
Submitted:
15 November 2023
Posted:
15 November 2023
You are already at the latest version
Abstract
Growing food autonomously on Mars is challenging due to the Martian soil's low nutrient content and high salinity. Understanding how plants adapt and evaluating their nutritional attributes are pivotal for sustained Mars missions. This research delved into the regeneration, stress tolerance, and dietary metrics of sweet potato (Ipomoea batatas) across different Mars Global Simulant (MGS-1) concentrations (0, 25, 50, and 75%). In our greenhouse experiment, 75% MGS-1 concentration most significantly inhibited sweet potato growth, storage root biomass, and chlorophyll content. This concentration also elevated plant tissues' H2O2, proline, and ascorbic acid levels. Higher MGS-1 exposures (50 and 75%) notably boosted vital amino acids and sugar groups in the plant's storage roots. Yet, increased MGS-1 concentrations notably diminished the total C: N ratio and elemental composition in both vines and storage roots. In summary, sweet potato exhibited optimal growth, antioxidant properties, yield, and nutrient profiles at 25% MGS-1 exposure, compared to higher concentrations. This study underscores the need for future interventions, like nutrient enhancement and controlled metal accessibility, to render it a suitable plant for space-based studies.
Keywords:
Mars Global Simulant
; Stress tolerance
; Antioxidant
; Amino acids
; Proline
; Sugar classes
1. Introduction
Exploring life beyond our planet requires understanding the limits of life’s adaptability. The recent advances in space research have opened a new chapter in the human exploration of Mars, fueled by governmental and private space initiatives [1,2]. As the ambition to establish a human presence on the Red Planet intensifies, supplying essentials to astronauts will become a daunting task. The logistical hurdles, enormous costs, and inherent risks associated with continual Earth-to-Mars supply make it imperative to consider producing food directly on Martian soil. The soil of Mars, known as regolith, is a unique blend, abundant in both micro- (Cr, Cu, Fe, Mn, Mo, Ni, and Zn) and macro-nutrients (C, Ca, H, K, N, Na, Mg, O, P, and S) that are vital for plant growth [3]. Yet, its high concentrations of salts, specifically sulfates and perchlorates, coupled with heavy metals, render it inhospitable for vegetation. Moreover, the natural properties of Martian soil—scarce nutrient content, the presence of phytotoxic elements, excessive salinity, and inadequate water retention capacity due to the lack of organic matter—indicate that raw Martian regolith is not inherently suitable for farming [4,5,6]. Thus, identifying the optimal soil composition and understanding plant adaptability to Martian conditions is crucial for potential colonization.
A prior study compared the growth of wild plants, crop plants, and legume plants on Lunar and Martian regolith simulants [7]. However, whether the simulants used in the experiment, truly represent actual regolith, remains questionable. Several other researchers have also ventured into studying the impact of Martian regolith on plant development, focusing on enhancing nutrient content and experimenting with various soil combinations. These studies encompass aspects such as seed germination, plant morphology, and biochemistry across diverse plant species [2,8,9,10,11,12,13]. A standout study by Roberto et al. [14] identified specific genes and proteins in Italian ryegrass that respond to stress in Martian soil simulants. However, a comprehensive understanding of the nutritional characteristics under Martian conditions remains uncharted, mainly highlighting the need to explore the dietary criteria for space sustenance.
Sweet potato (Ipomoea batatas (L.) Lam.), recognized as a dietary mainstay, ranks seventh in worldwide production [15]. Its potential as a "space crop" is derived from its impressive nutritional content, efficient water utilization, rapid growth cycle, ease of processing, abundant yields, and extended storage life [16,17,18]. The adaptability of sweet potato is remarkable, with its vines capable of regenerating new plants. Both its roots and tender leaves provide nutritious food sources [19,20]. A prior study demonstrated that, if supplemented with nutrient solutions, Lunar and Martian Regolith Simulants can support the growth of sweet potato [13].
The majority of the Martian regolith simulants were developed around one or two decades ago. Our understanding of the physical and chemical properties of Martian regolith has increased significantly in recent years. In our experiment, we used MGS-1 (Mars Global Simulant-1), a newly developed, high-fidelity mineralogical analog of Martian regolith, that bestows remarkable advancement over the previously developed Martian regolith simulants [21]. Against this backdrop, our study is structured to (i) evaluate the adaptability and regenerative capabilities of sweet potato under various MGS-1 conditions, (ii) understand its stress resilience through biochemical and amino acid analyses, and (iii) determine the nutritional value of its storage roots when grown in MGS-1 environments.
2. Results
2.1. Effect of MGS-1 exposure on growth and photosynthetic pigments
Producing essential consumables using extraterrestrial soil presents a significant challenge in the colonization of Mars. As such, pinpointing a viable method to produce these consumables is paramount. This study evaluated the growth response of sweet potato to various MGS-1 exposure levels to understand its growth, stress adaptation, and nutritional attributes. Compared to the control group, different MGS-1 concentrations significantly hindered sweet potato growth and storage root formation. The most pronounced phytotoxic effects, including stunted growth, diminished leaf size, and leaf margin necrosis, were evident at 75% MGS-1 exposure. Notably, at 100% exposure, sweet potato slips failed to grow and perished within 3 to 5 days post-transplantation (data not shown). Alongside this inhibited growth, there was a substantial reduction in the total storage root biomass by 40, 66.53, and 93.27% at 25, 50, and 75% MGS-1 exposures, respectively, when compared to the control (Table 1 and Figure S1). Indicators like plant height and biomass serve as primary markers of phytotoxicity under abiotic stress conditions. These findings suggest that MGS-1’s phytotoxicity might stem from its sterility, lack of biotic activity, and essential nutrients. Simulated soils from extraterrestrial environments tend to lack vital nutrients for plant growth and development.
The total chlorophyll content in leaf tissue was analyzed to evaluate the physiological response of sweet potato to varying MGS-1 exposure levels. Spectrophotometric results mirrored the growth findings, showing a significant decline in the chlorophyll content of sweet potato leaves due to MGS-1 exposure. After 120 days of treatment, the peak chlorophyll content, 41.89 µg/mg FW, was observed in the control leaves. This content decreased by 9.43, 16.41, and 5.37% at 25% exposure, 20.22, 38.70, and 15.61% at 50% exposure, and 46.43, 64.4, and 36.19% at 75% exposure for plants aged 40, 80, and 120 days, respectively (Figure 1A). Thus, chlorophyll content was not affected much when treated with 25% MGS-1, however chlorophyll content was reduced significantly upon treatment with 50% or 75% MGS-1. This decline in photosynthetic content among plants exposed to Martian regolith could be attributed to MGS-1 exposure inhibiting the enzymes vital for chlorophyll biosynthesis. Additionally, MGS-1 exposure reduced the total leaf area, possibly contributing to a decreased light harvesting rate and subsequent impairment of photosynthesis.
2.2. Effect of MGS-1 exposure on antioxidant system of sweet potato
Proline serves as a recognized physiological marker for environmental stress, reflecting the plant’s level of adaptation to such stressors. In this study, exposure to MGS-1 was found to significantly influence proline accumulation, particularly increasing the proline content in the leaves of sweet potato treated with MGS-1. When assessing the effects of various MGS-1 concentrations on proline accumulation, the most pronounced increases—33.56, 60.83 and 100% were noted at 50% exposure. This was followed by increases of 23.04, 55.31, and 86.17% at 75% exposure and 16.96, 25.89, and 59.82% at 25% exposure in plants aged 40, 80, and 120 days, respectively (Figure 1B). Notably, the proline accumulation trend in sweet potato was directly proportional to both MGS-1 concentrations and plant age, up to 50% exposure and 80 days of treatment. Beyond these points, the values began to decline.
Environmental stress can profoundly disrupt cellular redox balance, leading to the generation of reactive oxygen species (ROS) and subsequent oxidative damage in plants. In this context, the study examined the oxidative harm caused by MGS-1 by gauging H2O2 accumulation. The prominence of the dark brown hue in leaf tissue serves as a direct measure of ROS levels. Control leaf samples exhibited fewer dark brown spots across all three evaluated stages. Conversely, with MGS-1 exposure, there was a progressive increase in H2O2 accumulation in the leaves, correlating with both plant age and the levels of MGS-1. Notably, 120-day-old plants exposed to 75% MGS-1 demonstrated the highest H2O2 accumulation compared to other treatments (Figure 2 A and B).
This study also revealed a concentration-dependent rise in the total ascorbic acid content in storage roots due to MGS-1 exposure after 120 days harvest. The most substantial increase, 40.65%, was observed at 75% exposure, trailed by increments of 36.26% and 15.28% at 50% and 25% MGS-1 exposures, respectively (Table 1). This surge in ascorbic acid might be attributed to sweet potato’s defensive mechanisms, which neutralize the abundant ROS generated under MGS-1 stress.
2.3. Nutritional parameters
Similar to the results observed for stress markers, exposure to MGS-1 notably elevated sugar levels in storage root tissues. Among the MGS-1 concentrations tested, the highest levels of sugar compounds were detected at 75% exposure. Compared to the control, MGS-1 treatment resulted in increases of 382.06, 584.82, and 695.86% in fructose; 0.68, 228.57, and 650% in sucrose; and 16.69, 28.47 and 29.34% in glucose for 25, 50, and 75% MGS-1 exposures, respectively (Table 1). The observed pattern of sugar increase—glucose > fructose > sucrose—in the storage roots suggests that plants treated with MGS-1 might exhibit heightened activity of the acid-soluble invertase enzyme, facilitating the conversion of sucrose into glucose and fructose, compared to the control.
2.4. Amino acid profiling
Our study utilized Partial Least Squares Discriminant Analysis (PLS-DA) to compare amino acid profiles. The score plot distinctly separated control samples from those exposed to various MGS-1 levels, with PC1 accounting for 71.2% of the total variance (Figure 3A). This clear distinction underscores the profound impact of MGS-1 exposure on the amino acid profiles of storage roots. Notably, a 75% MGS-1 exposure greatly influenced the abundance of amino acids such as Gly, Ser, Phe, Lys, and Glu, each having a VIP score greater than 1, highlighting their significance under MGS-1 stress. At 50% MGS-1 exposure, other amino acids, namely Arg, His, Leu, and Pro, similarly exhibited VIP scores above 1 (Figure 4B). These findings suggest that environmental stress, such as that simulated by MGS-1 exposure, can induce increased levels of certain amino acids. Such enhancements have been previously noted to bolster rice plants’ defenses against salt stress by stabilizing metabolism and reducing ROS levels. Detailed data on the differences in amino acid levels due to MGS-1 exposure can be found in Figures S2 and S3. Furthermore, to ascertain the relationship between the control group and various MGS-1 exposures, we validated the amino acid expression patterns using correlation coefficients and hierarchical cluster analysis. The control samples displayed a strong correlation with 25% (0.92) and 50% (0.83) MGS-1 exposures, a moderate correlation with the 75% exposure (0.5), and a weaker correlation (0.31) between the 25% and 75% exposure levels (Figure 3C). Hierarchical cluster analysis further confirmed these correlations, aligning with the findings from the correlation coefficient analysis (Figure 3D). Taken together, these analyses solidify the notion that MGS-1 exposure profoundly influences amino acid composition in storage roots.
2.5. Elemental and total C:N analysis
The inclusion of MGS-1 in the growth medium had marked effects on mineral uptake and accumulation in both vines and storage roots compared to control plants. Under MGS-1 exposure, there was a pronounced increase in the accumulation of various elements such as Al (up to 204.16%), B (up to 82.64%), Fe (up to 1,193.86%), K (up to 66.22%), Mg (up to 339.18%), Mn (up to 946.6%), Si (up to 13.07%), and Zn (up to 104.47%). In contrast, there was a discernible reduction in the accumulation of Na (up to 52.88%) and P (up to 63.97%) in both the vines and storage roots compared to the control. Among the tested MGS-1 concentrations, K and Si reached peak accumulation at 25% exposure. Conversely, higher levels of B, Mg, Mn, and Zn were observed at 50% exposure. When considering bioaccumulation efficiency, vines demonstrated superior element accumulation rates compared to their corresponding storage root samples, irrespective of the MGS-1 treatment (Figure 4). Elements generally play roles in diverse plant metabolic and cellular functions. Hence, the interference of MGS-1 exposure with the uptake of these vital elements could disrupt the plants’ physiological processes, potentially hindering their growth.
In this study, contrary to the results from elemental analysis, MGS-1 exposure considerably diminished the total C:N ratio in both the vines and storage roots of the sweet potato plant. The highest observed C:N ratio (60.06%) was found in the storage roots under control conditions. Nevertheless, the C:N ratio consistently decreased as MGS-1 exposure concentrations increased, regardless of tissue type. Specifically, the C:N ratio fell by 40.82%, 49.25%, and 47.23% in storage roots and by 16%, 17.03%, and 34.35% in vines at 25%, 50%, and 75% MGS-1 exposure, respectively (Table 1).
3. Discussion
Plants have always garnered attention due to how they respond and maintain homeostasis [29]. Duri et al. [30] extensively reviewed the effects of Lunar and Martian regolith simulants to support plant growth. Unlike the Lunar regolith, no ‘real’ sample of Martian regolith has been brought back to Earth. Martian regolith lacks organic matter and contains very high levels of salts and a toxic level of perchlorate [30]. The reactive nitrogen on earth is primarily derived from the mineralization of organic matter [31]. Thus, supplementing Martian Regolith Simulant with organic matter or inorganic fertilizer is necessary to support the growth of crop plants. Our current study also indicates the challenges associated with crop cultivation on Mars. Our findings highlighted the inhibitory effects of MGS-1 on sweet potato growth and overall health and identified a combination of Martian Regolith Simulant and soil that could sustain the growth of sweet potato. However, we would like to point out here that our study was carried out with the assumption that the crop growth on Mars will be conducted in a closed environment that will resemble an Earth-like environment, including light conditions, atmospheric conditions (e.g., temperature and day-night length), and gravity.
The intriguing capability of plants to adapt and respond under varied environmental and growth conditions has always captivated the scientific community, especially botanists and agronomists [32]. Central to this adaptive mechanism is the modulation of photosynthetic pigments, which play a pivotal role in determining a plant’s physiological health and its efficiency in converting light energy into chemical energy [33]. In our study, the assessment of chlorophyll yielded significant insights. Notably, there was a pronounced reduction in total chlorophyll content in plants subjected to MGS-1 exposure. One probable explanation for this decrement could be the nutrient imbalances and the inherent challenges presented by the MGS-1 soil, which might have adversely affected the chloroplast’s function and diminished photosynthetic efficiency. Chlorophyll, being the chief pigment involved in light absorption and subsequent energy conversion, can be seen as an early indicator of stress. As highlighted by Glanz-Idan and Wolf, [34], and further corroborated by the extensive research of Zhou et al. [35], such declines in chlorophyll content are not just isolated events. Nutrient deficiencies, combined with hostile soil parameters, can impair chloroplast integrity, subsequently impacting the efficiency and organization of light-harvesting complexes [36,37]. Understanding these intricacies becomes even more critical when envisioning the challenges of cultivating crops in extraterrestrial settings like Mars [38].
The intricate dynamics of plant responses to stress encompass a spectrum of molecular, physiological, and biochemical adaptations. Delving deeper into this complex interplay, our study revealed compelling revelations regarding the stress markers of sweet potato under MGS-1 exposure. One of the most pronounced observations in this study was the amplified accumulation of proline in the leaves, mirroring findings from many scientific investigations on stress adaptation [39,40]. Proline, often heralded as a quintessential osmoprotectant, showcased an accumulation pattern directly proportional to the increasing concentrations of MGS-1 and the plant’s age. Such accumulations, which have been recurrently documented in various organisms, are emblematic of the plant’s endeavor to stabilize cellular structures and mitigate oxidative imbalances [41,42]. Proline is also well-known as an antioxidant, a metal chelator, and as a cellular signaling molecule [43].
An augmented ROS concentration, while being an innate response to stressors, can be deleterious, orchestrating a cascade of oxidative damages at the cellular level, which, if unchecked, can culminate in cell senescence or even death [44]. In the current experiment, MGS-1 exposure led to significant enhancement in the ROS level (Figure 2A and 2B). A recent transcriptome analysis revealed differential expression of genes implicated in mediating responses to different stresses, including high salt concentration, increased metal concentration, and elevated ROS levels, when model plant Arabidopsis thaliana was grown on diverse Lunar regoliths, that had been brought back to Earth by Apollo space missions [45]. It is important to mention here that both the Lunar and Martian regolith possess high salt and metal contents, hence plants grown directly on these regolith simulants may experience salt and metal stresses [30]. The intricate interactions of cellular defense mechanisms were further exemplified by the surge in ascorbic acid content within sweet potato storage roots as they grappled with escalating MGS-1 exposures. Ascorbic acid is well-known as an enzyme cofactor implicated in photosynthesis and also as an antioxidant [46].
When probing the effects of MGS-1 exposure on sweet potato, our investigation unraveled pronounced alterations in the sugar content within the storage root tissues. This metabolic recalibration, as evidenced by our results, suggests a robust surge in the activity of acid-soluble invertase enzymes, which facilitate the conversion of sucrose into its constituent monosaccharides, glucose, and fructose [47]. Such an elevation in sugar levels, while crucial for energy and carbon skeletons, may also be emblematic of their role as osmolytes, fortifying cells against the duress of stress [48]. However, sugars don’t merely play passive roles; their ascendance could also influence cellular dynamics. They serve as pivotal signaling moieties, orchestrating cellular machinations from modulating gene transcription to fine-tuning enzymatic cascades [49].
Venturing beyond sugars, our study delved deep into the amino acid tapestry of storage roots, unearthing profound shifts in their profiles under MGS-1’s influence. Amino acids are intricate players in plant physiology, especially during stress [50]. Our observations revealed that certain amino acids, likely acting as sentinel molecules, showcased heightened fluctuations, potentially encapsulating the plant’s strategic metabolic response to the rigors of MGS-1. Furthermore, amino acids like proline, glutamate, and glycine have been extolled for their role as osmoprotectants, safeguarding plants by helping maintain osmotic equilibrium amidst environmental adversities [51].
In our exploration, the inclusion of MGS-1 into the growth medium unveiled profound shifts in mineral dynamics, with both vines and storage roots undergoing marked changes in elemental uptake. Minerals and elements, often dubbed as plants’ lifeblood, anchor many physiological and biochemical pathways [52]. Thus, the shifts induced by MGS-1 exposure could ripple through these pathways, reshaping the plant’s metabolic and growth trajectories. Conversely, a particularly intriguing observation was the MGS-1-mediated modulation of the C:N ratio. Traditionally, a perturbed C:N ratio is emblematic of plants grappling with environmental duress or traversing nutrient-scarce landscapes [53]. This dynamic equilibrium between carbon and nitrogen is more than just a mere ratio; it is a window into the plant’s metabolic mechanism, offering glimpses of processes ranging from carbon fixation to nitrogen assimilation [54]. Moreover, the observed fluctuations in the C:N ratio, a barometer of plant metabolic health, warrant a deeper introspection. Such fluctuations could be symptomatic of broader metabolic recalibrations. These might encompass shifts in foundational pathways, such as the Calvin cycle, central to carbon fixation and nitrogen assimilation processes, pivotal for protein synthesis and growth [55,56].
4. Materials and Methods
4.1. Mars Global Simulant (MGS-1)
To assess the impact of Martian regolith on sweet potato growth, tolerance, and nutritional parameters, the Mars Global Simulant (MGS-1) was chosen for this study. The newly formulated MGS-1 soil was preferred because it has been modified to be free of hazardous components, distinguishing it from other commercially available Mars simulants [21]. The simulant was procured from the Exolith Laboratory located in Florida, USA. All analytical-grade chemicals used in this research were sourced from Sigma-Aldrich.
4.2. Experimental design
For our greenhouse study at the Bioplex facility of West Virginia State University, we procured and acclimatized healthy, uniformly-sized sweet potato slips of the Beauregard variety. After about two weeks, when they had developed new roots, we initiated a pot experiment using varying combinations of MGS-1 and potting mix: purely potting mix as a control, and then blends of 25%, 50%, and 75% MGS-1 exposure based on preliminary phytotoxicity screenings. Throughout the study, we maintained conditions at 28–36°C during the day and 20–28°C at night, with a 16-hour light/8-hour dark cycle and a consistent soil moisture level of 65–75%. We harvested plants at intervals of 40, 80, and 120 days, with the storage root’s total fresh weight (FW) measured electronically 120 days post-harvest.
4.1. Estimation of total chlorophyll
We quantified the total chlorophyll content in sweet potato leaf tissues following the method described by Strain et al. [22], albeit with slight modifications. Specifically, we collected 50 mg of fresh leaf tissues from each treatment. These were then homogenized in a solution of 80% (v/v) ice-cold acetone fortified with 1 mM KOH, using three times the volume of the solution to the tissue weight. After homogenizing, the mixture was centrifuged at 16,000 g for 2 minutes. The supernatant’s total chlorophyll content was subsequently determined using a spectrophotometer, measuring at wavelengths of 663 nm for chlorophyll a and 646 nm for chlorophyll b. The results are presented in mg/g FW.
4.3. Determination of proline content
The proline content was quantified using the acid ninhydrin method, referencing the procedure outlined by Shabnam et al. [23], though with minor modifications. We homogenized approximately 0.5 g of leaf tissues in 300 µl of ice-cold water. This mixture was then centrifuged at 16,000 g for 5 minutes. The resulting supernatants were carefully transferred to a new tube. We added 1.25% acid ninhydrin to these supernatants and then incubated the mixture in a water bath at 100°C for 30 minutes. After incubation, the reaction mixture was allowed to cool to room temperature. Using a spectrophotometer, the absorbance of the mixture was measured at 520 nm. By referencing a proline standard curve, we determined the proline concentration. The final results are expressed in terms of µmol/g FW.
4.4. 3,3’-diaminobenzidine (DAB) staining
The accumulation of ROS in sweet potato leaf samples, induced by MGS-1, was determined by measuring H2O2 levels using the 3,3′-diaminobenzidine (DAB) method [24]. Fresh leaf tissues were initially submerged in a freshly prepared DAB staining solution, which consisted of 1mg/ml of DAB (adjusted to pH 3.0) with the addition of 0.05% v/v Tween 20 and 2.5 ml of 200 mM Na2HPO4. These tissues were then subjected to a vacuum infiltration process at pressures ranging from 300 to 400 psi for 5 minutes. Following this, both the staining solution and the leaf tissues were incubated in dark conditions for 4 hours, with a gentle shaking at 90 rpm. Post-incubation, the staining solution was removed, and the samples were destained using a bleaching solution, a mixture of ethanol, acetic acid, and glycerol in a 3:1:1 ratio. This destaining process continued for 15 minutes at 95°C. The decolorized leaf samples were then examined under a light microscope (Zeiss Stemi 2000 – C, Germany). The presence of brown spots on these samples indicated the accumulation of H2O2.
4.5. Sample preparation for GC-MS and LC-MS
After 120 days of treatment, the control and MGS-1-treated storage root samples were isolated. They were then dried at 80°C for 24 hours. Subsequently, these oven-dried storage root samples were processed for various instrumental analyses—estimation of ascorbic acid and sugar classes. Storage root samples in powdered form were extracted using distilled water, then centrifuged and divided into two equal portions. One portion was designated for sugar analysis, while the other was allocated for amino acid quantitation. For the sugar analysis, the extracted supernatants were dried. They were then derivatized: initially with 75 μl of methoxyamine hydrochloride (40 mg/ml in pyridine) for 60 minutes at 50°C and subsequently with 75 μl of MSTFA+1%TMCS at 70°C for 120 minutes, with a further 2-hour incubation at room temperature. Before derivatization, an internal standard (10 μl of hentriacontanoic acid at 1 mg/mL concentration) was introduced to each sample. The chromatograms were obtained using a GC-MS system (Agilent Inc, CA, USA) outfitted with a ZB-5MS capillary column (60m×0.32mm I.D. and 0.25μm film thickness) from Phenomenex, CA, USA. During the process, a 1 μl aliquot was injected with a split ratio of 10:1. The Mass Hunter Quantitative Analysis B.08.00 software (Agilent Inc., CA, USA) was employed to evaluate the target peaks, as described by Salih et al. [25]. Authentic standards produced calibration curves spanning the range of 0.5 – 50 μg/ml. To ensure consistent comparisons across samples, all data were adjusted based on the internal standard in each chromatogram and the weight of the sample.
4.6. Amino acid profiling
The amino acid profile of the storage roots, under various MGS-1 exposure levels, was analyzed using Liquid Chromatography-Mass Spectrometry (LC-MS). The analysis employed the Vanquish system (TSQ Altis LC-MS/MS system, Thermo Scientific), utilizing a Hypersil Gold column (2.1 x 150 mm, 1.9 μ). The set flow rate was 600 μL/min. The mobile phases comprised 0.1% formic acid in water (A) and 0.1% in acetonitrile (B). The gradient was established as follows: from 0 to 0.5 minutes – 0% B, from 0.5 to 3.5 minutes – 60% B, from 3.5 to 5.5 minutes – 100% B, and from 5.5 to 7.5 minutes – 0% B. The injection volume was set at 1 μl, and the column chamber temperature was maintained at 500°C. Data collection was carried out in a positive SRM mode with a voltage of 3500V. Peak integration and quantification were conducted using the Thermo TraceFinder (4.1) software, as detailed by Qiu et al. [26]
4.7. Total C:N
The dried samples from both vines and storage roots were analyzed for total carbon (C) and nitrogen (N) content. The estimation was conducted using the Dumas method, as outlined in EN 13654-2. A Skalar Primacs SNC 100 analyzer (Skalar, the Netherlands) was utilized for this purpose, as documented by Analytical, [27]. The C:N ratio of the samples was calculated from the obtained values of total C and N.
4.8. Elemental analysis
Elemental analysis was performed as described by Ali et al. [28], with slight modifications. Approximately, 500 mg of powdered plant samples were measured and then subjected to acid digestion using a microwave digester (MARS 6, CEM Corporation, Matthews, NC, USA). The digested mixture was subsequently filtered and diluted using metal-free HPLC-grade water. The concentrations of the targeted elements in the samples were determined using Inductively Coupled Plasma – Mass Spectrometry (ICP – MS, X – series II, Thermo, U.S.).
4.9. Statistical analysis
All experiments were carried out in triplicate (n = 3). Data were subjected to one-way ANOVA, and mean values were compared using Tukey’s test. Analyses were performed using the SPSS software (version 20, SPSS Inc., www.spss.com).
5. Conclusions
The study explored the reactions of sweet potato when exposed to MGS-1, a substance simulating Martian soil. The findings revealed myriad physiological, biochemical, and molecular alterations in the plant. Interestingly, while a 25% MGS-1 concentration led to optimal plant growth and resilience, higher concentrations, especially at 75%, severely restricted growth and altered the plant’s biochemical makeup. Martian regolith is devoid of organic matter and contains a very high level of perchlorate, which is toxic for plant growth. This poses unique challenges in growing crops on Martian regolith [57]. These insights not only shed light on the inherent adaptive strategies of the plant but also emphasize the need for innovative agricultural methods on Mars. Potential solutions include enriching the soil with organic matter, introducing nitrogen-fixing bacteria, and leveraging decomposing microbes for nutrient recycling from plant and human waste [11]. For example, in a recent study [2], Martian regolith simulant supplemented with compost (MMS-1 Mojave Mars Simulant: compost 30:70, v:v) supported optimal photosynthesis, water use efficiency, quality traits, and overall crop performance in lettuce. In a recent study, supplementing Mars Regolith Simulant MMS-1 with green compost significantly enhanced the physicochemical properties of the growing substrate and improved the performance of potato plants [58]. A recent study elucidated molecular changes at transcriptional and proteomic levels in Italian ryegrass grown on the Martian regolith simulant MMS-1. This study exhibited changes in transcript and/or protein accumulation levels in genes involved in the phenylpropanoid pathway, translation, DNA methylation, and mediating stress responses [14]. As space colonization progresses, it becomes imperative to delve deeper into the molecular and genetic dynamics influencing crop plants’ adaptation and nutrition in Martian-like conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Effect of different concentrations of MGS1 exposure on A) growth and B) storage root and C) storage root fresh weight of I. batatas. Results are expressed as mean of triplicates ± SD. **P < 0.01, and ***P < 0.001. Figure S2: Effect of different MGS – 1 on amino acid profiling. Results are expressed as the means of three replicates ± SD and the asterisks indicate the significance at *P < 0.05, **P < 0.01, and ***P < 0.001 compared to the control according to the independent samples t test. Figure S3: Box-Whisker plot for the significantly different amino acids for the different exposure of MGS – 1. The significantly different amino acids obtained from ANOVA and post-hoc analysis. .
Author Contributions
UR and PN: conceptualization, project administration, funding acquisition, manuscript editing, supervision, KC: experimentation, data analysis, writing-original manuscript, MC: manuscript editing, proof reading, PS and HY: data analysis and experimental support. All authors have read and approved the final manuscript.
Funding
The authors thank NASA WV EPSCoR (NASA-EPSCoR-R3-21-0051) for their financial support.
Acknowledgments
The authors thank Roy J. Carver, Biotechnology Center, the University of Illinois Urbana-Champaign, for providing GC and LC–MS analysis. The authors would like to acknowledge the support of the Cornell Nutrient Analysis Laboratory, Cornell University, for offering ICP – MS analysis.
Conflicts of Interest
The authors have declared no conflict of interest.
References
- Rainwater, R.; Mukherjee, A. The legume-rhizobia symbiosis can be supported on Mars soil simulants. Plos one 2021, 16 (12), e0259957. [CrossRef]
- Duri, L. G.; El-Nakhel, C.; Caporale, A. G.; Ciriello, M.; Graziani, G.; Pannico, A.; Palladino, M.; Ritieni, A.; De Pascale, S.; Vingiani, S. Mars regolith simulant ameliorated by compost as in situ cultivation substrate improves lettuce growth and nutritional aspects. Plants 2020, 9 (5), 628. [CrossRef]
- McGuire, N. M.; Univerity, R. P. E. R. A.; DeMattio, D. Project HOME: Hydroponic Operations for Mars Exploration. 2020.
- Rosa Polonia, R. A.; Hufendick, B. T. Project HOME (Hydroponic Operations for Mars Exploration). 2020.
- Carr, M.; Head, J. Martian surface/near-surface water inventory: Sources, sinks, and changes with time. Geophysical Research Letters 2015, 42 (3), 726-732. [CrossRef]
- Plaut, J. J.; Picardi, G.; Safaeinili, A.; Ivanov, A. B.; Milkovich, S. M.; Cicchetti, A.; Kofman, W.; Mouginot, J.; Farrell, W. M.; Phillips, R. J. Subsurface radar sounding of the south polar layered deposits of Mars. science 2007, 316 (5821), 92-95. [CrossRef]
- Wamelink, W. W. G.; Frissel, Y. J.; Krijnen, J. H. W.; Verwoert, R. M.; Goedhart, W. P. Can Plants Grow on Mars and the Moon: A Growth Experiment on Mars and Moon Soil Simulants. PLoS ONE 2014, 9 (8), e103138. [CrossRef]
- DeMattio, D.; McGuire, N.; Rosa Polonia, R. A.; Hufendick, B. T. Project HOME hydroponic operations for Mars exploration. Beyond: Undergraduate Research Journal 2020, 4 (1), 5.
- Pandith, J. A.; Neekhra, S.; Ahmad, S.; Sheikh, R. A. Recent developments in space food for exploration missions: A review. Life Sciences in Space Research 2023, 36, 123-134. [CrossRef]
- Eichler, A.; Hadland, N.; Pickett, D.; Masaitis, D.; Handy, D.; Perez, A.; Batcheldor, D.; Wheeler, B.; Palmer, A. Challenging the agricultural viability of martian regolith simulants. Icarus 2021, 354, 114022. [CrossRef]
- Kasiviswanathan, P.; Swanner, E. D.; Halverson, L. J.; Vijayapalani, P. Farming on Mars: Treatment of basaltic regolith soil and briny water simulants sustains plant growth. Plos one 2022, 17 (8), e0272209. [CrossRef]
- Wamelink, G.; Frissel, J.; Krijnen, W.; Verwoert, M. Crop growth and viability of seeds on Mars and Moon soil simulants. Terraforming Mars 2021, 313-329. [CrossRef]
- Mortley, D. G.; Aglan, H. A.; Bonsi, C. K.; Hill, W. A. Growth of Sweetpotato in Lunar and Mars Simulants. In SAE Technical Paper, 2000.
- Berni, R.; Leclercq, C. C.; Roux, P.; Hausman, J. F.; Renaut, J.; Guerriero, G. A molecular study of Italian ryegrass grown on Martian regolith simulant. Science of the Total Environment 2023, 854. [CrossRef]
- Behera, S.; Chauhan, V. B. S.; Pati, K.; Bansode, V.; Nedunchezhiyan, M.; Verma, A. K.; Monalisa, K.; Naik, P. K.; Naik, S. K. Biology and biotechnological aspect of sweet potato (Ipomoea batatas L.): A commercially important tuber crop. Planta 2022, 256 (2), 40. [CrossRef]
- Kumar, V.; Singh, S.; Jakhwal, R.; Singh, B.; Tomar, H. SPACE FARMING: NEED FOR FRESH VEGETABLE CROP. [CrossRef]
- Laveriano-Santos, E. P.; López-Yerena, A.; Jaime-Rodríguez, C.; González-Coria, J.; Lamuela-Raventós, R. M.; Vallverdú-Queralt, A.; Romanyà, J.; Pérez, M. Sweet Potato Is Not Simply an Abundant Food Crop: A Comprehensive Review of Its Phytochemical Constituents, Biological Activities, and the Effects of Processing. Antioxidants (Basel) 2022, 11 (9). [CrossRef]
- Wilson, C. D.; Pace, R. D.; Bromfield, E.; Jones, G.; Lu, J. Y. Sweet potato in a vegetarian menu plan for NASA’s Advanced Life Support Program. Life Support Biosph Sci 1998, 5 (3), 347-351. From NASA.
- Bovell-Benjamin, A. C. Sweet potato: a review of its past, present, and future role in human nutrition. Advances in food and nutrition research 2007, 52, 1-59. [CrossRef]
- Tedesco, D.; de Almeida Moreira, B. R.; Júnior, M. R. B.; Maeda, M.; da Silva, R. P. Sustainable management of sweet potatoes: A review on practices, strategies, and opportunities in nutrition-sensitive agriculture, energy security, and quality of life. Agricultural Systems 2023, 210, 103693. [CrossRef]
- Cannon, K. M.; Britt, D. T.; Smith, T. M.; Fritsche, R. F.; Batcheldor, D. Mars global simulant MGS-1: A Rocknest-based open standard for basaltic martian regolith simulants. Icarus 2019, 317, 470-478. [CrossRef]
- Strain, H. H.; Cope, B. T.; Svec, W. A. [42] Analytical procedures for the isolation, identification, estimation, and investigation of the chlorophylls. In Methods in enzymology, Vol. 23; Elsevier, 1971; pp 452-476. [CrossRef]
- Shabnam, N.; Tripathi, I.; Sharmila, P.; Pardha-Saradhi, P. A rapid, ideal, and eco-friendlier protocol for quantifying proline. Protoplasma 2016, 253, 1577-1582. [CrossRef]
- Daudi, A.; O’Brien, J. A. Detection of hydrogen peroxide by DAB staining in Arabidopsis leaves. Bio-protocol 2012, 2 (18), e263-e263. [CrossRef]
- Salih, A. M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H. O.; Alabdallah, N. M.; Alansi, S.; Alshameri, A. Mass propagation of Juniperus procera Hoechst. Ex Endl. From seedling and screening of bioactive compounds in shoot and callus extract. BMC Plant Biology 2021, 21, 1-13. [CrossRef]
- Lee, J. Y.; Harney, D. J.; Teo, J. D.; Kwok, J. B.; Sutherland, G. T.; Larance, M.; Don, A. S. The major TMEM106B dementia risk allele affects TMEM106B protein levels, fibril formation, and myelin lipid homeostasis in the ageing human hippocampus. Molecular Neurodegeneration 2023, 18 (1), 63. [CrossRef]
- Vandecasteele, B.; Hofkens, M.; De Zaeytijd, J.; Visser, R.; Melis, P. Towards environmentally sustainable growing media for strawberry cultivation: Effect of biochar and fertigation on circular use of nutrients. Agricultural Water Management 2023, 284, 108361. [CrossRef]
- Mawari, G.; Kumar, N.; Sarkar, S.; Daga, M. K.; Singh, M. M.; Joshi, T. K.; Khan, N. A. Heavy Metal Accumulation in Fruits and Vegetables and Human Health Risk Assessment: Findings From Maharashtra, India. Environ Health Insights 2022, 16, 11786302221119151. From NLM. [CrossRef]
- Brooker, R.; Brown, L. K.; George, T. S.; Pakeman, R. J.; Palmer, S.; Ramsay, L.; Schöb, C.; Schurch, N.; Wilkinson, M. J. Active and adaptive plasticity in a changing climate. Trends in Plant Science 2022. [CrossRef]
- Duri, L. G.; Caporale, A. G.; Rouphael, Y.; Vingiani, S.; Palladino, M.; De Pascale, S.; Adamo, P. The Potential for Lunar and Martian Regolith Simulants to Sustain Plant Growth: A Multidisciplinary Overview. Frontiers in Astronomy and Space Sciences 2022, 8. [CrossRef]
- Stevens, J. C.; Manning, P.; Berg, D. V. J. L. L.; Graaf, D. C. C. M.; Wamelink, W. G. W.; Boxman, W. A.; Bleeker, A.; Vergeer, P.; Arroniz-Crespo, M.; Limpens, J.; et al. Ecosystem responses to reduced and oxidised nitrogen inputs in European terrestrial habitats. Environmental Pollution 2011, 159 (3), 665-676. [CrossRef]
- Mareri, L.; Parrotta, L.; Cai, G. Environmental stress and plants. MDPI: 2022; Vol. 23, p 5416. [CrossRef]
- Therby-Vale, R.; Lacombe, B.; Rhee, S. Y.; Nussaume, L.; Rouached, H. Mineral nutrient signaling controls photosynthesis: Focus on iron deficiency-induced chlorosis. Trends in plant science 2022, 27 (5), 502-509. [CrossRef]
- Glanz-Idan, N.; Wolf, S. Upregulation of photosynthesis in mineral nutrition-deficient tomato plants by reduced source-to-sink ratio. Plant signaling & behavior 2020, 15 (2), 1712543. [CrossRef]
- Zhou, J.; Yuan, W.; Di, B.; Zhang, G.; Zhu, J.; Zhou, P.; Ding, T.; Qian, J. Relationship among Electrical Signals, Chlorophyll Fluorescence, and Root Vitality of Strawberry Seedlings under Drought Stress. Agronomy 2022, 12 (6), 1428. [CrossRef]
- Szechyńska-Hebda, M.; Ghalami, R. Z.; Kamran, M.; Van Breusegem, F.; Karpiński, S. To be or not to be? Are Reactive oxygen species, antioxidants, and stress signalling universal determinants of life or death? Cells 2022, 11 (24), 4105. [CrossRef]
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I. K. How to cope with the challenges of environmental stresses in the era of global climate change: An update on ROS stave off in plants. International Journal of Molecular Sciences 2022, 23 (4), 1995. [CrossRef]
- Schuerger, A. C.; Wheeler, R. M.; Levine, H. G.; Paul, A.-L.; Ferl, R. J. Vegetable Health Challenges in Extraterrestrial Production. In Handbook of Vegetable and Herb Diseases, Springer, 2022; pp 1-49. [CrossRef]
- Nie, M.; Ning, N.; Chen, J.; Zhang, Y.; Li, S.; Zheng, L.; Zhang, H. Melatonin enhances salt tolerance in sorghum by modulating photosynthetic performance, osmoregulation, antioxidant defense, and ion homeostasis. Open Life Sciences 2023, 18 (1), 20220734. [CrossRef]
- Hafeez, A.; Rasheed, R.; Ashraf, M. A.; Qureshi, F. F.; Hussain, I.; Iqbal, M. Effect of heavy metals on growth, physiological and biochemical responses of plants. In Plants and their Interaction to Environmental Pollution, Elsevier, 2023; pp 139-159.
- Phillips, A. L. Interrogating the multi-stress tolerance of Oryza australiensis using novel genomic and phenomic strategies. 2022.
- Hashem, H. A.; Khalil, R. Insight into the Interaction of Strigolactones, Abscisic Acid, and Reactive Oxygen Species Signals. In Reactive Oxygen Species: Prospects in Plant Metabolism, Springer, 2023; pp 179-211.
- Hayat, S.; Hayat, Q.; Alyemeni, M. N.; Wani, A. S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signaling & Behavior 2012, 7 (11), 1456-1466. PMC. [CrossRef]
- Zentgraf, U.; Andrade-Galan, A. G.; Bieker, S. Specificity of H2O2 signaling in leaf senescence: is the ratio of H2O2 contents in different cellular compartments sensed in Arabidopsis plants? Cellular & Molecular Biology Letters 2022, 27 (1), 1-19. [CrossRef]
- Paul, A.-L.; Elardo, M. S.; Ferl, R. Plants grown in Apollo lunar regolith present stress-associated transcriptomes that inform prospects for lunar exploration. Communications Biology 2022, 5 (1). [CrossRef]
- Smirnoff, N.; Wheeler, L. G. Ascorbic Acid in Plants: Biosynthesis and Function. Critical Reviews in Biochemistry and Molecular Biology 2000, 35 (4), 291-314. [CrossRef]
- Li, Z.; Duan, S.; Lu, B.; Yang, C.; Ding, H.; Shen, H. Spraying alginate oligosaccharide improves photosynthetic performance and sugar accumulation in citrus by regulating antioxidant system and related gene expression. Frontiers in Plant Science 2023, 13, 1108848. [CrossRef]
- Faryal, S.; Ullah, R.; Khan, M. N.; Ali, B.; Hafeez, A.; Jaremko, M.; Qureshi, K. A. Thiourea-capped nanoapatites amplify osmotic stress tolerance in Zea mays L. by conserving photosynthetic pigments, osmolytes biosynthesis and antioxidant biosystems. Molecules 2022, 27 (18), 5744. [CrossRef]
- Khanna, K.; Ohri, P.; Bhardwaj, R. Decoding sugar regulation and homeostasis in plants: cracking functional roles under stresses. Journal of Plant Growth Regulation 2023, 42 (8), 4797-4817. [CrossRef]
- Liao, H.-S.; Chung, Y.-H.; Hsieh, M.-H. Glutamate: A multifunctional amino acid in plants. Plant Science 2022, 318, 111238. [CrossRef]
- Kocaman, A. Combined interactions of amino acids and organic acids in heavy metal binding in plants. Plant signaling & behavior 2023, 18 (1), 2064072. [CrossRef]
- Klem, K.; Oravec, M.; Holub, P.; Šimor, J.; Findurová, H.; Surá, K.; Veselá, B.; Hodaňová, P.; Jansen, M.; Urban, O. Interactive effects of nitrogen, UV and PAR on barley morphology and biochemistry are associated with the leaf C: N balance. Plant Physiology and Biochemistry 2022, 172, 111-124. [CrossRef]
- Zhong, Y.; Liu, J.; Jia, X.; Tang, Z.; Shangguan, Z.; Wang, R.; Yan, W. Environmental stress-discriminatory taxa are associated with high C and N cycling functional potentials in dryland grasslands. Science of The Total Environment 2022, 817, 152991. [CrossRef]
- Gu, X.; Leng, J.; Zhu, J.; Zhang, K.; Zhao, J.; Wu, P.; Xing, Q.; Tang, K.; Li, X.; Hu, B. Influence mechanism of C/N ratio on heterotrophic nitrification-aerobic denitrification process. Bioresource Technology 2022, 343, 126116. [CrossRef]
- Joos, L.; De Tender, C. Soil under stress: The importance of soil life and how it is influenced by (micro) plastic pollution. Computational and structural biotechnology journal 2022, 20, 1554-1566. [CrossRef]
- Li, J.; Liu, X.; Xu, L.; Li, W.; Yao, Q.; Yin, X.; Wang, Q.; Tan, W.; Xing, W.; Liu, D. Low nitrogen stress-induced transcriptome changes revealed the molecular response and tolerance characteristics in maintaining the C/N balance of sugar beet (Beta vulgaris L.). Frontiers in Plant Science 2023, 14, 1164151. [CrossRef]
- Oze, C.; Beisel, J.; Dabsys, E.; Dall, J.; North, G.; Scott, A.; Lopez, M. A.; Holmes, R.; Fendorf, S. Perchlorate and Agriculture on Mars. Soil Systems 2021, 5 (3), 37. [CrossRef]
- Caporale, G. A.; Paradiso, R.; Liuzzi, G.; Palladino, M.; Amitrano, C.; Arena, C.; Arouna, N.; Verrillo, M.; Cozzolino, V.; Pascale, D. S.; et al. Green compost amendment improves potato plant performance on Mars regolith simulant as substrate for cultivation in space. Plant and Soil 2023, 486 (1-2), 217-233. [CrossRef]
Figure 1.
Effect of different concentrations of MGS-1 exposure on A) total chlorophyll and B) proline of I. batatas leaves. Results are expressed as means of replicate ± SE, where ‘*’ indicates statistical significance (P<0.05), ‘**’ indicates statistical significance (P<0.01) and ‘***’ indicates statistical significance (P<0.001) according to Tukey’s test.
Figure 1.
Effect of different concentrations of MGS-1 exposure on A) total chlorophyll and B) proline of I. batatas leaves. Results are expressed as means of replicate ± SE, where ‘*’ indicates statistical significance (P<0.05), ‘**’ indicates statistical significance (P<0.01) and ‘***’ indicates statistical significance (P<0.001) according to Tukey’s test.
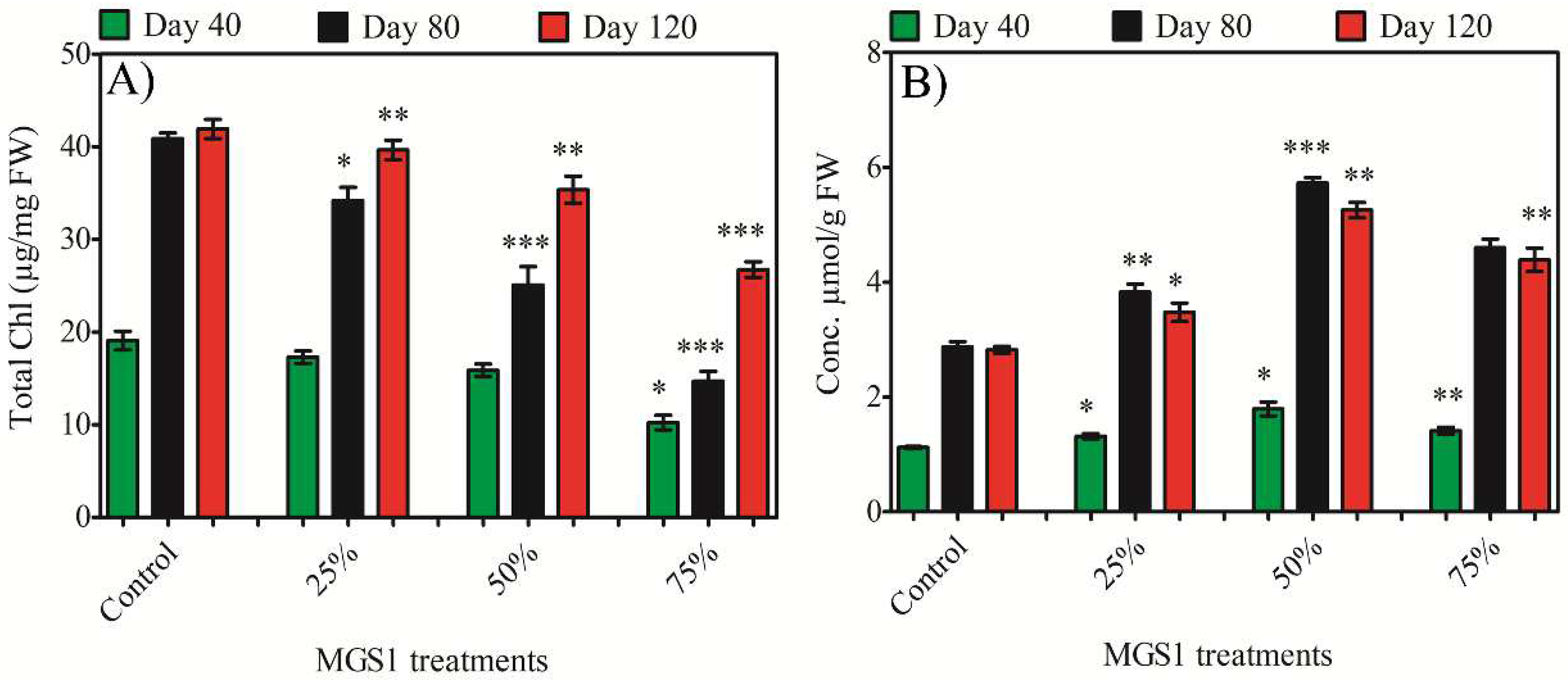
Figure 2.
Effect of different concentrations of MGS-1 exposure on H2O2 accumulation in I. batatas leaves (A) and microscopic images (B) of H2O2 accumulation. i) control, ii) 25%, iii) 50%, and iii) 75% of MGS1 exposure.
Figure 2.
Effect of different concentrations of MGS-1 exposure on H2O2 accumulation in I. batatas leaves (A) and microscopic images (B) of H2O2 accumulation. i) control, ii) 25%, iii) 50%, and iii) 75% of MGS1 exposure.

Figure 3.
Amino acid profiling of storage root tissues grown under control and different level of MGS-1 exposure. A) PLS-DA score plot, B) VIP plot, C) correlation plot analysis, and D) hierarchical cluster analysis.
Figure 3.
Amino acid profiling of storage root tissues grown under control and different level of MGS-1 exposure. A) PLS-DA score plot, B) VIP plot, C) correlation plot analysis, and D) hierarchical cluster analysis.
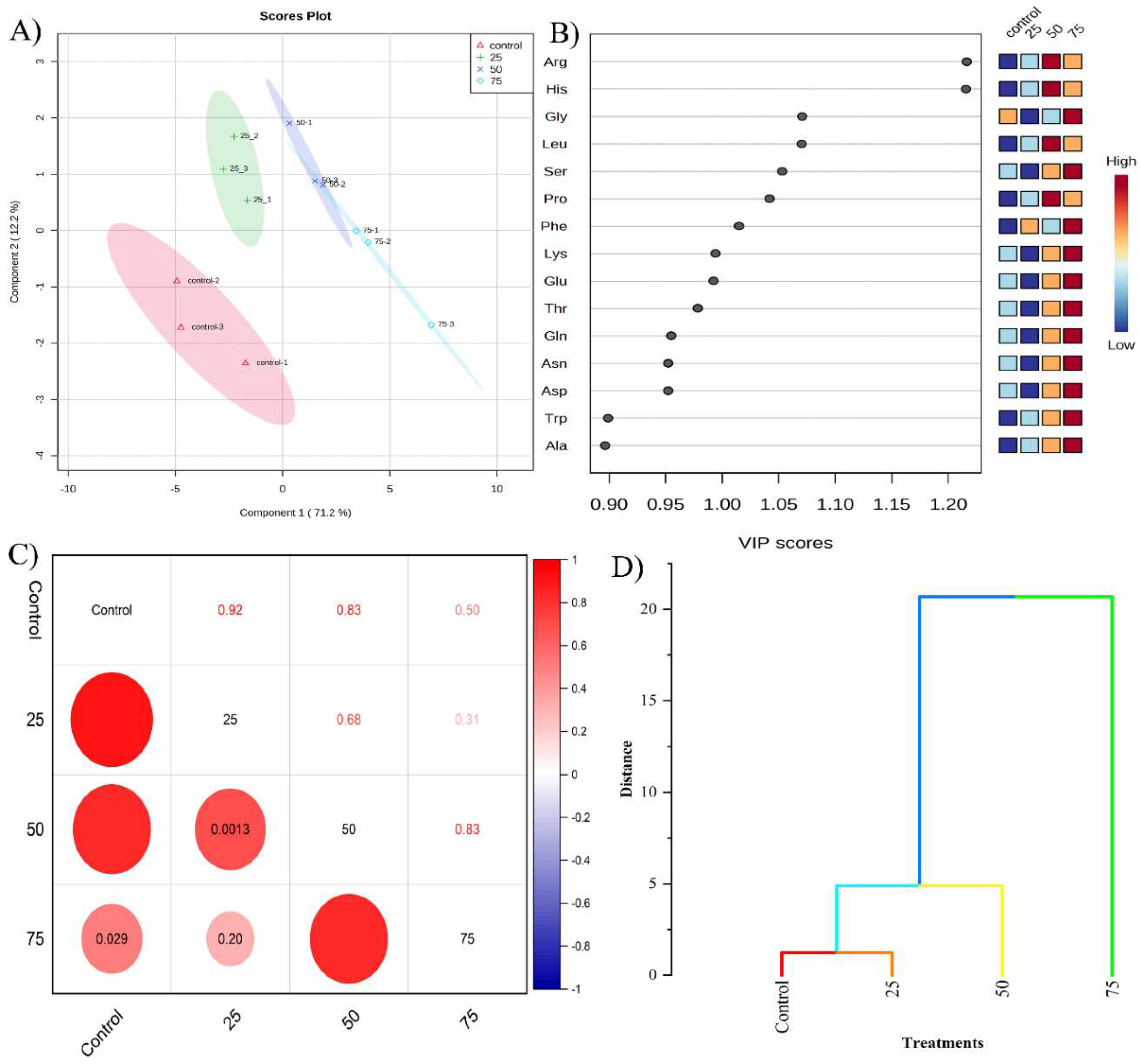
Figure 4.
Effect of different concentrations of MGS-1 exposure on the elemental accumulation in vines and storage roots of I. batatas plants A) Al, B) B, C) Fe, D) K, E) Mg, F) Mn, G) Na, H) P, I) Si, and J) Zn. Results are expressed as means of replicate ± SE, where ‘*’ indicates statistical significance (P<0.05), ‘**’ indicates statistical significance (P<0.01) and ‘***’ indicates statistical significance (P<0.001) according to Tukey’s test.
Figure 4.
Effect of different concentrations of MGS-1 exposure on the elemental accumulation in vines and storage roots of I. batatas plants A) Al, B) B, C) Fe, D) K, E) Mg, F) Mn, G) Na, H) P, I) Si, and J) Zn. Results are expressed as means of replicate ± SE, where ‘*’ indicates statistical significance (P<0.05), ‘**’ indicates statistical significance (P<0.01) and ‘***’ indicates statistical significance (P<0.001) according to Tukey’s test.
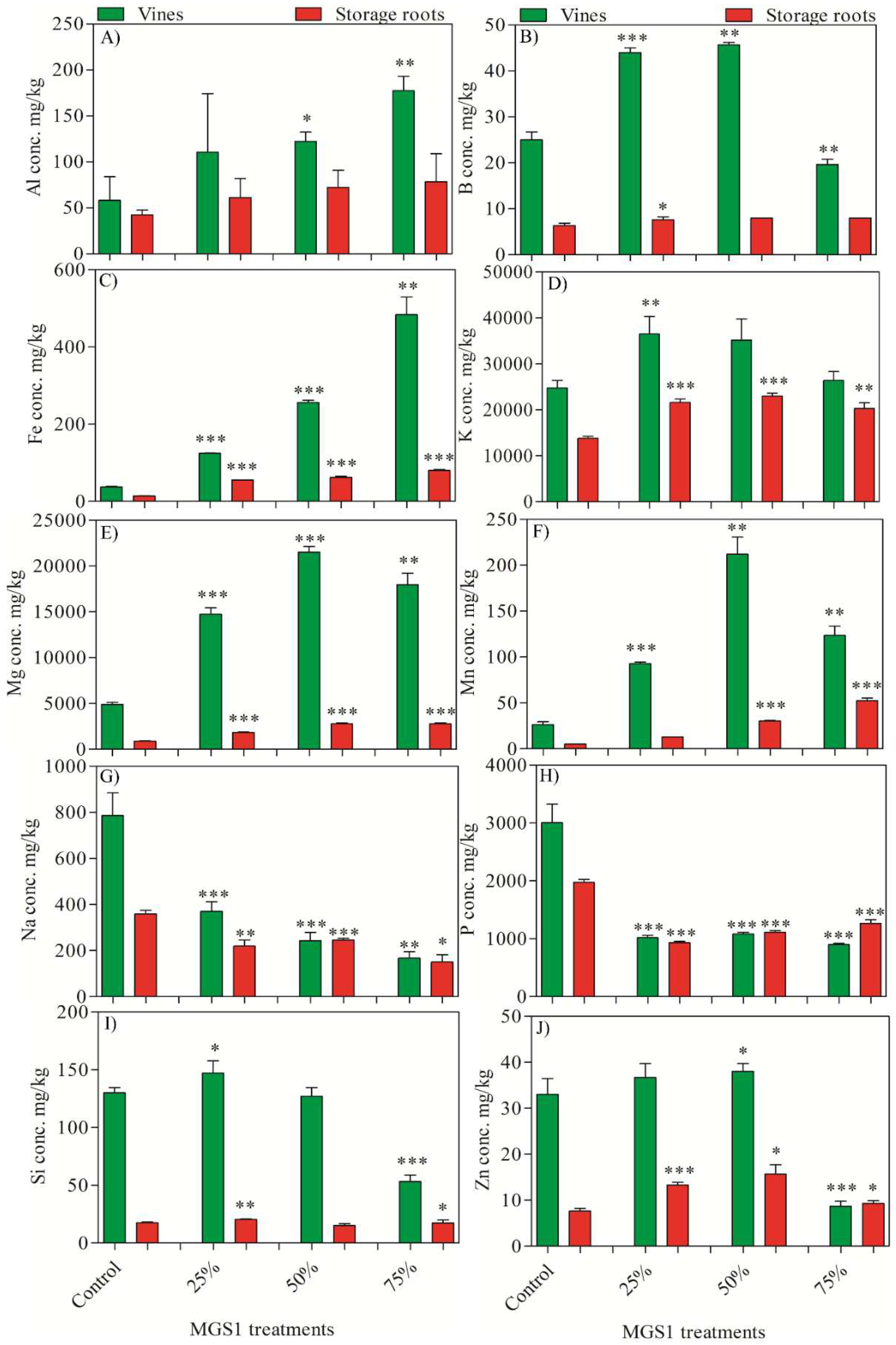

Table 1.
Effect of different concentrations of MGS-1 exposure on total sugar classes and C: N ratio in I. batatas.
Table 1.
Effect of different concentrations of MGS-1 exposure on total sugar classes and C: N ratio in I. batatas.
| Treatments | Storage root biomass (gm/plant) | Ascorbic acid (µg/mg DW) |
Sugar parameters (µg/mg DW) | Total C:N ratio (%) | |||
|---|---|---|---|---|---|---|---|
| Fructose | Sucrose | Glucose | Vines | SR | |||
| Control | 99±11.16 | 0.615±0.04 | 0.145±0.03 | 0.014±0.001 | 10.36±0.72 | 21.25±0.34 | 60.06±0.97 |
| 25% of MGS-1 | 59.4±11.64** | 0.709±0.06 | 0.699±0.04*** | 0.015±0.006 | 12.09±1.02 | 17.85±0.38*** | 35.54±0.18*** |
| 50% of MGS-1 | 33.13±6.66** | 0.838±0.14 | 0.993±0.03*** | 0.046±0.002*** | 13.31±0.46** | 17.63±0.84** | 30.48±0.04*** |
| 75% of MGS-1 | 6.66±2.30** | 0.865±0.08* | 1.154±0.01*** | 0.105±0.006*** | 13.4±0.77** | 13.95±0.13*** | 31.69±0.48*** |
Note: DW – dry weight, SR – Storage roots. Results are expressed as means of replicate ± SE, where ‘*’ indicates statistical significance (P<0.05), ‘**’ indicates statistical significance (P<0.01) and ‘***’ indicates statistical significance (P<0.001) according to Tukey’s test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



