
Submitted:
20 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
Legume plants enter symbiosis with soil nitrogen-fixing bacteria (rhizobia), thereby gaining access to the assimilable atmospheric nitrogen. This symbiosis is important for agriculture, thus, biofertilizers with effective strains of rhizobia are created for crop legumes to increase their yield and minimize the amounts of mineral fertilizers used. In this work, we sequenced and characterized the genome of Rhizobium leguminosarum bv. viciae strain RCAM1022, a component of the ‘Rhizotorfin’ biofertilizer produced in Russia and used for pea (Pisum sativum L.).
Keywords:
Rhizobium leguminosarum
; genome sequencing
; nitrogen fixation
1. Summary
Soil alphaproteobacteria united in the group of rhizobia, or nodule bacteria, are capable of fixing atmospheric nitrogen, i.e., converting it into an assimilable form. This process is carried out in symbiosis with legumes (family Fabaceae) within the specialized symbiotic organs called root nodules [1]. Owing to this symbiosis, the plant satisfies its need in nitrogen, while the bacteria gain access to the products of photosynthesis.
Legume-rhizobial symbiosis is quite specific [2] and involves a complex molecular dialogue between partners at all stages of its development; thus, certain plant species often interact only with a limited range of rhizobia species and even strains. Despite this, even bacteria that have successfully entered symbiosis differ in the positive effect they have on the plant, i.e., their symbiotic effectiveness [3]. Since there are many crops among legumes, and, moreover, they themselves are often used in crop rotations to enrich soils with assimilable nitrogen, studying the mechanisms underlying symbiotic effectiveness may be of great practical importance.
Host specificity and symbiotic effectiveness are multicomponent and complex traits [4]. Some bacteria may poorly fix nitrogen, but successfully colonize roots of the host plant, and vice versa. To increase fitness and yields of crops via symbiosis, it is necessary to select or design such ‘plant-bacterium’ pairs that exhibit both specificity and effectiveness.
For all major legume crops, biofertilizers containing specific (i.e., compatible with a particular plant species) rhizobial strains are developed. For pea (Pisum sativum L.), the biofertilizer named ‘Rhizotorfin’ is produced in Russia, and several field experiments have shown the positive effect of this preparation on pea yield. The strain used in ‘Rhizotorfin’, Rhizobium leguminosarum bv. viciae RCAM1022, has proved its effectiveness on various cultivars of pea [5,6,7]. However, the molecular basis of such effectiveness remains poorly studied. Here we sequenced, assembled and performed the basic annotation of this strain’s genome. Through comparative analysis with a database of known Rh. leguminosarum genomes, the closest strain was identified, being Rh. leguminosarum bv. viciae RCAM1026 [8]. Our study may be of use for future research into the mechanisms of effective interaction between rhizobia and legumes.
2. Data Description
2.1. Strain description
According to the catalog of the Russian Collection of Agricultural Microorganisms (RCAM) at the All-Russia Research Institute for Agricultural Microbiology (ARRIAM), Saint Petersburg, Russia, Rh. leguminosarum strain RCAM1022 was isolated from the roots of grain pea (cultivar ‘Moskovsky 572’) originating from the Moscow region in the Scientific Research Institute of Agriculture of the Non-Chernozem Zone in 1954 (the original name of the strain was 245a). The strain was deposited in the RCAM at the ARRIAM in 1968. Currently the strain (accession number RCAM1022) is stored at –80°C in the automated Tube Store (Liconic Instruments, Lichtenstein) in the RCAM at the ARRIAM (Saint-Petersburg, Russia).
Long-term field experiments have shown that this strain is characterized by effective nitrogen fixation [9]. RCAM1022 serves as a basis for the biofertilizer ‘Rhizotorfin’ (ULTRASTIM®) for pea (Pisum sativum L.). Several types of ‘Rhizotorfin’ with rhizobia specific to particular plant species increase the yield of pea, lentil, lathyrus, vetch and broad beans due to the formation of nodules with effective nitrogen fixation [5,6,7,9].
For present genome sequencing and analysis, the strain RCAM1022 was retrieved from the Russian Collection of Agricultural Microorganisms (RCAM) (All-Russia Research Institute for Agricultural Microbiology).
2.2. Genome Assembly and Annotation
To sequence the genome of RCAM1022 strain, a short-read sequencing library was prepared using the Illumina DNA Prep kit (Illumina, Inc., San Diego, CA) and sequenced using an Illumina NovaSeq 6000 (Illumina, Inc., San Diego, CA). In total, 18 Gbp (or 60,035,897) 2 × 150-bp reads were obtained. The raw reads were quality-trimmed (Phred33 score => 20), and adapter sequences were removed using the BBDuk tool from the BBMap suite ver. 39.03 (https://sourceforge.net/projects/bbmap/) [10]. After filtering, 112,324,092 high-quality paired reads were obtained. The genome was assembled using the Unicycler ver. 0.4.8 program [11] with default options. The assembly consists of 124 contigs, with a total length of 7,245,707 bp, N50 of 242,536 bp, longest contig is 600,404 bp and GC content of 60.85%. Of 124 contigs, 97 are longer than 200 bp.

Table 1.
The main characteristics of the draft genome assembly of the Rh. leguminosarum strain RCAM1022.
Table 1.
The main characteristics of the draft genome assembly of the Rh. leguminosarum strain RCAM1022.
| Total amount of contigs | 124 |
| Total length of contigs | 7,245,707 |
| N50 | 242,536 |
| N90 | 70,305 |
| Longest segment | 600,404 |
| Contigs longer than 200 bp | 97 |
| Coverage | 16x |
| GC% | 60.85% |
| Reads mapped onto assembly | 100% |
Prokka pipeline ver. 1.14.5 [12] was used to annotate the assembly. Final assembly contains 6,954 protein-coding genes, 2 rRNA operons, and 51 tRNA genes (Table 2).
MicrobeAnnotator ver. 2.0.5 [13] was used to describe the metabolic processes in which annotated genes are involved (Figure 1). Lysine metabolism, cofactor and vitamin metabolism, branched-chain amino acid metabolism, carbohydrate metabolism, serine and threonine metabolism are the most complete categories among annotated genes. Also, six nod, three fix and five nif genes were found in the annotated genome (Supplementary File S1).
The obtained assembly was compared with all Rh. leguminosarum strains available in NCBI databases using the fastANI ver.1.33 algorithm [14] incorporated in pyani python3.8 module [15]. According to the analysis, the closest related strain to the studied one is Rh. leguminosarum bv. viciae strain RCAM1026 [8] with average nucleotide identity of 99,993%. In total, 29 genes with variations in the nucleotide sequence between Rh. leguminosarum bv. viciae strain RCAM1022 and Rh. leguminosarum bv. viciae strain RCAM1026 were found. Among them, substitutions lead to a change in the amino acid sequence of 15 genes. These genes, in particular, encode the proteins such as Nodulation protein O, Glutathione amide reductase, Protein-export protein SecB, Periplasmic oligopeptide-binding protein, etc. (Supplementary File S2).
3. Methods
3.1. DNA Extraction
DNA was extracted from bacterial liquid culture, grown for 2 days in TY (Beringer, 1974) medium at 28°C. Five milliliters of bacterial culture was concentrated in 1.5 ml tube by successive quick (1-2 minutes) centrifugations at 2700 RCF (Eppendorf centrifuge 5430R, Eppendorf, Germany) and supernatant removing. DNA preparation was based on a method described in Wilson, 2001 [16] with essential additions necessary to reduce the concentration of polysaccharides. 500 μl of 1M NaCl was added to the sample and vortexed until full dilution of the precipitate. The tube was centrifuged for 10 minutes at 2700 RCF at room temperature (RT). Supernatant was carefully removed and the procedure was repeated two more times. Then the precipitate was diluted in 500 μl of Milli-Q Water (MQ water), and the sample was centrifuged for 10 min at 3000 RCF at RT. The supernatant was removed and the precipitate was diluted in 567 ml of TE buffer. Then further DNA extraction procedure was carried out in accordance with Wilson, 2001. At the final stage of isolation, the DNA precipitate was dried for 5 minutes at RT and then diluted in 50 μl of TE buffer by vortexing and overnight incubation at +4°C. Then DNA was stored at -20°C until the library preparation and sequencing.
3.2. DNA quality and quantity controls
DNA concentration was measured by dsDNA Quantitation Broad Range Kit (Invitrogen, USA) and NanoDrop OneC (Thermo Fisher Scientific, USA). DNA quality control (the presence of contaminating components) was performed by analyzing the sample at different absorbing ratios by NanoDrop OneC (Thermo Fisher Scientific, USA). Additionally, DNA was visualized on 1% agarose gel in TAE buffer using electrophoresis.
3.3. Whole Genome Sequencing, De Novo Genome Assembly, and Annotation
A whole-genome sequencing library was prepared using the Illumina DNA Prep kit (Illumina, Inc., San Diego, CA) following the manufacturer’s instructions.
The libraries were sequenced using the Illumina NovaSeq 6000 (Illumina, Inc., San Diego, CA) in Evrogen JSC (Moscow, Russia), and 60,035,897 2 × 150 paired end reads were generated. The quality of raw reads was accessed in the FASTQC program (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Trimming of adapter sequences from short read data was conducted by BBDuk tool from BBTools (https://sourceforge.net/projects/bbmap/) [10] package for Linux. Low-quality sequences (with Phred 33 score < Q20) were also removed with BBDuk. After processing, reads were contained from 50 to 150 bp. In total, 56,162,046 x 2 paired reads were obtained.
Genome assembly was performed using Unicycler – hybrid assembly pipeline, but we used Illumina-only read sets which were assembled using the SPAdes ver. 3.12.0 assembler [17] built into this pipeline. After assembling, the quality of the genome was checked by the QUAST ver. 5.2.0 [18] with mapping reads to the de novo assembled genome. The structural and functional annotation genome was determined by the Prokka [14] excluding the contigs shorter than 200 bp. Metabolics processes were obtained by MicrobeAnnotator ver. 2.0.5 [13] in ‘light’ mode.
To compare our assembly to all Rh. leguminosarum strains, assemblies were obtained from NCBI by taxid (Rh. leguminosarum taxid=384) using script genbank_get_genomes_by_taxon.py from pyani ver. 2.0. python ver. 3.8 module. Average nucleotide identity (ANI) was calculated by alignment-free comparison of the genomes with our assembly using FastANI tool ver.1.33 [14].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The list of Nod and Fix genes of Rh. leguminosarum RCAM1022; Table S2: The list of genes with variations between Rh. leguminosarum RCAM1022 and Rh. leguminosarum RCAM1026.
Author Contributions
Conceptualization, V.A.Z.; methodology, O.A.K; G.A.A., A.S.S.; validation, V.A.Z., A.S.S.; formal analysis, E.A.Z., O.A.K., V.A.Z., A.S.S.; investigation, E.A.Z., O.A.K., V.A.Z., A.S.S.; resources, V.A.Z.; data curation, E.A.Z.; writing—original draft preparation, O.A.K., E.A.Z.; writing—review and editing, V.A.Z., E.A.Z., A.S.S., O.A.K.; visualization, E.A.Z.; supervision, V.A.Z.; project administration, V.A.Z.; funding acquisition, V.A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The work of E.A.Z., A.S.S., O.A.K., G.A.A. and V.A.Z. was supported by the RSF grant # 22-16-00109.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
The assembly and sequence data have been uploaded to the NCBI database. The assembly is deposited under the BioProject accession number PRJNA1038702. The raw Illumina data are deposited under the accession number PRJNA1038712.
Acknowledgments
The authors thank Vera Safronova, the head of the Russian Collection of Agricultural Microorganisms (RCAM) (ARRIAM, Saint Petersburg, Russia), for providing information about the strain, Ekaterina Vasileva, Alla Saksaganskaia, Anastasiia Kimeklis (ARRIAM, Saint Petersburg, Russia) for discussion of methodological aspects, and Lyudmila Dvoryaninova (ARRIAM, Saint Petersburg, Russia) for her excellent technical assistance. The DNA isolation and quantity control were performed using equipment of the Core Center ‘Genomic Technologies, Proteomics and Cell Biology’ in ARRIAM (Saint Petersburg, Russia).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Oldroyd, G.E.D.; Downie, J.A. Coordinating Nodule Morphogenesis with Rhizobial Infection in Legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.; Lagunas, B.; Gifford, M.L. Determinants of Host Range Specificity in Legume-Rhizobia Symbiosis. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of Nitrogen Fixation in Rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [PubMed]
- Thrall, P.H.; Laine, A.L.; Broadhurst, L.M.; Bagnall, D.J.; Brockwell, J. Symbiotic Effectiveness of Rhizobial Mutualists Varies in Interactions with Native Australian Legume Genera. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Aleshin, M.A.; Subbotina, M.G. An Effect Of Doses Of Nitrogen And Of Rhizotorphin On Pea ( Pisum Sativum L.) Productivity In The Conditions Of Podzol Heavy-Loam Soil. Agrar. Sci. J. 2019, 8, 4–11. (In Russian) [Google Scholar] [CrossRef]
- Yurkov, A.P.; Lactionov, Y. V; Kojemyakov, A.P.; Stepanova, G. V Symbiotic Efficiency of Bacterial and Fungal Preparations for Forage Crops According to Seed Harvest. Kormoproizvodstvo 2017, 16–21. (In Russian).
- Kiselev, A.S.; Popolzukhina, N.A.; Popolzukhin, P.V.; Parshutkin, I.I.; Gaidar, A.A. Influence Of Inoculation On Photosynthesis And Symbiotic Parameters Of Pea And Its Crop Yield In The Conditions Of The Southern Forest Steppe Of Western Siberia. Vestn. NGAU 2018, 3, 23–30. (In Russian) [Google Scholar] [CrossRef]
- Afonin, A.; Sulima, A.; Zhernakov, A.; Zhukov, V. Draft Genome of the Strain RCAM1026 Rhizobium Leguminosarum Bv. Viciae. Genomics data 2017, 11, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Kuzakova, O.V.; Lomovatskaya, L.A.; Goncharova, A.M.; Romanenko, A.S. Effects Of Rhizobium Leguminosarum Bv. Viceae Strains Different In Their Symbiotic Effectiveness On Changes In Camp And Hydrogen Peroxide Concentrations In Cells Of Pea Seedlings. Russ. J. Plant Physiol. 2019, 66, 712–717. (In Russian) [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab.(LBNL), Berkeley, CA (United States), 2014; Vol. 13;
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, 1–22. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Perez, C.A.; Conrad, R.E.; Konstantinidis, K.T. MicrobeAnnotator: A User-Friendly, Comprehensive Functional Annotation Pipeline for Microbial Genomes. BMC Bioinformatics 2021, 22, 1–16. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and Taxonomy in Diagnostics for Food Security: Soft-Rotting Enterobacterial Plant Pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Wilson, K. Preparation of Genomic DNA from Bacteria. Curr. Protoc. Mol. Biol. 2001, 56, 2.4.1–2.4.5. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Metabolic summary of annotated protein-coding genes.
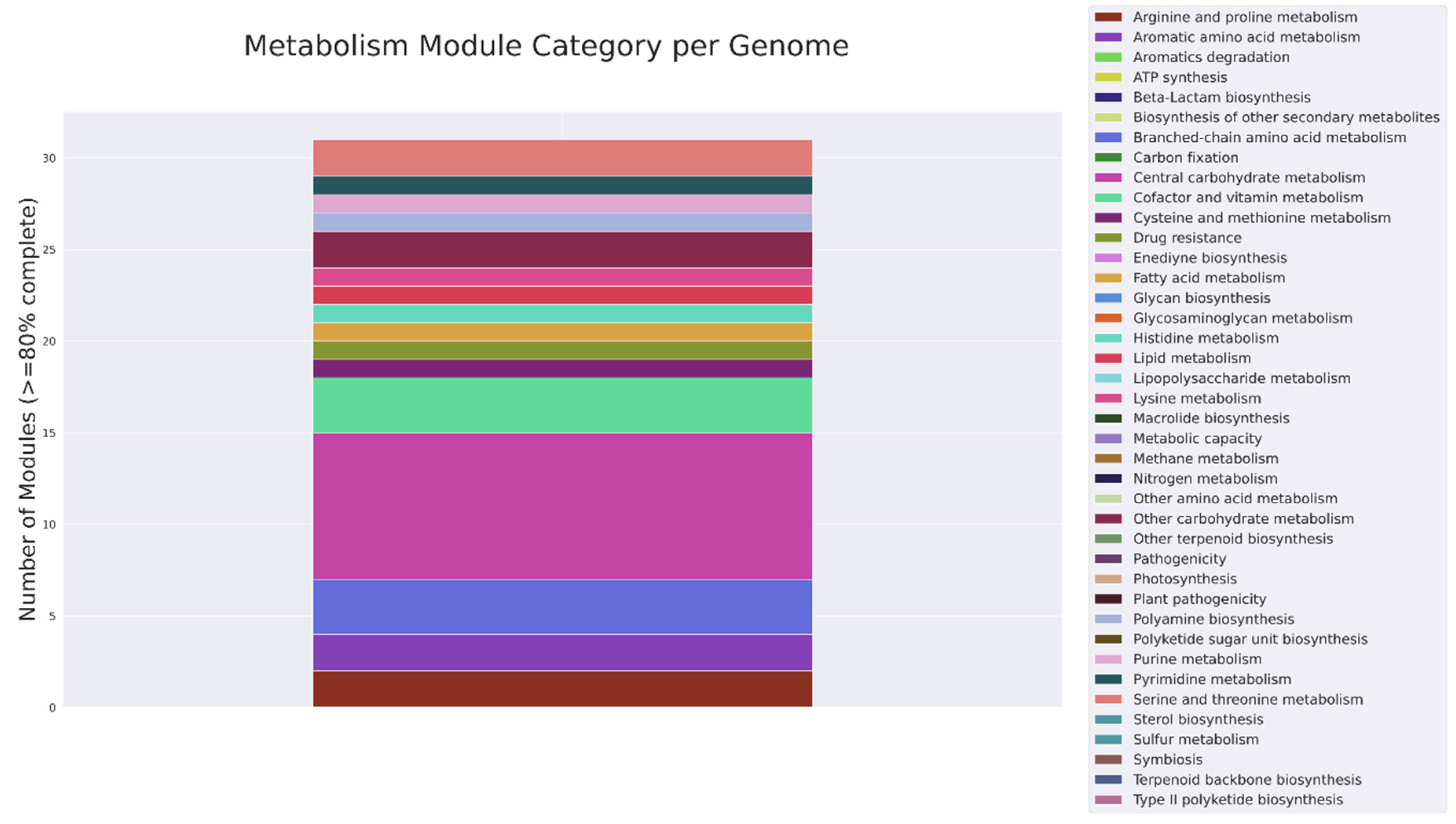
Table 2.
General description of annotated genome regions of Rh. leguminosarum strain RCAM1022.
| Contigs | 97 |
| Bases | 7240119 |
| CDS | 6901 |
| Protein coding genes | 6954 |
| rRNA | 2 |
| tRNA | 50 |
| tmRNA | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



