Submitted:
17 November 2023
Posted:
20 November 2023
You are already at the latest version
Abstract
The spatial and temporal variability in dietary preferences of juvenile three-spined stickleback Gasterosteus aculeatus, was studied across typical coastal habitats of Keret Archipelago, Kandalaksha Bay, the White Sea. Sampling of fish using beach seine was implemented in late July-early August and in late August, and zooplankton samples were collected to make a quantitative evaluation of abundance of potential prey items. Similarity Percentage (SIMPER) analysis implemented over the data obtained from stomach content analysis revealed five planktonic taxa mostly contributing to the diet of juvenile sticklebacks, of which copepod Temora longicornis was the most important prey item at marine sites, while Acartia longiremis prevailed in diet of fishes in marine lagoon. Calculation of selectivity index demonstrated that some taxa (e.g., A. longiremis) were always avoided by juvenile fishes, others (e.g. Microsetella norvegica) were firstly avoided and then selected in late August. Calculation of selectivity index was implemented per sample and per individual fish indicated that taxa like T. longicornis and were selected during the whole period of study, fishes demonstrated pronounced individual variation in prey selectivity. In general, data on selectivity indicated that despite juvenile sticklebacks feed on mostly available prey items, they can demonstrate taxonomic and size-specific prey selectivity.
Keywords:
food selectivity
; stickleback
; three-spined stickleback
; Gasterosteus aculeatus
; the White Sea
; zooplankton
; fish nursery grounds
; intertidal communities
1. Introduction
Survival of juvenile life stages are critical for the successful recruitment of fish populations [1,2,3]. At the spawning and nursery grounds, survival of juveniles can strongly be a function of habitat type and trophic relationships with adult fishes of the same and other species especially predation and competition [4]. Abundance and diet of adult and juvenile fishes at a particular spawning ground can be strongly dependent of the type of the spawning ground. Wave explosion, water turbidity, substrate, availability of shelter, predator pressure and availability of food resources are all factors shaping the quality of fish spawning grounds [5,6,7,8] .
The indented coastline of Kandalaksha Bay, the White Sea, is an important spawning area for many common species such as herring Clupea pallassii [9,10], smelt Osmerus dentex [11], and two species of sticklebacks: nine-spined stickleback Pungitius pungitius and three-spined stickleback Gasterosteus aculeatus [12]. The latter two species belong to an ecological group of ‘small fishes’ being important elements of coastal and pelagic food webs [13,14,15,16]. Studies of dietary preferences of sticklebacks, being mostly demersal fishes during the spawning season and pelagic during the rest of the year, are important for better understanding of benthic-pelagic coupling in temperate and subarctic marine ecosystems. Marine populations of the three-spined stickleback are migratory - fishes spend most of the summer in the inshore areas, and pelagic species during the other months of the year. Adults lay eggs in mid-June, larvae hatch within one week and then juveniles grow in the inshore and nearshore areas until late summer and early autumn [17]. Recent studies, based oncombinations of conventional stomach content analysis, carbon and nitrogen stable isotope analysis and environmental DNA analysis, demonstrate that the diet of adult sticklebacks at spawning grounds include benthic and planktonic taxa with the pronounced shifts within the spawning season [18,19,20].
Previous study of the diet of juvenile White Sea G. aculeatus at nursery grounds has focused on the most suitable habitat for the species – dense beds of eelgrass, Zostera marina [21]. However, as sticklebacks use several types of intertidal habitats as nursery grounds, including Z.marina, rockweeds Fucus spp. and Ascophillum nodosum, semi-isolated marine lagoons and even rocky shores [17,22], it is important to understand how each type of spawning and nursery ground affects the trophic behaviour of growing juveniles. By comparing the dietary preferences of juvenile fishes with the structure of nearshore zooplankton communities, this paper focuses on comparative analysis of feeding patterns of three-spined stickleback juveniles across various typical nursery grounds for the species in early and late phases of active juvenile growth in nearshore waters during their first summer.
2. Materials and Methods
Samples were collected in August 2014 over two periods of G. aculeatus spawning season, particularly 1.08 - 2.08 (hereafter – date 1, or early August), and 18.08-20.08 (date 2, or late August) at six sites differing by dominant intertidal habitat (Table 1, Figure 1). Additional information on sampling sites can be found in Supplementary Table 1. Sampling was implemented using a beach seine with length and high of wings 7.5 and 1.5 m, respectively, a mesh size was 5 mm from knot to knot in the wings and 1 mm in the codend. The seine was pulled from the shoreline with a rowing boat to approx. 30-metre distance from the shore and then pulled towards the shore to encircle the 120 square meter area.
In order to calculate the abundance of sticklebacks per square meter comparable with other studies of sticklebacks in the White Sea using the same sampling equipment [18,22], the catch efficiency coefficient equal to 0.6 (the ratio of fish caught related to the total number of fish in the catch area) was applied. In a few cases, sticklebacks were caught using hand nets. From the seine net, two samples were collected. One consisted of 50 ml of fish to evaluate the abundance and size structure of juvenile sticklebacks. The other was collected for evaluation of juveniles’ diet. The number of fish used for diet analysis is presented in Table 1. Exceptions were the samples from Keret and Krugliash at date 2, when the abundance of fish was low and all juveniles from the seine net were collected.
Simultaneously to juvenile stickleback sampling, we also collected zooplankton samples to analyze the density of planktonic food organisms. One replicate per site was collected with a plankton net (mesh size 93 µm) by filtering 100 liters of surface water.
The diet of juvenile sticklebacks was evaluated using standard stomach content analysis [18], and subsequent calculation of standard indices calculated either per sample (usually 30 fishes, see Table 1) or per individual fish.
The following indices were used - Feeding Intensity (FI), frequency of occurrence (%Fi), percent biomass (%Ii), percent number (%Qi), number of taxa from stomach content significantly contributing to the diet (D-index). Partial Feeding intensity Index (PFI), expressing the fullness contribution of each prey category, is then assigned summing up to the total stomach fullness [23,24]. The role of individual objects in diet (i.e. prey selectivity) was also calculated using the following equation (1) [25,26]:
where E is the selectivity index, ri is the proportion of prey species i in fish stomachs, and pi is the proportion of prey species i in the sea.
As a result, E index represents the relative composition of the prey items in the stomach contents and in the environment. The selectivity index was calculated based on either food spectrum of individual fishes within sample or averaged per sample. The selectivity index generally varies between −1 and 1, with negative values indicating avoidance, positive values indicating active selection. Following several recent studies on food selectivity in aquatic species [27], E index values between −0.25 and +0.25 were considered as indicating non-selective feeding, E ≥ +0.25 indicating preference of a specific prey items, and E ≤ −0.25 as avoidance of particular prey items.
Statistical analyses were performed using standard spreadsheet software (MS Excel 2013), STATISTICA v7.0, PAST v.411 [28], and R version 4.2.3. Separate factorial ANOVA analyses were implemented to reveal differences between sampling dates and sites using various parameters e.g. TL, FI, D-index, contribution of prey items to food spectrum. The assumptions of ANOVA analyses were evaluated with Fisher’s post hoc comparisons. Relationships between FI and abundance of juveniles and E index of each prey item and TL of juveniles were evaluated using linear regression analysis. A two-way ANOSIM test was applied to explore the differences between diet of juvenile sticklebacks by date and site (Clarke and Warwick 1994). ANOSIM generates a test statistic, R, and the magnitude of R is indicative of the degree of separation between groups, with a score of 1 indicating complete separation and 0 indicating no separation (Clarke and Green 1988; Clarke 1993). Similarity Percentage Analysis (SIMPER) was used with Bray-Curtis dissimilarity metric to examine which prey items mostly contributed to the diet of juvenile sticklebacks (Clarke 1993).
3. Results
3.1. Size Structure and Abundance of G. aculeatus Juveniles
Two-factor ANOVA results revealed some significant differences in juvenile length across the sites studied and the two periods of observation (F5, 1186=10.68, p<0.0001). Mean total length (TL) of juveniles increased by 15-25% during August at two eelgrass Zostera marina bed sites (Seldianaya: from 16.5 ± 0.2 to 20.7 ± 0.3; Letnaya: from 16.4 ±0.4 to 18.9 ± 0.1 mm). In Koliushkovaya lagoon, where dense eelgrass beds are also present, mean length increased from 16.7 ± 0.4 to 19.1 ± 0.2 mm (Figure 2). Increase in TL was also observed at rockweed site Podpakhta from 15.4 ± 0.3 to 16. 3± 0.3 the mm, yet significance level was lower than for Zostera and lagoon sites. At the other two sites TL did not change significantly. At these two sites, represented by rockweeds (Krugliash) and rocky shore (Keret) intertidal, density of juveniles was lower in late August compared to the eelgrass bed sites. Data on density of juveniles at all sites during two periods of study is shown in Figure 2.
3.2. Feeding Intensity of G. aculeatus
Feeding intensity (FI) significantly varied between early and late August (ANOVA, F 5,311=8.88, p<0.01), and between sites within each of the two periods of sampling (early August: F 5,174=5.74, p<0.001; late August: F 5,137=5.91, p<0.001). The proportion of empty stomachs in juvenile sticklebacks never exceeded 20%, and did not vary between data and station, with a 9% mean for all sites in early August and 6% in late August.
FI varied between early and late August at all sites except Podpakhta and Koliushkovaya (Figure 3). At Seldianaya, FI decreased by 2.7-times from 447.5 ± 91.6‱ in early August (maximum value across all six sites) to 165.9 ± 30.8‱. At the other three sites FI increased in late August, and at Krugliash (rockweeds site) mean FI increased by 4.7 times from 85.3 ± 22.6‱ to 401.8 ± 101.6‱. Based on regression analysis, FI of individual fishes was significantly inversely proportional to abundance of juveniles (F 1,321 = 9.429, R2 = 0.029 p = 0.0023).
3.3. Stomach Content Analysis
In total, 31 prey items were found in stomachs of juvenile sticklebacks. The number of taxa found in stomachs varied between sites and dates from 7 to 17, with an overall mean value of 13. The mean number of taxa per individual varied between 2.4 and 5.1 (mean = 3.9). Maximum taxonomic diversity in stomach contents was observed in fishes from Seldianaya, and a minimum in Koliushkovaya. The mean number of taxa mostly contributing to the diet (evaluated using D-index) per fish was 2.7 (max 3.5, min 1.9) based on abundance and 2.4 taxa (max 3.2, min 1.6) using biomass of food items. The two-factor ANOVA demonstrated that D-index and number of taxa in individual stomachs of stickleback juveniles varied between dates of sampling and sites (F5, 282=5.58, p<0.0001). The list of taxa mostly contributing to the diet varied between sites (Figure 4). The D-index did not show significant correlation with FI (R = -0.04, p>0.05 using abundance data, R = 0.07, p>0.05 using biomass data)
Similarity Percentage (SIMPER) analysis revealed five planktonic taxa mostly contributing to the diet of juvenile sticklebacks. Of these, 22% of the heterogeneity among samples was explained by the abundance of Temora longicornis (Müller O.F., 1785), 9% explained by Acartia longiremis (Lilljeborg, 1853), 13% by Podon leuckarti (G.O. Sars, 1862), 13% by Copepoda juv. (nauplii and copepoditii Copepoda), 6% by Microsetella norvegica (Boeck, 1865), and 13% by benthic Orthocladiinae larvae. Total heterogeneity among samples was 88%, and changes in biomass of above-mentioned species in stomach contents explained 75% of the spatial and temporal differences between samples.
The most important planktonic component of the diet of juveniles was T. longicornis, observed in stomach contents of juvenile sticklebacks at marine sites contributing on average 32% to the total biomass of stomach contents (varying between 17-60%). It contributed to the diet significantly differently between sites and sampling periods (ANOVA, F5-311=21.32, p<0.0001). The only site where T. longicornis did not contributed significantly to stomach contents was Koliushkovaya lagoon, where A. longiremis prevailed in diet of juvenile sticklebacks.
In early August, the highest proportion of this species in stomachs was found in Seldianaya (2-way ANOVA post-hoc p<0.05). In late August, the proportion of T. longicornis at Seldianaya was similar, and an increased proportion of this species in stomachs was observed at Keret (2-way ANOVA post hoc р<0.001) and Podpakhta (2-way ANOVA post hoc p<0.01).
The most important component of benthic feeding was Orthocladiinae larvae, contributing on average 17% to the stomach contents, reaching a maximum of 38%. No variation of consumption of this species by stickleback juveniles was observed between sites and date if a complete dataset is taken into analysis (2-way ANOVA, F5-311=3.46, all post-hoc p>0.05). However, among the early August samples, a higher proportion Orthocladiinae was observed in Seldianaya (ANOVA post hoc p<0.01).
In general, planktonic taxa prevailed in stomach contents both by density (up to 98%) and biomass (up to 95%) (see Figure 4 for details). Benthic species (nine taxa) significantly contributed to biomass (from 5% to 46%) only at a few sites and for a short period of time. Two-factor ANOVA demonstrated that proportion of planktonic components in the diet of fishes did not vary between two periods of observations (F5, 282=2.5, p=0.43), but varied between sites (F5, 282=2.5, p<0.05). The only site where benthic taxa significantly contributed to the diet of juvenile sticklebacks by biomass at both periods of observation was Krugliash (36% and 46% respectively). The length of the juveniles (TL) did not have a significant effect on the proportion of benthic versus planktonic taxa in stomach contents found using one-way ANOVA (F18, 275=1.53, p=0.07).
The 2-way ANOSIM analysis implemented using the Partial Fullness Index (PFI), revealed that both date and site of sampling significantly affect the diet of juvenile sticklebacks (Table 2). Similar analysis conducted on three other input data (%Ii, %Qi, presence-absence of taxa) also yielded highly significant (p<0.0001) R values for both date (ANOSIM R values between 0.51 and 0.52) and site (R values between 0.23 and 0.29) factors. The diet of sticklebacks varied between sites significantly with each of two sampling periods according to a one-way ANOSIM analysis (Table 2). Pairwise comparisons revealed that within the early August (Date 1) all sites differed significantly from each other. In late August (Date 2), two pairwise site comparisons did not reveal significant differences (Letnaya-Keret) and (Keret-Podpakhta); the remainder were significant (Table 2).
3.4. Food Resources: Zooplankton
In total, 25 zooplankton taxa were recorded during the study, varying between one and 15 taxa per site. Taxonomic diversity of planktonic communities at the marine sites was similar across the two periods of investigation. Among the most abundant taxa at marine sites were M. norvegica, T. longicornis, O.similis and P. leucartii. This group of taxa were observed at all sites except Koliushkovaya lagoon, yet the contribution of each taxon to the general abundance values were different across sites and the two periods of study (Figure 4). In Koliushkovaya lagoon only one zooplankton species was found - A. longiremis.
Diversity of zooplankton measured with D-index using abundance (N) and biomass (B) values varied between sites and dates. According to the 2-way ANOVA (factors ‘site’ and ‘date’), both abundance and biomass of zooplankton varied significantly between sites (ANOVA, F1-5 = 2.62, p<0.05), and biomass of zooplankton significantly varied between dates of sampling (ANOVA, F1-5 = 6.64, p<0.05). The highest abundance and biomass values of zooplankton were observed in the lagoon (ANOVA post hoc p<0.05). While no significant differences were found between zooplankton abundance at marine sites, eelgrass beds tend to show higher values in early August, while rocky shore and rockweed sites had higher zooplankton abundance in late August (Table 3)
3.5. Food Selectivity
Calculation of Ivlev’s selectivity index E based on food spectrum of individual fishes yielded the following results. In early August (Figure 5), negative values of E index lower than <-0.25 (i.e. avoidance of prey item) were observed for A. longiremis and O. similis at most sites indicating that these species were found in zooplankton communities but were absent in stomach contents of juvenile sticklebacks. The only exception for A. longiremis was marine lagoon Koliushkovaya where E values varied between zero and -0.25. In M. norvegica mean value lower than <-0.25 was observed only at eelgrass site Seldianaya; at other sites E varied between - 0.25 and + 0.25 for this species. Positive E values higher than 0.25 (prey selectivity) were observed for T. longicornis and P. leuckartii at rockweed site Podpakhta, and for T. longicornis at another rockweed site Krugliash. At lagoon site, Copepoda juv. were selected by sticklebacks against a background of high negative values of E observed at other sites.
In late August, sticklebacks still demonstrated avoidance of A. longiremis and O. similis at the most sites. Juvenile copepods (Copepoda juv.) were predated selectively at lagoon and rockweed site Krugliash. Selective predation of stickleback juveniles on M. norvegica was also observed at Krugliash as well as at Seldianaya. No clear selectivity of T. longicornis was observed in late August; P. leuckartii was avoided at Seldianaya and positively selected at rocky shore site Keret (Figure 5).
Using the E index based on values averaged per sample revealed that A. longiremis and O. similis were avoided by juvenile sticklebacks in early and late August. In A. longiremis values of E index varied between -0.14 and -1 and were -1 for O. similis at all sites and dates. Early August values of E for T. longicornis were positive at all sites reaching +1 and varied between - 0.2 and + 0.9 in late August. Similarly, E index was positive for P. leuckartii in early August, but a highly negative (-1) value was demonstrated for Seldianaya in late August in contrast with other sites (Figure 5). Linear regression analysis did not reveal relationships between TL of fish and E index of each prey item.
4. Discussion
4.1. August Planktonic Food Resources of Juvenile Sticklebacks
In general, the zooplankton of the studied sites was represented by a boreal warm-watermass related taxa assemblage typical of the coastal areas of the White Sea in summer [29]. Copepoda was the dominant group, reaching 90-100% of total biomass and density of the whole zooplankton community; with the addition of taxa belonging to Cladocera. These results correspond well to major studies of zooplankton in the region that describe Copepoda as the major contributing group to the White Sea zooplankton [30,31].
Absolute zooplankton biomass varied between 38.5 and 5040 mg/m3 with the general mean of 779 mg/m3. When Koliushkovaya lagoon is excluded from the analysis, due to exclusive dominance of A. longiremis, total biomass of zooplankton in ‘open’ marine sites was 281 mg/m3. These values correspond well to the within-decadal variation of zooplankton biomass in Kandalaksha Bay between 140 and 257 mg/m3 [32]. These studies also indicated that the zooplankton biomass is generally higher in coastal areas compared to the open sea [29].
Among the nursery grounds studied, the highest total biomass of zooplankton was observed at eelgrass (Zostera marina) beds. As dense eelgrass thickets significantly reduce the water currents, this community located on the edge of subtidal and intertidal zones form a unique habitat for both planktonic and benthic species [33,34]. Locally calm hydrodynamic conditions provide optimal habitat for zooplankton [35,36] and thus support the extensive food resources for juvenile sticklebacks for which eelgrass beds are also a most important nursery habitat.
4.2. Size Structure and Abundance of Juvenile Sticklebacks at Nursery Grounds
The studied G.aculeatus nursery grounds can be subdivided into three types on the basis of stickleback juvenile abundance: habitats with constant abundance throughout the juvenile nearshore growth season being either constantly high or low, and habitats where abundance of juveniles decreases from early to late August.
The exposed rocky shore – Keret – was the least optimal nursery ground, where the abundance of juvenile fishes was constantly low. On the contrary, eelgrass beds – Seldianaya, Letnaya and the marine lagoon – were the most optimal nursery grounds being also the most important spawning grounds for adult sticklebacks [22]. While rockweed sites – Krugliash and Podpakhta – demonstrated high abundance of juveniles in early August, these significantly decreased at the end of growing season. At two of three eelgrass bed sites studied (Seldianaya and marine lagoon Koliushkovaya), the abundance of juveniles was high during the whole growing season (Figure 2). The mean length of the juveniles at all three eelgrass sites also increased by the end of August indicating successful growth and survival of fishes during the summer. These results correspond well to the data from aquarium experiments that demonstrate that juvenile sticklebacks prefer eelgrass to rockweeds. Eelgrass beds are presumed to provide shelter for juvenile sticklebacks and other juvenile fishes from predators, thus the survival rate of juveniles in this nursery habitat is high [33,34,37,38,39].
4.3. Feeding Intensity in G. aculeatus Juveniles
Fishes with empty stomachs never exceeded 20%, indicating both suitable conditions for feeding and active feeding behavior at the sites studied. Previously, higher values of 60-80% of fishes with empty stomachs have been reported, these values linked to temperature conditions and the availability of food [40,41,42]. Our findings on FI variation among the sites studied and over the two periods of stickleback spawning season correspond well with data from Abdel-Malek, 1968 [42]. During that time, FI in juvenile sticklebacks varied between 50 and 456 ‱, very similar values to this study. Recent data on the Baltic Sea populations also demonstrated similar values [20]. Feeding intensity was not related directly to the type of nursery ground but was related to the abundance of juveniles. Fishes from samples with lower abundance were characterized by higher feeding intensity regardless of the period of sampling.
4.4. Stomach Content in Juvenile G. aculeatus
In total, 31 prey items were recorded in stomachs of juvenile sticklebacks, varying between seven and 17 at various nursery habitats. According to the studies conducted in 1960s [42] the number of taxa in stomach contents varied between five and 25, indicating the same level of diversity of prey objects in stickleback diet throughout a five-decade period between studies. In general, juvenile sticklebacks were found to be planktivorous, as planktonic taxa prevailed in stomach content by abundance and biomass. Among the diversity of prey items, nine benthic taxa were found in stomachs, rarely contributing up to 46% by biomass to the stomach content. Krugliash was the only site where benthic taxa were important for the stickleback diet during both early (36%) and late (46%) August.
Prevalence of planktonic diet in juvenile stickleback diet has previously been reported from the White Sea [21,37], the Baltic Sea [43] and freshwater lakes at British Isles [44]. In adult White Sea three-spined sticklebacks, diversity of prey items is also comparable [18], and similar values of diversity of prey items have been observed in nine-spined stickleback P. pungitius from lacustrine environments despite the fact that adult fishes were mostly considered in the study [45].
D-index and number of taxa in individual stomachs varied significantly between dates and sites and did not correlate to FI. Fish diet varied between early and late August and no relationship between the size of fishes (TL) and taxonomic richness of prey items in stomachs was observed. No clear differentiation between the taxonomic richness of prey items consumed by stickleback juveniles related to the particular type of marine nursery grounds was found. Nevertheless, within each sampling period, the diversity of prey items observed in stomachs was significantly different between most sites with minor exceptions indicating differences in the availability of prey items.
The highest number of effective taxa was observed in Letnaya and Podpakhta (4.6 ± 0.16 and 4.5 ± 0.16), contrasting with marine lagoon Koliushkovaya where the lowest number of effective taxa (2.52 ± 0.14) was found in fish diet. At the eelgrass site Seldianaya the diversity of prey items was high in early August, and on average in late August. In the stomachs of individual fishes, 2-5 prey items were observed, indicating that despite the broad taxonomic range of potential prey items found in stomachs at each site, these <5 prey items prevail in fish diet. According to our data, 70% of fish diet was formed by ‘effective’, and other 30% were randomly consumed. Across all the sites studied, two taxa were the most important contributors to the diet of juvenile sticklebacks, namely T. longicornis and Orthocladiinae, similar to the data from Seldianaya obtained earlier [21], with the only exception of the marine lagoon, where A.longiremis overwhelmingly prevailed in the zooplankton. In late August samples, the proportion of T. longicornis was higher, probably indicating that with the growth of individual fishes, their ability to catch this crustacean increases. Consumption of chironomid larvae (Orthocladiinae) by stickleback larvae probably indicates the patchy distribution of this prey item. Chironomids are common and an important element of fish diet in marine and freshwater ecosystems [4,44,45,46]. In White Sea juvenile sticklebacks, when Orthocladiinae are present in stomach contents, they represent the majority of the biomass of food consumed, while T. longicornis occurs evenly across the individual fishes within the sample. On average, three individuals of Orthicladiinae were observed per stomach of G. aculeatus, with a maximum of eight larvae. No differences in consumption rate of Orthocladiinae were observed between sites and dates of sample collection. Comparison of consumption of other effective taxa by stickleback juveniles revealed that consumption of P. leuckartii varied significantly among sites only, while consumption of T. longicornis, M. norvegica, Copepoda juv. and A. longiremis varied between both sampling dates and sites. Observed spatial and temporal variability in diet of juvenile sticklebacks correspond well to the previous findings [26,42,43,47].
4.5. Prey Selectivity by Juvenile Sticklebacks in the White Sea
Prey selectivity has previously been studied in both adult and juvenile three-spined sticklebacks, yet these studies were to a great extent implemented using aquarium experiments or in freshwater environments and more often were focused on adult fishes. Yet, these studies contributed significantly to an understanding of the role of local hydrology in trophic preferences [48], profitability and visual characteristics of prey items [49], and competition with other species within the same habitat [50,51], with a special focus on ecosystems where three-spined stickleback is a non-native species [52,53]. As the body size in sticklebacks is comparable to many macroinvertebrate predators (e.g., insect larvae), potential overlaps of their dietary preferences were also considered [54]. One of the most detailed descriptions of dietary shifts of sympatric species of sticklebacks, including G. aculeatus, P. pungitius, G. wheatlandi and Apeltes quadractus was implemented at tidal saltmarsh spawning grounds in Canada [55]. This study considered seasonal dynamics in selective feeding of sticklebacks from early summer when adults come to the spawning grounds in late August when both adults and juveniles were present in samples [55].
In our study, in early August, T. longicornis was selectively preferred by juvenile sticklebacks (E values up to 1) at all sites studied, based on E index averaged per sample. Considering individually calculated E index values, similar results or neutral selectivity were obtained. Selectivity for T. longicornis (mean body size 0.56 ± 0.003 mm) varied among sites in late August when both approaches of calculating E index were applied. While E index values for average fish varied from - 0.2 to + 0.9 at various sites, individual data yielded values between - 0.42 to + 0.34, both indicating neutral or even avoiding a pattern of selectivity for this species. Similar seasonal variation in selectivity by sticklebacks was observed for P. leuckartii. At most sites, this species was selected positively during early August. In late August, positive selectivity for this species was observed at Keret (reaching + 0.64 by individuals and + 0.8 by average fish). On the contrary, P. leuckartii was avoided by juveniles at Seldianaya in late August, according to both methods of calculating E index.
Two prey taxa (M. norvegica and Copepoda juv.), being the smallest organisms in stickleback diet considered, demonstrated opposed patterns of selectivity over the two periods of observation. Mean size of M. norvegica was 0.41 ± 0.001 mm), and Copepoda juv. were mainly represented by nauplii Copepoda (0.11 ± 0.001 mm). These taxa were among the prey items of mainly smaller juvenile sticklebacks. Both prey items were negatively or neutrally selected in early August (E from -0.75 to +0.06 for M. norvegica and from - 0.8 to - 0.16 for Copepoda juv.), and positively or neutrally selected in late August (E from + 0.23 to + 1 for M. norvegica and from - 0.19 to + 1 for Copepoda juv.). A plausible explanation for the change in dietary preferences of these fishes might be that this shift is linked to changes in abundance of prey items. In early August, both M. norvegica and Copepoda juv. each contributed about 15-20% to the total zooplankton biomass (Figure 4). Later, their abundance in the zooplankton decreased, but small sizes of juvenile sticklebacks were still present at the nursery grounds due to the extended spawning period (Figure 2). The observed decrease in abundance of M. norvegica and Copepoda juv. in the zooplankton might be related either to seasonal trends or be a consequence of selective feeding by juvenile sticklebacks.
Several dominant planktonic taxa were mostly avoided by sticklebacks. In both periods of observation, O. similis was avoided at all sites where this species was present in the zooplankton (see Figure 3 and Figure 4 for details). As the size of individual O. similis (0.4 ± 0.005 mm) is similar to M. norvegica, differential selectivity patterns for these two species can be linked to several factors. First, M. norvegica (as in many other Harpacticoida) is bright-coloured (red to orange). As the same part of the spectrum is recognizable by at least female sticklebacks during the mating season [56], this can potentially increase visibility of this species for juvenile sticklebacks. Second, these species demonstrate very different swimming behaviour. Despite spending most of the time motionless, cyclopoids belonging to genus Oithona (e.g. O. plumifera) are able to jump fast for up to 20 body lengths (speed 22.1 mm/s), and almost exclusively upwards [57]. This behaviour can be beneficial for O. similis to avoid attacks by sticklebacks. On the contrary, M. norvegica demonstrate very slow swimming velocity (0.6 mm/s) [58], and even T. longicornis,almost always selected by sticklebacks, has a speed of 5.9 mm/s in adults, and lower values during early life stages [59].
Another species selectively avoided (E index values mostly around -1) by juvenile sticklebacks was A. longiremis at nearly all sites and over both periods. In zooplankton communities this species reached about 60% of total biomass, and in the marine lagoon Koliushkovaya reached 100% of total biomass. Considering that FI of stickleback juveniles at this site was generally low (Figure 3), it is possible to assume that this is a case of forced feeding on the only available prey item despite high prey availability. The possible evidence for forced dietary preferences in the lagoon is that selectivity for the second prey item group available - Copepoda juv. – was highly positive despite remarkably lower abundance in the zooplankton. This probably indicates that when individual fish could find larvae of copepods (remarkably this group can also include early stages of A. longiremis to some extent), it preferred to feed on them, and if not, the only available food were adult A. longiremis.
While comparison of two methods of calculating E index mostly yielded similar results, three cases can be considered: (1) E index values calculated per individual and per average fish both simultaneously indicated either very high or very low positive selectivity, (2) each index indicated values within the ‘neutral’ selectivity zone between - 0.25 and + 0.25, (3) averaged fish index indicated positive selectivity while individual-based index indicated slightly negative or neutral selectivity. Individual variation of prey selectivity for P. leuckartii by G. aculeatus at various sites in early August is a good example of these three patterns (Figure 6). Thus, the conventional way of calculating E index, i.e., the values calculated per average fish at the nursery ground (i.e., local population), mostly indicated a general trend of prey selectivity by individual stickleback population at each nursery ground. On the contrary, individual E values provided more accurate estimates and indicated sites and dates where various fishes within the sample had different consumption of prey item by individuals within the local population.
5. Conclusions
This study provides several insights into the prey selectivity of juvenile three-spined sticklebacks Gasterosteus aculeatus at several types of nursery ground habitats during their early and late phases of active growth in inshore waters during their first summer in the White Sea.
Nursery grounds were classified into three categories based on the abundance of juvenile sticklebacks. Eelgrass beds were found to be the most optimal nursery grounds, with consistently high juvenile abundance and successful growth and survival. In contrast, exposed rocky shores were the least optimal, with consistently low juvenile abundance. Stomach content analysis indicated that the percentage of fish with empty stomachs was generally low, indicating suitable feeding conditions and active feeding behavior among juvenile sticklebacks. Feeding intensity was correlated with the abundance of juveniles, with higher feeding intensity observed in samples with lower abundance. Diet of juvenile sticklebacks was primarily planktonic. Benthic taxa played a significant role in the diet at specific sites, particularly in the rockweed habitat. The diversity of prey items varied between sites and sampling periods, with a few dominant prey items making up the majority of their diet. Juvenile sticklebacks exhibited prey selectivity, with preferences for certain prey items in early August. However, prey selectivity patterns varied among sites and time periods, indicating the influence of local factors and prey availability. Some prey items, such as A. longiremis, were selectively avoided, while others, like T. longicornis and P. leuckartii, were positively selected. Seasonal changes in prey availability appeared to influence the dietary preferences of juvenile sticklebacks. The study found that selectivity for specific prey items shifted between early and late August, potentially in response to changes in prey abundance.
Author Contributions
Conceptualization, methodology, writing—original draft preparation, A.S.G.-Y.; sampling, A.S.G.-Y., N.V.P., M.V.I. and T.S.I.; laboratory analysis and data analysis, A.S.G.-Y., N.V.P., E.A.G.-Y.; writing—review and editing, A.S.G.-Y., E.A.G.-Y., D.L.L. and M.V.I.; funding acquisition, M.V.I., A.S.G.-Y. and E.A.G.-Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation, (project No. RSF 22-24-00956).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Most of the raw data for the study are available in Supplementary Materials.
Acknowledgments
Authors would like to thank the staff of the Education and Research station “Belomorskaya” of Saint Petersburg State University and our colleagues at the Department of ichthyology and hydrobiology of Saint Petersburg State University for their support and collaboration. Also, we would like to thank Prof. James Scourse, Dr. Chris Lowe (University of Exeter) and two anonymous reviewers for their input towards increasing the reading quality of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Van De Wolfshaar, K.E.; Barbut, L.; Lacroix, G. From Spawning to First-Year Recruitment: The Fate of Juvenile Sole Growth and Survival under Future Climate Conditions in the North Sea. ICES J. Mar. Sci. 2022, 79, 495–505. [Google Scholar] [CrossRef]
- Doyle, M.J.; Strom, S.L.; Coyle, K.O.; Hermann, A.J.; Ladd, C.; Matarese, A.C.; Shotwell, S.K.; Hopcroft, R.R. Early Life History Phenology among Gulf of Alaska Fish Species: Strategies, Synchronies, and Sensitivities. Deep Sea Res. Part II: Top. Stud. Oceanogr. 2019, 165, 41–73. [Google Scholar] [CrossRef]
- Robert, D.; Shoji, J.; Sirois, P.; Takasuka, A.; Catalán, I.A.; Folkvord, A.; Ludsin, S.A.; Peck, M.A.; Sponaugle, S.; Ayón, P.M.; et al. Life in the Fast Lane: Revisiting the Fast Growth—High Survival Paradigm during the Early Life Stages of Fishes. Fish Fish. 2023, 24, 863–888. [Google Scholar] [CrossRef]
- Westrelin, S.; Balzani, P.; Haubrock, P.J.; Santoul, F. Interannual Variability in the Trophic Niche of Young-of-year Fish Belonging to Four Piscivorous Species Coexisting in a Natural Lake. Freshw. Biol. 2023, 68, 487–501. [Google Scholar] [CrossRef]
- Wells, R.A.; McNeil, W.J. Effect of Quality of the Spawning Bed on Growth and Development of Pink Salmon Embryos and Alevins; US Department of the Interior, Bureau of Commercial Fisheries, 1970;
- Van Deurs, M.; Persson, A.; Nilsson, P.A.; Jørgensen, C. Fish Resist Temptation from Junk Food: State-dependent Diet Choice in Reproductive Atlantic Cod Gadus Morhua Facing Seasonal Fluxes of Lipid-rich Prey. Oikos 2022, 2022, oik–08739. [Google Scholar] [CrossRef]
- Byström, P.; Persson, L.; Wahlström, E. COMPETING PREDATORS AND PREY: JUVENILE BOTTLENECKS IN WHOLE-LAKE EXPERIMENTS. Ecology 1998, 79, 2153–2167. [Google Scholar] [CrossRef]
- Okun, N.; Mehner, T. Distribution and Feeding of Juvenile Fish on Invertebrates in Littoral Reed (Phragmites) Stands. Ecol. Freshw. Fish 2005, 14, 139–149. [Google Scholar] [CrossRef]
- Lajus, D.L.; Alekseeva, Y.I.; Lajus, J.A. Herring Fisheries in the White Sea in the 18th–Beginning of the 20th Centuries: Spatial and Temporal Patterns and Factors Affecting the Catch Fluctuations. Fish. Res. 2007, 87, 255–259. [Google Scholar] [CrossRef]
- Evseenko, S.A.; Mishin, A.V. On the Distribution of Larvae and Localization of Spawning Stocks of White Sea Herring Clupea Pallasii Marisalbi. J. Ichthyol. 2011, 51, 776–787. [Google Scholar] [CrossRef]
- Semenova, A.V.; Stroganov, A.N.; Ponomareva, E.V.; Afanas’ev, K.I.; Vilkina, O.V. Large-Scale Genetic Structure and Diversity of Arctic Rainbow Smelt Osmerus Dentex Steindachner et Kner, 1870 throughout Its Distributional Range Based on Microsatellites. Polar Biol 2021, 44, 927–940. [Google Scholar] [CrossRef]
- Lajus, D.; Ivanova, T.; Rybkina, E.; Lajus, J.; Ivanov, M. Multidecadal Fluctuations of Threespine Stickleback in the White Sea and Their Correlation with Temperature. ICES J. Mar. Sci. 2021, 78, 653–665. [Google Scholar] [CrossRef]
- Cusa, M.; Berge, J.; Varpe, Ø. Seasonal Shifts in Feeding Patterns: Individual and Population Realized Specialization in a High Arctic Fish. Ecol. Evol. 2019, 9, 11112–11121. [Google Scholar] [CrossRef] [PubMed]
- Hure, M.; Mustać, B. Feeding Ecology of Sardina Pilchardus Considering Co-Occurring Small Pelagic Fish in the Eastern Adriatic Sea. Mar. Biodivers. 2020, 50, 40. [Google Scholar] [CrossRef]
- Kingsford, M.J. Drift Algae and Small Fish in Coastal Waters of Northeastern New Zealand. Mar. Ecol. Prog. Ser. 1992, 41–55. [Google Scholar] [CrossRef]
- Hoch, J.M.; Cabanelas Bermudez, A.; Coury, O.S.; Donahou, A.S.; Jeffers, C.N.; LaMartina, M.; Ramsaran, D.; Spadafore, S. The Influence of Personality on Small Fish Migration and Dispersal in the Everglades. Wetlands 2019, 39, 991–1002. [Google Scholar] [CrossRef]
- Lajus, D.L.; Golovin, P.V.; Zelenskaia, A.E.; Demchuk, A.S.; Dorgham, A.S.; Ivanov, M.V.; Ivanova, T.S.; Murzina, S.A.; Polyakova, N.V.; Rybkina, E.V. Threespine Stickleback of the White Sea: Population Characteristics and Role in the Ecosystem. Contemp. Probl. Ecol. 2020, 13, 132–145. [Google Scholar] [CrossRef]
- Genelt-Yanovskaya, A.S.; Polyakova, N.V.; Ivanov, M.V.; Nadtochii, E.V.; Ivanova, T.S.; Genelt-Yanovskiy, E.A.; Tiunov, A.V.; Lajus, D.L. Tracing the Food Web of Changing Arctic Ocean: Trophic Status of Highly Abundant Fish, Gasterosteus Aculeatus (L.), in the White Sea Recovered Using Stomach Content and Stable Isotope Analyses. Diversity 2022, 14, 955. [Google Scholar] [CrossRef]
- Jakubavičiūtė, E.; Casini, M.; Ložys, L.; Olsson, J. Seasonal Dynamics in the Diet of Pelagic Fish Species in the Southwest Baltic Proper. ICES J. Mar. Sci. 2016, 74, 750–758. [Google Scholar] [CrossRef]
- Demchuk, A.S.; Uspenskiy, A.A.; Golubkov, S.M. Abundance and Feeding of Fish in the Coastal Zone of the Neva Estuary, Eastern Gulf of Finland. Boreal Environ. Res. 2021, 26, 1. [Google Scholar]
- Demchuk, A.; Ivanov, M.; Ivanova, T.; Polyakova, N.; Mas-Martí, E.; Lajus, D. Feeding Patterns in Seagrass Beds of Three-Spined Stickleback Gasterosteus Aculeatus Juveniles at Different Growth Stages. J. Mar. Biol. Assoc. United Kingd. 2015, 95, 1635–1643. [Google Scholar] [CrossRef]
- Ivanova, T.S.; Ivanov, M.V.; Golovin, P.V.; Polyakova, N.V.; Lajus, D.L. The White Sea Threespine Stickleback Population: Spawning Habitats, Mortality, and Abundance. Evol. Ecol. Res. 2016, 17, 301–315. [Google Scholar]
- Amundsen, P.; Sánchez-Hernández, J. Feeding Studies Take Guts – Critical Review and Recommendations of Methods for Stomach Contents Analysis in Fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Hyslop, E.J. Stomach Contents Analysis—a Review of Methods and Their Application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Ivlev, V.S.
- Wootton, R.J. A Functional Biology of Sticklebacks; Calow, P., Ed.; 1st ed.; Croom Helm, 1984;
- Zeng, H.; Tang, F.; Li, Z.; Lu, W.; Zheng, Y. Feeding Selectivity and Diet Shift of Protosalanx Chinensis during Spring in Lake Dalong, Northeastern China. Water 2023, 15, 1847. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A. Past: Paleontological Statistics Software Package for Educaton and Data Anlysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Кoсoбoкoва, К.Н.; Перцoва, Н.М. Зooпланктoн Белoгo Мoря: Структура, Динамика И Экoлoгия Сooбществ. Система Белoгo Мoря. Вoдная Тoлща И Взаимoдействующие С Ней Атмoсфера, Криoсфера, Речнoй Стoк И Биoсфера 2012, 640–675.
- Бек, Т.А. Прибрежная Зoна в Экoсистеме Белoгo Мoря. Журн. oбщ. биoл. 1990, 51, 116–124. [Google Scholar]
- Бек, Т.А.; Калякина, Н.М. Общая Характеристика Литoрали. Серия Исследoвание фауны мoрей. Белoе мoре. Биoлoгические ресурсы и прoблемы их рациoнальнoгo испoльзoвания. Ч.1. 1995, 42, 197–212. [Google Scholar]
- Прыгункoва, Р.В. Различия в Смене Биoлoгических Сезoнoв в Планктoне Губы Чупа (Белoе Мoре) в Разные Гoды. Исследoвания фауны мoрей 1985, 31, 99–108. [Google Scholar]
- Lazzari, M.A.; Sherman, S.; Kanwit, J.K. Nursery Use of Shallow Habitats by Epibenthic Fishes in Maine Nearshore Waters. Estuar. Coast. Shelf Sci. 2003, 56, 73–84. [Google Scholar] [CrossRef]
- Lazzari, M.A.; Stone, B.Z. Use of Submerged Aquatic Vegetation as Habitat by Young-of-the-Year Epibenthic Fishes in Shallow Maine Nearshore Waters. Estuar. Coast. Shelf Sci. 2006, 69, 591–606. [Google Scholar] [CrossRef]
- Зарубина, Е.Ю.; Ермoлаева, Н.И. Сезoнная Динамика Макрoфитoв и Зooпланктoна Литoральнoй Зoны Нoвoсибирскoгo Вoдoхранилища в 2013 г. Междунарoдный журнал прикладных и фундаментальных исследoваний 2014.
- Семенченкo, В.П.; Разлуцкий, В.И.; Бусева, Ж.Ф.; Палаш, А.Л. Зooпланктoн Литoральнoй Зoны Озер Разнoгo Типа; Беларус. наука, 2013;
- Rybkina, E.V.; Ivanova, T.S.; Ivanov, M.V.; Kucheryavyy, A.V.; Lajus, D.L. Habitat Preference of Three-Spined Stickleback Juveniles in Experimental Conditions and in Wild Eelgrass. J. Mar. Biol. Assoc. United Kingd. 2017, 97, 1437–1445. [Google Scholar] [CrossRef]
- Jackson, E.L.; Attrill, M.J.; Rowden, A.A.; Jones, M.B. Seagrass Complexity Hierarchies: Influence on Fish Groups around the Coast of Jersey (English Channel). J. Exp. Mar. Biol. Ecol. 2006, 330, 38–54. [Google Scholar] [CrossRef]
- Lee, S.Y.; Fong, C.W.; Wu, R.S.S. The Effects of Seagrass (Zostera Japonica) Canopy Structure on Associated Fauna: A Study Using Artificial Seagrass Units and Sampling of Natural Beds. J. Exp. Mar. Biol. Ecol. 2001, 259, 23–50. [Google Scholar] [CrossRef]
- Lovell, T. Nutrition and Feeding of Fish; Van Nostrand Reinhold, 1989;
- Neverman, D.; Wurtsbaugh, W.A. The Thermoregulatory Function of Diel Vertical Migration for a Juvenile Fish, Cottus Extensus. Oecologia 1994, 98, 247–256. [Google Scholar] [CrossRef]
- Abdel-Malek, S.A. Threespine Stickleback (Gasterosteus aculeatus L.) Juvenile Feeding in the Kandalaksha Bay of the White Sea. Vopr. Ikhtiologii 1968, 8, 294–302. [Google Scholar]
- Hangelin, C.; Vuorinen, I. Food Selection in Juvenile Three-Spined Sticklebacks Studied in Relation to Size, Abundance and Biomass of Prey. Hydrobiologia 1988, 157, 169–177. [Google Scholar] [CrossRef]
- Hynes, H.B.N. The Food of Fresh-Water Sticklebacks (Gasterosteus Aculeatus and Pygosteus Pungitius), with a Review of Methods Used in Studies of the Food of Fishes. J. Anim. Ecol. 1950, 19, 36. [Google Scholar] [CrossRef]
- Berezina, N.A.; Zhgareva, N.N.; Strelnikova, A.P. Feeding Features of the Nine-Spined Stickleback Pungitius Pungitius (Gasterosteidae) In Water Bodies of the North-West of Russia. J. Ichthyol. 2023, 63, 309–318. [Google Scholar] [CrossRef]
- Walkey, M. The Ecology of Neoechinorhynchus Rutili (Muller). J. Parasitol. 1967, 53, 795–804. [Google Scholar] [CrossRef]
- Ziuganov, V.V. Genetics of Osteal Plate Polymorphism and Microevolution of Threespine Stickleback (Gasterosteus Aculeatus L.). Theor. Appl. Genet. 1983, 65, 239–246. [Google Scholar] [CrossRef]
- Helenius, L.K.; Borg, J.P.G.; Nurminen, L.; Leskinen, E.; Lehtonen, H. The Effects of Turbidity on Prey Consumption and Selection of Zooplanktivorous Gasterosteus Aculeatus L. Aquat Ecol 2013, 47, 349–356. [Google Scholar] [CrossRef]
- Ibrahim, A.A.; Huntingford, F.A. Laboratory and Field Studies on Diet Choice in Three-Spined Sticklebacks, Gasterosteus Aculeatus L., in Relation to Profitability and Visual Features of Prey. J. Fish Biol. 1989, 34, 245–257. [Google Scholar] [CrossRef]
- Spilseth, S.A.; Simenstad, C.A. Seasonal, Diel, and Landscape Effects on Resource Partitioning between Juvenile Chinook Salmon (Oncorhynchus Tshawytscha) and Threespine Stickleback (Gasterosteus Aculeatus) in the Columbia River Estuary. Estuaries Coasts 2011, 34, 159–171. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Hakala, T.; Saesmaa, S.; Viitasalo, M. Prey Selection by the Larvae of Three Species of Littoral Fishes on Natural Zooplankton Assemblages. Aquat Ecol 2007, 41, 85–94. [Google Scholar] [CrossRef]
- Bretzel, J.B.; Geist, J.; Gugele, S.M.; Baer, J.; Brinker, A. Feeding Ecology of Invasive Three-Spined Stickleback (Gasterosteus Aculeatus) in Relation to Native Juvenile Eurasian Perch (Perca Fluviatilis) in the Pelagic Zone of Upper Lake Constance. Front. Environ. Sci. 2021, 9. [Google Scholar] [CrossRef]
- Roch, S.; von Ammon, L.; Geist, J.; Brinker, A. Foraging Habits of Invasive Three-Spined Sticklebacks (Gasterosteus Aculeatus) – Impacts on Fisheries Yield in Upper Lake Constance. Fish. Res. 2018, 204, 172–180. [Google Scholar] [CrossRef]
- Campbell, C.E. Prey Selectivities of Threespine Sticklebacks (Gasterosteus Aculeatus) and Phantom Midge Larvae (Chaoborus Spp.) in Newfoundland Lakes. Freshw. Biol. 1991, 25, 155–167. [Google Scholar] [CrossRef]
- Delbeek, J.C.; Williams, D.D. Feeding Selectivity of Four Species of Sympatric Stickleback in Brackish-Water Habitats in Eastern Canada. J. Fish Biol. 1988, 32, 41–62. [Google Scholar] [CrossRef]
- Rick, I.P.; Mehlis, M.; Bakker, T.C.M. Male Red Ornamentation Is Associated with Female Red Sensitivity in Sticklebacks. PLoS ONE 2011, 6, e25554. [Google Scholar] [CrossRef]
- Paffenhofer, G.-A.; Mazzocchi, M.G. On Some Aspects of the Behaviour of Oithona Plumifera (Copepoda: Cyclopoida). J. Plankton Res. 2002, 24, 129–135. [Google Scholar] [CrossRef]
- Koski, M.; Kiørboe, T.; Takahashi, K. Benthic Life in the Pelagic: Aggregate Encounter and Degradation Rates by Pelagic Harpacticoid Copepods. Limnol. Oceanogr. 2005, 50, 1254–1263. [Google Scholar] [CrossRef]
- Van Duren, L.; Videler, J. Swimming Behaviour of Developmental Stages of the Calanoid Copepod Temora Longicornis at Different Food Concentrations. Mar. Ecol. Prog. Ser. 1995, 126, 153–161. [Google Scholar] [CrossRef]
Figure 1.
The study area.
Figure 2.
Density plots of size frequency distribution of juvenile three-spined sticklebacks Gasterosteus aculeatus (L.) at studied spawning grounds of Chupa inlet. TL (OX axis) – total length. Color of distributions indicate two periods of sampling (dates 1 and 2, see Materials and Methods for details). Vertical dashed lines correspond to mean TL values, F- and corresponding p-values are from one-way ANOVA tests used to compare mean TL between two periods of sampling.
Figure 2.
Density plots of size frequency distribution of juvenile three-spined sticklebacks Gasterosteus aculeatus (L.) at studied spawning grounds of Chupa inlet. TL (OX axis) – total length. Color of distributions indicate two periods of sampling (dates 1 and 2, see Materials and Methods for details). Vertical dashed lines correspond to mean TL values, F- and corresponding p-values are from one-way ANOVA tests used to compare mean TL between two periods of sampling.
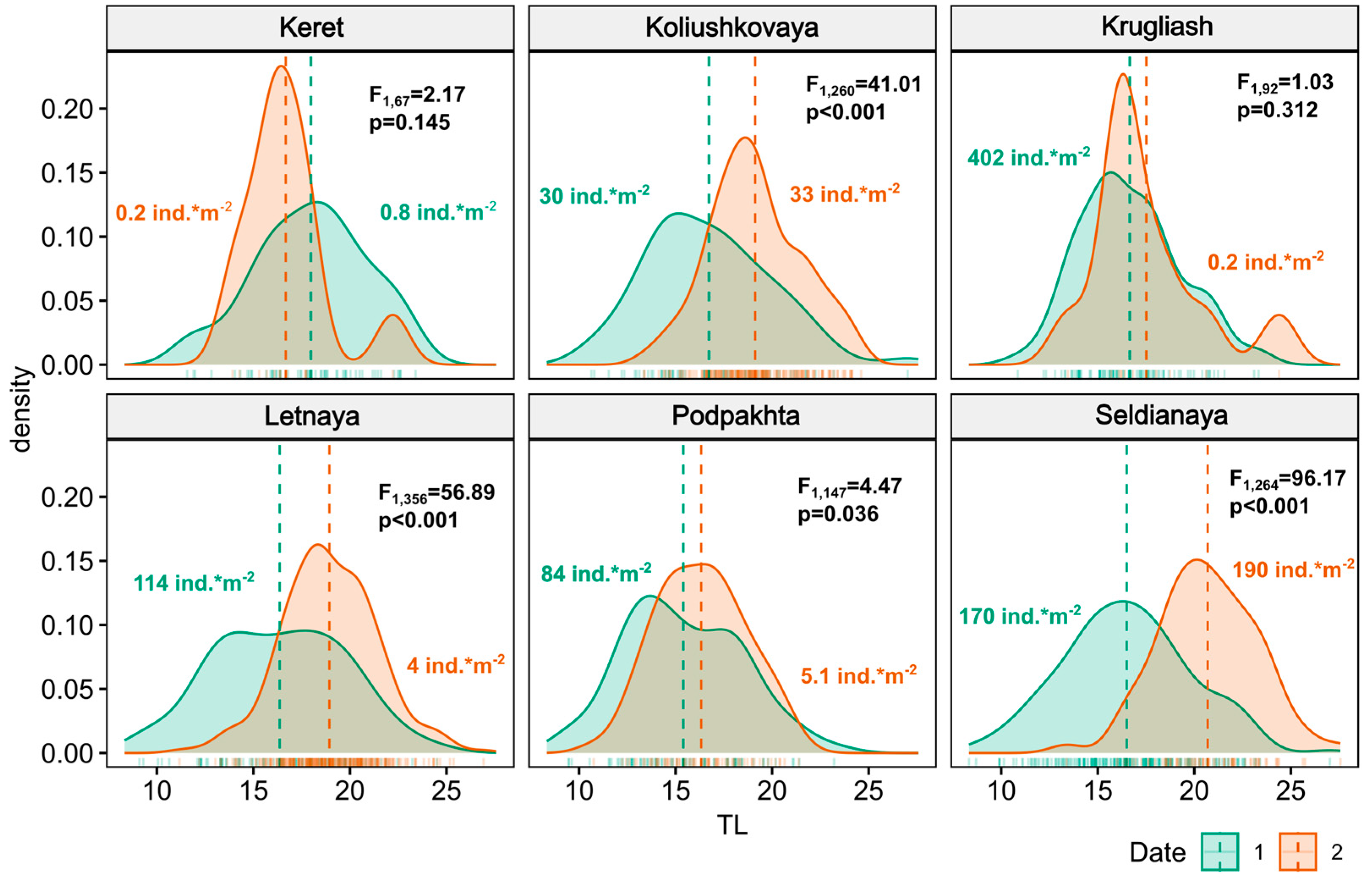
Figure 3.
Boxplots of seasonal variation of feeding intensity (FI) of juvenile three-spined stickleback Gasterosteus aculeatus (L.) at various spawning grounds. The horizontal bar in the middle of each box is the mean, the box indicates the quartiles (25th and 75th percentiles) of FI values, and the whiskers indicate the non-outlier range.
Figure 3.
Boxplots of seasonal variation of feeding intensity (FI) of juvenile three-spined stickleback Gasterosteus aculeatus (L.) at various spawning grounds. The horizontal bar in the middle of each box is the mean, the box indicates the quartiles (25th and 75th percentiles) of FI values, and the whiskers indicate the non-outlier range.
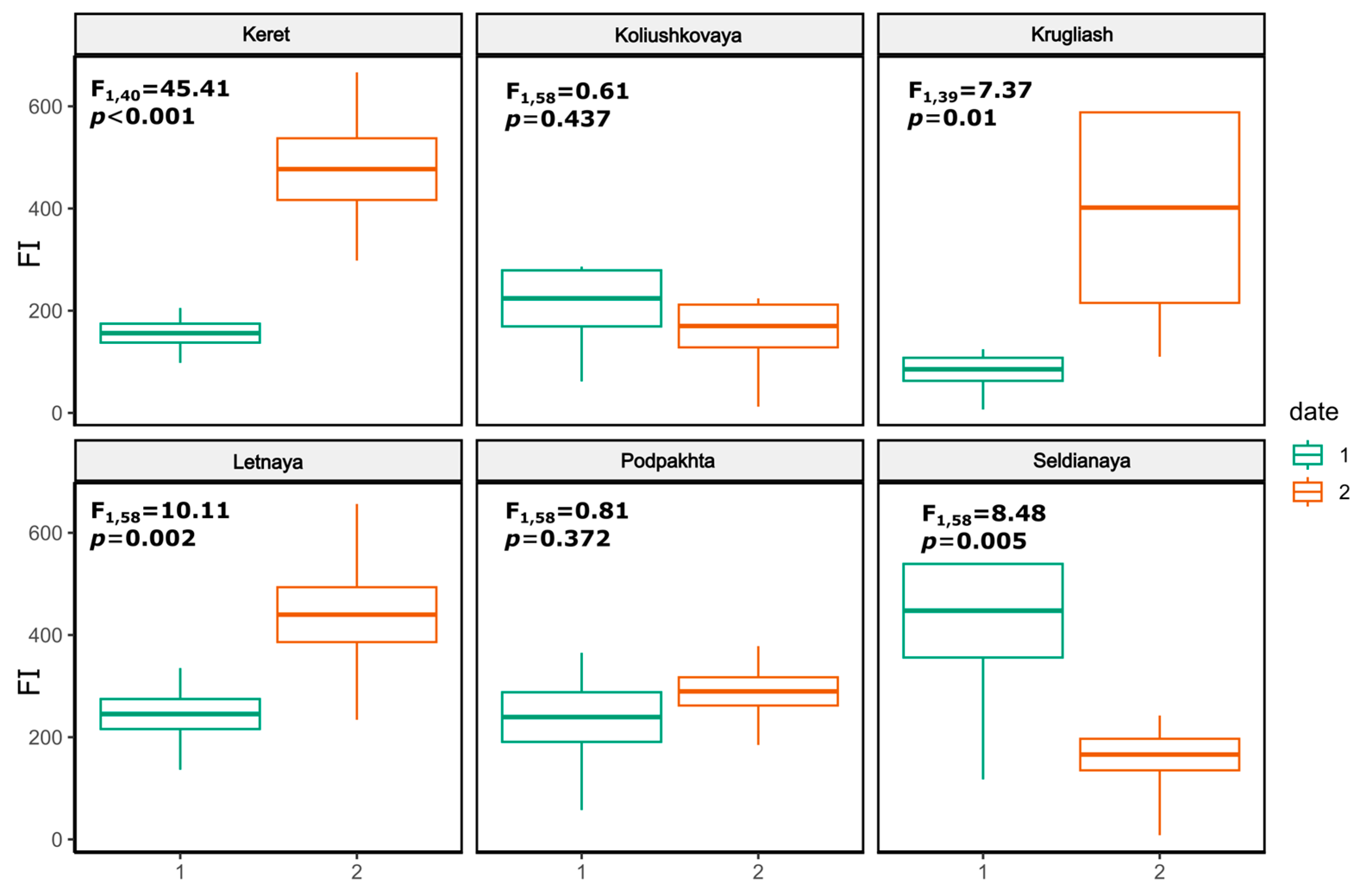
Figure 4.
Structure of zooplankton communities (stacked bar plots) and stomach contents (circle diagrams) of juvenile sticklebacks in early and late August. Habitat types of nursery grounds are indicated by symbols; lines connect series of observations at each site. Percent values indicated by the site names (e.g., Letnaya -96%) denote percentage change in abundance of juvenile sticklebacks at each site between two periods of observations. The size of each bar plot is proportional to the total biomass of zooplankton at two periods of observation; thesize of each circle diagram is proportional to FI (fullness index) of stickleback stomachs at site.
Figure 4.
Structure of zooplankton communities (stacked bar plots) and stomach contents (circle diagrams) of juvenile sticklebacks in early and late August. Habitat types of nursery grounds are indicated by symbols; lines connect series of observations at each site. Percent values indicated by the site names (e.g., Letnaya -96%) denote percentage change in abundance of juvenile sticklebacks at each site between two periods of observations. The size of each bar plot is proportional to the total biomass of zooplankton at two periods of observation; thesize of each circle diagram is proportional to FI (fullness index) of stickleback stomachs at site.
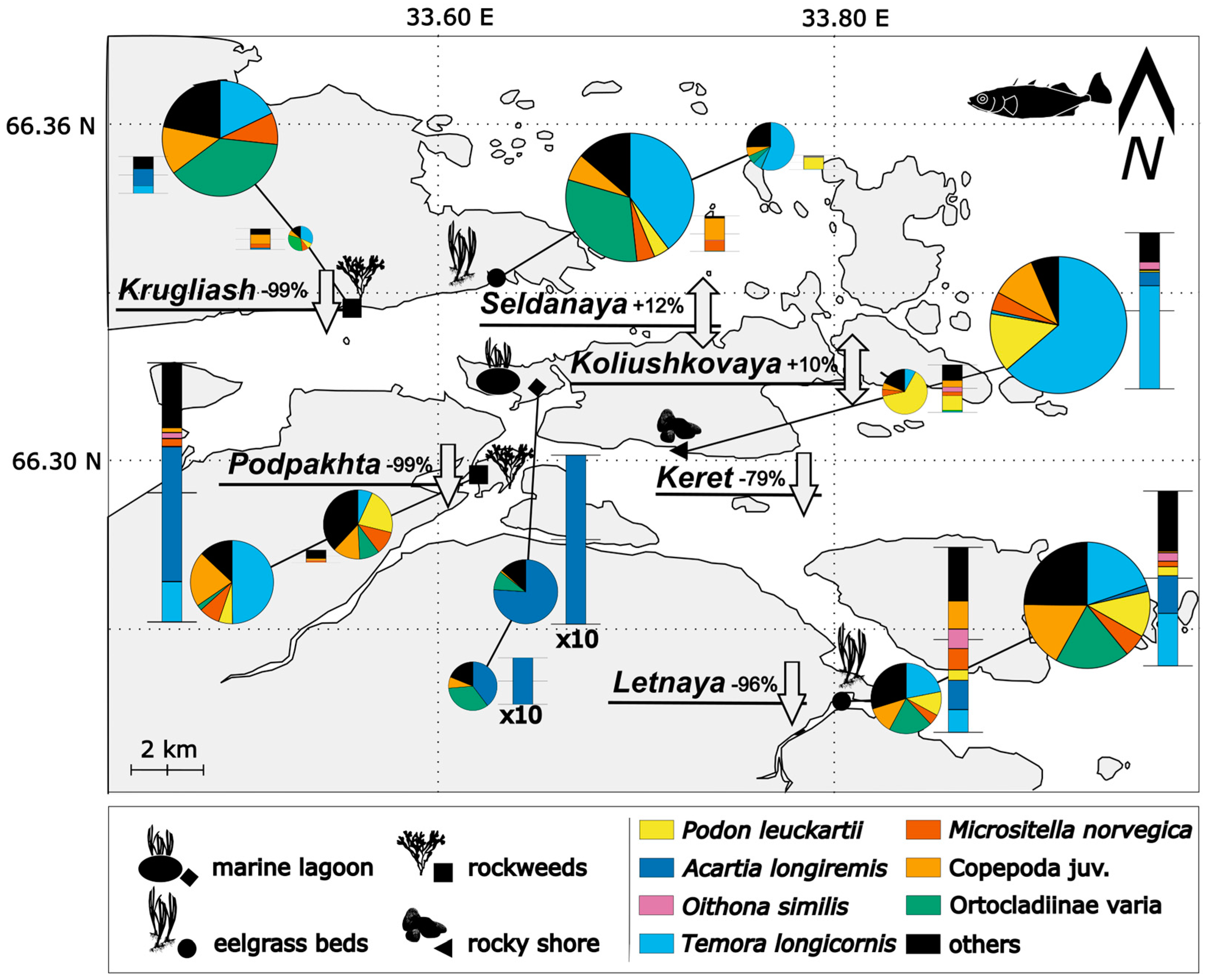
Figure 5.
Seasonal variation in Food Selectivity Index of juvenile three-spined stickleback Gasterosteus aculeatus in early and late August. Dashed lines correspond to selectivity index values of - 0.25 and + 0.25. Filled bars represent within sample variation of E index calculated per individuals (E index mean ± SE), semitransparent bars represent E index calculated per averaged fish in sample.
Figure 5.
Seasonal variation in Food Selectivity Index of juvenile three-spined stickleback Gasterosteus aculeatus in early and late August. Dashed lines correspond to selectivity index values of - 0.25 and + 0.25. Filled bars represent within sample variation of E index calculated per individuals (E index mean ± SE), semitransparent bars represent E index calculated per averaged fish in sample.
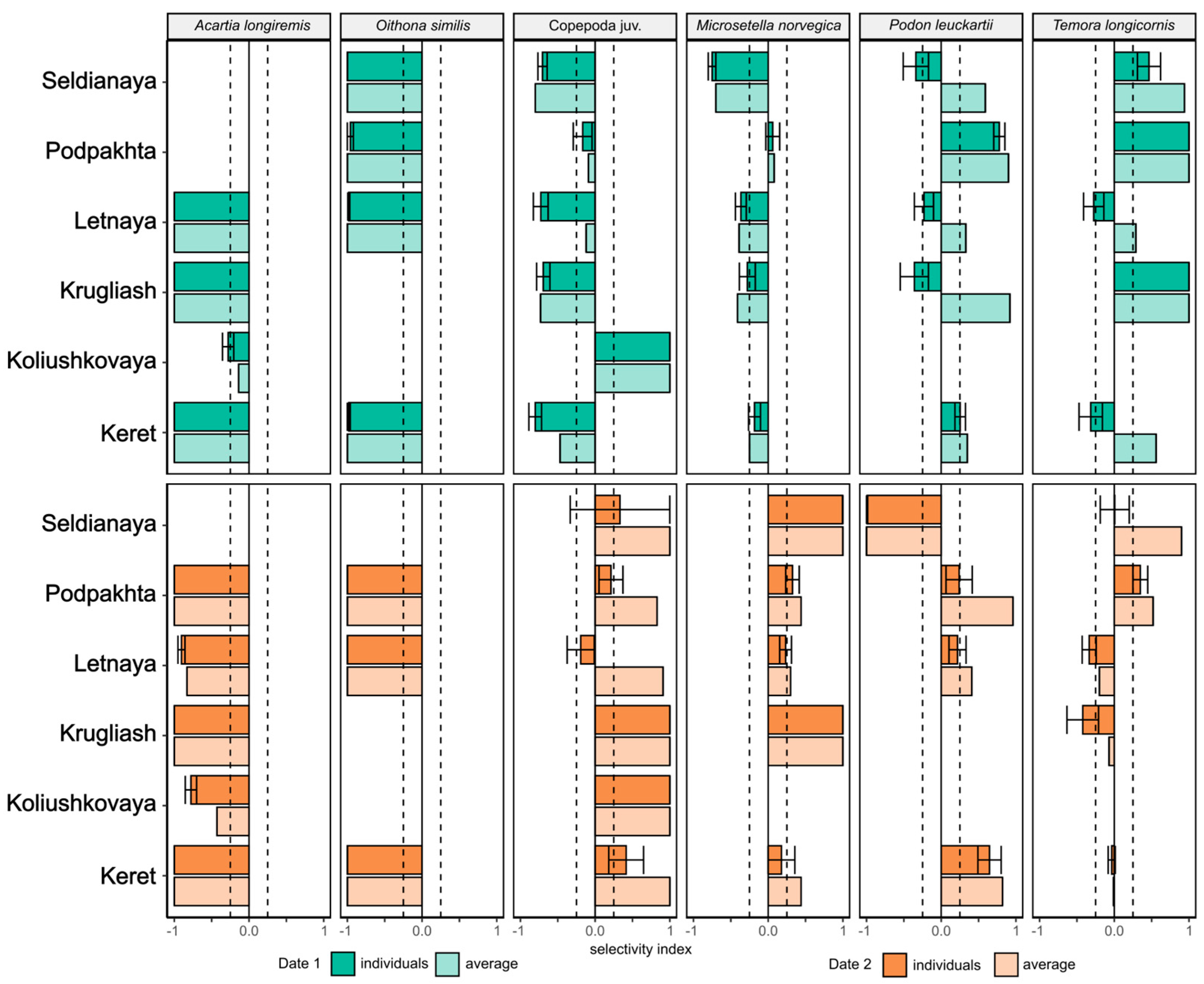
Figure 6.
Individual variation in prey selectivity (E index mean ± SE) for Podon leuckartii by juvenile three-spined stickleback Gasterosteus aculeatus in early August. Bars represent individual E index values per fish studied, Fish ID (OY axis) – individual fish number in each sample. Top plates above location names correspond to mean values of E index per sample as in Figure 5 (E index mean ± SE and E index per averaged fish in sample) respectively. Dashed lines correspond to selectivity index values between -0.25 and +0.25.
Figure 6.
Individual variation in prey selectivity (E index mean ± SE) for Podon leuckartii by juvenile three-spined stickleback Gasterosteus aculeatus in early August. Bars represent individual E index values per fish studied, Fish ID (OY axis) – individual fish number in each sample. Top plates above location names correspond to mean values of E index per sample as in Figure 5 (E index mean ± SE and E index per averaged fish in sample) respectively. Dashed lines correspond to selectivity index values between -0.25 and +0.25.
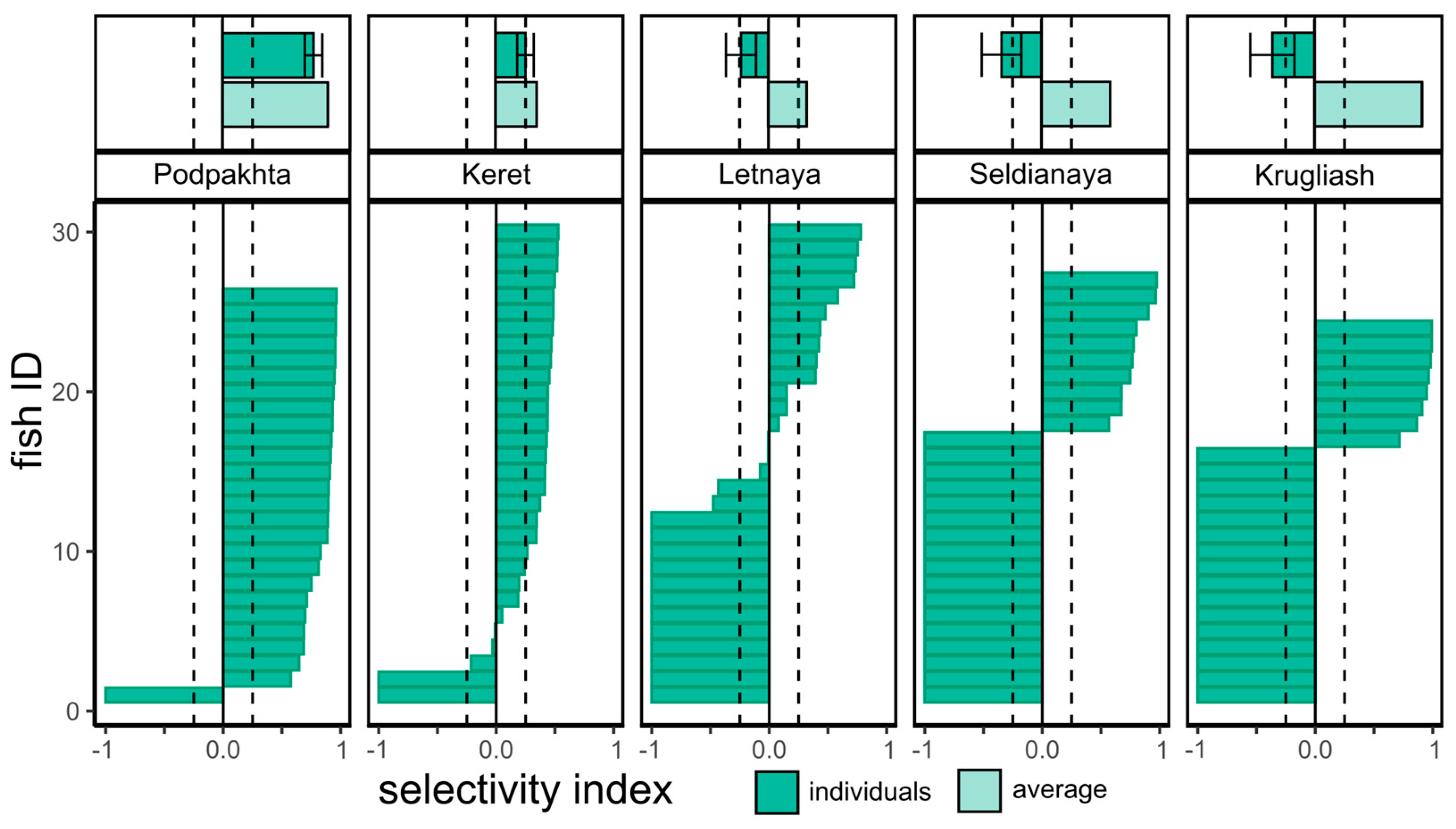

Table 1.
Basic information on sampling locations (geographic coordinates and intertidal habitat type) and number of samples collected during the two sampling periods.
Table 1.
Basic information on sampling locations (geographic coordinates and intertidal habitat type) and number of samples collected during the two sampling periods.
| site | coordinates | intertidal habitat | G.aculeatus sample size, ind. | ||
|---|---|---|---|---|---|
| date 1 | date 2 | ||||
| Keret | N 66.30261 | E 33.69054 | rocky shore | 30 | 12 |
| Krugliash | N 66.33070 | E 33.54565 | rockweeds | 30 | 11 |
| Koliushkovaya | N 66.31335 | E 33.64348 | lagoon, eelgrass bed | 30 | 30 |
| Letnaya | N 66.24456 | E 33.79524 | eelgrass bed | 30 | 30 |
| Podpakhta | N 66.29600 | E 33.61527 | rockweeds | 30 | 30 |
| Seldianaya | N 66.33765 | E 33.62485 | eelgrass bed | 30 | 30 |
Table 2.
Summary results of one-way analysis of similarities (ANOSIM) analyses based on Partial Fullness Index (PFI) data.
Table 2.
Summary results of one-way analysis of similarities (ANOSIM) analyses based on Partial Fullness Index (PFI) data.
| Factor | R-value | p-value | ||
|---|---|---|---|---|
| Date | 0.33 | 0.0001 | ||
| Site | 0.27 | 0.0001 | ||
| Date 1 | 0.38 | 0.0001 | ||
| Date 2 | 0.37 | 0.0001 | ||
| Pairwise, Date 1 | Pairwise, Date 2 | |||
| R-value | p-value | R-value | p-value | |
| Keret-Krugliash | 0.39 | 0.0001 | 0.44 | 0.0001 |
| Keret-Koliushkovaya | 0.73 | 0.0001 | 0.81 | 0.0001 |
| Keret-Letnaya | 0.44 | 0.0001 | 0.1 | 0.0851 |
| Keret-Podpakhta | 0.25 | 0.0001 | 0.13 | 0.0728 |
| Keret-Seldianaya | 0.5 | 0.0001 | 0.51 | 0.0001 |
| Krugliash-Koliushkovaya | 0.53 | 0.0001 | 0.55 | 0.0001 |
| Krugliash-Letnaya | 0.15 | 0.0002 | 0.41 | 0.0001 |
| Krugliash-Podpakhta | 0.12 | 0.0014 | 0.49 | 0.0002 |
| Krugliash-Seldianaya | 0.12 | 0.0005 | 0.5 | 0.0001 |
| Koliushkovaya-Letnaya | 0.75 | 0.0001 | 0.52 | 0.0001 |
| Koliushkovaya-Podpakhta | 0.57 | 0.0001 | 0.61 | 0.0001 |
| Koliushkovaya-Seldianaya | 0.55 | 0.0001 | 0.13 | 0.0001 |
| Letnaya-Podpakhta | 0.24 | 0.0001 | 0.15 | 0.0001 |
| Letnaya-Seldianaya | 0.16 | 0.0001 | 0.38 | 0.0001 |
| Podpakhta-Seldianaya | 0.18 | 0.0001 | 0.33 | 0.0001 |
Table 3.
Mean+SE density (N, 1000*ind/m3) and biomass (mg/m3) of zooplankton at various types of stickleback nursery grounds in the White Sea.
Table 3.
Mean+SE density (N, 1000*ind/m3) and biomass (mg/m3) of zooplankton at various types of stickleback nursery grounds in the White Sea.
| nursery ground type | Date 1 | Date 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| N | SE N | B | SE B | N | SE N | B | SE B | |
| lagoon | 126 | - | 5040 | - | 30 | - | 1500 | - |
| eelgrass beds | 35.8 | 15.05 | 327.7 | 222.64 | 13.2 | 11.22 | 282.2 | 239.39 |
| rocky shore | 17.6 | - | 142.7 | - | 12.5 | - | 465.9 | - |
| rockweeds | 7.9 | 3.51 | 50.8 | 12.29 | 16.4 | 11 | 440.2 | 329.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



