
Submitted:
20 November 2023
Posted:
21 November 2023
You are already at the latest version
Abstract
The Vitamin D Receptor (VDR) mediates the actions of vitamin D, which has important roles in bone homeostasis, growth/differentiation of cells, immune functions and reduction of inflammation. Emerging evidences suggest that epigenetic modifications of the VDR gene, particularly DNA methylation, may contribute to the onset and progression of many human disorders. This review aims to summarize the available information on the role of VDR methylation signatures in different pathological contexts, including autoimmune diseases, infectious diseases, cancer and others. The reversible nature of DNA methylation could enable the development of therapeutic strategies, offering new avenues for the management of these worldwide diseases.
Keywords:
Vitamin D receptor (VDR)
; Epigenetics
; DNA methylation
; VDR-related diseases
1. Introduction
The Vitamin D receptor (VDR) belongs to the nuclear receptor superfamily and can mediate the actions of the biologically active form of vitamin D [1]. VDR, together with the Retinoid X receptor (RXR), forms a heterodimer capable of regulating more than 900 genes [2]. VDR-RXR binds vitamin D-sensitive positive or negative promoter elements (VDRE and nVDRE), which are made up of repeated core binding motifs [3]. When VDR-RXR binds to VDRE or nVDRE, it can activate or repress the transcription of target genes, by recruiting transcriptional coactivators and corepressor, respectively [4]. This VDR-induced regulation orchestrates key biological processes, including tumor progression, the immune system regulation, cell proliferation and differentiation [5]. The VDR gene is located on the long arm of chromosome 12 (12q12-q14) and spans a genomic region of approximately 75 Kb. The gene comprises eight exons (2-9) that code for a protein of 427 amino acids, while six noncoding exons (1a-1f) are located in the 5’ regulatory region [6,7] (Figure 1). The human VDR primary promoter spans exon 1a and is characterized by the lack of a TATA initiator box, its GC-rich nature, and the presence of putative binding sites for a variety of transcription factors [6]. Three other alternative promoters are present in the VDR gene, located at exons 1f, 1c and 9, some of which are tissue-specific [3] (Figure 1). Genetic variability, along with epigenetic modifications, can modulate VDR expression, contributing to the onset and progression of several diseases, generally known as “VDR-related” [3,8]. Among the epigenetic signatures, DNA methylation appears to be the most suitable biomarker for the detection and treatment of human diseases. DNA methylation is a mechanism that involves the transfer of a methyl group to the C5 position of cytosine, forming 5-methylcytosine, by enzymes belonging to the DNA methyltransferases (DNMTs), capable of transferring CH3 from S-adenyl Methionine (SAM) to the cytosine residue [9]. According to the predictive model of CpG islands (CGIs) made by Bock and colleagues [10], the VDR methylation can occurs on 9 bona fide CGIs, which lie along the promoter regions and the gene body. In particular, two CGIs seem to play a crucial role in human diseases, CGI 1062 which overlaps exon 1a and comprises 56 CpG sites, and CGI 1060 overlapping with one of the alternative promoters (Figure 1) [10,11]. In addition, other GC-rich regions within the VDR gene have been identified using predictive methylation analytical tools, such as DataBase of CpG islands and Analytical Tool (DBCAT) (Figure 1) [12]. Given the complexity of the CpG content in the VDR gene sequence, the global characterization of the methylation pattern in different pathological conditions remains a major challenge to be fully explored. Indeed, dynamic VDR methylation signatures in specific CpGs may play a pivotal role in both the diagnosis and prognosis of several VDR-related diseases. The main purpose of this literature review is to provide an up-to-date overview of the role of VDR methylation in different human disorders, including autoimmune and infectious diseases, cancer and others. A deeper understanding of disease-specific epigenetic patterns is critical to developing targeted and effective novel epigenetic therapies to improve the clinical management and responses to vitamin D.
2. Search Strategy
A global search of publications was conducted using the international bibliographic database PubMed. The identified articles were selected until July 2023. The keywords used for search were: “VDR methylation”, “VDR methylation in diseases”, “VDR methylation in autoimmune diseases”, “VDR methylation AND Rheumatoid Arthritis”, “VDR methylation AND Multiple Sclerosis”, “VDR methylation AND Bechet’s disease”, “VDR methylation in infectious diseases”, “VDR methylation AND Tuberculosis disease”, “VDR methylation AND Hand Foot and Mouth disease”, “VDR methylation AND HIV”, “VDR methylation in cancer”, “VDR gene methylation AND breast cancer”, “VDR methylation AND Adrenocortical carcinoma”, “VDR methylation AND Hepatocellular carcinoma”, “VDR methylation AND Parathyroid adenomas”, “VDR methylation AND colorectal cancer”, “VDR methylation AND Osteoporosis”, “VDR methylation AND Male infertility”, “VDR methylation AND Recurrent Kidney stone formation”, “VDR methylation AND Type 2 Diabetes Mellitus”. The search was conducted without restriction of ethnicity or geographic area and language filters were applied to eliminate non-English language articles. In addition, the following exclusion criteria were used: abstracts, short communications, letters to the editor and dissertations.
3. Autoimmune Diseases
Vitamin D modulates immune responses by regulating B and T-cell (TCs) proliferation and the production of inflammatory cytokines and immunoglobulin, including auto-antibodies [13]. Vitamin D plays a role in the activity of CD4+ TCs, enhancing the generation of regulatory T cells (Tregs) and Th2 cells and suppressing pro-inflammatory TCs, specifically Th17 and Th1 [13,14]. This regulatory mechanism is able of creating a tolerogenic immune status which can be altered when the level of vitamin D decreases, leading to immune dysregulation and increased susceptibility to autoimmune disorders [15,16]. The function of the vitamin D is mediated by its binding with VDR on the membrane of immune system cells such as dendritic cells, monocytes and B and T lymphocytes [17]. Consequently, the genetic variability and epigenetic modifications in the VDR could alter immune homeostasis, significantly impacting the onset and progression of autoimmune diseases, such as Rheumatoid Arthritis (RA), Multiple Sclerosis (MS) and Behcet's Disease (BD) (Table 1) [18,19,20,21]. RA is a progressive autoimmune disease characterized by chronic joint inflammation and pain, attributed to the influx of CD4+ TCs into the synovial lining and the increase of macrophage-like and fibroblast-like cells that produce degradative enzymes and proinflammatory cytokines [22]. Although many studies have investigated the role of VDR genetic variability in RA susceptibility, only a limited number of researches have been focused on the epigenetic signatures of this gene. Zhang et al., investigated the methylation levels of the VDR primary promoter in a Chinese cohort comprising 122 RA patients and 123 controls (CTRs). The study identified a significant reduction of VDR methylation levels in RA patients compared to CTRs, suggesting that this signature may represent a potential disease biomarker [23]. In contrast with these results, a second epigenetic study revealed no significant differences in the methylation levels of 10 CpG sites within the VDR primary promoter between a group of 35 RA patients and 41 healthy subjects [19]. However, this controversial result could be attributed to both the small size of the study cohort and the small number of CpG sites analyzed. Vitamin D and its receptor also play a pivotal role in multiple sclerosis (MS), as low levels of vitamin D or alterations in the VDR gene are a risk factor for this condition [24]. MS is a chronic and severe systemic autoimmune disease with abnormal immune system reactions, which damage central nervous system components [25]. The pathogenesis is still unclear and recent data argue in favor of an involvement of epigenetic mechanisms in MS susceptibility [26]. So far, only one study has investigated the role of VDR methylation in T cells from relapsing-remitting MS (RRMS) patients compared to CTRs. The results revealed a significant increase in DNA methylation levels within an alternative promoter of the VDR, while the VDR primary promoter remains extensively unmethylated [20]. The specific pattern of VDR methylation in the alternative promoter at exon 1c suggests an important role of this region in MS susceptibility, to be further explored. One hypothesis suggested by the authors is the presence of a 4-exon transcript (GenBank accession: AK129594), which is a natural antisense transcript (NAT), that could modulate VDR expression levels also trough self-regulatory circuits, but this regulatory mechanism is still unclear [20]. Epigenetic changes of the VDR gene have also been investigated in Behcet's disease (BD), a chronic recurrent multisystem inflammatory condition caused by several pro-inflammatory cytokines regulated by vitamin D [21]. The study by Shirvani et al., analyzed the methylation levels of CpG sites in the VDR primary promoter (800 bp upstream and 200 bp downstream of the ATG start site) in peripheral blood mononuclear cells (PBMCs) from BD patients respect to CTRs. Despite the decreased VDR expression levels observed in BD patients, the methylation level of the VDR gene reflect no differences between the two groups. This result may be explained by the effects of other GC-rich regions along the VDR gene not analyzed in this study or by additional epigenetic mechanisms leading to the observed VDR downregulation [27]. Overall, the study on the role of VDR methylation pattern in autoimmune diseases remains still a controversial and under-explored area in current research. The main lack may derive from the complexity of these disorders, whose underlying mechanisms have yet to be fully characterized.
4. Infectious Diseases
Vitamin D plays a pivotal role also in innate immune response being able to stimulate immune cells to fight pathogens, such as Mycobacterium tuberculosis, enterovirus 71 and human immunodeficiency virus (HIV) [31,42,43]. The vitamin D-bound VDR induces the transcription of genes encoding antimicrobial peptides, such as cathelicidin and β-defensin 2, that contain VDRE in their promoters. These peptides exhibit broad-spectrum antimicrobial activity against bacteria, viruses, and fungi and their decrease has often been linked to low Vitamin D levels and increased susceptibility to infectious diseases [44,45,46]. A close interconnection exists between the progression of Tuberculosis (TB) and deficiency of vitamin D [47]. TB is caused by Mycobacterium tuberculosis infection and its elimination depends on expression of several antimicrobial peptides and cytokines, which are activated by VDR [47,48]. A pilot study by Yang et al., identified an inverse correlation between the VDR hypermethylation pattern and the expression level of VDR, IL-1β, IL-6 and TNF-α genes in RAW 264.7 cells previously infected with Mycobacterium, suggesting a protective effect of VDR hypomethylation signature against Mycobacterium infection [49]. A subsequent study performed by the same research group analyzed VDR methylation levels of a 269 bp region in the primary promoter, comprising 16 CpG sites, in TB patients compared to CTRs. Results showed an increase in VDR methylation levels in TB patients vs. CTRs, confirming the potential involvement of this factor in the pathogenesis and progression of TB [28,49] (Table 1). Accordingly, another investigation carried out in children with active TB demonstrated an inverse correlation between VDR methylation and its expression levels. Most importantly, hypermethylation of CpG sites in the VDR promoter region showed a significant diagnostic accuracy in discriminating TB disease, as determined by Receiver Operating Characteristics (ROC) curve analysis [37] (Table 1). All these evidences support the hypothesis that increased VDR methylation leads cells of innate immunity to secrete insufficient antimicrobial peptides to fight TB disease [12]. Only one epidemiological study conducted by Wang et al. in a Chinese population does not support this model. The research analyzed 60 CpG sites within the VDR primary promoter in 122 TB patients vs. 118 CTRs showing an opposite pattern of methylation respect to the previous reports: VDR promoter hypomethylation in TB patients and VDR promoter hypermethylation in CTRs (Table 1). The discrepancy in the VDR methylation pattern observed in these three TB studies could result from the different regions of the VDR analyzed. While the first two studies analyzed the VDR methylation in alternative promoter regions Wang et al., focused their attention on the primary promoter. However, the latest study did not assess the downstream immune responses modulated by the binding of vitamin D to VDR and the impact of VDR methylation on its expression levels [29]. The methylation of VDR have also been implicated in the gravity of Hand, Foot, and Mouth Disease (HFMD) caused by enterovirus 71 (EV71) (Table 1) [30]. Li et al., evaluated the methylation status of VDR primary promoter (from -638 bp to -545 bp of the ATG start site) in 58 children with severe HFMD, 58 children with mild HFMD, and 60 matched healthy controls. The results have shown a hypermethylation of VDR primary promoter in mild HFMD patients compared to children with severe HFMD, suggesting that VDR methylation is an early event in EV71-associated HFMD [30]. Among infectious diseases, recent data on the role of VDR epigenetic mechanisms in HIV disease are also emerging (Table 1). HIV is a retrovirus capable of infecting human cells, leading to an acquired immunodeficiency syndrome (AIDS) with a weakened immune system [50]. HIV seems to promote VDR methylation at its promoter region, inducing down-regulation of VDR expression levels in TCs [31]. A pilot study analyzed the methylation levels of CpG sites in the VDR primary promoter region in HIV-infected TCs, showing increased methylation of CpG sites and increased expression of DNA methyltransferase 3b (DNMT3b) [31]. HIV-induced hypermethylation of VDR in TCs led to a reduction in VDR expression levels which promotes TCs apoptosis [31]. In conclusion, the majority of the studies reported here agree that VDR hypermethylation is a hallmark of increased susceptibility to infectious diseases. The identification of the complete VDR epigenetic signature is therefore essential to understand the maintaining of a balanced immune homeostasis essential to counteract efficiently pathogens.
5. Cancer
The development and progression of cancer depend on multiple molecular mechanisms in which vitamin D and its receptor are highly involved [51]. In the pathological context of cancer, the VDR can directly or indirectly regulate the expression of >200 genes that influence cell proliferation, differentiation and apoptosis, as well as immunomodulation and angiogenesis [52,53]. Several molecular pathways in cancer development are intimately linked to vitamin D and VDRE regions. c-MYC, a protein that regulate the cell cycle often altered in cancer, contains VDRE in its promoter [54]. It was reported that ligand-activated VDR could downregulate c-MYC expression through a direct bond with two VDREs [54,55]. Low levels of vitamin D and 1,25-(OH)2-D3/VDR can also limit the hyperproliferative action of β-catenin in the epidermal cancer, interfering with the essential Wnt/β-catenin signaling [54,56]. Since genetic and epigenetic changes can influence the risk of developing cancer, many studies have been focused on the epigenetic signatures of the VDR gene [57]. Increased VDR methylation levels have been observed in several cancer types and lead to reduced mRNA and protein expression levels [32,57], supporting the loss of an antiproliferative role of VDR [58]. VDR hypermethylation has been associated with hepatocellular carcinoma (HCC) [34], adrenocortical carcinoma (ACC) [33], colorectal cancer [36] and breast cancers [32] (Table 1). Breast cancer is the most common cancer among women, affecting millions of people worldwide [59]. Its development is influenced by a complex interplay of genetic, epigenetic and environmental factors [60]. 1α, 25(OH)(2)-Vitamin D3, by binding to its receptor, regulates the differentiation of the normal mammary gland and may be a useful parameter in treating or preventing breast cancer [32,61]. However, many breast cancer cells are resistant to 1α, 25(OH)(2)-Vitamin D3, but how this insensitivity may be involved in pathogenesis remains unclear [32]. Marik et al., analyzed the mechanism of resistance and the role of epigenetic silencing of VDR by promoter hypermethylation [32]. An in silico analysis of the VDR gene identified three CGIs in an area ranging from -789 bp upstream to +380 bp downstream of the transcription start site (TSS) relative to primary promoter [32]. The subsequent in vitro analysis conducted on different breast cancer and immortalized normal breast epithelial cell lines, previously treated with 1α, 25(OH)(2)-Vitamin D3, confirmed the insensitivity to calcitriol in breast cancer cells. Additionally, breast cancer cell lines showed an increase of VDR methylation levels than normal cells. To validate these preliminary data, the methylation analysis was also performed on breast cancer tissue compared to normal breast tissue. In breast cancers, VDR promoter was significantly hypermethylated (65%) than in normal breast tissue (15%), supporting the proposed mechanism of silencing VDR expression through promoter hypermethylation, which appears linked to 1α, 25(OH)(2)-Vitamin D3 sensitivity [32]. Aberrant VDR promoter methylation has also been observed in human ACC, resulting in dysregulation of steroid biosynthesis and adrenal growth [62,63]. A methylation analysis of a CGI located in VDR primary promoter, including 42 CpG sites, was performed on ACC samples, revealing a higher methylation pattern of 27 CpG sites from 3/8 ACC samples. In addition, hypermethylation of 27 VDR CpG sites has been correlated with a lower VDR mRNA expression [33]. Accordingly, another study showed that a percentage of VDR promoter methylation was significantly higher in the HCC group compared to both the Chronic Liver Disease (CLD) and CTRs [34]. In contrast, Varshney et al., reported no promoter methylation of the VDR in parathyroid adenomas although a 93% decrease in VDR expression in the same samples, highlighting the hypothesis of other mechanisms that may regulate VDR levels [35]. Colorectal cancer is a third example in which the role of VDR promoter hypermethylation has been investigated. Colorectal carcinogenesis results from genetic and epigenetic changes as intestinal epithelial cells progress from average to malignant phenotype [64]. A significant association was found between VDR methylation and VDR expression in colorectal cancer progression [36]. A study conducted by Afshan and colleagues in 75 colorectal cancer samples has detected the hypermethylation of the VDR promoter in 28 (37.33%) of 75 cases [36]. Furthermore, methylation levels and decreased VDR expression were correlated with the different grades and stages of carcinoma progression, from poorly differentiated colorectal cancer to advanced stages [36]. Overall, studies conducted to date agree that VDR hypermethylation may favour cancer onset and progression, however these analyses have been performed on a relatively small number of samples in few cancer types. It is therefore necessary to fill these gaps before considering VDR methylation signature as a biomarker of different grades and stages of cancer progression.
6. Other Diseases
Vitamin D deficiency is a common background in many other human disorders, such as bone diseases, male infertility, type 2 diabetes mellitus (T2DM) and kidney stone formation [65,66,67,68]. The shared low vitamin D status prompted researchers to investigate the epigenetic signatures of VDR in all these pathological conditions (Table 1). Recent studies have explored DNA methylation pattern of several genes, including VDR, involved in osteogenic differentiation, osteogenesis, bone remodelling and other bone metabolism-related processes [69,70]. To address this topic, our research group investigated the interconnections between transcriptomic profile and epigenetics of the VDR gene in a Caucasian cohort consisting of 25 osteoporotic patients (OPs) and 25 healthy subjects (CTRs) [37]. Our analysis identified lower VDR expression levels in PBMCs from OPs compared to CTRs, which positively correlates with BMD and t-score values, thus associating with a more severe pathological phenotype. VDR altered expression levels led us to investigate VDR methylation signature at 6 CpGs within a 104-bp region located in a VDR promoter CGI (CGI 1062; Figure 1) by pyrosequencing analysis. Unfortunately, no statistically significant difference was found in the VDR methylation pattern between OPs and CTRs, nor any association between the methylation pattern of VDR and its altered expression levels. These results could be attributable to the presence of additional CGIs/CpG sites not taken into consideration, or to other epigenetic signatures that modulating VDR expression levels in OP disease [37]. The expression of VDR in the male reproductive system and spermatozoa, led researchers to investigate the VDR methylation signatures in association with reproductive disorders, including male infertility [38,71] These conditions are linked to the supposed positive role of vitamin D in sperm maturation, which would result in increased intracellular calcium concentration, sperm motility and induction of the acrosome reaction [71]. In this pathological context, Vladoiu and colleagues reported higher VDR methylation levels in three promoter CGIs (Island 1- 192 bp, Island 2- 187 bp, Island 3- 160 bp; Figure 1) in infertile males with lower vitamin D levels, compared to males with higher vitamin D levels and healthy subjects [38]. Additionally, VDR methylation percentage increases with the severity of the diagnosis, correlating with lower sperm motility, lower sperm concentration, and altered sperm morphology [38]. These promising results were also confirmed in a subsequent study conducted on a cohort of Egyptian men with idiopathic infertility. Published data reported a negative correlation between seminal methylation status of CGI2, located in the VDR promoter region, and both sperm concentration and progressive motility, strengthening the potential role of this epigenetic mechanism in male infertility [39]. Methylation of VDR has also recently been investigated in the pathological state of recurrent kidney stone formation, given the significant association between the genetic variability of VDR and urinary stone formation risk [40,72]. In the study from Khatami et al., VDR methylation analysis was performed in 30 consecutive recurrent kidney stone formers and 30 age and gender-matched CTRs, revealing a hypermethylation pattern of two VDR promoter regions, including 5 and 11 CpG sites, in cases respect to CTRs [40]. However, these preliminary analyses need to be further explored as they represent the only bibliographic data available to date on the role of VDR methylation in recurrent stone formation. In the contest of T2DM, VDR methylation has also been analysed in association with physical activity in a 1:1 matching case-control study [41]. HRM approach was performed to determine the methylation levels of VDR promoter in a 300 bp region comprising 27 CpGs, both in 272 T2DM patients and healthy CTRs. Interestingly, the proactively physical activity seems to reduce the risk of T2DM, especially in people with low VDR methylation levels. These results suggests that VDR methylation could attenuates the association between physical activity and T2DM. Moreover, increased methylation levels of VDR were also associated with decreased levels of serum insulin, confirming its pivotal role in modulating T2DM phenotype [41]. All these preliminary data encourage additional researches on the role of VDR methylation in the above-mentioned diseases, still poorly explored. To date, the major limitation is the small region of the VDR gene analyzed, which comprise a few CGIs/CpG sites located mainly in the promoter, thus neglecting the entire gene that may have other methylation-sensitive sequences. In addition, further studies on larger sample size are needed to confirm the methylation pattern of the VDR, as well as functional studies to confirm the role of DNA methylation in these human disorders.
7. New Advances in Therapeutic Strategies
Methylation signatures are increasingly being studied as part of new pharmacological strategies due to their ability to be potentially reversible. Several demethylating agents capable of removing DNA methylation aberrations have been identified in the pathological context of autoimmune diseases, infectious diseases and cancer [73,74]. It was demonstrated in vitro and in vivo that Methotrexate (MTX), a drug commonly used to treat various diseases, including RA, is able to modulate DNA methylation [75,76]. MTX inhibits methionine S-adenosyltransferase (MAT), the enzyme responsible for the synthesis of S-adenosyl methionine (SAM), which is able to donate its methyl group in numerous biologically important reactions, including DNA methylation [76]. To date, only one study has explored the role of MTX as an agent capable of modulating the methylation pattern of the VDR for the treatment of RA, unfortunately without any positive results [23]. 5-Aza-2'-deoxycytidine (ZdCyd) is a second demethylating agent tested in the field of infectious diseases [31]. An in vitro study performed on TCs previously activated with HIV, showed an upregulation of DNMT3b, an increased methylation of the VDR promoter and a downregulation of VDR levels. Treatment with ZdCyd inhibits the effect of HIV, reverting VDR expression pattern [31]. Another class of demethylating agents are chemotherapeutic compounds, normally used as a therapeutic strategy in the treatment of cancer, which are also capable of establishing covalent complexes with DNMTs, leading to depletion of active enzymes [77]. Among them, 5-azacytidine (ZCyd) was investigated as treatment agent in calcitriol-resistant breast cancer cell lines. Interestingly, ZCyd induces the hypermethylation of the VDR promoter with the consequent decreased VDR expression levels. Therefore, ZCyd was able to inhibit the epigenetic silencing of VDR and restore calcitriol sensitivity of breast cancer cell lines, thus representing a potential drug in anti-cancer therapy [32]. In the field of HCC an interesting potential therapeutic molecule is represented by curcumin as epigenetic agent. Curcumin is able to covalently block the catalytic site of DNMTs, thus generating an inhibitory effect on DNA methylation signature [78]. In curcumin-treated HepG2 cells VDR methylation percentage is decreased and VDR expression pattern is reactivated [34]. All these therapeutic strategies proposed to inhibit aberrant VDR methylation represent interesting interventions, which unfortunately have several disadvantages. These compounds show cytotoxicity at high doses or long-term treatment [77]. Their active form has a short half-life in circulation and are unstable in aqueous solution [79]. Sometimes, demethylating agents lack of specificity for target genes and this may cause undesirable effects. Finally, some of these compounds, such as ZCyd and ZdCyd, require actively proliferating cells to interact with DNA[79]. Further studies on the role of VDR methylation in various human disorders and the mechanism of action of demethylating agents are essential to translate from in vitro and in vivo studies to clinical practice.
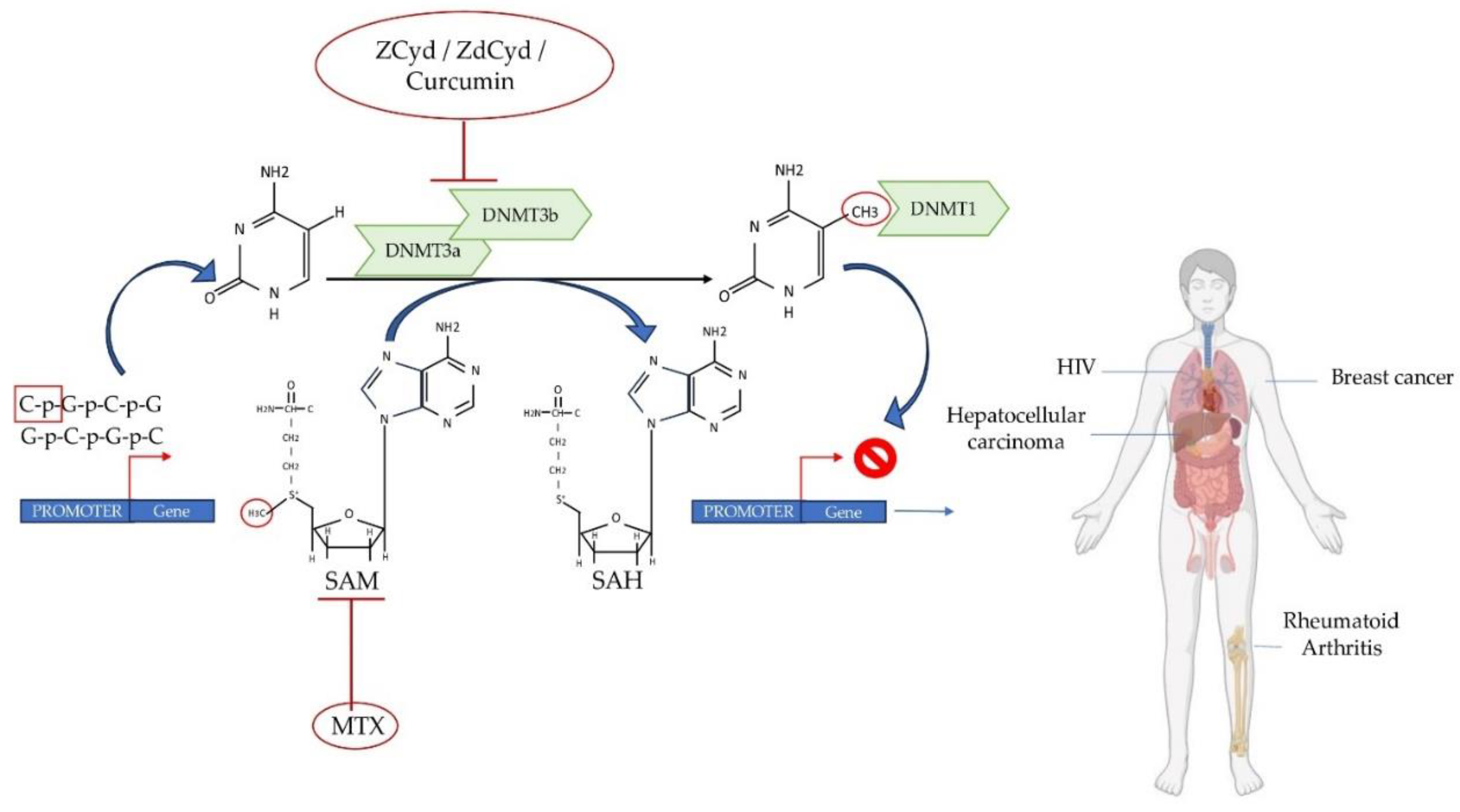
Figure 2.
Summary of the potential mechanisms of action of DNA methylation inhibitors in RA, HIV and cancer. The transfer of a methyl group to the C5 position of cytosine at CpG sites, facilitated by enzymes belonging to DNMTs, results in the formation of 5-methylcytosine. DNMT3a/3b can extract CH3 from the methyl donor SAM to produce SAH. MTX, utilized as an anti-inflammatory drug, has the ability to inhibit SAM. Demethylating agents such as ZCyd, ZdCyd and curcumin can inhibit DNMT3a and DNMT3b activities. DNA methyltransferases, DNMTs; SAM, S-adenosylmethionine; SAH, S-adenosylhomocysteine; ZCyd, 5-Azacytidine; ZdCyd, 5-Aza-2'-deoxycytidine MTX, Created with the support of BioRender.com.
Figure 2.
Summary of the potential mechanisms of action of DNA methylation inhibitors in RA, HIV and cancer. The transfer of a methyl group to the C5 position of cytosine at CpG sites, facilitated by enzymes belonging to DNMTs, results in the formation of 5-methylcytosine. DNMT3a/3b can extract CH3 from the methyl donor SAM to produce SAH. MTX, utilized as an anti-inflammatory drug, has the ability to inhibit SAM. Demethylating agents such as ZCyd, ZdCyd and curcumin can inhibit DNMT3a and DNMT3b activities. DNA methyltransferases, DNMTs; SAM, S-adenosylmethionine; SAH, S-adenosylhomocysteine; ZCyd, 5-Azacytidine; ZdCyd, 5-Aza-2'-deoxycytidine MTX, Created with the support of BioRender.com.
8. Conclusions
The VDR epigenetic signature through DNA methylation mechanism emerges as a critical factor in the susceptibility to several human diseases. To date, there are many gaps and conflicting data on the role of VDR methylation in autoimmune diseases, mainly attributable to the lack of a complete understanding of their pathogenic mechanisms Nevertheless, many studies on infectious diseases and cancer agree that hypermethylation of the VDR promoter is associated with reduced VDR expression levels. This signature appears to increase susceptibility to infectious diseases and inhibit the antitumour effects of vitamin D, thereby increasing cancer risk and progression. However, the use of different detection methods to investigate VDR methylation together with the small sample size and stratification, remains major limitations to be overcome. The key role of VDR in all these pathological conditions underlines the pleiotropy of this factor, prompting researchers to characterize in deep the VDR methylation signature also to develop new therapeutic strategies. So far, therapeutic strategies proposed to inhibit aberrant VDR methylation have primarily been tested in vivo and in vitro models, which need to be strengthened to translate preclinical efficacy into practise improving clinical management and responses to vitamin D in human disorders.
Author Contributions
Conceptualization, A.B., V.V.V. and U.T.; methodology, B.G; A.F., E. P., V.V.V.; resources, B.G. and A.F.; data curation, B.G., A.F., V.V.V.; writing—original draft preparation, B.G. and A.F.; writing—review and editing, A.B. and V.V.V.; supervision, A.B. and V.V.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study were extracted from the articles cited in the text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol Rev 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Portales-Castillo, I.; Simic, P. PTH, FGF-23, Klotho and Vitamin D as Regulators of Calcium and Phosphorus: Genetics, Epigenetics and Beyond. Front Endocrinol (Lausanne) 2022, 13, 992666. [Google Scholar] [CrossRef] [PubMed]
- Saccone, D.; Asani, F.; Bornman, L. Regulation of the Vitamin D Receptor Gene by Environment, Genetics and Epigenetics. Gene 2015, 561, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.-C.; Jurutka, P.W. Molecular Mechanisms of Vitamin D Action. Calcif Tissue Int 2013, 92, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Pike, J.W.; Meyer, M.B. The Vitamin D Receptor: New Paradigms for the Regulation of Gene Expression by 1,25-Dihydroxyvitamin D(3). Endocrinol Metab Clin North Am 2010, 39, 255–269, table of contents. [Google Scholar] [CrossRef]
- Miyamoto, K.; Kesterson, R.A.; Yamamoto, H.; Taketani, Y.; Nishiwaki, E.; Tatsumi, S.; Inoue, Y.; Morita, K.; Takeda, E.; Pike, J.W. Structural Organization of the Human Vitamin D Receptor Chromosomal Gene and Its Promoter. Mol Endocrinol 1997, 11, 1165–1179. [Google Scholar] [CrossRef]
- Jehan, F.; d’Alésio, A.; Garabédian, M. Exons and Functional Regions of the Human Vitamin D Receptor Gene around and within the Main 1a Promoter Are Well Conserved among Mammals. J Steroid Biochem Mol Biol 2007, 103, 361–367. [Google Scholar] [CrossRef]
- White, J.H. Vitamin D Signaling, Infectious Diseases, and Regulation of Innate Immunity. Infect Immun 2008, 76, 3837–3843. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.; Walter, J.; Paulsen, M.; Lengauer, T. CpG Island Mapping by Epigenome Prediction. PLoS Comput Biol 2007, 3, e110. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Saccone, D.S.; Tugizimana, F.; Asani, F.F.; Jeffery, T.J.; Bornman, L. Methylation of the Vitamin D Receptor (VDR) Gene, Together with Genetic Variation, Race, and Environment Influence the Signaling Efficacy of the Toll-Like Receptor 2/1-VDR Pathway. Front Immunol 2017, 8, 1048. [Google Scholar] [CrossRef] [PubMed]
- Maruthai, K.; Sankar, S.; Subramanian, M. Methylation Status of VDR Gene and Its Association with Vitamin D Status and VDR Gene Expression in Pediatric Tuberculosis Disease. Immunol Invest 2022, 51, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Tonacci, A.; Negrini, S.; Greco, M.; Borro, M.; Puppo, F.; Gangemi, S. Emerging Role of Vitamin D in Autoimmune Diseases: An Update on Evidence and Therapeutic Implications. Autoimmun Rev 2019, 18, 102350. [Google Scholar] [CrossRef] [PubMed]
- Dardalhon, V.; Korn, T.; Kuchroo, V.K.; Anderson, A.C. Role of Th1 and Th17 Cells in Organ-Specific Autoimmunity. J Autoimmun 2008, 31, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Szodoray, P.; Nakken, B.; Gaal, J.; Jonsson, R.; Szegedi, A.; Zold, E.; Szegedi, G.; Brun, J.G.; Gesztelyi, R.; Zeher, M.; et al. The Complex Role of Vitamin D in Autoimmune Diseases. Scand J Immunol 2008, 68, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Illescas-Montes, R.; Melguizo-Rodríguez, L.; Ruiz, C.; Costela-Ruiz, V.J. Vitamin D and Autoimmune Diseases. Life Sci 2019, 233, 116744. [Google Scholar] [CrossRef] [PubMed]
- Ao, T.; Kikuta, J.; Ishii, M. The Effects of Vitamin D on Immune System and Inflammatory Diseases. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Fletcher, J.; Bishop, E.L.; Harrison, S.R.; Swift, A.; Cooper, S.C.; Dimeloe, S.K.; Raza, K.; Hewison, M. Autoimmune Disease and Interconnections with Vitamin D. Endocr Connect 2022, 11. [Google Scholar] [CrossRef]
- Puncevičienė, E.; Gaiževska, J.; Sabaliauskaitė, R.; Šnipaitienė, K.; Vencevičienė, L.; Vitkus, D.; Jarmalaitė, S.; Butrimienė, I. Analysis of Epigenetic Changes in Vitamin D Pathway Genes in Rheumatoid Arthritis Patients. Acta Med Litu 2022, 29, 78–90. [Google Scholar] [CrossRef]
- Ayuso, T.; Aznar, P.; Soriano, L.; Olaskoaga, A.; Roldán, M.; Otano, M.; Ajuria, I.; Soriano, G.; Lacruz, F.; Mendioroz, M. Vitamin D Receptor Gene Is Epigenetically Altered and Transcriptionally Up-Regulated in Multiple Sclerosis. PLoS One 2017, 12, e0174726. [Google Scholar] [CrossRef]
- Bulur, I.; Onder, M. Behçet Disease: New Aspects. Clin Dermatol 2017, 35, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. PATHOGENESIS OF RHEUMATOID ARTHRITIS: THE INTERSECTION OF GENETICS AND EPIGENETICS. Trans Am Clin Climatol Assoc 2018, 129, 171–182. [Google Scholar] [PubMed]
- Zhang, T.-P.; Li, H.-M.; Huang, Q.; Wang, L.; Li, X.-M. Vitamin D Metabolic Pathway Genes Polymorphisms and Their Methylation Levels in Association With Rheumatoid Arthritis. Front Immunol 2021, 12, 731565. [Google Scholar] [CrossRef]
- Scazzone, C.; Agnello, L.; Bivona, G.; Lo Sasso, B.; Ciaccio, M. Vitamin D and Genetic Susceptibility to Multiple Sclerosis. Biochem Genet 2021, 59, 1–30. [Google Scholar] [CrossRef]
- Yamout, B.I.; Alroughani, R. Multiple Sclerosis. Semin Neurol 2018, 38, 212–225. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple Sclerosis - a Review. Eur J Neurol 2019, 26, 27–40. [Google Scholar] [CrossRef]
- Shirvani, S.S.; Nouri, M.; Sakhinia, E.; Babaloo, Z.; Jadideslam, G.; Shahriar, A.; Farhadi, J.; Khabbazi, A. The Expression and Methylation Status of Vitamin D Receptor Gene in Behcet’s Disease. Immun Inflamm Dis 2019, 7, 308–317. [Google Scholar] [CrossRef]
- Jiang, C.; Zhu, J.; Liu, Y.; Luan, X.; Jiang, Y.; Jiang, G.; Fan, J. The Methylation State of VDR Gene in Pulmonary Tuberculosis Patients. J Thorac Dis 2017, 9, 4353–4357. [Google Scholar] [CrossRef]
- Wang, M.; Kong, W.; He, B.; Li, Z.; Song, H.; Shi, P.; Wang, J. Vitamin D and the Promoter Methylation of Its Metabolic Pathway Genes in Association with the Risk and Prognosis of Tuberculosis. Clin Epigenetics 2018, 10, 118. [Google Scholar] [CrossRef]
- Li, Y.-P.; Deng, H.-L.; Wang, W.-J.; Wang, M.-Q.; Li, M.; Zhang, Y.-F.; Wang, J.; Dang, S.-S. Vitamin D Receptor Gene Methylation in Patients with Hand, Foot, and Mouth Disease Caused by Enterovirus 71. Arch Virol 2020, 165, 1979–1985. [Google Scholar] [CrossRef]
- Chandel, N.; Husain, M.; Goel, H.; Salhan, D.; Lan, X.; Malhotra, A.; McGowan, J.; Singhal, P.C. VDR Hypermethylation and HIV-Induced T Cell Loss. J Leukoc Biol 2013, 93, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Marik, R.; Fackler, M.; Gabrielson, E.; Zeiger, M.A.; Sukumar, S.; Stearns, V.; Umbricht, C.B. DNA Methylation-Related Vitamin D Receptor Insensitivity in Breast Cancer. Cancer Biol Ther 2010, 10, 44–53. [Google Scholar] [CrossRef]
- Pilon, C.; Rebellato, A.; Urbanet, R.; Guzzardo, V.; Cappellesso, R.; Sasano, H.; Fassina, A.; Fallo, F. Methylation Status of Vitamin D Receptor Gene Promoter in Benign and Malignant Adrenal Tumors. Int J Endocrinol 2015, 2015, 375349. [Google Scholar] [CrossRef]
- Abdalla, M.; Khairy, E.; Louka, M.L.; Ali-Labib, R.; Ibrahim, E.A.-S. Vitamin D Receptor Gene Methylation in Hepatocellular Carcinoma. Gene 2018, 653, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Varshney, S.; Bhadada, S.K.; Sachdeva, N.; Arya, A.K.; Saikia, U.N.; Behera, A.; Rao, S.D. Methylation Status of the CpG Islands in Vitamin D and Calcium-Sensing Receptor Gene Promoters Does Not Explain the Reduced Gene Expressions in Parathyroid Adenomas. J Clin Endocrinol Metab 2013, 98, E1631–5. [Google Scholar] [CrossRef] [PubMed]
- Afshan, F.U.; Masood, A.; Nissar, B.; Chowdri, N.A.; Naykoo, N.A.; Majid, M.; Ganai, B.A. Promoter Hypermethylation Regulates Vitamin D Receptor (VDR) Expression in Colorectal Cancer-A Study from Kashmir Valley. Cancer Genet 2021, 252–253, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Gasperini, B.; Visconti, V.V.; Ciccacci, C.; Falvino, A.; Gasbarra, E.; Iundusi, R.; Brandi, M.L.; Botta, A.; Tarantino, U. Role of the Vitamin D Receptor (VDR) in the Pathogenesis of Osteoporosis: A Genetic, Epigenetic and Molecular Pilot Study. Genes (Basel) 2023, 14. [Google Scholar] [CrossRef]
- Vladoiu, S.; Botezatu, A.; Anton, G.; Manda, D.; Paun, D.L.; Oros, S.; Rosca, R.; Dinu Draganescu, D. THE INVOLVEMENT OF VDR PROMOTER METHYLATION, CDX-2 VDR POLYMORPHISM AND VITAMIN D LEVELS IN MALE INFERTILITY. Acta Endocrinol (Buchar) 2017, 13, 294–301. [Google Scholar] [CrossRef]
- Hussein, T.M.; Eldabah, N.; Zayed, H.A.; Genedy, R.M. Assessment of Serum Vitamin D Level and Seminal Vitamin D Receptor Gene Methylation in a Sample of Egyptian Men with Idiopathic Infertility. Andrologia 2021, 53, e14172. [Google Scholar] [CrossRef]
- Khatami, F.; Gorji, A.; Khoshchehreh, M.; Mashhadi, R.; Pishkuhi, M.A.; Khajavi, A.; Shabestari, A.N.; Aghamir, S.M.K. The Correlation between Promoter Hypermethylation of VDR, CLDN, and CasR Genes and Recurrent Stone Formation. BMC Med Genomics 2022, 15, 109. [Google Scholar] [CrossRef]
- Yu, S.; Feng, Y.; Qu, C.; Yu, F.; Mao, Z.; Wang, C.; Li, W.; Li, X. Vitamin D Receptor Methylation Attenuates the Association between Physical Activity and Type 2 Diabetes Mellitus: A Case-Control Study. J Diabetes 2022, 14, 97–103. [Google Scholar] [CrossRef]
- Tyagi, G.; Singh, P.; Varma-Basil, M.; Bose, M. Role of Vitamins B, C, and D in the Fight against Tuberculosis. Int J Mycobacteriol 2017, 6, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-P.; Deng, H.-L.; Xu, L.-H.; Wang, M.-Q.; Li, M.; Zhang, X.; Dang, S.-S. Association of Polymorphisms in the Vitamin D Receptor Gene with Severity of Hand, Foot, and Mouth Disease Caused by Enterovirus 71. J Med Virol 2019, 91, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.A.; Miller, C.W.; El-Abbassi, A.M.; Cutchins, D.C.; Cutchins, C.; Grant, W.B.; Peiris, A.N. Antimicrobial Implications of Vitamin D. Dermatoendocrinol 2011, 3, 220–229. [Google Scholar] [CrossRef] [PubMed]
- White, J.H. Emerging Roles of Vitamin D-Induced Antimicrobial Peptides in Antiviral Innate Immunity. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Peric, M.; Koglin, S.; Kim, S.-M.; Morizane, S.; Besch, R.; Prinz, J.C.; Ruzicka, T.; Gallo, R.L.; Schauber, J. IL-17A Enhances Vitamin D3-Induced Expression of Cathelicidin Antimicrobial Peptide in Human Keratinocytes. J Immunol 2008, 181, 8504–8512. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.A.; Fernandes, D.C.R.O.; Braga, A.C.O.; Cavalcante, G.C.; Sortica, V.A.; Hutz, M.H.; Leal, D.F.V.B.; Fernades, M.R.; Santana-da-Silva, M.N.; Lopes Valente, S.E.; et al. Investigation of Genetic Susceptibility to Mycobacterium Tuberculosis (VDR and IL10 Genes) in a Population with a High Level of Substructure in the Brazilian Amazon Region. Int J Infect Dis 2020, 98, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Möller, M.; de Wit, E.; Hoal, E.G. Past, Present and Future Directions in Human Genetic Susceptibility to Tuberculosis. FEMS Immunol Med Microbiol 2010, 58, 3–26. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X.; Yin, W.; Xie, D.; He, W.; Jiang, G.; Fan, J. 5-Aza-2′-Deoxycytidine Enhances the Antimicrobial Response of Vitamin D Receptor against Mycobacterium Tuberculosis. RSC Adv 2016, 6, 61740–61746. [Google Scholar] [CrossRef]
- Patton, L.L. HIV Disease. Dent Clin North Am 2003, 47, 467–492. [Google Scholar] [CrossRef]
- Young, M.R.I.; Xiong, Y. Influence of Vitamin D on Cancer Risk and Treatment: Why the Variability? Trends Cancer Res 2018, 13, 43–53. [Google Scholar]
- Chiang, K.-C.; Yeh, C.-N.; Chen, M.-F.; Chen, T.C. Hepatocellular Carcinoma and Vitamin D: A Review. J Gastroenterol Hepatol 2011, 26, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.un N.; Khan, T.A.; Maqbool, S.A. Vitamin D Receptor Cdx-2 Polymorphism and Premenopausal Breast Cancer Risk in Southern Pakistani Patients. PLoS One 2015, 10, e0122657. [Google Scholar] [CrossRef] [PubMed]
- Larriba, M.J.; González-Sancho, J.M.; Barbáchano, A.; Niell, N.; Ferrer-Mayorga, G.; Muñoz, A. Vitamin D Is a Multilevel Repressor of Wnt/b-Catenin Signaling in Cancer Cells. Cancers (Basel) 2013, 5, 1242–1260. [Google Scholar] [CrossRef] [PubMed]
- Toropainen, S.; Väisänen, S.; Heikkinen, S.; Carlberg, C. The Down-Regulation of the Human MYC Gene by the Nuclear Hormone 1alpha,25-Dihydroxyvitamin D3 Is Associated with Cycling of Corepressors and Histone Deacetylases. J Mol Biol 2010, 400, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Larriba, M.J.; Valle, N.; Pálmer, H.G.; Ordóñez-Morán, P.; Alvarez-Díaz, S.; Becker, K.-F.; Gamallo, C.; de Herreros, A.G.; González-Sancho, J.M.; Muñoz, A. The Inhibition of Wnt/Beta-Catenin Signalling by 1alpha,25-Dihydroxyvitamin D3 Is Abrogated by Snail1 in Human Colon Cancer Cells. Endocr Relat Cancer 2007, 14, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Fetahu, I.S.; Höbaus, J.; Kállay, E. Vitamin D and the Epigenome. Front Physiol 2014, 5, 164. [Google Scholar] [CrossRef] [PubMed]
- Pilon, C.; Urbanet, R.; Williams, T.A.; Maekawa, T.; Vettore, S.; Sirianni, R.; Pezzi, V.; Mulatero, P.; Fassina, A.; Sasano, H.; et al. 1α,25-Dihydroxyvitamin D₃ Inhibits the Human H295R Cell Proliferation by Cell Cycle Arrest: A Model for a Protective Role of Vitamin D Receptor against Adrenocortical Cancer. J Steroid Biochem Mol Biol 2014, 140, 26–33. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast Cancer: Biology, Biomarkers, and Treatments. Int Immunopharmacol 2020, 84, 106535. [Google Scholar] [CrossRef]
- Segovia-Mendoza, M.; García-Quiroz, J.; Díaz, L.; García-Becerra, R. Combinations of Calcitriol with Anticancer Treatments for Breast Cancer: An Update. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Lerario, A.M.; Moraitis, A.; Hammer, G.D. Genetics and Epigenetics of Adrenocortical Tumors. Mol Cell Endocrinol 2014, 386, 67–84. [Google Scholar] [CrossRef]
- Howard, B.; Wang, Y.; Xekouki, P.; Faucz, F.R.; Jain, M.; Zhang, L.; Meltzer, P.G.; Stratakis, C.A.; Kebebew, E. Integrated Analysis of Genome-Wide Methylation and Gene Expression Shows Epigenetic Regulation of CYP11B2 in Aldosteronomas. J Clin Endocrinol Metab 2014, 99, E536–43. [Google Scholar] [CrossRef] [PubMed]
- Ayers, D.; Boughanem, H.; Macías-González, M. Epigenetic Influences in the Obesity/Colorectal Cancer Axis: A Novel Theragnostic Avenue. J Oncol 2019, 2019, 7406078. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, L.; Carmeliet, G. Integrated View on the Role of Vitamin D Actions on Bone and Growth Plate Homeostasis. JBMR Plus 2021, 5, e10577. [Google Scholar] [CrossRef]
- Adamczewska, D.; Słowikowska-Hilczer, J.; Walczak-Jędrzejowska, R. The Association between Vitamin D and the Components of Male Fertility: A Systematic Review. Biomedicines 2022, 11. [Google Scholar] [CrossRef]
- Szymczak-Pajor, I.; Śliwińska, A. Analysis of Association between Vitamin D Deficiency and Insulin Resistance. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Letavernier, E.; Daudon, M. Vitamin D, Hypercalciuria and Kidney Stones. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Xu, F.; Li, W.; Yang, X.; Na, L.; Chen, L.; Liu, G. The Roles of Epigenetics Regulation in Bone Metabolism and Osteoporosis. Front Cell Dev Biol 2020, 8, 619301. [Google Scholar] [CrossRef]
- Visconti, V.V.; Cariati, I.; Fittipaldi, S.; Iundusi, R.; Gasbarra, E.; Tarantino, U.; Botta, A. DNA Methylation Signatures of Bone Metabolism in Osteoporosis and Osteoarthritis Aging-Related Diseases: An Updated Review. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Blomberg Jensen, M.; Bjerrum, P.J.; Jessen, T.E.; Nielsen, J.E.; Joensen, U.N.; Olesen, I.A.; Petersen, J.H.; Juul, A.; Dissing, S.; Jørgensen, N. Vitamin D Is Positively Associated with Sperm Motility and Increases Intracellular Calcium in Human Spermatozoa. Hum Reprod 2011, 26, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Shabestari, A.N.; Baghdadabad, L.Z.; Khatami, F.; Reis, L.O.; Pishkuhi, M.A.; Kazem Aghamir, S.M. Genetic Polymorphisms and Kidney Stones Around the Globe: A Systematic Review and Meta-Analysis. Front Genet 2022, 13, 913908. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Nielsen, H.M.; Hansen, L.L. Epigenetics and Cancer Treatment. Eur J Pharmacol 2009, 625, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Yoo, C.B.; Weisenberger, D.J.; Chuang, J.; Wozniak, C.; Liang, G.; Marquez, V.E.; Greer, S.; Orntoft, T.F.; Thykjaer, T.; et al. Preferential Response of Cancer Cells to Zebularine. Cancer Cell 2004, 6, 151–158. [Google Scholar] [CrossRef]
- Hanoodi, M.; Mittal, M. Methotrexate; 2023.
- Wang, Y.-C.; Chiang, E.-P.I. Low-Dose Methotrexate Inhibits Methionine S-Adenosyltransferase in Vitro and in Vivo. Mol Med 2012, 18, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Christman, J.K. 5-Azacytidine and 5-Aza-2’-Deoxycytidine as Inhibitors of DNA Methylation: Mechanistic Studies and Their Implications for Cancer Therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef]
- Fu, S.; Kurzrock, R. Development of Curcumin as an Epigenetic Agent. Cancer 2010, 116, 4670–4676. [Google Scholar] [CrossRef]
- Howell, P.M.; Liu, Z.; Khong, H.T. Demethylating Agents in the Treatment of Cancer. Pharmaceuticals (Basel) 2010, 3, 2022–2044. [Google Scholar] [CrossRef]
Figure 1.
Structure of the human VDR gene. The human VDR is located on the long arm of chromosome 12 (12q12-q14) and comprises eight exons (2-9) along with six non-coding exons (1f-1c). The green box represents the primary promoter and the orange boxes indicate the relative positions of VDR alternative promoters. The black boxes represent the bona fide CGIs mapped according to the CGIs predictive model by Bock and colleagues. The red boxes symbolize CGIs identified by bioinformatic tools.
Figure 1.
Structure of the human VDR gene. The human VDR is located on the long arm of chromosome 12 (12q12-q14) and comprises eight exons (2-9) along with six non-coding exons (1f-1c). The green box represents the primary promoter and the orange boxes indicate the relative positions of VDR alternative promoters. The black boxes represent the bona fide CGIs mapped according to the CGIs predictive model by Bock and colleagues. The red boxes symbolize CGIs identified by bioinformatic tools.
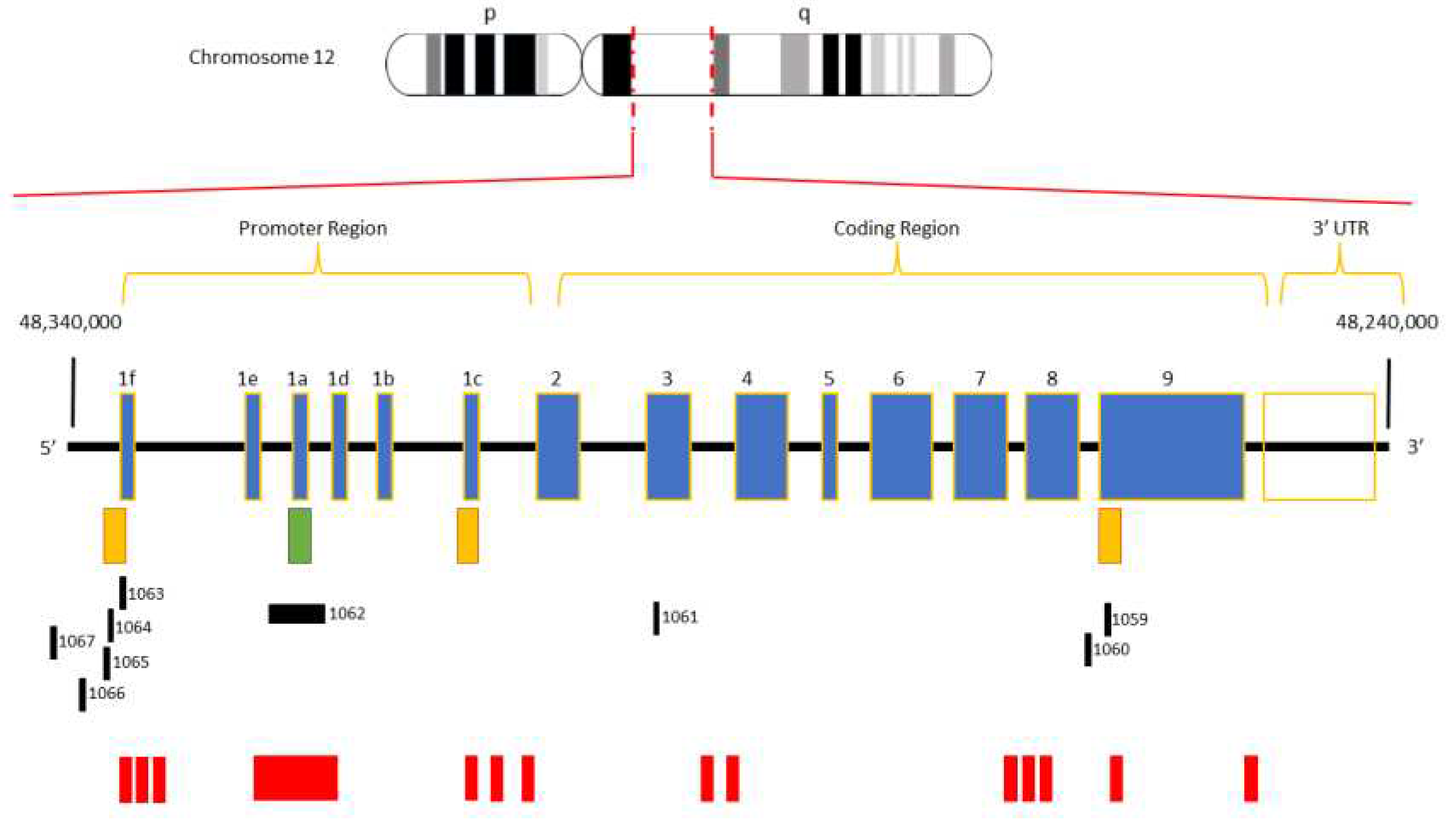

Table 1.
Summary of the main studies on VDR gene methylation in autoimmune and infectious diseases, cancer and others pathological conditions discussed in the text.
Table 1.
Summary of the main studies on VDR gene methylation in autoimmune and infectious diseases, cancer and others pathological conditions discussed in the text.
| Disease | Sample Size | Method of Analysis | Genomic Context (VDR gene) |
Analysis Outcome | Reference | |
|---|---|---|---|---|---|---|
| Autoimmune diseases | Rheumatoid Arthritis | 122 RA vs 123 CTRs |
MethylTarget | CpG sites in the promoter region |
VDR methylation levels in RA patients were significantly reduced compared to CTRs |
[23] |
| 35 RA vs 41 CTRs |
Pyrosequencing | 10 CpG sites in the promoter region | Methylation analysis revealed no significant differences between RA patients compared to CTRs | [19] | ||
| Multiple Sclerosis |
23 RRMS
vs 12 CTRs |
Bisulfite cloning
sequencing |
23 CpG sites in main promoter 10 CpG sites in alternative promoter located at non-coding exon 1c | Methylation levels in VDR alternative promoter were significantly higher in RRMS patients compared to CTRs | [20] | |
| Behcet's disease | 48 BD vs 60 CTRs |
MeDIP-qPCR | All CpG sites in the promoter region from −800 bp to +200 bp relative to the TSS | Methylation analysis revealed no significant differences between BD patients compared to CTRs | [27] | |
| Infectious diseases | Tuberculosis disease | 27 TB vs 30 CTRs |
Bisulfite cloning
sequencing |
16 CpG sites in VDR | TB patients were in the hypermethylation state compared to CTRs | [28] |
| 43 TB vs 33 CTRs |
MS-PCR | The location of CpG sites and CGIs present in the VDR sequence were identified by DBCAT | Methylation analysis revealed an hypermethylation in TB patients and hypomethylation in CTRs | [12] | ||
| 122 TB vs 118 CTRs |
Illumina MiSeq | 60 CpG sites in the promoter region (48299590 - 48298885) |
Methylation levels were significantly lower in TB patients compared to CTRs | [29] | ||
| Hand, foot, and mouth disease | 116 HFMD vs 60 CTRs | MethylTarget | 12 CpG in promoter region from -638 bp to -545 bp relative to the TSS | Methylation levels of VDR promoter in HFMD were lower compared to CTRs | [30] | |
| HIV | TCs obtained from healthy volunteers | Pyrosequencing | CpG in VDR promoter region from -512 bp to -28 bp relative to the TSS | HIV-infected TCs showed increased methylation in CpG sites | [31] | |
|
Cancer |
Breast cancer | 15 BCT vs 7 NBT |
MS-PCR | 3 CGIs in VDR promoter region from -789 bp to +380 bp relative to the TSS | Methylation levels of VDR promoter in BCT were significantly higher compared to NBT | [32] |
| Adrenocortical carcinoma |
23 AT vs 3 NAT |
BSP | 42 CpG sites in VDR promoter region from -693 bp to -65 bp relative to the TSS | 27/42 CpG sites were methylated in 3 ACCs | [33] | |
| Hepatocellular carcinoma |
15 HC vs 15 CLD vs 15 NT | MS-PCR | VDR promoter | Methylation levels of VDR promoter in HCC were significantly higher compared to other studies groups | [34] | |
| Parathyroid adenomas |
15 PAT vs 4 NPT |
BSP | 31 CpG sites in VDR promoter region from -538 bp to -79 bp relative to the TSS | There was no significant methylation in the promoter region of VDR in parathyroid adenomatous tissues. | [35] | |
| Colorectal cancer |
75 CCT
vs 75 NE |
MS-PCR | VDR promoter | Hypermethylation of VDR was detected in 28 (37,33%) of 75 cases | [36] | |
| Others | Osteoporosis | 25 OP vs 25 CTRs |
Pyrosequencing | 6 CpG sites in VDR promoter | No statistically significant difference was found in the methylation pattern between OP and CTRs | [37] |
| Male infertility | 69 ID vs 37 CTRs |
MS-PCR | 3 CGIs in VDR promoter | VDR methylation percentage was increased with the severity of the diagnosis, correlating with lower sperm motility and concentration, and altered sperm morphology | [38] | |
| 60 IID vs 60 CTRs |
MS-PCR | 1 CGI in VDR promoter | Methylation levels of VDR promoter in IID were significantly higher compared to CTRs | [39] | ||
| Recurrent kidney stone formation |
30 consecutive recurrent kidney stone formers vs 30 CTRs | MS-HRM | 16 CpG sites in VDR promoter | Two VDR promoter regions was hypermethylated in patients with consecutive recurrent kidney stone formers compared to CTRs | [40] | |
|
Type 2 diabetes
mellitus |
272 T2DM vs 272 CTRs | MS-HRM | 27 CpG sites in VDR promoter | Increased methylation levels of VDR were associated with decreased levels of serum insulin |
[41] |
Abbreviations: RA, Rheumatoid Arthritis; CTRs, control groups; VDR, Vitamin D Receptor; RRMS, Relapsing-Remitting Multiple Sclerosis; BD, Behcet's Disease; MeDIP-qPCR, Methylated DNA Immunoprecipitation-Real Time PCR; TSS, transcription start site; TB, Tuberculosis; MS-PCR, Methylation Specific- Polymerase Chain Reaction; CGIs, CpG islands; DBCAT, DataBase of CGIs and Analytical Tool; HFMD, Hand, Foot, and Mouth Disease; HIV, Human Immunodeficiency Virus; TCs, Tcell; BCT, breast cancer tissue; NBT, normal breast tissue; AT, adrenal tumors; NAT, normal adrenal tissue; BSP, Bisulfite Sequencing PCR; ACC, adrenocortical carcinoma; HC, hepatocellular carcinoma; CLD, Chronic Liver Disease; NT, normal tissue; PAT, parathyroid adenomas tissue; NPT, normal parathyroid tissue; CCT, colorectal cancer tissue; NE, normal epithelium; OP, Osteoporosis; ID, Infertility Disease; IID, Idiopathic Infertility Disease; MS-HRM, Methylation-Sensitive High Resolution Melting; T2DM, Type 2 Diabetes Mellitus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



