
Submitted:
15 November 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
Insect behavior is coordinated mainly by smell, through the diverse odor-binding proteins (OBP) that allow them to identify and recognize their environment. Sensory information collected through smell is then analyzed and interpreted in the brain, allowing for correct insect functioning. The behavior of honey bees (Apis mellifera L.) can be affected by different pathogens, such as deformed wing virus (DWV). In particular, DWV variant A (DWV-A) is capable of altering olfactory sensitivity, reducing the gene expression of different OBPs, including those associated with nursing behavior. DWV is also capable of replicating itself in the sensory lobes of the brain, further compromising the processing of sensory information. This study evaluated the behavioral response of nurse honey bees exposed to a pheromone compound and the alterations in the gene expression of pre- and post-synaptic neuronal genes neuroxins-1 and neurogilin-1 in the bee heads and OBP proteins in antennae of nurse bees inoculated with DWV-A. The behavioral response of nurse bees exposed to the larval pheromone compounds benzyl alcohol was analyzed using a Y-tube olfactometer. The viral load, gene expression of OBP5 and OBP11 in antennae and the neuroxins-1 and neurogilin-1 in the bee heads were analyzed by qPCR. High viral loads significantly reduced the ability of 10- and 15-day-old nurse honey bees to choose the correct pheromone compound. Also, the gene expression of OBP5, OBP11, neuroxin-1 and neurogilin-1 in nurse honey bees decreased when they were highly infected with DWV-A. These results suggest that a DWV-A infection can disturb information processing and cause nurse honey bees to reduce their activity inside the hive, altering internal cohesion.
Keywords:
Nurse bees
; DWV-A
; OBPs
; synaptic genes
; olfactory responses
1. Introduction
Bees, particularly Apis mellifera L., have an excellent olfactory system, which modulates highly cooperative social behavior in and outside the hive, such as brood care, foraging, hive maintenance, and cleaning, among other activities. This peripheral sensory system is located mainly in the antennae and is key in the chemosensory detection of pheromones and environmental odorants [1,2]. The sensitivity and specificity of the olfactory system is greatly attributed to the presence of odorant binding proteins (OBPs) [3]. The OBPs are small water-soluble molecules located in the sensory dendrite of the antennae that selectively bind to different environmental compounds [4,5]. The number and diversity of OBPs vary among insect species; in A. mellifera, a total of 21 OBPs have been described, of which only nine are specifically expressed in the antennae [6]. Among these proteins, OBP5 and OBP11 are particularly related to the larval care process inside beehives [7,8]. These OBPs have an affinity for the volatile compounds of the warning pheromone generated by diseased larvae [7,8]. Some of the specific compounds emitted and perceived by these proteins are phenethyl acetate, phenyl ethanol and benzyl alcohol [9]. Of these three compounds, benzyl alcohol is the most competitive compound for the OBP5 protein recognition site [7].
The decline of honey bees and other bee populations has been reported throughout the last decade [10,11], partially associated with diseases and pathogens specific to these species [12]. Viral pathogens are of great relevance because they can remain in the hives without generating detectable clinical symptoms, making their recognition difficult [13,14]. Also, viral diseases in conjunction with different stress factors, such as food quality and availability, climatic conditions and other pathogens, such as varroa destructor Anderson and Trueman, bacteria or fungi can greatly contribute to the loss of bee hives [10,13,14]. One of the most important viral diseases is deformed wing virus (DWV), which characteristically causes wing deformities in infected adult bees [15]. DWV has also proven to cause early mortality in infected bees [15], as well as a long term increase in viral titers, which can generate alterations in olfactory sensitivity and foraging tasks in infected workers [16].
Three epidemiologically important variants of DWV have been described: DWV-A, DWV-B and DWV-C [17,18]. DWV-A has proven to be the predominant variant in Chile [19,20] and is able to replicate itself in the basal regions of the antennal epithelium [21]. It has also been demonstrated that the presence of this pathogen in the antennae of A. mellifera results in a decrease in olfactory sensitivity to different scents of plant species, as well as inducing a decrease in gene expression, specially of OBP5 and OBP11 [22]. In addition, DWV can alter the memory of A. mellifera because this pathogen is capable of replicating itself in brain regions, including the neuropils responsible for vision and smell [23]. Co-infestation with the varroa mite has proven to generate gene depletion of the pre- and postsynaptic proteins neurexin (AmNrx-1) and neuroligin (AmNlg-1), respectively, decreasing information processing and associative memory [24]. Nevertheless, the down-regulation of these genes has only been observed in the interaction with varroa mite; it has not yet been determined whether the virus alone is capable of altering their expression. Therefore, the aim of this study was to evaluate the behavioral response of nurse honey bees exposed to pheromone compounds related to larvae care and also to evaluate the alterations in the gene expressions of the pre- and post-synaptic neuronal genes neuroxins-1 and neurogilin-1 in bee heads and antennae. OBP proteins in the antennae of nurse bees inoculated with DWV-A were also assessed. The results of this study will aid in understanding the effects of DWV-A infections at the individual and social level within in the colony.
2. Materials and Methods
2.1. Inoculum preparation
The deformed wing virus variant A (DWV-A) isolate was obtained from infected colonies following the methodology described by Gusachenko et al. [25]. Briefly, a pool of 20 honey bees was homogenized in a stomacher bag with phosphate buffered saline (1x PBS) for 90s at high speed in stomacher 80 equipment (Seward, London, UK). Samples were centrifuged at 1500 g for 10 min, followed by 10000 g for 10 min, both at 4°C. The supernatant was purified by filtration with a 0.22 µm filter (PES, Merck Millipore, Darmstadt, Germany). Subsequently, 200 µl of the supernatant was used for RNA extraction, cDNA synthesis and the subsequent quantification of the viral load of the inoculum.
2.2. Honey bee inoculation
Adult A. mellifera specimens for the trial were supplied by the experimental apiary located at the experimental center "El Nogal" (36 ° 35′58.25 ″ S - 72 ° 04′51.77 ″ W), Universidad de Concepción, Chillán, Chile. Brood frames were maintained under controlled conditions at 30 ° C ± 1; 60% ± 3 RH. Subsequently, newly emerged bees were carefully collected and randomly confined in plastic cages (base = 8 cm diameter, mouth = 10 cm diameter, and height = 15 cm). The experimental procedure was then carried out in a similar way to that previously published in Silva et al. [22]. In brief, one group of young bees was individually and orally inoculated (I-DWV treatment) with 5 µl of a viral suspension (1.0 × 109 copy number per bee) in a 60% sucrose solution. The bees that did not consume the total viral inoculum suspension were discarded from the experiment. Another group of bees was not inoculated with the viral suspension (N-DWV treatment), which were referred to as a control group. Then, 700 worker bees were maintained in ten plastic cages, with 70 bees per cage (each individual was considered an independent replicate for future experiments). These bees were supplemented with 3 g of pollen replacer and 60% sucrose syrup ad libitum [26].
2.3. Stimulus preparation
For the behavioral assay, a pure standard of benzyl alcohol (≥99% GC, Sigma-Aldrich, Munich, Germany), and Mentha piperita essential oil were used as stimuli considering the previous results report by Silva et al. [22]. In summary, M. piperita leaves were collected from ornamental specimens in Chillán, Ñuble Region, Chile. Then, 300 g of this material were air-dried at room temperature for 48 h, after which they were boiled for 6 h by hydrodistillation using a Clevenger apparatus Finally, anhydrous sodium sulfate (Na2SO4) was added to remove water from the samples and they were stored at 8 °C in complete darkness. Each stimulus was diluted in ethanol (≥99% GC, Sigma-Aldrich, Munich, Germany) at a concentration of 100 µL/mL [27].
2.4. Olfactometric bioassays
The behavioral response of A. mellifera to volatiles of M. piperita essential oil and benzyl alcohol were conducted using a glass Y-tube olfactometer (21 cm long with an internal diameter of 3 cm), with glass odor chambers (15 cm long with an internal diameter of 3 cm) that were connected to the end of the Y-tube arms to deliver the stimulus using filtered air at 280 ml min-1 with a positive pressure air pump. A volume of 10 μl of each stimulus diluted in ethanol at a concentration of 100 µL/mL [27] was applied to a strip of filter paper (1 × 7.5 cm); 30 s were allowed for the solvents to evaporate, and then these were placed inside an odor chamber. Thirty worker honey bees that were 5-, 10-, 15-, and 20-days-old (120 worker honey bees in total) were used per trial and health condition (I-DWV or N-DWV) Each worker honey bee was allowed to move freely inside the olfactometer for 6 min. The first choice made by each insect in that time frame to either arm of the Y-tube was considered a positive choice. The worker honey bees were subjected to choice tests between benzyl alcohol vs. air and essential oil vs. benzyl alcohol. After each bee chose, they were stored at -80°C until further analysis.
2.5. RNA extraction and cDNA synthesis
Between 2 to 5 worker honey bees were selected for RNA extraction depending on the distribution of choices made by bees in the olfactory test. Then, the antennae and head of these previously selected bees were used for RNA extraction according to Kim et al. [21] and Mondet et al. [28], with some modifications Briefly, selected antennae and head were cooled to -80 °C and then grounded by adding 200 µL of TrizolTM (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Afterwards, 5 µL of Carrier RNA were added for antennae and head respectively, according to the manufacturer's recommendations (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Absolute ethanol (100%) and 200 µL of chloroform were added to the homogenate and incubated at 4 °C for 10 min. This was then centrifuged for 5 min at 10,000 × g at 4 °C and finally, the supernatant was collected for RNA extraction. RNA extraction, from antennae and head, was performed following the instructions provided by E.Z.N.A. Total RNA Kit I (Omega Bio-Tek, Norcross, GA, USA). RNA quality and yield was determined with a spectrometer (Infinite 200 PRO NanoQuant, Tecan Group, Männedorf, Switzerland). The extracted RNA was subsequently used for cDNA first strand synthesis using a M-MLV reverse transcriptase enzyme (Invitrogen, Life Technologies, Carlsbad, CA, USA), according to the manufacturer's instructions. The cDNA samples were stored at -20 °C for later use.
2.6. Real-Time PCR quantification for viral load and gene expression
The quantification of the viral load (DWV-A) and expression of the neuronal genes, such as AmNrX-1 and AmNlG-1 [24], as well as Amobp5 (OBP5) and Amobp11 (OBP11) [2] was carried out using specific primers (Table 1). The qPCR reaction was carried out using the 1x KAPA SYBR FAST Universal 2x qPCR Master Mix (Kapa Biosystems, Wilmington, MA, USA), following the supplier's instructions. Samples were brought to a reaction volume of 15 µL, including 20 ng of cDNA, 530 nM of each primer and sterile filtered molecular grade water until 15 µL were reached. The thermal reaction conditions were 95 °C for 5 min, followed by 40 cycles at 95 °C for 15 s, 60 °C for 15 s and 72 °C for 30 s. Real-time PCR assays were performed on a Stratagene Mx3000P thermal cycler (Agilent Technologies, Santa Clara, CA, USA) and data were analyzed using MxPro software (Stratagene, Agilent Technologies, Santa Clara, CA, USA). The relative expression of each gene was calculated after normalization with an endogenous gene (β-actin) (Table 1), as described by Pfaffl [29]. To determine the viral load in each segment of the worker bees (head and antennae), absolute quantification of DWV-A was performed. Briefly, a standard curve using a purified PCR product (Wizard®VR SV gel and PCR clean-up system, Promega, Madison, WI, USA), belonging to the viral target sequence [19], was used. Then, the purified amplicon was quantified by spectrophotometry (EpochTM Microplate Spectrophotometer, BioTek, Winooski, VT, USA) to calculate the copy number, according to Wu et al. [30]. Therefore, linear standard curves (95-100 % efficiency) were then generated using serial dilution (1.0 × 101 to 1.0 × 109) of the purified cDNA viral copy number. Subsequently, Ct values were plotted against copy number values (log10). Therefore, the sample copy number was estimated using the Ct values and comparisons with the linear equation of the standard curve and the normalization values of the β-actin gene cleanup [31]. Then, the data were expressed as the number of DWV-A copies per bee considering the dilutions that were performed in the cDNA synthesis and qPCR reaction.
2.7. Data analysis
Considering that the viral load quantified in the antennae and head was correlated in bees that were inoculated and non-inoculated with DWV-A, a general linear model (GLM) was used to design a nested multivariate ANOVA (MANOVA). Thus, viral load data quantified in the antennae and the head were considered response variables. Bee age (5-, 10-, 15- and 20-days-old) and viral status (bees inoculated (I-DWV) and non-inoculated (N-DWV) with DWV-A) were considered independent variables, with the viral condition nested in the bee age variable. The Bonferroni test (p < 0.05) was then run to separate means between treatments. A chi-square test (p < 0.05) was used to analyze the statistical difference in the responses (preference) of inoculated (I-DWV) and non-inoculated (N-DWV) honey bees at 5-, 10-, 15- and 20-days-old (preference) in the olfactometric Y-tube test with benzyl alcohol and M. piperita essential oil. Also, a binary (1 and 0) logistic regression was run to determine the likelihood of preference of the young bees to the pheromone compound in the Y-tube test in function of the DWV-A load and bee age. The preference for the pheromone compound was marked as positive (yes = 1) and no preference or preference for the other compound was considered a negative response (no = 0). The data on all bees tested in the experiments (including the viral load of the thorax and abdomen of inoculated and non-inoculated bees ) was included in the logistic regression. Statistical differences in the gene expressions of AmNrX-1, AmNlG-1, Amelobp5 (OBP5) and Amelobp11 (OBP11) were also estimated by nested MANOVA because Amelobp5 and Amelobp11, as well as AmNrX-1 and AmNlG-1 expression values were correlated. Thus, relative gene expressions were considered response variables in function of the independent variables, such as bee age and viral status (nested variable). Then, a Bonferroni test (p < 0.05) was run to separate the means between treatments. All analyses were carried out with STATISTICA 7.0 software (StatSoft, Tulsa, OK, USA).
3. Results
3.1. Viral load in antenna and head
The viral load detected in honey bees varied significantly between inoculated (I-DWV) and non-inoculated (N-DWV) bees, but also, these significant changes in viral load were influenced by bee age (Wilks λ = 0.09; F = 157.76; df = 6/414; p < 0.001) in both the antenna and head (Figure 1). Viral loads were found to be high and significant (1.0 x 109 viral copy number per bee) in I-DWV bees that were 10- to 20-days old, compared with non-inoculated (N-DWV) bees that also showed a basal viral level (1.0 x 104 to 1.0x 105 viral copy number per bee) that was not significant (Figure 1A). The viral loads in bee heads also varied among treatments in bees from 5- to 20-days old; we detected that I-DWV bees showed high viral loads (1.0 x 107 to 1.0 x 1010 viral copy number per bee) compared to N-DWV bees (1.0 x 104 to 1.0 x 104 viral copy number per bee) (Figure 1B).
3.2. Olfactory tests
In the Y-tube olfactometer, non-inoculated honey bees (N-DWV) were generally more attracted to the pheromone compound, benzyl alcohol, than worker honey bees that were intentionally inoculated with the DWV-A (I-DWV), except in the youngest bees 5 days after pupal emergence(Table 2). Thus, circa 50 to 60% of N-DWV bees responded significantly to the pheromone compound, measured in 10-, 15- and 20-day-old bees (Table 2). On the other hand, a low proportion (10 to 23%) of the inoculated bees (I-DWV) preferred the pheromone compound. In fact, most non-inoculated bees (N-DWV) showed no preference when tested in the Y-tube olfactometer (Table 2). Also, when the preference between benzyl alcohol or M. piperita essential oil was tested in the Y-tube test, significant differences were only observed in 10- and 15-day-old bees, in which case, a significantly higher proportion (almost 50%) of N-DWV bees were attracted to the benzyl In contrast, I-DWV bees of the same ages had a low preference (10 to 20%) for the pheromone compound (Table 3). No difference in behavioral preference was observed between N-DWV and I-DWV bees that were 5- and 20-days-old when the pheromone and the M. piperita essential oil were used as stimuli (Table 3): In fact, 20-day-old non-inoculated bees (N-DWV) significantly preferred the essential oil over the pheromone compound; a response that was not found in DWV-A inoculated bees (I-DWV) (Table 3). Furthermore, a significant proportion of I-DWV bees had no preference for the benzyl alcohol or the M. piperita essential oil (Table 3).
The regression logistic analysis showed that the probability of preference to the pheromone compound vs air by nurse honey bees tested in the Y-tube was significantly influenced by the DWV-load (Wald´s χ2 = 28.99, p < 0.001, Odds ratio = 0.79) and the bee age (Wald´s χ2 = 7.47, p = 0.006, Odds ratio = 1.08). The probability that nurse honey bees responded to the pheromone compound vs the essential oil was also significantly influenced by the DWV-load (Wald´s χ2 = 27.28, p < 0.001, Odds ratio = 0.72), but not by the bee age (Wald´s χ2 = 0.72, p = 0.395, Odds ratio = 0.98) (Figure 2B):
3.3. Gene expression
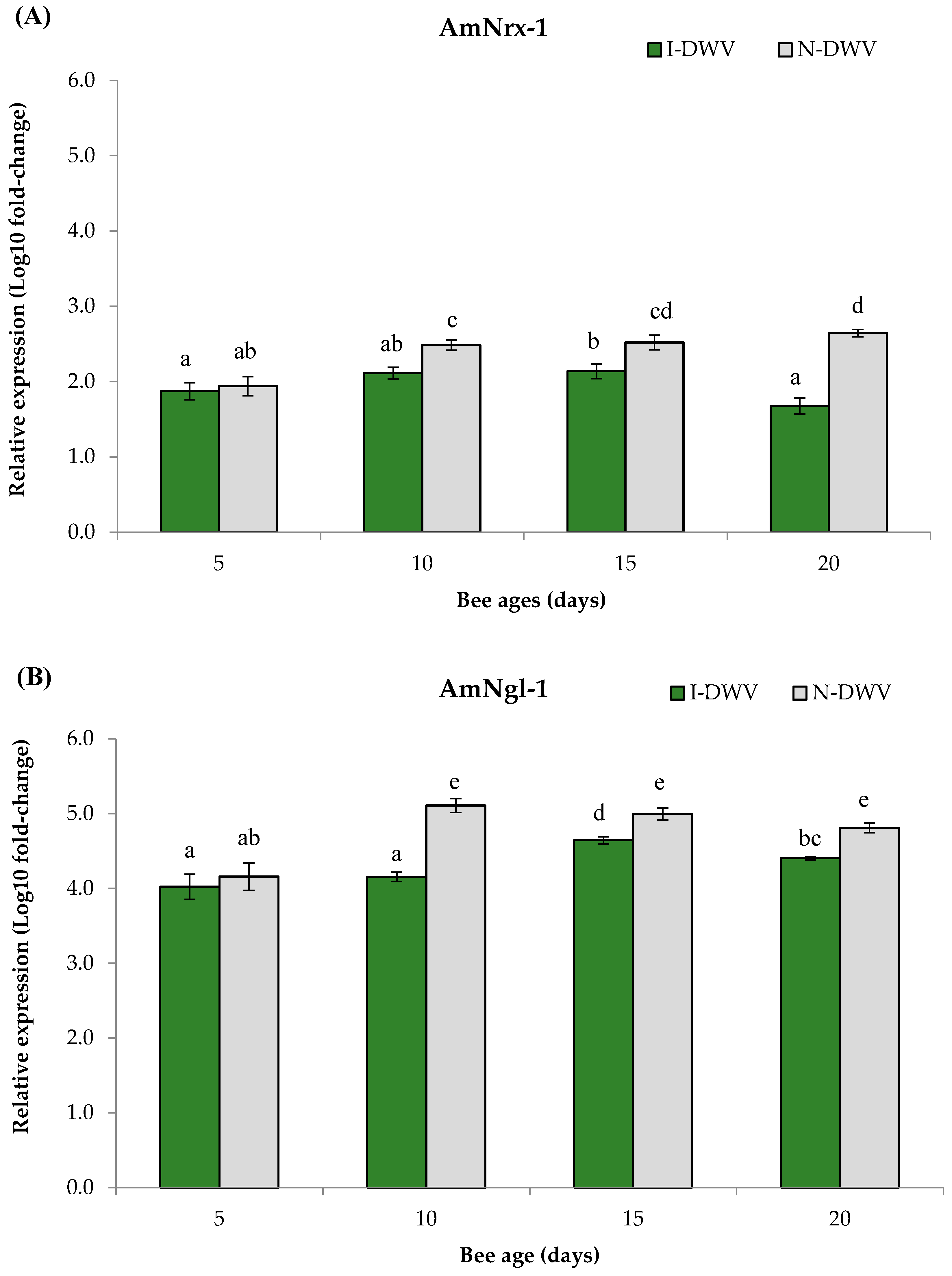
Regarding the gene expressions related to OBP proteins and the synaptic neuronal genes in worker honey bees, we detected significant differences in all genes analyzed in bees that were inoculated and non-inoculated with DWV-A. Nevertheless, the viral condition nested in the bee age affected the observed changes in the gene expressions AmelObp5 and AmelObp11 (MANOVA Wilks λ= 0.48; F = 22.57; df = 8, 414; p < 0.001), and also AmNrx-1 and AmNlg-1 (MANOVA Wilks λ = 0.60; F = 14.81; df = 8, 414; p < 0.001). We observed in antennae that the genes AmelObp5 and AmelObp11 were significantly down-regulated in inoculated bees (I-DWV) compared to non-inoculated bees (N-DWV) (Figure 3A and 3B). The significantly lower expression was observed in I-DWV bees from 10- to 20-days-old,ranging between 15 and 22% in Amelobp5 and 15% to 65% in Amelobp11 (Figure 3A and 3B). Similarly, when the synaptic genes AmNrx-1 and AmNlg-1 were analyzed they were also down-regulated in I-DWV bees (Figure 4A and 4B). I-DWV bees also showed lower expression levels for both aforementioned genes in the heads of 10- to 20-day-old worker bees. These low expressions ranged from 19 to 53% and 9 to 21% for AmNrx-1 and AmNgl-1, respectively (Figure 4A and 4B).
3.2. Figures, Tables and Schemes

Table 1.
Primers used in this study.
| Primers Names | Sequence | Reference |
|---|---|---|
| AmNrx-1 | F- CTGCTTCGAGCGACGACTAT | Morfin et al., 2019 |
| R- ACGACCGGATGGATGATTGG | ||
| AmNlg-1 | F- ATGTCGAGGATGCTGCGACTGGA | Morfin et al., 2008 |
| R-TACCTGTGCACTATCTCCTGTTGTA | ||
| DWV-A | F-TATCTTCATTAAAGCCACCTGGAA | Yang et al., 2005 |
| R- TTTCCTCATTAACTGTGTCGTTGAT | ||
| AmelOBP5 | F- ATGCGGAAATCGTGCTTGCA | Li et al., 2015 |
| R- TGCCATTACTCACGGGAAGA | ||
| AmelOBP1 | F- TGAGGATGTCGAAGCTACGGAA | Li et al., 2015 |
| R- CACGGAGCAATAAACGCTATGG | ||
| β -actin | F- ATGCCAACACTGTCCTTTCTGG | Yang et al., 2005 |
| R- GACCCACCAATCCATACGGA |
Table 2.
Proportion of worker bees that were inoculated (I-DWV) with deformed wing virus variant A vs. non-inoculated (N-DWV) bees that responded to the pheromone compound benzyl alcohol versus air in the Y-tube test. A chi-square test (p < 0.05) was run to detect significant differences in bees that were inoculated and non-inoculated with DWV according to their ages.
Table 2.
Proportion of worker bees that were inoculated (I-DWV) with deformed wing virus variant A vs. non-inoculated (N-DWV) bees that responded to the pheromone compound benzyl alcohol versus air in the Y-tube test. A chi-square test (p < 0.05) was run to detect significant differences in bees that were inoculated and non-inoculated with DWV according to their ages.
| Bee age (days) | Pheromone (benzyl alcohol) vs. Air | |||||||||
| Status | Pheromone | χ2 | p-value | Air | χ2 | p-value | No preference | χ2 | p-value | |
| 5 | I-DWV | 0.33 | 1.36 | 0.243 | 0.30 | 3.75 | 0.053 | 0.37 | 6.70 | 0.010 |
| N-DWV | 0.20 | 0.10 | 0.70 | |||||||
| 10 | I-DWV | 0.10 | 13.02 | <0.001 | 0.30 | 3.75 | 0.053 | 0.60 | 3.27 | 0.071 |
| N-DWV | 0.53 | 0.10 | 0.37 | |||||||
| 15 | I-DWV | 0.13 | 10.80 | 0.001 | 0.13 | 0.00 | 0.992 | 0.73 | 9.64 | 0.002 |
| N-DWV | 0.53 | 0.13 | 0.33 | |||||||
| 20 | I-DWV | 0.23 | 8.30 | 0.004 | 0.23 | 1.92 | 0.166 | 0.53 | 3.36 | 0.067 |
| N-DWV | 0.60 | 0.10 | 0.30 | |||||||
Table 3.
Proportion of worker bees that responded to the pheromone compound benzyl alcohol or Mentha piperita essential oil in the Y-tube test (A) when worker bees were inoculated (I-DWV) or non-inoculated (N-DWV) with deformed wing virus variant A. A chi-square test (p < 0.05) was run to detect significant differences in bees that were inoculated and non-inoculated with DWV according to their age.
Table 3.
Proportion of worker bees that responded to the pheromone compound benzyl alcohol or Mentha piperita essential oil in the Y-tube test (A) when worker bees were inoculated (I-DWV) or non-inoculated (N-DWV) with deformed wing virus variant A. A chi-square test (p < 0.05) was run to detect significant differences in bees that were inoculated and non-inoculated with DWV according to their age.
| Bee age (days) | Pheromone (benzyl alcohol) vs. essential oil | |||||||||
| Status | Pheromone | χ2 | p-value | Essential oil | χ2 | p-value | No preference | χ2 | p-value | |
| 5 | I-DWV | 0.23 | 0.09 | 0.766 | 0.07 | 1.46 | 0.228 | 0.70 | 1.15 | 0.284 |
| N-DWV | 0.27 | 0.17 | 0.57 | |||||||
| 10 | I-DWV | 0.20 | 7.18 | 0.007 | 0.07 | 1.46 | 0.228 | 0.73 | 11.28 | 0.001 |
| N-DWV | 0.53 | 0.17 | 0.30 | |||||||
| 15 | I-DWV | 0.10 | 11.43 | 0.001 | 0.27 | 0.32 | 0.573 | 0.63 | 13.61 | <0.001 |
| N-DWV | 0.50 | 0.33 | 0.17 | |||||||
| 20 | I-DWV | 0.07 | 1.46 | 0.228 | 0.27 | 11.28 | 0.001 | 0.67 | 17.78 | <0.001 |
| N-DWV | 0.17 | 0.70 | 0.13 | |||||||
Figure 1.
DWV-A load measured in antennae (A) and head (B) of Apis mellifera of different ages that were inoculated (I-DWV) and non-inoculated (N-DWV). Means (±SE) with different letters indicate significant difference according to the Bonferroni test (p < 0.05).
Figure 1.
DWV-A load measured in antennae (A) and head (B) of Apis mellifera of different ages that were inoculated (I-DWV) and non-inoculated (N-DWV). Means (±SE) with different letters indicate significant difference according to the Bonferroni test (p < 0.05).
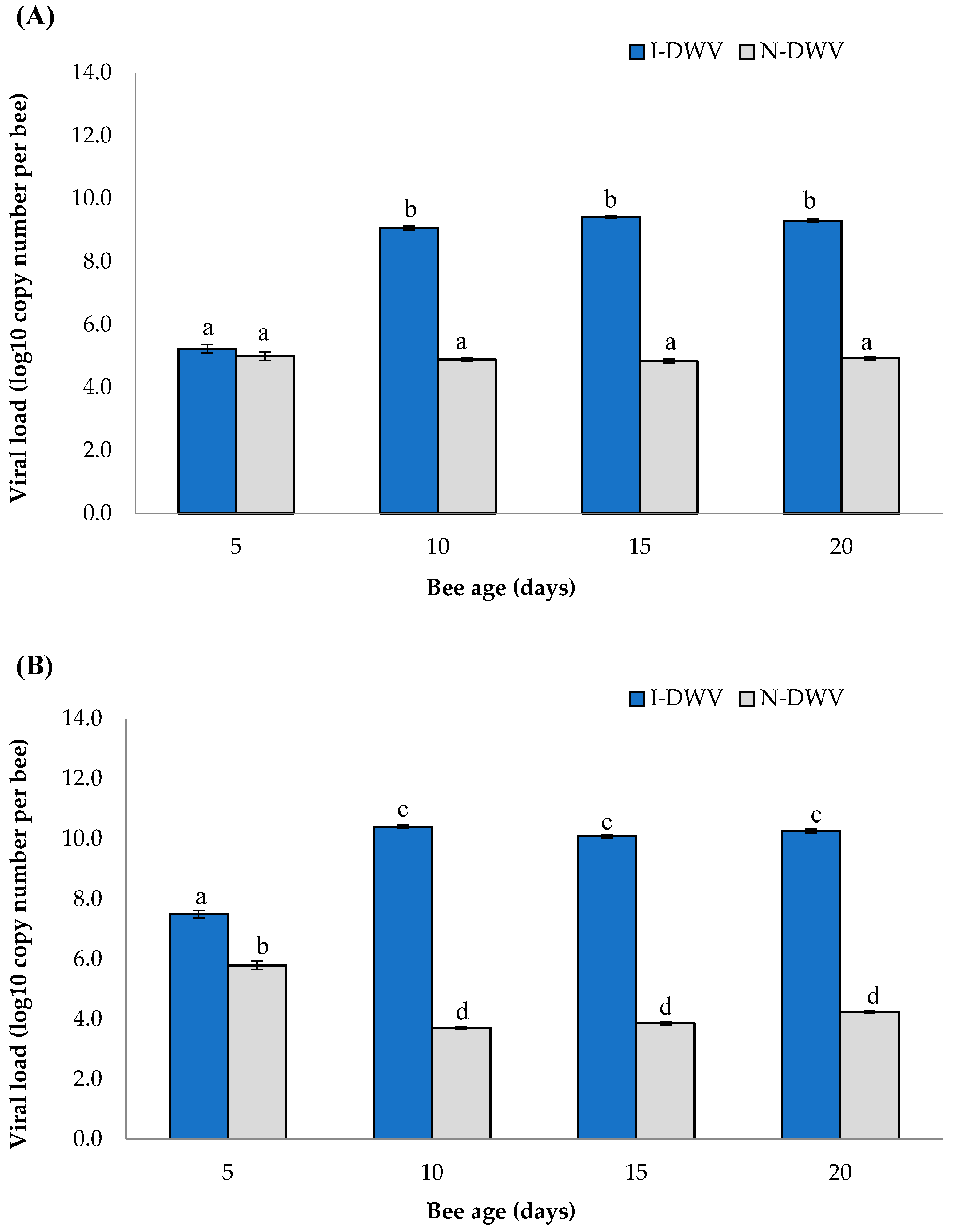
Figure 2.
Three-dimensional logistic regression curve representing the probability of the preference of honey bees to a pheromone compound (benzyl alcohol) vs air (A) and a pheromone vs an essential oil (B) in function of the DWV-A load and bee age in the Y-tube test. When the bees were exposed to the pheromone compound vs air, the bees that responded by preferring the pheromone were marked as positive (yes = 1, n = 76), while those that showed no preference were considered negative responses (no = 0, n = 140). When bees were exposed to the pheromone vs the essential oil, the preference for the pheromone was considered positive (yes = 1, n = 63), while bees that showed no preference or preferred another compound were recorded as negative (no = 0, n = 153).
Figure 2.
Three-dimensional logistic regression curve representing the probability of the preference of honey bees to a pheromone compound (benzyl alcohol) vs air (A) and a pheromone vs an essential oil (B) in function of the DWV-A load and bee age in the Y-tube test. When the bees were exposed to the pheromone compound vs air, the bees that responded by preferring the pheromone were marked as positive (yes = 1, n = 76), while those that showed no preference were considered negative responses (no = 0, n = 140). When bees were exposed to the pheromone vs the essential oil, the preference for the pheromone was considered positive (yes = 1, n = 63), while bees that showed no preference or preferred another compound were recorded as negative (no = 0, n = 153).
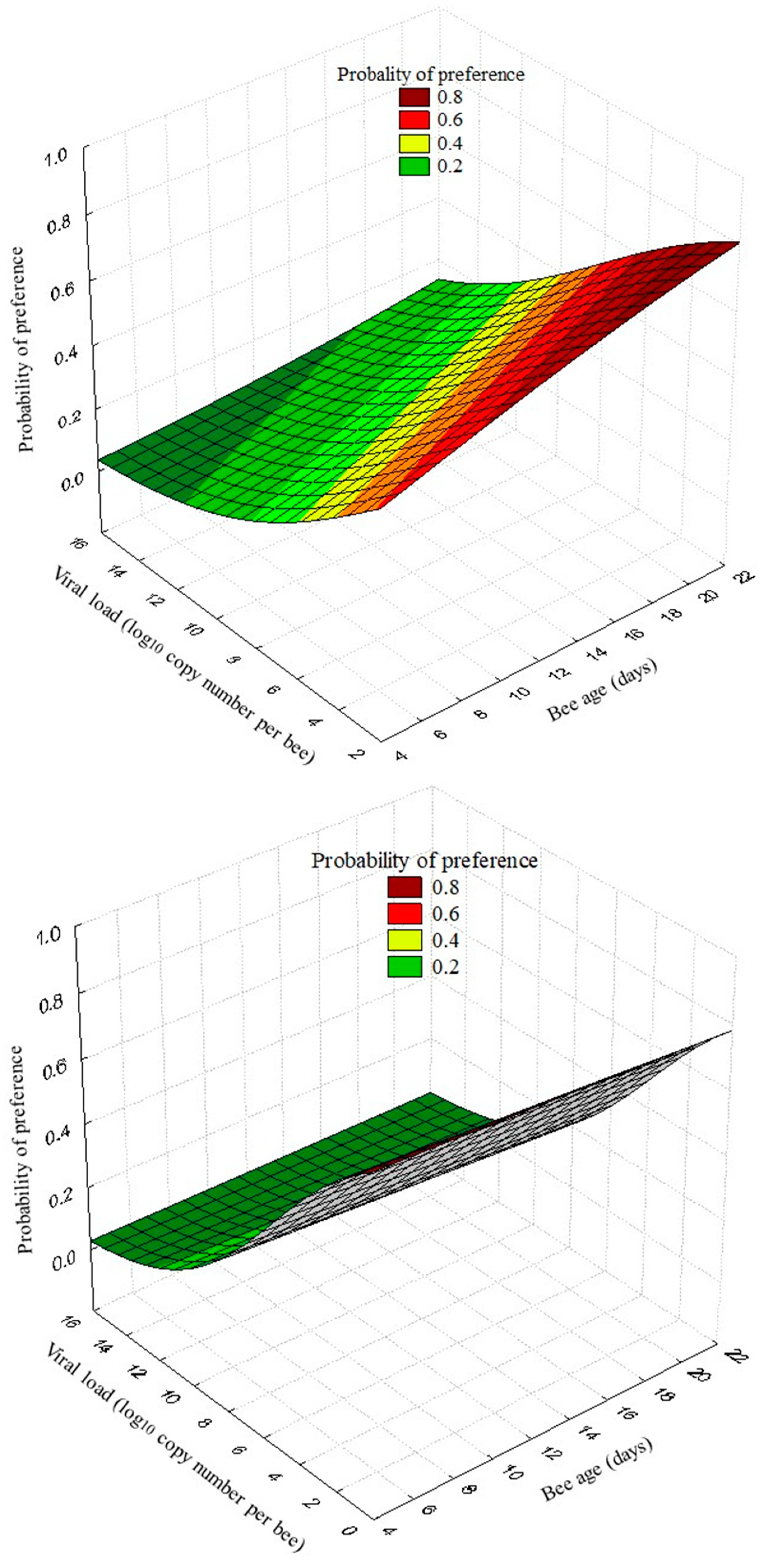
Figure 3.
Gene expression of genes coding for OBPs in adult Apis mellifera antennae of different ages and that were inoculated (I-DWV) and non-inoculated (N-DWV) with DWV-A. Means (±SE) with different letters indicate significant difference according to the Bonferroni test (p < 0.05) in (A) AmelObp5,(OBP5) and (B) AmelObp11 (OBP11) genes relative expression.
Figure 3.
Gene expression of genes coding for OBPs in adult Apis mellifera antennae of different ages and that were inoculated (I-DWV) and non-inoculated (N-DWV) with DWV-A. Means (±SE) with different letters indicate significant difference according to the Bonferroni test (p < 0.05) in (A) AmelObp5,(OBP5) and (B) AmelObp11 (OBP11) genes relative expression.
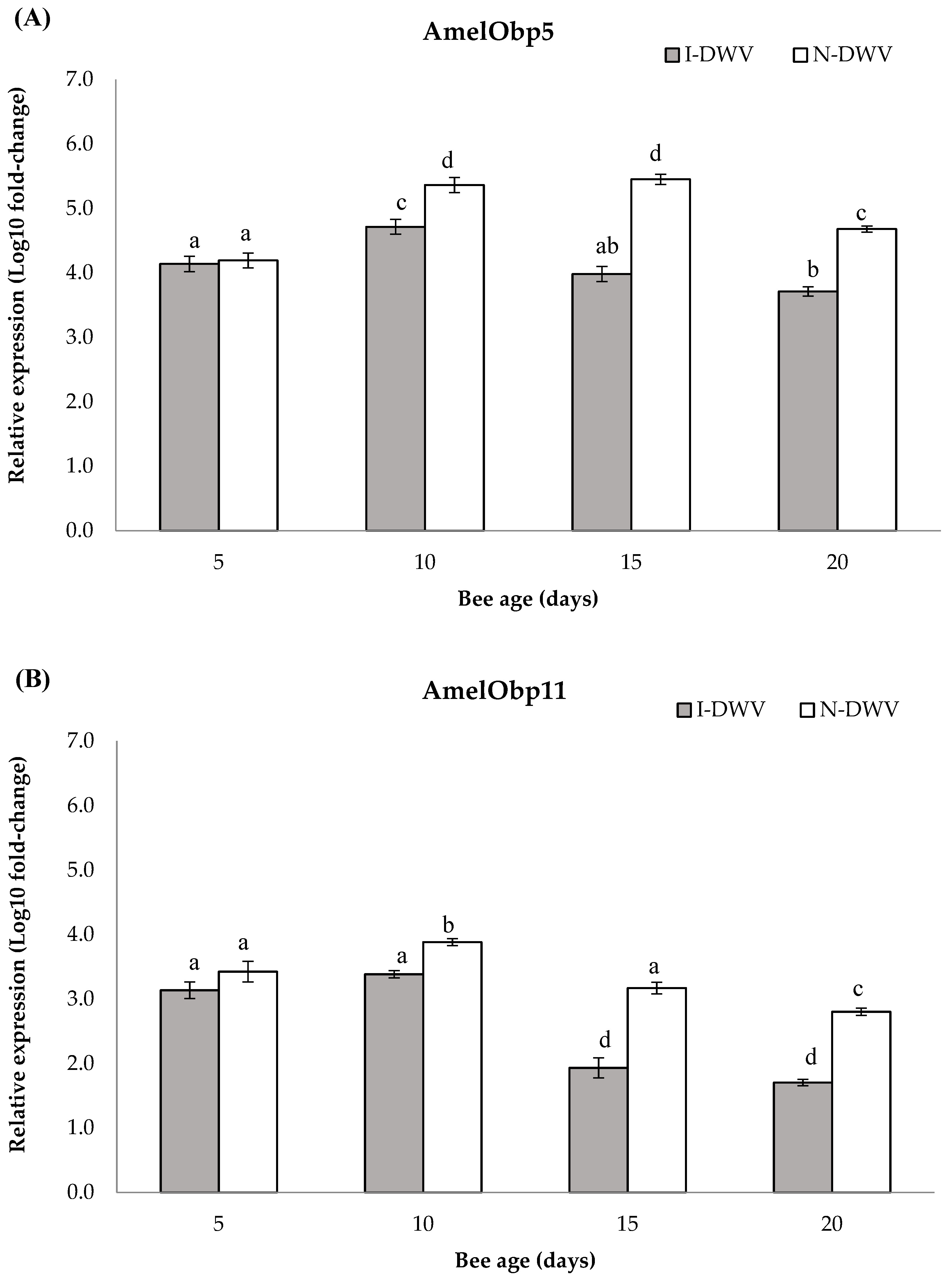
Figure 4.
Gene expression of genes coding for neural proteins in adult Apis mellifera heads that were inoculated (I-DWV) and non-inoculated (N-DWV) with DWV-A and had different ages. Means (±SE) with different letters indicate significant differences according to the Bonferroni test (p < 0.05) in (A) AmNrx-1 (Neurexin) and (B) AmNgl-1 (Neuroligin) genes relative expressions.
Figure 4.
Gene expression of genes coding for neural proteins in adult Apis mellifera heads that were inoculated (I-DWV) and non-inoculated (N-DWV) with DWV-A and had different ages. Means (±SE) with different letters indicate significant differences according to the Bonferroni test (p < 0.05) in (A) AmNrx-1 (Neurexin) and (B) AmNgl-1 (Neuroligin) genes relative expressions.
4. Discussion
It is widely known that olfaction coordinates a large part of insect behaviors, allowing them to select and recognize environmental scents and pheromones [1,2]. Therefore, the ability to respond behaviorally to an odor source greatly depends on the proper functioning of the olfactory system and the different molecules that interact with this system [32].
Odorant binding proteins (OBPs) have been described as essential molecules for scent recognition [33]." OR "Odorant binding proteins (OBPs) have been described as some of the most important molecules for scent recognition [33]. These proteins, after participating in the recognition of environmental compounds, bind to a transmembrane receptor, generating a stimulus in the form of a synapse that is interpreted in the brain; thus, it is understood that these proteins act as the first bridge between the environment and insects [33]. Their role in behavioral responses has been evidenced in Drosophila melanogaster Meigen mutants where OBP genes have proven to be responsible for ethanol perception being silenced, resulting in insects that are deficient in this protein, therefore showing abnormal behaviors with respect to aroma compared to wild insects [34]. A similar case has been described in other insects, such as Spodoptera litura F., where the gene coding for the pheromone binding proteins (PBP) SlitPBP1 and SlitPBP2 was silenced; thus, the insect showed lower behavioral responses. The mutant insects in this case were less attracted to the pheromone used as a stimulus, which was associated with a decrease in the perception and olfactory sensitivity to specific compounds perceived by the PBPs [35]. We have previously demonstrated that the presence of DWV-A at high loads in the antennae generated a reduction in olfactory perception evaluated in EAG assays, and a reduction in the expression of the genes Amobp5 and Amobp11 coding for OBP5 and OBP11 proteins [22]. Additionally, it has been studied that these genes are significantly expressed in antennae of 10- and 15-day-old bees [7,8]. These results have allowed linking these proteins to nurse bee behavior, in which the insects are dedicated to the task of caring for larvae, considering that the OBP5 protein has a high affinity for compounds of the alarm pheromone of diseased larvae, including phenethyl acetate, phenylethanol and benzyl alcohol, with benzyl alcohol being the most competitive for the recognition site of the protein [7]. Therefore, the decreases in olfactory preferences observed in our study may be partially explained by the gene knockdown of OBP5 and OBP11 proteins in the antennae, which are responsible for the recognition of benzyl alcohol. Thus, young honey bees inoculated with the DWV-A showed a reduction in their ability to detect compounds associated with these proteins, as a consequence of the decrease in the gene expressions coding for olfactory proteins. It is important to understand that insect behaviors are complex responses to environmental recognition; thus, honey bees depend on neural processes to perform these complex behaviors, including foraging, hive coordination and hygienic behavior [36]. Therefore, alteration of the normal functioning of neural processes compromises insect behaviors in response to perceived stimuli in the environment [37]. Young honey bees artificially inoculated with DWV have been shown to exhibit learning and memory impairment [37]. In addition, Morfin et al. [24] demonstrated that co-infection of the ectoparasite V. destructor and deformed wing virus (DWV) decreases memory and learning ability in PER assays 48 h post viral inoculation; they also observed that bees with high viral loads, co-infected with V. destructor decreased the gene expressions associated with pre- and postsynaptic proteins AmNrx-1 (neurexin) and AmNlg-1 (neuroligin). Thus, they associated the decrease in these genes with the loss and reduction in the learning capacity of bees with high viral loads that are also parasitized with V. destructor. Nevertheless, our study demonstrates that the increase in the viral load in the insect brain in the absence of the V. destructor mite causes decreases in these pre- and post-synaptic gene expressions. This is one of the few studies to date that evidenced this consequence with only the presence of high viral loads.
Shah et al. [23] reported that DWV alters the cell structure of critical regions of the brain, including the neuropils responsible for vision and olfaction, and is also capable of dividing different sections of the sensory perception lobes, which could compromise physiological functions provided that this pathogen alters the expression of different genes. Therefore, there is evidence that DWV affects the capabilities and physiological functioning of the sensory organs of bees [21,22,38], which could explain why this alteration in behavioral responses was observed in our study. Furthermore, the logistic regression analysis showed a direct relationship between the viral load detected in the body (Table S1 and S2, Supplementary Material) and the probability of preference to specific aromas by young honey bees (Figure 2). This study indicated that a high level of DWV-A in bees reduced the probability that these bees choose the pheromone compound. Nevertheless, the reduction in the behavioral responses of these bees may also be the consequence of multiple altered factors resulting from the presence of the DWV, suggesting that an alteration in the sensory system, specifically the decrease of sensory perception in the antennae and alterations in the processes of interpretation of synaptic signals, specifically in the genetic alteration of neuronal genes, could lead these insects to present altered and reduced behaviors. Though their behaviors were not totally deficient because we observed behavioral responses in both treatments, in bees that were infected and non-infected with DWV-A.
The decrease in the behavioral response of DWV-A infected bees could be related to the observed decrease in OBPs and synaptic genes. The decline in the response of healthy 20-day-old bees exposed to the pheromone and essential oil component could be explained, in part, by the age-related distribution of tasks within the hive, known as temporal polyethism [39,40]. The existence of four castes within the hive was proposed in 1986, including cell cleaning (1 to 3 days old), care and maintenance of the larvae by nurse bees (4 to 12 days old), food processing and nest maintenance, which were known as middle-aged bees (13 to 20 days old) and foraging (more than 21 days old) [41]. Johnson [42] subsequently confirmed the existence of the four castes within the hive, indicating that as the bees matured their tasks became more limited, thus the 4-12-day-old bees focused solely on brood care tasks, while the 12-20-day-old bees specialized in nectar processing and nest maintenance in preparation for foraging outside the hive. Therefore, it is logical to expect that bees between 15 and 20 days old (in this study) would be more attracted to plant-derived compounds associated with foraging, such as the essential oil of M. piperita, than to the pheromone component that was more related to the larval scents. This could explain, in part, why the behavioral response and preference of inoculated and non-inoculated bees was the same at the end of the trial (20-day-old bees), since non-inoculated bees had a greater preference for the aroma of the plant-derived essential oil over the pheromone compound (Table 3). Therefore, the low preference for the pheromone compound would be equal to the preference of the infected bees, and this behavior would be expected given the physiological development of the species itself and the castes within the hive, as described by Johnson [42]. Nevertheless, in bees inoculated with DWV-A we did not observe a significant preference for the essential oil in 20-day-old bees. Therefore, we thought that the infection caused by this pathogen would have serious consequences on the physiological development of the insects, affecting the social specialization associated with temporal polyethism, which provides benefits at the group level due to the division of labor that permits increased productivity and reliable task performance. We demonstrated here that DWV-A significantly affects young bees (10-15 days-old) in the recognition or preference of a specific compound (benzyl alcohol) related to the tasks of nurse honey bees. Therefore, DWV-A infected nurse bees demonstrated a low perception of the compounds released by brood that are necessary to maintain their care, which indicates that this brood is poorly cared for, possibly leading to a delay in the development of the colony or its total decline. It is also still unknown whether DWV affects the preference or attraction for other compounds or pheromones within the colony or aromas present in the flowers that attract foraging honey bees. Therefore, at least two questions remain. Do DWV-A and/or other DWV variants significantly affect foraging honey bees and their ability to recognize aromas in the field? Can DWV cause disorientation in the field leading to a loss in foraging bees because they are unable to locate their colony of origin? Further studies are required to answers these questions.
5. Conclusions
In summary, the high viral load recorded in the antennae and heads decreased the necessary behaviors of nurse bees. DWV-A also caused a down-regulation of OBPs in the antennae and the synaptic genes neuroxins-1 and neurogilin-1 in bee heads. Therefore, a DWV-A infection could compromise the functioning of smell and processing of peripheral information, altering the behaviors of nurse bees associated with the aromas of the hive, and negatively affecting the survival of bees at the individual and colony levels.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
M.V. conceived of this research; M.V., and D.S. designed experiments; D.S. and A.J. performed the experiments and the analysis in labs; D.S., N.A., R.C., A.J., Z.N. and M.V. wrote the manuscript and participated in the revisions. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the FONDECYT N◦ 1211688 grant from the National Research and Development Agency (ANID), Chile.
Data Availability Statement
Not Aplicable
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare that they have no conflict of interests. The funders played no role in the design of the study, the collection, analyses, or interpretation of the data, in the writing of the manuscript, or the decision to publish the results.
References
- Groot, A. T.; Dekker, T.; Heckel, D. The Genetic Basis of Pheromone Evolution in Moths. Annu. Rev. Entomol 2016, 61(1): 99–117. [CrossRef]
- Li, Q and Liberles, S.D. Aversion and attraction through olfaction. Curr. Biol. 2015, 25(3):120-129. [CrossRef]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [CrossRef]
- Zhou, J.-.-J., Vieira, F.G., He, X.-.-L., Smadja, C., Liu, R., Rozas, J. and Field, L.M. Genome annotation and comparative analyses of the odorant-binding proteins and chemosensory proteins in the pea aphid Acyrthosiphon pisum. Insect Molecular Biology 2010, 19: 113-122. [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93: 184-200. [CrossRef]
- Forêt, S.; Maleszka, R. Function and evolution of a gene family encoding odorant binding-like proteins in a social insect, the honey bee (Apis mellifera). Genome Res. 2006, 16(11): 1404–1413. [CrossRef]
- Zhao, H.; Luo, Y.; Lee, J.; Zhang, X.; Liang, Q.; Zeng, X. The Odorant-binding protein gene obp11 shows different spatiotemporal roles in the olfactory system of Apis mellifera ligustica and Apis cerana cerana. Sociobiology. 2013, 60, 429–435. [CrossRef]
- Zhao, H.X.; Zeng, X.N.; Liang, Q.; Zhang, X.F.; Huang, W.Z.; Chen, H.S.; Luo, Y.X. Study of the obp5 gene in Apis mellifera ligustica and Apis cerana cerana. Genet. Mol. Res. 2015, 14 (2), 6482–6494. [CrossRef]
- Plettner, E., Eliash, N., Singh, N.K. The chemical ecology of host-parasite interaction as a target of Varroa destructor control agents. Apidologie 2017, 48, 78–92. [CrossRef]
- Potts, S.G.; Biesmeier, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: trends, impacts and drivers. Trend. Ecol. Evol. 2010, 25, 345-353. [CrossRef]
- Hall, D.M.; Steiner, R. Insect pollinator conservation policy innovations at subnational levels: Lessons for lawmakers. Environ. Sci. Policy 2019, 93, 118-128. [CrossRef]
- vanEngelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony Collapse Disorder: A Descriptive Study. PLOS ONE 2009, 4(8): e6481. [CrossRef]
- Martin, S.J.; Hardy, J.; Villalobos, E.; Martin-Hernández, R.; Nikaido, S.; Higes, M. Do the honeybee pathogens Nosema ceranae and deformed wing virus act synergistically?. Environ Microbiol Rep. 2013, 5-4.
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226.
- Brettell, LE; Mardoqueo, GJ; Schroeder, DC; Jones, IM; da Silva, JR; Vicente-Rubiano, M.; Martin, SJ. A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees. Insects, 2017 , 8 , 28.
- Benaets, K.; Van Geystelen, A.; Cardoen, D.; De Smet, L.; de Graaf, DC; Schoofs, L.; Larmuseau, MH; Brettell, LE; Martín, SJ; Wenseleers, T. Covert deformed wing virus infections have long-term deleterious effects on honeybee foraging and survival. Proc. Biol. Science, 2017 , 284. [CrossRef]
- Kevill, J. L.; de Souza, F.S.; Sharples, C.; Oliver, R.; Schroeder, D.C.; Martin, S.J. DWV-A Lethal to Honey Bees (Apis mellifera): A Colony Level Survey of DWV Variants (A, B, and C) in England, Wales, and 32 States across the US. Viruses 2019, 11(5): 426-426. [CrossRef]
- McMahon, D.P.; Natsopoulou, M.E.; Doublet,V.; Furst, M.; Weging, S.; Brown, M.J.; Gogol-Doring, A.; Paxton, R.J.; Mallinger, R.E.; Gratton, C. Species richness of wild bees, but not the use of managed honeybees, increases fruit set of a pollinator-dependent crop. Journal of Applied Ecology 2016, 52 (2): 323-330.
- Riveros, G.; Arismendi, N.; Zapata, N.; Evans, D.; Pérez, I.; Aldea, P.; Vargas, M. Occurrence, prevalence and viral load of deformed wing virus variants in Apis mellifera colonies in Chile. J. Apic. Res. 2019, 59, 63–68. [CrossRef]
- Vargas, M.; Arismendi, N.; Riveros, G.; Zapata, N.; Bruna, A.; Vidal, M.; Rodríguez, M.; Gerding, M. Viral and intestinal diseases detected in Apis mellifera in Central and Southern Chile. Chil. J. Agric. Res. 2017, 77, 243–249. [CrossRef]
- Kim, S. H.; Mercer, A.; Mitchell, A.; de Miranda, J.R.; Ward, V.; Mondet, F.; Bostina, M. Viral infections alter antennal epithelium ultrastructure in honey bees. J. Invertebr. Pathol. 2019, 168, 107252. [CrossRef]
- Silva, D.; Ceballos, R.; Arismendi, N.; Dalmon, A.; Vargas, M. Variant A of the Deformed Wings Virus Alters the Olfactory Sensitivity and the Expression of Odorant Binding Proteins on Antennas of Apis mellifera. Insects 2021, 12, 895. [CrossRef]
- Shah, K.S.; Evans, E.C.; Pizzorno, M.C. Localization of deformed wing virus (DWV) in the brains of the honeybee, Apis mellifera Linnaeus. Virol. 2009, 6, 182. [CrossRef]
- Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. The Combined Effects of Varroa destructor Parasitism and Exposure to Neonicotinoids Affects Honey Bee (Apis mellifera L.) Memory and Gene Expression. Biology 2019, 9(9):237. [CrossRef]
- Gusachenko, O.N.; Woodford, L.; Balbirnie-Cumming, K.; Ryabov, E.V.; Evans, D.J. Evidence for and against deformed wing virus spillover from honey bees to bumble bees: A reverse genetic analysis. Sci. Rep. 2020, 10, 16847. [CrossRef]
- Arismendi, N.; Caro, S.; Castro, M.P.; Vargas, M.; Riveros, G.; Venegas, T. Impact of mixed infections of gut parasites Lotmaria passim and Nosema ceranae on the lifespan and immune-related biomarkers in Apis mellifera. Insects 2020, 11, 420. [CrossRef]
- Liu, J.; Chen, M.; Ma, W.; Zheng, L.; Zhang, B.; Zhao, H.; Jiang, Y. Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera. Agronomy 2023, 13, 339. [CrossRef]
- Mondet. F.; Alaux, C.; Severac, D.; Rohmer, M.; Mercer, A.; Le Conte, Y. Antennae hold a key to Varroa-sensitive hygiene behaviour in honey bees. Sci. Rep. 2015 ,5: 10454. [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2003–2007. [CrossRef]
- Wu, Y.; Dong, X.; Kadowaki, T. Characterization of the copy number and variants of deformed wing virus (DWV) in the pairs of honey bee pupa and infesting Varroa destructor or Tropilaelaps mercedesae. Front. Microbiol. 2017, 8, 1558. [CrossRef]
- Yang, X.; Cox-Foster, D. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [CrossRef]
- Renou, M.; Anton, S. Insect olfactory communication in a complex and changing world. Current Opinion in Insect Science 2020, 42. 1-7. [CrossRef]
- Nakamura, A.; Chahad-Ehlers, S.; Lima, A. et al. Reference genes for accessing differential expression among developmental stages and analysis of differential expression of OBP genes in Anastrepha obliqua. Sci Rep 2016, 6, 17480. [CrossRef]
- Zhou, J.-J.; Zhang, G.-A.; Huang, W.; Birkett, M.A.; Field, L.M.; Pickett, J.A.; Pelosi, P. Revisiting the odorant-binding protein LUSH of Drosophila melanogaster: evidence for odour recognition and discrimination, FEBS Lett. 2004, 558 (1) 23–26. [CrossRef]
- Zhu, G.; Zheng, M.-Y.; Sun, J.-B.; Khuhro, S.A.; Yan, Q.; Huang, Y.; Syed, Z.; Dong, S.-L. CRISPR/Cas9 mediated gene knockout reveals a more important role of PBP1 than PBP2 in the perception of female sex pheromone components in Spodoptera litura. Insect Biochem. Mol. Biol. 2019, 115, 103244. [CrossRef]
- Dukas, R. Mortality rates of honey bees in the wild. Insect. Soc. 2008, 55, 252–255. [CrossRef]
- Iqbal J. and Mueller U. Virus infection causes specific learning deficits in honeybee foragers. Proc. R. Soc. B. 2007, 274:1517-1521.
- Chen, P.; Lu, Y.-H.; Lin, Y.-H.; Wu, C.-P.; Tang, C.-K.; Wei, S.-C.; Wu, Y.-L. Deformed Wing Virus Infection Affects the Neurological Function of Apis mellifera by Altering Extracellular Adenosine Signaling. Insect Biochem. Mol. Biol. 2021, 139, 103674. [CrossRef]
- Robinson, G.E. Genomics and integrative analyses of division of labor in honeybee colonies. Am. Nat. 2002, 160:S160–S172.
- Seid, M.A.; Traniello, J.F.A. Age-related repertoire expansion and division of labor in Pheidole dentata (Hymenoptera: Formicidae): a new perspective on temporal polyethism and behavioral plasticity in ants. Behav. Ecol. Sociobiol. 2006, 60, 631–644. [CrossRef]
- Seeley, T.D. Division of labor among worker honey bees. Ethology 1986, 71:249–251.
- Johnson, B.R. Within-nest temporal polyethism in the honey bee. Behav. Ecol. Sociobiol. 2008, 62, 777–784. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



