Submitted:
15 November 2023
Posted:
16 November 2023
You are already at the latest version
Abstract
Studies show Human Milk Oligosaccharides (HMOs) influence on infant growth were conflicted. This study aims to determine the relationship between HMO profiles and weight growth indicators of infants ages 0-4 months in Indonesia. A prospective longitudinal study was conducted among 120 mother-infant pairs aged 0 – 4 months from the hospital and Public Health Center in Jakarta between August 2021 – May 2022. The gene analysis for the Secretor and Lewis status of the mother were performed by using targeted Next Generation Sequencing (NGS) and Sanger at baseline. The HMO profiles (2'FL, LNFP I, LNT, LNnT, 3'SL, and 6'SL), breast milk calories, and fat were examined at 0, 2, and 4 months. Weight for age z-score, weight for length z-score, infection episode, and mother's body mass index were examined every month. Data were analyzed using Pearson or Spearman correlation test (correlation coefficient), general linear model for repeated measures ANOVA test and Friedman test (mean difference between and within groups, respectively) and logistic regression unadjusted OR (association between dependent and independent variables), with a p-value <0.05 considered statistically significant. Infants' weight velocities were higher among high LNT and 3'SL levels in the second month, and in high 2'FL and LNFP I mothers in the fourth month. HMO profile (2’FL, LNFP I, and LNT) of lactating mothers have a positive relationship with weight growth indicators, and innovative interventions to enhance HMO concentration among mothers should be considered as one of the ways to improve infant growth.
Keywords:
human milk oligosaccharides
; growth indicator
; secretor status
1. Introduction
Human breast milk comprises carbohydrates, protein, lipids, vitamins, minerals, digestive enzymes, and hormones. Additionally, it contains numerous immune cells, including macrophages, stem cells, and bioactive molecules [1]. One example of a bioactive substrate is human milk oligosaccharides (HMOs), which consist of a diverse range of structures. HMOs are the third most prevalent category of molecules in human milk, following lactose (70 g/L) and lipids (40 g/L) [2]. The concentration of HMOs has been documented to range from 20 to 25 g/L in colostrum [3], gradually decreasing to 5-25 g/L over a six-month breastfeeding period [4]. It is predicted that over 200 HMOs probably exist, with only approximately 100 fully characterized. Initially referred to as the 'Bifidus factor' in human milk, HMOs have mainly been acknowledged for their capacity to promote the growth of bifidobacteria [2].
According to a study by Larsson et al [5], there was a positive correlation between infants' weight gain velocity and the concentrations of HMOs, and the amount of fucose bound to HMOs (p = 0.025). Additionally, the presence of 2'-fucosyllactose (2'FL) in human milk was also positively associated with weight gain velocity during the same period. Conversely, the levels of lacto-N-neotetraose (LNnT) were found to be lower in the group of infants with high weight gain. Another investigation conducted in The Gambia, including 33 pairs of mothers and infants, demonstrated the value of 3′-sialyl lactose (3’SL) as a reliable marker for assessing the weight-for-age z score (WAZ) in infants. Nevertheless, the findings of this investigation also show that lacto-N-fucopentaose I (LNFP I) made a significant favorable impact on the height-for-age z score (HAZ) at 20 weeks (p = 0.05) [6]. In contrast to the findings of Sprenger et al. [7], this study did not detect any statistically significant differences in body weight, body length, body mass index, and head circumference among infants aged up to four months who were fed breast milk containing either low or high concentrations and compositions of gen α-1-2-fucosyltransferase (FUT2)-associated HMOs.
The influence of genetics on the characteristics of HMOs has been widely acknowledged [2]. The composition of HMOs is greatly affected by four different maternal phenotypes, which are determined by the expression profiles of two specific genes: the α-1-2-fucosyltransferase (FUT2) gene and the α-1-3-4-fucosyltransferase (FUT3) gene. The FUT2 gene is responsible for distinguishing individuals with secretor positive (Se+) and secretor negative (Se-) status, whereas the FUT3 gene is indicative of the Lewis Group (Le+ or Le-) [2]. Women defined as Se+Le+ show higher concentrations, more complex profiles, and secrete all HMOs compared to women who are Se−Le− [2]. On the contrary, people identified as non-secretors lack the FUT2 enzyme, leading to a milk composition that either lacks 2'FL or has it in minimum quantities [8]. Though there have been reports of regional and racial variations, it is estimated that 20% of people worldwide have dormant secretor genes (secretor negative) [9].
HMOs are classified into three categories [10]: (1) neutral fucosylated HMOs, e.g., 2’FL and LNFP I; (2) neutral non-fucosylated HMOs, e.g., lacto-N-tetraose (LNT) and LNnT; and (3) acidic sialylated HMOs, e.g., 3’SL and 6’sialyllactose (6’SL). Neutral fucosylated compounds represent 35–50% of the total HMO content; neutral non-fucosylated compounds represent 42–55%; and acidic sialylated compounds represent 12–14% of the total HMOs. Only the FUT2 gene is relevant to the synthesis of 2’FL, as it is the most abundant HMO, with 2.38 g/L on average [11].
Both the World Health Organization (WHO) and the United Nations Children’s Fund (UNICEF) recommend exclusive breastfeeding during the first six months postpartum and continued breastfeeding until 24 months of age [12]. Whereas the European Society for Paediatric Gastroenterology, Hepatology, and Nutrition (ESPGHAN) has formulated recommendations that exclusive or full breastfeeding should be promoted for at least four months and exclusive or predominant breastfeeding for approximately six months as a desirable goal [13].
Environmental, genetic, and hormonal factors influence infant growth [14]. Frequent illness can impair healthy growth, with infections being the most common in the first two years of life, especially diarrhea and respiratory infections. In 2006, the WHO released child growth standards from birth to five years of age to describe the growth of healthy infants who were exclusively or predominantly breast-fed for at least three months [15]. The weight growth standards refer to the WAZ, weight-for-length (WLZ), and weight velocity.
Studies showed that HMOs are essential during the development of the newborn. The mechanisms of action of HMOs include stimulation of the growth of commensal bacteria in the gut [16], antimicrobial and antiviral activity [17], mucosal barrier maturation, modulation of pathogen recognition, and effects on immune function [18]. HMOs have recently been linked to growth in early infancy, assuming the HMOs have potentially positive impact on the microbiota and the immune system of the infant [5].
The relationship between HMO profiles and infant growth, especially in the critical period in the first months of life, is assumed to influence later life. Based on the explanation above, this study will elaborate on the relationships between breast milk HMO profiles and weight growth indicators in Indonesian infants aged 0–4 months.
2. Materials and Methods
2.1. Study Design, Participants and Milk Sampling
This report was a follow-up study on recently published data [19]. The study was conducted in private women's and children's hospitals and public health centers in South Jakarta between August 2021 and May 2022. The study flow chart is depicted in Fig. 1. One hundred and twenty mother-infant pairs were enrolled between the infant’s birth and one month of age. Inclusion criteria were singleton birth, birth with an APGAR score ≥7 in the first minute and 9/10 in five minutes, delivery at the gestational age of >37 weeks, birth weight >2500 g, receiving exclusive breast milk in the first three days of life, mothers aged between 21 to 35 years, and mothers willing to breastfeed for a minimum of four months. Exclusion criteria were: congenital illness or malformation that could affect growth; history of type 1 or 2 diabetes before and after pregnancy; history of gestational diabetes; presenting conditions that contraindicate breastfeeding; and smoking.
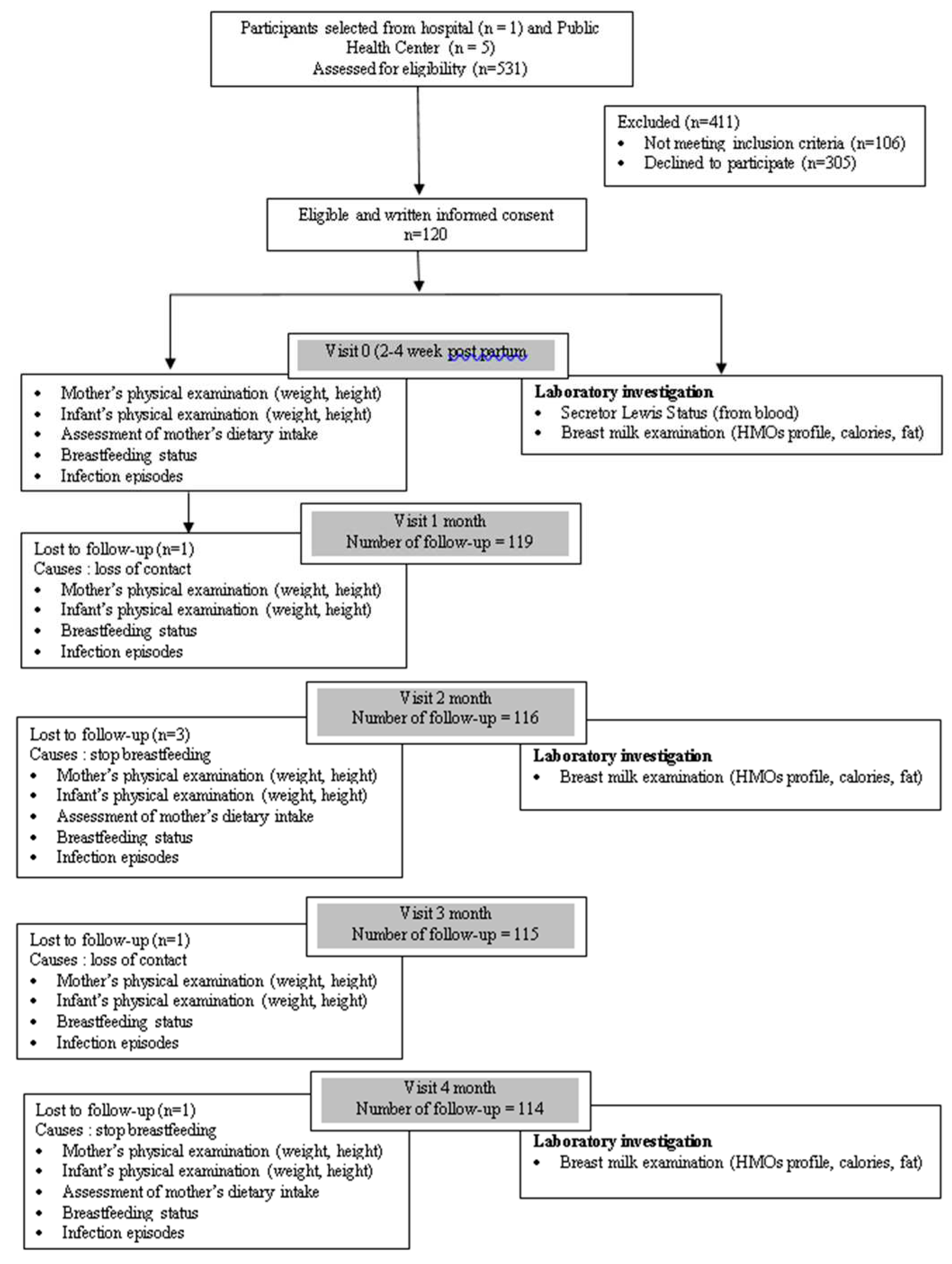
Figure 1.
Study flow chart of the prospective longitudinal study.
To exclude even minor diurnal effects, the milk samples were generally collected during the morning hours between 6:00 and 11:00 a.m [20]. Samples were taken approximately 1½ hours after the last feeding. Beforehand, mothers were trained on how to collect the breastmilk by hand or pump and encouraged to wash their hands before the procedure. Breast skin was cleaned with warm water to reduce bacterial concentration, and the first drops were discharged [21]. Full milk expression from a single breast was collected and homogenized, and a 10 mL sample was taken for analysis, with the remainder being returned to the mother [22]. The breast milk was collected into a 15-mL polypropylene tube, stored at -18°C in a box with dry ice, transferred to the laboratory, and then stored at -80 °C until analysis [23].
As representatives of the FUT2-dependent HMOs, we measured 2’FL and LNFP I; as representatives of the sialylated HMOs, we measured 3’SL and 6’SL; and as representatives of the HMO core structures that can be further decorated with sialic acid or fucose, we measured LNT and LNnT. Authentic reference standards 2’FL, LNFP I, LNT, LNnT, 3’SL, and 6’SL were purchased from Biosynth Carbosynth, England. The HMO analysis was done in the District Health Laboratory Jakarta using the liquid chromatography-mass spectroscopy (LC-MS/MS) method. The breast milk calories and fats were also analyzed. The mothers provided breast milk samples three times: at baseline (2-4 weeks), second, and fourth months of age.
The genotyping assessment was done in the Integrated Laboratorium Faculty of Medicine at Universitas Indonesia. Targeted Next Generation Sequencing (NGS) by the MiSeq system Illumina was used to analyze the FUT2 and FUT3 genes. Thirty samples (25%) need to be reassessed with the Sanger sequencing method. The peripheral blood was collected in tubes containing ethylene diamine tetraacetic acid, and the white blood cells were isolated. This study used the DNA Sequencing Library Prep Kit (CUST-SEQ). FUT Library Prep, a customized kit to purify blood genomic DNA, target and amplify human genes FUT2 and FUT3, performed the amplicons’ end repair, ligated the amplicons with dual index adapters, and purified and enriched the prepared amplicons library compatible with Illumina sequencing. FUT2 analysis was done by examination of the rs1047781 (A385T), while FUT3 analysis was done by examination of the rs28362459 (T59G) and the rs3745635 (G508A).
Infant weight assessment was measured with a mobile digital baby scale, Seca 334. The infant was placed in the middle of the scale with no clothes on, and an average of two readings was taken, and measurements were read to the nearest 0.1 kg. Infant length was measured with a measuring rod for baby scale Seca 233, which is attached to baby scale Seca 334. Two people are required to do the measurement. No shoes, socks, or headgear should be worn. One person holds the crown of the head against the headboard, and the Frankfort plane forms a 900-degree angle with the backboard. This person also ensures that the head, shoulders, and buttocks touch the backboard or flat board. The other person keeps the legs straight against the backboard and slides the footboard against the bottom of the feet with the toes pointing upwards; if having difficulty, the leg(s) can be pressed down at the knee to flex the foot up, and one leg can also be used. An average of three readings is taken, and measurements are read to the nearest 0.1 cm. For WAZ, WLZ, and weight velocity assessment, the weight and length data were compared with the WHO growth chart for WAZ, WLZ, and growth velocity.
Mother’s weight was measured with Seca 876 flat scales for mobile use. The mothers were asked to remove their footwear (shoes or sandals) and headgear (hats, caps, or hair bows). The weight was recorded in kilograms and measured with Seca 217. The mothers were asked to stand on the board, facing the examiner. The mothers were asked to stand with feet together, heels against the backboard, and knees straight, look straight ahead, and not look up. The measurement was moved gently down onto the head of the mother. The mother was asked to breathe in and stand tall—the reading of height in centimeters at the exact point The mothers were asked to step away from the measuring board, and the height measurement was recorded in centimeters. The BMI was calculated based on weight and height measurements. Dietary intake was assessed by a two-day food record and recall before breast milk collection and was analyzed with Nutrisurvey by training dietision.
2.2. Statistical Analysis
All data was edited, coded, and entered into the computer. SPSS for Windows version 26.0 was used for data analysis. A univariate analysis will be conducted to determine the distribution of each variable. The test of normality of the data was performed with the Kolmogorov-Sminov test at the significance level of p > 0.05 as normally distributed data. The continuous data will be presented as mean±standard deviation and for categorical data in n (%) for normally distributed data and geometric mean±standard deviation for non-normally distributed data. For bivariate analysis, a Pearson correlation test will be performed if the data is normally distributed or a Spearman rank test if the data is not normally distributed. The Mann-Whitney test was used to compare the means of two non-normal distribution groups. For repeated measurement, the means of groups were analyzed with repeated ANOVA tests if the data were normally distributed or Friedman if the data were non-normally distributed. Simple logistic regression was used to measure the relationship between categorical maternal and infant factors and categorical dependent variables (HMO profiles and growth indicators). A general linear model for repeated measures was used to observe the difference between independent and dependent variables at 0, 2, and 4 months.
3. Results
3.1. Baseline Characteristics of the Prospective Study
3.2. Relationship between HMO Profiles and Weight Growth Indicators of Infants Aged 0–4 Months
When the statistical analysis with Spearman’s correlation test was performed on all the data, no significance was found between the HMO profile and weight growth indicator. In the secretor positive group, in the baseline between 2’FL and WAZ, there tended to be a correlation (p = 0.089) and reached a statistically significant negative, weak correlation (r = -0.294, p = 0.016) in the second month. Other results tend to show that a correlation was found between LNFP I with WLZ at baseline (p = 0.082), 2’FL with WLZ in the second month (p = 0.063), and between LNnT with WLZ in the fourth month (p = 0.079). In the weak secretor group, in the second month, a significant weak positive correlation was found between 2’FL and WLZ (r=0.338, p = 0.018). In the fourth month, a significant weak positive correlation was found between 2’FL with WAZ (r = 0.359, p = 0.013 and between 2’FL with WLZ (r=0.292, p = 0.046), as shown in Table 2.
In baseline (Table 3), there was a significant negative moderate correlation between 2’FL and WAZ (r = -0.525, p = 0.025, also a significant positive moderate correlation between LNFP I (r=0.478, p = 0.045) with WLZ. In the second month, LNT and WLZ tend to correlate with a p-value of 0.066 (borderline significant). In the fourth month, none of the HMOs were significantly associated ed with weight growth outcomes. Other types of HMOs were not significantly correlated with any weight growth parameters at baseline, the second month, or the fourth month.
Table 4 showed a significant relationship between 2’FL in the fourth month and weight velocity (p = 0.041) with a general linear model for repeated measures analysis. There also tends to be a relationship between 2’FL and WAZ in the fourth month (p = 0.057) and between LNFP I and WLZ in the fourth month (p = 0.078).
3.3. HMO Profiles During Follow-up
Figure 2A,B show the changes in numeric variables during follow-up. Through 4 months of follow-up, all six HMOs (2’FL, LNT, LNnT, LNFP I, 3’SL, and 6’SL) reached statistical differences, with all p=<0.001. Fat intake of mothers came significance with a p-value of 0,030. Breastmilk calories are also significantly different, with a p-value of 0.002. While all growth indicators (WAZ, WLZ, and weight velocity) have p = 0.039, <0.001, and <0.001.
3.4. The Difference in HMO Profiles between Secretor Status
A Mann-Whitney test was performed to analyze the difference in HMO profiles between the secretor positive and the weak secretor, as shown in Table 5. Consistency was demonstrated in the 2’FL value with p-values of 0.014, 0.082, and 0.013 in the baseline, second, and fourth months. Others also showed a significant association between secretor status and LNFP I at baseline (p = 0.060); in the second month, LNnT (p = 0,064) and LNnT (p = 0.060).
4. Discussion
4.1. Secretor Status
The allele frequency of FUT2 and FUT3 gene SNPs is dependent on ethnicity [24]. Many published studies have determined secretor status in the presence or near absence of 2'FL using blood and saliva samples with hemagglutination inhibition techniques [9,25]. The prevalence status from this study is based on rs1047781 because these SNPs are most representative in East Asian and South East Asia and most likely represent a broad ethnicity in Indonesia. The NCBI dbSNP determined wild-type AA (70 mothers, 58.3%) as secretors and AT (50 mothers, 41.7%) as weak secretors, so the total secretor status in this study was 100%. FUT2/FUT3 genotyping is the most accurate method for distinguishing phenotypes [26]. Secretor-gene-specific antigen expression may not fully develop in newborns during the first few months postpartum. Therefore, A, B, and H-secretor typing were not performed for the respective babies, as seen in this study.
Targeted NGS was used in this study to determine FUT2 and FUT3. Thirty blood samples (25%) required further analysis using the Sanger technique as recommended. The targeted NGS for genotyping analysis was customized. During the analytical process, there might be changes in indexing that disrupt the reading analysis. The targeted NGS was also the first done in the HMOs study, different from other studies that used less simple methods [21]. An absence of H antigen in salivary secretions and on mucosal surfaces of the body characterizes non-secretors [27]. In European and African populations, the most common inactivating variant is rs601338 (W154X), whereas, in Asian populations, the missense SNP rs1047781 leads to a truncated protein with weak bioactivity [28].
4.2. Relationship between HMO Profiles and Weight Growth Indicators in Infants Aged 0–4 Months
As mentioned in earlier studies, the six HMO profiles chosen to be examined in this study were based on their effect on growth. If calculated, the sum of these six types of HMOs in mature milk accounts for 30–50% of total HMOs. This type of HMO has also recently been produced industrially and added to milk formula.
This study showed a significant relationship between 2’FL in the fourth month and weight velocity (p = 0.041) with a general linear model for repeated measures analysis. There also tends to be a relationship between 2’FL and WAZ in the fourth month (p = 0.057) and between LNFP I and WLZ in the fourth month (p = 0.078). The analysis in the secretor positive group showed that in the baseline between 2’FL and WAZ, there tended to be a correlation and reached a statistically significant negative, weak correlation in the second month. There was also a significant positive and weak correlation between LNT and WLZ in the second month. Other results tend to show that a correlation was found between LNFP I with WLZ at baseline, 2’FL with WLZ in the second month, and between LNnT with WLZ in the fourth month.
In the weak secretor group, in the second month, a significant weak positive correlation was found between 2’FL and WLZ. In the fourth month, a significant weak positive correlation was found between 2’FL and WAZ and between 2’FL and WLZ. In the Se+Le group, there was a relationship between 2'FL in WAZ at baseline and between LNFP I and WLZ. The result shows that the secretor status was more impactful than the Lewis status because, although the Lewis status was negative, the secretor, even with less enzyme activity, still impacted the infant's growth.
The study results showed a relationship between HMO profiles and weight growth indicators, especially according to the secretor status. Although the results seem scattered during follow-up, the 2’FL showed the most consistent relationship with the weight growth indicator. Six types of HMOs were assessed compared with three weight growth indicators, making it quite difficult to achieve consistency during follow-up. Not to mention, the value of HMOs also showed changes during follow-up. Another consideration was assumed since the HMO analysis was done for the first time in Indonesia; there were no comparisons for HMO analysis.
A review from Sprenger et al. [29] showed a positive association was found between 2’FL and WAZ, as indicated in a study from Lagstrὄm et al. [30], and with weight velocity in a report from Larsson et al. [5]. Interestingly, theoretically, 2’FL has a positive effect on the growth of infants, but this study showed a contrary result. The one mechanism possible explained by Lee et al. [31] was that 2’FL supplementation significantly decreased body weight gain, energy intake, fat mass, and improved inflammatory profiles [32].
Few studies [5,6,7,29,30] examined the association between HMO concentrations and growth measures in infants. They report being rather heterogeneous, making comparisons quite tricky. Growth in infants is a complex process that involves numerous pathways. Not to mention, we cannot eliminate the possibility of the effect of the COVID-19 infection, which is found in some mothers and infants.
4.3. The Changes in HMOs During Follow-Up
The median concentration of 2’FL, LNT, and LNnT at baseline and the second month is in range in comparison with the study from Soyyilmaz et al. [33]. At the same time, LNFP I, 3’SL, and 6’SL showed lower concentrations. In the fourth month, the median of LNFP I, LNnT, and 6’SL was higher, while the median of 2’FL, LNT, and 3’SL was lower than the study review [33]. The increased synthesis of 3’FL and 3’SL in the mammary gland is likely the result of decreased competition for sugar nucleotide substrates in the absence of active FUT2 enzymes. This absence of FUT2 activity increases the effective GDP-fucose substrate availability and allows for greater rates of fucosylation by other fucosyltransferase enzymes, such as the FUT3 enzyme that produces 3’FL. If not all of the additionally available substrate undergoes alternative fucosylation, increased concentrations of the remaining un-fucosylated precursor oligosaccharides will remain, as with LNT [34].
HMO abundances were prominently affected by stages of lactation and maternal genotype. Total HMO concentrations decline from 20–25 g/L a few days before and after parturition to 5–25 g/L over a six-month lactation period [4] The results of the current studies suggest that concentrations of most HMOs decrease throughout lactation [4]. The average decrease of approximately 40% of the initial HMOS concentration in breast milk occurs during the first three months [35]. In early life, infants have an immature intestinal immune system, making them more vulnerable to infection by opportunistic pathogens in early lactation [36]. The high HMO level in colostrum may protect the infant in this sensitive stage of its development [37]. The high concentration of HMOs in an infant's early stage is essential for growth and immunity requirements [38]. The higher concentration of total HMOs in breast milk in the early postpartum period is consistent with the protective functions of colostrum and early milk when the neonate is immunologically immature, and the gut microbiota is not yet fully established [20]. The fact that these HMOs were stable or increased throughout lactation suggests that they may have essential biological functions that extend beyond the first few months of life.
5. Conclusions
All the HMOs showed a significant decrease over time, especially for 2’FL, where a considerable change was decreased from two to four months. All of the mothers in this study had secretor status (58.3% were secretor positive; 41.7% were weak secretors) and Lewis-positive gene status, respectively. There was a significant relationship between 2’FL in the fourth month and weight velocity (p = 0.041) with a general linear model for repeated measures analysis. There also tends to be a relationship between 2’FL and WAZ in the fourth month (p = 0.057) and between LNFP I and WLZ in the fourth month (p = 0.078).
Author Contributions
V.S. was the principal investigator. Conceptualization, B.H., and V.S.; methodology, A.H.; software, V.S.; validation, V.S., and B.H.,; formal analysis, A.H.; investigation, V.S, A., and N.S.M.; resources, Z.M., and D.S.,; data curation, V.S., and B.H.; writing—original draft preparation, writing—review and editing, V.S., and B.H.; supervision, D.S., N.S.M., Z.M., A., and A.H.; project administration, V.S.; funding acquisition, B.H. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by Wyeth Sduenam.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Committee for Research Ethics, Faculty of Medicine, Universitas Indonesia (No. KET-738/UN2.F1/ETIK/PPM.00.02/2020) and registered at www.ClinicalTrial.gov (NCT04515264).
Informed Consent Statement
Written informed consent has been obtained from the patients to publish this paper.
Acknowledgments
The authors like to thank the subjects for their participation.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Martin, C.; Ling, P.R.; Blackburn. G. Review of Infant Feeding: Key Features of Breast Milk and Infant Formula. Nutrients 2016, 118(5), 279. [CrossRef]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 1;22(9), 1147–62. [CrossRef]
- Gabrielli, O.; Zampini, L.; Galeazzi, T; Padella, L.; Santoro, L.; Peila, C. et al. Preterm Milk Oligosaccharides During the First Month of Lactation. PEDIATRICS 2011, 1; 128(6), e1520–31. [CrossRef]
- Austin, S.; De Castro, C.; Bénet, T.; Hou, Y.; Sun, H.; Thakkar, S. et al. Temporal Change of the Content of 10 Oligosaccharides in the Milk of Chinese Urban Mothers. Nutrients 2016, 8;8(6), 346. [CrossRef]
- Larsson, M.W.; Lind, M.V.; Laursen, R.P.; Yonemitsu, C.; Larnkjær, A.; Mølgaard, C. et al. Human Milk Oligosaccharide Composition Is Associated With Excessive Weight Gain During Exclusive Breastfeeding—An Explorative Study. Front Pediatr, 2019, 18;7, 297. [CrossRef]
- Davis, J.C.C.; Lewis, Z.T.; Krishnan, S.; Bernstein, R.M, Moore, S.E.; Prentice, A.M. et al. Growth and Morbidity of Gambian Infants are Influenced by Maternal Milk Oligosaccharides and Infant Gut Microbiota. Sci Rep, 2017, 7(1), 40466. [CrossRef]
- Sprenger, N.; Lee, L.Y.; De Castro, C.A.; Steenhout, P.; Thakkar, S.K. Longitudinal change of selected human milk oligosaccharides and association to infants’ growth, an observatory, single center, longitudinal cohort study. PLOS ONE, 2017, 9;12(2), e0171814. [CrossRef]
- Sprenger, N.; Odenwald, H.; Kukkonen, A.K.; Kuitunen, M.; Savilahti, E.; Kunz, C. FUT2-dependent breast milk oligosaccharides and allergy at 2 and 5 years of age in infants with high hereditary allergy risk. Eur J Nutr, 2017, 56(3), 1293–301. [CrossRef]
- Saboor, M.; Ullah, A.; Qamar, K.; Mir, A.; and Moinuddin. Frequency of ABH secretors and non secretors: A cross sectional study in Karachi. Pak J Med Sci, 2014, 30(1), 189–93. [CrossRef]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum Dev, 2015, 91(11), 619–22. [CrossRef]
- Castanys-Muñoz, E.; Martin, M.J.; Prieto, P.A. 2’-fucosyllactose: an abundant, genetically determined soluble glycan present in human milk. Nutr Rev, 2013, 71(12), 773–89. [CrossRef]
- WHO. Infant and young child feeding [Internet]. [cited 2019 Nov 2]. Available from: https://www.who.int/news-room/fact-sheets/detail/infant-and-young-child-feeding.
- Fewtrell, M.; Bronsky, J.; Campoy, C.; Domellöf, M.; Embleton, N.; Fidler Mis, N. et al. Complementary Feeding: A Position Paper by the European Society for Paediatric Gastroenterology, Hepatology, and Nutrition (ESPGHAN) Committee on Nutrition. J Pediatr Gastroenterol Nutr, 2017, 64(1), 119–32. [CrossRef]
- Wei, C.; Gregory, J.W. Physiology of normal growth. Paediatr Child Health, 2009, 19(5), 236–40. [CrossRef]
- WHO Multicentre Growth Reference Study Group. WHO Child Growth Standards: Methods and Development. Length/height-for-age, weightfor-age, weight-for-length, weight-for-height and body mass index-for age: methods and development. [Internet]. Available from: http://www.who.int/childgrowth/standards/technical_report/en/index.html.
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G. et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome, 2015, 3(1), 13. [CrossRef]
- Hoeflinger, J.L.; Davis, S.R.; Chow, J.; Miller, M.J. In Vitro Impact of Human Milk Oligosaccharides on Enterobacteriaceae Growth. J Agric Food Chem, 2015, 63(12), 3295–302. [CrossRef]
- Xiao, L.; van’t Land, B.; Engen, P.A.; Naqib, A.; Green, S.J.; Nato, A. et al. Human milk oligosaccharides protect against the development of autoimmune diabetes in NOD-mice. Sci Rep, 2018, 8(1), 3829. [CrossRef]
- Sudarma, V.; Sunardi, D.; Marzuki, N.S.; Munasir, Z.; Asmarinah.; Hidayat, A.; Hegar, B. Human Milk Oligosaccharide Profiles and the Secretor and Lewis Gene Status of Indonesian Lactating Mothers. Pediatr Gastroenterol Hepatol Nutr, 2023, 26(5), 266-276. [CrossRef]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J. et al. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br J Nutr, 2010, 14;104(9), 1261–71. [CrossRef]
- Cabrera-Rubio, R.; Kunz, C.; Rudloff, S.; García-Mantrana, I.; Crehuá-Gaudiza, E. Martínez-Costa, C. et al. Association of Maternal Secretor Status and Human Milk Oligosaccharides With Milk Microbiota: An Observational Pilot Study. J Pediatr Gastroenterol Nutr, 68(2), 256–63. [CrossRef]
- Austin, S.; De Castro, C.A.; Sprenger, N.; Binia, A.; Affolter, M.; Garcia-Rodenas, C.L. et al. Human Milk Oligosaccharides in the Milk of Mothers Delivering Term versus Preterm Infants. Nutrients, 2019, 5;11(6), 1282. [CrossRef]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J. et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J Nutr, 2018, 1;148(11), 1733–42. [CrossRef]
- Günaydın, G.; Nordgren, J.; Sharma, S.; Hammarström, L. Association of elevated rotavirus-specific antibody titers with HBGA secretor status in Swedish individuals: The FUT2 gene as a putative susceptibility determinant for infection. Virus Res, 2016, 211, 64–8. [CrossRef]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Boekhorst, J.; Schneeberger, S.; Karanja, S. et al. Maternal Human Milk Oligosaccharide Profile Modulates the Impact of an Intervention with Iron and Galacto-Oligosaccharides in Kenyan Infants. Nutrients, 2019, 29;11(11), 2596. [CrossRef]
- Vandenplas, Y.; Berger, B.; Carnielli, V.; Ksiazyk, J.; Lagström, H.; Sanchez Luna, M. et al. Human Milk Oligosaccharides: 2′-Fucosyllactose (2′-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients, 2018, 24;10(9), 1161. [CrossRef]
- Williams, J.E.; McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Brooker, S.L.; Kamau-Mbuthia, E.W. et al. Key genetic variants associated with variation of milk oligosaccharides from diverse human populations. Genomics, 2021, 113(4), 1867–75. [CrossRef]
- Silva, L.M.; Carvalho, A.S.; Guillon, P.; Seixas, S.; Azevedo, M.; Almeida, R. et al. Infection-associated FUT2 (Fucosyltransferase 2) genetic variation and impact on functionality assessed by in vivo studies. Glycoconj J, 2010, 27(1), 61–8. [CrossRef]
- Sprenger, N.; Tytgat, H.L.P.; Binia, A.; Austin, S.; Singhal, A. Biology of human milk oligosaccharides: From basic science to clinical evidence. J Hum Nutr Diet, 2022, 35(2), 280–99. [CrossRef]
- Lagström, H.; Rautava, S.; Ollila, H.; Kaljonen, A.; Turta, O.; Mäkelä, J. et al. Associations between human milk oligosaccharides and growth in infancy and early childhood. Am J Clin Nutr, 2020, 1;111(4), 769–78. [CrossRef]
- Lee, S.; Goodson, M.; Vang, W.; Kalanetra, K.; Barile, D.; Raybould, H. 2′-Fucosyllactose Supplementation Improves Gut-Brain Signaling and Diet-Induced Obese Phenotype and Changes the Gut Microbiota in High Fat-Fed Mice. Nutrients, 2020, 5;12(4), 1003. [CrossRef]
- Lee, S.A.; Lim, J.Y.; Kim, B.S.; Cho, S.J.; Kim, N.Y.; Kim, O.B. et al. Comparison of the gut microbiota profile in breast-fed and formula-fed Korean infants using pyrosequencing. Nutr Res Pract, 2015;9(3),242. [CrossRef]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients, 2021, 9;13(8), 2737. [CrossRef]
- Samuel, T.M.; Binia, A.; de Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M. et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci Rep, 2019, 9(1), 11767. [CrossRef]
- Albrecht, S.; Schols, H.A.; van den Heuvel, EGHM.; Voragen, A.G.J.; Gruppen, H. Occurrence of oligosaccharides in feces of breast-fed babies in their first six months of life and the corresponding breast milk. Carbohydr Res, 2011, 346(16), 2540–50. [CrossRef]
- Labbok, M.H.; Clark, D.; Goldman, A.S. Breastfeeding: maintaining an irreplaceable immunological resource. Nat Rev Immunol, 2004, 4(7), 565–72. [CrossRef]
- Kunz, C.; Rudloff, S.; Baier, W.; Klein, N.; Strobel, S. Oligosaccharides in human milk: structural, functional, and metabolic aspects. Annu Rev Nutr 2000, 20, 699–722. [Google Scholar] [CrossRef] [PubMed]
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J. et al. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front Pediatr, 2018, 10;6, 239. [CrossRef]
Figure 2.
2A and 2B. HMOs profiles during follow up; p<0.001 (Friedman test).
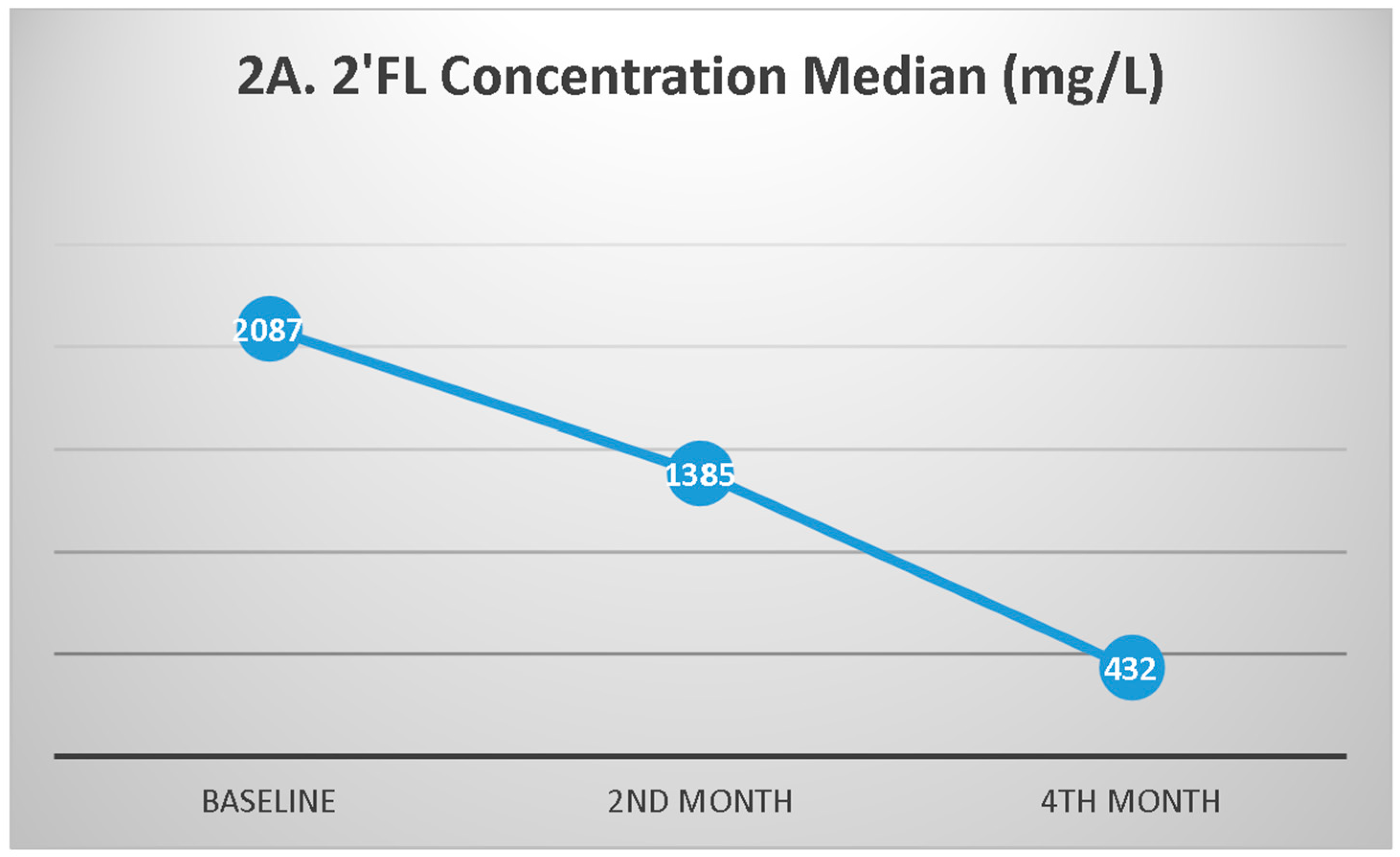
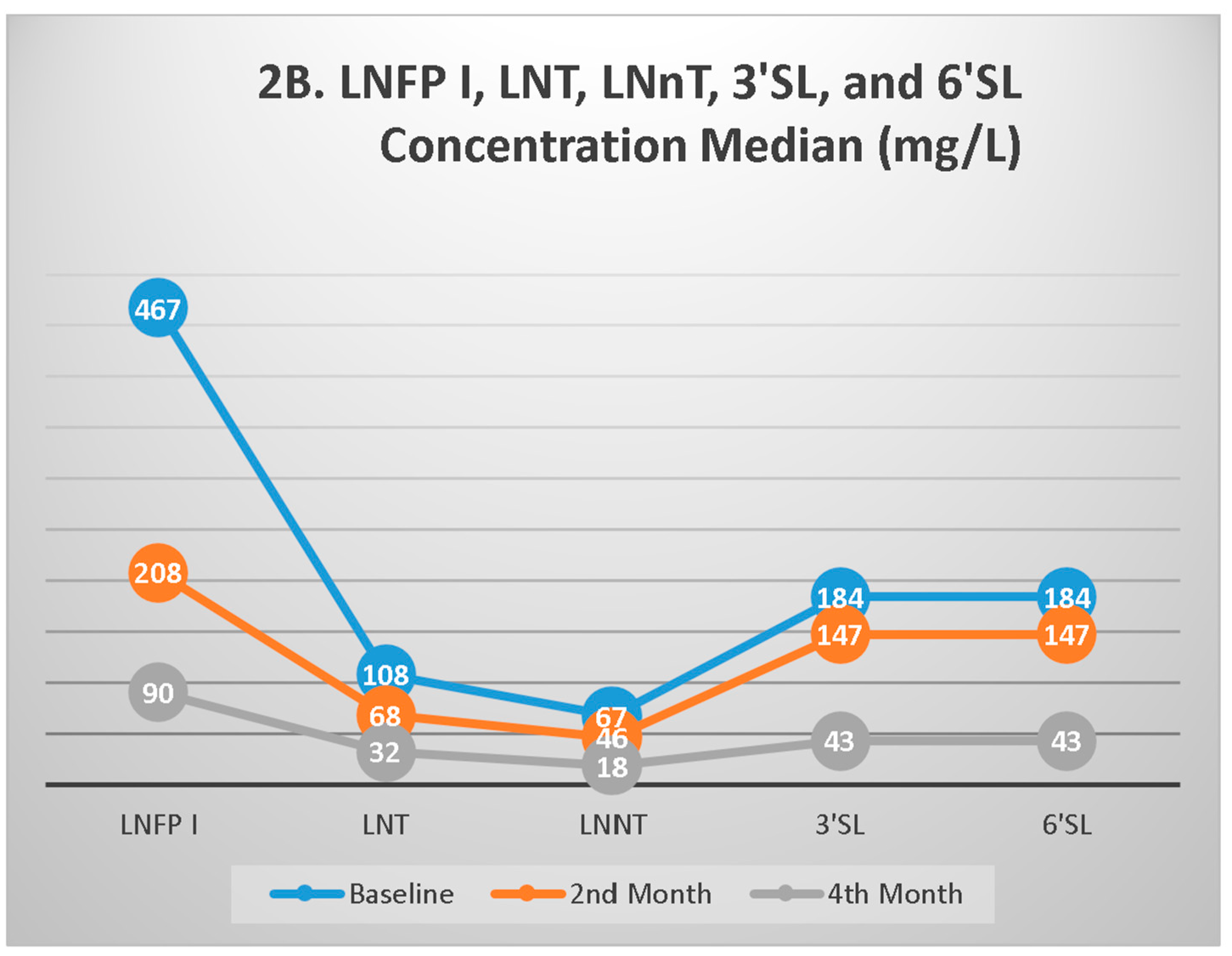

Table 1.
Baseline Characteristics of the Subjects.
| Characteristics | n (%) |
| Maternal variables | |
| Nutritional status | |
| BMI<25 kg/m2 | 79 (65.8) |
| BMI ≥25 kg/m2 | 41 (34.2) |
| Economic status | |
| Low-middle income | 61 (50.8) |
| High-income | 59 (49.2) |
Economic status: low-middle <Rp 1.600.000,-/person/month; high-income: ≥Rp 1.600.000,-/person/month.
Table 2.
The Correlations between HMO Profiles and Weight Growth Indicators of Infants Aged 0 – 4 Months.
Table 2.
The Correlations between HMO Profiles and Weight Growth Indicators of Infants Aged 0 – 4 Months.
| All | Secretor Positive | Weak Secretor | ||||
| r | p-value | r | p-value | r | p-value | |
| WAZ Baseline | ||||||
| 2’FL | -0.145 | 0.113 | -0.205 | 0.089 | -0.029 | 0.840 |
| LNFP I | -0.068 | 0.459 | -0.971 | 0.558 | 0.021 | 0.883 |
| LNT | 0.048 | 0.601 | 0.098 | 0.419 | -0.010 | 0.944 |
| LNnT | 0.060 | 0.517 | 0.061 | 0.618 | 0.092 | 0.527 |
| 3’SL | 0.043 | 0.641 | 0.150 | 0.216 | -0.066 | 0.650 |
| 6’SL | 0.053 | 0.567 | 0.164 | 0.175 | -0.067 | 0.643 |
| WAZ 2nd month | ||||||
| 2’FL | -0.131 | 0.162 | -0.294 | 0.016* | 0.142 | 0.331 |
| LNFP I | -0.094 | 0.313 | -0.157 | 0.204 | -0.091 | 0.536 |
| LNT | 0.036 | 0.701 | 0.083 | 0.506 | -0.101 | 0.489 |
| LNnT | 0.016 | 0.865 | -0.005 | 0.968 | -0.061 | 0.680 |
| 3’SL | 0.018 | 0.850 | -0.071 | 0.570 | 0.105 | 0.471 |
| 6’SL | 0.014 | 0.879 | -0.074 | 0.550 | 0.104 | 0.478 |
| WAZ 4th month | ||||||
| 2’FL | 0.028 | 0.768 | -0.157 | 0.204 | 0.359 | 0.013* |
| LNFP I | -0.019 | 0.839 | 0.016 | 0.895 | -0.052 | 0.728 |
| LNT | -0.006 | 0.947 | 0.071 | 0.568 | -0.157 | 0.291 |
| LNnT | 0.030 | 0.751 | 0.003 | 0.978 | -0.043 | 0.774 |
| 3’SL | -0.089 | 0.344 | -0.063 | 0.613 | -0.113 | 0.450 |
| 6’SL | -0.080 | 0.400 | -0.061 | 0.623 | -0.089 | 0.552 |
| WLZ 0 month | ||||||
| 2’FL | -0.033 | 0.720 | -0.033 | 0.786 | 0.044 | 0.759 |
| LNFP I | -0.081 | 0.381 | -0.209 | 0.082 | 0.150 | 0.298 |
| LNT | 0.001 | 0.993 | -0.030 | 0.807 | 0.004 | 0.977 |
| LNnT | -0.032 | 0.726 | -0.067 | 0.579 | 0.047 | 0.745 |
| 3’SL | 0.038 | 0.676 | 0.064 | 0.596 | -0.029 | 0.842 |
| 6’SL | 0.034 | 0.712 | 0.059 | 0.629 | -0.025 | 0.863 |
| WLZ 2nd month | ||||||
| 2’FL | 0.002 | 0.984 | -0.229 | 0.063 | 0.338 | 0.018* |
| LNFP I | -0.002 | 0.819 | 0.273 | 0.437 | -0.171 | 0.240 |
| LNT | 0.105 | 0.262 | 0.131 | 0.025* | -0.080 | 0.587 |
| LNnT | 0.044 | 0.636 | 0.096 | 0.289 | -0.070 | 0.631 |
| 3’SL | 0.043 | 0.684 | 0.115 | 0.352 | 0.022 | 0.879 |
| 6’SL | 0.031 | 0.742 | 0.101 | 0.416 | 0.020 | 0.894 |
| WLZ 4th month | ||||||
| 2’FL | 0.040 | 0.672 | -0.127 | 0.306 | 0.292 | 0.046* |
| LNFP I | 0.072 | 0.449 | 0.163 | 0.188 | -0.054 | 0.718 |
| LNT | 0.069 | 0.468 | 0.198 | 0.107 | -0.148 | 0.321 |
| LNnT | 0.118 | 0.210 | 0.216 | 0.079 | -0.004 | 0.981 |
| 3’SL | 0.062 | 0.511 | 0.108 | 0.386 | -0.004 | 0.979 |
| 6’SL | 0.053 | 0.574 | 0.108 | 0.384 | -0.029 | 0.844 |
|
WV 2nd month (total g/2 mo) |
||||||
| 2’FL | -0.045 | 0.663 | -0.161 | 0.193 | 0.062 | 0.672 |
| LNFP I | -0.091 | 0.330 | -0.143 | 0.249 | -0.105 | 0.474 |
| LNT | -0.016 | 0.867 | 0.048 | 0.700 | -0.174 | 0.231 |
| LNnT | -0.020 | 0.833 | -0.011 | 0.929 | -0.156 | 0.285 |
| 3’SL | 0.111 | 0.233 | -0.001 | 0.991 | -0.016 | 0.914 |
| 6’SL | 0.101 | 0.281 | -0.005 | 0.968 | -0.012 | 0.933 |
Abbreviations: WAZ: Weight for age Z score; WLZ: weight for length Z score; WV: weight velocity. *p<0.05; Spearman’s correlation test. Baseline data: infants aged 2 – 4 weeks
Table 3.
The Correlation between HMO Profiles and Weight Growth Indicators According to Secretor Lewis Group Status of Infants Aged 0 – 4 Months.
Table 3.
The Correlation between HMO Profiles and Weight Growth Indicators According to Secretor Lewis Group Status of Infants Aged 0 – 4 Months.
| Se+Le+ | Se+Le- | |||
| r | p-value | r | p-value | |
| WAZ Baseline | ||||
| 2’FL | -0.083 | 0.406 | -0.525 | 0.025* |
| LNFP I | -0.098 | 0.328 | 1 | 0.693 |
| LNT | 0.023 | 0.822 | 0.188 | 0.456 |
| LNnT | 0.015 | 0.883 | 0.243 | 0.331 |
| 3’SL | 0.024 | 0.814 | 0.205 | 0.415 |
| 6’SL | 0.034 | 0.738 | 0.208 | 0.408 |
| WAZ 2nd month | ||||
| 2’FL | -0.134 | 0.187 | -0.213 | 0.411 |
| LNFP I | -0.060 | 0.557 | -0.309 | 0.228 |
| LNT | 0.040 | 0.693 | 0.066 | 0.801 |
| LNnT | -0.006 | 0.954 | 0.179 | 0.492 |
| 3’SL | 0.026 | 0.797 | -0.012 | 0.962 |
| 6’SL | 0.024 | 0.815 | -0.011 | 0.996 |
| WAZ 4th month | ||||
| 2’FL | -0.015 | 0.884 | -0.275 | 0.286 |
| LNFP I | -0.009 | 0.927 | -0.027 | 0.918 |
| LNT | 0.009 | 0.932 | -0.081 | 0.758 |
| LNnT | 0.030 | 0.770 | 0.042 | 0.874 |
| 3’SL | -0.113 | 0.272 | -0.015 | 0.995 |
| 6’SL | -0.099 | 0.336 | -0.017 | 0.948 |
| WLZ Baseline | ||||
| 2’FL | -0.040 | 0.690 | -0.007 | 0.977 |
| LNFP I | -0.159 | 0.111 | 0.478 | 0.045* |
| LNT | -0.026 | 0.793 | 0.170 | 0.500 |
| LNnT | -0.043 | 0.670 | -0.017 | 0.946 |
| 3’SL | 0.051 | 0.608 | -0.129 | 0.611 |
| 6’SL | 0.045 | 0.654 | -0.127 | 0.617 |
| WLZ 2nd month | ||||
| 2’FL | 0.018 | 0.860 | -0.255 | 0.323 |
| LNFP I | 0.042 | 0.680 | -0.438 | 0.079 |
| LNT | 0.186 | 0.066 | -0.140 | 0.593 |
| LNnT | 0.106 | 0.297 | -0.092 | 0.726 |
| 3’SL | 0.049 | 0.633 | -0.014 | 0.957 |
| 6’SL | 0.035 | 0.728 | -0.012 | 0.963 |
| WLZ 4th month | ||||
| 2’FL | 0.014 | 0.894 | 0.229 | 0.377 |
| LNFP I | 0.050 | 0.630 | 0.257 | 0.319 |
| LNT | 0.054 | 0.600 | 0.130 | 0.619 |
| LNnT | 0.089 | 0.386 | 0.350 | 0.168 |
| 3’SL | 0.033 | 0.748 | 0.221 | 0.395 |
| 6’SL | 0.025 | 0.804 | 0.223 | 0.390 |
| WV 2nd month (total g/2 mo) | ||||
| 2’FL | -0.062 | 0.540 | -0.037 | 0.888 |
| LNFP I | -0.066 | 0.516 | -0.314 | 0.220 |
| LNT | 0.004 | 0.971 | -0.118 | 0.653 |
| LNnT | 0.012 | 0.904 | -0.007 | 0.978 |
| 3’SL | 0.070 | 0.491 | 0.291 | 0.257 |
| 6’SL | 0.060 | 0.555 | 0.292 | 0.256 |
Abbreviations: WAZ: Weight for age Z score; WLZ: weight for length Z score; WV: weight velocity. Se+Le+: Secretor status positive and weak, Lewis positive. Se+Le-: Secretor status positive and weak, Lewis negative. *p<0.05; Spearman’s correlation test.
Table 4.
General Linear Model for Repeated Measures between HMO Profiles at Second Month and Weight Growth Indicators.
Table 4.
General Linear Model for Repeated Measures between HMO Profiles at Second Month and Weight Growth Indicators.
| HMOs profiles | Mean (CI 95%) | p-value | Mean difference (95% CI) | |
| WAZ | ||||
| 2nd month | 2’FL ≥med | -0.50 (-0.74 – 0.25) | 0.143 | -0.26 (-0.61 – 0.09) |
| 2’FL <med | -0.24 (-0.49 – 0.01) | |||
| 4th month | 2’FL ≥med | -0.54 (-0.8 – 0.28) | 0.057 | -0.36 (-0.73 – 0.01) |
| 2’FL <med | -0.18 (-0.44 – 0.09 | |||
| 2nd month | LNFP I ≥med | -0.45 (-0.70 – 0.21) | 0.333 | -0.17 (-0.52 – 0.18) |
| LNFP I <med | -0.28 (-0.53 – 0.03) | |||
| 4th month | LNFP I ≥med | -0.49 (-0.76 – 0.23) | 0.145 | -0.28 (-0.65 – 0.10) |
| LNFP I <med | -0.22 (-0.48 – 0.05) | |||
| 2nd month | LNT ≥med | -0.39 (-0.64 – 0.15) | 0.793 | -0.05 (-0.40 – 0.30) |
| LNT <med | -0.35 (-0.59 – 0.10) | |||
| 4th month | LNT ≥med | -0.50 (-0.76 – 0.23) | 0.141 | -0.28 (-0.65 – 0.09) |
| LNT <med | -0.22 (-0.48 – 0.05) | |||
| 2nd month | LNnT ≥med | -0.42 (-0.67 – 0.17) | 0.583 | -0.10 (-0.45 – 0.25) |
| LNnT <med | -0.32 (-0.57 – 0.08) | |||
| 4th month | LNnT ≥med | -0.47 (-0.74 – 0.20) | 0.254 | -0.22 (-0.59 – 0.16) |
| LNnT <med | -0.25 (-0.51 – 0.01) | |||
| 2nd month | 3’SL ≥med | -0.36 (-0.61 – 0.11) | 0.914 | 0.02 (-0.33 – 0.37) |
| 3’SL <med | -0.38 (-0.63 – 0.13) | |||
| 4th month | 3’SL ≥med | -0.36 (-0.63 – 0.09) | 0.991 | -0.002 (-0.38 – 0.37) |
| 3’SL <med | -0.36 (-0.62 – 0.10) | |||
| 2nd month | 6’SL ≥med | -0.36 (-0.61 – 0.11) | 0.914 | 0.02 (-0.33 – 0.37) |
| 6’SL <med | -0.38 (-0.63 – 0.13) | |||
| 4th month | 6’SL ≥med | -0.36 (-0.63 – 0.09) | 0.991 | -0.002 (-0.38 – 0.37) |
| 6’SL <med | -0.36 (-0.62 – 0.09) | |||
| WLZ | ||||
| 2nd month | 2’FL ≥med | -0.29 (-0.58 – 0.01) | 0.687 | 0.08 (-0.33 – 0.50) |
| 2’FL <med | -0.38 (-0.67 – 0.08) | |||
| 4th month | 2’FL ≥med | -0.52 (-0.81 – 0.23) | 0.228 | -0.25 (-0.66 – 0.16) |
| 2’FL <med | -0.27 (-0.56 – 0.02) | |||
| 2nd month | LNFP I ≥med | -0.42 (-0.71 – 0.13) | 0.391 | -0.18 (-0.59 – 0.23) |
| LNFP I <med | -0.24 (-0.54 – 0.05) | |||
| 4th month | LNFP I ≥med | -0.58 (-0.86 – 0.29) | 0.078 | -0.37 (-0.77 – 0.04) |
| LNFP I <med | -0.21 (-0.50 – 0.08) | |||
| 2nd month | LNT ≥med | -0.28 (-0.57 – 0.02) | 0.581 | 0.12 (-0.30 – 0.53) |
| LNT <med | -0.39 (-0.68 – 0.1) | |||
| 4th month | LNT ≥med | -0.48 (-0.77 – 0.19) | 0.439 | -0.16 (-0.58 – 0.25) |
| LNT <med | -0.32 (-0.61 – 0.03) | |||
| 2nd month | LNnT ≥med | -0.36 (-0.65 – 0.06) | 0.824 | -0.05 (-0.46 – 0.37) |
| LNnT <med | -0.31 (-0.60 – 0.02) | |||
| 4th month | LNnT ≥med | -0.49 (-0.79 – 0.20) | 0.382 | -0.18 (-0.60 – 0.23) |
| LNnT <med | -0.31 (-0.60 – 0.02) | |||
| 2nd month | 3’SL ≥med | -0.31 (-0.60 – 0.2) | 0.826 | 0.05 (-0.37 – 0.46) |
| 3’SL <med | -0.36 (-0.65 – 0.06) | |||
| 4th month | 3’SL ≥med | -0.39 (-0.69 – 0.10) | 0.969 | 0.008 (-0.41 – 0.42) |
| 3’SL <med | -0.40 (-0.69 – 0.11) | |||
| 2nd month | 6’SL ≥med | -0.31 (-0.60 – 0.02) | 0.826 | 0.05 (-0.37 – 0.46) |
| 6’SL <med | -0.36 (-0.65 – 0.06) | |||
| 4th month | 6’SL ≥med | -0.39 (-0.69 – 0.10) | 0.969 | 0.008 (-0.41 – 0.42) |
| 6’SL <med | -0.40 (-0.69 – 0.11) | |||
| 4th month | LNFP I ≥med | 3404 (3189 – 3618) | 0.202 | -198 (-504 – 108) |
| LNFP I <med | 3602 (3384 – 3820) | |||
| 2nd month | LNT ≥med | 2145 (1971 – 2319) | 0.826 | -27.3 (-274 – 219) |
| LNT <med | 2172 (1998 – 2347) | |||
| 4th month | LNT ≥med | 3401 (3185 – 3617) | 0.198 | -200 (-505 – 106) |
| LNT <med | 3601 (3385 – 3817) | |||
| 2nd month | LNnT ≥med | 2112 (1936 – 2287) | 0.459 | -92.3 (-338 – 154) |
| LNnT <med | 2204 (2032 – 2376) | |||
| 4th month | LNnT ≥med | 3384 (3167 – 3602) | 0.139 | -229 (-534 – 75) |
| LNnT <med | 3614 (3300 – 3827) | |||
| 2nd month | 3’SL ≥med | 2158 (1984 – 2332) | 0.992 | -1.19 (-248 – 245) |
| 3’SL <med | 2159 (1985 – 2334) | |||
| 4th month | 3’SL ≥med | 3498 (3280 - 3715) | 0.968 | -6.23 (-314 – 302) |
| 3’SL <med | 3504 (3286 – 3722) | |||
| 2nd month | 6’SL ≥med | 2158 (1984 – 2332) | 0.992 | -1.19 (-248 – 245) |
| 6’SL <med | 2159 (1985 – 2334) | |||
| 4th month | 6’SL ≥med | 3498 (3280 – 3715) | 0.968 | -6.23 (-314 – 302) |
| 6’SL <med | 3504 (3286 – 3722) |
*p<0.05. Abbreviations: WAZ: Weight for age Z score; WLZ: weight for length Z score; WV: weight velocity. Median baseline (mg/L): 2’FL (2618), LNFP I (466), LNT (107), LNnT (68,3), 3’SL (187), 6’SL (185). Median 2nd month (mg/L): 2’FL (1385), LNFP I (206), LNT (68,2), LNnT (46,1), 3’SL (147), 6’SL (147). Median 4th month (mg/L): 2’FL (431), LNFP I (89.9), LNT (32,2), LNnT (18.1), 3’SL (43.3), 6’SL (43.6). Category of economic status: 1. low-middle <Rp 1.600.000,-; 2. high-income: ≥Rp 1.600.000,-.
Table 5.
Differences in HMOs Profiles during Follow-up between Secretor Status.
| HMOs profiles | Secretor positive Median (IQR) |
Weak Secretor Median (IQR) |
p-value |
| 2’FL | |||
| Baseline | 2288 (764, 4316) | 1900 (48.6, 2670) | 0.014* |
| 2nd month | 1581 (215, 2710) | 870 (49, 2386) | 0.082 |
| 4th month | 554 (156, 759) | 207 (11.5, 540) | 0.013* |
| LNFP I | |||
| Baseline | 590 (164, 1401) | 410 (112, 1002) | 0.060 |
| 2nd month | 198 (131, 446) | 231 (155, 833) | 0.391 |
| 4th month | 96 (25.8, 488) | 60.4 (22.9, 288) | 0.523 |
| LNT | |||
| Baseline | 116 (50, 1808) | 107 (61.4, 223) | 0.484 |
| 2nd month | 63.2 (41.3, 98.1) | 89.7 (40.8, 140.9) | 0.064 |
| 4th month | 31.4 (16.5, 87.1) | 32.6 (21, 62.9) | 0.979 |
| LNnT | |||
| Baseline | 75.6 (36.1, 118.7) | 64.8 (36, 109) | 0.542 |
| 2nd month | 42.2 (30.4, 59.2) | 54.5 (30.2, 76.7) | 0.060 |
| 4th month | 20.3 (9.6, 50) | 15.7 (10.5, 29.1) | 0.392 |
| 3’SL | |||
| Baseline | 185 (132) | 184 (142, 225) | 0.435 |
| 2nd month | 139 (102, 179) | 153 (116, 191) | 0.365 |
| 4th month | 43.4 (39.3, 59.0) | 43.3 (38.6, 51.5) | 0.734 |
| 6’SL | |||
| Baseline | 185 (132, 260) | 184 (142, 223) | 0.546 |
| 2nd month | 139 (102, 178) | 152 (116, 191) | 0.346 |
| 4th month | 43.4 (39.4, 58.9) | 44.2 (38.9, 51.3) | 0.852 |
Values are mean±SDs, Median (IQRs). *p-value <0.05. Statistical analysis was performed with the Mann-Whitney test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



