
Submitted:
16 November 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
Radiation therapy (RT) is one of the main approaches of cancer therapy, including non-small cell lung cancer (NSCLC). Radiation-induced DNA double-stranded breaks (DSBs), single-stranded DNA breaks (SSBs) and oxidized nucleobases cause replication fork stalling, chromosomal instability or point mutagenesis and can lead to cell death. Translesion DNA synthesis (TLS) is one of the DNA damage tolerance (DDT) pathways promoting replication of damaged DNA. Here, we explored the effects of the knockout of primase-polymerase PRIMPOL (PRIMPOL −/−) and TLS polymerase POLI (POLI −/−) genes in two different monoclonal lines derived from human NSCLC A549 cells. Our study aimed to investigate the impact of their absence on the susceptibility of these cells to ionizing radiation (IR)-induced stress. Without IR, cells that do not have PRIMPOL or POLI experience a considerable rise in apoptosis by day 7 of continuous cultivation and a noteworthy decline in cell migration under spatial confinement when compared to the parental wild-type cells. Despite this, we have found that PRIMPOL −/− exhibited higher clonogenic survival compared to wild-type cell lines in response to IR stress. Both of these cell lines demonstrated remarkable resilience to radiation stress, surpassing the survival rate of the original A549 cells. POLI deficiency reduces cellular γH2AX foci accumulation, while the presence of highest number of γH2AX foci in PRIMPOL−/− cells was ATM-independent. We report for the first time that increased post-irradiation clonogenic survival of PRIMPOL −/− A549 cells might be associated with the increase of CD133+ but not CD44+ populations of cancer stem-like cells.
Keywords:
non-small cell lung cancer
; ionizing radiation
; DNA repair
; PrimPol
; DNA damage tolerance
; translesion synthesis
; Pol iota (Polι)
; clonogenic survival
; cancer stem-like cells
1. Introduction
Radiation therapy allows cancer cells elimination through the formation of DNA damage. DNA double-stranded breaks (DSBs) can be repaired by homologous recombination (HR) [1] or non-homologous end joining (NHEJ) [2]. Single-stranded DNA (ssDNA) lesions are subject to repair by nucleotide excision repair [3] and base excision repair [4], while DNA mismatch repair functions to correct replication errors in newly synthesized DNA [5]. During the S phase of the cell cycle unrepaired SSBs stalls the progression of DNA replication and causes fork reversal [6]. Translesion DNA synthesis (TLS) emerged as a mechanism of DNA damage tolerance (DDT) that enables the completion of DNA replication leaving the unrepaired damage for future elimination [7].
During TLS, replicative DNA polymerase is temporarily replaced by a specialized TLS polymerase that has the ability to replicate across DNA lesions. TLS polymerases can use the damaged template for synthesis and bypass DNA lesions, may incorporate the non-complementary nucleotides thereby facilitating replication fork progression [8]. These polymerases include B-family Pol ζ and Y-family Pol η, Pol κ, Pol ι and Rev1.
Pol ι efficiently bypasses a variety of DNA lesions including lesions induced by oxidative stress but possesses very low accuracy of nucleotide incorporation [9,10]. Pol ι efficiently incorporates deoxynucleotides opposite the 3' T of [6-4] PPs, abasic sites [11], and opposite N2-adducted guanines [11], products of cytosine oxidation [12] and 8-oxoguanine (8-oxoG) [13], the most abundant and mutagenic oxidative lesion in DNA. Pol ι also plays TLS-independent functions in DDT.
Several SNPs in POLI (encoding Polι) have been linked to the development of melanoma, prostate cancer, lung adenocarcinoma and squamous cell carcinoma [14,15,16]. POLI expression was elevated in breast cancer [13], bladder cancer [17] and oesophageal squamous cell carcinoma (ESCC) [18]. At the same time, Pol ι inhibited tumorigenesis and mutagenesis in some cancers, despite its extreme replication infidelity. Polι deficiency in mice leads to increased susceptibility to UV-induced skin tumors [19,20]. Pol ι also protects from chemically induced lung cancer. Polι was found to be the Par2 (pulmonary adenoma resistance 2) gene responsible for adenoma and adenocarcinoma susceptibility in mice [21,22,23]. In addition, the activation of Polι triggers the Erk and JNK signaling pathways, which play a pivotal role in promoting invasion and metastasis via EGFR-ERK-mediated epithelial to mesenchymal transition [24,25]. Ultimately, this leads to the increase in proliferation and a grim prognosis for individuals with ESCC [18,25,26]. However, whether Polι plays a role in proliferation and ionizing radiation-induced DNA damage response (DDR) of NSCLC cells remains unclear.
Recently, a novel primase-polymerase called PrimPol has been identified in eukaryotic cells [27]. PrimPol promotes DNA repriming downstream of a lesion or replication block to facilitate fork restart following ultraviolet (UV) damage and is important for inter-strand cross-link traverse [28]. PrimPol synthesizes DNA primers de novo, restarting replication in the nucleus and mitochondria after damaged DNA regions or secondary structures—G quadruplexes and R loops [28,29,30,31,32,33,34,35]. PrimPol also exhibits properties of a translesion DNA polymerase, effectively incorporating nucleotides opposite a number of small DNA lesions, including 8-oxoguanine, O6-methylguanine, 5-formyluracil, and apurine-apyrimidine sites in vitro [36,37,38]. PrimPol also quite effectively incorporates complementary dCMPs opposite the intra-strand cisplatin crosslink in vitro [39]. Deletion of the Polι gene in a mouse embryonic fibroblast cell line slows down the replication of nuclear and mitochondrial DNA under physiological conditions and causes sensitivity to a number of genotoxic agents: UV radiation, methyl methanesulfonate, hydroxyurea, benz[a]pyrene and cisplatin [29,32,34,40,41,42,43]. PRIMPOL −/− cells were more sensitive to UV-C irradiation in colony survival assays than wild-type and Pol η-deficient cells [40]. Mutations in PRIMPOL have also been found in human cancers [44,45,46,47,48,49] and compiled in Genomic Data Commons Data Portal, GDC [50] and Catalogue of Somatic Mutations in Cancer, COSMIC databases [51]. The essential role of PRIMPOL in ensuring the smooth progression of DNA replication forks highlights the tremendous potential of inhibiting this non-essential mammalian enzyme. This compelling strategy offers a promising avenue for effectively treating various conditions by disrupting DNA synthesis.
The lack of a particular type of DNA polymerase is anticipated to have a profound effect on the DDR. This is expected to increase the replication stress caused by DNA damage [52,53], leading to targeting differentiated and cancer stem cells/stem cell-like cells (CSCs) [54] by potential activation of apoptosis. However, the impact of PRIMPOL and POLI elimination in NSCLC cells exposed to ionizing radiation has not been elucidated.
In this study, we aimed to evaluate the impact of PRIMPOL and POLI elimination on human A549 NSCLC cell line exposed to X-ray irradiation stress. We observed significant differences in cell survival, apoptosis, DNA repair and the expression of cancer stem cells (CSCs) markers in PRIMPOL and POLI knockout cells compared to wild-type cells.
2. Results
2.1. Evaluation of Invasion, Cell Cycle Progression and Cell Death of PRIMPOL-/- and POLI -/- Cell Lines.
To analyze functions of PrimPol and Polι, two crucial enzymes involved in DNA damage tolerance, we investigated A549 cell lines harbouring PRIMPOL and POLI knockouts generated using CRISPR-Cas9 technology [55].
The T-Distributed Stochastic Neighbor Embedding (t-SNE) machine learning algorithm has enabled the analysis of single-cell expression distribution in different cancers, leveraging the CancerSEA database [56]. This analysis revealed possible participation of PRIMPOL in various crucial biological functions such as cell invasion, cell cycle regulation, apoptosis, and hypoxia response. Currently, there is a concerning lack of direct experimental proof exploring the significance of PRIMPOL and POLI in these essential biological functions of NSCLC cells.
Hence, we tested first KO cell lines for cancer cell biophysical (mechanobiological) trait, persistency in cell migration under spatial confinement. Boyden chambers assay is a commonly employed assay that measures the capacity of cell motility and invasiveness toward a chemo-attractant gradient under spatial confinement of porous matrix. Comparing to parental A549 wild-type cells, both PRIMPOL-/- and POLI −/− cell lines showed significant decrease in cell motility and invasiveness through 8 µm pores toward a chemo-attractant gradient (Figure 1).
Knockout of genes involved in DDT response could potentially slow down the progression of replication fork following irradiation. To examine this cell cycle analysis using PI staining was conducted 48 and 72 h of cultivation without irradiation and after 6 Gy X-rays exposure.
Without irradiation, PRIMPOL and POLI knockouts themselves did not influence the redistribution of cells between cell cycle phases at 48 h of cultivation. Decrease in proportion of POLI -/- cells in S phase was significant, while demonstrating only the same trend for PRIMPOL-/- cells by 72 h of cultivation (Figure 2b).
6 Gy irradiation induced the decrease of the proportion of wild-type cells in G1 phase, while both PRIMPOL-/- and POLI -/- cells did not change this proportion significantly compared to non-irradiated cells (Figure 2a,b vs. c,d). The decrease in the S phase after IR was almost equal in all studied cells (1.4 - 3 times compared to corresponding non-irradiated cells). Comparing to non-irradiated cells, all cell lines demonstrated more than 2 times accumulation in G2 phase following 6 Gy exposure. However, the proportion of PRIMPOL-/- and POLI -/- cells accumulated in G2 phase were significantly lower compared to wild-type cells (for POLI p<0.05 and p<0.01 at 48 and 72 h post-irradiation, respectively; for PRIMPOL-/- p<0.01 at 72 h post-irradiation) (Figure 2c,d).
Next, we used X-ray irradiation, a widely utilized technique in cancer treatment and a well-established stimulus for activating apoptosis. The quantification of the proportion of apoptotic cells was conducted using well-established YO-PRO-1 and PI staining initially after irradiation (0 h time-point) or 48 h post-irradiation [57].
Without irradiation, in initial time-point the knockout cells displayed nearly double the baseline apoptosis compared to the parental cells, however, this distinction did not reach statistical significance (Fig. 3a-c, time 0 h). It was observed that, after 48 hours of cultivation, POLI −/− cells exhibited remarkably reduced apoptosis, which was in stark contrast to the PRIMPOL −/− cells (Fig.2 b-c, time 48h), when compared to their respective initial values (0 h). Of note, elimination of PRIMPOL and POLI leads to significant upregulation of cellular basal apoptosis compared to wild-type (WT) parental cells by day 7 (Fig.3d) of their cultivation.
The effect of PRIMPOL and POLI knockouts on cell apoptosis was also quantified at 48 h after exposure to 8 Gy of X-rays. By that time, both WT and POLI −/− cells demonstrated significant increases in apoptosis over corresponding controls (p < 0.01, Figure 3a,c), while there was no difference between control and irradiated PRIMPOL −/− cells. (Figure 3b). Notable, the total proportion of YO-PRO-1 positive cells was almost equal in all irradiated cell lines. These experimental findings unequivocally show that the lack of PRIMPOL and POLI did not lead to a heightened rate of apoptosis (compared to unirradiated control at the initial time of the experiment, 0 h) 48 hours after being exposed to a significant dose of ionizing radiation.
2.2. Radiosensitivity of PRIMPOL-/- and POLI -/- Cell Lines Using Clonogenic Assay
The colony formation assay, or clonogenic assay, is an in vitro quantitative technique designed to measure the ability of a single adherent cell to survive over time and expand into a clonal population [58]. Under DNA-damaging conditions induced by X-ray exposure, the dose-related higher clonogenic survival (survival fraction) will indicate higher resistance (lower sensitivity) to DNA-damaging insult.
A conventional colony formation assay was performed to determine the impact of PRIMPOL and POLI knockouts on cells’ radiosensitivity. Both PRIMPOL -/- and POLI −/− cell lines showed decreased radiosensitivity compared to parental A549 wild-type cells growing under anchorage-dependent conditions (Figure 4). However, the statistical significance was obtained only for PRIMPOL -/- cells after irradiation with 6 Gy (**p < 0.01).
2.3. Comparative Analysis of γH2AX Kinetics, Phosphorylated Ataxia Telangiectasia Mutated (pATM) and Rad51 in wild-type, PRIMPOL-/- and POLI -/- Cell Lines
A rapid phosphorylation of the minor histone variant H2AX occurs in response to DNA double-stranded breaks (DSBs). This process takes place at Ser-139 in mammals, leading to the creation of γH2AX [59]. The quantification of individual γH2AX foci through fluorescence microscopy is now the preferred method for detecting DNA DSBs, especially when the radiation dose does not exceed 2 Gy. It has been determined that each break is equivalent to one γH2AX focus. In order to investigate the influence of PRIMPOL and POLI knockout on the repair of DNA double-strand breaks (DSBs) following 2 Gy irradiation, we conducted an analysis of the kinetics of γH2AX foci (Figure 5a). The majority of knockout cells exhibited pan-nuclear γH2AX staining, indicating a significant buildup of replication stress (nuclear images not displayed). The analysis of γH2AX foci number yielded contrasting results for the knockout cell lines. The POLI -/- cells exhibited the lowest level of γH2AX foci, both at their baseline and after exposure to 2 Gy. On the other hand, the PRIMPOL-/- cells displayed significantly higher numbers of foci compared to POLI-/- and wild-type (for 4 hours only) cells, even without any irradiation present (Figure 5a). Compared to other cell lines, PRIMPOL cells exhibited the highest number of γH2AX foci immediately after irradiation (1 hour) (*p < 0.05 compared to wild-type). Moreover, the dephosphorylation kinetics of γH2AX foci in PRIMPOL-/- cells were found to be the slowest. Even 24 h after irradiation these cells had almost 7 times higher number of γH2AX foci compared to POLI-/- cells. Thus, the PRIMPOL and POLI knockout had a different impact on the kinetics of γH2AX foci number in A549 cells after X-ray irradiation.
The ATM protein, or ataxia-telangiectasia mutated, plays a crucial role in regulating the DDR. It specifically handles the repair of DNA DSBs. By phosphorylating Serine 139 on the C-terminal tail of the histone variant H2AX, it provides a significant and noteworthy contribution to DNA repair [60]. The activation of the ATM protein by DNA double-strand breaks (DSBs) was investigated to assess the dynamics of pATM foci in A549 wild-type, PRIMPOL-/- and POLI -/- cells. Unlike the kinetics of γH2AX foci, all the studied cell lines demonstrated the same kinetics of pATM foci (Figure 5b). This data suggests that phosphorylation of γH2AX in PRIMPOL -/- cells was ATM-independent.
Rad51 is the central homologous recombination (HR) enzyme and responsible for replication fork processing and restart during replication stress [61]. In order to determine if the elimination of PRIMPOL and POLI affects the formation of Rad51 foci after exposure to radiation, the number of Rad51 foci was analyzed over time. The formation of Rad51 foci is contingent upon the cell cycle and linked to the gradual process of homologous recombination (HR). Thus, the highest count of Rad51 foci was detected no sooner than 6 hours after irradiation and returned to basal levels by the end of 24 hours (Figure 5c). Despite the analysis revealing similar kinetics for all cell lines (Figure 5c), POLI -/- cells exhibited a decreased number of Rad51 foci compared to wild-type and PRIMPOL-/- cells exposed to 2 Gy IR. Both unexposed PRIMPOL-/- and POLI -/- cells demonstrated 1.5-2 times lower numbers of foci compared to wild-type cells. However, the statistical significance was seen only between POLI -/- and wild-type cells after 24 h of incubation (p<0.05). Thus, our data indicates that POLI can be directly associated with the expression of Rad51.
2.4. The Proportion of Cancer Cells with Stem-like Cells in PRIMPOL-/- and POLI -/- Cell Lines with and without IR Exposure.
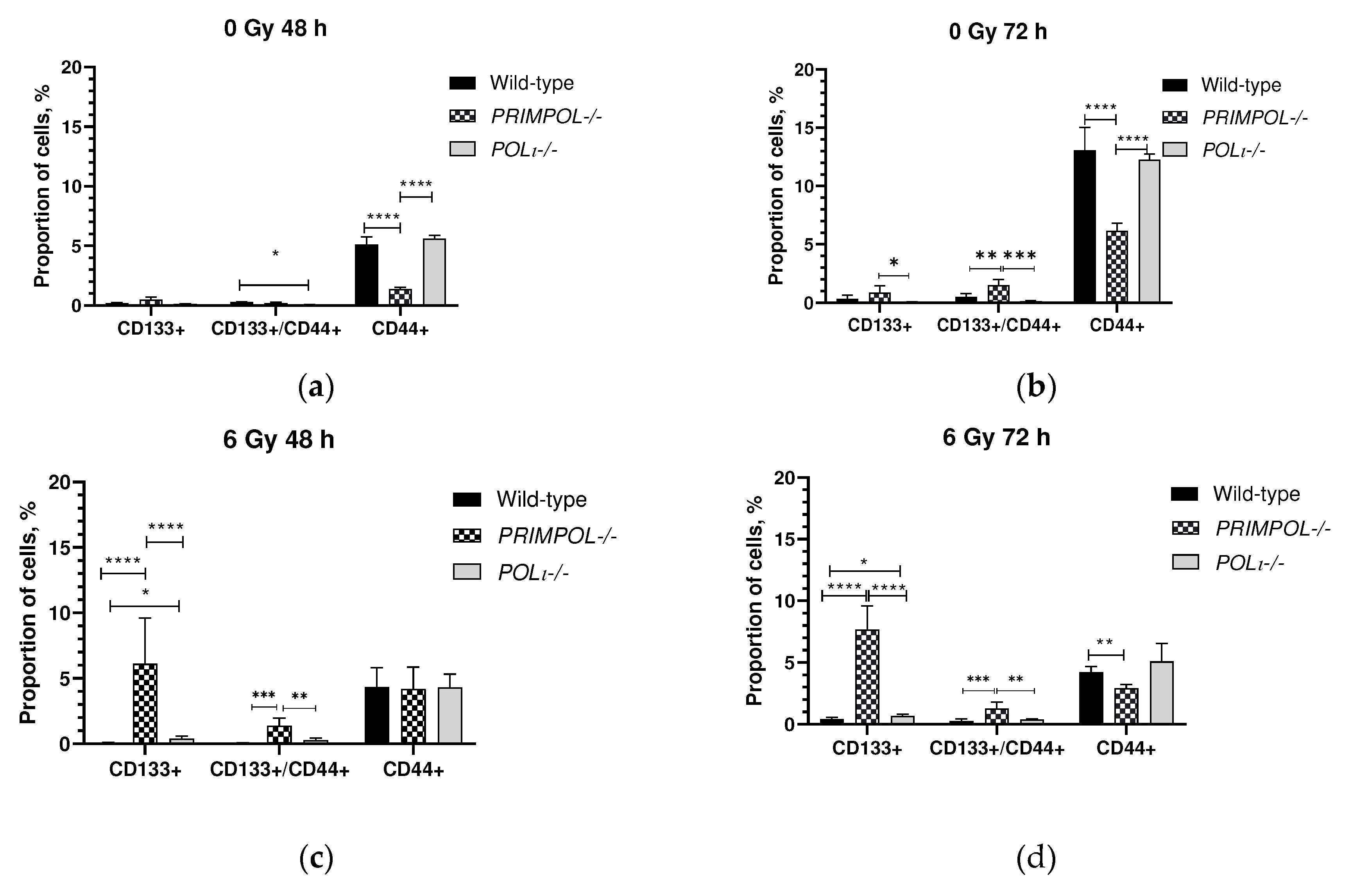
Clonogenic activity is a sensitive indicator of the presence of undifferentiated cancer stem-like cells (CSCs) [62]. In order to determine if there is an association between increased clonogenic survival of PRIMPOL-/- cells (Fig.4) and possible presence of CSC-like cells, we analyzed the proportion of CD44+ and CD133+ populations 48 and 72 h after being exposed to 6 Gy. Extending the incubation period from 48 h to 72 h has resulted in a greater than 2-fold increase in the proportion of CD44+ cells across all examined cell lines (Figure 6a,b). Nevertheless, in the absence of irradiation, it was observed that PRIMPOL-/- cells exhibited a strikingly lower number of CD44+ cells when compared to both wild-type and POLI -/- cells (p < 0.0001). 6 Gy exposure did not show any significant effect on the proportion of CD44+ cells (Figure 6c,d). Simultaneously, irradiation was found to significantly increase the population of CD133+ PRIMPOL-/- cells (p < 0.0001, 48 and 72 hours after 6 Gy, as shown in Figure 6c,d). Interestingly, irradiation also had an impact on CD133+/CD44+ cells, as their proportion was statistically significantly higher in PRIMPOL-/- cells compared to wild-type (p < 0.001) and POLI -/- cells (p < 0.01).
3. Discussion
PrimPol plays a crucial role in facilitating the resumption of DNA replication after encountering DNA lesions, such as DNA interstrand cross-links [28]. This remarkable ability is demonstrated through its involvement in DNA replication-restart pathways [41,63,64,65]. Pol iota (Polι) seems to be primarily responsible for regulating a DDT pathway choice that determines how DNA damage is tolerated [66]. This pathway choice involves a competitive interaction between PrimPol-mediated repriming and fork reversal. It should be noted that this function is distinct from Pol iota's role in DNA replication and DNA damage repair [9]. Instead, it serves as an important protective enzyme, even though it does play a significant role in TLS. Together, these data suggest that the significance of specialized DNA polymerases in the DDR extends beyond their participation in TLS. TLS is generally regarded as mechanistically simple and straightforward, but error-prone DDT pathway as it has high potential for mutagenesis [67]. These include dormant origin firing, homologous recombination (HR) and specialized DNA polymerases involved in TLS bypass of replication-stalling lesions [68,69]. Heterogeneous fidelity of TLS polymerases together with different combinations of polymerases/lesions/sequence contexts can result in distinct sets of mutated genes and thus the development of specific cancers [70]. Recent studies indicated that TLS primase/polymerase PrimPol is possibly associated with the development and chemosensitivity of many cancers [32,71]. In present study, we aimed to evaluate the impact of PRIMPOL and POLI genes on metastatic potential and radiosensitivity of human NSCLC cells.
As was previously shown by others, the ability to migrate, structure and mechanics of cancer cells are linked directly to their metastatic potential [72,73,74,75,76]. Recently, machine learning algorithm-based single-cell analysis revealed possible participation of PRIMPOL in various crucial biological functions such as cell invasion, cell cycle regulation, apoptosis, and hypoxia response [56].
Recent evidence has shown a compelling connection between the ability of cancer cells to migrate through 8 µm pores and their capacity to indent and display metastatic behavior [77]. Hence, we employed this method to assess the motility and invasiveness of both wild type and polymerase knock-out cells in response to a serum gradient while confined within a porous matrix of Boyden chambers. Our data has uncovered a significant decline in a vital biophysical (mechanobiological) characteristic of cancer cells - their ability to withstand migration while being confined to a specific area for three consecutive days. This decline was observed in both knockout (KO) cell lines, as shown in Figure 1, when compared to their syngeneic wild type counterparts. It is true that both KO cell lines had a decrease in S-phase cells after 72 hours of cultivation (Fig. 2b). However, it is highly improbable that cell cycle disruptions are the cause of the observed reduced migration. Figure 3 (a-c) illustrates the remarkable increase in baseline apoptosis in KO cells. The levels reach a peak nearly twice as high as those in their parental counterparts. On the seventh day of cell cultivation, the frequency of basal apoptosis increases significantly in both knockout cell lines, as shown in Figure 3d. This finding can be pinpointed as the possible cause behind the observed migratory decline effect. Thus, our current study offers experimental evidence affirming the AI-based algorithm's prediction of a positive correlation between PRIMPOL expression and migration/invasiveness, as well as cell cycle progression [56]. Additionally, we have demonstrated an inverse relationship between PRIMPOL expression and the level of apoptosis in NSCLC cells.
Furthermore, we have effectively presented similar discoveries and connections regarding the role of POLI deletions in these essential biological functions of the same NSCLC cells. It was previously demonstrated that Polι promotes cell migration and invasion of ESCC and breast cancer cells [18,24,25,78]. Our data strongly supports the effectiveness of small interfering RNA (siRNA) targeting the POLI gene in inhibiting invasion of the A549 lung cancer cell line [79]. It is important to mention that this treatment has had contrasting effects on apoptosis as compared to the results we obtained from knocking out POLI gene (Fig. 3d) of the same cell line. The use of siRNA to partially silence this gene had little impact on the cellular MTT-detected activity, which was mistakenly attributed to apoptosis by these authors. Our groundbreaking study reveals that completely knocking out POLI gene significantly enhances the occurrence of true apoptosis, establishing a clear inverse correlation with the activity of the POLI gene.
Although PRIMPOL and POLI knockouts augment basal apoptosis, their effect on reproductive cell survival following exposure to ionizing radiation (IR) was increased (Fig. 4). Notably, this increase reached statistical significance only in PRIMPOL-/- cells after irradiation with 6 Gy, indicating their increased DDT. While our current study did not prioritize it, the data we collected do not rule out the possibility of anastasis. Anastasis is a remarkable cellular recovery phenomenon that saves dying cells from the edge of death [80]. The unique response to IR stress, characterized by activated metastasis, epithelial mesenchymal transition (EMT), and DNA damage repair mechanisms promoting anastasis [81], may still occur differentially in PRIMPOL and POLI knockouts. Thus, our data is in line with the clinical observation that the expression of PRIMPOL is decreased in lung adenocarcinoma when compared to normal tissues. This decrease was significantly linked to patients' prognosis [56].
The absence of specialized DNA polymerases can have a significant impact on various aspects of DDR. It is highly likely that without these enzymes, there would be an increase in replication stress caused by DNA damage [52,53]. To evaluate whether the efficiency of DNA DSBs repair involved into increased DDT of PRIMPOL -/- and POLI -/- cell clones, we evaluated γH2AX and pATM foci kinetics.
ATM, ATR, and DNA-PK are three phosphoinositide 3-kinase (PI3K)-related kinases that phosphorylate H2AX in response to DNA damage [82]. While ATM is considered as a major physiological mediator of H2AX phosphorylation in response to DSB formation [83], ATR phosphorylates H2AX in response to single-stranded DNA breaks and during replication stress, such as replication fork arrest [84]. We observed no discrepancies in the number of pATM foci between the cell lines being compared. This compelling finding suggests that the absence of PRIMPOL and POLI does not have any impact on the cellular capacity to repair 'true' DNA DSBs.
The levels of γH2AX are a crucial indicator of DNA damage throughout the entire organism. Examining these levels can provide invaluable insights into the degree of replication stress in the absence of specialized DNA polymerases. Therefore, we examined the accumulation of γH2AX in cancer cells lacking either PRIMPOL or POLI and exposed to ionizing radiation (IR), a commonly used stimulus that induces replication stress. We observed a significant disparity in the number of γH2AX foci between the compared cells (Fig.5a). We observed a decrease in γH2AX levels in cells that lacked POLI, whereas the absence of PRIMPOL led to a significant increase in both the baseline and residual γH2AX foci numbers induced by ionizing radiation (Fig. 5a). Of note, PRRIMPOL knock-out also augments the Rad51 foci, albeit during the first four hours after irradiation (Fig. 5c). Upon irradiation PrimPol is able to bind to replication protein A (RPA) [85]. RPA is the single-stranded DNA (ssDNA)-binding protein and one of the proteins required for the maintenance of genetic information [86]. Since the pATM foci number remained unchanged among populations (Fig. 5b), the remarkable increase in the number of γH2AX foci in PRIMPOL-/- cells could be linked to the known deficiencies in fork restart after replication arrest [85].
POLI expression in Esophageal Squamous Cell Carcinoma (ESCC) was negatively correlated with overall survival of ESCC patients with postoperative adjuvant radiotherapy, suggesting its role in ESCC radioresistance [87]. Recently, the requirement of POLI for optimal γH2AX and 53BP1 accumulation has been shown [66]. The study conducted by Sabrina F. Mansilla et al. yielded similar results, demonstrating a reduction in γH2AX and 53BP1 levels following treatment of Polι-depleted cells with cisplatin [66]. Of note, POLI knock-out also mitigated the accumulation of pan-nuclear Rad51 foci during the first 4 and 24 h after irradiation (Fig. 5c). Our data indicates that POLI deletion might be directly associated with either translational or post-translational down-regulation of RAD51 as was shown previously [66,87]. Important, RAD51 is one of three well-known proteins (behind BRCA1, BRCA2) that negatively regulate the PRIMPOL expression. The role of Rad51 in homologous recombination is well-established. It is worth mentioning that this protein is also linked to fork reversal (FR) [88] and is a favored binding partner of POLι in non-differentiated cancer stem-like cells [54]. Consequently, the concurrent reduction of Rad51 and elimination of POLι are highly likely to stimulate a greater reliance on PrimPol-dependent repriming [89]. This process might significantly impair the p53-POLι-mediated FR-DDT pathway in cell populations that resemble CSCs. If that's true, then it supports the idea that POLI has a critical influence on the selection of the differential DDT pathway [54]. Such a change in the DDT pathway could potentially explain why POLI -/- cells have a lower clonogenic survival rate than PRIMPOL -/- cells following exposure to IR stress in our study.
In the absence of POLI, PRIMPOL to take over and resulted in enhanced repriming, DNA replication acceleration, and bypassing of the checkpoint activation during the S phase. Consequently, this led to a surge in chromosome instability during the M phase [66]. Our findings (Fig. 2c,d) support the observation of G2/M arrest instead of S phase arrest after exposure to IR. According to our data, POLI knocked-out cells likely have a remarkable ability to amplify replication stress after being exposed to IR. This finding challenges the conventional understanding of other specialized DNA polymerases, which typically counter this stress instead.
Cancer stem-like cells (CSCs) are a subpopulation of tumor cells that can drive tumor initiation and recurrence [90]. PRIMPOL mutations have been discovered in human cancers, but what's even more fascinating is its over-expression in CD 133-positive CSC-like glioblastoma subtypes [91]. These points to potential functions in combating replication stress in diseased tissues. Since PrimPol seems to have crucial functions in sustaining RF progression, despite not being a necessary enzyme in mammals, the development of PrimPol inhibitors to interrupt DNA synthesis could potentially offer a viable approach for treating various conditions. Here, we present evidence that a dosage of 6 Gy IR could trigger a remarkable surge in the proportion of CD133+ and CD133+/CD44+ populations in PRIMPOL-/- cells. The data indicate that their greater clonogenic survival (Fig.4) could be linked to the existence of these CSC-like cells. Additional research is necessary to understand better the mechanisms that determine the choice of the DDT pathway and the subsequent induction of CSC-like cells by PRIMPOL-/- cells in response to IR stress. It is critical to exercise caution when developing inhibitors for specialized DNA polymerases in cancer treatment, as our data shows a strong indication for potential complications.
4. Materials and Methods
4.1. Cell Cultures and Cultivation conditions
The PRIMPOL and POLI knocked out A549 cells were kindly provided by Dr. A. Makarova (Institute of Gene Biology of Russian Academy of Sciences, Moscow, Russia) and Prof. D.O. Zharkov (ICBFM SB RAS, Novosibirsk, Russia). Cells were obtained using the CRISPRrispr/Cas9 system and the pSpCas9(BB)-2A-GFP vector as previously described [55]. The absence of PrimPol and Polι in cells was verified by immunoblotting (Figure S1) Cells were cultured in DMEM with high glucose content GlutaMAX™ Supplement (Gibco, USA), 10% fetal bovine serum, 100 U/mL penicillin, and 100 U/mL streptomycin at 37°C and 5% CO2.
4.2. Trans-Well Migration Assay Using Boyden Chambers
Briefly, cells were serum starved overnight and 104 cells were seeded in 0,1 ml of serum-free cell-type specific media in the upper compartment of transwell inserts (EDM Milli-pore, Billerica, MA) with 8 µm pores. Serum-supplemented growth media was placed in the lower chamber, serving as a chemoattractant. Cells were incubated in the wells for 72 h at 37 °C and 5% CO2. The cells from the upper and lower compartments were collected, by incubation with 0,2 and 0,3 ml of trypsin-EDTA solution (0.25%; Biological Industries, Cromwell, CT), respectively, for 5 min at 37 °C and under 5% CO2. After trypsin neutralization with serum-containing growth media the cells collected from upper and lower chamber were counted using hemocytometer. For each cell type 6–9 independent experiments were performed, and the percentage of cancer cells from all cell lines that cross through 8 µm Boyden chamber membranes was determined.
4.3. Apoptosis Analysis
To quantify the proportion of early-stage apoptotic cells the commercial kit “Vybrant Apoptosis Assay Kit #4” with YO-PRO-1 and PI for Flow Cytometry (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA; catalog number: V13243) was used. The cells were stained according to the supplemented manufacturer protocol. Immediately (0 hours) or 48 hours post-radiation, cells were collected and washed in cold phosphate-buffered saline (PBS). A total of 1 μL of YO-PRO-1 stock solution and 1 μL of PI stock solution were added to each 1 mL containing 1 × 106 cells. Cells were incubated on ice for 20–30 min and analyzed by flow cytometry (BD FACSCalibur, Becton Dickinson, San Jose, CA, USA) using 488 nm excitation with green fluorescence emission for YO-PRO-1 (i.e., 530/30 bandpass) and red fluorescence emission for PI (i.e., 610/20 bandpass). A total of 50,000 events were acquired for each sample and analyzed with FlowJo™ Software (Becton Dickinson, San Jose, CA, USA).
4.4. Colony Formation and Soft Agar Assay
Cells were exposed to 0 Gy, 2 Gy, 4 Gy, and 6 Gy of X-ray irradiation, plated on 60 mm Petri dishes in 150, 500, 1000, and 2000 cells/well, respectively and incubated at 37°C 5% CO2. After two weeks, cells were fixed with 100% methanol for 15 min at room temperature, followed by Giemsa staining for 15 min. Only colonies containing more than 50 cells were counted. Plating efficiency (PE) and survival fractions (SF) were calculated using the following equations:
PE = number of colonies formed/number of cells seeded× 100%
SF = number of colonies formed /( number of cells seeded × PE)
4.5. γH2AX, pATM and Rad51 Foci Analysis
Cells were seeded in 96-well plates, fixed 1–24 h after irradiation in 4% paraformaldehyde for 15 min at room temperature and permeabilized in 0.3% Triton-X 100 (in PBS, pH 7.4) supplemented with 2% bovine serum albumin (BSA). Cells were incubated for 1 h at room temperature with primary mouse monoclonal antibody against γH2AX (dilution 1:200, clone JBW 301, Cat. # 05-636, Merck-Millipore, Burlington, VT, USA), primary mouse monoclonal antibody against pATM (dilution 1:200, clone 10H11.E12, Merck Millipore, Burlington, VT, USA) and with primary rabbit polyclonal antibody against Rad51 (dilution 1:200, Merck Millipore, Burlington, VT, USA) diluted in PBS with 1% BSA and 0.3% Triton-X 100. After several rinses with PBS, cells were incubated for 1 h with secondary antibodies IgG (H + L) goat anti-mouse (Alexa Fluor 555 conjugated, dilution 1:800; Cat. # A-21424, Merck-Millipore, Burlington, VT, USA), goat anti-rabbit (Alexa Fluor 488 conjugated, dilution 1:500; Cat. # A-11008, Merck Millipore, Burlington, VT, USA) diluted in PBS (pH 7.4) with 1% BSA. Cells were imaged using Nikon Eclipse Ni-U microscope (Nikon, Tokyo, Japan) equipped with a high-definition camera ProgResMFcool (Jenoptik AG, Jena, Germany). Filter sets used were UV-2E/C (340–380 nm excitation and 435–485 nm emission), B-2E/C (465–495 nm excitation and 515–555 nm emission), and Y-2E/C (540–580 nm excitation and 600–660 nm emission). A total of 300–400 cells were imaged for each data point. Foci were counted by manual scoring.
4.6. Analysis of the Proportion of Cancer Stem Cells by Flow Cytometry
Cells were collected by trypsinization, washed in ice-cold PBS (pH = 7.4) and 1 × 106 cells per sample were incubated with Anti-CD133 Antibody, Alexa Fluor® 488 conjugated (MAB4310X, Sigma-Aldrich, Darmstadt, Germany) and with monoclonal Anti-CD44−PE antibody (SAB4700187, Sigma-Aldrich, Darmstadt, Germany) for 30 min at 4 °C. Cells were analyzed by flow cytometry (BD FACSCalibur, Becton Dickinson, San Jose, CA, USA). A total of 50,000 events were acquired for each sample and the proportion of positive cells was analyzed with BD CellQuest Pro 5.1 software (Becton Dickinson, San Jose, CA, USA).
4.7. Cell Cycle Analysis by Flow Cytometry
Exponentially growing cells were irradiated (6 Gy) and incubated for 48 and 72 hours post-radiation. Cells were then harvested and 1 × 106 cells per sample were resuspended in ice-cold PBS. Cells were fixed in ice-cold 70% ethanol for 30 min at 4 °C and then kept at −20 °C until analyzed. Before analysis, specimens were washed twice with PBS, and resuspended in 0.5 mg/ml PI containing 50 μl of 100 μg/ml RNase for 30 min in the dark. Cells were analyzed by flow cytometry (BD FACSCalibur, Becton Dickinson, San Jose, CA, USA). A total of 50,000 events were acquired for each sample and the percentage of cells in the different phases of the cell cycle was analyzed with FlowJo™ Software (Becton Dickinson, San Jose, CA, USA).
4.8. Statistics
Statistics were performed using GraphPad Prism 9.0.2.161 (GraphPad Software, San Diego, CA, USA) software. Statistical significance was tested using the Student t-test. The results are represented as means ± SEM of more than three independent experiments. Significance levels were denoted by asterisks: * p < 0.05, ** p < 0.01, *** p < 0.001, ****p < 0.0001.
5. Conclusions
Here, we explored the effects of the absence of PRIMPOL (PRIMPOL −/−) and POLI (POLI −/−) genes, which encode two TLS polymerases, in two different monoclonal lines derived from human NSCLC A549 cells. Without IR, cells that do not have PRIMPOL or POLI experience a considerable rise in apoptosis and a noteworthy decline in cell migration under spatial confinement when compared to the parental wild-type cells. Despite this, we have found that PRIMPOL −/− exhibited a remarkably higher clonogenic survival compared to POLI -/- cell lines in response to IR stress. Both of these cell lines demonstrated remarkable resilience to radiation stress, surpassing the survival rate of the original A549 cells. The observed effect may be due to the significantly higher number of both acute and residual γH2AX foci in PIMPOL-deficient cells compared to the wild type parental and POLI-deficient cells. In fact, the latter displayed the lowest capacity to accumulate γH2AX foci 48 hours after exposure to ionizing radiation. In addition, we are excited to share groundbreaking discoveries that propose a connection between improved survival of PRIMPOL −/− cells after radiation and a rise in cancer stem-like cells characterized by an increase in CD133+ populations, rather than CD44+ populations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.P.; formal analysis, M.P., Z.N.; investigation, M.P., T.B., A.G., Z.N. and I.M.; writing—original draft preparation, M.P.; writing—review and editing and supervision, S.L.; visualization, M.P., Z.N.; project administration, M.P.; funding acquisition, M.P. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Higher Education of the Russian Federation (Goszadaniye) 075-03-2023-106, project № FSMG-2023-0015. This work was also supported (for S.L. and Z.N.) by grant from the Russian Science Foundation (No. 23-24-00601).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The materials and data are available from the corresponding authors.
Acknowledgments
The authors would like to thank Dr. A. Makarova (Institute of Gene Biology of Russian Academy of Sciences, Moscow, Russia) and Prof. D.O. Zharkov (ICBFM SB RAS, Novosibirsk, Russia) for the provided cell lines used in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wright, W.D., S.S. Shah, and W.-D. Heyer, Homologous recombination and the repair of DNA double-strand breaks. Journal of Biological Chemistry, 2018. 293(27): p. 10524-10535. [CrossRef]
- Lieber, M.R., The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu Rev Biochem, 2010. 79: p. 181-211. [CrossRef]
- Marteijn, J.A., et al., Understanding nucleotide excision repair and its roles in cancer and ageing. Nat Rev Mol Cell Biol, 2014. 15(7): p. 465-81. [CrossRef]
- Krokan, H.E. and M. Bjoras, Base excision repair. Cold Spring Harb Perspect Biol, 2013. 5(4): p. a012583.
- Li, G.M., Mechanisms and functions of DNA mismatch repair. Cell Res, 2008. 18(1): p. 85-98. [CrossRef]
- Qiu, S., et al., Replication Fork Reversal and Protection. Front Cell Dev Biol, 2021. 9: p. 670392. [CrossRef]
- Vaisman, A., J.P. McDonald, and R. Woodgate, Translesion DNA Synthesis. EcoSal Plus, 2012. 5(1).
- Sale, J.E., Competition, collaboration and coordination--determining how cells bypass DNA damage. J Cell Sci, 2012. 125(Pt 7): p. 1633-43.
- Vaisman, A. and R. Woodgate, Mysterious and fascinating: DNA polymerase i remains enigmatic 20 years after its discovery. DNA Repair (Amst), 2020. 93: p. 102914. [CrossRef]
- Makarova, A.V. and A.V. Kulbachinskiy, Structure of human DNA polymerase iota and the mechanism of DNA synthesis. Biochemistry (Mosc), 2012. 77(6): p. 547-61. [CrossRef]
- Washington, M.T., et al., Efficient and error-free replication past a minor-groove DNA adduct by the sequential action of human DNA polymerases iota and kappa. Mol Cell Biol, 2004. 24(13): p. 5687-93.
- Vaisman, A. and R. Woodgate, Unique misinsertion specificity of poliota may decrease the mutagenic potential of deaminated cytosines. EMBO J, 2001. 20(22): p. 6520-9. [CrossRef]
- Yang, J., et al., Altered DNA polymerase iota expression in breast cancer cells leads to a reduction in DNA replication fidelity and a higher rate of mutagenesis. Cancer Res, 2004. 64(16): p. 5597-607.
- Luedeke, M., et al., Predisposition for TMPRSS2-ERG fusion in prostate cancer by variants in DNA repair genes. Cancer Epidemiol Biomarkers Prev, 2009. 18(11): p. 3030-5. [CrossRef]
- Sakiyama, T., et al., Association of amino acid substitution polymorphisms in DNA repair genes TP53, POLI, REV1 and LIG4 with lung cancer risk. Int J Cancer, 2005. 114(5): p. 730-7.
- Silvestrov, P., et al., DNArCdb: A database of cancer biomarkers in DNA repair genes that includes variants related to multiple cancer phenotypes. DNA Repair (Amst), 2018. 70: p. 10-17. [CrossRef]
- Yuan, F., et al., Overexpressed DNA polymerase iota regulated by JNK/c-Jun contributes to hypermutagenesis in bladder cancer. PLoS One, 2013. 8(7): p. e69317. [CrossRef]
- Zou, S., et al., DNA polymerase iota (Pol iota) promotes invasion and metastasis of esophageal squamous cell carcinoma. Oncotarget, 2016. 7(22): p. 32274-85. [CrossRef]
- Dumstorf, C.A., et al., Participation of mouse DNA polymerase iota in strand-biased mutagenic bypass of UV photoproducts and suppression of skin cancer. Proc Natl Acad Sci U S A, 2006. 103(48): p. 18083-8.
- Ohkumo, T., et al., UV-B radiation induces epithelial tumors in mice lacking DNA polymerase eta and mesenchymal tumors in mice deficient for DNA polymerase iota. Mol Cell Biol, 2006. 26(20): p. 7696-706.
- Lee, G.H. and H. Matsushita, Genetic linkage between Pol iota deficiency and increased susceptibility to lung tumors in mice. Cancer Sci, 2005. 96(5): p. 256-9.
- Iguchi, M., et al., The error-prone DNA polymerase iota provides quantitative resistance to lung tumorigenesis and mutagenesis in mice. Oncogene, 2014. 33(27): p. 3612-7.
- Wang, M., et al., Pol iota is a candidate for the mouse pulmonary adenoma resistance 2 locus, a major modifier of chemically induced lung neoplasia. Cancer Res, 2004. 64(6): p. 1924-31.
- Zou, S., et al., DNA polymerase iota (Pol iota) promotes the migration and invasion of breast cancer cell via EGFR-ERK-mediated epithelial to mesenchymal transition. Cancer Biomark, 2019. 24(3): p. 363-370. [CrossRef]
- He, C., et al., Phosphorylation of ETS-1 is a critical event in DNA polymerase iota-induced invasion and metastasis of esophageal squamous cell carcinoma. Cancer Sci, 2017. 108(12): p. 2503-2510. [CrossRef]
- Su, Z., et al., DNA Polymerase Iota Promotes Esophageal Squamous Cell Carcinoma Proliferation Through Erk-OGT-Induced G6PD Overactivation. Front Oncol, 2021. 11: p. 706337. [CrossRef]
- Guilliam, T.A., et al., Primase-polymerases are a functionally diverse superfamily of replication and repair enzymes. Nucleic Acids Res, 2015. 43(14): p. 6651-64. [CrossRef]
- Gonzalez-Acosta, D., et al., PrimPol-mediated repriming facilitates replication traverse of DNA interstrand crosslinks. EMBO J, 2021. 40(14): p. e106355.
- Piberger, A.L., et al., PrimPol-dependent single-stranded gap formation mediates homologous recombination at bulky DNA adducts. Nat Commun, 2020. 11(1): p. 5863.
- Butler, T.J., et al., Mitochondrial genetic variation is enriched in G-quadruplex regions that stall DNA synthesis in vitro. Hum Mol Genet, 2020. 29(8): p. 1292-1309. [CrossRef]
- Svikovic, S., et al., R-loop formation during S phase is restricted by PrimPol-mediated repriming. EMBO J, 2019. 38(3).
- Quinet, A., et al., PRIMPOL-Mediated Adaptive Response Suppresses Replication Fork Reversal in BRCA-Deficient Cells. Mol Cell, 2020. 77(3): p. 461-474 e9. [CrossRef]
- Torregrosa-Munumer, R., et al., PrimPol is required for replication reinitiation after mtDNA damage. Proc Natl Acad Sci U S A, 2017. 114(43): p. 11398-11403. [CrossRef]
- Kobayashi, K., et al., Repriming by PrimPol is critical for DNA replication restart downstream of lesions and chain-terminating nucleosides. Cell Cycle, 2016. 15(15): p. 1997-2008.
- Taglialatela, A., et al., REV1-Polzeta maintains the viability of homologous recombination-deficient cancer cells through mutagenic repair of PRIMPOL-dependent ssDNA gaps. Mol Cell, 2021. 81(19): p. 4008-4025 e7.
- Makarova, A.V., et al., In vitro lesion bypass by human PrimPol. DNA Repair (Amst), 2018. 70: p. 18-24. [CrossRef]
- Guilliam, T.A., et al., Human PrimPol is a highly error-prone polymerase regulated by single-stranded DNA binding proteins. Nucleic Acids Res, 2015. 43(2): p. 1056-68. [CrossRef]
- Zafar, M.K., et al., Kinetic analysis of human PrimPol DNA polymerase activity reveals a generally error-prone enzyme capable of accurately bypassing 7,8-dihydro-8-oxo-2'-deoxyguanosine. Biochemistry, 2014. 53(41): p. 6584-94. [CrossRef]
- Boldinova, E.O., et al., Translesion activity of PrimPol on DNA with cisplatin and DNA-protein cross-links. Sci Rep, 2021. 11(1): p. 17588. [CrossRef]
- Bailey, L.J., et al., PrimPol-deficient cells exhibit a pronounced G2 checkpoint response following UV damage. Cell Cycle, 2016. 15(7): p. 908-18.
- Mouron, S., et al., Repriming of DNA synthesis at stalled replication forks by human PrimPol. Nat Struct Mol Biol, 2013. 20(12): p. 1383-9. [CrossRef]
- Schiavone, D., et al., PrimPol Is Required for Replicative Tolerance of G Quadruplexes in Vertebrate Cells. Mol Cell, 2016. 61(1): p. 161-9. [CrossRef]
- Bailey, L.J., J. Bianchi, and A.J. Doherty, PrimPol is required for the maintenance of efficient nuclear and mitochondrial DNA replication in human cells. Nucleic Acids Res, 2019. 47(8): p. 4026-4038. [CrossRef]
- Diaz-Talavera, A., et al., A cancer-associated point mutation disables the steric gate of human PrimPol. Sci Rep, 2019. 9(1): p. 1121. [CrossRef]
- Rechkoblit, O., et al., Structure and mechanism of human PrimPol, a DNA polymerase with primase activity. Sci Adv, 2016. 2(10): p. e1601317. [CrossRef]
- Boldinova, E.O., C.A.C. Manukyan capital A, and C. Makarova capital A, The DNA ligands Arg47 and Arg76 are crucial for catalysis by human PrimPol. DNA Repair (Amst), 2021. 100: p. 103048.
- Kasho, K., et al., A unique arginine cluster in PolDIP2 enhances nucleotide binding and DNA synthesis by PrimPol. Nucleic Acids Res, 2021. 49(4): p. 2179-2191. [CrossRef]
- Carvalho, G., et al., Human PrimPol Discrimination against Dideoxynucleotides during Primer Synthesis. Genes (Basel), 2021. 12(10). [CrossRef]
- Guilliam, T.A., et al., Molecular basis for PrimPol recruitment to replication forks by RPA. Nat Commun, 2017. 8: p. 15222. [CrossRef]
- Grossman, R.L., et al., Toward a Shared Vision for Cancer Genomic Data. N Engl J Med, 2016. 375(12): p. 1109-12. [CrossRef]
- Tate, J.G., et al., COSMIC: the Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res, 2019. 47(D1): p. D941-D947. [CrossRef]
- Tonzi, P. and T.T. Huang, Role of Y-family translesion DNA polymerases in replication stress: Implications for new cancer therapeutic targets. DNA Repair (Amst), 2019. 78: p. 20-26. [CrossRef]
- Bertolin, A.P., S.F. Mansilla, and V. Gottifredi, The identification of translesion DNA synthesis regulators: Inhibitors in the spotlight. DNA Repair (Amst), 2015. 32: p. 158-164. [CrossRef]
- Ihle, M., et al., Impact of the interplay between stemness features, p53 and pol iota on replication pathway choices. Nucleic Acids Res, 2021. 49(13): p. 7457-7475.
- Gromova A.S., B.E.O., Kim D.V., Chuprov-Netochin R.N., Leonov S.V., Pustopalova M.V., Zharkov D.O., Makarova A.V. , Response of PRIMPOL-Knockout Human Lung Adenocarcinoma A549 Cells to Genotoxic Stress. Biochemistry, 2023. in press.
- Deng, L., et al., Multi-omics analysis of DNA replication-associated primase polymerase (PRIMPOL) in pan-cancer: a potential target for prognosis and immune response. Eur J Med Res, 2023. 28(1): p. 207. [CrossRef]
- Fujisawa, S., et al., Evaluation of YO-PRO-1 as an early marker of apoptosis following radiofrequency ablation of colon cancer liver metastases. Cytotechnology, 2014. 66(2): p. 259-73. [CrossRef]
- Franken, N.A., et al., Clonogenic assay of cells in vitro. Nat Protoc, 2006. 1(5): p. 2315-9. [CrossRef]
- Mah, L.J., A. El-Osta, and T.C. Karagiannis, gammaH2AX: a sensitive molecular marker of DNA damage and repair. Leukemia, 2010. 24(4): p. 679-86.
- Burma, S., et al., ATM phosphorylates histone H2AX in response to DNA double-strand breaks. J Biol Chem, 2001. 276(45): p. 42462-7. [CrossRef]
- Mason, J.M., et al., Non-enzymatic roles of human RAD51 at stalled replication forks. Nat Commun, 2019. 10(1): p. 4410. [CrossRef]
- Rajendran, V. and M.V. Jain, In Vitro Tumorigenic Assay: Colony Forming Assay for Cancer Stem Cells. Methods Mol Biol, 2018. 1692: p. 89-95.
- Bianchi, J., et al., PrimPol bypasses UV photoproducts during eukaryotic chromosomal DNA replication. Mol Cell, 2013. 52(4): p. 566-73. [CrossRef]
- Garcia-Gomez, S., et al., PrimPol, an archaic primase/polymerase operating in human cells. Mol Cell, 2013. 52(4): p. 541-53. [CrossRef]
- Keen, B.A., et al., Molecular dissection of the domain architecture and catalytic activities of human PrimPol. Nucleic Acids Res, 2014. 42(9): p. 5830-45. [CrossRef]
- Mansilla, S.F., et al., Polymerase iota (Pol iota) prevents PrimPol-mediated nascent DNA synthesis and chromosome instability. Sci Adv, 2023. 9(15): p. eade7997.
- Sale, J.E., Translesion DNA synthesis and mutagenesis in eukaryotes. Cold Spring Harb Perspect Biol, 2013. 5(3): p. a012708. [CrossRef]
- Livneh, Z., O. Ziv, and S. Shachar, Multiple two-polymerase mechanisms in mammalian translesion DNA synthesis. Cell Cycle, 2010. 9(4): p. 729-35. [CrossRef]
- Prakash, S., R.E. Johnson, and L. Prakash, Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function. Annu Rev Biochem, 2005. 74: p. 317-53. [CrossRef]
- Makridakis, N.M. and J.K. Reichardt, Translesion DNA polymerases and cancer. Front Genet, 2012. 3: p. 174. [CrossRef]
- Pilzecker, B., et al., PrimPol prevents APOBEC/AID family mediated DNA mutagenesis. Nucleic Acids Res, 2016. 44(10): p. 4734-44. [CrossRef]
- Cross, S.E., et al., Nanomechanical analysis of cells from cancer patients. Nat Nanotechnol, 2007. 2(12): p. 780-3.
- Guck, J., et al., Optical deformability as an inherent cell marker for testing malignant transformation and metastatic competence. Biophys J, 2005. 88(5): p. 3689-98.
- Swaminathan, V., et al., Mechanical stiffness grades metastatic potential in patient tumor cells and in cancer cell lines. Cancer Res, 2011. 71(15): p. 5075-80.
- Kraning-Rush, C.M., J.P. Califano, and C.A. Reinhart-King, Cellular traction stresses increase with increasing metastatic potential. PLoS One, 2012. 7(2): p. e32572. [CrossRef]
- Merkher, Y. and D. Weihs, Proximity of Metastatic Cells Enhances Their Mechanobiological Invasiveness. Ann Biomed Eng, 2017. 45(6): p. 1399-1406. [CrossRef]
- Merkher, Y., et al., Rapid Cancer Diagnosis and Early Prognosis of Metastatic Risk Based on Mechanical Invasiveness of Sampled Cells. Ann Biomed Eng, 2020. 48(12): p. 2846-2858. [CrossRef]
- Sun, H., et al., Elevated DNA polymerase iota (Poli) is involved in the acquisition of aggressive phenotypes of human esophageal squamous cell cancer. Int J Clin Exp Pathol, 2015. 8(4): p. 3591-601.
- Li, L., et al., siRNA of DNA polymerase iota inhibits the migration and invasion in the lung cancer cell A549. Acta Biochim Biophys Sin (Shanghai), 2018. 50(9): p. 929-933. [CrossRef]
- Tang, H.M., et al., Molecular signature of anastasis for reversal of apoptosis. F1000Res, 2017. 6: p. 43.
- Chakraborty, S., et al., Integration of EMT and cellular survival instincts in reprogramming of programmed cell death to anastasis. Cancer Metastasis Rev, 2020. 39(2): p. 553-566. [CrossRef]
- Blackford, A.N. and S.P. Jackson, ATM, ATR, and DNA-PK: The Trinity at the Heart of the DNA Damage Response. Mol Cell, 2017. 66(6): p. 801-817. [CrossRef]
- Bakkenist, C.J. and M.B. Kastan, DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature, 2003. 421(6922): p. 499-506. [CrossRef]
- Ward, I.M., K. Minn, and J. Chen, UV-induced ataxia-telangiectasia-mutated and Rad3-related (ATR) activation requires replication stress. J Biol Chem, 2004. 279(11): p. 9677-80. [CrossRef]
- Wan, L., et al., hPrimpol1/CCDC111 is a human DNA primase-polymerase required for the maintenance of genome integrity. EMBO Rep, 2013. 14(12): p. 1104-12.
- Wold, M.S., Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem, 1997. 66: p. 61-92. [CrossRef]
- Li, X., et al., Polymerase iota (POLI) confers radioresistance of esophageal squamous cell carcinoma by regulating RAD51 stability and facilitating homologous recombination. Cell Death Discov, 2023. 9(1): p. 291. [CrossRef]
- Zellweger, R., et al., Rad51-mediated replication fork reversal is a global response to genotoxic treatments in human cells. J Cell Biol, 2015. 208(5): p. 563-79.
- Vallerga, M.B., et al., Rad51 recombinase prevents Mre11 nuclease-dependent degradation and excessive PrimPol-mediated elongation of nascent DNA after UV irradiation. Proc Natl Acad Sci U S A, 2015. 112(48): p. E6624-33.
- Walcher, L., et al., Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front Immunol, 2020. 11: p. 1280. [CrossRef]
- Yan, X., et al., A CD133-related gene expression signature identifies an aggressive glioblastoma subtype with excessive mutations. Proc Natl Acad Sci U S A, 2011. 108(4): p. 1591-6. [CrossRef]
Figure 1.
The capacity of cell motility and invasiveness of A549 wild-type, PRIMPOL-/- and POLI -/- cells toward a chemo-attractant gradient assessed by Boyden chambers (with 8 µm pores) assay. ****p < 0.0001. Data are means ± SEM.
Figure 1.
The capacity of cell motility and invasiveness of A549 wild-type, PRIMPOL-/- and POLI -/- cells toward a chemo-attractant gradient assessed by Boyden chambers (with 8 µm pores) assay. ****p < 0.0001. Data are means ± SEM.
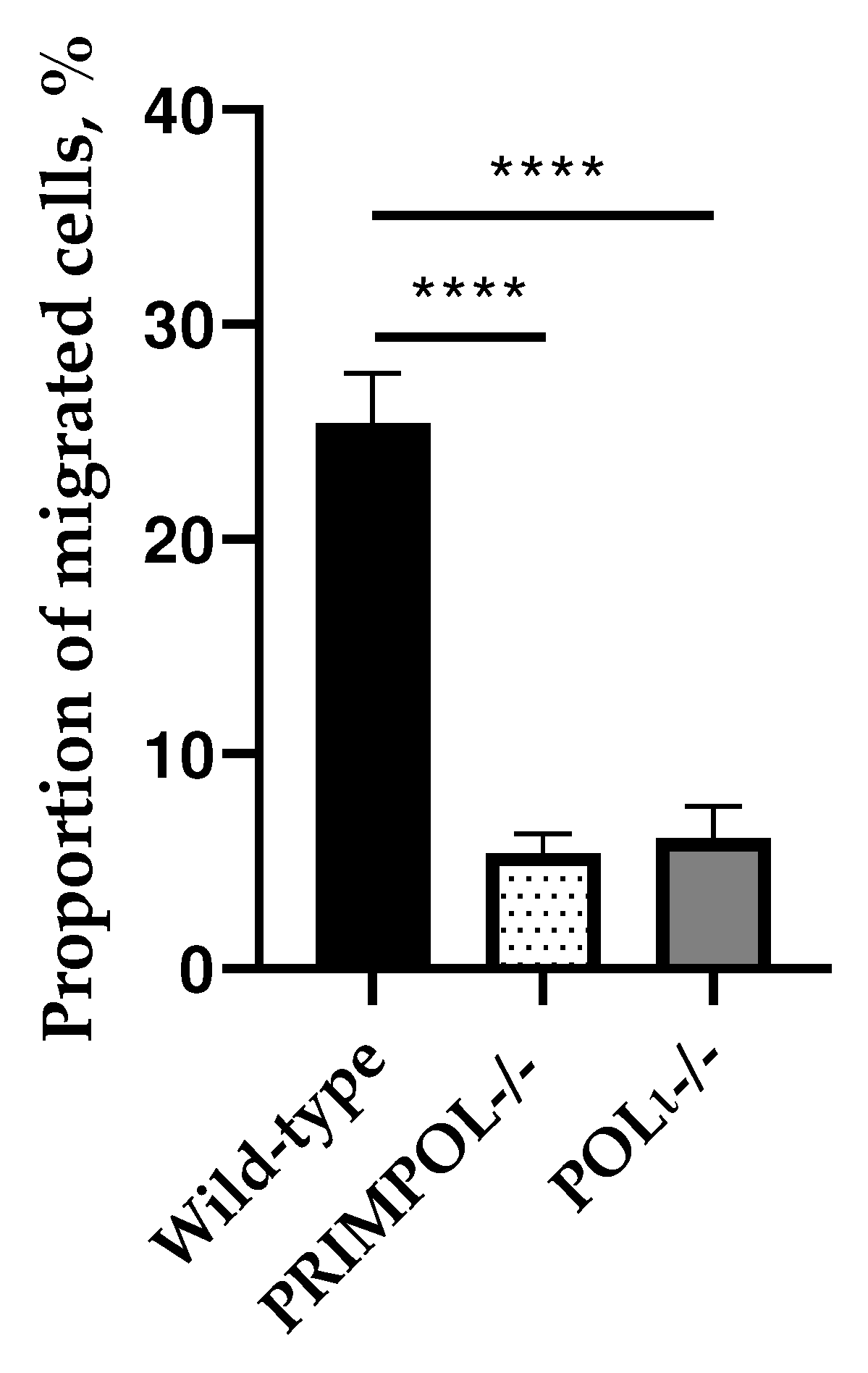
Figure 2.
Cell cycle analysis of A549 wild-type, PRIMPOL-/- and POLI -/-(a) 48 h and (b) 72 h without irradiation, and (c) 48 h and (d) 72 h after 6 Gy X-rays exposure. *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001. A total of 50,000 events were acquired for each sample. Data are means ± SEM of more than three independent experiments.
Figure 2.
Cell cycle analysis of A549 wild-type, PRIMPOL-/- and POLI -/-(a) 48 h and (b) 72 h without irradiation, and (c) 48 h and (d) 72 h after 6 Gy X-rays exposure. *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001. A total of 50,000 events were acquired for each sample. Data are means ± SEM of more than three independent experiments.
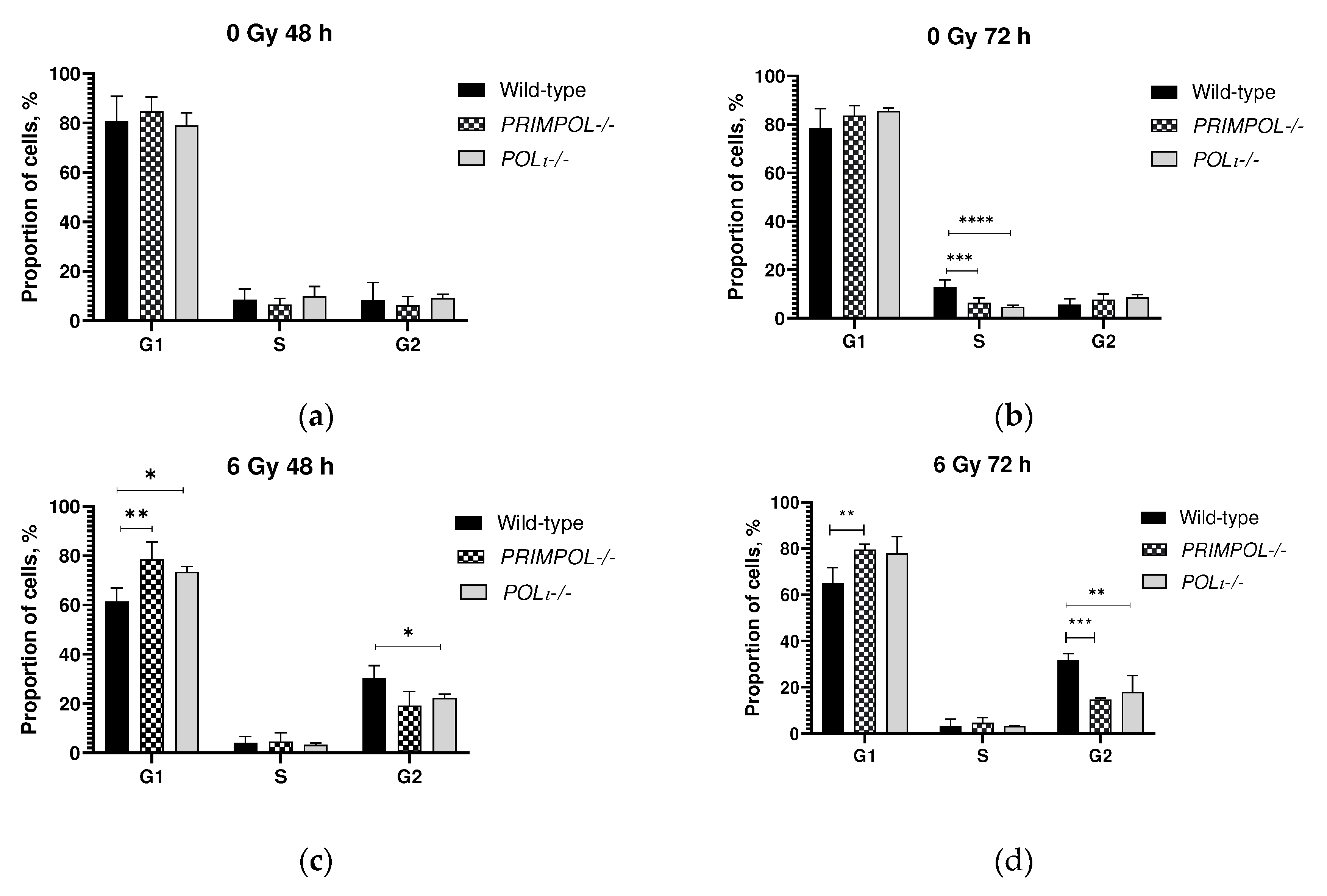
Figure 3.
Apoptosis analysis of non-irradiated and irradiated (8 Gy exposure) (a) wild-type; (b) PRIMPOL -/- and (c) POLI −/− A549 cells at the initial time of the exposure and 48 h post-irradiation. (d) Apoptosis analysis of non-irradiated cells at 7 days of cultivation, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Data are means ± SEM.
Figure 3.
Apoptosis analysis of non-irradiated and irradiated (8 Gy exposure) (a) wild-type; (b) PRIMPOL -/- and (c) POLI −/− A549 cells at the initial time of the exposure and 48 h post-irradiation. (d) Apoptosis analysis of non-irradiated cells at 7 days of cultivation, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Data are means ± SEM.
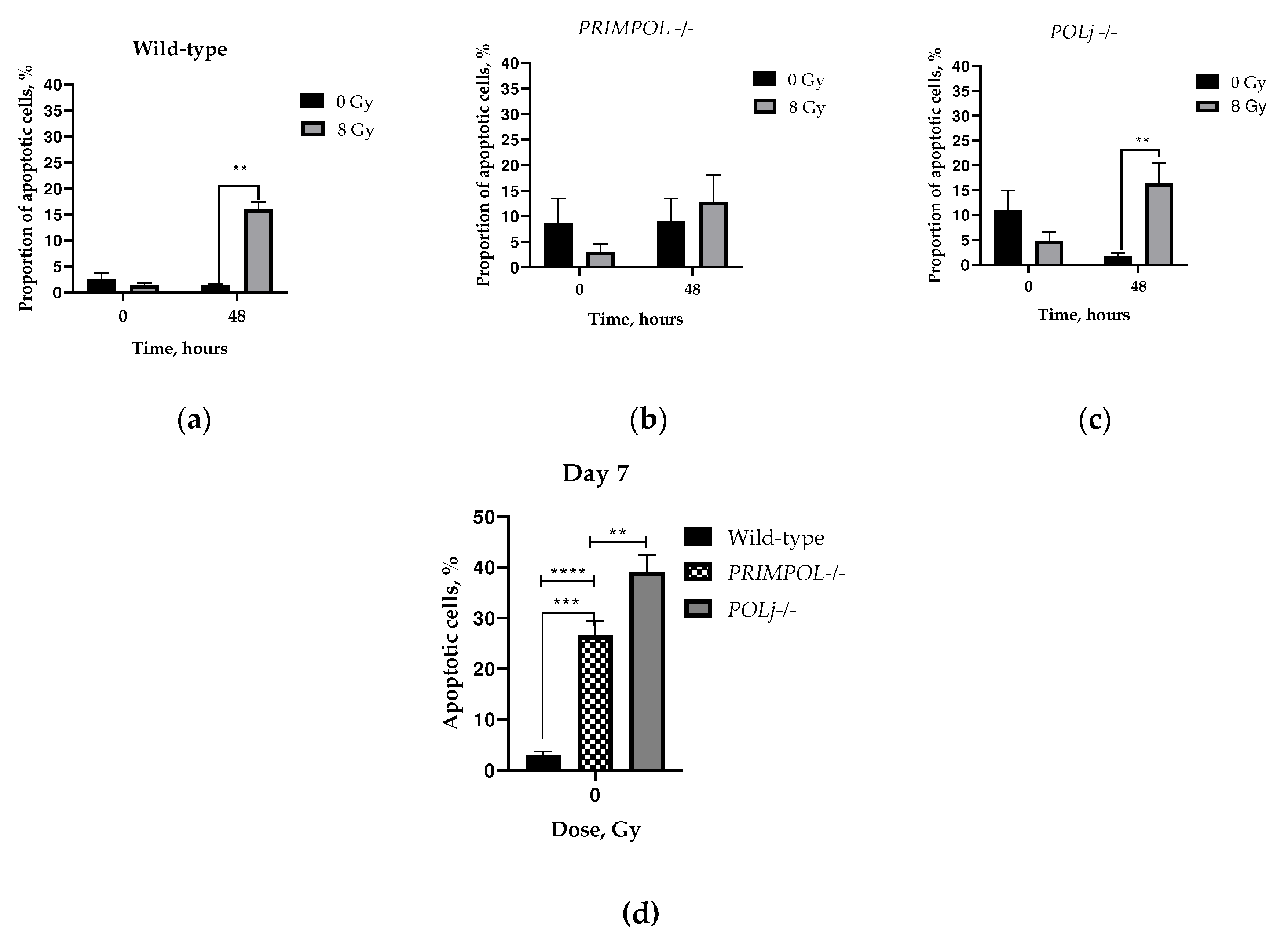
Figure 4.
Radiosensitivity of A549 wild-type, PRIMPOL-/- and POLI -/- cells assessed by colony formation assay in anchorage-dependent fashion. ** p < 0.01. Data are means ± SEM.
Figure 4.
Radiosensitivity of A549 wild-type, PRIMPOL-/- and POLI -/- cells assessed by colony formation assay in anchorage-dependent fashion. ** p < 0.01. Data are means ± SEM.
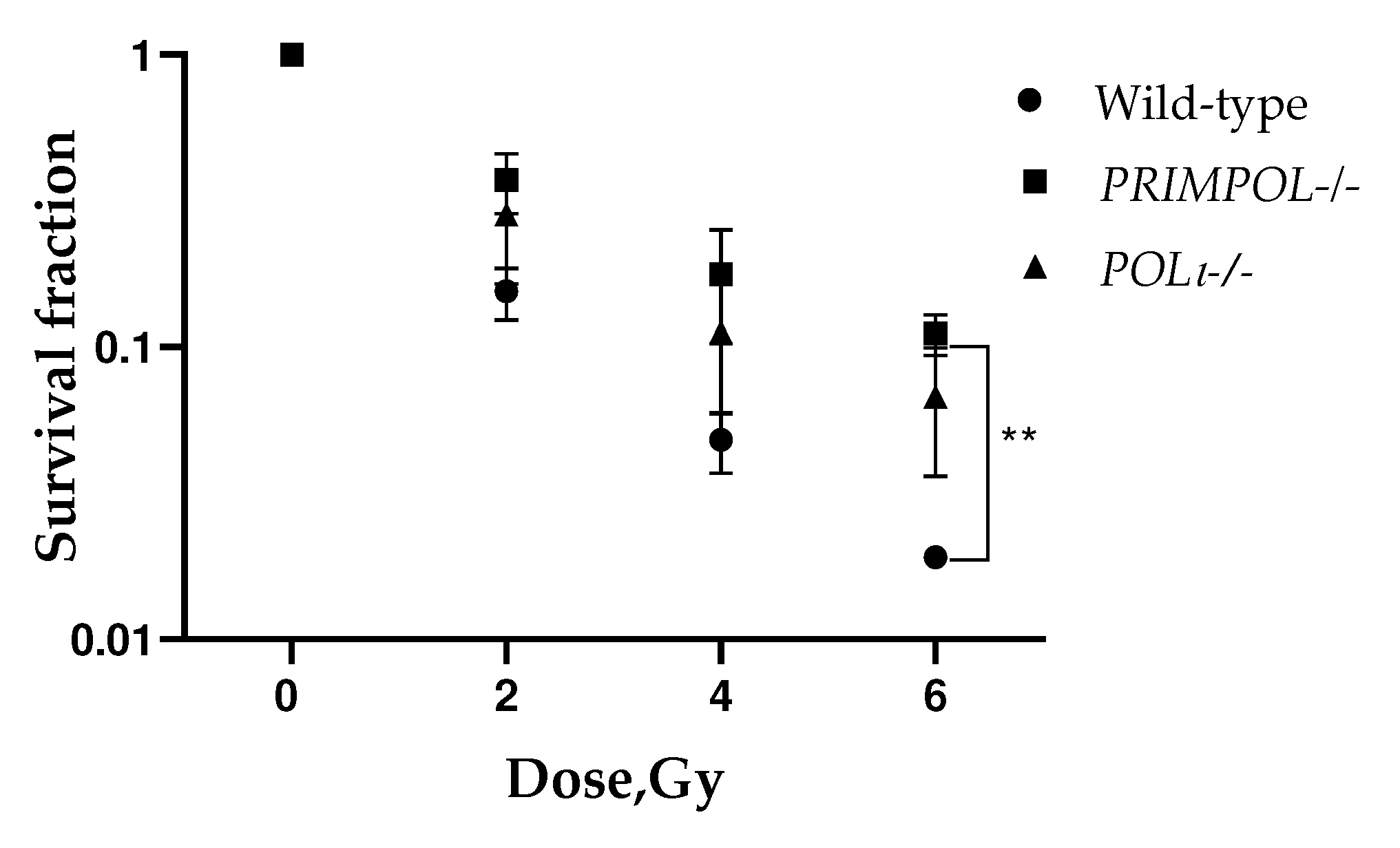
Figure 5.
Kinetics of γH2AX (a), phosphorylated ataxia telangiectasia mutated (pATM) (b) and Rad51 (c) foci changes in A549 wild-type, PRIMPOL-/- and POLI -/- cells. *p < 0.05, **p < 0.01, ****p < 0.0001. Data are means ± SEM.
Figure 5.
Kinetics of γH2AX (a), phosphorylated ataxia telangiectasia mutated (pATM) (b) and Rad51 (c) foci changes in A549 wild-type, PRIMPOL-/- and POLI -/- cells. *p < 0.05, **p < 0.01, ****p < 0.0001. Data are means ± SEM.
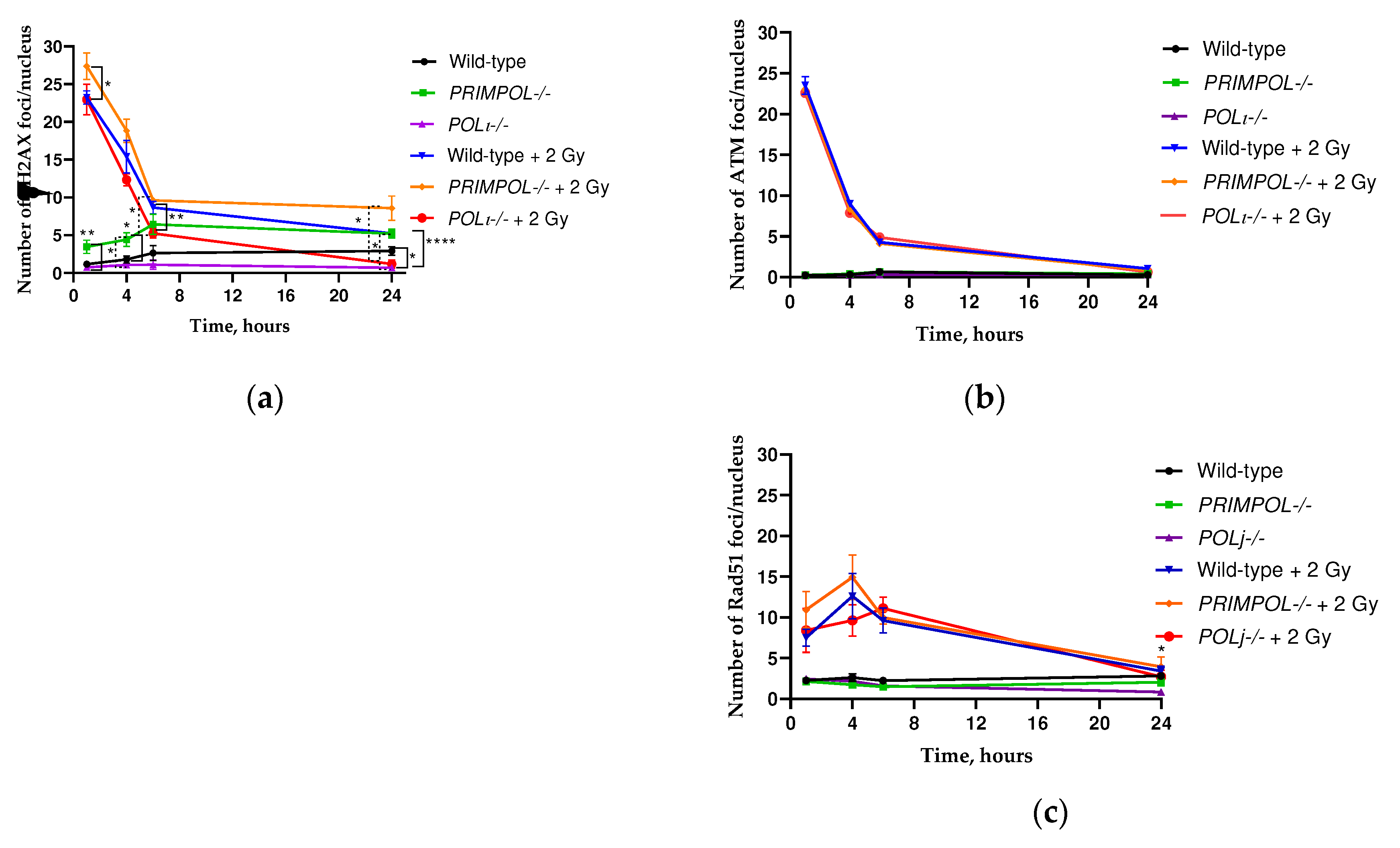
Figure 6.
The proportion of CD44+, CD133+ and CD133+/ CD44+ populations in wild-type, PRIMPOL-/- and POLI -/- cells at (a,c) 48 h and (b,d) 72 h of cultivation after (a, b) 0 Gy and (c, d) 6 Gy X-ray exposure. *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001. Data are means ± SEM.
Figure 6.
The proportion of CD44+, CD133+ and CD133+/ CD44+ populations in wild-type, PRIMPOL-/- and POLI -/- cells at (a,c) 48 h and (b,d) 72 h of cultivation after (a, b) 0 Gy and (c, d) 6 Gy X-ray exposure. *p < 0.05, **p < 0.01, ***p < 0.001, **** p < 0.0001. Data are means ± SEM.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



