
Submitted:
20 November 2023
Posted:
21 November 2023
You are already at the latest version
Abstract
The leading role of Italy in the cultivation of durum wheat stimulated intense breeding activities in the country from the beginning of the 20th century, much earlier than any other country involved in durum wheat production. Older, genetically more heterogeneous landraces became replaced with new highly productive, superior quality varieties, and this led an inevitable reduction in overall genetic diversity among new cultivars, which makes the genetic variability of the old cultivars preserved particularly valuable and important. The aim of this paper was to assist future breeding programmes by providing a detailed description of the history of durum wheat breeding in Italy and of the changes in yield, quality and related traits that subsequently occurred, starting from the most diffuse landraces present between 1900 and 1920 up until the present day. The parallel evolution of breeding techniques, breeding goals and agricultural systems in this period is also described, and some future breeding goals suggested. In the current context of climate change and of rapidly mutating pathogen populations, preserving the yield level with the continuous introduction of new cultivars by exploiting the reservoir of largely unused genetic variation stored in old cultivars and landraces could be as important as increasing grain yield and quality.
Keywords:
Triticum durum
; breeding
; grain yield
; grain quality
; physiological traits
1. Introduction
Durum wheat (Triticum turgidum subsp. durum (Desf.) Husn) is grown on around 17 million ha worldwide, primarily in the Mediterranean Basin, Canada and the United States (Eurostat). Its global production for 2020 was assessed at 33.6 million tons, and approximately half of this came from the Mediterranean Basin [1]. Within Europe, Italy is the front-runner in durum wheat production, with a cultivation area averaging 1.23 million ha (EUROSTAT, average for the last three years), equivalent to 53% of the EU’s surface area dedicated to durum wheat and 57% of the EU’s overall production.
The first ever durum wheat breeding program was launched in Italy back in the early 1900s, an unsurprising fact considering the economic significance of the pasta industry and the country’s leading position in Europe in terms of durum wheat production [2]. The nation’s careful and prolonged breeding work has generated a valuable germplasm that is widely cultivated both nationally and internationally [3].
The aims of this review are three-fold: i. to describe the rich and interesting history of durum wheat breeding in Italy, highlighting the strong interplay between breeding and the evolution of agricultural systems; ii. to analyze the effects of breeding on grain yield, yield-related traits and grain quality; and iii. to propose future avenues and directions for breeders to pursue.
2. Varietal evolution
2.1. Landraces and local populations (1800-1920)
Before Mendel’s discoveries in the 19th century on the principles of genetic inheritance and before Strampelli’s work at the turn of the century on genetic improvement (initially on bread wheat and subsequently on durum), durum wheat cultivation in Italy was mostly based on indigenous landraces that had evolved and adapted to the agroecosystem of Southern Italy (primarily the islands, Sicily and Sardinia). They came about through natural selection and/or unconscious selection by farmers [4]. In 1927, Emanuele De Cillis, in his book “I Grani d’Italia", described the most widespread landraces of wheat at that time [5] (Table 1).
Most of these landraces belonged to the mediterraneum type [6,7,8] (Figure 1), characterized by early vigour, tall plants (up to 180 cm) prone to lodging, late flowering and high grain filling rates. After the war between Italy and the Ottoman Empire in 1912, North African (Libyan in particular), Syrian and Palestinian landraces (e.g., “Aziziah” and “Eiti”) arrived in Italy, thus contributing to the maintenance of a large genetic base. Many of them belonged to the so-called syriacum type, characterized by profuse tillering, shorter plants (below 120 cm) and earlier anthesis than the mediterraneum types due to the different climatic conditions of their respective areas of origin [7,8].
These landraces formed an important part of a self-supporting rural economy based on complex agricultural systems, where animals provided labour and manure for fertilization, and rotations with legumes provided animal feed whilst increasing soil nitrogen. Fallowing was also common, particularly on the islands.
2.2. First period: genealogical selection from local and exotic populations (1920–1950)
In the 30 years spanning 1920 to 1950, Italy was the first country within the Mediterranean basin to begin ‘conscious’ breeding—primarily focused on the genealogical selection of pure lines derived from previously cultivated local varieties. The only exception was “Garigliano”, a pure line ‘created’ by Strampelli, who, for the first time, used controlled hybridization to create genetic variability. These more uniform new cultivars gradually replaced the original landraces in Italy, whereas other countries continued to grow durum landraces until the so-called "Green Revolution” in the late 60s [9].
The most successful cultivar of this period was "Senatore Cappelli" (from here on referred to as “Cappelli”), released by Strampelli in 1915, and subsequently cultivated across up to 60% of the area dedicated to durum wheat in Italy. Cultivation of the “Cappelli” variety also spread to other Mediterranean countries, namely Spain and Turkey. Although it is generally assumed that “Cappelli” originated from the North African population “Jean Rhetifah”, recent studies highlighted genetic distance between “Cappelli” and “Jean Rhetifah” [10,11,12], and instead confirmed its genetic similarity to the Tunisian “Bidi”.
“Cappelli” was appreciated for its high grain yield (2.6 t ha-1 on a plot basis according to Strampelli), being 33% higher than the preceding landraces according to Maliani [13], for its adaptability, high number of kernels per spike and spikelet, and excellent semolina quality [14]. It was not the only cultivar developed during those years, and Strampelli was not the only active breeder, others being De Cillis, Conti, Barbieri and Casale (Table 2).
Despite its undisputed success, “Cappelli”—a typical component of the mediterranean group and hence tall and late—faced challenges such as rust and lodging susceptibility, which were long-"ignored" partly because its low sowing density and low soil fertility requirement did not exacerbate the lodging problem. Moreover the distribution of nitrogen fertilizers was quite limited (4.0 kg ha-1 of N2, 7.8 of P2O5 and 0.5 of K2O, referring to the total agricultural area of about 21 million ha) [15]—low compared with contemporary fertilization rates.
During this period, the area dedicated to durum wheat cultivation in Italy also experienced significant variation in surface area (from 1.2 million ha in the period 1926–1930 to 1.3 million ha in the period 1946–50, ISTAT data) and geographic distribution. Initially restrained to the islands, South and Central Italy, in the late 1940s durum wheat cultivation expanded to other regions within Central Italy, namely Tuscany, Marche and Umbria [16].
With the “Battle for Wheat”, the average yield raised from the 0.9 t ha-1 of the 1920s to 1.2 t ha-1 in the latter half of the 1930s. However, during the 1940s, yields dropped to 1920s levels due to the impact of the war. This suggests that the agronomic component may have been more decisive than the genetic component in determining yields during that period [17]. After the war, the insufficiency of durum wheat production in Italy in both quantitative and qualitative terms also occurred because it was primarily concentrated in the southern regions, often on infertile and resource-limited soils. The limited yields did not encourage the necessary investment. Thus, to ensure an adequate supply, milling and pasta industries blended locally grown durum wheat with durum wheat from other parts of the world, such as the United States, Canada and Argentina (e.g., “Candeal-Taganrog”, a selection of the original Russian population). These imports exhibited characteristics such as a yellowish colour and a strong gluten [18].
2.3. Second period: intra- and inter-specific hybridization and mutagenesis (1950–1973)
Between the 1950s and 1960s, the genetic improvement in durum wheat in Italy was governed by the crossbreeding of lines from the mediterranean group, mainly from the North Africa (like “Cappelli”) and the syriacum group (“Aziziah”, “Eiti”, “Sinai”, “Tripolino”), with the aim of generating variability for subsequent selection. Nazareno Strampelli had already proposed this breeding technique, creating the “Garigliano” cultivar as early as 1927 through the crossbreeding of “Cappelli” with “Tripolino” [19].
Many of the new cultivars released during this period (“Capeiti 8”, “Patrizio 6” [20], “Casale 92”, “Sincape 9”, “Grifoni 235”) were based on “Cappelli” (Table 3), meaning that this cultivar appears in the pedigree of almost all durum wheat cultivars bred in Italy and elsewhere [21].
The crossbreeding between mediterranean and syriacum types reduced plant height to less than 120 cm, and thus the lodging incidence, and brought anthesis forward [22]. This marked the beginning of a gradual genetic improvement to enhance earliness, making Italian varieties generally earlier than those grown in other Mediterranean areas [23].
The best results were obtained with “Capeiti 8” and “Patrizio 6”, which outperformed “Cappelli” in yield [15], earliness and lodging resistance, bringing about the decline of “Cappelli” by the mid-1960s [7]. The reduction in plant height, initially appreciated solely for its positive effect on lodging, also led to an initial, significant increase in the harvest index (HI), a dominant theme in durum wheat genetic improvement, from about 0.32 in “Cappelli” to about 0.40 in “Capeiti 8” [24,25].
On the other hand, both the milling and pasta-making qualities of “Capeiti 8” and “Patrizio 6” were considerably worse than those of “Cappelli” [26], in part because of the expansion of durum wheat cultivation into less suitable areas compared with those where “Cappelli” was grown [27]. In addition to a lower grain weight [22,24,25], the new cultivars exhibited deficiencies in the plasticity of dough [16], as well as greater lipoxidase activity[7], negatively impacting the amber coloration of semolina.
Durum breeding was very intense in Italy in the following years up until 1973. “Capeiti 8” was widely used as a parent for intraspecific crosses, producing cultivars, including “Maristella”, “Nuragus” and “Ichnusa” [28] (Table 3) in Sardinia, and “Hymera” and “Trinakria” in Sicily [29]. “Maristella” and “Trinakria” were known for their good quality, although not as good as Cappelli. “Appulo”, selected by Dionigi in Apulia, was the most successful cultivar of this group. After being registered in the Varietal Register in 1973, the following year it ranked among the top four most widely cultivated Italian cultivars together with “Capeiti 8”, “Patrizio” and “Cappelli” (ISTAT). Using results from about 200 field trials, Rivoira [30] estimated a 12% increase in yield with the transition from “Cappelli” to “Capeiti 8”, and another 7% from “Capeiti 8” to “Appulo”, “Maristella” and “Isa”.
Interspecific hybridization with other Triticum species, like T. dicoccum, T. turgidum and T. sphaerococcum, was also common during this period to improve resistance to pathogens, cold and lodging, and to increase spike fertility. These efforts resulted in the creation of varieties such as “Lambro” and “Belfuggito”, characterized by high yield and cold resistance, but not widely cultivated [31].
In the same years, mutagenesis was applied to durum breeding by the groups led by D’Amato and Scarascia-Mugnozza at the CNEN (National Committee for Nuclear Energy, now ENEA) to obtain the cultivars making up the Castel- group: “Castelporziano” and “Castelfusano” (a “Cappelli” mutant), “Casteldelmonte” (a “Grifoni” mutant) and “Castelnuovo” (a “Garigliano” mutant) [25,32,33]. “Castelporziano” was more lodging-resistant than “Cappelli” thanks to a single partially dominant gene capable of reducing height by 34% [34].
During this period, the cultivated area for durum wheat in Italy increased from 1.4 to 1.6 million ha due to its expansion across the south as well as in more northern regions such as Marche, Umbria, Lombardy, Veneto and Emilia Romagna. Altogether, the more favourable pedoclimatic conditions of these regions, the improved cultivars, and the increase in nitrogen fertilization, made possible the reduction in plant height, boosted national grain yields from 1.12 to 1.9 t ha-1 (ISTAT).
2.4. Final period: semi-dwarf cultivars (1974–today)
The last period of durum breeding began with the production of the first semi-dwarf cultivars, which continue to be grown today. The success of dwarfing genes, responsible for a reduction in height and an increase HI and ear fertility at similar biomass levels, arose from a strong synergy between breeding and management: these new cultivars were able to exploit the high nitrogen rates made available by the contemporary replacement of animal waste with industrially produced ammonia as the primary source of nitrogen; their lower competitiveness towards weeds was compensated through the use of herbicides; and they were better suited to the increasing levels of mechanization in farming practices. Interestingly, the scheme followed by Norman Borlaug, the father of the Green Revolution, was practically the same as Strampelli’s, and benefited from the preceding diffusion of Strampelli’s varieties all over the world. What was different was the organization level (Borlaug worked at CIMMYT [35]) and the reference agro-ecosystems, which were not yet prepared to exploit the potential of semi-dwarf cultivars at the time of Strampelli.
The semi-dwarf Rht-B1b allele, the central protagonist of the 1960s’ Green Revolution of bread wheat, appeared for the first time in commercial durum wheat cultivars in the 1970s as the outcome of the breeding work carried out by Vallega and Zitelli, ENEA (Italian National Agency for New Technologies, Energy, and Sustainable Economic Development), the Experimental Institute for Cereal Research and the Experimental-Agricultural Regional Center in Sardegna. These first semi-dwarf cultivars were “Valgerardo”, “Valnova”, “Creso” and “Karel” [36] (Table 4). The Japanese bread wheat “Norin 10” was the donor of the Rht alleles, and durum wheats from Central and North America were the donors of various resistances to pathogens. “Creso” originated from a cross between a segregating population from CIMMYT (Centro Internacional de Mejoramiento de Maiz Y Trigo, Mexico) with Cp B 144, a Cappelli mutant. Registered in 1974, “Creso” spread across Italy quickly, especially in the central and northern regions due to its lateness, gradually replacing “Capeiti 8” and “Appulo”. Its diffusion reached a peak in 1987, when it was responsible for 43% of national durum wheat production (ISTAT). It was one of the most longaeval durum wheat cultivars, remaining within the top 10 Italian cultivars until 2005, i.e. for about 30 years.
The take up of “Val” group varieties (“Valgerardo”, “Valnova”, “Valforte”, etc.) was limited, primarily due to their inadequate commercialization rather than to genuine genetic inferiority [18]. These cultivars did not spread as widely as “Creso”, in spite of being earlier than the latter and hence more suited for southern regions. During this period, the cultivar “Karel” (“Mex” x “198” x “Maristella”), established in Sardinia at the CRAS (Centro Regionale Agrario Sperimentale) by Deidda in 1979, exhibited higher production levels and production stability compared with “Creso”, albeit with smaller kernels and medium-low pasta-making quality. Results from a three-year varietal comparison trial conducted across 33 locations in central and southern Italy [37] found the earliness of Karel to be similar to that of “Appulo”, “Maristella” and “Trinakria”, but it outperformed other dwarf and semi-dwarf cultivars in production levels (averaging 4.5 t ha-1 over three years and across all locations), along with “Valforte” (4.3 t ha-1) and “Valgerardo” (4.1 t ha-1).
The productivity enhancements achieved with these new cultivars compared with “Capeiti 8” varied from 21 to 33% [37]. Besides the height variations, studies reported a higher ratio of useful to total tillering and greater synchrony in the development of secondary tillers compared with that for primary tillers [30]. Overall, these new durum wheat cultivars achieved production levels comparable to those of bread wheat, while maintaining the high technological quality required by the pasta industry.
The rapid evolution of durum breeding led, in 1974, to the foundation of the National Network for Comparison of Durum Wheat Varieties, coordinated by the Research Unit for the Quality Enhancement of Cereals (CREA) in Rome, which aimed to orient farmers in their cultivar choice. This network is still active and organizes 50–60 variety comparison trials each year across six areas encompassing the different environmental conditions of Italy, with the scope of comparing new cultivars against the most cultivated ones. The parameters evaluated include data on heading date, plant height, grain yield and yield components, quality and other agronomic traits, providing a useful historical dataset with which to analyze the evolution of durum wheat productivity.
New legal regulations accompanied the entering of semi-dwarf wheats into the panorama of durum wheat cultivars, such as Italian decree n. 580/67, which encouraged the use of durum for “pasta pureness”, regulation (EEC) n. 1143/76, which established a community support system for various types of cereals including durum wheat, Italian decree n. 1096/71 "Discipline of seed activity", which established Variety Registers, the registration of cultivars and the official certification of seeds as preliminary conditions for their commerce.
The success of the recently established varieties and the enactment of these new legal regulations spurred increased investments in durum wheat improvement and ultimately led to a decline in the contribution of public breeders in varietal development, which were progressively substituted by private seed companies (Table 4). This change made the marketing and commercialization activities carried out by seed companies crucial for the success of a cultivar, which did not always mirror its actual agronomic value. As a result of this intense breeding work, the entire available gene pool for tetraploid wheats underwent recombination, and a plethora of highly productive new cultivars incorporating novel germplasm, primarily sourced from CIMMYT were registered on the national register [38,39,40,41].
Breeding techniques still mostly rely on the conventional schemes adopted for autogamous species, i.e., the creation of genetic variability through controlled hybridization and the subsequent selection of pure lines. Marker-assisted selection has been a useful aid since 1990, particularly for the so-called “breeding by design” (involving the assembling of specific alleles necessary to obtain a previously defined “ideotype”), although it is mostly effective when “qualitative” traits controlled by just a single or few genes are involved (e.g. disease resistances, grain quality or development traits).
Breeding techniques still mostly rely on the conventional schemes adopted for autogamous species, i.e., the creation of genetic variability through controlled hybridization and the subsequent selection of pure lines. Marker-assisted selection has been a useful aid since 1990, particularly for the so-called “breeding by design” (involving the assembling of specific alleles necessary to obtain a previously defined “ideotype”), although it is mostly effective when “qualitative” traits controlled by just a single or few genes are involved (e.g. disease resistances, grain quality or development traits).
The adoption of new cultivars did not keep pace with the rhythm of appearance of new cultivars, as demonstrated by the dynamics of seed certification (Table 5). For example, “Duilio”, one of the most used cultivars until 2010, reached its maximum diffusion in 1993 despite being registered nearly a decade earlier in 1984; “Simeto”, registered four years later in 1988, reached its maximum diffusion (at 33%, expressed as the percentage of the seed used considering the top ten cultivars) in 1996. Among the cultivars registered in the 1990s, “Colosseo”, “Iride”, “Ciccio” and “Claudio” were the most diffuse. Apart from “Colosseo” (which became moderately widespread in the same year as its registration and reached its maximum diffusion just three years later at 10%,), “Ciccio”, “Iride” and “Claudio” only appeared 3–6 years after their respective registrations. “Iride” was the most widespread, peaking at 13.2% in 2008, i.e., 12 years after being registered.
In 2017, four years after being registered in the Varietal Register, “Antalis” ascended to the fourth position among the most produced seeds. This success reached its zenith in 2020 when “Antalis” claimed the top spot as the most produced cultivar. “Simeto”, “Iride”, and “Claudio”, each with a registration history spanning over two decades in the Varietal Register, consistently maintained prominent positions among the most diffused cultivars until 2020. Overall, the varietal landscape seems to be heading towards greater diversification, considering that the top ten cultivars for seed production represented a whopping 84% of total seeds produced in 1992, but only 58% in 2010 and 46% in 2018 [42].
The great diffusion of “Antalis” was justified by its greater productivity in the varietal comparisons carried out from 2015 to 2021, together with “Ramirez”, “Monastir”, “Claudio” and “Kanakis.
The steady rise in the number of cultivars being registered is testament to this intense breeding activity: 58 cultivars in 1981 [30], 134 in 2002, 174 in 2011, and an enormous 327 in 2022; thus an average of 14 new cultivars per year over the last decade. Alongside the new varieties, several older varieties are still present, including “Cappelli”, “Castelporziano”, “Appulo”, “Valnova” and “Creso”, all registered before 1975, as well as a group of 25 older constitution registered as “varietà da conservazione”, including some of the pure lines diffused earlier on (“Bidì”, “Tripolino”, “Ruscia”, “Capeiti”, etc.). Cultivars imported from other countries, mainly France, appeared in the Italian Register as early as 1995, and their presence has increased in recent years. Indeed, one of the most diffuse cultivars in the last decade, “Antalis”, was developed in France.
Grain yields showed a continuous and almost linear increase from the introduction of semi-dwarf cultivars until 2020, an increase that already began as early as in the second half of the 1960s (Figure 2). Although lower than the 96% average increase recorded between 1926–30 and 1976–80, when semi-dwarf cultivars were first cultivated, the 76% average increase observed from 1976–80 to 2016–20 is still relevant. On the other hand, it cannot be attributed to varietal evolution only, because ISTAT data refer to farm or ‘actual’ yields, and hence they reflect the combined effect of management, environment, cultivar and geographic distribution. Sowing rates of 350–400 viable seeds m-2 and fertilization rates of 100–180 kg of nitrogen ha-1 are common for modern cultivars, compared with 200 seeds m-2 and 40–60 kg of nitrogen ha-1 for the old cultivars.
The cultivation area assigned to durum wheat continued to grow after the appearance of semi-dwarf cultivars in parallel with the increase in yields, reaching a peak of 1.8 million ha in the period 1986–1990. A progressive decline in the area invested in durum wheat cultivation in Italy was observed after 1990, despite the MacSharry Reform (1765/92), which introduced an additional payment to durum wheat producers, compensating them for income loss due to aligning the price of durum wheat with other cereals. Unfortunately, the MacSharry Reform also led to a deterioration in farming practices, which was responsible for the deterioration in the quality of European durum wheat [43]. Starting from 1995, pressures by seed producer lobbies led the Ministry of Agriculture to impose the use of 100% certified seed to access the additional aid for durum wheat production areas, thus increasing the seed renewal rate from 30% in the early 1990s to over 70% [44].
The reduction in the area dedicated to durum wheat cultivation after 2005 primarily affected regions traditionally suited for durum wheat cultivation, such as Southern Italy and the islands. Several factors contributed to this decrease, including the significant price volatility of durum wheat, with extended periods of low profitability often failing to cover production costs. The introduction of decoupling from the new European Union regulations, which no longer tied premium payments to crop selection, also had a significant impact. Puglia became the top-ranking region, providing 27% of the national surface dedicated to durum wheat, followed by Sicily (21%), Basilicata (9%), Marche (8%) and Emilia Romagna (6%) (mean of 2016–2020 data, ISTAT). This decrease in cultivated area resulted in a decrease in the national production of durum wheat, exacerbating the incapacity of Italy to meet the needs of the processing industry [45].
3. Evolution of grain yield and related traits
Grain yield is a complex, low heritable trait, affected by a number of physiological mechanisms. Although empirical selection for grain yield was effective in the past, improving plant types through physiological breeding [46] may prove to be the more effective approach nowadays, when the relative rates of yield increase are declining [47]. The premise for designing improved plant types is knowledge about well-characterized genetic resources to use in crossing strategies. An understanding of the changes in grain yield and related traits resulting from durum wheat breeding in Italy and a characterization of the genotypes involved can, therefore, help physiological breeding.
3.1. Grain yield
Evaluating the effect of breeding on grain yield is a difficult task because of the strong interaction between genotype, environmental conditions and management. Even when cultivars from different eras of breeding are grown side-by-side and, by consequence, subjected to the same environment, evaluation of the genotypic component can be biased by management, since old durum wheat cultivars cannot be grown at the same plant population density and with the same rates of nitrogen fertilization commonly adopted for modern semi-dwarf cultivars due to their susceptibility to lodging. Moreover, both the biotic and abiotic environment may change over the /time span of comparison, so that adaptation to these changes is also included in the apparent “breeding progress” [47,48].
Giunta et al., [49] compared cultivars from different eras of breeding in Sardinia with their pertaining sowing rate, two sowing dates (normal and late) and two nitrogen rates, one suitable for old and the other suitable for modern cultivars. A 37% increase in yield potential was calculated for a comparison of old cultivars (pure lines grown until 1950) with an intermediate group including cultivars obtained by crossing mediterraneum and syriacum germplasm, plus an additional 19% considering the introduction of Rht genes, corresponding to a mean increase of 23.9 kg ha-1 year-1. The greater productivity of modern cultivars was expressed at both N rates and sowing dates, although modern cultivars benefited more from the higher nitrogen supply. Similar results were obtained by De Vita et al. [50], who estimated an genetic yield gain of 19.9 kg ha−1 year−1 for cultivars covering the same historical period as Giunta et al. [49], grown under different fertilization rates and with vs without irrigation in Southern Italy. Grain yield variation between cultivars ranged from 3.26 to 5.43 t ha-1, i.e. the same range observed by Giunta et al. [49] between old and modern cultivars. In both experiments the genetic gain was most clearly associated with a higher kernel number m−2 [49,50].
These comparisons, although informative, mainly reflect the effect of Rht introgression, but they cannot quantify effects of breeding on the grain yield of the semi-dwarf cultivars released in the last 50 years. To the best of our knowledge, no side-by-side comparison of durum wheat semi-dwarf cultivars released after 1974 exists in the present literature.
Historical data available from the National Net of Cultivar Comparisons/National Network for Comparison of Durum Wheat Varieties have been used, however, to try to isolate the breeding effect from other sources of variation (environment, geographical distribution, management, etc.) by means of the “check cultivars” included in all trials. Bianchi and Mariani (1993) [51], using data from the trials conducted between 1974 and 1992, for which the check cultivars were “Appulo”, “Capeiti 8” and “Trinakria”, quantified an average yield increase of 1.4 t ha-1 during the considered period, equivalent to an annual increment of approximately 0.07 t ha-1. It is worth noting that the maximum recorded yields exceeded 5.5 t ha-1 during this period. Bias was introduced into the evaluation of their genetic progress by the tall stature of the check cultivars that tended to lodge when grown at high planting densities and with high doses of nitrogen fertilizers. The cultivars that contributed the most to this genetic improvement included “Duilio” and “Simeto” [51].
A similar approach was subsequently used by Salis [52] with the 153 cultivars compared between 1990 and 2010 in 39 field trials, involving 25 cultivars in each of the two Sardinian fields belonging to the national evaluation network. The check cultivars in this period were the semi-dwarf cultivars “Creso”, “Duilio” and “Simeto”. The analysis allowed for the identification of the most productive cultivars released before the 1980s and in different five-year periods, spanning 1981–85 to 2001–05. A continuous increase in grain yield in the considered period was highlighted (Figure 3). “Duetto”, the best among the cultivars released between 2001 and 2005, produced an average 23% (more than 8 t ha-1) more than the check cultivars. Although the check cultivars were semi-dwarf varieties in this case, an overestimation of genetic progress may have arisen from the generally observed ‘decay’ in productivity of a given cultivar (the check cultivars in this case), often deriving from a progressive increase in susceptibility to insects and diseases [48].
The record yield of 10.56 t ha-1 recorded with cultivar “Fiore” was assumed to represent the potential yield for this type of environment, but cultivar “Iride” also produced more than 10 t ha-1 in the same year (2005). Similar potential yields have been reported for bread wheat grown in the UK [47], confirming that the two wheat species share a comparable yield potential. Comparing this potential yield with the attainable water-limited yields (assumed to be equal to the average field yields) and the farm or actual yields as reported in the ISTAT database (Figure 4) reveals large differences. While the gap between potential and attainable yield can mostly be attributed to the notable variability in the amounts and patterns of rainfall in Mediterranean environments, the gap between attainable and farm yields highlights the importance of actions aimed at supporting farmer activities (infra-structure, education, extension services, etc.).
3.2. Plant height, lodging resistance, HI
The gradual reduction in plant height associated with an increase in the harvest index has been a primary breeding objective, affecting lodging susceptibility, sink capacity and biomass partitioning [53,54]. The initial reduction in plant height, of about 30 cm, was obtained crossing mediterraneum with syriacum germplasm (cultivars released between 1950 and 1970, or the so-called ‘intermediate cultivars’), followed by a further reduction of about 20 cm attained by the introduction of the Rht1 gene[49]. These events caused a height decrease of approx. 0.51 cm year−1 for cultivars grown between 1900 and 2000 [54]. This progressive reduction in height also occurred in other countries such as Spain [1] and Argentina [55].
The strong correlation between decrease in plant height and increase in harvest index (HI) led to a notable increase in HI even before the introgression of Rht genes. Starting from average values of 0.25-0.28 for the old cultivars, the HI of the intermediate cultivars was 10% higher, rising a further 4% in the modern, semi-dwarf cultivars released at the end of 20th century, reaching maximum values of around 0.40 [49,56]. The resulting estimated average increase in HI was 0.19±0.017% per year, comparable to the 0.2% increase per year calculated by Brancourt-Hulmel et al. (2003) [57] in bread wheat cultivars released in France between 1946 and 1992.
The reasons why the decrease in plant height translated into an increase in HI are that total biomass was almost unaffected [49,54], and that the reduced stem growth rates allow for more resources to be allocated to the developing ear, resulting in a greater number of fertile florets and grains per ear [47], and ultimately in an estimated increase of 35–41 kernels m−2 year-1 [49,54]. Kernel weight was almost unaffected by the year of release of durum wheat cultivars, or showed just a slight decrease, but grain yield, being closely associated to kernel number m-2, increased by 20–24 kg ha−1 year-1 [49,54].
Following the increase in wheat yield, over 70% of commercial wheat worldwide now contains Rht1 (located on the B genome of bread and durum wheat) or Rht2 (on the D genome of bread wheat) [58]. One of the pleiotropic effects of Rht1 and Rht2 is to reduce plant height by decreasing cell size and reducing cell elongation due to insensitivity to endogenous gibberellins. This results in a shorter coleoptile length and smaller leaf areas in seedlings, undesirable traits in semi-arid and arid environments where they lead to poor emergence and reduced early growth if sowing conditions are unfavourable [58,59]. A gibberellin-sensitive dwarfing gene (Rht14) was identified in the old durum wheat cultivar “Castelporziano”, and it was proposed that it could be used to cope with this problem [59]. Rht14 exerts no negative effect on coleoptile length and seedling establishment, although it negatively affects kernel weight and some quality traits [59].
Lodging incidence also changed as a consequence of the reduction in plant height. Not only does lodging reduce grain yield due to inefficient radiation use but it also worsens grain quality [60]. Lodging depends on plant height [61], and may be caused by stem buckling [62] or by the failure of the root–soil complex. Breeding has not significantly altered stem or anchorage strength [63], hence the improved resistance to lodging of modern cultivars comes from the reduction in plant height. Reducing plant height through the introgression of the syriacum germplasm was less efficient at reducing lodging than the introgression of Rht genes because while the lodging incidence of intermediate cultivars was still lower than that of old cultivars, it remained higher than that of semi-dwarf ones [49].
3.3. Leaf and canopy traits
Breeding for grain yield and earliness indirectly affected a number of leaf and canopy traits involved in carbon assimilation and transpiration. In particular, the progressive reduction in plant height increased canopy aerodynamic resistance, ultimately resulting in a higher canopy temperature of the modern Italian durum wheat cultivars compared with old ones [64]. This result was confirmed, although only for the grain filling period, in a larger set including landraces and modern cultivars from the whole Mediterranean basin [1]. At the leaf level, breeding acted in the opposite direction by decreasing the stomatal resistance of modern cultivars [64]. The combination of a lower canopy temperature and a higher stomatal resistance of older cultivars may be advantageous for the crop’s water requirements and water use efficiency, especially during grain filling [1].
Modern cultivars also produce smaller and thicker flag leaves than older cultivars [64], similarly to what was noted in UK bread wheat [65], likely as a consequence of the presence of semi-dwarfing alleles at Rht-B1 [66,67]. Overall, these changes should be considered positive for photosynthesis because a lower leaf area of the flag leaf may help improve light capture by lower leaves, and the thickness was associated with a greater density of the photosynthetic machinery, as shown by the parallel increase in the nitrogen content per unit of leaf surface [64]. It should be noted that the change in this latter trait had already occurred in the cultivars obtained by crossing mediterraneum with syriacum germplasm before the introgression of “Norin 10” genes. Consistent with this observations, the photosynthetic efficiency of the flag leaf, often correlated with Rubisco activity, is one of the factors associated with grain yield (GY) in modern wheat cultivars [68,69,70].
No effect of breeding was observed on the green area index or the leaf area index [71], but senescence began later relative to anthesis date, and developed at a faster rate in the modern than in the old cultivars. The earlier anthesis in modern cultivars is one of the reasons for their later onset of senescence, because it shifts the anthesis-maturity period to less stressful conditions. As a result, modern cultivars maintain their greenness over a greater proportion of their anthesis-to-maturity period [64].
It is interesting that in both bread wheat and durum wheat the morphological changes in the flag leaf seen with this ‘breeding progress’ (e.g., smaller, more erect, higher N per unit area, and higher chlorophyll concentration) were not unlike those reported for winter wheat progress in the United Kingdom [72].
3.4. Phenology
The timing of phenological events is one of the most critical factors for crop adaptation and yield in a specific environment [73,74], and an optimal window for flowering can be defined for each environment. In Mediterranean environments this optimal window is bound by spring frost, on one side, and by terminal water and temperature stress on the other. The lateness of old cultivars with respect to this optimal window led to an advance in anthesis date as a consequence of breeding [49]. In contrast with what was observed in bread wheat [75], in durum wheat the advance in the anthesis date was attained before the introgression of Rht genes thanks to crosses with syriacum germplasm [49], while the subsequent introgression of Rht genes appears to have delayed flowering time with respect to intermediate cultivars [76].
The genetic regulation of flowering time in wheat is determined by the interplay of at least 20 genes scattered across the whole genome [77]. These genes have been classified according to whether they respond to vernalization or to photoperiod or whether they confer earliness per se [78,79,80]. Understanding which of these genes were involved in the change in anthesis date in the shift from old to modern cultivars and which ones are governing the development of modern cultivars may be useful to improve adaptation and yield potential even further [81]. According to Motzo and Giunta (2007) [76], the lateness of old cultivars was due to their greater photoperiod sensitivity and cold requirements, whereas the greater earliness per se was the reason for the earlier flowering time of the intermediate group of cultivars. Rht genes reduced photoperiod sensitivity but decreased earliness per se. This residual earliness per se is fundamental to modern cultivars to preserve and increase the length of the terminal spikelet–anthesis period because the longer this period, the greater the ear fertility [82]. Interestingly, some quantitative cold requirements are still present in modern Italian cultivars, although they are generally classified as spring types [76], partly because the traditional autumn sowings ensure the cold requirement is met [76]. In any case, the lower cold requirement of modern cultivars compared with old ones has the drawback of increasing their inter-annual variability in anthesis date by increasing the role of temperature in the control of their development [83].
The reduced duration of the vegetative phase of modern cultivars [54] moved the grain filling period to less stressful conditions and lengthened its duration whenever the terminal drought caused the simultaneous senescence of all cultivars, contributing to the increase in HI. Therefore, the above-mentioned relative constancy of grain weight was the result of an almost full compensation between rate and duration of grain filling, as shown by the negative correlation between the maximum rate of grain filling (that ranged between 2.4 and 3.3 mg d-1) and the year of cultivar release of six Italian cultivars constituted between 1910 and 1996 [84]. The high rate of grain filling of old cultivars confers tolerance to terminal drought [85], i.e. the ability to maintain high grain weights despite the high temperatures and short grain filling caused by their lateness [86].
3.5. Nitrogen uptake and use-efficiency
In contrast with what was observed in bread wheat [87], durum wheat breeding induced an increase in total nitrogen uptake, necessary to sustain the increases in yield and harvest index [49]. Grains have a higher nitrogen concentration than straw, and the greater proportion of dry matter allocated to grains as a consequence of the increase in HI was, therefore, the reason for the increase in total N uptake, despite the parallel reduction in grain protein percentage [88]. This is why similar time trends were observed between HI and NHI, with the latter increasing from 0.71 in old cultivars to 0.80 in semi-dwarf ones [49].
Although substantial progress has been made, wheat is the least efficient major crop in terms of nitrogen use, with its nitrogen use efficiency (NUE, grain yield vs total available N in the soil) around 25% lower than that of maize and rice [89]. From an agronomic point of view, fertilizer use efficiency (fertilizer recovered in total biomass as a fraction of fertilizer applied) is a useful index to consider, estimated to be only 33% in wheat [90].
A recent study by Lupini et al. (2021) [91], comparing NUE and tolerance to water and nitrogen stress in old and modern durum wheat varieties, found that modern cultivars outperformed old ones under conventional nitrogen and water regimes, whereas old varieties were better able to cope with both water and nitrogen stress, particularly at the stem elongation and milk-ripening stages of development, when nitrate and ammonium transporter gene expression in the root was the highest. NUE in this paper was calculated as total biomass production per unit of N taken up, to avoid in the penalization conferred to old cultivars from their low HIs [92]. These findings confirm recent discoveries suggesting the possible indirect selection of wheat varieties with better NUE through the choice of high-yielding varieties [10,93]. In particular, Taranto et al. [10] showed that major well-known genes and/or QTLs (quantitative trait loci) influencing traits such as plant height (RHT), earliness (VRN, PPD) and grain quality could explain the differences between old and modern durum wheat varieties, and are often located in the same genomic regions where genes related to nitrogen metabolism are found.
3.6. Grain and semolina quality
Durum wheat quality is evaluated according to the final product to be obtained. Before the 20th century no distinction was made between bread and durum wheat, and durum was used for both bread and pasta production. It was De Cillis (1942) [5,17] who first demonstrated that pasta made using vitreous grains, which have a higher protein content, had a better cooking quality than pasta made from starchy grains with a lower protein content. Nowadays, durum wheat is mainly used for pasta production, although in the south of Italy it is traditionally used for breadmaking too. In both cases, protein percentage and gluten strength are the traits defining the quality of durum wheat, with grain protein percentage contributing the most (40%) to the EUQuality Index for durum wheat (European Commission Regulation No. 2237/2003, 23 December 2003).
One of the trade-offs for an increase in grain yield obtained with breeding is the decrease in grain protein percentage, estimated at 0.14–0.19% year-1 [54,94]. This has led to an average difference in protein percentage of about 1–1.5% between landraces and modern Italian cultivars [49,95], with protein content in old cultivars ranging between 11.9–14% against a variation in the range of 10.5–12% for modern cultivars in a five-year field trial which compared 14 old against 14 modern cultivars [83]. This superiority of old cultivars in protein percentage persists even when they are grown in soils characterized by lower fertility and with lower fertilization rates than modern ones [83]. As stated above, this difference cannot be attributed to a lower capacity of modern cultivars to take up nitrogen, nor to a reduced NHI, but rather to the decrease in the amount of nitrogen allocated to each grain caused by the higher number of grains m-2 set up by modern cultivars, responsible for their high grain yields [83].
Gluten strength describes the continuity and strength of the protein network associated with gluten viscosity and elasticity. Strong and tenacious gluten is needed when pasta is the final product, whereas strong but extensible gluten is needed in the case of breadmaking. Given similar protein percentages, gluten strength depends on the types of glutenin and gliadin proteins present (genetically determined) and on their ratio [96], and it can be quantified via the Gluten Index (0–100%) [97,98]. Breeding activity on durum wheat was mainly directed at ameliorating pasta-making quality, and compensated for the above-mentioned decrease in protein percentage by improving gluten strength through the incorporation of favourable combinations of alleles for both high (HMW-GS) and low (LMW-GS) molecular weight glutenin subunits, and by an increase in the expression of B-type LMW-GS [99]. As a consequence, the glutenin-gliadin ratio increased, the Gluten index increased from 6–32% in landraces to 55–87% in modern cultivars, and the technological quality of dough in general improved [50,95,99,100,101]. Similar results were obtained in Spain [94]. The higher gluten strength of modern cultivars, combined with their lower protein percentage, resulted in a lower or similar spaghetti cooking quality [100,102] depending on the pasta drying temperature—low (<60°C), high (60–80 °C) or ultra-high (80–100°C)—with consequent changes in the relative importance of quantity vs quality of proteins. While the quality of pasta dried at low temperatures is governed by both protein quantity and quality, the quality of pasta dried at high and ultra-high drying temperatures is governed by protein quantity only [103].
The successful breeding work on gluten strength had the negative side-effect of decreasing the large genetic variability in gluten characteristics present in the landraces to the very low levels retained in modern cultivars [9]. This means that landraces provide potential sources of genetic variation for gluten characteristics [104], particularly when bread is the final product in mind [2,95].
Durum wheat bread is characterized by a higher protein content, greater yellowness and by a longer shelf-life than common bread. It remains highly traditional in the Mediterranean region, in Italy in particular [105,106,107]. However, bread made with durum wheat cannot achieve the same volume as bread made with bread wheat because although its gluten is more tenacious, it is less strong and elastic as a consequence of the absence of the glutenin D-genome, which confers extensibility to bread wheat [108,109,110]. Substituting the glutenin alleles of durum wheat was a less effective breeding strategy than transferring bread wheat chromosome 1D, responsible for gluten and dough strength, and/or chromosome 5D, responsible for kernel softness [106,110,111,112,113]. The improved gluten extensibility and superior baking quality obtained by Canadian breeders by crossing durum with emmer wheat (T. dicoccum, tetraploid) improved the breadmaking quality of durum wheat without sacrificing its strong gluten and pasta-cooking quality [114,115].
The importance of pasta colour for the market led breeders to try and enhance the carotenoid content of durum kernels and semolina through the introgression of a yellow pigment gene (Yp) [116]. In contrast with De Vita et al. (2007) [54], who did not find any difference in colour between old and modern cultivars, several studies ascertained modern cultivars to have a higher yellow index than old cultivars [94,117]
4. Future prospects: lessons from the past
4.1. Grain yield
Increasing biomass production seems to be the most promising strategy for increasing grain yield [118] given the limited scope for increasing HI above the actual potential 50% due to the negative consequences of further reductions in plant height on grain yield [119]. It was suggested that the optimal plant height to maximize yield often falls between 0.7 and 1.0 m. Greater biomass already appears to be contributing to genetic yield progress in modern bread wheat cultivars released since about 1990 (e.g., [72,120]).
Biomass can be increased by lengthening the growth cycle. Antedate sowings can lengthen the growing season if phenology is also changed through the introduction of some cold requirement and/or with an increase in photoperiodic sensitivity to avoid excessively early anthesis. Crop simulations reveal that an early sowing system combined with slower-developing wheat genotypes could exploit a longer growing season and increase grain yield in spite of the recent climate changes [121]. In semi-arid and arid environments, advancing sowing date would also require increasing sowing depth to guarantee the seed sufficient humidity for germination. Gibberellin-sensitive Rht genes, such as Rht14 in the old “Castelporziano” cultivar, are needed in this case, because their longer coleoptiles are associated with good seedling emergence.
4.2. Phenology: is the advance in anthesis date still desirable?
Between 2011 and 2020, the average surface temperature increased by 1.1°C compared with the average temperature of the late 19th century (before the industrial revolution) and it is now warmer than any other period in the last 100,000 years (IPCC 2021). The earliness of most durum wheat cultivars, partly due to limited or absent vernalization sensitivity, could already represent a problem today, but even more so in the future. The increasing temperatures associated with climate change may cause an excessive advance in anthesis date and a shortening of the whole growing period (the time from sowing to maturity), becoming the main yield-reduction factor [122]. Both modelling and field data have demonstrated that the combination of a delayed anthesis and a higher rate of grain filling compared with current cultivars is an effective adaptation strategy in warmer climates because it increases both grain and protein yield, provided that nitrogen supply is not limiting [123]. This combination of traits is present in many old cultivars [84,86]. The positive effect of a delayed anthesis on grain yield is likely the consequence of the lengthening of the sowing–anthesis period that, by allowing for a greater biomass at anthesis, may result in an increased grain set [124].
4.3. Grain quality: are grain yield and grain protein mutually exclusive?
Durum wheat grain protein and grain yield are often negatively correlated [83,125]. The comparison between old (lower grain yields and higher grain protein percentages) and modern (the opposite combination) cultivars has identified in an imbalance between the increase in total nitrogen uptake and increase in grain number m-2 (the main determinant of grain yield), and the reason for this trade-off is associated with breeding. In modern cultivars, a rather limited source of nitrogen is diluted by high grain numbers [126]. Grain nitrogen comes partly from the N absorbed before anthesis, which is remobilized to the grains from senescing organs, and partly from post-anthesis N uptake. It seems that the good capacity to absorb N after anthesis endows a cultivar with the ability to make the most of favourable weather conditions in the spring by combining a high grain N with a high grain yield [127,128]. Traits which improve post-anthesis N uptake are a high potential grain weight [129] and a prolonged root activity after anthesis [128].
Breeding for gluten strength in durum wheat focused on pasta-making quality so that only a part of the genetic diversity of durum wheat has been captured in the modern varieties generated through breeding over the last century, and the modern germplasm is characterized by a drastically limited variation at the loci and in the allelic combinations controlling gluten strength [9]. Landraces and old cultivars represent, therefore, a potential source of genetic variability, and could be used to identify the best gluten composition and the appropriate gluten strength, particularly for final products other than pasta, such as bread [2,99,104,130].
A wide range of micro-nutrients (iron and zinc primarily) has been observed in durum landraces [131,132], making them potentially useful for improving the nutritional value of modern cultivars, which are instead characterized by a rather limited range of micro-nutrients [133].
The volatile organic compounds responsible for bread aroma, a very complex and important trait for breadmaking, are different in old vs modern cultivars, once again offering new opportunities for future breeding with this quality in mind [104].
5. Conclusions
The effective process of genetic improvement, by replacing older and genetically heterogeneous varieties with new highly productive and superior quality varieties, inevitably involved a reduction in overall genetic diversity [9]. In the current context of climate change and rapidly mutating pathogen populations, preserving the yield level through the continuous introduction of new cultivars, exploiting the reservoir of largely unused genetic variation stored in old cultivars and landraces, could be as important as increasing grain yield and quality [134]. The accurate phenotyping of agronomic and physiological traits and the study of the relationships between agronomic performance and genetic structure will be fundamental for exploiting the genetic variability present in old constitutions.
Author Contributions
“Conceptualization, M.L.C. and R.M.; methodology, F.G.; data curation, M.L.C.; writing—original draft preparation, M.L.C.; writing—review and editing, M.L.C., R.M., F.G.; supervision, R.M.. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This study was carried out within the Agritech National Research Center (Spoke 7) and received funding from the European Union Next-GenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR) – MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4 – D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Data Availability Statement
Publicly available datasets were analyzed in this study
Conflicts of Interest
The authors declare no conflict of interest.
References
- Royo, C.; Ammar, K.; Villegas, D.; Soriano, J.M. Agronomic, Physiological and Genetic Changes Associated With Evolution, Migration and Modern Breeding in Durum Wheat. Front. Plant Sci. 2021, 12, 674470. [Google Scholar] [CrossRef] [PubMed]
- Mefleh, M.; Conte, P.; Fadda, C.; Giunta, F.; Piga, A.; Hassoun, G.; Motzo, R. From Ancient to Old and Modern Durum Wheat Varieties: Interaction among Cultivar Traits, Management, and Technological Quality. J. Sci. Food Agric. 2019, 99, 2059–2067. [Google Scholar] [CrossRef]
- Bagnara, D.; Scarascia-Mugnozza, G.T. Outlook in Breeding for Yield in Durum Wheat. In Proceedings of the Proceedings of the Symposium on genetics and breeding of durum wheat; p. 1975.
- Azeez, M.A.; Adubi, A.O.; Durodola, F.A. Landraces and Crop Genetic Improvement. In Rediscovery of Landraces as a Resource for the Future; IntechOpen, 2018; ISBN 1-78923-725-4. [Google Scholar]
- De Cillis, E. I Grani d’Italia. Tipografia della Camera dei deputati. 1927. [Google Scholar]
- Grignac, P. Contribution à l’étude de Triticum Durum Desf, Toulouse, 1965.
- Bozzini, A. Genetica e Miglioramento Genetico Dei Frumenti Duri; Tipografia del libro, 1970. [Google Scholar]
- Annicchiarico, P.; Pecetti, L.; Damania, A.B. Relationships between Phenotypic Variation and Climatic Factors at Collecting Sites in Durum Wheat Landraces. Hereditas 1995, 122, 163–167. [Google Scholar] [CrossRef]
- Nazco, R.; Peña, R.J.; Ammar, K.; Villegas, D.; Crossa, J.; Royo, C. Durum Wheat (Triticum Durum Desf.) Mediterranean Landraces as Sources of Variability for Allelic Combinations at Glu-1/Glu-3 Loci Affecting Gluten Strength and Pasta Cooking Quality. Genet. Resour. Crop Evol. 2014, 61, 1219–1236. [Google Scholar] [CrossRef]
- Taranto, F.; D’Agostino, N.; Rodriguez, M.; Pavan, S.; Minervini, A.P.; Pecchioni, N.; Papa, R.; De Vita, P. Whole Genome Scan Reveals Molecular Signatures of Divergence and Selection Related to Important Traits in Durum Wheat Germplasm. Front. Genet. 2020, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Marzario, S.; Logozzo, G.; David, J.L.; Zeuli, P.S.; Gioia, T. Molecular Genotyping (SSR) and Agronomic Phenotyping for Utilization of Durum Wheat (Triticum Durum Desf.) Ex Situ Collection from Southern Italy: A Combined Approach Including Pedigreed Varieties. Genes 2018, 9, 465. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Maliani, C. Nuove Varietà Di Grano Duro e Loro Caratteristiche. Quad. Accad. Georgofili VI 1998, 57–64. [Google Scholar]
- Porceddu, E. Evoluzione Varietale e Problemi Attuali Del Miglioramento Genetico Dei Cereali Vernini. Riv. Agron. 1987, 21, 33–54. [Google Scholar]
- Rivoira, G.; Deidda, M.; Marras, G.F. Evoluzione e Problemi Attuali Della Coltivazione Dei Cereali Vernini. Riv Agron 1987, 21, 5–32. [Google Scholar]
- Iannelli, P.; Pezzali, M. [Durum Wheat: You Have Worked Hard, but You Have to Insist in This Way to Be Self-Sufficient].[Italian]. Inf. Agrar. 1976. [Google Scholar]
- Bozzini, A.; Corazza, L.; D’Egidio, M.G.; Di Fonzo, N.; La Fiandra, D.; Pogna, N.E.; Poma, I. Durum Wheat (Triticum Turgidum Spp. Durum). In Proceedings of the “Italian contribution to plant genetics and breeding”, University of Tuscia. Book prepared at the occasion of the XV Congress of the European Association for Research on Plant Breeding-Eucarpia, 21-25 Sep; 1998; pp. 181–194. [Google Scholar]
- Bianchi, A.; Boggini, G. Evoluzione Varietale. Quad. Accad. Dei Georgofili VI 1998, 25–42. [Google Scholar]
- Strampelli, N. Origini, Sviluppi, Lavori e Risultati. Milano Italy Ist. Naz. Genet. Cerealic. Alfieri Lacroix 1932. [Google Scholar]
- Casale, F. Tre Novità Agrarie. Soc. Ital. Genet. Agrar. 1955, 5, 339–350. [Google Scholar]
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic Diversity and Population Structure of Tetraploid Wheats (Triticum Turgidum L.) Estimated by SSR, DArT and Pedigree Data. Plos One 2013, 8, e67280. [Google Scholar] [CrossRef] [PubMed]
- Caruso, P.; Majo, I. Contributo al Miglioramento Genetico Del Grano Duro. Gen. Agr. 1964, XVII, 107–131. [Google Scholar]
- Pecetti, L.; Boggini, G.; Doust, M.A.; Annicchiarico, P. Performance of Durum Wheat Landrace from Jordan and Morocco in Two Mediterranean Environments (Northern Syria and Sicily). J. Genet. Breed. 1996, 50, 41–46. [Google Scholar]
- Polano, P. Contributo al Miglioramento Della Produzione Del Grano Duro in Sardegna. Studi Sassar. 1958, VI, 1–28. [Google Scholar]
- Bogyo, T.P.; Scarascia-Mugnozza, G.T.; Sigurbjörnsson, B.; Bagnara, D. ADAPTATION STUDIES WITH RADIATION-INDUCED DURUM WHEAT MUTANTS; Washington State Univ; Pullman, 1969. [Google Scholar]
- Vallega, J. Historical Perspective of Wheat Breeding in Italy. In Proceedings of the 4. FAO/Rockefeller Foundation Wheat Seminar, Tehran, Iran, 21 May 1973; 1974. [Google Scholar]
- Vallega, J.; Zitelli, G. New High Yielding Italian Durum Wheat Varieties. Ann. Ist. Sper. Cerealic. Roma Italy 1975. [Google Scholar]
- Barbieri, A.; Deidda, M. Contributo al Miglioramento Genetico Del Grano Duro. Sementi Elette 1968, 3, 1–3. [Google Scholar]
- Ballatore, G.P. Due Nuove Cultivar Di Grano Duro, 1970.
- Rivoira, G. Alcune Considerazioni Agronomiche Sulle Nuove Cultivars Italiane Di Grano Duro. Genetica agraria 1982, V, 142–192. [Google Scholar]
- Alessandroni, A.; Rusmini, B.; Scalfati, M.C. Influenza Di Specie Del Genere Triticum e Di Altri Generi Nelle Discendenze Di Incroci Interspecifici e Intergenerici per Il Miglioramento Genetico Di Triticum Durum; Instituto nazionale di genetica per la cerealicoltura” N. Strampelli, 1967. [Google Scholar]
- Scarascia Mugnozza, G.T. Due Linee Di Frumento Duro Ottenute per Mutazione Radioindotta. Com. Naz. Energ. Nucl. Italy 1968, 4, 3–8. [Google Scholar]
- Bozzini, A.; Girotto, G. Grani Duri Nell’Italia Centrale. L’Informatore Agrar. 1971, 48, 7335–7340. [Google Scholar]
- Bozzini, A.; Scarascia-Mugnozza, G.T. A Dominant Short Straw Mutation Induced by Thermal Neutrons in Durum Wheat. Wheat Inf. Serv. 1967, 23, 5–6. [Google Scholar]
- Lumpkin, T.A. How a Gene from Japan Revolutionized the World of Wheat: CIMMYT’s Quest for Combining Genes to Mitigate Threats to Global Food Security. In Proceedings of the Advances in Wheat Genetics: From Genome to Field: Proceedings of the 12th International Wheat Genetics Symposium, Springer Japan, 2015; pp. 13–20.
- Bozzini, A.; Bagnara, D. Creso, Mida e Tito, Nuovi Grani Duri Del Laboratorio Agricoltura Del CNEN. L’informatore Agrar. 1974, 20, 15869–15871. [Google Scholar]
- Rivoira, G. Risultati Agronomici Conseguiti Dall’attività Collegiale Di Ricerca Del Subprogetto “Frumento Duro”; Enna: 1979; pp. 149–173.
- Porceddu, E.; Spagnoletti Zeuli, P.L.; Scarascia Mugnozza, G.T. Genetical Variability in F3 Families from Genetically Distant Durum Wheat (Triticum Durum) Varieties. Genet. Agrar. 1980. [Google Scholar]
- Blanco, A.; SCARASCIA MUGNOZZA, G. Messapia: Una Nuova Varietà Di Frumento Duro Precoce e Produttiva. Inf. Agrar. 1989, 37, 61–63. [Google Scholar]
- Blanco, A.; Simeone, R.; Resta, P. The Addition of Dasypyrum Villosum (L.) Candargy Chromosomes to Durum Wheat (Triticum Durum Desf.). Theor. Appl. Genet. 1987, 74, 328–333. [Google Scholar] [CrossRef]
- Blanco, A.; Giorgi, B.; Perrino, P.; Simeone, R. Genetic Resources and Breeding of Durum Wheat Quality. Agric. E Ric. 1991, 41–58. [Google Scholar]
- Porfiri, O. La ricerca varietale al servizio delle filiere.
- Conforti, P. Un’analisi Delle Ipotesi Di Riduzione Del Sostegno al Grano Duro Nell’Unione Europea e Dei Suoi Effetti in Italia. INEA Work. Pap. 2002, 82. [Google Scholar]
- Lorenzetti, R.; Strampelli, N.; Degani, L.; Raimondi, G.; Wittrich, A. The Wheat Science: The Green Revolution of Nazareno Strampelli. No Title 2000. [Google Scholar]
- Quaranta, F.; Belocchi, A.; Fornara, M.; Ripa, C.; D’Egidio, M.G. Le varietà di frumento duro in Italia. 2007.
- Reynolds, M.; Langridge, P. Physiological Breeding. Curr. Opin. Plant Biol. 2016, 31, 162–171. [Google Scholar] [CrossRef]
- Fischer, R.A.; Edmeades, G.O. Breeding and Cereal Yield Progress. Crop Sci. 2010, 50, S-85–S-98. [Google Scholar] [CrossRef]
- Connor, D.J.; Mínguez, M.I. Evolution Not Revolution of Farming Systems Will Best Feed and Green the World. Glob. Food Secur. 2012, 1, 106–113. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Trends since 1900 in the Yield Potential of Italian-Bred Durum Wheat Cultivars. Eur. J. Agron. 2007, 27, 12–24. [Google Scholar] [CrossRef]
- De Vita, P.; Mastrangelo, A.M.; Codianni, P.; Fornara, M. Bio-Agronomic Evaluation of Old and Modern Wheat, Spelt and Emmer Genotypes for Low-Input Farming in Mediterranean Environment. Ital. J. Agron. 2007, 2, 291. [Google Scholar] [CrossRef]
- Bianchi, A.; Mariani, B.M. La Rete Nazionale Di Prove Di Confronto Tra Varietà Di Frumento Duro: Un Bilancio Dopo Venti Anni. L’informatore Agrar. 1993, 34, 5–7. [Google Scholar]
- Salis, G.; Motzo, R.; Giunta, F. Salis, G. (2011). Analisi Di Venti Anni Di Prove Di Confronto Tra Varietà Di Frumento Duro (Triticum Turgidum Ssp. Durum L.), Università degli studi di Sassari, Dipartimento di Agraria, Sez. di agronomia, coltivazioni erbacee e genetica, Corso di Laurea in Scienze e Tecnologie agrarie: Sassari, 2011.
- Porceddu, E.; Blanco, A. Evolution of Durum Wheat Breeding in Italy. In Proceedings of the International Symposium on Genetics and Breeding of Durum Wheat; Porceddu, E., Qualset, C.O., Damania, A.B., Eds.; 2014; pp. 157–173. [Google Scholar]
- De Vita, P.; Nicosia, O.L.D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding Progress in Morpho-Physiological, Agronomical and Qualitative Traits of Durum Wheat Cultivars Released in Italy during the 20th Century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Achilli, A.L.; Roncallo, P.F.; Echenique, V. Genetic Gains in Grain Yield and Agronomic Traits of Argentinian Durum Wheat from 1934 to 2015. Agronomy 2022, 12, 2151. [Google Scholar] [CrossRef]
- Royo, C.; Álvaro, F.; Martos, V.; Ramdani, A.; Isidro, J.; Villegas, D.; García Del Moral, L.F. Genetic Changes in Durum Wheat Yield Components and Associated Traits in Italian and Spanish Varieties during the 20th Century. Euphytica 2007, 155, 259–270. [Google Scholar] [CrossRef]
- Brancourt-Hulmel, M.; Doussinault, G.; Lecomte, C.; Bérard, P.; Le Buanec, B.; Trottet, M. Genetic Improvement of Agronomic Traits of Winter Wheat Cultivars Released in France from 1946 to 1992. Crop Sci. 2003, 43, 37–45. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Ellis, M.H.; Bonnett, D.G.; Condon, A.G.; Falk, D.; Richards, R.A. The Rht13 Dwarfing Gene Reduces Peduncle Length and Plant Height to Increase Grain Number and Yield of Wheat. Field Crops Res. 2011, 124, 323–331. [Google Scholar] [CrossRef]
- Duan, S.; Zhao, Z.; Qiao, Y.; Cui, C.; Morgunov, A.; Condon, A.G.; Chen, L.; Hu, Y.-G. GAR Dwarf Gene Rht14 Reduced Plant Height and Affected Agronomic Traits in Durum Wheat (Triticum Durum). Field Crops Res. 2020, 248, 107721. [Google Scholar] [CrossRef]
- Berry, P.M.; Spink, J. Predicting Yield Losses Caused by Lodging in Wheat. Field Crops Res. 2012, 137, 19–26. [Google Scholar] [CrossRef]
- Berry, P.M.; Sterling, M.; Spink, J.H.; Baker, C.J.; Sylvester-Bradley, R.; Mooney, S.J.; Tams, A.R.; Ennos, A.R. Understanding and Reducing Lodging in Cereals. Adv. Agron. 2004, 84, 215–269. [Google Scholar]
- Neenan, M.; Spencer-Smith, J.L. An Analysis of the Problem of Lodging with Particular Reference to Wheat and Barley. J. Agric. Sci. 1975, 85, 495–507. [Google Scholar] [CrossRef]
- CROOK, M.J.; Ennos, A.R. The Mechanics of Root Lodging in Winter Wheat, Triticum Aestivum L. J. Exp. Bot. 1993, 44, 1219–1224. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Has Long-Term Selection for Yield in Durum Wheat Also Induced Changes in Leaf and Canopy Traits? Field Crops Res. 2008, 106, 68–76. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Sylvester-Bradley, R.; Weightman, R.; Snape, J.W. Identifying Physiological Traits Associated with Improved Drought Resistance in Winter Wheat. Field Crops Res. 2007, 103, 11–24. [Google Scholar] [CrossRef]
- Gale, M.D.; Youssefian, S. Dwarfing Genes in Wheat. Prog. Plant Breed. 1985, 1, 1–35. [Google Scholar]
- LeCain, D.R.; Morgan, J.A.; Zerbi, G. Leaf Anatomy and Gas Exchange in Nearly Isogenic Semidwarf and Tall Winter Wheat. Crop Sci. 1989, 29, 1246–1251. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.-M.; Condon, A.G.; Saavedra, A.L. Wheat Yield Progress Associated with Higher Stomatal Conductance and Photosynthetic Rate, and Cooler Canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Gaju, O.; DeSilva, J.; Carvalho, P.; Hawkesford, M.J.; Griffiths, S.; Greenland, A.; Foulkes, M.J. Leaf Photosynthesis and Associations with Grain Yield, Biomass and Nitrogen-Use Efficiency in Landraces, Synthetic-Derived Lines and Cultivars in Wheat. Field Crops Res. 2016, 193, 1–15. [Google Scholar] [CrossRef]
- Carmo-Silva, E.; Andralojc, P.J.; Scales, J.C.; Driever, S.M.; Mead, A.; Lawson, T.; Raines, C.A.; Parry, M.A. Phenotyping of Field-Grown Wheat in the UK Highlights Contribution of Light Response of Photosynthesis and Flag Leaf Longevity to Grain Yield. J. Exp. Bot. 2017, 68, 3473–3486. [Google Scholar] [CrossRef]
- Álvaro, F.; Isidro, J.; Villegas, D.; García Del Moral, L.F.; Royo, C. Breeding Effects on Grain Filling, Biomass Partitioning, and Remobilization in Mediterranean Durum Wheat. Agron. J. 2008, 100, 361–370. [Google Scholar] [CrossRef]
- Shearman, V.J.; Sylvester-Bradley, R.; Scott, R.K.; Foulkes, M.J. Physiological Processes Associated with Wheat Yield Progress in the UK. Crop Sci. 2005, 45, 175–185. [Google Scholar] [CrossRef]
- Richards, R.A. Crop Improvement for Temperate Australia: Future Opportunities. Field Crops Res. 1991, 26, 141–169. [Google Scholar] [CrossRef]
- Shorter, R.; Lawn, R.J.; Hammer, G.L. Improving Genotypic Adaptation in Crops–a Role for Breeders, Physiologists and Modellers. Exp. Agric. 1991, 155–175. [Google Scholar] [CrossRef]
- Canevara, M.G.; Romani, M.; Corbellini, M.; Perenzin, M.; Borghi, B. Evolutionary Trends in Morphological, Physiological, Agronomical and Qualitative Traits of Triticum Aestivum L. Cultivars Bred in Italy since 1900. Eur. J. Agron. 1994, 3, 175–185. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F. The Effect of Breeding on the Phenology of Italian Durum Wheats: From Landraces to Modern Cultivars. Eur. J. Agron. 2007, 26, 462–470. [Google Scholar] [CrossRef]
- Snape, J.W.; Quarrie, S.A.; Laurie, D.A. Comparative Mapping and Its Use for the Genetic Analysis of Agronomic Characters in Wheat. Euphytica 1996, 89, 27–31. [Google Scholar] [CrossRef]
- Worland, A.J. The Influence of Flowering Time Genes on Environmental Adaptability in European Wheats. Euphytica 1996, 89, 49–57. [Google Scholar] [CrossRef]
- Law, C.N.; Worland, A.J. Genetic Analysis of Some Flowering Time and Adaptive Traits in Wheat. New Phytol. 1997, 137, 19–28. [Google Scholar] [CrossRef]
- Laurie, D.A.; Griffiths, S.; Dunford, R.P.; Christodoulou, V.; Taylor, S.A.; Cockram, J.; Beales, J.; Turner, A. Comparative Genetic Approaches to the Identification of Flowering Time Genes in Temperate Cereals. Field Crops Res. 2004, 90, 87–99. [Google Scholar] [CrossRef]
- Slafer, G.A.; Abeledo, L.G.; Miralles, D.J.; Gonzalez, F.G.; Whitechurch, E.M. Photoperiod Sensitivity during Stem Elongation as an Avenue to Raise Potential Yield in Wheat. In Wheat in a Global Environment: Proceedings of the 6th International Wheat Conference, 5–9 June 2000, Budapest, Hungary; Bedö, Z., Láng, L., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 487–496. ISBN 978-94-017-3674-9. [Google Scholar]
- González, F.G.; Slafer, G.A.; Miralles, D.J. Grain and Floret Number in Response to Photoperiod during Stem Elongation in Fully and Slightly Vernalized Wheats. Field Crops Res. 2003, 81, 17–27. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Motzo, R. Grain Yield and Grain Protein of Old and Modern Durum Wheat Cultivars Grown under Different Cropping Systems. Field Crops Res. 2019, 230, 107–120. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F.; Pruneddu, G. The Response of Rate and Duration of Grain Filling to Long-Term Selection for Yield in Italian Durum Wheats. Crop Pasture Sci. 2010, 61, 162. [Google Scholar] [CrossRef]
- Bruckner, P.L.; Frohberg, R.C. Bruckner, P. L., & Frohberg, R. C. (1987). Rate and Duration of Grain Fill in Spring Wheat 1. Crop Science, 27(3), 451-455. Crop Sci. 1987, 27, 451–455. [Google Scholar]
- Giunta, F.; Mefleh, M.; Pruneddu, G.; Motzo, R. Role of Nitrogen Uptake and Grain Number on the Determination of Grain Nitrogen Content in Old Durum Wheat Cultivars. Agronomy 2020, 11, 42. [Google Scholar] [CrossRef]
- Calderini, D.F.; Torres-León, S.; Slafer, G.A. Consequences of Wheat Breeding on Nitrogen and Phosphorus Yield, Grain Nitrogen and Phosphorus Concentration and Associated Traits. Ann. Bot. 1995, 76, 315–322. [Google Scholar] [CrossRef]
- Sinclair, T.R. Historical Changes in Harvest Index and Crop Nitrogen Accumulation. Crop Sci. 1998, 38, 638–643. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, H.; Chen, X.; et al. Pursuing Sustainable Productivity with Millions of Smallholder Farmers. Nature 2018, 555, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Raun, W.R.; Johnson, G.V. Improving Nitrogen Use Efficiency for Cereal Production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Lupini, A.; Preiti, G.; Badagliacca, G.; Abenavoli, M.R.; Sunseri, F.; Monti, M.; Bacchi, M. Nitrogen Use Efficiency in Durum Wheat Under Different Nitrogen and Water Regimes in the Mediterranean Basin. Front. Plant Sci. 2021, 11, 607226. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J. Genetic Variation in Traits for Nitrogen Use Efficiency in Wheat. J. Exp. Bot. 2017, 68, 2627–2632. [Google Scholar] [CrossRef]
- Cormier, F.; Faure, S.; Dubreuil, P.; Heumez, E.; Beauchêne, K.; Lafarge, S.; Praud, S.; Le Gouis, J. A Multi-Environmental Study of Recent Breeding Progress on Nitrogen Use Efficiency in Wheat (Triticum Aestivum L.). Theor. Appl. Genet. 2013, 126, 3035–3048. [Google Scholar] [CrossRef]
- Subira, J.; Peña, R.J. Breeding Progress in the Pasta-Making Quality of Durum Wheat Cultivars Released in Italy and Spain during the 20th Century. 2014.
- Motzo, R.; Fois, S.; Giunta, F. Relationship between Grain Yield and Quality of Durum Wheats from Different Eras of Breeding. Euphytica 2004, 140, 147–154. [Google Scholar] [CrossRef]
- Sissons, M. Role of Durum Wheat Composition on the Quality of Pasta and Bread. Food 2008, 2, 75–90. [Google Scholar]
- Edwards, N.M.; Gianibelli, M.C.; McCaig, T.N.; Clarke, J.M.; Ames, N.P.; Larroque, O.R.; Dexter, J.E. Relationships between Dough Strength, Polymeric Protein Quantity and Composition for Diverse Durum Wheat Genotypes. J. Cereal Sci. 2007, 45, 140–149. [Google Scholar] [CrossRef]
- Clarke, F.R.; Clarke, J.M.; Ames, N.A.; Knox, R.E.; Ross, R.J. Gluten Index Compared with SDS-Sedimentation Volume for Early Generation Selection for Gluten Strength in Durum Wheat. Can. J. Plant Sci. 2010, 90, 1–11. [Google Scholar] [CrossRef]
- De Santis, M.A.; Giuliani, M.M.; Giuzio, L.; Vita, P.D.; Lovegrove, A.; Shewry, P.R.; Flagella, Z. Differences in Gluten Protein Composition between Old and Modern Durum Wheat Genotypes in Relation to 20th Century Breeding in Italy. Eur. J. Agron. 2017, 87, 19–29. [Google Scholar] [CrossRef]
- Fois, S.; Schlichting, L.; Marchylo, B.; Dexter, J.; Motzo, R.; Giunta, F. Environmental Conditions Affect Semolina Quality in Durum Wheat ( Triticum Turgidum Ssp. Durum L.) Cultivars with Different Gluten Strength and Gluten Protein Composition. J. Sci. Food Agric. 2011, 91, 2664–2673. [Google Scholar] [CrossRef]
- De Santis, M.A.; Giuliani, M.M.; Giuzio, L.; De Vita, P.; Flagella, Z. Assessment of Grain Protein Composition in Old and Modern Italian Durum Wheat Genotypes. Ital. J. Agron. 2018, 40–43. [Google Scholar] [CrossRef]
- Padalino, L.; Mastromatteo, M.; Lecce, L.; Spinelli, S.; Contò, F.; Del Nobile, M.A. Chemical Composition, Sensory and Cooking Quality Evaluation of Durum Wheat Spaghetti Enriched with Pea Flour. Int. J. Food Sci. Technol. 2014, 49, 1544–1556. [Google Scholar] [CrossRef]
- D’egidio, M. G.; Mariani, B. M.; Nardi, S.; Novaro, P.; Cubadda, R. , Chemical and Technological Variables and Their Relationships: A Predictive Equation for Pasta Cooking Quality. Cereal Chem. 673 275-281 1990, 67, 275–281. [Google Scholar]
- De Vita, P.; Pecchioni, N. Considerazioni CREA Sulla Qualità Del Grano Italiano, Con Particolare Riferimento Contenuto Proteico.; Roma, 2016; p. 12.
- Arzani, A. Chapter 7 - Emmer (Triticum Turgidum Ssp. Dicoccum) Flour and Bread. In Flour and Breads and their Fortification in Health and Disease Prevention, 2nd ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press, 2019; pp. 89–98. ISBN 978-0-12-814639-2. [Google Scholar]
- Peña, R.J.; Trethowan, R.; Pfeiffer, W.H.; Van Ginkel, M. Quality (End-Use) Improvement in Wheat. J. Crop Prod. 2002, 5, 1–37. [Google Scholar] [CrossRef]
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops That Feed the World 6. Past Successes and Future Challenges to the Role Played by Maize in Global Food Security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- Dexter, J.E.; Matsuo, R.R. INFLUENCE OF PROTEIN CONTENT ON SOME DURUM WHEAT QUALITY PARAMETERS. Can. J. Plant Sci. 1977, 57, 717–727. [Google Scholar] [CrossRef]
- Boyacioglu, M. H.; D’appolonia, B. L. Characterization and Utilization of Durum Wheat for Breadmaking. II. Study of Flour Blends and Various Additives. Cereal Chem. 1994, 71, 28–33. [Google Scholar]
- Sissons Mike; Pleming Denise,; Margiotta Benedetta,; D’Egidio Maria Grazia; Lafiandra Domenico Effect of the Introduction of D-Genome Related Gluten Proteins on Durum Wheat Pasta and Bread Making Quality. Crop Pasture Sci. 2014, 65, 27–37. [CrossRef]
- Liu, C-Y; Shepherd, K. W.; Rathjen, A.J. Improvement of Durum Wheat Pastamaking and Breadmaking Qualities. Cereal Chem 1996, 73, 155–166. [Google Scholar]
- Morris, C.F.; Casper, J.; Kiszonas, A.M.; Fuerst, E.P.; Murray, J.; Simeone, M.C.; Lafiandra, D. Soft Kernel Durum Wheat—A New Bakery Ingredient? Cereal Foods World 2015, 60, 76–83. [Google Scholar] [CrossRef]
- Boehm, J.D.; Zhang, M.; Cai, X.; Morris, C.F. Molecular and Cytogenetic Characterization of the 5DS–5BS Chromosome Translocation Conditioning Soft Kernel Texture in Durum Wheat. Plant Genome 2017, 10, plantgenome2017.04.0031. [Google Scholar] [CrossRef]
- Schlchting, L.M.; Dexter, J.E.; Preston, K.R.; Edwards, N.M.; Marchylo, B.A.; Hucl, P. Quality Characterization of Linesfrom a Cross between Emmer (T. Dicoccum) and Durum(T. Durum)Wheat. In Proceedings of 2nd International Workshop.Durum Wheat and Pasta Quality: Recent Achievements and NewTrends; D’Egidio, M.G., Ed.; Industria Grafica Failli Fausto: Guidonia-Montecelio, Italy, 2003; pp. 71–76. [Google Scholar]
- Marchylo, B.A.; Dexter, J.E.; Clarke, F.R.; Clarke, J.M.; Preston, K.R. Relationships among Bread-Making Quality, Gluten Strength, Physical Dough Properties, and Pasta Cooking Quality for Some Canadian Durum Wheat Genotypes. Can. J. Plant Sci. 2001, 81, 611–620. [Google Scholar] [CrossRef]
- Scarascia-Mugnozza, G.T. The Contribution of Italian Wheat Geneticists: From Nazareno Strampelli to Francesco D’Amato. In Proceedings of the International Congress ‘in the Wake of the Double Helix: From the Green Revolution to the Gene Revolution’; Tuberosa, R., Phillips, R.L., Gale, M., Eds.; Avenue Media: Bologna, Italy, 2005; pp. 53–75. [Google Scholar]
- Dinelli, G.; Marotti, I.; Silvestro, R.D.; Bosi, S.; Bregola, V.; Accorsi, M.; Loreto, A.D.; Benedettelli, S.; Ghiselli, L.; Catizone, P. Agronomic, Nutritional and Nutraceutical Aspects of Durum Wheat (Triticum Durum Desf.) Cultivars under Low Input Agricultural Management. Ital. J. Agron. 2013, 8. [Google Scholar] [CrossRef]
- Reynolds, M.; Foulkes, J.; Furbank, R.; Griffiths, S.; King, J.; Murchie, E.; Parry, M.; Slafer, G. Achieving Yield Gains in Wheat. Plant Cell Environ. 2012, 35, 1799–1823. [Google Scholar] [CrossRef]
- Berry, P.M.; Sylvester-Bradley, R.; Berry, S. Ideotype Design for Lodging-Resistant Wheat. Euphytica 2007, 154, 165–179. [Google Scholar] [CrossRef]
- Donmez, E.; Sears, R.G.; Shroyer, J.P.; Paulsen, G.M. Genetic Gain in Yield Attributes of Winter Wheat in the Great Plains. Crop Sci. 2001, 41, 1412–1419. [Google Scholar] [CrossRef]
- Hunt, J.R.; Lilley, J.M.; Trevaskis, B.; Flohr, B.M.; Peake, A.; Fletcher, A.; Zwart, A.B.; Gobbett, D.; Kirkegaard, J.A. Early Sowing Systems Can Boost Australian Wheat Yields despite Recent Climate Change. Nat. Clim. Change 2019, 9, 244–247. [Google Scholar] [CrossRef]
- Asseng, S.; Zhu, Y.; Wang, E.; Zhang, W. Crop Modeling for Climate Change Impact and Adaptation. In Crop physiology; Elsevier, 2015; pp. 505–546.
- Asseng, S.; Kheir, A.M.S.; Kassie, B.T.; Hoogenboom, G.; Abdelaal, A.I.N.; Haman, D.Z.; Ruane, A.C. Can Egypt Become Self-Sufficient in Wheat? Environ. Res. Lett. 2018, 13, 094012. [Google Scholar] [CrossRef]
- Fischer, R.A. Wheat Physiology: A Review of Recent Developments. Crop Pasture Sci. 2011, 62, 95. [Google Scholar] [CrossRef]
- Rharrabti, Y.; Villegas, D.; García del Moral, L.F.; Aparicio, N.; Elhani, S.; Royo, C. Environmental and Genetic Determination of Protein Content and Grain Yield in Durum Wheat under Mediterranean Conditions. Plant Breed. 2001, 120, 381–388. [Google Scholar] [CrossRef]
- Acreche, M.M.; Slafer, G.A. Variation of Grain Nitrogen Content in Relation with Grain Yield in Old and Modern Spanish Wheats Grown under a Wide Range of Agronomic Conditions in a Mediterranean Region. J. Agric. Sci. 2009, 147, 657–667. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Nemeh, A.; Pruneddu, G. Durum Wheat Cultivars Grown in Mediterranean Environments Can Combine High Grain Nitrogen Content with High Grain Yield. Eur. J. Agron. 2022, 136, 126512. [Google Scholar] [CrossRef]
- Bogard, M.; Allard, V.; Brancourt-Hulmel, M.; Heumez, E.; Machet, J.-M.; Jeuffroy, M.-H.; Gate, P.; Martre, P.; Le Gouis, J. Deviation from the Grain Protein Concentration–Grain Yield Negative Relationship Is Highly Correlated to Post-Anthesis N Uptake in Winter Wheat. J. Exp. Bot. 2010, 61, 4303–4312. [Google Scholar] [CrossRef]
- Mi, G.; Tang, L.; Zhang, F.; Zhang, J. Is Nitrogen Uptake after Anthesis in Wheat Regulated by Sink Size? Field Crops Res. 2000, 68, 183–190. [Google Scholar] [CrossRef]
- Royo, C.; Elias, E.M.; Manthey, F.A.; Carena, M.J. Cereals—Handbook of Plant Breeding. 2009.
- Sciacca, F.; Allegra, M.; Licciardello, S.; Roccuzzo, G.; Torrisi, B.; Virzì, N.; Brambilla, M.; Romano, E.; Palumbo, M. Potential Use of Sicilian Landraces in Biofortification of Modern Durum Wheat Varieties: Evaluation of Caryopsis Micronutrient Concentrations. Cereal Res. Commun. 2018, 46, 124–134. [Google Scholar] [CrossRef]
- Hernandez-Espinosa, N.; Laddomada, B.; Payne, T.; Huerta-Espino, J.; Govindan, V.; Ammar, K.; Ibba, M.I.; Pasqualone, A.; Guzman, C. Nutritional Quality Characterization of a Set of Durum Wheat Landraces from Iran and Mexico. LWT 2020, 124, 109198. [Google Scholar] [CrossRef]
- Magallanes-López, A.M.; Hernandez-Espinosa, N.; Velu, G.; Posadas-Romano, G.; Ordoñez-Villegas, V.M.G.; Crossa, J.; Ammar, K.; Guzmán, C. Variability in Iron, Zinc and Phytic Acid Content in a Worldwide Collection of Commercial Durum Wheat Cultivars and the Effect of Reduced Irrigation on These Traits. Food Chem. 2017, 237, 499–505. [Google Scholar] [CrossRef]
- Connor, D.J.; Loomis, R.S.; Cassman, K.G. Crop Ecology: Productivity and Management in Agricultural Systems; Cambridge University Press, 2011; ISBN 1-139-50032-5. [Google Scholar]
Figure 1.
Schematic representation of durum wheat from the syriacum typicum and mediterraneum typicum groups (modified from Ali Dib et al. [9]).
Figure 1.
Schematic representation of durum wheat from the syriacum typicum and mediterraneum typicum groups (modified from Ali Dib et al. [9]).
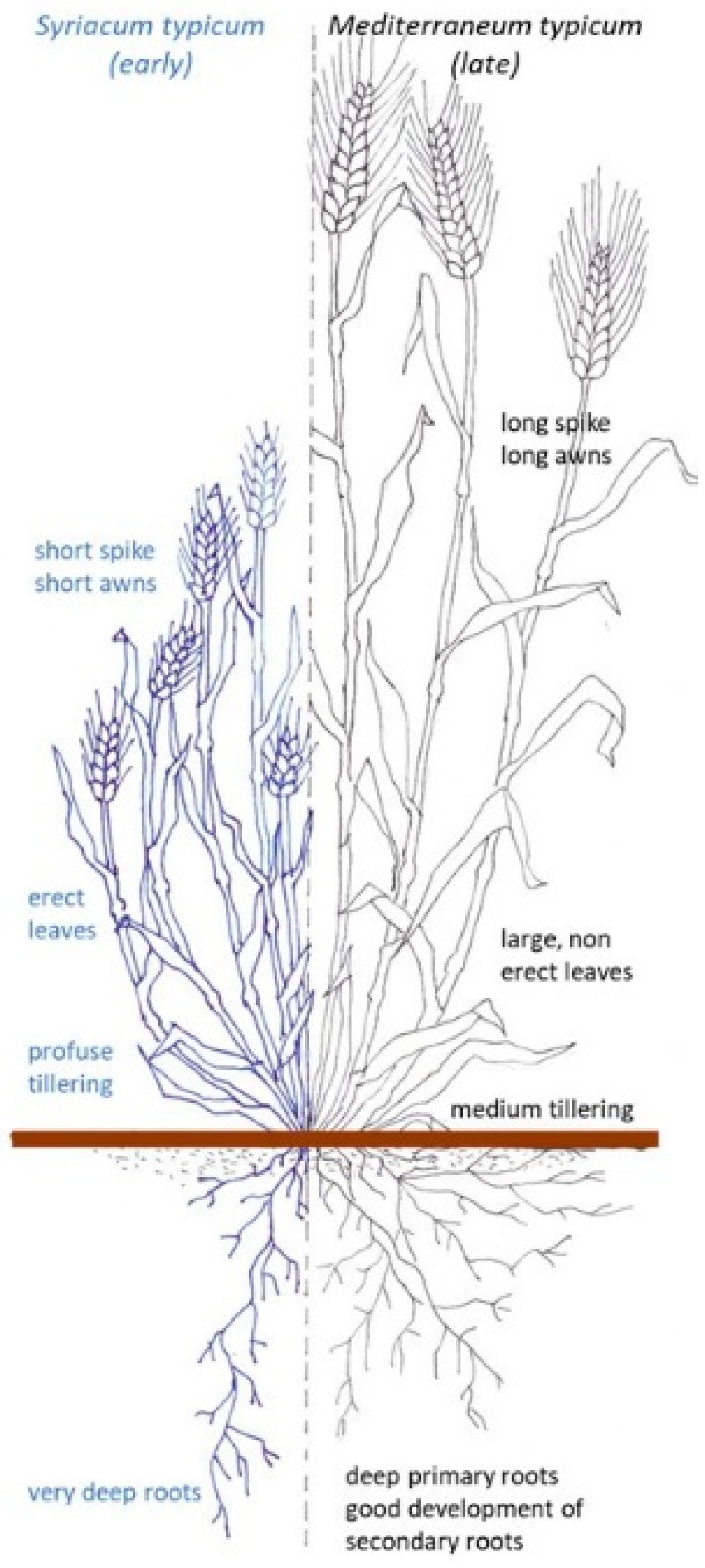
Figure 2.
Evolution in surface area and grain yield according to ISTAT data. Data are means of five-year periods.
Figure 2.
Evolution in surface area and grain yield according to ISTAT data. Data are means of five-year periods.
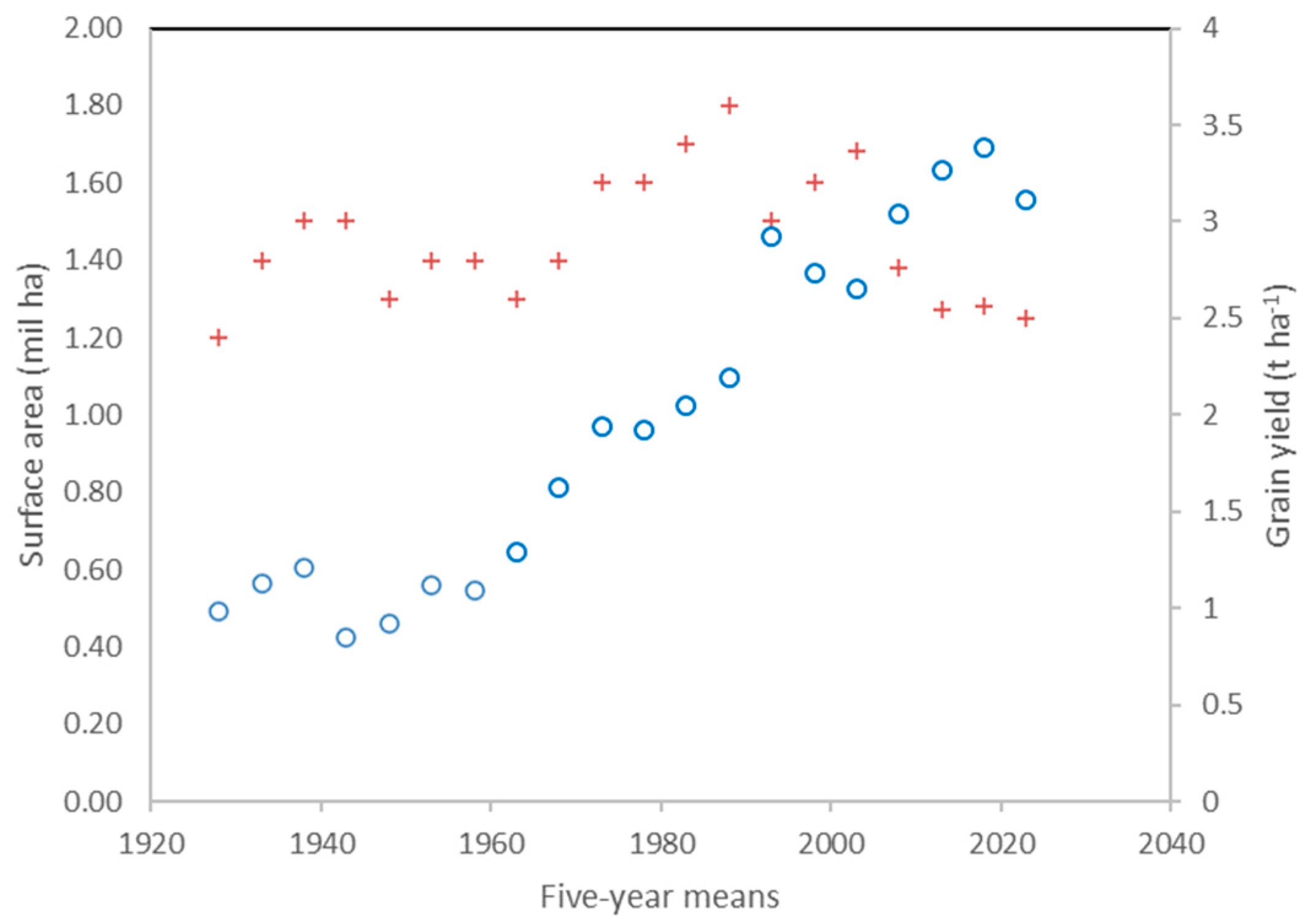
Figure 3.
Productivity indices (yield expressed as a percentage of the controls) of the best cultivars within each five-year period of constitution and average grain yield for the trials in which they were present (modified from Salis [52]).
Figure 3.
Productivity indices (yield expressed as a percentage of the controls) of the best cultivars within each five-year period of constitution and average grain yield for the trials in which they were present (modified from Salis [52]).
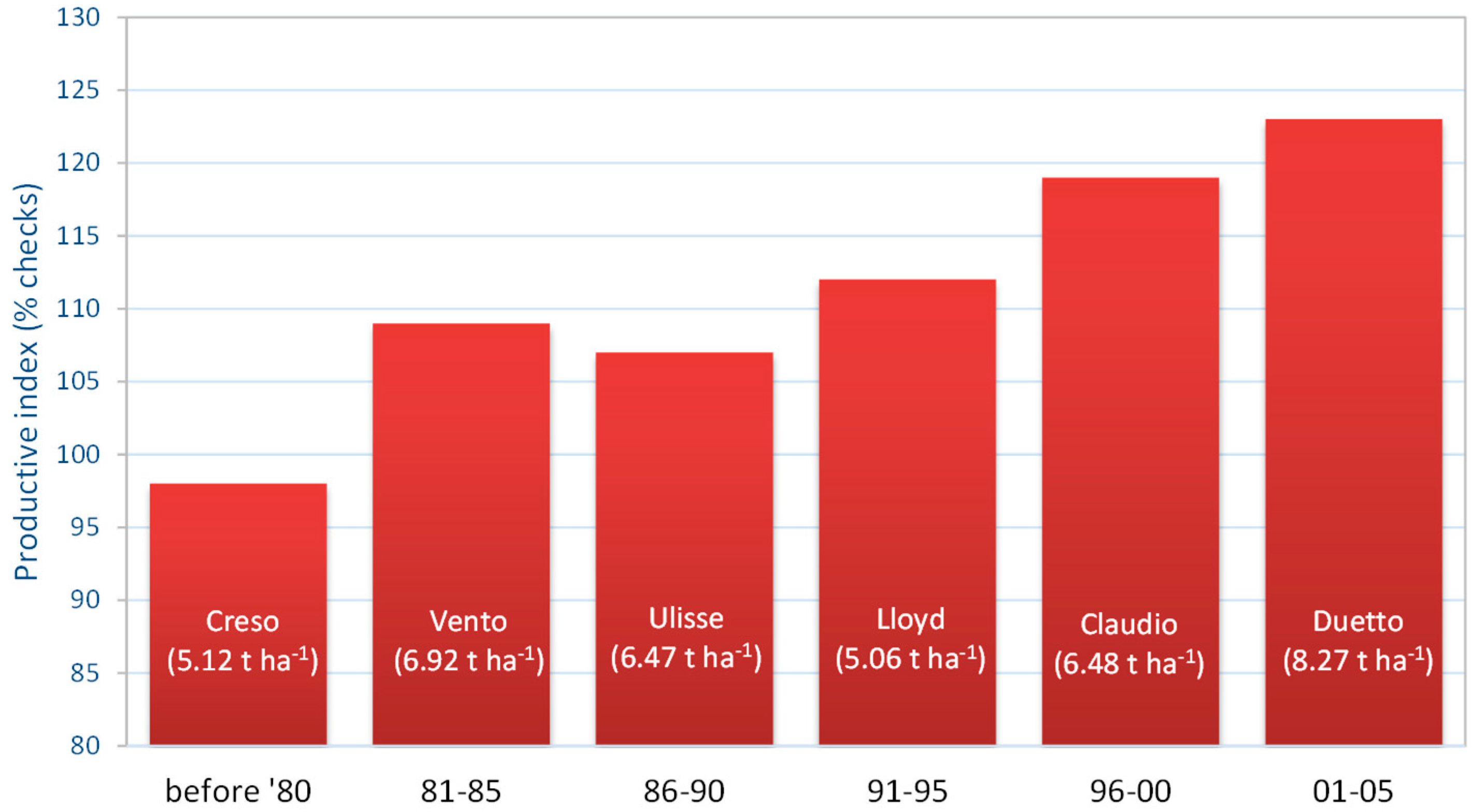
Figure 4.
Potential, attainable, and farm yields in two Sardinian sites in absolute values and as percentage (numbers inside the bars). Potential yield was considered equal to the record yield in the 20 years of experimental trials, i.e. 10.56 t ha-1, attainable yields were field means, farm yields were obtained by ISTAT (modified from Salis [52]).
Figure 4.
Potential, attainable, and farm yields in two Sardinian sites in absolute values and as percentage (numbers inside the bars). Potential yield was considered equal to the record yield in the 20 years of experimental trials, i.e. 10.56 t ha-1, attainable yields were field means, farm yields were obtained by ISTAT (modified from Salis [52]).
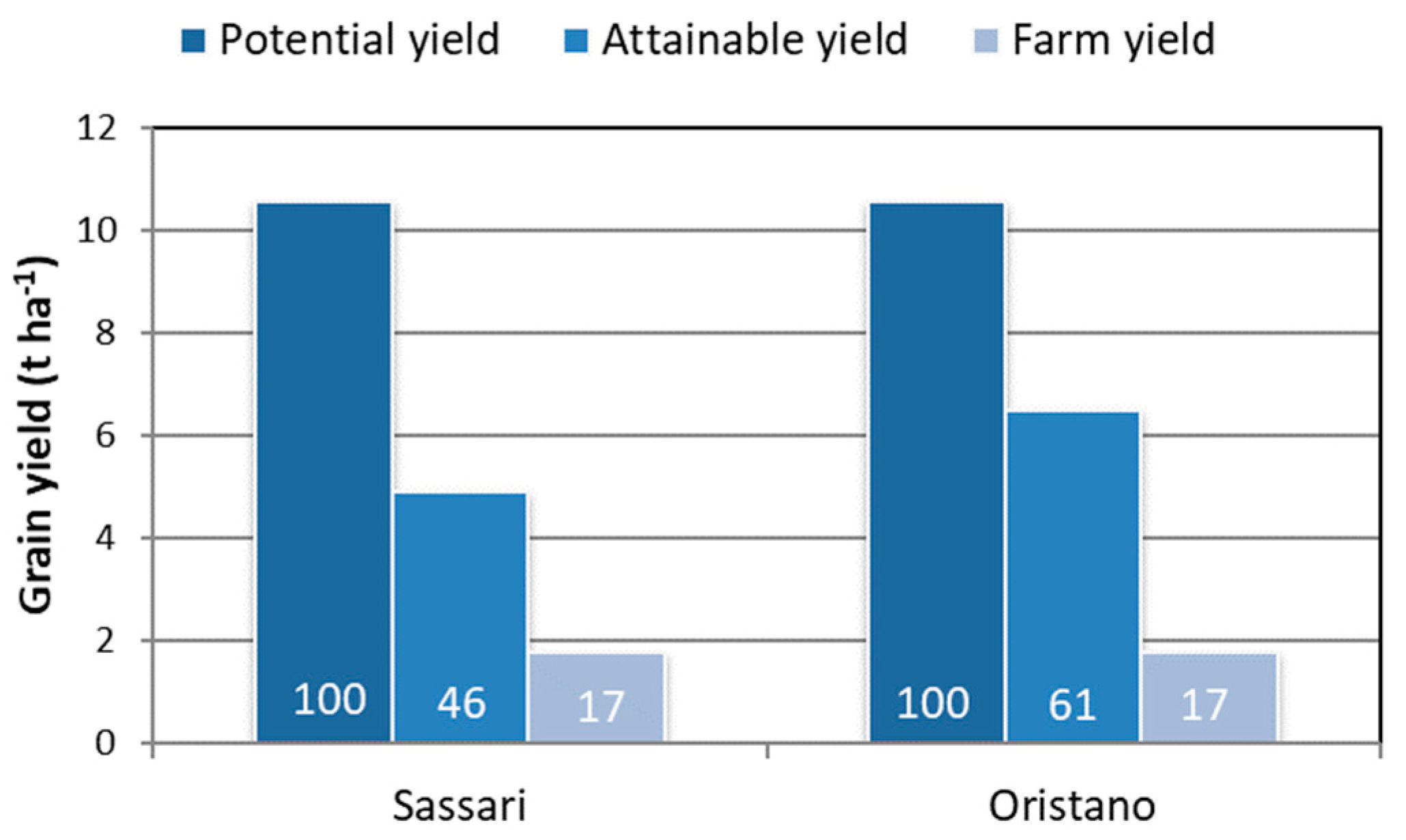

Table 1.
The main landraces of durum wheat cultivated in Italy until the 1920s.
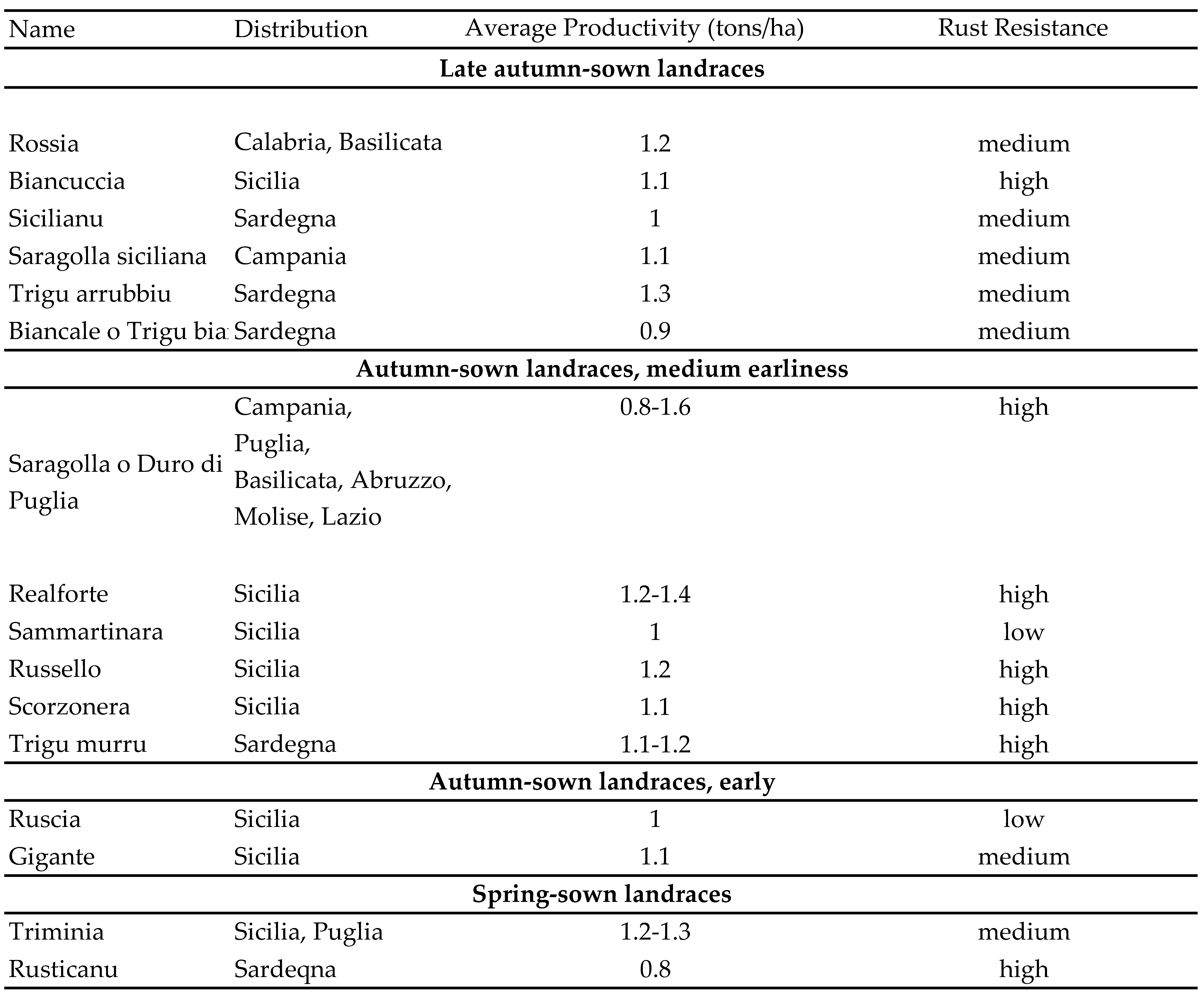 |
Table 2.
Durum wheat cultivars based on single pure lines developed in Italy between the 1920s and 1950s through genealogical selection from landraces (exception “Garigliano”).
Table 2.
Durum wheat cultivars based on single pure lines developed in Italy between the 1920s and 1950s through genealogical selection from landraces (exception “Garigliano”).
 |
Table 3.
Durum wheat cultivars selected in Italy between the 1950s and the first half of the 1970s from segregating populations obtained through inter- and intra- specific hybridization.
Table 3.
Durum wheat cultivars selected in Italy between the 1950s and the first half of the 1970s from segregating populations obtained through inter- and intra- specific hybridization.
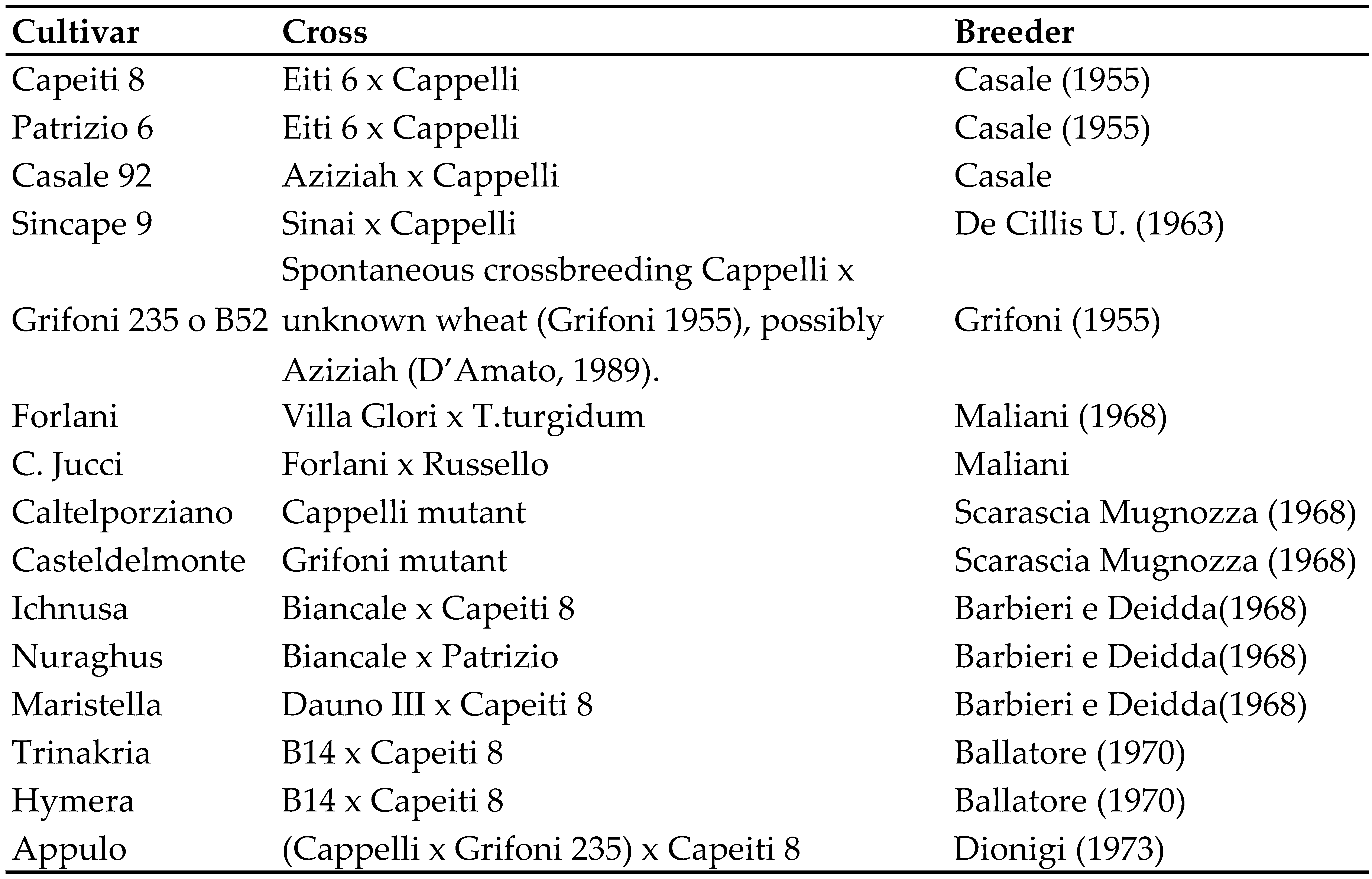 |
Table 4.
Main semi-dwarf cultivars released between 1974 and 2000.
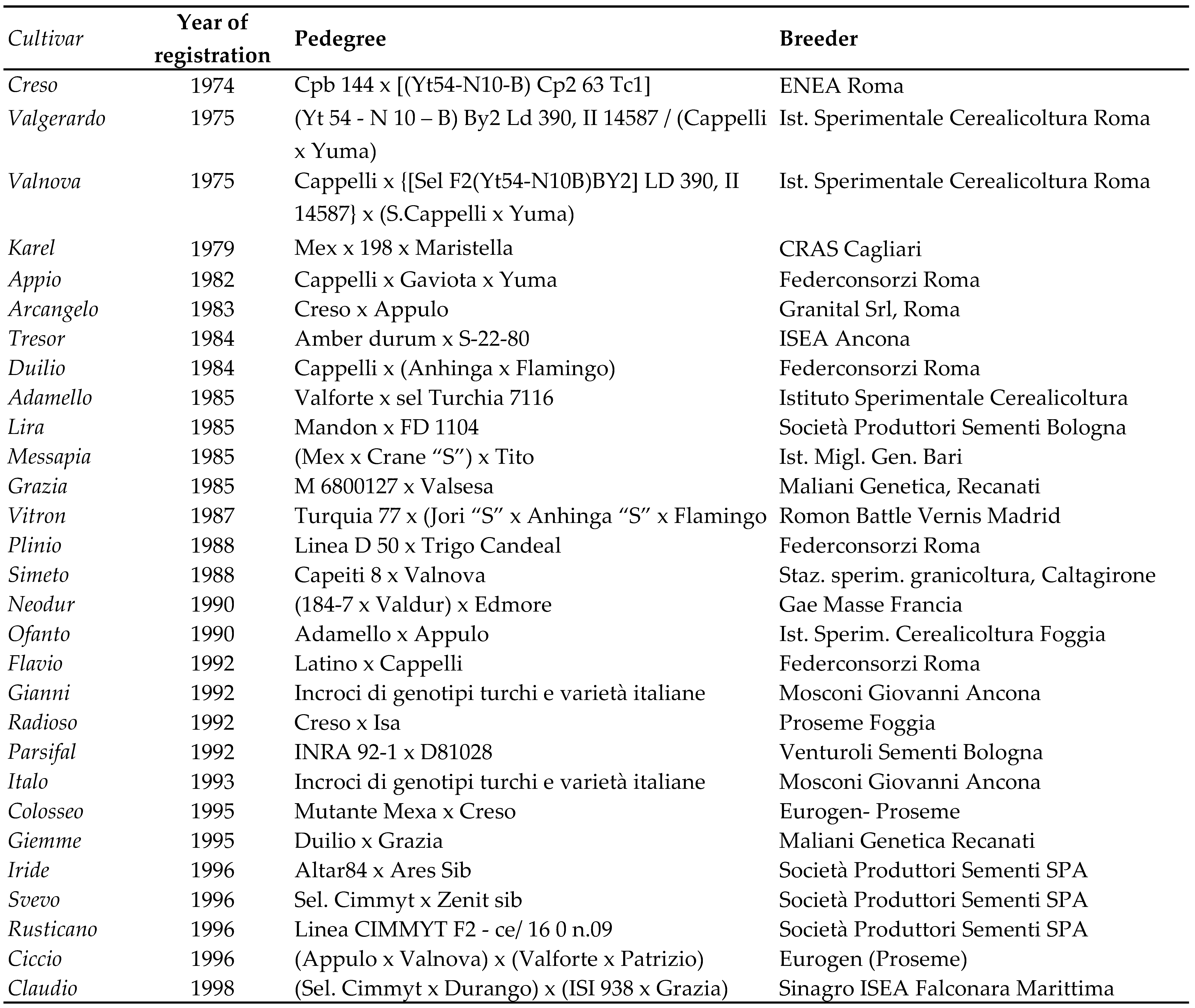 |
Table 5.
The percentage of seed production for the top ten cultivars over the total production of the top ten cultivars between 1992 and 2010 in Italy (ISTAT data).
Table 5.
The percentage of seed production for the top ten cultivars over the total production of the top ten cultivars between 1992 and 2010 in Italy (ISTAT data).
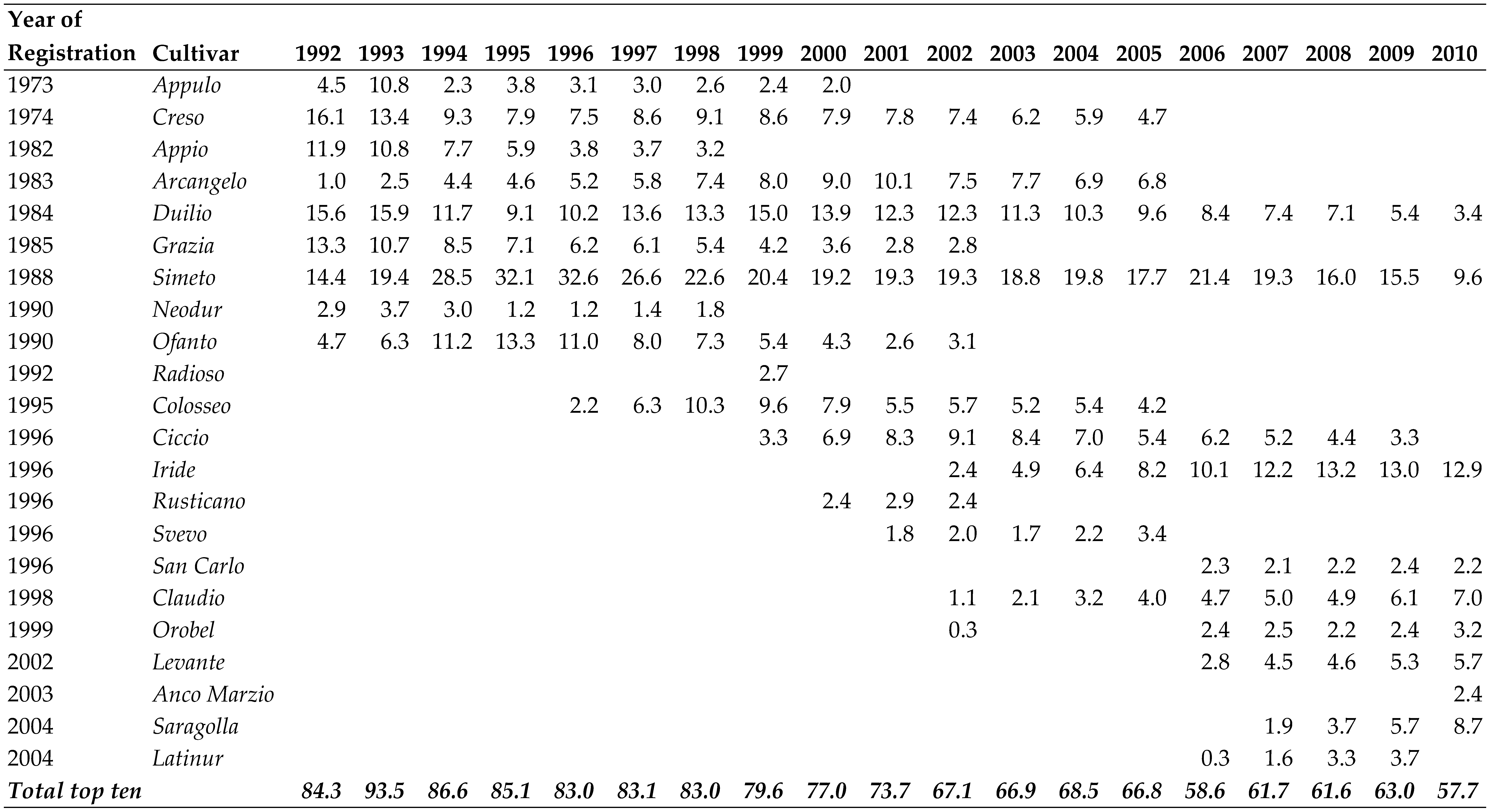 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



