, 44, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; et al. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Mandel, I.; Paperna, T.; Glass-Marmor, L.; Volkowich, A.; Badarny, S.; Schwartz, I.; Vardi, P.; Koren, I.; Miller, A. Tight junction proteins expression and modulation in immune cells and multiple sclerosis. J. Cell. Mol. Med. 2012, 16, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Argaw, A.T.; Gurfein, B.T.; Zhang, Y.; Zameer, A.; John, G.R. VEGF-mediated disruption of endothelial CLN-5 promotes blood-brain barrier breakdown. Proc. Natl. Acad. Sci. U S A 2009, 106, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Shirakura, K.; Okada, Y.; Takeda, H.; Endo, K.; Tamura, M.; et al. Claudin-5-binders enhance permeation of solutes across the blood-brain barrier in a mammalian model. J. Pharmacol. Exp. Ther. 2017, 363, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef]
- Feng, S.; Cen, J.; Huang, Y.; Shen, H.; Yao, L.; Wang, Y.; Chen, Z. Matrix metalloproteinase-2 and -9 secreted by leukemic cells increase the permeability of blood-brain barrier by disrupting tight junction proteins. PLoS One 2011, 6, e20599. [Google Scholar] [CrossRef]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef]
- Stewart, P.A.; Hayakawa, K.; Akers, M.A.; Vinters, H.V. A morphometric study of the blood-brain barrier in Alzheimer’s disease. Lab. Invest. 1992, 67, 734–742. [Google Scholar]
- Kazmierski, R.; Michalak, S.; Wencel-Warot, A.; Nowinski, W.L. Serum tight-junction proteins predict hemorrhagic transformation in ischemic stroke patients. Neurology 2012, 79, 1677–1685. [Google Scholar] [CrossRef]
- Kılıç, F.; Işık, Ü.; Demirdaş, A.; Doğuç, D.K.; Bozkurt, M. Serum zonulin and claudin-5 levels in patients with bipolar disorder. J. Affect. Disord. 2020, 266, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Işık, Ü.; Aydoğan Avşar, P.; Aktepe, E.; Doğuç, D.K.; Kılıç, F.; Büyükbayram, H. Serum zonulin and claudin-5 levels in children with obsessive-compulsive disorder. Nord. J. Psychiatry 2020, 74, 346–351. [Google Scholar] [CrossRef]
- Stewart, P.A.; Magliocco, M.; Hayakawa, K.; Farrell, C.L.; Del Maestro, R.F.; Girvin, J,; Kaufmann, J.C.; Vinters, H.V.; Gilbert, J. A quantitative analysis of blood-brain barrier ultrastructure in the aging human. Microvasc. Res. 1987, 33, 270–282. [CrossRef]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Shinohara, M.; Yamazaki, A.; Murray, M.E.; Liesinger, A.M.; Heckman, M.G.; Lesser, E.R.; Parisi, J.E.; Petersen, R.C.; et al. Selective loss of cortical endothelial tight junction proteins during Alzheimer’s disease progression. Brain 2019, 142, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovic, A.V.; Situ, M.; Citalan-Madrid, A.F.; Stamatovic, S.M.; Xiang, J.; Keep, R.F. Blood-brain barrier dysfunction in normal aging and neurodegeneration: Mechanisms, impact, and treatments. Stroke 2023, 54, 661–672. [Google Scholar] [CrossRef]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. U S A 1999, 96511–96516. [Google Scholar] [CrossRef]
- Scalise, A.A.; Kakogiannos, N.; Zanardi, F.; Iannelli, F.; Giannotta, M. The blood-brain and gut-vascular barriers: from the perspective of claudins. Tissue Barriers 2021, 9, 1926190. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.; Hanley, N.; Reschke, C.R.; Reddy, A.; Mäe, M.A.; Connolly, R.; Behan, C.; O’Keeffe, E.; Bolger, I.; Hudson, N.; et al. Microvascular stabilization via blood-brain barrier regulation prevents seizure activity. Nat. Commun. 2022, 13, 2003. [Google Scholar] [CrossRef]
- Greene, C.; Kealy, J.; Humphries, M.M.; Gong, Y.; Hou, J.; Hudson, N.; Cassidy, L.M.; Martiniano, R.; Shashi, V.; Hooper, S.R.; et al. Dose-dependent expression of claudin-5 is a modifying factor in schizophrenia. Mol. Psychiatry. 2018, 23, 2156–2166. [Google Scholar] [CrossRef]
- Dion-Albert, L.; Cadoret, A.; Doney, E.; Kaufmann, F.N.; Dudek, K.A.; Daigle, B.; Parise, L.F.; Cathomas, F.; Samba, N.; Hudson, N.; et al. Vascular and blood-brain barrier-related changes underlie stress responses and resilience in female mice and depression in human tissue. Nat. Commun. 2022, 13, 164. [Google Scholar] [CrossRef] [PubMed]
- Viggiano, D.; Wagner, C.A.; Martino, G.; Nedergaard, M.; Zoccali, C.; Unwin, R.; Capasso, G. Mechanisms of cognitive dysfunction in CKD. Nat. Rev. Nephrol. 2020, 16, 452–469. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Winblad, B.; Marengoni, A.; Klarin, I.; Fastbom, J.; Fratiglioni, L. Heart failure and risk of dementia and Alzheimer disease: a population-based cohort study. Arch. Intern. Med. 2006, 166, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Manemann, S.M.; Knopman, D.S.; St Sauver, J.; Bielinski, S.J.; Chamberlain, A.M.; Weston, S.A.; Jiang, R.; Roger, V.L. Alzheimer’s disease and related dementias and heart failure: A community study. J. Am. Geriatr. Soc. 2022, 70, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, M.A.; Dean, D.C.; Bendlin, B.B.; Jarjour, N.N.; Esnault, S.; Zetterberg, H.; Heslegrave, A.; Evans, M.D.; Davidson, R.J.; Busse, W.W. Neuroimaging and biomarker evidence of neurodegeneration in asthma. J. Allergy Clin. Immunol. 2022, 149, 589–98.e6. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Li, S.; Chung, S.H.; Zhu, L.; Stayt, J.; Su, T.; Couraud, P.; Romero, I.A.; Weksler, B.; Gillies, M.C. Tyrosine phosphorylation of VE-cadherin and claudin-5 is associated with TGF-β1-induced permeability of centrally derived vascular endothelium. Eur. J. Cell Biol. 2011, 90, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Awan, F.M.; Anjum, S.; Obaid, A.; Ali, A.; Paracha, R.Z.; Janjua, H.A. In-silico analysis of claudin-5 reveals novel putative sites for post-translational modifications: Insights into potential molecular determinants of blood-brain barrier breach during HIV-1 infiltration. Infect. Genet. Evol. 2014, 27, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.S.; Utshigisawa, T.; Ogura, H.; Aoki, T.; Kawakami, T.; Ohga, S.; Ohara, A.; Ito, E.; Yamamoto, T.; Kanno, H. Clinical and genetic diagnosis of thirteen Japanese patients with hereditary spherocytosis. Hum. Genome Var. 2022, 9, 1. [Google Scholar] [CrossRef]
- Tachibana, K.; Hashimoto, Y.; Shirakura, K.; Okada, Y.; Hirayama, R.; Iwashita, Y.; Nishino, I.; Ago, Y.; Takeda, H.; Kuniyasu, H.; Kondoh, M. Safety and efficacy of an anti-claudin-5 monoclonal antibody to increase blood-brain barrier permeability for drug delivery to the brain in a non-human primate. J. Control. Release 2021, 336, 105–111. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Zhou, W.; Hamauchi, K.; Shirakura, K.; Doi, T.; Yagi, K.; Sawasaki, T.; Okada, Y.; Kondoh, M.; Takeda, H. Engineered membrane protein antigens successfully induce antibodies against extracellular regions of claudin-5. Sci. Rep. 2018, 8, 8383. [Google Scholar] [CrossRef]
- Zhou, W.; Takeda, H. Cell-free production of proteoliposomes for functional analysis and antibody development targeting membrane proteins. J. Vis. Exp. 2020, 163, e61871. [Google Scholar] [CrossRef]
- Wilson, D.H.; Rissin, D.M.; Kan, C.W.; Fournier, D.R.; Piech, T.; Campbell, T.G.; Meyer, R.E.; Fishburn, M.W.; Cabrera, C.; Patel, P.P.; et al. The Simoa HD-1 Analyzer: A novel fully automated digital immunoassay analyzer with single-molecule sensitivity and multiplexing. J. Lab. Autom. 2016, 21, 533–547. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sensitivity and specificity of a plasma human claudin (CLDN) 5 protein detection system using Simoa. (A) Schematic illustration of singleplex detection of human CLDN-5 using Simoa. First, the capture CLDN-5 monoclonal antibody (mAb) (clone R9) was immobilized on carboxylated paramagnetic beads. CLDN-5 protein was captured on the antibody-coupled beads and then sandwiched with the biotinylated detector CLDN-5 mAb. Next, the beads were labeled with streptavidin–β-galactosidase. Finally, the immunocomplex beads were mixed with the enzyme substrate resorufin β-D-galactopyranoside and loaded onto the disc microarray. Fluorescent signals were analyzed automatically to obtain the average enzyme amount per bead. (B) Response curve of the Simoa assay for the detection of human CLDN-5. Data are given as mean ± standard deviation (n = 5). Standard curves were generated by using a four-parameter logistic model, and R-squared values were calculated. (C) Analysis of the specificity of CLDN-5 detection using Simoa. Proteoliposomes containing human CLDN-1 to -6 were tested at concentrations of 0.03, 0.3, and 3 ng/mL. Data are given as mean ± standard deviation (n = 3).
Figure 1.
Sensitivity and specificity of a plasma human claudin (CLDN) 5 protein detection system using Simoa. (A) Schematic illustration of singleplex detection of human CLDN-5 using Simoa. First, the capture CLDN-5 monoclonal antibody (mAb) (clone R9) was immobilized on carboxylated paramagnetic beads. CLDN-5 protein was captured on the antibody-coupled beads and then sandwiched with the biotinylated detector CLDN-5 mAb. Next, the beads were labeled with streptavidin–β-galactosidase. Finally, the immunocomplex beads were mixed with the enzyme substrate resorufin β-D-galactopyranoside and loaded onto the disc microarray. Fluorescent signals were analyzed automatically to obtain the average enzyme amount per bead. (B) Response curve of the Simoa assay for the detection of human CLDN-5. Data are given as mean ± standard deviation (n = 5). Standard curves were generated by using a four-parameter logistic model, and R-squared values were calculated. (C) Analysis of the specificity of CLDN-5 detection using Simoa. Proteoliposomes containing human CLDN-1 to -6 were tested at concentrations of 0.03, 0.3, and 3 ng/mL. Data are given as mean ± standard deviation (n = 3).
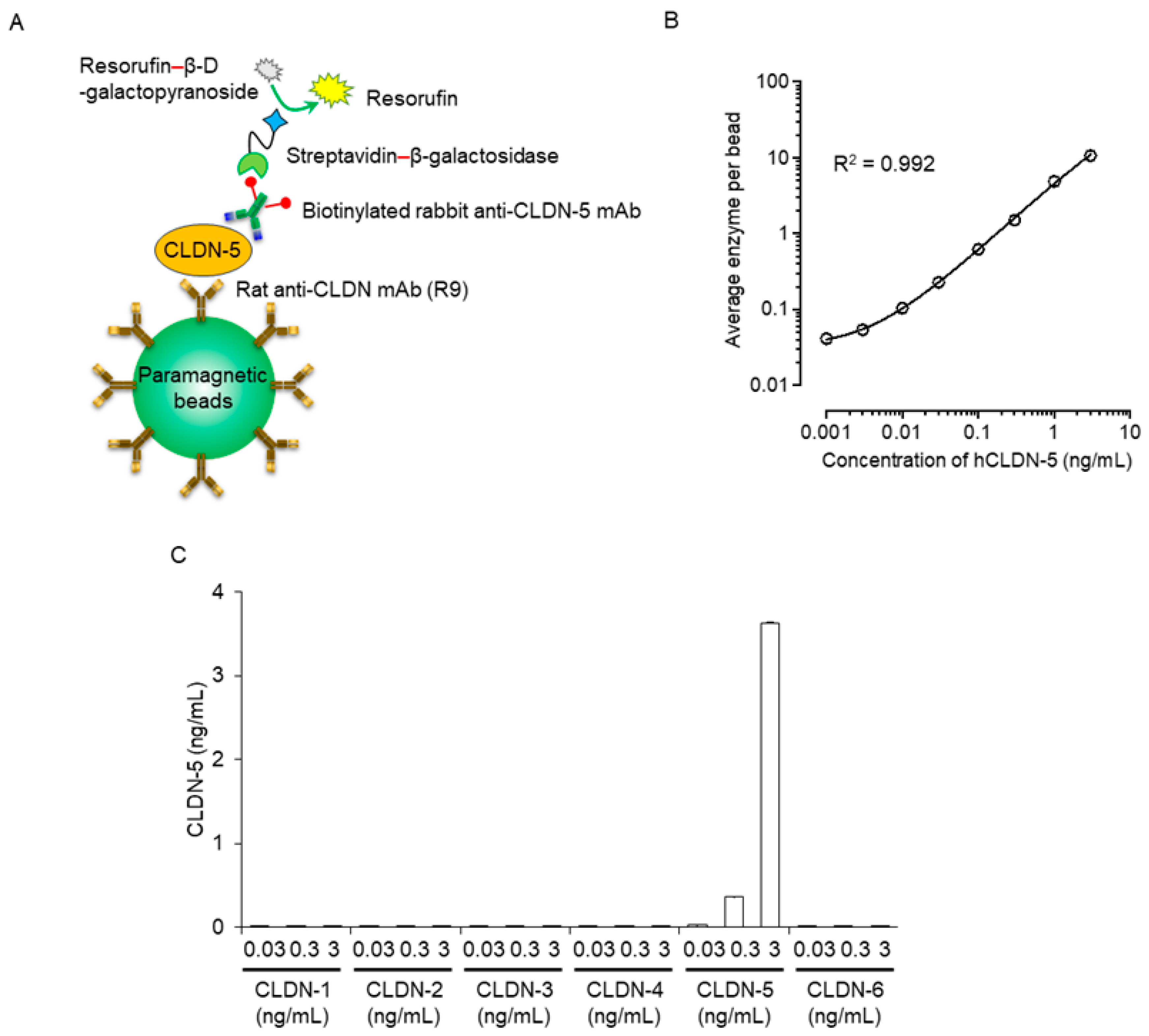
Figure 2.
Comparison of plasma CLDN-5 or pTau-181 levels among cognitively normal controls and patients with mild cognitive impairment (MCI) or Alzheimer disease (AD). Plasma samples were collected from cognitively normal individuals, MCI patients and AD patients in the NCGG Biobank (n = 100 per group). Plasma (A) CLDN-5 and (B) pTau-181 levels were measured by using a Simoa system as described in the Materials and Methods. The horizontal bar indicates the median value and the error bars the 25th and 75th percentiles of each group. The statistical analysis was performed using Kruskal–Wallis nonparametric ANOVA with the Dunn multiple-comparisons test.
Figure 2.
Comparison of plasma CLDN-5 or pTau-181 levels among cognitively normal controls and patients with mild cognitive impairment (MCI) or Alzheimer disease (AD). Plasma samples were collected from cognitively normal individuals, MCI patients and AD patients in the NCGG Biobank (n = 100 per group). Plasma (A) CLDN-5 and (B) pTau-181 levels were measured by using a Simoa system as described in the Materials and Methods. The horizontal bar indicates the median value and the error bars the 25th and 75th percentiles of each group. The statistical analysis was performed using Kruskal–Wallis nonparametric ANOVA with the Dunn multiple-comparisons test.
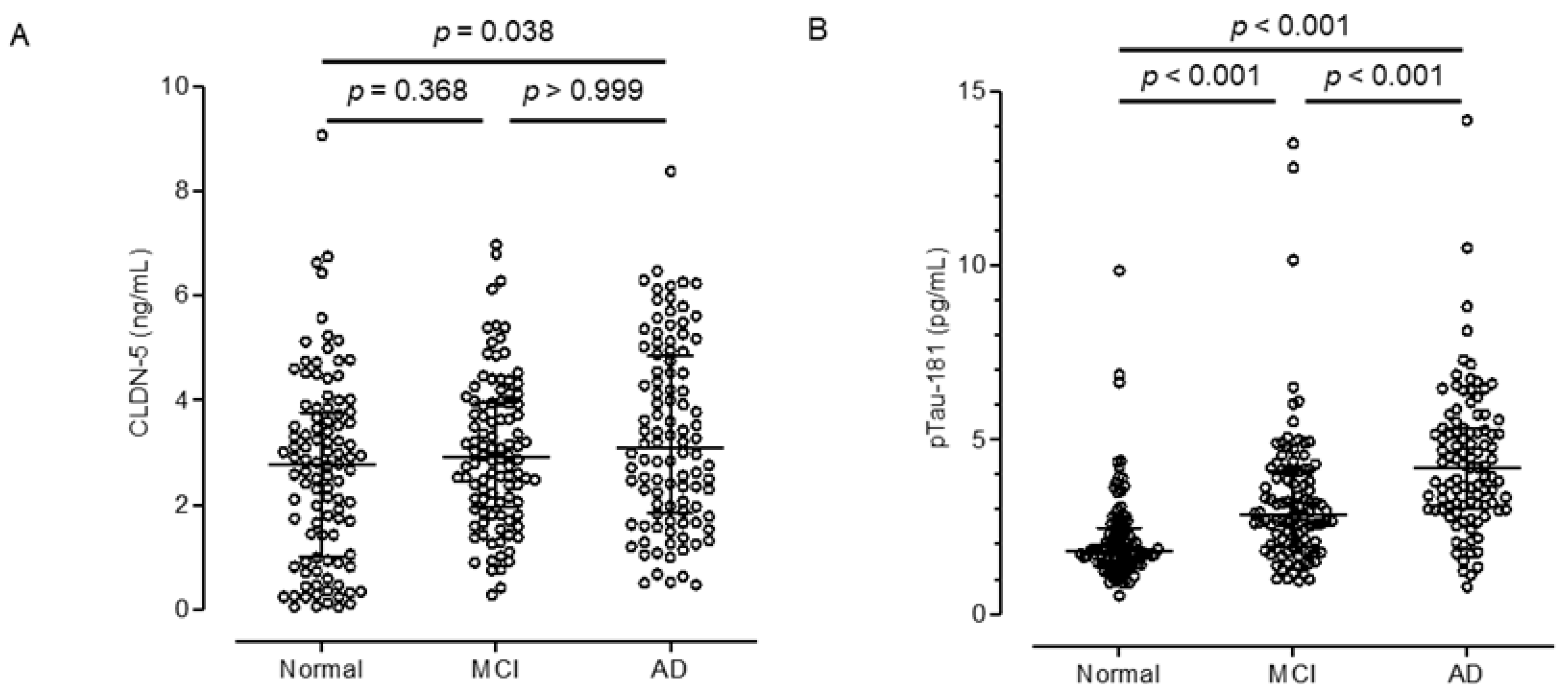
Figure 3.
Association of the plasma CLDN-5 level with age or plasma pTau-181 level. Spearman rank-correlation analyses of plasma CLDN-5 concentration and age are shown for (A) cognitively normal controls, (B) patients with MCI, and (C) patients with AD; Those of plasma pTau-181 and age are shown for (D) cognitively normal controls, (E) patients with MCI, and (F) patients with AD; and those of plasma CLDN-5 and pTau-181 concentrations are shown in (G) cognitively normal controls, (H) patients with MCI, and (I) patients with AD.
Figure 3.
Association of the plasma CLDN-5 level with age or plasma pTau-181 level. Spearman rank-correlation analyses of plasma CLDN-5 concentration and age are shown for (A) cognitively normal controls, (B) patients with MCI, and (C) patients with AD; Those of plasma pTau-181 and age are shown for (D) cognitively normal controls, (E) patients with MCI, and (F) patients with AD; and those of plasma CLDN-5 and pTau-181 concentrations are shown in (G) cognitively normal controls, (H) patients with MCI, and (I) patients with AD.
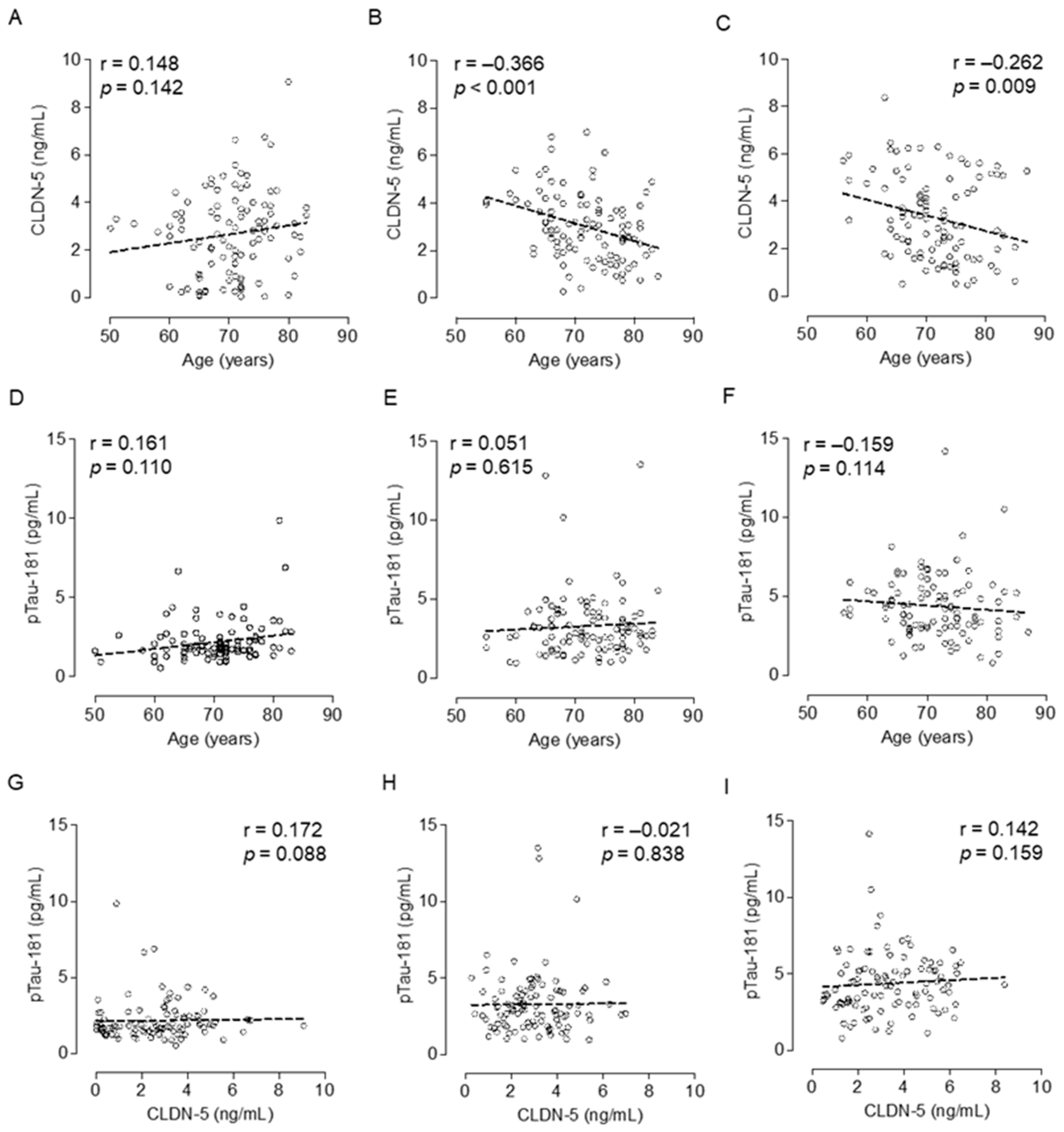

Table 1.
Precision and accuracy of quantification of human CLDN-5 concentration series.
| Theoretical concentration (ng/mL) | Actual concentration (ng/mL) | Mean (ng/mL) |
SD | CV (%) |
Accuracy | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||||
| 0 | ND | ND | ND | 0.000 | ND | 0.000 | NA | NA | NA |
| 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.000 | 13.80 | 117.18 |
| 0.003 | 0.003 | 0.004 | 0.003 | 0.003 | 0.003 | 0.003 | 0.000 | 15.51 | 95.37 |
| 0.01 | 0.010 | 0.011 | 0.011 | 0.010 | 0.009 | 0.010 | 0.001 | 7.61 | 102.21 |
| 0.03 | 0.029 | 0.033 | 0.030 | 0.029 | 0.029 | 0.030 | 0.002 | 5.74 | 100.19 |
| 0.1 | 0.103 | 0.108 | 0.096 | 0.105 | 0.090 | 0.100 | 0.007 | 7.47 | 100.27 |
| 0.3 | 0.290 | 0.315 | 0.293 | 0.280 | 0.208 | 0.277 | 0.041 | 14.73 | 92.46 |
| 1 | 1.094 | 1.152 | 1.082 | 1.083 | 1.048 | 1.092 | 0.038 | 3.47 | 109.19 |
| 3 | 3.008 | 3.180 | 2.888 | 2.517 | 2.981 | 2.915 | 0.246 | 8.44 | 97.16 |
CV, coefficient of variation; NA, not applicable; ND, not detected; SD, standard deviation.
Table 2.
Characteristics of patients who provided the plasma samples from Japan’s NCGG Biobank used in this study.
Table 2.
Characteristics of patients who provided the plasma samples from Japan’s NCGG Biobank used in this study.
| Characteristic | Cognitively normal (n = 100) |
MCI (n = 100) |
AD (n = 100) |
|---|---|---|---|
| Age, y | 70.3 ± 6.7 | 71.7 ± 6.6 | 71.5 ± 6.7 |
| Female, n (%) | 50 (50.0) | 50 (50.0) | 50 (50.0) |
| MMSE score | 29.5 ± 0.7 | 22.2 ± 1.9 | 16.0 ± 3.8 |
AD, Alzheimer’s disease; MCI, mild cognitive impairment; MMSE, Mini-Mental State Examination. Data are shown as mean ± S.D.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



