Submitted:
15 November 2023
Posted:
16 November 2023
You are already at the latest version
Abstract
Insulin icodec of Novo Nordisk is a novel long-acting insulin analogue that exhibits an extended duration of action, providing a promising treatment option for individuals with diabetes. It has been reported that the incorporation of fatty acid moieties into insulin icodec plays a crucial role in its prolonged action, as these fatty acid chains facilitate the formation of stable hexameric structures, thereby delaying insulin absorption and promoting sustained release. Yet, the underlying biophysics still is elusive of the roles of the three site-specific mutations (Y14A\_E, Y37B\_H, F46B\_H) of insulin icodec in its prolonged activity. Thus, through a comprehensive structural and biophysical analysis of the insulin (both native and icodec) structures bound to its receptor, this article delves deep into the biophysics underlying the molecular design of insulin icodec, and identified a delicate biophysical mechanism through which two missense mutations of insulin icodec (Y37B\_H and F46B\_H) contribute to its prolonged duration of action. Overall, this structural and biophysical investigation provides valuable insights into the mechanisms underlying the relationship between three site-specific mutations and prolonged duration of action of insulin icodec, while understanding these modifications at a structural and biophysical level can aid in the rational design of future long-acting insulin analogues, offering further enhanced therapeutic options for diabetic patients.
Keywords:
Insulin icodec
; Molecular design
; Site-specific mutation
; Structural biophysics
1. Introduction
Ligand-receptor binding affinity is an essential parameter in both computational and experimental drug discovery & design [1]. Thanks to the continued development of experimental structural biology and the fifty-three-year old Protein Data Bank (PDB) [2,3], a high-throughput comprehensive structural biophysical analysis becomes possible [4] for specific ligand-receptor complex structures deposited in PDB, such that our understanding of the structural and biophysical basis of their interfacial stability is able to help us modify the binding affinity of certain drug target and its interacting partners.
Take insulin icodec for example [5,6,7,8]. Insulin icodec of Novo Nordisk is a long-acting insulin analogue for better management of blood sugar levels in people with diabetes [9,10,11]. Insulin icodec is designed to provide a steady release of insulin throughout the day, mimicking the natural insulin production in the body [12,13,14]. Insulin icodec is typically administered through injection once a week, which helps lower blood sugar levels by allowing glucose to enter the body’s cells, where it is subsequently used for energy production [15,16]. Moreover, insulin icodec has a distinct pharmacokinetic profile compared to other long-acting insulin analogs. It exhibits a long duration of action, with a half-life of approximately 196 hours, leading to improved glycemic control and reduced hypoglycemia risk [17,18]. Interestingly, Icosema (of Novo Nordisk, too) [19,20,21,22] represents a combination medication that consists of insulin icodec and semaglutide, which belongs to a class of medications called glucagon-like peptide-1 (GLP-1) receptor agonists and helps regulate blood sugar levels by stimulating the release of insulin, reducing the production of glucagon (a hormone that increases blood sugar levels), and slowing down the digestion process for weight reduction [23,24,25].
Given the two-dimensional structures of native human insulin and insulin icodec as shown in Figure 1, this article summarizes below a list of structural modifications of insulin icodec compared to native human insulin [26,27,28]:
Overall, these structural modifications (Figure 2) in insulin icodec result in a more stable and longer-acting insulin analogue compared to regular insulin, providing a more consistent and sustained blood glucose-lowering effect:
- insulin icodec is able to form aggregates or clusters at the subcutaneous injection site, gradually releasing into the bloodstream over an extended period.
- insulin icodec undergoes structural modifications that increases its stability and solubility and preventing enzyme-mediated degradation and rapid clearance [36].
- insulin icodec is conjugated with a fatty acid at position B30 (K50B_C20, Figure 2). After injection, the fatty acid chain in insulin icodec interacts with albumin in the subcutaneous tissue, forming reversible albumin-insulin complexes. These complexes act as a reservoir, gradually releasing insulin icodec into the bloodstream, increasing its fat solubility and allowing it to bind to fatty acid-binding proteins, forming a depot of the insulin icodec reversibly bound to albumin.
- the incorporation of fatty acid chains facilitate the formation of stable hexameric structures, thereby delaying insulin absorption and promoting sustained release.
With respect to the addition of a fatty acid (K50B_C20, Figure 2), it is conceivable that the deletion of threonine at position B30 (T51B_del, Figure 2) is for K50B_C20 to take place easier and more efficiently than without the deletion of threonine at position B30. Nonetheless, the underlying biophysics remains elusive of the roles of the three site-specific mutations (Y14A_E, Y37B_H, F46B_H) of insulin icodec in its prolonged activity. Therefore, through a comprehensive structural and biophysical analysis of the insulin (both native and icodec) structures bound to its receptor, this article delves deep into the biophysics underlying the molecular design of insulin icodec, and identified a delicate biophysical mechanism through which two missense mutations of insulin icodec (Y37B_H and F46B_H) contribute to its prolonged duration of action.
2. Materials and Methods
2.1. A summary of insulin receptor-related structures in PDB
As listed in Table 1, as of November 16, 2023, there are a variety of experimental complex structures of insulin (analogues) bound to its receptor (IR), such as PDB entries 7PG3 (insulin receptor bound to 3 insulins), 7PG4 (insulin receptor bound to 2 insulins), 6SOF (insulin receptor bound to 4 insulins).
Among the insulin receptor-related structures listed in Table 1, PDB entry 6SOF [37] is the only experimental complex structure of insulin bound to IR, where all four distinct binding sites of the IR dimer are saturated by four insulin molecules. Therefore, PDB entry 6SOF [37] is chosen here as the structural template for subsequent structural modelling of insulin icodec bound to IR.
2.2. Construction of a complex structural model of insulin icodec bound to IR
Towards the construction of a complex structural model of insulin icodec bound to IR, first, the amino acid sequences of the two chains of native human insulin and IR (according to PDB entry 6SOF [37], supplementary file nati.pdb) are listed in italics in fasta format as below,
>InsulinA
GIVEQCCTSICSLYQLENYCN
>InsulinB
FVNQHLCGSHLVEALYLVCGERGFFYTPKT
>IRpartOne
HLYPGEVCPGMDIRNNLTRLHELENCSVIEGHLQILLMFKTRPEDFRDLSFPKLIMITDYLLLFRVYGLESLKDLFPNLTVIRGSRLFFNYALVIFEMVHLKELGLYNLMNITRGSVRIEKNNELCYLATIDWSRILDSVEDNYIVLNKDDNEECGDICPGTAKGKTNCPATVINGQFVERCWTHSHCQKVCPTICKSHGCTAEGLCCHSECLGNCSQPDDPTKCVACRNFYLDGRCVETCPPPYYHFQDWRCVNFSFCQDLHHKCKNSRRQGCHQYVIHNNKCIPECPSGYTMNSSNLLCTPCLGPCPKVCHLLEGEKTIDSVTSAQELRGCTVINGSLIINIRGGNNLAAELEANLGLIEEISGYLKIRRSYALVSLSFFRKLRLIRGETLEIGNYSFYALDNQNLRQLWDWSKHNLTITQGKLFFHYNPKLCLSEIHKMEEVSGTKGRQERNDIALKTNGDQASCENELLKFSYIRTSFDKILLRWEPYWPPDFRDLLGFMLFYKEAPYQNVTEFDGQDACGSNSWTVVDIDPPLRSNDPKSQNHPGWLMRGLKPWTQYAIFVKTLVTFSDERRTYGAKSDIIYVQTDATNPSVPLDPISVSNSSSQIILKWKPPSDPNGNITHYLVFWERQAEDSELFELDYCLKGLKLPSRTWSPPFESEDSQKHNQSEYEDSAGECCSCPKTDSQILKELEESSFRKTFEDYLHNVVFVPRPS
>IRpartTwo
HRPFEKVVNKESLVISGLRHFTGYRIELQACNQDTPEERCSVAAYVSARTMPEAKADDIVGPVTHEIFENNVVHLMWQEPKEPNGLIVLYEVSYRRYGDEELHLCVSRKHFALERGCRLRGLSPGNYSVRIRATSLAGNGSWTEPTYFYVTDYLDVPSNIAK
Second, the amino acid sequences of the two chains of insulin icodec and IR are listed in italics in fasta format as below,
>InsulinIcodecA
GIVEQCCTSICSLEQLENYCN
>InsulinIcodecB
FVNQHLCGSHLVEALHLVCGERGFFHTPKT
>IRpartOne
HLYPGEVCPGMDIRNNLTRLHELENCSVIEGHLQILLMFKTRPEDFRDLSFPKLIMITDYLLLFRVYGLESLKDLFPNLTVIRGSRLFFNYALVIFEMVHLKELGLYNLMNITRGSVRIEKNNELCYLATIDWSRILDSVEDNYIVLNKDDNEECGDICPGTAKGKTNCPATVINGQFVERCWTHSHCQKVCPTICKSHGCTAEGLCCHSECLGNCSQPDDPTKCVACRNFYLDGRCVETCPPPYYHFQDWRCVNFSFCQDLHHKCKNSRRQGCHQYVIHNNKCIPECPSGYTMNSSNLLCTPCLGPCPKVCHLLEGEKTIDSVTSAQELRGCTVINGSLIINIRGGNNLAAELEANLGLIEEISGYLKIRRSYALVSLSFFRKLRLIRGETLEIGNYSFYALDNQNLRQLWDWSKHNLTITQGKLFFHYNPKLCLSEIHKMEEVSGTKGRQERNDIALKTNGDQASCENELLKFSYIRTSFDKILLRWEPYWPPDFRDLLGFMLFYKEAPYQNVTEFDGQDACGSNSWTVVDIDPPLRSNDPKSQNHPGWLMRGLKPWTQYAIFVKTLVTFSDERRTYGAKSDIIYVQTDATNPSVPLDPISVSNSSSQIILKWKPPSDPNGNITHYLVFWERQAEDSELFELDYCLKGLKLPSRTWSPPFESEDSQKHNQSEYEDSAGECCSCPKTDSQILKELEESSFRKTFEDYLHNVVFVPRPS
>IRpartTwo
HRPFEKVVNKESLVISGLRHFTGYRIELQACNQDTPEERCSVAAYVSARTMPEAKADDIVGPVTHEIFENNVVHLMWQEPKEPNGLIVLYEVSYRRYGDEELHLCVSRKHFALERGCRLRGLSPGNYSVRIRATSLAGNGSWTEPTYFYVTDYLDVPSNIAK
Lastly, to build a reliable structural model of the insulin icodec bound to IR, the amino acid sequences above (>InsulinIcodecA, >InsulinIcodecB, >IRpartOne and >IRpartTwo) were plugged into the SWISS-MODEL homology modeling server [38,39] and the Modeller software [40] to build a homology structural model with reasonable accuracy, supplementary file icod.pdb, which was subsequently subject to a set of comprehensive structural biophysical analysis as described previously in [4].
2.3. A comprehensive structural and biophysical analysis of insulin icodec bound to IR
As described previously in [4], a comprehensive structural and biophysical analysis was conducted for the structural model of the insulin icodec bound to IR (supplementary file icod.pdb). Specifically, the salt bridge analysis was conducted with an in-house python script only for titrateable residues (Asp, Glu, Lys, Arg and His), 4.0 Å was used as the cutoff distance for the two oppositely charged groups [4]. The hydrogen bond analysis was also conducted for only side chain nuclei with an in-house python script, and employed two geometric criteria: (a) a cutoff value of the angle formed by acceptor (A), donor (D) and hydrogen (H) () of 30°; (b) a cutoff value of donor-acceptor distance at 3.0 Å. That is, a hydrogen bond is only considered to be formed if is no larger than 30° and the donor-acceptor distance is not larger than 3.0 Å [4].
3. Results
As described above, the addition of a fatty acid to insulin icodec (K50B_C20, Figure 2) is for it to interact with albumin and form reversible albumin-insulin complexes, i.e., a reservoir for gradually releasing insulin icodec into the bloodstream. Moreover, it is highly likely that the deletion of threonine at position B30 (T51B_del, Figure 2) is for K50B_C20 to take place easier and more efficiently. As a result, the comprehensive structural and biophysical analysis here focuses on the three site-specific mutations (Y14A_E, Y37B_H, F46B_H) of insulin icodec.
Coincidentally, the three mutations (Y14A_E, Y37B_H, F46B_H) invariably fall into the same category of the substitution of an amino acid residue with an amino acid residue with a hydrophilic side chain. Briefly, with a set of comprehensive structural and biophysical analysis [4] for supplementary files icod.pdb and nati.pdb, this article here for the first time reports that
Of the three mutations (Y14A_E, Y37B_H, F46B_H), a total of three electrostatic interactions (including salt bridges and hydrogen bond) were identified and listed as below in Table 2.
Specifically, before Y37B_H (Figure 1 and Figure 2), i.e., in the case of native human insulin (Table 3), only one salt bridge (3.204 Å long) was formed between C_LYS_921 (of IR) and F_GLU_1804 (of regular insulin) at the binding interface of native human insulin bound to IR (supplementary file nati.pdb). Similarly, after Y37B_H (Figure 1 and Figure 2), i.e., in the case of insulin icodec (Table 4), only one salt bridge (3.669 Å long) was formed between C_LYS_921 (of IR) and F_GLU_1804 (of insulin icodec) at the binding interface of insulin icodec bound to IR (supplementary file icod.pdb). Interestingly, apart from the interfacial salt bridge between C_LYS_921 (of IR) and F_GLU_1804 (of insulin icodec), there is also one intra-molecular salt bridge (3.659 Å long) between F_HIS_1799 (of insulin icodec) and F_GLU_1804 (of insulin icodec).
Thus, it is obvious that in the case of native human insulin (Table 3), F_GLU_1804 (of regular insulin) focuses on binding to C_LYS_921 (of IR) alone with a 3.204 Å long interfacial salt bridge between regular insulin and IR, while in the case of insulin icodec (Table 4), F_GLU_1804 (of insulin icodec) has to bind both C_LYS_921 (of IR) and F_HIS_1799 (of insulin icodec) with two salt bridges, as listed in Table 4.
Furthermore, the existence of F_HIS_1799 (of insulin icodec) is entirely due to the site-directed mutation Y37B_H (Figure 1 and Figure 2). Therefore, it is entirely due to the site-directed mutation Y37B_H that the salt bridge at the binding interface between insulin and IR becomes weaker (from 3.204 Å long to 3.669 Å long), but is still not disrupted by the site-directed mutation Y37B_H, such that the binding affinity of ligand-receptor is lowered but not eliminated by the site-directed mutation Y37B_H (Figure 1 and Figure 2), and thereby ensuring downstream signal transduction for the prolonged blood glucose-lowering effect of insulin icodec.
With respect to the site-directed mutation F46B_H (Figure 1 and Figure 2), it is not involved in any salt bridge or hydrogen bond within insulin icodec itself, nor is it involved in any salt bridge or hydrogen bond at the binding interface between insulin icodec and IR, according comprehensive structural and biophysical analysis [4] of insulin icodec bound to IR (supplementary file icod.pdb). Nonetheless, a close inspection with PyMol [41] of supplementary file icod.pdb revealed that the positively charged side chain of His1808 of insulin icodec (due to the site-specific mutation F46B_H, Figure 2) is only 6.4 Å away (Figure 4) from the positively charged side chain of R900 (Arg900) of IR. Despite the cutoff distance (4.0 Å) as used in [4], a distance 6.4 Å is not sufficient to be defined as strong an electrostatic interaction as a salt bridge, it still is able to induce a weak electrostatic repulsive force between the positively charged side chains of His1808 of insulin icodec and that of R900 of IR, and thereby reducing the binding affinity between insulin icodec and IR. Moreover, in case the site-specific mutation is not F46B_H, Figure 2), but F46B_E, i.e., it is not a histidine, but a glutamate, it is conceivable that the distance between Glu1808 of insulin icodec and R900 of IR is shorter than 6.4 Å (Figure 4), make it highly likely for a novel stable salt bridge to be formed between E1808 (Glu1808) of insulin icodec and R900 (Arg900) of IR, according to the 4 Å distance cutoff as used in [4].
Finally, with respect to the site-directed mutation Y14A_E (Figure 1 and Figure 2), , it mutation does not seem to impact the binding between insulin icodec and IR, as it is a fully solvent-exposed amino acid residue, as shown by Figure 5. Thus, as of November 16, 2023 from structural and biophysical perspective, it is not clear yet why site-directed mutation Y14A_E is included in the design of insulin icodec. Here, this article is only able to speculate that Y14A_E is for the overall structural stability of icodec insulin for it to exert its prolonged blood glucose-lowering effect for patients with diabetes.
4. Conclusion and Discussion
For the first time, through a comprehensive structural and biophysical analysis of the insulin (both native and icodec) structures bound to its receptor, this article puts forward a delicate biophysical mechanism through which two missense mutations of insulin icodec (Y37B_H and F46B_H) contribute to its prolonged duration of action. In addition to the biophysics underlying the molecular design of insulin icodec, this article also argues that this structural and biophysical understanding is of help for rational design of future long-acting insulin analogues, offering further enhanced therapeutic options for diabetic patients.
5. Ethical statement
No ethical approval is required.
6. Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the author used OpenAI’s ChatGPT in order to improve the readability of the manuscript, and to make it as concise and short as possible. After using this tool, the author reviewed and edited the content as needed and takes full responsibility for the content of the publication.
Author Contributions
Conceptualization, W.L.; methodology, W.L.; software, W.L.; validation, W.L.; formal analysis, W.L.; investigation, W.L.; resources, W.L.; data duration, W.L.; writing–original draft preparation, W.L.; writing–review and editing, W.L.; visualization, W.L.; supervision, W.L.; project administration, W.L.; funding acquisition, not applicable.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Fuji, H.; Qi, F.; Qu, L.; Takaesu, Y.; Hoshino, T. Prediction of Ligand Binding Affinity to Target Proteins by Molecular Mechanics Theoretical Calculation. Chemical and Pharmaceutical Bulletin 2017, 65, 461–468. [Google Scholar] [CrossRef]
- Berman, H.; Henrick, K.; Nakamura, H. Announcing the worldwide Protein Data Bank. Nature Structural & Molecular Biology 2003, 10, 980–980. [Google Scholar]
- Li, W. Half-a-century Burial of ρ, θ and φ in PDB 2021.
- Li, W. How do SMA-linked mutations of SMN1 lead to structural/functional deficiency of the SMA protein? PLOS ONE 2017, 12, e0178519. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, E.; Pridal, L.; Glendorf, T.; Hansen, B.F.; Hubálek, F.; Kjeldsen, T.; Kristensen, N.R.; Lützen, A.; Lyby, K.; Madsen, P.; Pedersen, T.Å.; Ribel-Madsen, R.; Stidsen, C.E.; Haahr, H. Molecular and pharmacological characterization of insulin icodec: a new basal insulin analog designed for once-weekly dosing. BMJ Open Diabetes Research & Care 2021, 9, e002301. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Once-weekly basal insulin icodec: Looking ONWARDS from pharmacology to clinical trials. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2022, 16, 102615. [Google Scholar] [CrossRef]
- Bajaj, H.S.; Bergenstal, R.M.; Christoffersen, A.; Davies, M.J.; Gowda, A.; Isendahl, J.; Lingvay, I.; Senior, P.A.; Silver, R.J.; Trevisan, R.; Rosenstock, J. Switching to Once-Weekly Insulin Icodec Versus Once-Daily Insulin Glargine U100 in Type 2 Diabetes Inadequately Controlled on Daily Basal Insulin: A Phase 2 Randomized Controlled Trial. Diabetes Care 2021, 44, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, J.; Bain, S.C.; Gowda, A.; Jódar, E.; Liang, B.; Lingvay, I.; Nishida, T.; Trevisan, R.; Mosenzon, O. Weekly Icodec versus Daily Glargine U100 in Type 2 Diabetes without Previous Insulin. New England Journal of Medicine 2023, 389, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Philis-Tsimikas, A.; Bajaj, H.S.; Begtrup, K.; Cailleteau, R.; Gowda, A.; Lingvay, I.; Mathieu, C.; Russell-Jones, D.; Rosenstock, J. Rationale and design of the phase 3a development programme (ONWARDS 1–6 trials) investigating once-weekly insulin icodec in diabetes. Diabetes, Obesity and Metabolism 2022, 25, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Philis-Tsimikas, A.; Asong, M.; Franek, E.; Jia, T.; Rosenstock, J.; Stachlewska, K.; Watada, H.; Kellerer, M. Switching to once-weekly insulin icodec versus once-daily insulin degludec in individuals with basal insulin-treated type 2 diabetes (ONWARDS 2): a phase 3a, randomised, open label, multicentre, treat-to-target trial. The Lancet Diabetes & Endocrinology 2023, 11, 414–425. [Google Scholar] [CrossRef]
- Mathieu, C.; Ásbjörnsdóttir, B.; Bajaj, H.S.; Lane, W.; Matos, A.L.S.A.; Murthy, S.; Stachlewska, K.; Rosenstock, J. Switching to once-weekly insulin icodec versus once-daily insulin glargine U100 in individuals with basal-bolus insulin-treated type 2 diabetes (ONWARDS 4): a phase 3a, randomised, open-label, multicentre, treat-to-target, non-inferiority trial. The Lancet 2023, 401, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Lingvay, I.; Asong, M.; Desouza, C.; Gourdy, P.; Kar, S.; Vianna, A.; Vilsbøll, T.; Vinther, S.; Mu, Y. Once-Weekly Insulin Icodec vs Once-Daily Insulin Degludec in Adults With Insulin-Naive Type 2 Diabetes. JAMA 2023, 330, 228. [Google Scholar] [CrossRef] [PubMed]
- Lingvay, I.; Buse, J.B.; Franek, E.; Hansen, M.V.; Koefoed, M.M.; Mathieu, C.; Pettus, J.; Stachlewska, K.; Rosenstock, J. A Randomized, Open-Label Comparison of Once-Weekly Insulin Icodec Titration Strategies Versus Once-Daily Insulin Glargine U100. Diabetes Care 2021, 44, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, T.B.; Hubálek, F.; Hjørringgaard, C.U.; Tagmose, T.M.; Nishimura, E.; Stidsen, C.E.; Porsgaard, T.; Fledelius, C.; Refsgaard, H.H.F.; Gram-Nielsen, S.; Naver, H.; Pridal, L.; Hoeg-Jensen, T.; Jeppesen, C.B.; Manfè, V.; Ludvigsen, S.; Lautrup-Larsen, I.; Madsen, P. Molecular Engineering of Insulin Icodec, the First Acylated Insulin Analog for Once-Weekly Administration in Humans. Journal of Medicinal Chemistry 2021, 64, 8942–8950. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, J.; Bajaj, H.S.; Janež, A.; Silver, R.; Begtrup, K.; Hansen, M.V.; Jia, T.; Goldenberg, R. Once-Weekly Insulin for Type 2 Diabetes without Previous Insulin Treatment. New England Journal of Medicine 2020, 383, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, H.S.; Goldenberg, R.M. Insulin Icodec Weekly: A Basal Insulin Analogue for Type 2 Diabetes. European Endocrinology 2023, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- DiMarchi, R.D.; Mayer, J.P. Icodec Advances the Prospect of Once-Weekly Insulin Injection. Journal of Medicinal Chemistry 2021, 64, 8939–8941. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, J.; Prato, S.D. Basal weekly insulins: the way of the future! Metabolism 2022, 126, 154924. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.; Bhattacharya, S.; Kapoor, N. Contemporary Classification of Glucagon-Like Peptide 1 Receptor Agonists (GLP1RAs). Diabetes Therapy 2021, 12, 2133–2147. [Google Scholar] [CrossRef] [PubMed]
- Pratley, R.; Amod, A.; Hoff, S.T.; Kadowaki, T.; Lingvay, I.; Nauck, M.; Pedersen, K.B.; Saugstrup, T.; Meier, J.J. Oral semaglutide versus subcutaneous liraglutide and placebo in type 2 diabetes (PIONEER 4): a randomised, double-blind, phase 3a trial. The Lancet 2019, 394, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.L.; Beutel, T.R.; Trujillo, J.M. Oral semaglutide in type 2 diabetes. Journal of Diabetes and its Complications 2020, 34, 107520. [Google Scholar] [CrossRef] [PubMed]
- Li, W. Strengthening Semaglutide-GLP-1R Binding Affinity via a Val27-Arg28 Exchange in the Peptide Backbone of Semaglutide: A Computational Structural Approach. Journal of Computational Biophysics and Chemistry 2021, 20, 495–499. [Google Scholar] [CrossRef]
- Nadkarni, P.; Chepurny, O.G.; Holz, G.G. Regulation of Glucose Homeostasis by GLP-1. In Progress in Molecular Biology and Translational Science; Elsevier, 2014; pp. 23–65.
- Lau, J.; Bloch, P.; Schäffer, L.; Pettersson, I.; Spetzler, J.; Kofoed, J.; Madsen, K.; Knudsen, L.B.; McGuire, J.; Steensgaard, D.B.; Strauss, H.M.; Gram, D.X.; Knudsen, S.M.; Nielsen, F.S.; Thygesen, P.; Reedtz-Runge, S.; Kruse, T. Discovery of the Once-Weekly Glucagon-Like Peptide-1 (GLP-1) Analogue Semaglutide. Journal of Medicinal Chemistry 2015, 58, 7370–7380. [Google Scholar] [CrossRef] [PubMed]
- Gabery, S.; Salinas, C.G.; Paulsen, S.J.; Ahnfelt-Rønne, J.; Alanentalo, T.; Baquero, A.F.; Buckley, S.T.; Farkas, E.; Fekete, C.; Frederiksen, K.S.; Helms, H.C.C.; Jeppesen, J.F.; John, L.M.; Pyke, C.; Nøhr, J.; Lu, T.T.; Polex-Wolf, J.; Prevot, V.; Raun, K.; Simonsen, L.; Sun, G.; Szilvásy-Szabó, A.; Willenbrock, H.; Secher, A.; Knudsen, L.B. Semaglutide lowers body weight in rodents via distributed neural pathways. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Plum-Mörschel, L.; Andersen, L.R.; Hansen, S.; Hövelmann, U.; Krawietz, P.; Kristensen, N.R.; Lehrskov, L.L.; Haahr, H. Pharmacokinetic and Pharmacodynamic Characteristics of Insulin Icodec After Subcutaneous Administration in the Thigh, Abdomen or Upper Arm in Individuals with Type 2 Diabetes Mellitus. Clinical Drug Investigation 2023, 43, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Belal, H.; Gandhi, G.Y. In uncontrolled T2DM treated with a basal-bolus insulin regimen, weekly icodec was noninferior to daily glargine for HbA1c at 26 wk. Annals of Internal Medicine 2023, 176, JC94. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.L.; Bassetti, M.; Mangoni, A.A. Drugs in Context Editorial: Review of 2020 and what lies ahead in therapeutic interventions. Drugs in Context 2021, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Griffin, T.P.; Dinneen, S.F. In T2DM, weekly insulin icodec did not differ from daily glargine for reducing HbA1c or significant/severe hypoglycemia. Annals of Internal Medicine 2021, 174, JC34. [Google Scholar] [CrossRef] [PubMed]
- Pieber, T.R.; Arfelt, K.N.; Cailleteau, R.; Hart, M.; Kar, S.; Mursic, I.; Svehlikova, E.; Urschitz, M.; Haahr, H. Hypoglycaemia frequency and physiological response after double or triple doses of once-weekly insulin icodec vs once-daily insulin glargine U100 in type 2 diabetes: a randomised crossover trial. Diabetologia 2023, 66, 1413–1430. [Google Scholar] [CrossRef] [PubMed]
- Zerihun, K.; Mhanna, M.; Ayesh, H.; Ghazaleh, S.; Khader, Y.; Beran, A.; Aldhafeeri, A.; Sharma, S.; Iqbal, A.; Legesse, H.; Jaume, J. Efficacy and Safety of Insulin Icodec Versus Glargine U100: A Meta-Analysis of Randomized Controlled Trials. American Journal of Therapeutics 2022, 30, e480–e483. [Google Scholar] [CrossRef] [PubMed]
- Bellary, S.; Barnett, A.H. Insulin icodec: evolution or revolution in diabetes therapy? The Lancet Diabetes & Endocrinology 2023, 11, 379–380. [Google Scholar] [CrossRef]
- e Silva, R.R.; de Miranda Gauza, M.; Guisso, M.E.S.; da Silva, J.O.N.; Kohara, S.K. Once-Weekly Insulin Icodec vs. Once-Daily Insulin Glargine U100 for type 2 diabetes: a systematic review and meta-analysis of phase 2 randomized controlled trials. Archives of Endocrinology and Metabolism 2023, 67. [Google Scholar] [CrossRef] [PubMed]
- Pieber, T.R.; Asong, M.; Fluhr, G.; Höller, V.; Kristensen, N.R.; Larsen, J.H.; Ribel-Madsen, R.; Svehlikova, E.; Vinther, S.; Voortman, M.; Haahr, H. Pharmacokinetic and pharmacodynamic properties of once-weekly insulin icodec in individuals with type 2 diabetes. Diabetes, Obesity and Metabolism 2023. [Google Scholar] [CrossRef]
- Russell-Jones, D.; Babazono, T.; Cailleteau, R.; Engberg, S.; Irace, C.; Kjaersgaard, M.I.S.; Mathieu, C.; Rosenstock, J.; Woo, V.; Klonoff, D.C. Once-weekly insulin icodec versus once-daily insulin degludec as part of a basal-bolus regimen in individuals with type 1 diabetes (ONWARDS 6): a phase 3a, randomised, open-label, treat-to-target trial. The Lancet 2023. [Google Scholar] [CrossRef] [PubMed]
- Feher, J. Digestion and Absorption of the Macronutrients. In Quantitative Human Physiology; Elsevier, 2017; pp. 821–833. [CrossRef]
- Gutmann, T.; Schäfer, I.B.; Poojari, C.; Brankatschk, B.; Vattulainen, I.; Strauss, M.; Ünal Coskun. Cryo-EM structure of the complete and ligand-saturated insulin receptor ectodomain. Journal of Cell Biology 2019, 219. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; Lepore, R.; Schwede, T. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Research 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; Schwede, T. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Research 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Sali, A. Protein Structure Modeling with MODELLER. In Methods in Molecular Biology; Springer US, 2020; pp. 239–255.
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsletter On Protein Crystallography 2002, 40, 82–92. [Google Scholar]
Figure 1.
A brief illustration of the two-dimensional structures of native human insulin and insulin icodec of Novo Nordisk [29,30,31]. In this figure, the amino acid residues with pink backgrounds represents the positions of the three site-specific mutations (Y14A_E, Y37B_H, F46B_H) of insulin icodec.
Figure 1.
A brief illustration of the two-dimensional structures of native human insulin and insulin icodec of Novo Nordisk [29,30,31]. In this figure, the amino acid residues with pink backgrounds represents the positions of the three site-specific mutations (Y14A_E, Y37B_H, F46B_H) of insulin icodec.
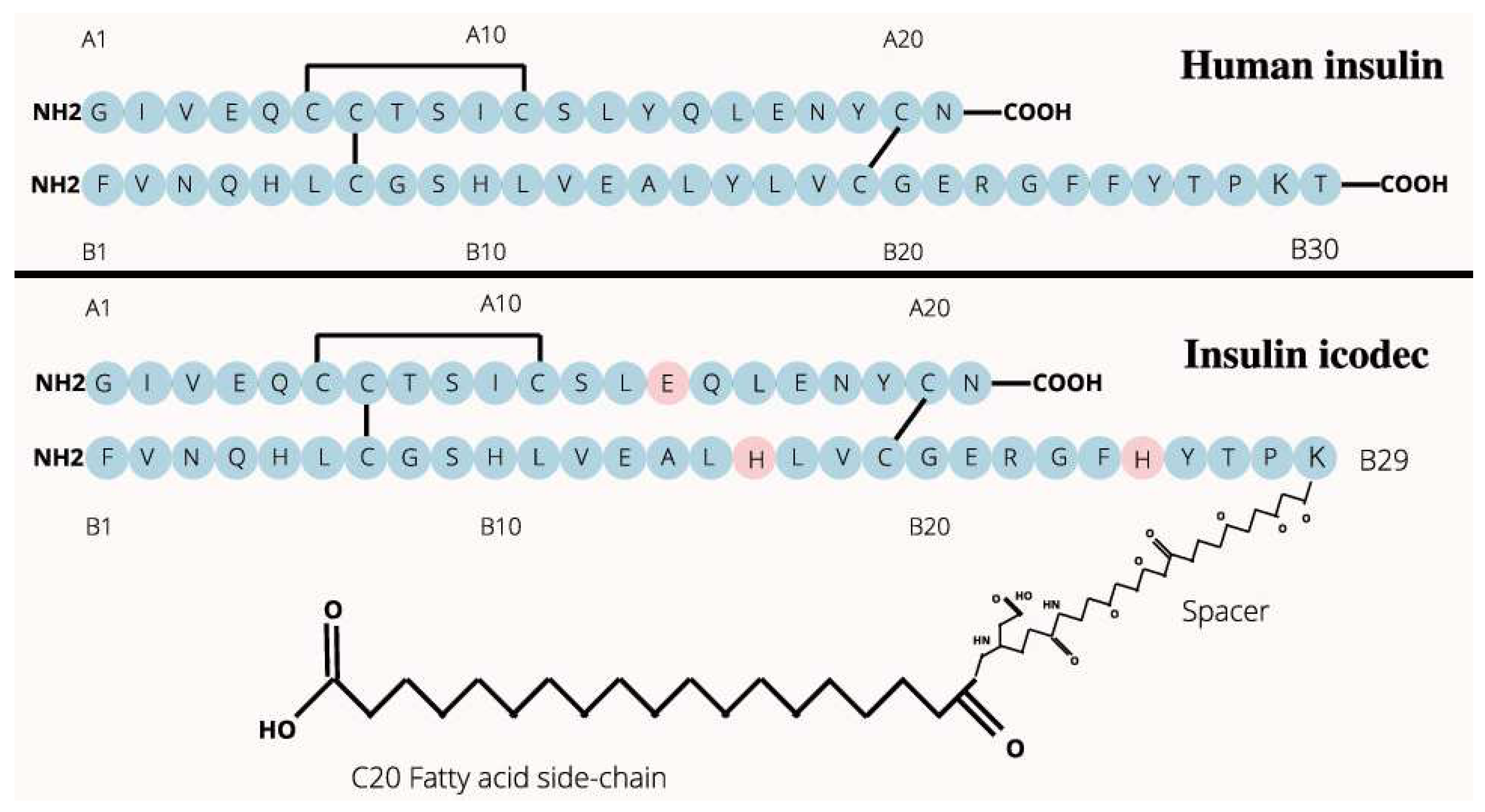
Figure 2.
A summary of the structural modification of insulin icodec in comparison to native human insulin. In this figure, Y14A_E (i.e., replacement of Tyr14 (Y14) at position A14 (position 14 of chain A) by a histidine), Y37B_H (i.e., replacement of Tyr16 (Y16) at position B16 by a histidine) and F46B_H (i.e., replacement of Phe25 (F25) at position B25 by a histidine) represent three site-specific missense mutations of insulin icodec, T51B_del represents deletion of Thr30 (T30) at position B30, while K50B_C20 represents the addition of a 20-carbon fatty acid to the lysine amino acid (K50B) at position B29 [32,33,34,35].
Figure 2.
A summary of the structural modification of insulin icodec in comparison to native human insulin. In this figure, Y14A_E (i.e., replacement of Tyr14 (Y14) at position A14 (position 14 of chain A) by a histidine), Y37B_H (i.e., replacement of Tyr16 (Y16) at position B16 by a histidine) and F46B_H (i.e., replacement of Phe25 (F25) at position B25 by a histidine) represent three site-specific missense mutations of insulin icodec, T51B_del represents deletion of Thr30 (T30) at position B30, while K50B_C20 represents the addition of a 20-carbon fatty acid to the lysine amino acid (K50B) at position B29 [32,33,34,35].
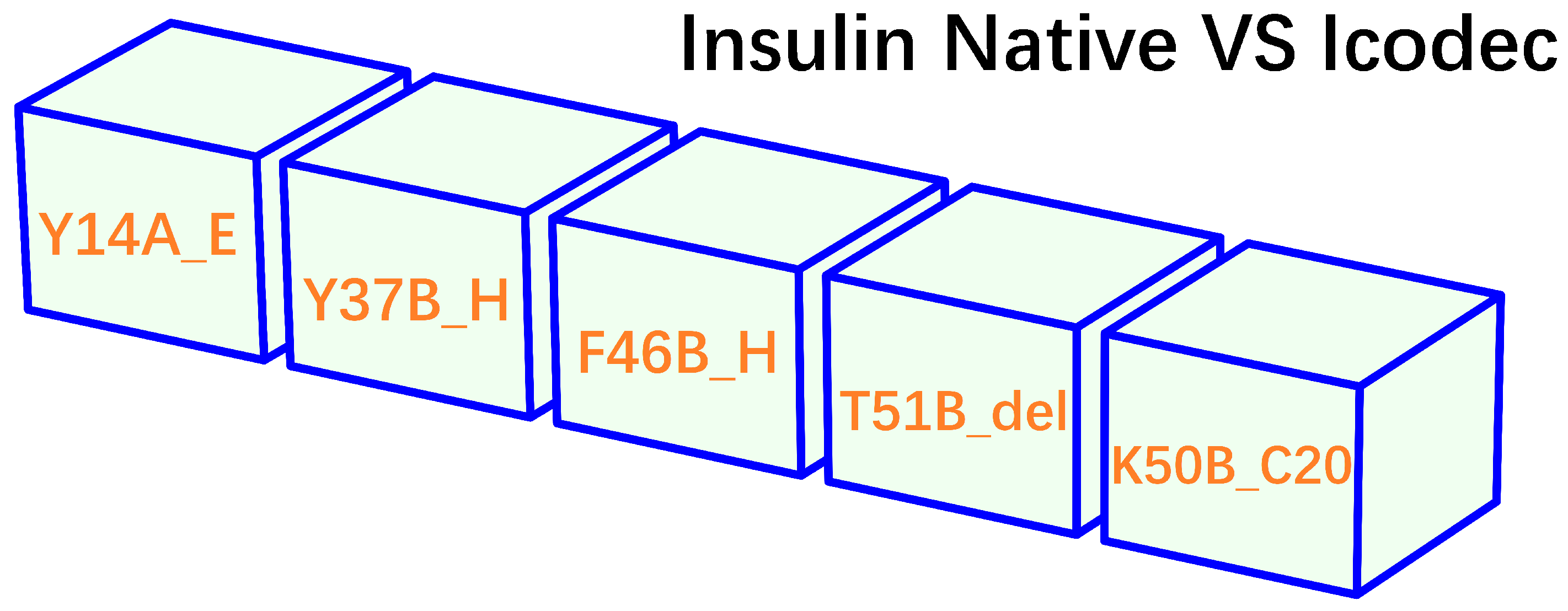
Figure 3.
Glu1804 of insulin icodec forms two salt bridges, one with His1799 of insulin icodec (due to the Y37B_H substitution), and another one with Lys921 of IR at their binding interface. This figure is prepared by PyMol [41].
Figure 3.
Glu1804 of insulin icodec forms two salt bridges, one with His1799 of insulin icodec (due to the Y37B_H substitution), and another one with Lys921 of IR at their binding interface. This figure is prepared by PyMol [41].
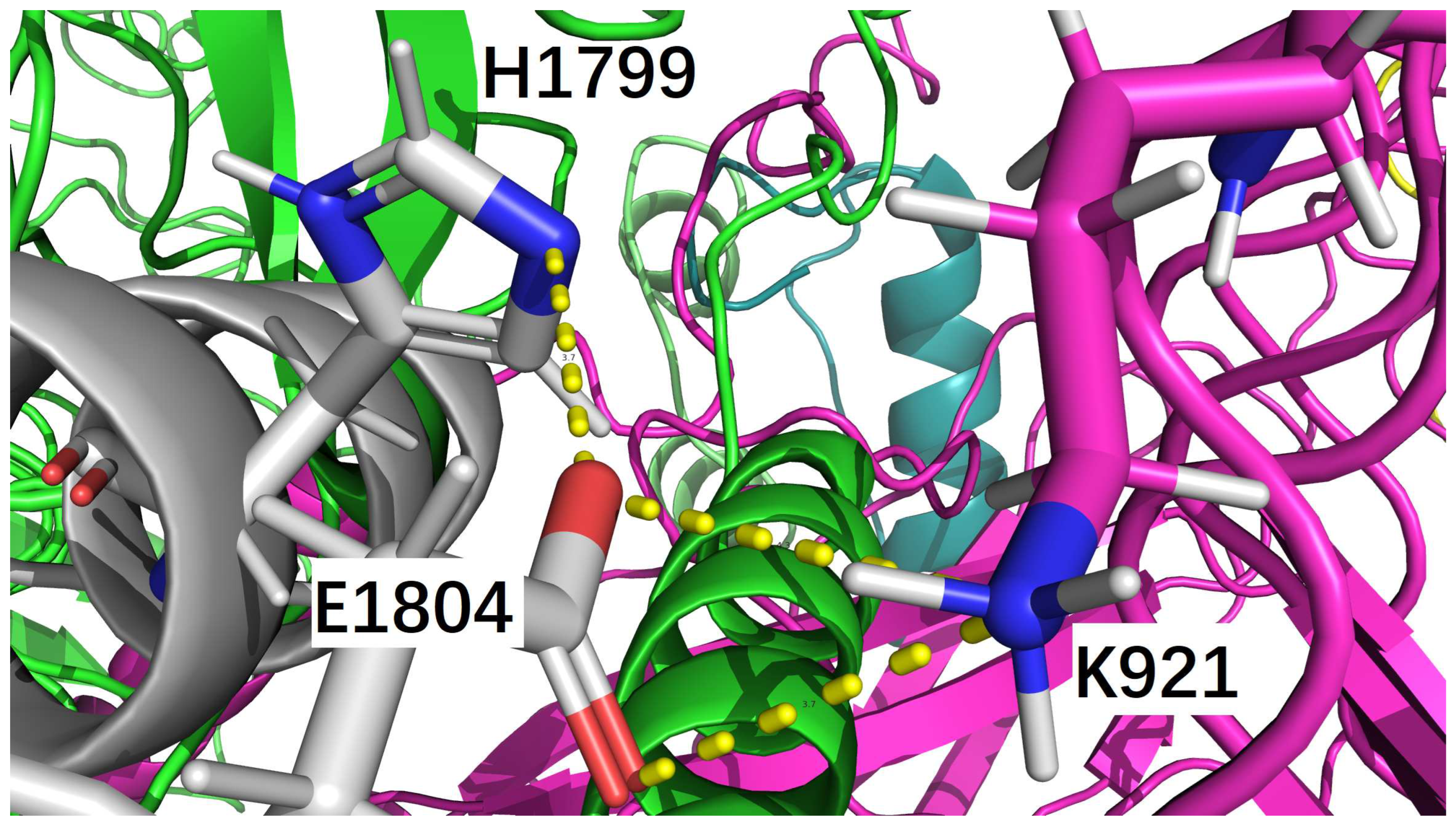
Figure 4.
A 6.4 Å long electrostatic repulsive force between between H1808 of insulin icodec (His1808, due to the site-specific mutation F46B_H, Figure 2) and R900 of IR. This figure is prepared by PyMol [41] according to supplementary file icod.pdb.
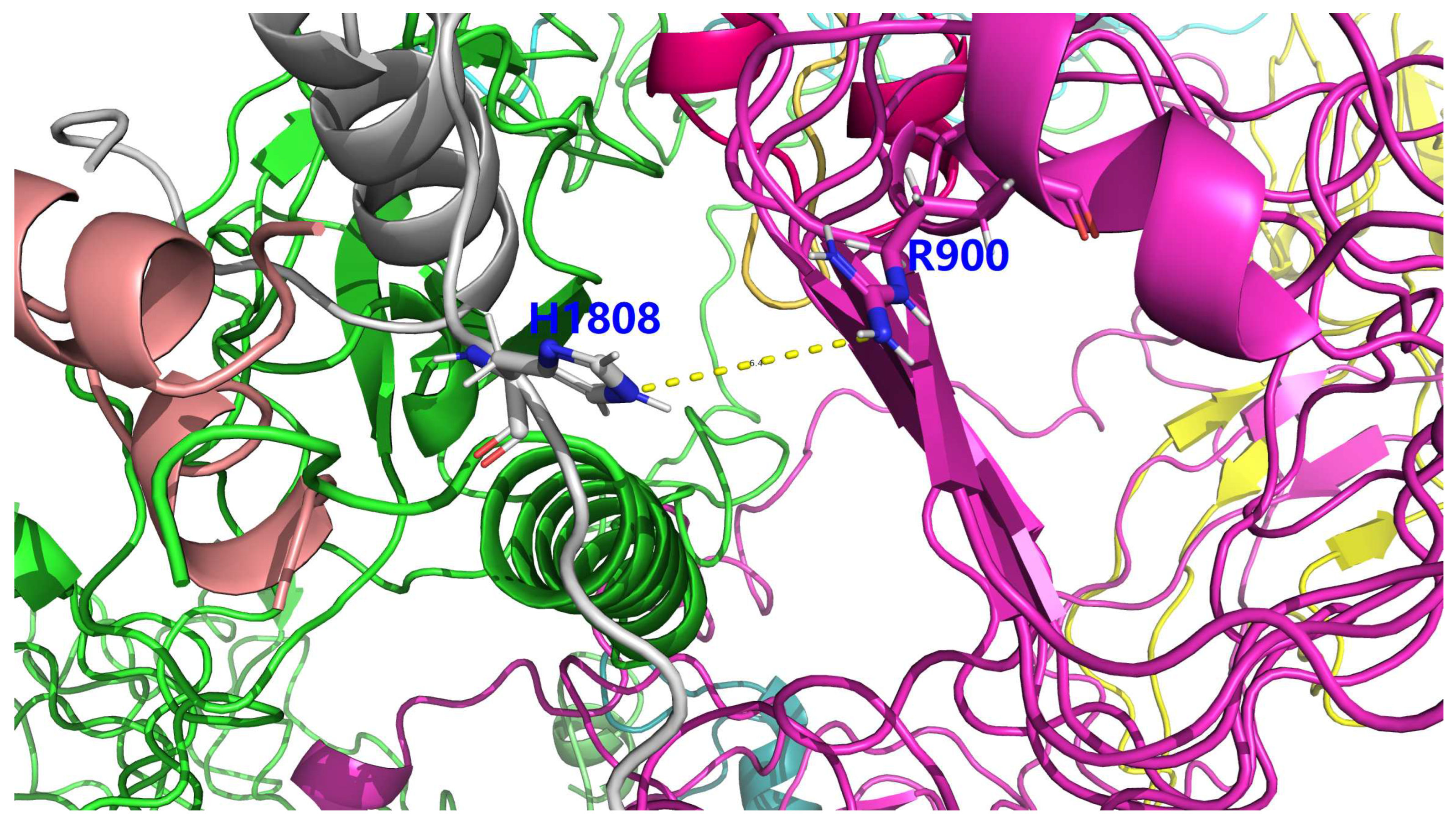
Figure 5.
A fully solvent-exposed Glu14 of insulin icodec according to supplementary file icod.pdb. This figure is prepared by PyMol [41].
Figure 5.
A fully solvent-exposed Glu14 of insulin icodec according to supplementary file icod.pdb. This figure is prepared by PyMol [41].
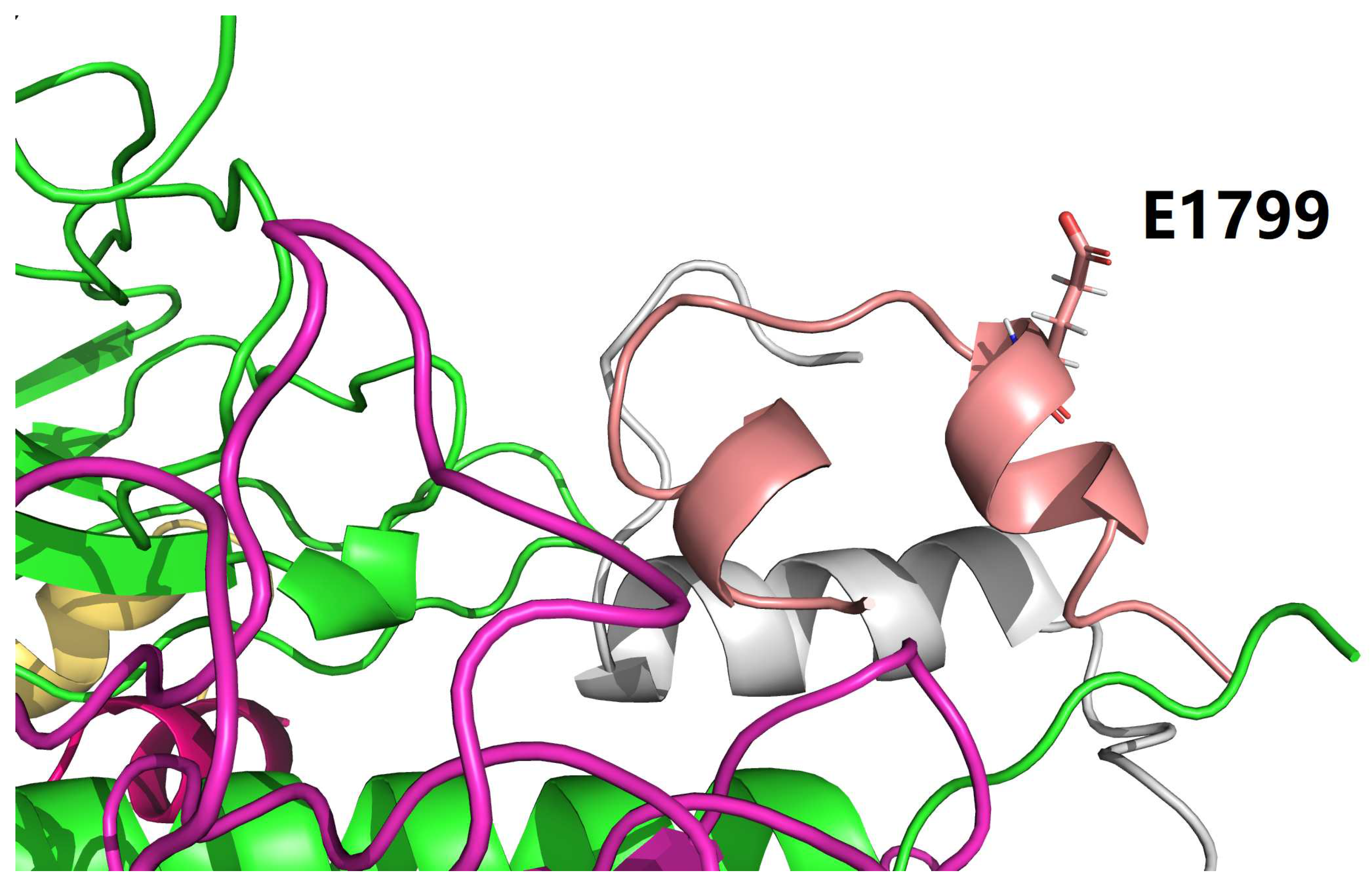

Table 1.
Experimentally determined IR-related structures (released newest from oldest) in the Protein Data Bank (PDB [2]) as of November 16, 2023, QUERY code: UniProt Molecule Name = "Insulin receptor".
Table 1.
Experimentally determined IR-related structures (released newest from oldest) in the Protein Data Bank (PDB [2]) as of November 16, 2023, QUERY code: UniProt Molecule Name = "Insulin receptor".
| PDB ID | Structure Title (release date from newest to oldest) |
|---|---|
| 8DWN | Crystal structure of bis-phosphorylated insulin receptor kinase domain |
| 7YQ3 | human insulin receptor bound with A43 DNA aptamer and insulin |
| 7YQ4 | human insulin receptor bound with A62 DNA aptamer and insulin - locally refined |
| 7YQ5 | human insulin receptor bound with A62 DNA aptamer and insulin |
| 7YQ6 | human insulin receptor bound with A62 DNA aptamer |
| 8EYX | Cryo-EM structure of 4 insulins bound full-length mouse IR mutant with physically decoupled alpha CTs (C684S/C685S/C687S; denoted as IR-3CS) Asymmetric conformation 1 |
| 8EYY | Cryo-EM structure of 4 insulins bound full-length mouse IR mutant with physically decoupled alpha CTs (C684S/C685S/C687S, denoted as IR-3CS) Asymmetric conformation 2 |
| 8EZ0 | Cryo-EM structure of 4 insulins bound full-length mouse IR mutant with physically decoupled alpha CTs (C684S/C685S/C687S; denoted as IR-3CS) Symmetric conformation |
| 8GUY | human insulin receptor bound with two insulin molecules |
| 7U6D | Head region of insulin receptor ectodomain (A-isoform) bound to the non-insulin agonist IM459 |
| 7U6E | Head region of insulin receptor ectodomain (A-isoform) bound to the non-insulin agonist IM462 |
| 7PHT | Structure of Insulin receptor’s transmembrane domain |
| 8DTL | Cryo-EM structure of insulin receptor (IR) bound with S597 peptide |
| 8DTM | Cryo-EM structure of insulin receptor (IR) bound with S597 component 2 |
| 7S0Q | Head region of a complex of IGF-I with the ectodomain of a hybrid insulin receptor / type 1 insulin-like growth factor receptor |
| 7S8V | Leg region of a complex of IGF-I with the ectodomain of a hybrid insulin receptor / type 1 insulin-like growth factor receptor |
| 7SL1 | Full-length insulin receptor bound with site 1 binding deficient mutant insulin (A-V3E) |
| 7SL2 | Full-length insulin receptor bound with site 2 binding deficient mutant insulin (A-L13R) – asymmetric conformation |
| 7SL3 | Full-length insulin receptor bound with site 2 binding deficient mutant insulin (A-L13R) – symmetric conformation |
| 7SL4 | Full-length insulin receptor bound with site 2 binding deficient mutant insulin (B-L17R) – asymmetric conformation |
| 7SL6 | Full-length insulin receptor bound with site 2 binding deficient mutant insulin (B-L17R) – symmetric conformation |
| 7SL7 | Full-length insulin receptor bound with both site 1 binding deficient mutant insulin (A-V3E) and site 2 binding deficient mutant insulin (A-L13R) |
| 7STH | Full-length insulin receptor bound with unsaturated insulin WT (2 insulin bound) symmetric conformation |
| 7STI | Full-length insulin receptor bound with unsaturated insulin WT (1 insulin bound) asymmetric conformation |
| 7STJ | Full-length insulin receptor bound with unsaturated insulin WT (2 insulins bound) asymmetric conformation (Conformation 1) |
| 7STK | Full-length insulin receptor bound with unsaturated insulin WT (2 insulins bound) asymmetric conformation (Conformation 2) |
| 7MQO | The insulin receptor ectodomain in complex with a venom hybrid insulin analogue - "head" region |
| 7MQR | The insulin receptor ectodomain in complex with four venom hybrid insulins - symmetric conformation |
| 7MQS | The insulin receptor ectodomain in complex with three venom hybrid insulin molecules - asymmetric conformation |
| 7MD4 | Insulin receptor ectodomain dimer complexed with two IRPA-3 partial agonists |
| 7MD5 | Insulin receptor ectodomain dimer complexed with two IRPA-9 partial agonists |
| 7PG0 | Low resolution Cryo-EM structure of full-length insulin receptor bound to 3 insulin with visible ddm micelle, conf 1 |
| 7PG2 | Low resolution Cryo-EM structure of full-length insulin receptor bound to 3 insulin, conf 1 |
| 7PG3 | Low resolution Cryo-EM structure of the full-length insulin receptor bound to 3 insulin, conf 2 |
| 7PG4 | Low resolution Cryo-EM structure of the full-length insulin receptor bound to 2 insulin, conf 3 |
| 7QID | tentative model of the human insulin receptor ectodomain bound by three insulin |
| 7KD6 | Insulin Receptor L1-CR plus alphaCT fragment in co-complex with Fv 83-7 and single-chain insulin SCI-b |
| 7BW7 | Cryo-EM Structure for the Ectodomain of the Full-length Human Insulin Receptor in Complex with 1 Insulin. |
| 7BW8 | Cryo-EM Structure for the Insulin Binding Region in the Ectodomain of the Full-length Human Insulin Receptor in Complex with 1 Insulin |
| 7BWA | Cryo-EM Structure for the Ectodomain of the Full-length Human Insulin Receptor in Complex with 2 Insulin |
| 6VEP | Human insulin in complex with the human insulin microreceptor in turn in complex with Fv 83-7 |
| 6VEQ | Con-Ins G1 in complex with the human insulin microreceptor in turn in complex with Fv 83-7 |
| 6SOF | human insulin receptor ectodomain bound by 4 insulin |
| 6PXV | Cryo-EM structure of full-length insulin receptor bound to 4 insulin. 3D refinement was focused on the extracellular region. |
| 6PXW | Cryo-EM structure of full-length insulin receptor bound to 4 insulin. 3D refinement was focused on the top part of the receptor complex. |
| 6HN4 | Leucine-zippered human insulin receptor ectodomain with single bound insulin - "lower" membrane-proximal part |
| 6HN5 | Leucine-zippered human insulin receptor ectodomain with single bound insulin - "upper" membrane-distal part |
| 6CE7 | Insulin Receptor ectodomain in complex with one insulin molecule |
| 6CE9 | Insulin Receptor ectodomain in complex with two insulin molecules |
| 6CEB | Insulin Receptor ectodomain in complex with two insulin molecules - C1 symmetry |
| 5U1M | Structure of the IRS-1 PTB Domain Bound to the Juxtamembrane Region of the Insulin Receptor |
| 5KQV | Insulin receptor ectodomain construct comprising domains L1,CR,L2, FnIII-1 and alphaCT peptide in complex with bovine insulin and FAB 83-14 (REVISED STRUCTURE) |
| 5TQ1 | Phospholipase C gamma-1 C-terminal SH2 domain bound to a phosphopeptide derived from the insulin receptor |
| 5J3H | Human insulin receptor domains L1-CR in complex with peptide S519C16 and 83-7 Fv |
| 5HHW | Crystal structure of insulin receptor kinase domain in complex with cis-(R)-7-(3-(azetidin-1-ylmethyl)cyclobutyl)-5-(3-((tetrahydro-2H-pyran-2-yl)methoxy)phenyl)-7H -pyrrolo[2,3-d]pyrimidin-4-amine. |
| 4ZXB | Structure of the human insulin receptor ectodomain, IRDeltabeta construct, in complex with four Fab molecules |
| 5E1S | The Crystal structure of INSR Tyrosine Kinase in complex with the Inhibitor BI 885578 |
| 4XSS | Insulin-like growth factor I in complex with site 1 of a hybrid insulin receptor / Type 1 insulin-like growth factor receptor |
| 4XST | Structure of the endoglycosidase-H treated L1-CR domains of the human insulin receptor in complex with residues 697-719 of the human insulin receptor (A-isoform) |
| 4XLV | Crystal structure of the activated insulin receptor tyrosine kinase dimer |
| 4OGA | Insulin in complex with Site 1 of the human insulin receptor |
| 2MFR | Solution structure of the transmembrane domain of the insulin receptor in micelles |
| 4IBM | Crystal structure of insulin receptor kinase domain in complex with an inhibitor Irfin-1 |
| 3W11 | Insulin receptor ectodomain construct comprising domains L1-CR in complex with human insulin, Alpha-CT peptide(704-719) and FAB 83-7 |
| 3W12 | Insulin receptor ectodomain construct comprising domains L1-CR in complex with high-affinity insulin analogue [D-PRO-B26]-DTI-NH2, alpha-CT peptide(704-719) and FAB 83-7 |
| 3W13 | Insulin receptor ectodomain construct comprising domains L1-CR in complex with high-affinity insulin analogue [D-PRO-B26]-DTI-NH2, alphact peptide(693-719) and FAB 83-7 |
| 3ETA | Kinase domain of insulin receptor complexed with a pyrrolo pyridine inhibitor |
| 3EKN | Insulin receptor kinase complexed with an inhibitor |
| 3EKK | Insulin receptor kinase complexed with an inhibitor |
| 2Z8C | Phosphorylated insulin receptor tyrosine kinase in complex with (4-[5-carbamoyl-4-(3-methylanilino)pyrimidin-2-yl]aminophenyl)acetic acid |
| 3BU3 | Crystal structure of the insulin receptor kinase in complex with IRS2 KRLB peptide |
| 3BU5 | Crystal structure of the insulin receptor kinase in complex with IRS2 KRLB peptide and ATP |
| 3BU6 | Crystal structure of the insulin receptor kinase in complex with IRS2 KRLB phosphopeptide |
| 2HR7 | Insulin receptor (domains 1-3) |
| 2B4S | Crystal structure of a complex between PTP1B and the insulin receptor tyrosine kinase |
| 2AUH | Crystal structure of the Grb14 BPS region in complex with the insulin receptor tyrosine kinase |
| 1RQQ | Crystal Structure of the Insulin Receptor Kinase in Complex with the SH2 Domain of APS |
| 1LK2 | 1.35A crystal structure of H-2Kb complexed with the GNYSFYAL peptide |
| 1P14 | Crystal structure of a catalytic-loop mutant of the insulin receptor tyrosine kinase |
| 1I44 | CRYSTALLOGRAPHIC STUDIES OF AN ACTIVATION LOOP MUTANT OF THE INSULIN RECEPTOR TYROSINE KINASE |
| 1GAG | CRYSTAL STRUCTURE OF THE INSULIN RECEPTOR KINASE IN COMPLEX WITH A BISUBSTRATE INHIBITOR |
| 1IR3 | PHOSPHORYLATED INSULIN RECEPTOR TYROSINE KINASE IN COMPLEX WITH PEPTIDE SUBSTRATE AND ATP ANALOG |
| 1IRK | CRYSTAL STRUCTURE OF THE TYROSINE KINASE DOMAIN OF THE HUMAN INSULIN RECEPTOR |
Table 2.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary files icod.pdb and nati.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
Table 2.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary files icod.pdb and nati.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
| PDB file name | Residue A | Atom A | Residue B | Atom B | Distance (Å) |
|---|---|---|---|---|---|
| icod.pdb | C_LYS_921 | NZ | F_GLU_1804 | OE1 | 3.659 |
| icod.pdb | F_HIS_1799 | NE2 | F_GLU_1804 | OE2 | 3.669 |
| nati.pdb | C_LYS_921 | NZ | F_GLU_1804 | OE2 | 3.204 |
Table 3.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary file nati.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
Table 3.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary file nati.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
| PDB file name | Residue A | Atom A | Residue B | Atom B | Distance (Å) |
|---|---|---|---|---|---|
| nati.pdb | C_LYS_921 | NZ | F_GLU_1804 | OE2 | 3.204 |
Table 4.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary file icod.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
Table 4.
Interfacial salt bridging network analysis [4] of the structural model of the insulin icodec bound to IR (supplementary file icod.pdb). In this table, the residue naming scheme is Chain ID_residue name_residue number.
| PDB file name | Residue A | Atom A | Residue B | Atom B | Distance (Å) |
|---|---|---|---|---|---|
| icod.pdb | C_LYS_921 | NZ | F_GLU_1804 | OE1 | 3.659 |
| icod.pdb | F_HIS_1799 | NE2 | F_GLU_1804 | OE2 | 3.669 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



