
Submitted:
18 November 2023
Posted:
20 November 2023
You are already at the latest version
Abstract
The ADP-ribosylation factor 6 (Arf6), as the only member of the Arf family III protein, has been extensively studied for its diverse biological functions in animals. Previously, the Arf6 protein in Magnaporthe oryzae was found to be crucial for endocytosis and polarity establishment during asexual development. However, its role remains unclear in Sclerotinia sclerotiorum. Here, we identified and characterized SsArf6 in S. sclerotiorum using a reverse genetic approach. Deletion of SsArf6 impaired hyphal growth and development, and produced more branches. Interestingly, knockout of SsArf6 enhanced the tolerance of S. sclerotiorum to oxidative stress but the sensitivity to osmotic stress, and increased melanin accumulation, indicative of different roles of SsArf6 in various stress responses. Moreover, the appressorium formation was severely impaired, and fungal virulence to host plants was significantly reduced. Overall, our findings demonstrate the essential role of SsArf6 in hyphal development, stress responses, appressorium formation, and fungal virulence to host plants.
Keywords:
SsArf6
; hyphal growth
; appressorium formation
; stress response
; virulence
; melanin
1. Introduction
Sclerotinia sclerotiorum, as a notorious soilborne plant pathogenic fungus, which has an extremely broad host range, is capable of infecting economically significant crops such as rapeseed, sunflower, and soybeans, resulting in substantial agricultural losses [1,2,3,4,5]. In China, one of the main disease of oilseed rape is Sclerotinia stem rot (SSR) caused by S. sclerotiorum, leading to losses of approximate 8.4 billion RMB annually[6,7]. S. sclerotiorum produces sclerotia that can survive in the field for multiple years [8,9]. In addition, it employs different infection ways, including infection of host plant tissues through the hypha by germination of sclerotia. When conditions are favorable, sclerotia can also germinate to form apothecia, releasing many ascospores to infect the hosts [1,10].
Previous studies indicate that the interaction mechanisms between S. sclerotiorum and its host are intricate. To establish successful colonization, S. sclerotiorum undergoes specific morphological changes in its hyphal tips, leading to the formation of appressoria [11,12,13]. Appressoria play a crucial role in adhering to the host surface and penetrating the cuticle [14]. In the past few decades, some proteins related to the development and formation of appressoria have been identified and characterized in the S. sclerotiorum, including mitogen-activated protein kinase (MAPK) signaling pathway proteins SsSte12 [15] and SsFkh1 [16], autophagy-related proteins SsATG8, SsNBR1 [17], SsFoxE3 [18], and SsAtg1 [19], as well as TOR signaling pathway protein SsTOR[20]. Additionally, secreted protein SsRhs1 [21], Ssams2 [22], SsNsd1 [23]and Magnaporthe appressoria-specific (MAS) protein Sscnd1 [24] has also been confirmed to participate in appressoria formation.
Growing evidences suggest that the infection of the host by S. sclerotiorum happens in stages [14,25]. During the early stages of infection, besides directly invading host tissues through appressoria, S. sclerotiorum also secretes virulent factors, for instance oxalic acid [26,27], cell wall-degrading enzymes [28,29], and effectors [30,31] to impede mechanisms for recognizing and defending against pathogens [32]. Once colonization is achieved, S. sclerotiorum enters into the necrotrophic phase, characterized by the production of substantial amounts of reactive oxygen species (ROS) and virulent factors, which induce rapid cell death and the progression of necrotic symptoms [3,14,25,32,33]. Currently, due to the lack of resistant varieties, chemical method is mainly used to control SSR [34]. However, S. sclerotiorum has been reported to develop resistance to fungicides [35,36]. This has raised significant attention towards the control and prevention of S. sclerotiorum.
ADP-ribosylation factor 6 (Arf6) is a member of the ADP-ribosylation factors (Arf) family, which can regulate the endomembrane recycling and actin cytoskeleton remodeling at the cell surface [37,38]. In mammals, Arf6 plays a crucial role in regulating neutrophil energy metabolism [39], cancer cell invasion, metastasis and proliferation [40], as well as membrane lipid homeostasis [41]. Additionally, a homolog of Arf6 in Aspergillus nidulans, named ArfB, functions in endocytosis to play important roles in polarity establishment during isotropic growth and polarity maintenance during hyphal extension [42]. The Arf6's homolog in Magnaporthe oryzae also has similar functions [38]. However, the biological function of Arf6 in S. sclerotiorum remains unclear.
Through reverse genetic approaches, we characterized the roles of SsArf6 in S. sclerotiorum, and found that SsArf6 is highly conserved among various plant pathogenic fungi. Knockout of SsArf6 led to hindered mycelium growth and development, producing more branches, aerial hyphae and fewer sclerotia. Surprisingly, SsArf6 negatively regulates melanin accumulation and hydrogen peroxide resistance. In addition, deletion of SsArf6 significantly increased the sensitivity of S. sclerotiorum to osmotic stress. Most importantly, the appressorium formation and virulence to host plants exhibited severe impairments in knockout mutants. Together, our results suggesting that SsArf6 plays a significant role in the formation of appressorium, hyphal development, resistance to abiotic stress in S. sclerotiorum and fungal virulence to host plants.
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
Cultured in a constant temperature incubator at 20℃, the wild-type strain (WT), knockout mutant, and complemented strains were grown on potato dextrose agar (PDA) plates. The PDA plates were supplemented with 200 μg/mL hygromycin B (Roche) for the knockout mutants and PDA plates supplemented with 100 μg/mL G418 Sulfate (Geneticin) (Yeasen) for the complemented strain.
2.2. Plant Materials and Growth Conditions
Arabidopsis thaliana, oilseed rape (Brassica napus) and Nicotiana benthamiana seedlings were cultivated in a growth room at a temperature of 22℃, with a photoperiod of 16 hours of light followed by 8 hours of darkness. Except for Brassica napus, which were cultivated for 5 weeks, all other plants were cultivated for 4 weeks, were selected for the virulence assays with S. sclerotiorum.
2.3. Phylogenetic Tree Construction and Sequence Analysis
Homologous sequences of Arf6 in Zymoseptoria tritici, S. sclerotiorum, Ustilago maydis, Botrytis cinerea, Magnaporthe oryzae, Fusarium oxysporum and Fusarium graminearum were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/). The Arf6 gff3 files were got from the ensemblFungi database (https://fungi.ensembl.org/). The phylogenetic tree was constructed using MEGA11 programs with 1,000 bootstrap replicates and the neighbor joining method[43]. The gene feature was visualized by GSDS2.0 [44], and the alignment of Arf6 proteins was performed using Clustal Omega [45] and results viewed with Jalview [46].
2.4. Knockout and Complementation of SsArf6
The split-marker method was employed to knockout the SsArf6 (sscle_03g022330) gene in S. sclerotiorum. As previously described [47], two rounds of PCR were used to construct the replacement fragments SsArf6-UP-HY and SsArf6-Down-YG, which were used to replace the target gene SsArf6. The PCR products were co-transformed into wild-type (WT) strain protoplasts. SsArf6 knockout transformants were selected on PDA plates containing 200 μg/mL hygromycin B, and at least three rounds of hyphae tip transfer and protoplast purification were performed to obtain knockout mutant homozygotes.
In genetic complementation, a 1746-bp fragment consisting of upstream and full-length genomic DNA of SsArf6 was amplified specifically through PCR. The resulting fragment was individually fused with NEO fragments, which were amplified from the pCH-EF-1 plasmid (provided by D. Jiang from Huazhong Agricultural University). The resulting PCR products were co-transformed into protoplasts of the knockout mutant strain. Validation was achieved through PCR using six sets of primers (Table S1), and the transformed knockout and complementation strains were selected for further experimentation.
2.5. DNA and RNA Manipulation
The mycelium of S. sclerotiorum was cultivated on PDA medium containing cellophane for 48 hours, and then ground into fine powder in liquid nitrogen. Genomic DNA was extracted from both WT and mutant strains using the cetyltrimethylammonium bromide (CTAB) method [48]. The extracted WT genomic DNA was used as a template for amplifying the full-length sequence and the flanking sequence of SsArf6. The extracted genomic DNA from the mutants was used to confirm the knockout and complementation of SsArf6. To evaluate the expression of SsArf6 in the knockout and complementation strains at the transcriptional level, the mycelium was collected after 48 hours of cultivation on PDA medium containing cellophane, and total RNA was extracted using the SteadyPure Plant RNA Extraction Kit (Accurate Biology). The first-strand cDNA was synthesized from the total RNA using the Evo M-MLV RT-PCR Kit (Accurate Biology) as the template. Semi-quantitative RT-PCR was performed with cDNA as the template for 28 PCR cycles. All primers used in this study were listed in Table S1.
2.6. Colony Morphology Observation
The growth lengths of the WT, knockout mutant (ΔSsarf6) and complemented strain (SsArf6-C) on potato dextrose agar (PDA) medium were measured every 24 hours. The colony morphologies of the WT, ΔSsarf6, and SsArf6-C strains on PDA medium were documented using a digital camera at 24 hours, 48 hours, and 8 days of cultivation respectively.
2.7. Stress Treatment
The WT, ΔSsarf6, and SsArf6-C strains were treated with 5mM, 10mM and 15mM of H2O2; cell wall inhibitor Congo Red (CR), 0.02% of Sodium Dodecyl Sulfate (SDS); osmotic stressors 0.5M NaCl, KCl and 1M sorbitol and glucose. After 48 hours, the colony morphology was recorded using a digital camera and the inhibition rate was calculated. The inhibition rate (%) was calculated as 100 × (the colony radius of the strain on pure PDA subtracted by the colony radius of the strain under different stressors) divided by the colony radius of the strain on pure PDA.
2.8. Analysis of Compound Appressoria, OA and Virulence to Host Plants
The WT, ΔSsarf6, and SsArf6-C strains were inoculated onto a PDA medium containing 0.005% (W/V) Bromophenol Blue. After 48 hours, the color change was recorded using a digital camera. For appressorium formation observation, the WT, ΔSsarf6, and SsArf6-C strain blocks were inoculated onto a slide, and after 24 hours of cultivation, the morphology of the compound appressoria was observed and recorded under an optical microscope (Axio Imager 2, ZEISS, Oberkochen, Germany). After a cultivation period of 36 hours, the appressorium of WT, ΔSsarf6, and SsArf6-C strain was recorded using a digital camera.
To observe the morphology of the compound appressoria formed on onion epidermal cells, the S. sclerotiorum strains were inoculated onto the onion epidermis cultured for 16 hours as previously described [49]. After that, the onion epidermis was soaked in a 0.5% Trypan Blue solution for 30 minutes. Next, a bleaching solution was prepared in a ratio of ethanol: acetic acid: glycerol of 3:1:1 and used to decolorize the samples. The morphology of the composite attached cells was then observed and recorded under an optical microscope. For the virulence analysis, the WT, ΔSsarf6, and SsArf6-C strains were inoculated onto detached leaves of A. thaliana, B. napus and N. benthamiana. After 36 hours of infection, the infection morphology was recorded using a digital camera, and the lesion areas are analyzed using Image J software [50].
3. Results
3.1. Identification of the Arf6 homolog in S. sclerotiorum
Previously, when using a forward genetic approach to screen hypo-virulent mutant of S. sclerotiorum, we got one mutant with a candidate gene which was speculated to be a guanine exchange factor for Arf6(SsEFA6, sscle_12g090120), related to the pathogenic capacity of S. sclerotiorum (unpublished data). To determine whether Arf6 is also associated with pathogenicity, we then performed a genomic blasting and identified sscle_03g022330 as the homologous gene of MoArf6 in S. sclerotiorum, and named as SsArf6, which comprised four exons and three introns, spanning a length of 794 base pairs and encoding 186 amino acids. As shown in Figure 1A, SsArf6 homologous proteins were prevalent among plant pathogenic fungi. When conducting gene structure analysis of these homologues, we discovered that UmArf6 contained only one CDS region, while SsArf6 and BcArf6 had up to four CDSs (Figure 1B). In addition, protein sequence alignment revealed evolutionary conservation of SsArf6, which had a high similarity with Arf6 homologs in F. oxysporum, F. graminearum, B. cinerea, Z. tritici, and U. maydis, with 86.02%, 84.95%, 85.48%, 98.39%, 76.92%, and 76.84% amino acid sequence identity respectively (Figure 1C).
3.2. Knockout of SsArf6 leads to aberrant mycelium growth, increased melanin accumulation and decreased sclerotium production
In order to investigate the biological function of SsArf6 in S. sclerotiorum, we generated the SsArf6 deletion mutant, ΔSsarf6 (Figure S1A), using a split-marker method based on homologous recombination. The results indicated that SsArf6 was completely replaced by hph (Figure S1B, Figure S3). The growth of ΔSsarf6 strain mycelium was hindered, with an average growth rate of 1.27cm/24h, significantly lower than that of the WT strain (Figure 2A, B). In addition, the ΔSsarf6 strains produced more branches and melanin compared to WT strains (Figure 2C, D). After incubation on PDA medium for 8 days, there were noticeable differences in colony morphology among WT and ΔSsarf6 strains. Specifically, the ΔSsarf6 strain formed many aerial hyphae (Figure 2E) but fewer sclerotia (Figure 2F, G) compared to WT. Furthermore, the phenotype of SsArf6-C, which was generated through in situ complementation in the ΔSsarf6 background using a knock-in method (Figure S2A, B, Figure S3), was similar to WT (Figure 2). These findings strongly indicate that SsArf6 plays an important role in hyphal growth, melanin accumulation and sclerotium production.
3.3. SsArf6 is implicated in abiotic stress response in S. sclerotiorum
To explore the response of SsArf6 to different stresses, S. sclerotiorum was treated with various cell wall inhibitors and osmotic stresses. When exogenous 0.5M NaCl, 0.5M KCl, 1M glucose and 1M sorbitol were applied, the colony radii of ΔSsarf6 were significantly reduced, and the inhibition rate were significantly higher than that of WT and SsArf6-C strains (Figure 3A, B). However, in the medium supplemented with cell wall inhibitors Congo red (CR) and Sodium Dodecyl Sulfate (SDS), ΔSsarf6 demonstrated a stress response that was comparable to both the WT and SsArf6-C strains (Figure 3C, D) .
Former studies suggested that fungal melanin is frequently located intracellularly and has antioxidant properties, as it can neutralize reactive oxygen species and other free radicals [51]. Therefore, we assessed the antioxidant capacity of the ΔSsarf6 mutant, and found that they exhibited significant tolerance to hydrogen peroxide compared to the WT and SsArf6-C strains (Figure 4A, B), indicative of the essential role of SsArf6 in regulating the abiotic stress responses in S. sclerotiorum.
3.4. SsArf6 is involved in compound appressoria development
To successfully colonize, S. sclerotiorum undergoes specific morphological changes in its hyphal tips to form appressoria to break down the host cuticle [14]. Therefore, we analyzed the formation of compound appressoria in ΔSsarf6 mutants, and found that the production of compound appressoria by ΔSsarf6 mutants was significantly abnormal compared to the WT and SsArf6-C strain. Both WT and SsArf6-C strain formed compound appressoria on the slide surface as well as onion epidermal cells, while ΔSsarf6 mutants did not (Figure 5A, B, C). Furthermore, we also assessed the secretion capacity of ΔSsarf6 mutants for oxalic acid and revealed that the secretion of oxalic acid was normal in the ΔSsArf6 strain, with no significant differences compared to the WT and SsArf6-C strains (Figure 5D). These results suggest that SsArf6 is essential for the formation of complex appressoria and might be associated with the pathogenic ability of S. sclerotium.
3.5. SsArf6 is essential for virulence to host plants
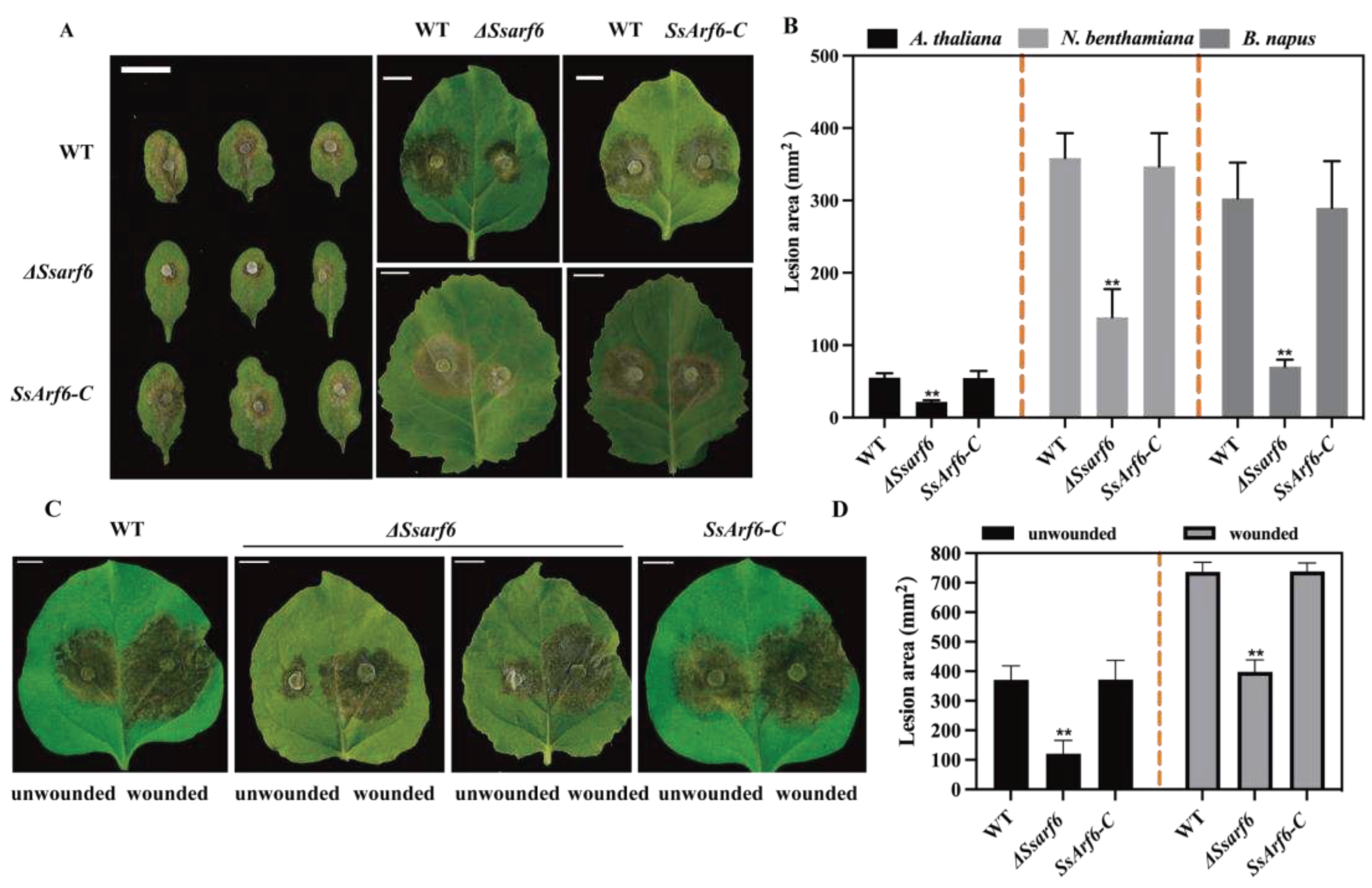
To confirm whether SsArf6 is related to the pathogenicity of S. sclerotiorum, we inoculated WT, ΔSsarf6, and SsArf6-C strain on detached leaves of A. thaliana, B. napus and N. Benthamian. After 36 hours of infection, the ΔSsarf6, WT and SsArf6-C strain caused water-soaked pale brown lesions on the leaves. However, the ΔSsarf6 mutants showed a significant decrease in lesion area compared to the WT and SsArf6-C strains on leaf blades of different hosts (Figure 6A, B). In addition, when inoculated onto the wounded N. Benthamiana detached leaves, we observed a significant increase in the infection area of ΔSsarf6 after 36 hours, compared to the intact detached leaves, under the same infection conditions, but the infection area of ΔSsarf6 was still significantly less than that of WT and SsArf6-C strains (Figure 6C, D). These findings strongly indicate that SsArf6 plays an essential role in the fungal virulence to hosts, except for its function in appressoria formation of S. sclerotiorum.
4. Discussion
Arf6 is a conserved protein that plays a vital role in the development of fungi, deletion of Arf6 in M. oryzae and A. nidulans results in slower mycelium growth, increased number of mycelial branches [38,42]. In this study, we identified a homologous protein SsArf6 in S. sclerotiorum, which shares a high degree of similarity with Arf6 proteins in other plant pathogenic fungi. Knockout of SsArf6 led to impaired hyphal development, increased branching and melanin accumulation, excessive growth of aerial hyphae as well as negatively impacted sclerotia yield in S. sclerotiorum.
Fungal melanin, as a potent antioxidant, protects cells by scavenging hydrogen peroxide, hydroxyl radicals, and superoxide anions [52]. To investigate whether or not SsArf6 mutant be tolerant to oxidative stress due to its melanin accumulation, we simulated oxidative stress by adding different concentrations of hydrogen peroxide exogenously to evaluate the antioxidant capacity of the SsArf6 deletion mutant. As expected, ΔSsarf6 mutants indeed exhibited significant tolerance to oxidative stress caused by hydrogen peroxide, compared to WT and SsArf6-C. Additionally, we observed that ΔSsarf6 mutants were more sensitive to hyperosmotic stress but unaffected by cell wall inhibitory agents, indicating that SsArf6 plays an important but different roles in responding to different abiotic stress.
Appressoria and oxalic acid are essential for the interaction between S. sclerotiorum and its host.The formation of appressoria helps to break down the physical barriers of the host, such as the cell wall and cuticle [53], and oxalic acid is a key virulence factor in the invasion process, as its secretion enhances the activity of hydrolytic enzymes [26], induces programmed cell death in plants [54], and inhibits host defenses [27]. The absence of SsArf6 resulted in abnormal appressoria development while it did not affect oxalic acid secretion in S. sclerotiorum, suggests that SsArf6 is involved in the interaction between S. sclerotiorum and its host other than oxalic acid pathway.
In M. oryzae, Arf6 was reported to be not essential for its pathogenicity [38]. When fungal virulence of the SsArf6 deletion mutant was assessed, we observed significantly decreases compared to WT and SsArf6-C strain. Furthermore, when infection assays were performed on wounded leaves of N. benthamiana, the area of infection by the ΔSsarf6 mutant significantly increased compared to unwounded leaves 36 hours post inoculation, although it still remained to be significantly lower than that of the WT and SsArf6-C strains under the same infection conditions. These evidences indicated an essential role of SsArf6 in fungal virulence to hosts, except for its function in appressoria formation of S. sclerotiorum. Further characterization is needed regarding the specific mechanisms by which SsArf6 regulates the formation of appressoria and pathogenicity.
Taken together, SsArf6 is involved in mycelial growth, appressorium development, and stress response in S. sclerotiorum, and contributes to the infection process and fungal virulence to host plants. Our study provides evidence for understanding of the role of Arf6 in the interaction between S. sclerotiorum and its hosts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Knockout of SsArf6 in S. sclerotiorum. (A) The schematic diagram of the strategy used for knockout of SsArf6. The SsArf6, Hygromycin phosphotransferase (hph), and G418 resistance gene NEO were denoted by yellow, red, and green rectangles, respectively. The labeled primers in the diagram were used for amplifying flanking sequences of SsArf6 or facilitating mutant screening. (B) The verification of knockout of SsArf6 by PCR. Genomic DNA obtained from WT, knockout mutant ΔSsarf6 were utilized as templates for PCR. A total of six primer pairs were employed to detect the insertion of hph and its upstream and down-stream fragments, as well as to confirm the knockout of SsArf6. The sizes of the amplified bands were indicated within brackets. The lanes labeled as M represent the DNA marker. Figure S2: Complementation of SsArf6 in ΔSsarf6 mutants of S. sclerotiorum. (A) The schematic diagram of the strategy used for the complementation of ΔSsarf6. The SsArf6, Hygromycin phosphotransferase (hph), and G418 resistance gene NEO were denoted by yellow, red, and green rectangles, respectively. The labeled primers in the diagram were used for amplifying full-length genomic DNA of SsArf6 or facilitating transformant screening. (B) The verification of complementation of SsArf6 by PCR. Genomic DNA obtained from the deletion mutant ΔSsArf6, and complemented strain SsArf6-C were utilized as templates for PCR. A total of six primer pairs were employed to confirm the complementation of SsArf6. The sizes of the amplified bands were indicated within brackets. The lanes labeled as M represent the DNA marker. Figure S3: The semi-quantitative RT-PCR analysis of the mutant strains ΔSsArf6 and SsArf6-C. Table S1: Primers in this study.
Author Contributions
K.W. and S.X. planned and designed the research. K.W. carried out most of the experiments. K.W., S.W. and S.X. wrote the original draft of the manuscript. S.W. and Q. X. contributed to the mutagenized population of S. sclerotiorum. T.W. analyzed and validated the data. All authors contributed to and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China (grant 31971836).
Data Availability Statement
The GenBank accession numbers (species names) for organisms used in this study are as follows: Zymoseptoria tritici (ZtArf6, XP_003857013.1), Sclerotinia sclerotiorum (SsArf6, APA07463.1), Ustilago maydis(UmArf6, XP_011391884.1), Botrytis cinerea (BcArf6, XP_001547581.1), Magnaporthe oryzae (MoArf6, XP_003715902.1), Fusarium oxysporum (FoArf6, XP_018253040.1), Fusarium graminearum (FgArf6, XP_011321112.1), were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/). The Arf6 gff3 files were got from the ensemblFungi database (https://fungi.ensembl.org/).
Acknowledgments
We cordially thank Xin Li (University of British Columbia) for the critical suggestions regarding the experiments, Daohong Jiang (Huazhong Agricultural University) for sharing pCH-EF-1 plasmid, and Jeffrey Rollins (University of Florida) for sharing WT S. sclerotiorum strain 1980. We would like to thank the National Natural Science Foundation of China (grant 31971836) for funds awarded to S.X.
Conflicts of Interest
All authors declare that there is no conflict of interest.
References
- Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The notorious soilborne pathogenic fungus Sclerotinia sclerotiorum: An update on genes studied with mutant analysis. Pathogens 2019, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Ding, L.N.; Li, T.; Guo, X.J.; Li, M.; Liu, X.Y.; Cao, J.; Tan, X.L. Sclerotinia stem rot resistance in rapeseed: Recent progress and future prospects. J. Agric. Food Chem. 2021, 69, 2965–2978. [Google Scholar] [CrossRef] [PubMed]
- Fass, M.I.; Rivarola, M.; Ehrenbolger, G.F.; Maringolo, C.A.; Montecchia, J.F.; Quiroz, F.; García-García, F.; Blázquez, J.D.; Hopp, H.E.; Heinz, R.A.; et al. Exploring sunflower responses to Sclerotinia head rot at early stages of infection using RNA-seq analysis. Sci. Rep. 2020, 10, 13347. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.D.; Hartman, G.L.; Mueller, D.S.; Leitz, R.A.; Nickell, C.D.; Pedersen, W.L. Yield and seed quality of soybean cultivars infected with Sclerotinia sclerotiorum. Plant Dis. 1998, 82, 826–829. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Hua, W.; Yin, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed research and production in China. CROP.J 2017, 5, 127–135. [Google Scholar] [CrossRef]
- Zhang, X.; Cheng, J.; Lin, Y.; Fu, Y.; Xie, J.; Li, B.; Bian, X.; Feng, Y.; Liang, W.; Tang, Q.; et al. Editing homologous copies of an essential gene affords crop resistance against two cosmopolitan necrotrophic pathogens. Plant Biotechnol. J. 2021, 19, 2349–2361. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Harper, G.E.; Frampton, C.M.; Stewart, A. Factors influencing survival of sclerotia of Sclerotium cepivorum in New Zealand soils. N. Z. J. Crop Hortic. Sci 2002, 30, 29–35. [Google Scholar] [CrossRef]
- Willetts, H.J.; Wong, J.A.L. The biology of Sclerotinia sclerotiorum,S. trifoliorum, andS. minor with emphasis on specific nomenclature. Bot. Rev. 1980, 46, 101–165. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, K.; Li, M.; Hu, H.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. SsAGM1-mediated uridine diphosphate-N-acetylglucosamine synthesis is essential for development, stress response, and pathogenicity of Sclerotinia sclerotiorum. Front.Microbiol. 2022, 13, 938784. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Buchenauer, H.; Han, Q.; Zhang, X.; Kang, Z. Ultrastructural and cytochemical studies on the infection process of Sclerotinia sclerotiorum in oilseed rape. J. Plant Dis. Prot. 2008, 115, 9–16. [Google Scholar] [CrossRef]
- Uloth, M.B.; Clode, P.L.; You, M.P.; Barbetti, M.J. Attack modes and defence reactions in pathosystems involving Sclerotinia sclerotiorum, Brassica carinata, B. juncea and B. napus. Ann. Bot. 2015, 117, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Rollins, J.A. Mechanisms of broad host range necrotrophic pathogenesis in Sclerotinia sclerotiorum. Phytopathology. 2018, 108, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Li, J.; Yu, B.; Liu, L.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. Transcription factor SsSte12 was involved in mycelium growth and development in Sclerotinia sclerotiorum. Front.Microbiol. 2018, 9, 2476. [Google Scholar] [CrossRef] [PubMed]
- Cong, J.; Xiao, K.; Jiao, W.; Zhang, C.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. The coupling between cell wall integrity mediated by MAPK kinases and SsFkh1 is involved in sclerotia formation and pathogenicity of Sclerotinia sclerotiorum. Front.Microbiol. 2022, 13, 816091. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Lai, W.; Huang, K.; Li, Y.; Wang, Z.; Chen, X.; Wang, A. SsATG8 and SsNBR1 mediated-autophagy is required for fungal development, proteasomal stress response and virulence in Sclerotinia sclerotiorum. Fungal Genet. Biol. 2021, 157, 103632. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Yu, H.; Cong, J.; Xiao, K.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. Transcription factor SsFoxE3 activating SsAtg8 is critical for sclerotia, compound appressoria formation, and pathogenicity in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2022, 23, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Yu, H.; Chen, X.; Xiao, K.; Jia, D.; Wang, F.; Zhang, Y.; Pan, H. The SsAtg1 activating autophagy is required for sclerotia formation and pathogenicity in Sclerotinia sclerotiorum. J. Fungi 2022, 8, 1314. [Google Scholar] [CrossRef]
- Jiao, W.; Ding, W.; Rollins, J.A.; Liu, J.; Zhang, Y.; Zhang, X.; Pan, H. Cross-talk and multiple control of Target of Rapamycin (TOR) in Sclerotinia sclerotiorum. Microbiol. Spectrum 2023, 11, e0001323. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, J.; Zhu, W.; Yang, Y.; Mei, J.; Bi, C.; Qian, W.; Qing, L.; Tan, W. Ss-Rhs1, a secretory Rhs repeat-containing protein, is required for the virulence of Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 18, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Q.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. Ssams2, a gene encoding GATA transcription factor, is required for appressoria formation and chromosome segregation in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 3031. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Li, L.; Liu, J.; Zhang, Y.; Pan, H. Proteomics analysis of SsNsd1-mediated compound appressoria formation in Sclerotinia sclerotiorum. Int. J. Mol. Sci. 2018, 19, 2946. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, Y.; Yan, B.; Liao, H.; Dong, M.; Meng, X.; Wan, H.; Qian, W. Host-induced gene silencing of a multifunction gene Sscnd1 enhances plant resistance against Sclerotinia sclerotiorum. Front.Microbiol. 2021, 12, 693334. [Google Scholar] [CrossRef] [PubMed]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Monazzah, M.; Rabiei, Z.; Enferadi, S.T. The effect of oxalic acid, the pathogenicity factor of Sclerotinia Sclerotiorum on the two susceptible and moderately resistant lines of sunflower. Iran. J. Biotechnol. 2018, 16, e1832. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog. 2011, 7, e1002107. [Google Scholar] [CrossRef]
- Wei, W.; Pierre-Pierre, N.; Peng, H.; Ellur, V.; Vandemark, G.J.; Chen, W. The D-galacturonic acid catabolic pathway genes differentially regulate virulence and salinity response in Sclerotinia sclerotiorum. Fungal Genet. Biol. 2020, 145, 103482. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yajima, W.; Das, D.; Suresh, M.R.; Kav, N.N. Isolation, expression and characterization of two single-chain variable fragment antibodies against an endo-polygalacturonase secreted by Sclerotinia sclerotiorum. Protein Expr Purif. 2009, 64, 237–243. [Google Scholar] [CrossRef]
- Fan, H.; Yang, W.; Nie, J.; Zhang, W.; Wu, J.; Wu, D.; Wang, Y. A novel effector protein SsERP1 inhibits plant ethylene signaling to promote Sclerotinia sclerotiorum infection. J. Fungi 2021, 7. [Google Scholar] [CrossRef]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Ma, L.; Coutu, C.; Bekkaoui, D.; Hegedus, D.D. Receptor-like kinases BAK1 and SOBIR1 are required for necrotizing activity of a novel group of Sclerotinia sclerotiorum necrosis-inducing effectors. Front. Plant Sci. 2020, 11, 1021. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Li, W.; Tran, L.P.; Mostofa, M.G. Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the pathogenomic features of a global pathogen. Cells 2023, 12. [Google Scholar] [CrossRef]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in the Sclerotinia sclerotiorum transcriptome during infection of Brassica napus. BMC Genomics 2017, 18, 266. [Google Scholar] [CrossRef]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef]
- Zhou, F.; Zhang, X.L.; Li, J.L.; Zhu, F.X. Dimethachlon Resistance in Sclerotinia sclerotiorum in China. Plant Dis. 2014, 98, 1221–1226. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, Y.-P.; Chen, C.-J.; Zhou, M.-G. Detection of resistance in Sclerotinia sclerotiorum to carbendazim and dimethachlon in Jiangsu Province of China. Australas. Plant Pathol. 2014, 43, 307–312. [Google Scholar] [CrossRef]
- Donaldson, J.G. Multiple roles for Arf6: sorting, structuring, and signaling at the plasma membrane. J. Biol. Chem. 2003, 278, 41573–41576. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhou, T.; Chen, L.; Zheng, S.; Chen, S.; Zhang, D.; Li, G.; Wang, Z. Arf6 controls endocytosis and polarity during asexual development of Magnaporthe oryzae. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed]
- Gamara, J.; Davis, L.; Leong, A.Z.; Page, N.; Rollet-Labelle, E.; Zhao, C.; Hongu, T.; Funakoshi, Y.; Kanaho, Y.; Aoudji, F.; et al. Arf6 regulates energy metabolism in neutrophils. Free Radical Biol. Med. 2021, 172, 550–561. [Google Scholar] [CrossRef]
- Li, R.; Peng, C.; Zhang, X.; Wu, Y.; Pan, S.; Xiao, Y. Roles of Arf6 in cancer cell invasion, metastasis and proliferation. Life Sci 2017, 182, 80–84. [Google Scholar] [CrossRef]
- Schweitzer, J.K.; Sedgwick, A.E.; D'Souza-Schorey, C. ARF6-mediated endocytic recycling impacts cell movement, cell division and lipid homeostasis. Semin. Cell Dev. Biol. 2011, 22, 39–47. [Google Scholar] [CrossRef]
- Lee, S.C.; Schmidtke, S.N.; Dangott, L.J.; Shaw, B.D. Aspergillus nidulans ArfB plays a role in endocytosis and polarized growth. Eukaryot Cell. 2008, 7, 1278–1288. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S. Targeted gene replacement in fungi using a split-marker approach. Methods Mol. Biol. 2012, 835, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ao, K.; Tian, L.; Qiu, Y.; Huang, X.; Liu, X.; Hoy, R.; Zhang, Y.; Rashid, K.Y.; Xia, S.; et al. A forward genetic screen in Sclerotinia sclerotiorum revealed the transcriptional regulation of its sclerotial melanization pathway. Mol. Plant-Microbe Interact. 2022, 35, 244–256. [Google Scholar] [CrossRef]
- Yang, C.; Tang, L.; Qin, L.; Zhong, W.; Tang, X.; Gong, X.; Xie, W.; Li, Y.; Xia, S. mRNA turnover protein 4 is vital for fungal pathogenicity and response to oxidative stress in Sclerotinia sclerotiorum. Pathogens 2023, 12, 281. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Smith, D.F.Q.; Casadevall, A. The role of melanin in fungal pathogenesis for animal hosts. Curr. Top. Microbiol. Immunol. 2019, 422, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Meng, X.; Mo, C.; Wei, X.; Ma, A. Melanin of fungi: from classification to application. World J. Microbiol. Biotechnol. 2022, 38, 228. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Fu, Y.; Xie, J.; Li, B.; Chen, T.; Lin, Y.; Chen, W.; Jiang, D.; Cheng, J. Sclerotinia sclerotiorum SsCut1 modulates virulence and cutinase activity. J Fungi 2022, 8, 526. [Google Scholar] [CrossRef]
- Kim, K.S.; Min, J.Y.; Dickman, M.B. Oxalic acid is an elicitor of plant programmed cell death during Sclerotinia sclerotiorum disease development. Mol. Plant-Microbe Interact. 2008, 21, 605–612. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic and sequence analysis of SsArf6. (A) Phylogenetic analysis of Arf6 between different species. (B) The Arf6 gene features in F. oxysporum, F. graminearum, M. oryzae, S. sclerotiorum, B. cinerea, Z. tritici, and U. maydis illustrated schematically by GSDS2.0. (C) Multiple sequence alignment of FoArf6, FgArf6, PoArf6, SsArf6, BcArf6, ZtArf6, and UmArf6. The alignment results were visualized using Jalview clustal Colour Scheme.
Figure 1.
Phylogenetic and sequence analysis of SsArf6. (A) Phylogenetic analysis of Arf6 between different species. (B) The Arf6 gene features in F. oxysporum, F. graminearum, M. oryzae, S. sclerotiorum, B. cinerea, Z. tritici, and U. maydis illustrated schematically by GSDS2.0. (C) Multiple sequence alignment of FoArf6, FgArf6, PoArf6, SsArf6, BcArf6, ZtArf6, and UmArf6. The alignment results were visualized using Jalview clustal Colour Scheme.
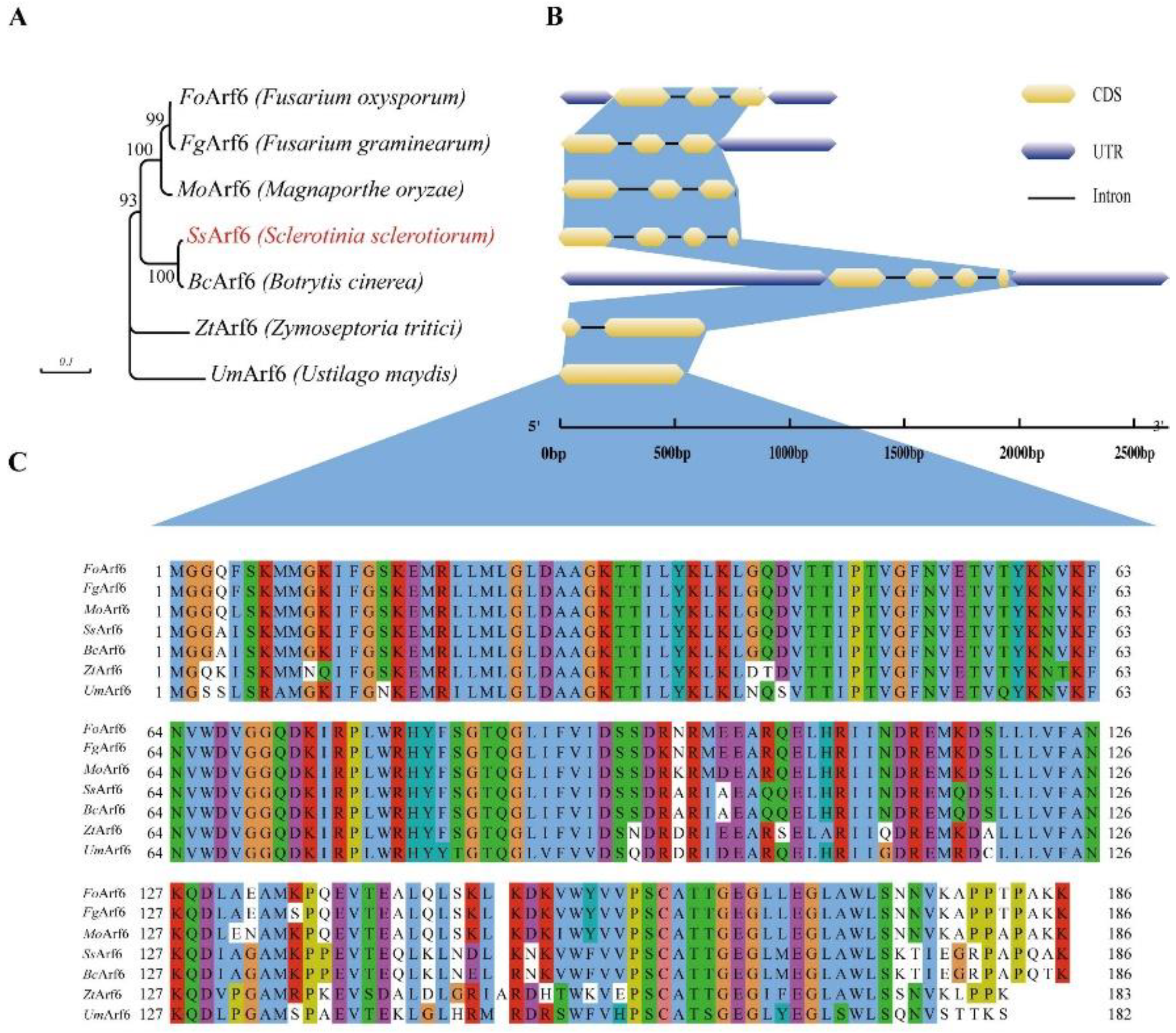
Figure 2.
SsArf6 contributes to mycelial development, melanin accumulation and sclerotia production in S. sclerotiorum. (A) The colony morphology of WT, ΔSsarf6, and SsArf6-C strains grown on PDA culture medium for 24h and 48h. (B) The mean length and growth rate of mycelia of WT, ΔSsArf6, and SsArf6-C strains 48 hours later. (C) Melanin accumulation of WT, ΔSsarf6, and SsArf6-C strains grown on PDA culture medium for 5d. (D) Branching patterns of mycelia of WT, ΔSsarf6, and SsArf6-C strains. (E) The colony morphology of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium for 8 days.(F, G) The number of sclerotia per plate. WT refers to the wild-type strain; ΔSsarf6, the knockout strain; and SsArf6-C, the complemented strain. The experiment was repeated three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between WT and knockout mutant or complemented strains was analyzed using the student’s t-test (* p < 0.05).
Figure 2.
SsArf6 contributes to mycelial development, melanin accumulation and sclerotia production in S. sclerotiorum. (A) The colony morphology of WT, ΔSsarf6, and SsArf6-C strains grown on PDA culture medium for 24h and 48h. (B) The mean length and growth rate of mycelia of WT, ΔSsArf6, and SsArf6-C strains 48 hours later. (C) Melanin accumulation of WT, ΔSsarf6, and SsArf6-C strains grown on PDA culture medium for 5d. (D) Branching patterns of mycelia of WT, ΔSsarf6, and SsArf6-C strains. (E) The colony morphology of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium for 8 days.(F, G) The number of sclerotia per plate. WT refers to the wild-type strain; ΔSsarf6, the knockout strain; and SsArf6-C, the complemented strain. The experiment was repeated three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between WT and knockout mutant or complemented strains was analyzed using the student’s t-test (* p < 0.05).
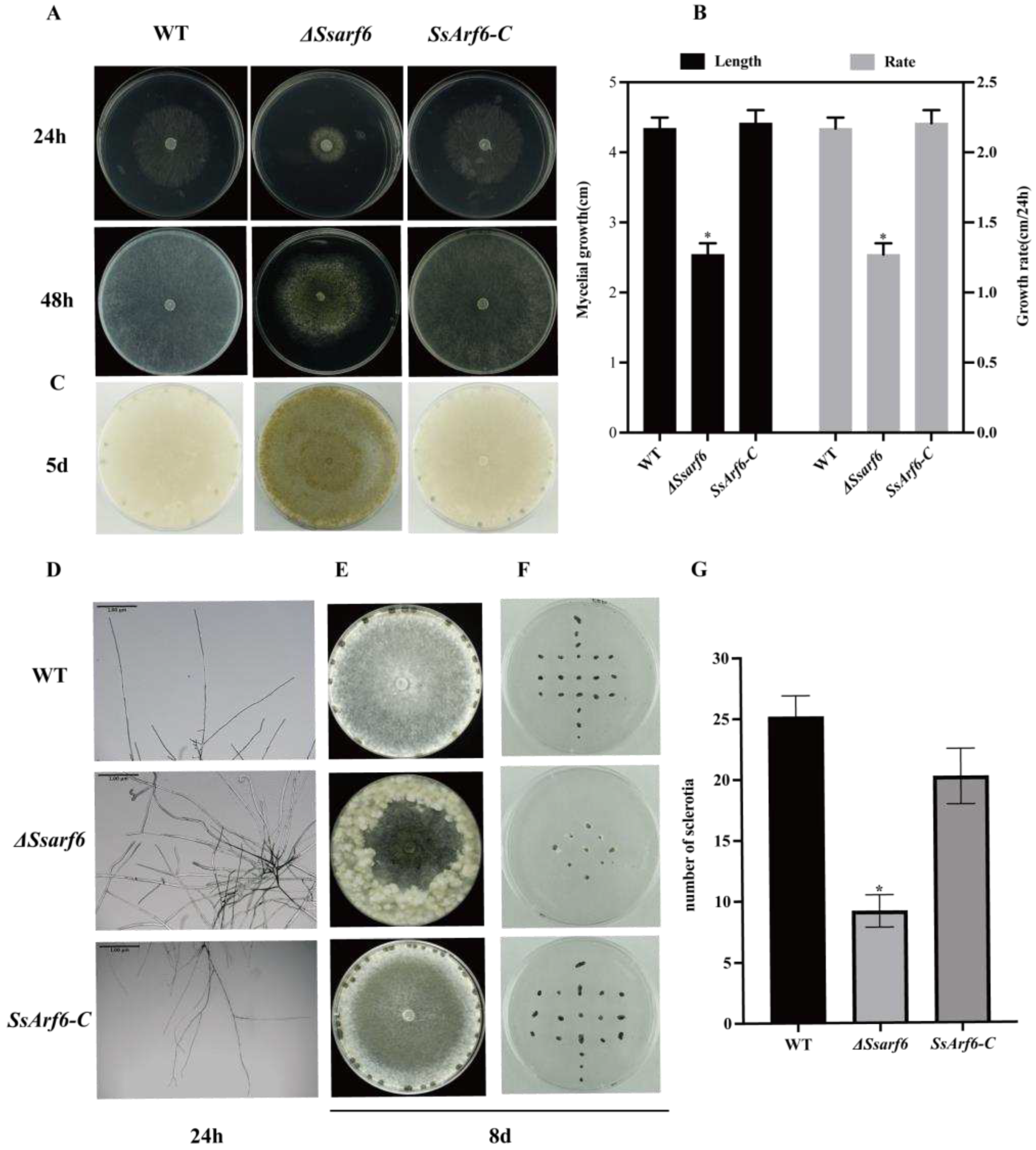
Figure 3.
SsArf6 contributes to the responses to osmotic stress in S. sclerotiorum. (A, B) The colony morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing different osmotic stressors, 0.5M NaCl, 0.5M KCl, 1M glucose and 1M sorbitol, for 48h. (C, D) The colony morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing different cell wall inhibitors, 0.5mg/mL Congo Red (CR) and 0.02% Sodium Dodecyl Sulfate (SDS) for 48h. ΔSsarf6 represents the knockout strain; SsArf6-C, the complemented strain. The experiment was conducted three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between the WT and knockout mutant or complemented strains was analyzed using the student’s t-test (** p < 0.01).
Figure 3.
SsArf6 contributes to the responses to osmotic stress in S. sclerotiorum. (A, B) The colony morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing different osmotic stressors, 0.5M NaCl, 0.5M KCl, 1M glucose and 1M sorbitol, for 48h. (C, D) The colony morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing different cell wall inhibitors, 0.5mg/mL Congo Red (CR) and 0.02% Sodium Dodecyl Sulfate (SDS) for 48h. ΔSsarf6 represents the knockout strain; SsArf6-C, the complemented strain. The experiment was conducted three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between the WT and knockout mutant or complemented strains was analyzed using the student’s t-test (** p < 0.01).
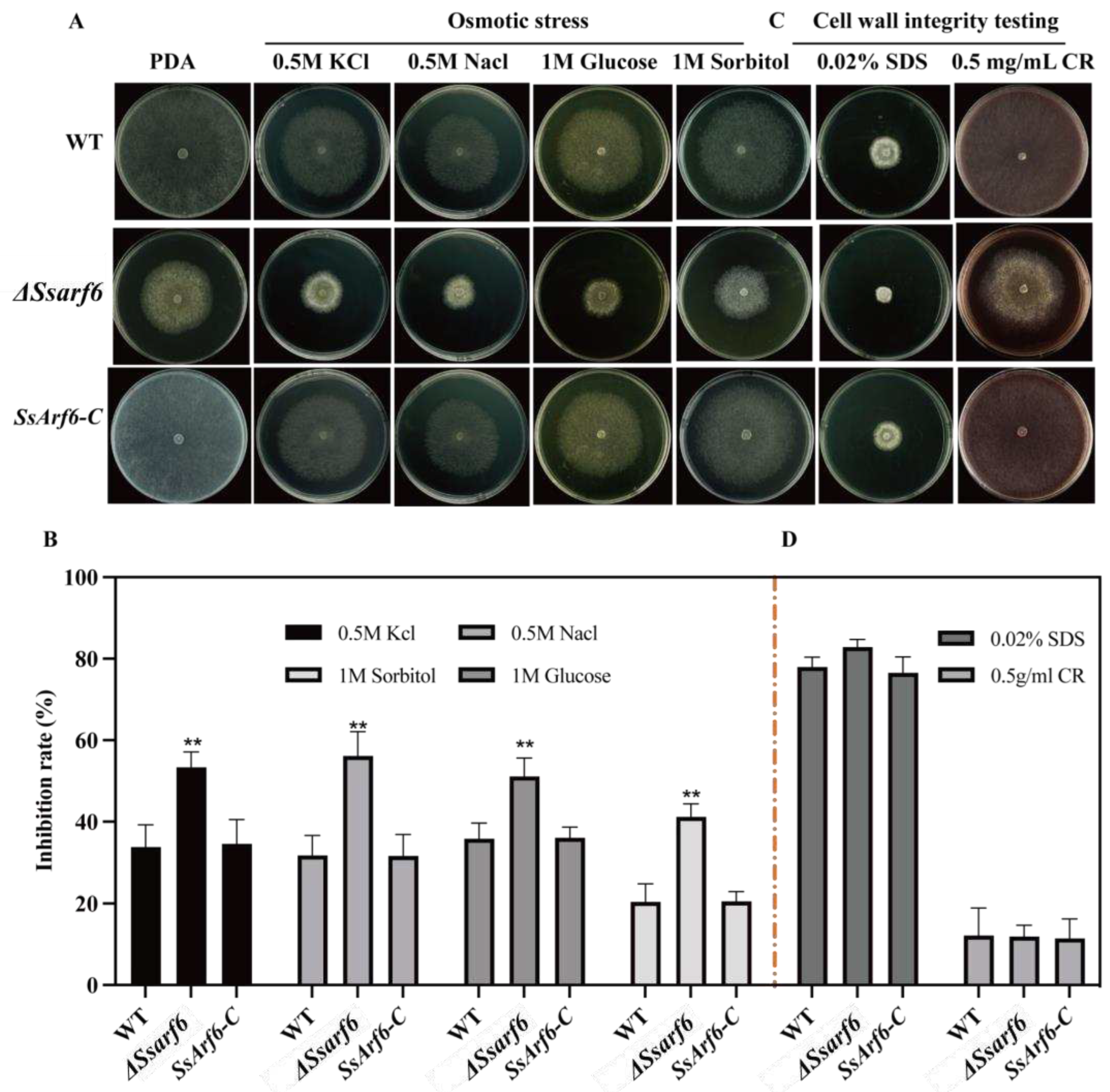
Figure 4.
SsArf6 negatively regulates the resistance of S. sclerotiorum to hydrogen peroxide. (A, B) The mycelium morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing 5mM, 10mM, and 15mM H2O2 for 48h. The experiment was conducted three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between the WT and knockout mutant or complemented strains was analyzed using the student’s t-test (** p < 0.01).
Figure 4.
SsArf6 negatively regulates the resistance of S. sclerotiorum to hydrogen peroxide. (A, B) The mycelium morphology and inhibition rate of WT, ΔSsarf6, and SsArf6-C strains grown on PDA medium containing 5mM, 10mM, and 15mM H2O2 for 48h. The experiment was conducted three times with similar results. Error bars represent the standard deviation (SD). The statistical significance between the WT and knockout mutant or complemented strains was analyzed using the student’s t-test (** p < 0.01).
Figure 5.
SsArf6 is involved in compound appressoria formation. (A) The Appressorium morphology after 48 hours on glass slides by WT, ΔSsarf6, and SsArf6-C strains (B) Appressorium formed after 24 hours on glass slides by WT, ΔSsarf6, and SsArf6-C strains observed under an optical microscope. Bar = 1 µm. (C) Invasion assay of WT, ΔSsarf6, and SsArf6-C on onion epidermis. Invasion mycelial were stained by trypan blue. (D) Analysis of oxalic acid secretion ability of WT, ΔSsarf6, and SsArf6-C strains, with colony morphology after 48h cultivation on PDA medium containing bromophenol blue. When the pH of Bromophenol Blue is greater than or equal to 3.0 and less than 4.6, it appears yellow, and when the pH is greater than or equal to 4.6, it appears blue. WT represents the wild-type strain; ΔSsarf6, the knockout strain; SsArf6-C, the complemented strain.
Figure 5.
SsArf6 is involved in compound appressoria formation. (A) The Appressorium morphology after 48 hours on glass slides by WT, ΔSsarf6, and SsArf6-C strains (B) Appressorium formed after 24 hours on glass slides by WT, ΔSsarf6, and SsArf6-C strains observed under an optical microscope. Bar = 1 µm. (C) Invasion assay of WT, ΔSsarf6, and SsArf6-C on onion epidermis. Invasion mycelial were stained by trypan blue. (D) Analysis of oxalic acid secretion ability of WT, ΔSsarf6, and SsArf6-C strains, with colony morphology after 48h cultivation on PDA medium containing bromophenol blue. When the pH of Bromophenol Blue is greater than or equal to 3.0 and less than 4.6, it appears yellow, and when the pH is greater than or equal to 4.6, it appears blue. WT represents the wild-type strain; ΔSsarf6, the knockout strain; SsArf6-C, the complemented strain.
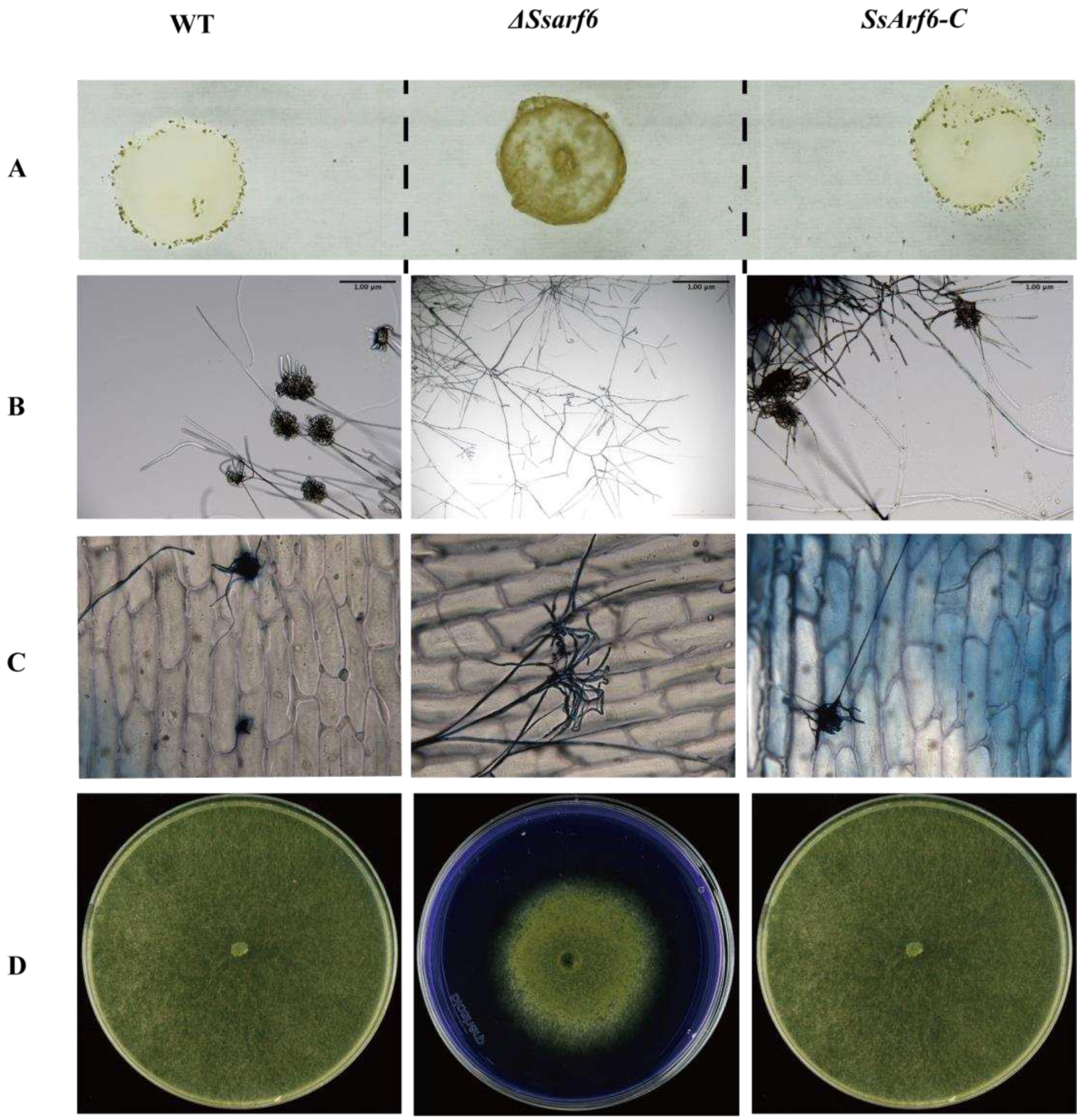
Figure 6.
SsArf6 contributes to the virulence of S. sclerotiorum. (A) Inoculated lesions of WT, ΔSsarf6 and Ssarf6-C of detached leaves of A. thaliana, B. napus and N. benthamiana. (B) Lesion areas of WT, ΔSsarf6 and SsArf6-C on leaves of A. thaliana, B. napus and N. benthamiana. (C) Inoculated lesions of ΔSsarf6 detached unwounded/wounded leaves of N. benthamiana. (D) Lesion areas of ΔSsarf6 on wounded/unwounded leaves of N. benthamiana. Data were recorded at 36 h post inoculation. Bar = 10 mm. Image J was used to analyze the lesion area. The experiment was repeated three times with similar results. Error bars represent SD. The statistical significance between WT and knockout mutant strain or complemented strains was analyzed using Student's t-test (**p<0.01). WT represents the wild-type strain; ΔSsarf6, the knockout strain; SsArf6-C, the complemented strain.
Figure 6.
SsArf6 contributes to the virulence of S. sclerotiorum. (A) Inoculated lesions of WT, ΔSsarf6 and Ssarf6-C of detached leaves of A. thaliana, B. napus and N. benthamiana. (B) Lesion areas of WT, ΔSsarf6 and SsArf6-C on leaves of A. thaliana, B. napus and N. benthamiana. (C) Inoculated lesions of ΔSsarf6 detached unwounded/wounded leaves of N. benthamiana. (D) Lesion areas of ΔSsarf6 on wounded/unwounded leaves of N. benthamiana. Data were recorded at 36 h post inoculation. Bar = 10 mm. Image J was used to analyze the lesion area. The experiment was repeated three times with similar results. Error bars represent SD. The statistical significance between WT and knockout mutant strain or complemented strains was analyzed using Student's t-test (**p<0.01). WT represents the wild-type strain; ΔSsarf6, the knockout strain; SsArf6-C, the complemented strain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



