
Submitted:
21 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
High concentrations of sodium (Na+), chloride (Cl-), calcium (Ca2+), and sulphate (SO42-) are fre-quently found in saline soils. Crop plants cannot successfully develop and produce because salt stress impairs the uptake of calcium, potassium, and water into plant cells. Different intracellular and extracellular ionic concentration changes by salinity, including those of calcium, potassium, and protons. These cations serve as stress signaling molecules in addition to being essential for ionic homeostasis and nutrition. Maintaining an appropriate K+: Na+ ratio is one crucial plant mechanism for salt tolerance, which is a complicated trait. Another important mechanism is the ability for fast extrusion of Na+ from the cytosol. Calcium is established as a ubiquitous secondary messenger, which transmits various stress signals into metabolic alterations that cause adaptive responses. When plants are under stress, the cytosolic-free Ca2+ concentration can rise to 10 times or more from its resting level of 50-100 nanomolar. Reactive oxygen species (ROS) are linked to the calcium alterations, and are produced by stress. Depending on the type, frequency and intensity of the stress, the cytosolic calcium signals oscillate, are transient, or persist for a longer period and exhibit specific "signatures". Both the input and efflux of Ca2+ from the cytosol affect the length and amplitude of the signal. According to several reports, under stress calcium alterations can occur not only in the cytoplasm of the cell but also in the cell walls, nucleus and other cell organelles and the calcium waves propagate through the whole plant. Here, we will focus on how wheat and other important crops absorb Na+, K+ and Cl- when plants are under salt stress, as well as how calcium, potassium, and pH cause intracellular signaling and homeostasis. Similar mechanisms in the model plant Arabidopsis will also be considered.
Keywords:
cereals
; chloride
; cytosolic calcium
; K+
; Na+
; pH
; salt stress
; signaling
; wheat
1. Introduction
Soil salinity is harmful for plant growth and development as it inhibits the uptake of essential nutrients such as potassium and calcium [1]. Salinity also causes reduced water uptake, changed metabolism, ionic imbalance and toxicity [2]. In dry and warm areas, coastal areas, and where irrigation with saline water is common, salinity is often a serious problem. Sodium (Na+) and chloride (Cl-) are the most abundant elements in saline soils and cause most harmful effects. The earth crust contains 3% sodium and the seas and oceans contain more than 5% [3]. Saline soils are sometimes alkaline, which also is harmful for plants [4].
Except for halophytes and C4 plants, Na+ is not an essential element for growth or reproduction [5]. On the other hand, chloride is an essential macronutrient for many higher plants, although high chloride concentration might have a negative impact on plant growth [6,7]. Chloride has been underestimated as a toxic element, and in some plant species it is as harmful as sodium [8]. Most plant species, which are glycophytes, cannot survive in high salinity soils, but halophytes grow even better under salinity. High salinity causes both osmotic stress and ion toxicity, which in turn also induce oxidative stress [9,10].
Plants have several mechanisms to cope with salt stress and keep ion homeostasis under salinity. This review will focus on how important crops, especially wheat, rice, barley, pea and beans take up sodium, chloride and potassium, how they sense the salt stress and transfer the information by cytosolic calcium and pH signaling to metabolic downstream reactions leading to tolerance. Some findings show that also potassium can take part in the signaling, and will be discussed. Numerous studies are included in the present review that focus on cytosolic ion alterations of calcium, sodium and pH in shoot and root protoplasts, live cells without cell walls, but also sodium, potassium and chloride changes in intact plants subjected to salt stress. Under stress systemic-calcium signaling occurs within the whole plant by calcium waves from the local perception to distal target cells and organs. Here we will describe the recent advances concerning the systemic signaling, such as amino acid-induced calcium signaling.
Measurements of the influx and concentrations of different ions can be done by epi-fluorescence microscopy, video imaging and fluorescent probes, which specifically bind to the ion of interest. Fluorescence tracers can also be expressed in the cells by genetic modifying.
It is still unclear how the plant senses salt. According to investigations conducted on Arabidopsis and rice, Na+ should reach the cytosol prior to a further sensing process. Recent findings suggest that Na+ can bind to a plasma membrane sphingolipid, which then triggers a calcium influx [1], but also other possibilities for sodium sensing will be mentioned.
2. Na+ uptake and accumulation under salt stress
2.1. The hydraulic conductivity (Lp) affects ion transport
The water flow through a plant decreases under salinity and affects the ion transport. Lu and Fricke [12] investigated the root hydraulic conductivity (Lp) in wheat under different NaCl concentrations and found that Lp of cortex cells was differently affected by NaCl concentrations under day and night, and in the main axis of roots and lateral roots. The aquaporin-inhibitor H2O2 reduced Lp during the night, showing that these proteins were important for hydraulic conductivity. The authors suggested that the changes in root Lp in response to salt stress depended on altered activity of aquaporins in root and leaves. Aquaporins play key roles in hydraulic regulation of other types of abiotic stress too. They are intrinsic protein channels in plasma membranes, ER, vacuoles and plastids, facilitating diffusion of water and small neutral molecules, and dissolved gases like CO2 and ammonia. Interestingly, aquaporins can be regulated by signaling intermediates, cytosolic Ca2+, pH and reactive oxygen species (ROS) [13]. Genetically modified aquaporins could be future candidates for improving salt tolerance in plants.
2.2. Uptake of Na+ and Cl- at whole plant level
In most plant species Na+ and Cl- can easily be absorbed by both the main root and lateral roots. The ions are transported through root hairs into the epidermis, cortex, endodermal cells, and into the parenchyma cells, layers of pericycle and thereafter, into the xylem for further passive transport by the transpiration stream in the shoot. Solutes and water can travel by the epidermis and cortex cells into the xylem in three ways, apoplastically by the cell walls and extracellular spaces, symplastically from cell to cell by cytoplasm and plasmodesmata, openings in the cell walls, and by a transmembrane pathway by plasma membranes and cell walls [14]. The endodermal cells form a central ring structure where the radial walls are thickened (Casparin strips) with hydrophobic suberin that prevents the apoplastic ion transport. In the symplastic pathway ions have to pass one or several membranes.
Kronzucker and Britto, [15] investigated which way plants used for uptake of ions. Reports show that monocots, like wheat and rice, more often take up Na+ and Cl- by the apoplastic way. In rice 50 % of both Na+ and Cl- uptakes in shoot were apoplastic [15,16]. In Arabidopsis and other dicots some part of the ion uptake was significantly apoplastic. The solute permeability coefficients in Arabidopsis and rice were rather similar. By the symplastic pathway the ions have to pass several different channels or transporter proteins.
2.3. Ion uptake across a membrane
The driving force for movement of an ion across a membrane depends on two components: one electrical and one chemical, depending on differences in charges and ion activities across the membrane [17]. This electrochemical potential difference for movement of an ion across a membrane can be described by:
where Vm is the electrical potential difference with reference to the cytosol and extra-cytosolic sides, Jcyt and Jext are the activities of the ion in the cytosol and at external side and z is the valence of the ion. F, R, T have their usual meanings.
delta µNa+ = zFVm + RT ln [Jcyt /J ext],
Since the membrane potential difference across the plasma membrane is approximately around -140 mV, a positive ion like Na+ can easily pass into the cytosol even at low concentrations [18].
2.4. Cellular uptake of Na+
Na+ is transported into a plant cell by many different channels and transport proteins, such as nonselective cation channels, NSCCs, low affinity cation transporter, LCT1, cation-chloride cotransporters, CCCs, high affinity-potassium transporters, HAKs, HKT1, HKT2 and Shaker-type K channel, AKT1.
2.4.1. Cytosolic uptake of Na+ in wheat and rice by NSCCs.
The cytosolic uptake of Na+ in wheat and rice under salinity is mainly mediated by nonselective ion channels NSCCs, but also by transport proteins [3,15,19]. Monocots like wheat and rice use many different transporters. Davenport and Tester, [20] showed that the NSCCs are the primary pathways for Na+ influx into wheat roots, as the influx was inhibited by calcium, a mechanism that was considered specific for that type of channels. However, later reports suggested that both K+ and Ca2+ may inhibit Na+ influx by NSCCs and also influx by LCT1 [15].
NSCCS are divided in depolarization-activated, hyperpolarization-activated and voltage-insensitive channels. In barley the voltage-insensitive NSCCs show both inward and outward currents [21]. NSCCs can also be characterized and named by their ligands and stimuli as Cyclic Nucleotide Gated Channels, CNGCs [22,23], and Glutamate-Like Receptor proteins, GLRs [24].
2.4.2. Cytosolic uptake of Na+ in wheat and rice by CCCs, HKTs and AKT
Except for NSCCs, Na+ uptake can be mediated by cation-chloride cotransporters, CCCs, but other uptake mechanisms are proposed as well. In wheat and rice high affinity-potassium transporters, HAKs, HKT1, HKT2, low affinity-cation-transporter, LCT1, and the Shaker-type K channel, AKT1, were shown to transport sodium [25,26], and references therein. In wheat LCT1 was suggested to transport Ca2+and Cd2+ [27]. In barley HKTs were showed to improve salt tolerance, as overexpression caused increased tolerance [28]. The HvHKT1;1 in barley showed a higher selectivity of K+/Na+ and a high retention of K+ and Ca2+ in root cells under salinity [29].
2.4.3. Cytosolic Na+ uptake in Arabidopsis
2.5. Long-distance translocation of sodium
2.5.1. Sodium transport in rice by HKTs
Many reports show that HKT1 is mediating Na+ translocation from roots to shoot in rice, under salt stress. Cell-specific expression analysis by in situ-PCR revealed that HKT1 was induced in the root epidermis, vascular cylinder and shoot mesophyll cells of the sensitive rice, indicating a transport of Na+ from root to shoot, where it is more damaging [33]. HKT1 induction also occurred in shoot phloem, in the transition from phloem to mesophyll, and in the mesophyll cells of rice, suggesting a recirculating of K+ in the leaf. Horie et al. [34] showed that HKT1 mediates Na+ influx and transport, but not K+ influx and transport and that HKT2 might transport both Na+ and K+. In the tolerant rice an induction of OsHKT2 and OsVHA was obtained in the root epidermis, exodermis and xylem tissue, which indicates a K+ + Na+ uptake and transport through the root xylem [33]. This cultivar could confer salt-tolerance by decreasing expression of HKT1 and increase expression of HKT2 in the shoot, leading to a low Na+/K+ concentration ratio, which is important for salt tolerance and ionic homeostasis [35].
Investigations show that HKT1 does not transport Na+ from root to shoot in all species. AtHKT1 in Arabidopsis can prevent xylem loading and translocation to the shoot [36], but in salt sensitive rice, Na+ is translocated to the shoot by OsHKT1 [33]. OsHKT1;5 is suggested to recirculate Na+ by phloem to the root tissue [37,38].
2.5.2. Sodium transport in barley by HKTs
Different transport mechanisms of HKT1:5 was demonstrated in the cultivated barley, Hordeum vulgare, and the halophyte ecotype Hordeum marinum, as the halophyte took up less Na+ into the shoot and higher K+ into the root, compared with Hordeum vulgare when subjected to 400 mM NaCl [39]. Thus, the halophyte maintained a higher K+/Na+ ratio, and also used less energy for salt tolerance than the cultivated ecotype. In several investigations salt tolerance in the halophyte barley Hordeum maritimum was compared with salt tolerance in Hordeum vulgare. Results from those investigations are discussed in Section 4.
2.5.3. Sodium transport in Arabidopsis by HKTs
An investigation in Arabidopsis suggested that high expression of the AtHKT1;1 in the mature part of the Arabidopsis root stele decreased Na+ accumulation into the shoot and increased salt tolerance [40]. Moreover, other research on Arabidopsis indicated that AtHKT1;1 controls the root accumulation of Na+ and retrieves Na+ from xylem, but is not mediating root influx or recirculation in the phloem [41].
2.5.4. The CCCs, cation-chloride cotransporters
Loss of function experiments with CCCs showed that this protein can transport K+, Na+ and Cl- in symport [42]. CCCs are suggested to be involved in long-distance translocation of Na+ [43]. It is uncertain if the CCC are retaining Na+ and Cl- at the xylem parenchyma cells, where CCCs are expressed, or translocate these ions into the xylem. (More information on CCCs can be found in section 4).
2.6. Salt tolerance
2.6.1. Plants tolerate salt stress by different mechanisms
Salinity stress in plants causes osmotic stress, Na+ and Cl+ toxicity, and negative effects on ion homeostasis, as high salt concentration can decrease the uptake of K+, Ca2+ and NO3 [26,44]. The osmotic stress is prior to the ionic stress and, in barley, more serious for the plant to combat than ionic stress, as it might be connected with formation of ROS [45]. Osmotic stress depends on that the osmotic potential is more negative outside the roots than inside under high salt concentration, which makes water uptake more difficult.
Plants have developed several mechanisms to resist salinity [31,44]. The first minutes to days osmotic stress causes reduced water uptake and plant growth reduction. Plants can stand the first phase by reduction of the cell expansion and closing their stomata, but under ionic stress cytosolic Na+ and Cl- concentrations can be toxic and affect the metabolism that after long time may cause cell death.
As reported for barley, salt tolerance does not only depend on a low uptake of Na+, but also on the plant’s ability to retain K+ and Ca2+ [29,46]. Plant might have a higher selectivity for K+ over Na+, by accumulating a low amount of Na+ in root cells, and increase their K+ concentration at high external Na+, which would lead to K+/Na+ homeostasis [44]. Higher plants can also exclude Na+ and Cl- from the leaves, by preventing xylem loading, recirculate Na+ in the phloem, or transport these ions into the vacuoles to avoid toxic concentrations in the cytosol.
2.6.2. Halophytes and glycophytes.
Halophytes can survive much higher concentrations of NaCl, 100–200 mM Na+ and Cl-, than glycophytes, and the tissue concentration can be much higher, >500 mM [47]. Halophytes even grow better under high salt. With some exceptions, glycophytes and halophytes might own similar tolerance mechanisms, but the reaction strength is higher, or start more rapidly in halophytes [47,48]. Halophytes usually transport larger amount of Na+ and/or K+ into the vacuole than the glycophytes, to use as a cheap osmoticum. Some halophytes have salt glands which can extrude Na+ from the leaf cells [44].
High salt conditions can cause K+ deficiency both in glycophytes and halophytes [49]. One reason for this might be the competition between K+ and Na+ at the uptake sites, as these ions have similar ionic radius and ion hydration energy [50,51]. The selectivity for uptake of K+ over Na+ is different in different plants, usually Na+ can inhibit the uptake of K+, but the opposite is less common, at least if K+ concentration is less than 75 mM [52] Another reason is that the presence of high Na+ concentration causes a depolarization of the plasma membrane leading to K+ efflux from the cells by GORK and NSCCS channels [53]; Figure 1).
Depolarization also causes an influx of calcium [54]. The depolarization is not always negative as it may lead to an activation of the H+ATPase, and the protons pumped out to apoplast can be used by the SOS1 antiporter. A more recent report suggests that GORK, an outward-rectifying K+ channel, may operate to switch off energy-consuming anabolic reactions and instead use the energy for stress adaptation [55]. The K+ efflux could be a signaling mechanism under salt stress, for instance, an increased H+ATPase activity and K+ influx from the vacuole to compensate for the K+ loss. The SKOR channel, STELAR K+-outward rectifier might also be involved in K+ efflux induced by depolarization.
2.6.3. Regulation of Na+ transport at the at xylem/parenchyma cell border
Not only a low cytosolic Na+ in the plant cells is important for tolerance, but also prevention of ion loading into the xylem at the parenchyma-xylem border, for further transport to the shoot. It was shown that both root and shoot of the halophyte quinoa had a higher K+ concentration than root and shoot of the glycophyte pea, and under salinity this concentration was even higher [56]. Despite a higher concentration of Na+ in quinoa roots than in pea roots, the K+/Na+ ratio was higher in quinoa.
To explain these results electrophysiological measurements of K+ and H+ fluxes from mechanically isolated root xylem of pea and quinoa were performed [57,58]. Addition of 20 mM NaCl and ABA to the xylem medium, mimicking the natural xylem solution [59], caused a strong K+ efflux from stelar cells of pea, but no efflux from quinoa [56], reflecting a K+ retention in quinoa roots. In pea K+ was translocated to the shoot to compensate for a lower uptake of K+ in the root. There was also a H+ efflux from stelar tissue of both species, but the efflux was more pronounced in pea. ABA accumulates in the root under salt stress [60]. There it might stimulate the SOS protein [61]. Addition of 50 µM ABA induced a net H+ uptake into xylem-parenchyma cells of both species and a net K+ efflux, suggesting an ion exchanger at the xylem-parenchyma interface [62]. The results corroborate results showing a higher concentration of K+ in the shoots of quinoa than in pea under salt stress [56]. Quinoa keeps more K+ as osmotic regulation in the roots than pea does. The high concentration of Na+ found in the shoot of quinoa may depend on that quinoa uses Na+ as osmoregulation in the shoot.
2.6.4. Different barley cultivars differ in salt tolerance
Metabolomic and transcriptomic analyses of barley genotypes showed that the halophyte Hordeum marinum under salt stress used more Na+ and K+ ions for osmotic regulation and root tolerance than the cultivated Hordeum vulgare, and also increased the glycolysis and TCA cycle to get a high energy supply, necessary for shoot tolerance [63].
2.6.5. Tolerant rice cultivars have different salt-tolerance mechanisms
In three rice cultivars showing different salt tolerance, the most sensitive cultivar, cv. VD20, accumulated less Na+ than the other two cultivars [64]. The most tolerant cv. AGPPS114 accumulated more Na+ and also contained higher concentrations of proline and glycine betaine as osmotic regulation than the other two cultivars. Moreover, VD20 showed a higher expression of the HKTs transporters, HKT1;4 and HKT1;5, than the other cultivars, as analyzed by real time PCR. The authors reported that the tolerant cultivar showed higher expression of the SOS1 and NHX1, than the sensitive cultivars, which might explain the salt tolerance of the former cultivar.
2.6.6. SOS1 role in salt tolerance
A low concentration of Na+ in the cytosol is important for salt tolerance. Plants are able to transport Na+ from the cytosol into apoplast by the Na+/H+ antiporter SOS1. The localization of SOS1 in the plasma membrane was revealed by confocal imaging of a SOS1-green fluorescent protein fusion in transgene Arabidopsis [61]. Expression analyses showed that SOS1 was present in plasma membranes of root tips and in parenchyma cells of the xylem/symport boundary. Thus, the SOS1 protein should have another function too: to reduce transport of Na+ from xylem parenchyma cells into the xylem vessels, which also would be of importance for salt tolerance. Conflicting results were recent published from an investigation on gene expression of SOS1 and Na+-flux measurements, which stated that it is only the SOS1 transporters in the outer root tissue that are excluding Na+ from root cytosol, but that SOS1 operating in the stele actively loads Na+ to the xylem transpiration stream [65].
Cytosolic Na+ concentration and fluxes in living cells can be analyzed by dual-wavelength fluorescence microscopy and the sodium-binding fluorescent dye SBFI, AM [66]. By use of this technique, measurements on mesophyll protoplasts from Arabidopsis shoots showed that the sos1 mutant took up more Na+ into the cytosol than did the Wt protoplasts when external solution contained 100 mM NaCl [67]. Moreover, the Arabidopsis nhx mutant, localized in tonoplast and has a functional SOS1, also took up more Na+ into cytosol than the Wt, probable because both the Na+ /H antiporters are involved in the efflux of Na+ from cytosol. The main function of NHX is believed to be a regulator of pH and intracellular homeostasis [68]. In Arabidopsis there are 4 isoforms of NHX and nhx triple and quadruple knockouts showed reduced growth. A lack of any vacuolar-NHX activity resulted in reduced Na+ uptake and no K+ uptake, suggesting that these antiporters take up K+ but also some Na+. They also reported a Na+-uptake transporter, which was independent of proton transport.
2.6.7. Na+ and K+ transport into the vacuole
Since long time it has been stressed that the Na+/H+ antiporter, NHX in the tonoplast and in some endomembranes is important for salt tolerance, as it might transport Na+ out from cytosol. However, [69] reported that K+ concentration decreased when Na+ concentration was increased in the vacuole. Moreover, a coordinated transport by NHX and KEAs, K+ -efflux antiporters was reported to result in salt tolerance [70]. Thus, NHX might be more important for K+ transport into the vacuole and for pH regulation, even if NHX to a lesser degree also mediates transport of Na+ [68]. Recent findings suggest that the CCX, a cation/Ca2+ exchanger, is suggested as a better candidate for salt tolerance than NHX, as it deceases high concentrations of Na+ in cytosol and reduces reactive oxygen species [71].
2.7. Measurements of cytosolic ion changes in different species/cultivars under salinity
To compare the influx kinetics of Na+ species D’Onofrio and coworkers (2005) [72], Kader and Lindberg (2005) [73] (Figure 2) and Sun et al. [56] used epifluorescence microscopy and the Na+-binding fluorescent dye SBFI, AM, the acetoxymethyl ester of the benzofuran isophtalate, SBFI. The fluorescent Na+ indicator CoroNA Green, AM can also be used to measure Na+ concentrations in the cytosol. Dyes in AM-ester form can penetrate plasma membrane and enter into cytosol, where they split by esterases into the Na+-binding fluorescent form. Ratiometric measurements at two excitation wavelengths make the result more reliably, than one-wavelength measurement, as different dye concentrations, photobleaching and thickness of the cell show little effects [66,74].
2.7.1. Cytosolic Na+ influx and efflux from salt tolerant and sensitive species, quince, sugar beet and wheat, differ
D’Onofrio et al. [72] showed that the amplitude and duration of Na+ influx into protoplasts from species that are highly salt tolerant, such as quince (Cydonia oblonga Mill), salt tolerant sugar beet (Beta vulgaris L.cv. Monohill), and less tolerant wheat (Triticum aestivum L. cv. Kadett) were in the order: wheat > sugar beet > quince. The quince protoplasts took up sodium only from a calcium free buffer containing 200-400 mM NaCl. The Na+ influx was transient in quince, but in wheat and sugar beet it increased to a certain level and was then stable. As 1.0 mM external Ca2+ inhibited the influx, it is likely that Na+ influx at high salt were mediated by nonselective-cation channels [30,75]. Only the halophyte quince was able to a rapid efflux of Na+ from the cytosol.
2.7.2. Cytosolic Na+ and pH changes are different in the halophyte quinoa and the glycophyte pea
Different dynamics were obtained in the cytosolic influx of Na+ in the glycophyte pea, Pisum sativum, and the halophyte quinoa, Chenopodium quinoa [56]. The cytosolic Na+ concentration, [Na+cyt], was analyzed in mesophyll protoplasts after cultivation of seedlings with and without 100 mM NaCl, and upon addition of NaCl to the protoplast medium.
Addition of 100 mM NaCl to control protoplasts of quinoa caused a transient [Na+ cyt] influx and the maximal concentration was obtained after 360 s, at the same time as for salt adapted quince [56,72], but when NaCl was added to salt-cultivated quinoa protoplasts, the maximal Na+ influx was obtained later (after 450-500 s) and was mainly transient [56].
Addition NaCl (50 mM) to pea-control protoplasts caused another reaction: the influx increased and was then stable at the same Na+ level. In addition, NaCl addition to pea protoplasts from seedlings pretreated with salinity showed a gradual increase for a long time.
The different reaction obtained in salinity-cultivated quinoa, suggests an adaptation mechanism, and might depend on a rapid activation of the H+-ATPase, which was absent in pea, as the Na+ influx in pea was increasing with time [76]. Quinoa grows optimally at 100-200 mM NaCl, but is tolerant not only to salt stress, but also to drought and frost, and thus suitable for cultivation at areas where no other plants survive [77].
2.7.3. Cytosolic Na+ influx and efflux in tolerant and sensitive rice
The same difference in Na+-influx dynamics were also obtained when comparing the influx in tolerant rice cv. Pokkali with sensitive rice cv. BRRIDhan29 [73].
Pharmacological analysis indicated that NSCCs were the main pathways for Na+ influx in the tolerant rice cultivar, but that both NSCCs and high-affinity potassium channels HKTs contributed to influx in the sensitive one. The transient influx of Na+ suggests that tolerant rice has a mechanism for fast extrusion of Na+ that protects the cytosol from ion toxicity. This rice cultivar also has less PM permeability to Na+, compared to salt-sensitive rice [78]. Inhibitor analysis suggested that tolerant rice transported Na+ from the cytosol into the vacuole by the Na+/H+ antiporter NHX in the tonoplast, but the sensitive cultivar transported Na+ out of the protoplast by the Na+/H+ antiporter SOS1 in plasma membrane [79,80,81,82].
Expression analyses by real-time RT-PCR of the OsVHA genes, which encode the H+-ATPase in the tonoplast demonstrated that the OsVHA transcripts were induced immediately after Na+ stress in the tolerant rice cultivar, but in the sensitive one, OsVHA it was low and delayed 6h. The tonoplast H+-ATPase is supposed to be important for salinity tolerance as it builds up an electrochemical gradient for transport of cations into the vacuole [34,83] These findings confirmed the results from protoplast experiments that showed a fast efflux of sodium from the cytosol of the tolerant rice, but little sodium efflux from the sensitive rice [33,73].
3. Cl- uptake and transport under salinity
Cl- is primarily present in the soil solution as the chloride monovalent anion, and unlike other important soil anions like nitrate and sulphate, Cl- is not chemically changed by soil bacteria. Under salt (NaCl) stress, Cl- passively enters root cells and radially enters into xylem vessels for translocation to the shoot. At a low concentration, Cl- is taken up in the root by Cl-/2H+ symporters in a secondary active way. At high concentrations in the cytoplasm, both Na+ and Cl- ions are biologically toxic to plants [84]. Reports show that in glycophytic species the root-Cl- efflux is strongly associated with shoot Cl- exclusion and salt tolerance [85,86,87]. A low concentration of Cl- in the roots or shoots of several plants, such as faba bean, barley, Lotus, and Chrysanthemum, is positively correlated with salt tolerance [8,88,89]. The salt tolerance of many plants is linked to an effective management of Cl- uptake and transport. Teakle and Tyerman [84] highlighted key Cl- -transport characteristics related to plant salt tolerance: (i) decreased net Cl- uptake by roots, (ii) decreased net xylem Cl- loading, (iii) intercellular Cl- compartmentation, (iv) intracellular Cl- compartmentation, and (v) phloem recirculation and translocation within the plant. Effective Cl- exclusion from roots or shoots could prevent excessive Cl- accumulation in plant tissues. In Figure 3, the overall mechanisms of Cl-transport were briefly outlined.
3.1. Antagonism between Cl- and other anions in wheat and other cereals, and in tomatoes and rose plants
According to Abdelgadir et al. [90] there is an antagonistic relationship between Cl− and nitrate (NO3-) when external Cl- concentrations are too high, which inhibits wheat development and yield [91]. Interestingly, phosphate (PO43-) and Cl- have also been reported to face close competition for anion-anion absorption. Such competition has also been documented for tomatoes and rose plants, in addition to cereals [92]. It appears that Cl- tends to impede crop growth and development under NaCl salinity by causing phosphorus and sulphur deficiencies by blocking PO43- and SO42- absorption. This can occur through two different mechanisms: either Cl- leaks from protein pores, which quantitatively dislocate SO42- or PO43- and cause a dramatic drop in their uptake, or antagonistic competition for a binding site at transport proteins of salt ions between Cl- and counter-cations.
3.2. Wheat leaves might accumulate less Na+ and Cl− than leaves of barley, canola, and chickpea
Under salinity wheat is significantly suffering from poor plant development and water uptake, but its leaves exhibit far lower levels of Na+ and Cl− than those of barley and canola [93]. The Cl− content in wheat leaves increased to 84% with a rise in subsoil NaCl from -126 to -270 osmotic potential values (ECe values from 3.5 to 7.5 dS/m). This study demonstrates that at all subsoil salinity levels, wheat has a superior salt exclusion mechanism at the roots than canola or chickpea. It was hypothesized that there was limited transfer of Na+ and Cl− from roots to shoots in wheat plants, since the concentration of Na+ and Cl− in wheat leaves was much lower than that of barley, canola, and chickpea leaves at the highest level of subsoil NaCl [93].
3.3. Chloride channels and -transporters under salinity
Inadequate compartmentalization of Cl- between the apoplasm, the cytoplasm, and the vacuole in the leaves might cause salt toxicity [84]. In root cells, Cl- is transported inward via Cl-/H+ co-transporters, anion channels and nitrate transporters (NRTs). Different anion channels are involved in the passive transfer of Cl- during salinity. Under salt stress conditions, the rate-limiting step for Cl- accumulation in the shoot is the root-to-shoot xylem transport pathway [94].
3.3.1. Voltage-dependent chloride channels CLCs
Prokaryotic and eukaryotic organisms contain voltage-gated chloride channels of the CLC-type family, which mediate passive Cl- transport driven by the electrochemical gradient. Under salt stress, these channels mediate Cl- homeostasis in plants.
The regulation of net Cl- absorption under salinity depends on anion channels that open to allow anion outflow. In barley plants pre-treated with 200 mM NaCl, Yamashita et al. [95] discovered an increase in Cl- permeability of plasma membrane vesicles from the roots. Various investigations have reported the presence of Cl- efflux channels in root plasma membranes [96,97,98,99]. Electrophysiological studies found that the plasma membrane of A. thaliana cortical cells and epidermal root hairs exhibited substantial levels of depolarization-activated anion efflux channels [100,101].
Uptake of Na+ and Cl- and their translocation from root to shoot may differ in different wheat species and cultivars
Two wheat species, Triticum aestivum, cv. Tanit, and T. durum, cv., were compared by Ouergh et al. [102]. Cultivar Ben Bachir showed a noticeable difference in the ion partitioning between the roots and the shoots. Tanit roots and Ben Bachir shoots were the primary locations where Na+ and Cl- ions accumulated specifically. Compared to Ben Bachir wheat plants, Tanit plants accumulated significantly less Na+ and Cl- ions in their leaves. Ben Bachir leaves accumulated more Na+ and Cl- ions under salinity, and this was accompanied by a shift in the osmotic potential values to more negative values. Ben Bachir demonstrated traits of NaCl-tolerant species. Furthermore, photosynthetic properties of the resistant Ben Bachir cultivar were unaffected by salinity stress.
Cl-uptake and transport rates may vary among genotypes of wheat and increase linearly with increasing salt. There is no connection between the accumulation of Na+ and Cl- ions or the total of Na+ and K+. Rates of Cl-transport from roots to shoots in all genotypes were increased with increasing salinity. The pattern of root Cl- concentration was identical to that of Na+ [103].
Experiments with wheat protoplasts from the epidermis and cortical cells demonstrated the presence of outward rectifying anion channels that are triggered by depolarization (OR-DAACs). While the characteristics of these channels are similar across species, the rates of activation and rectification vary [104,105].
Cl- influx channels in rice by CLCs
Rice OsCLC1, a voltage-dependent Cl- channel demonstrates an adaptational response to elevated Cl-, but involvement in nitrate absorption is less probable in highly salinized environments. OsCLC1 coordinates the control of anion and cation homeostasis in salt-treated rice, contributing to salinity adaptation. OsCLC1 transcript levels were downregulated in the leaves and roots of the salt-sensitive Cl- accumulating rice line IR29 in response to salt stress, but expression was briefly upregulated in the salt-tolerant Cl- excluding rice line Pokkali [106]. OsCLC-1 and OsCLC-2 are homologs of the tobacco CLC-Nt1. OsCLC-2 was exclusively expressed in the roots, nodes, internodes, and leaf sheaths, while OsCLC-1 was expressed in the majority of tissues. Treatment with NaCl enhanced the level of OsCLC-1 expression but not OsCLC-2 [107].
CLC- channels in tonoplast of soybean and cotton decreases Cl-/NO3- ratio
Salinity caused tonoplast-localized GsCLC-c2 in soybean to transport Cl- more efficiently than NO3- into the vacuoles in a pH-independent way. Therefore, GsCLC-c2 could lower the Cl/NO3 ratio by maintaining a relatively high NO3 level in the aerial parts of salt-stressed plants. Thus, is likely that GsCLC-c2 reduces salt stress in planta by preventing excess Cl- within the vacuoles of root cells and preventing Cl- from translocation to shoots [108].
Cotton plants subjected to chloride concentrations (by NaCl or KCl) activate the tonoplast-chloride channel GhCLCg-1. A silencing of GhCLCg-1 increased the Cl- concentrations in the roots, stems, and leaves and the Na+/K+ ratio in the stems and leaves, resulting in reduced salt tolerance. Thus, GhCLCg-1 can positively control salt tolerance by regulating ion accumulation of upland cotton [109]. GhCLCg-1 transcript levels were greater in roots and leaves during salt stress but not in stems. By increasing the concentration of Cl- in the roots and reducing the quantity of Cl- delivered to the shoots, Gossypium hirsutum ghclg1 and A. thaliana atclcg mutants altered plant sensitivity to chloride and elevated Cl- concentrations [110,111].
Voltage-dependent influx channels in barley and maize
Patch clamp studies with protoplasts of barley [112] and maize [113] root xylem parenchyma cells identified multiple Cl--permeable anion channels, including an inwardly rectifying anion channel (X-IRAC), activated by plasma membrane hyperpolarization, a slowly activating anion channel (X-SLAC), and a quickly activating anion channel (X-QUAC).
Voltage-dependent SLAC/SLAH channels
The slowly activating (S-type) channel called SLow-type Anion Channel Associated/SLAC1 is activated by depolarization and is controlled by the external anion activity [114,115,116]. SLAC1 mediates the efflux of chloride or nitrate. In xylem-pericycle cells, the anion channels SLAH1 and SLAH3 of Arabidopsis co-localize. The heteromeric channels function jointly to form chloride-conducting channels. Soil salinity reduces SLAH1 expression, which results in Cl- exclusion from the shoot. AtSLAH1 expression significantly decreased when the soil was highly salinized. AtSLAH1 loss-of-function mutants only had a xylem-sap chloride concentration that was half that of the wild-type. Under salt stress, plants modify the assembly and differential expression of the SLAH1/SLAH3 anion-channel subunits to regulate the distribution of NO3- and Cl- between the root and shoot. Without considerably influencing the inclusion of NO3-, SLAH1 abundance contributes significantly to the Cl- exclusion [117].
The involvement of SLAC/SLAH in chloride efflux from barley and Arabidopsis
The protein family known as Homologues (SLAC/SLAH) interacts with K+-channels like KAT1 and AKT2 and regulates K+ homeostasis in plants [118,119]. According to Planes et al. [120], A. thaliana root-epidermal cells emit Cl- when exposed to ABA. Microelectrode measurements of SLAH3-dependent Cl- currents in root epidermal cells suggest that S-type channels may play a role in controlling net Cl- uptake in plants. Salinity-sensitive barley cultivars had more Cl- in the root cortical cells than salt-tolerant types had [121]. It was shown that transcripts of the anion channels HvSLAH1 and HvSLAC1, which facilitate Cl- efflux, were positively correlated with grain yield in the barley leaves and this shows that in barley, shoot Cl- exclusion and salt tolerance are related [122]. Furthermore, the downregulation of SLAH1 by salt stress suggests that the expression of SLAH3 and SLAH1, which is linked to xylem Cl- loading, may play a role in salt tolerance [123].
Calcium-activated Cl- efflux channel in sorghum
It was observed that a voltage-dependent anion channel in mesophyll cells, activated by an increase in cytosolic calcium, mediated chloride-ion efflux depending on the extremely negative membrane potential, producing an outward-directed anion gradient. Cl- was shown to be extruded from the roots of salt-stressed sorghum plants [124].
Intracellular chloride-channels in Arabidopsis
In the epidermal cells of Arabidopsis hypocotyls, a voltage-dependent anion channel with two functional modes: rapid and slow modes was discovered [125]. Two different forms of cytosol-negative potential-activated (hyperpolarization-activated) anion channels, VCl for Cl- and VMal for malate, are present in the tonoplast [126]. In electrophysiological studies using planar lipid bilayers, VCCN1 was shown to perform as a voltage-dependent Cl- channel, and it was consequently postulated that it mediates Cl- import into the thylakoid lumen of Arabidopsis thaliana [127].
Chloride-efflux channels in guard cells
A guard cell anion channel (GCACl) was observed in the plasma membrane
of isolated faba bean protoplasts [128]. The control of salt efflux from guard cells by voltage dependent anion channels was validated by Cl- currents of individual guard cells obtained in Patch-clamp experiments. The sensitivity of GCACl to chloride is within the physiological range for the number of exogenous anions [129].
3.3.2. Stretch-activated Cl- efflux channels in guard cells and pollen tubes
Along with voltage-gated channels, cells of higher plants have stretch-activated channels in the plasma membrane. The existence of stretch-activated Cl-, Ca2+, and K+ channels in the plasma membrane of guard cells was demonstrated by Cosgrove and Hedrich [130]. Due to the fact that stomatal guard cells have a variety of mechano-sensitive ion channels, they may be able to translate a change of salinity-induced xylem pressure into a change in stomatal apertures [131,132]. MSL10 (mechanosensitive channel-like 10) prefers Cl- over Na+ (6:1) and may be involved in Cl- efflux to release membrane tension after the channel opens [133]. Pollen tubes from mutant MSL8 plants rupture more frequently, which is consistent with the observed efflux of Cl- during pollen tube formation and suggests that MSL8 is involved in maintaining cell osmolarity [134,135].
3.3.3. Ion channels in xylem parenchyma cells of the root
To release ions into the xylem, anion channels in the xylem parenchyma of roots may work similarly to those in guard cells [97]. Activation of the KORC (K+ outward rectifying conductance) channel, which was found in both the xylem parenchyma cells of barley and stelar cells of maize, causes Cl- efflux via this channel. The downhill gradient and the negative membrane potential of the plasma membrane, induce anion efflux from the cytoplasm into the extracellular space. This leads to a depolarization, which may facilitate a propagation of signals in the cytoplasm by the plasmodesmata connections between cells [131].
3.3.4. ALMT channels
R-type anion proteins are encoded by the ALuminium-Activated Malate channel ALMT family in plants. It has been suggested that the maize plasma-membrane channel ZmALMT1, which is selective to sulphate, Cl-, and NO3- is mostly expressed in mature root tissues [136]. Anion channels ALMT6 and ALMT9 are permeable to a variety of anions, including Cl and malate. Smaller stomatal openings in AtALMT9 plants make them more resistant to osmotic stress, and AtALMT9 also mediates inward-chloride currents that are activated by the presence of low amounts of cytoplasmic malate [136]. It is known that the tonoplast transporters/channels CLCa, CLCc, ALMT9, and ALMT6 all mediate anion inflow into the vacuole, suggesting that they may all work similarly to DTX33 and DTX35 (detoxifying efflux carrier 33 and 35).
Calcium, boron, malate and aluminium affect the accumulation of Cl-.in wheat
Every wheat genotype under investigation showed a notable capacity to regulate the accumulation of Cl-. As the Ca2+ concentration rose, so did the Cl- concentration. This may be related to the rise in K+ that occurs when Ca2+ increases. Furthermore, it was discovered that no variations in Cl- concentration were seen between many genotypes, including tetraploid and hexaploidy [137].
The leaves of wheat plants stressed with salt alone showed an increase in both Na+ and Cl- concentrations; however, the leaves of wheat plants stressed with salt and a high dose of boron showed a significant decrease in Cl- concentration. As a result, under salinity, boron has a positive effect on the concentrations of harmful ions [138,139].
The selectivity of aluminum-activated malate transporter1 (TaALMT1) for malate over Cl− is complex in wheat varieties resistant to aluminum (Al). Without Al, TaALMT1 is partially active and might be involved in Cl− fluxes across the plasma membrane. The channel becomes less selective for malate than for Cl− at high external Cl−, but the transport is more selective for malate than for Cl− at low external Cl−. The ability of the root tips to excrete malate may be diminished if Al stress and salinity co-occurred due to the shift in reversal potential to more negative potentials, depolarization brought on by salinity, and potential competition between Cl− and malate at the cytoplasmic face of the channel [84,140].
3.5. Cl- transporters under salinity
3.5.1. CLCs transporting Cl- in antiport with protons
A strategy for shoot- Cl- exclusion, and for regulating cytoplasmic Cl- at a tolerable range involves sequestering of Cl- into the vacuole [84,141,142,143,144,145]. Electrophysiological studies show that the plasma membrane of root-hair cells, beside Cl- channels, contains electrogenic
Cl-/2H+ cotransporters, that mediate either Cl- influx or Cl- efflux across the plasma membrane. There are evidences that the Cl-/nH+ antiporters also mediate Cl- influx and Cl- effluxes across the tonoplast.
CLCs in maize, soybean and Arabidopsis
It was shown that both maize and Arabidopsis confer salt tolerance by regulating Cl homeostasis. ZmCLC-d overexpression in Arabidopsis improved salt tolerance [146]. A maize CLC gene, ZmCLCg, was linked with salt tolerance in a GWAS analysis. Therefore, the salt-hypersensitive phenotype lacking ZmCLCg obtained a prominent Cl- accumulation in shoot tissue [147].
GmCLC1, a vacuolar pH-dependent Cl- transporter in soybean that also functions as an H+/ Cl- antiporter, controls the accumulation of Cl- in shoots and improves soybean salt tolerance [148,149,150]. The overexpression of wild soybean GsCLC-c2 enhances Cl- accumulation in the root, enhances Cl- exclusion from the shoot, and raises NaCl resistance [108]. Na+ exclusion and salt tolerance are influenced by soybean GmSALT3, an endoplasmic-reticulum transporter in cells connected to the root vasculature. Additionally, it facilitates Cl- exclusion from the shoot and therefore GmSALT3/CHX1 is regarded as a K+/H+ exchanger [151,152,153].
Mesophyll cells, hydathodes, and phloem of the leaf exhibit high expression of AtCLCg, a vacuolar Cl- transporter and homolog of AtCLCc [154].
Subcellular CLCs transporters in Arabidopsis
Seven CLCs found in Arabidopsis thaliana are localized to the organellar membrane [155]. The spinach AtCLC-f ortholog is localized in the outer membrane of chloroplasts. The subcellular localization and amino acid sequence imply that AtCLCg may be involved in detoxifying the cytoplasm of leaf cells during NaCl stress by sequestering Cl--ions in the vacuole. GhCLCg-1 overexpression in Arabidopsis thaliana improved salt tolerance [110]. In the absence of AtCLCg, a significant amount of Cl- is either in the cytoplasm or in the apoplasm where it inhibits enzyme reactions or increases water loss, respectively [84,156]. When Arabidopsis was subjected to NaCl stress they showed elevated expression of AtCLCg. This process is crucial because it enables the plant to expel ions when their ion concentration rise too high [157].
The AtCLCg protein regulates the phloem recirculation of Cl-, thus preventing overaccumulation of this ion in photosynthetically active leaves during salt stress. This procedure may help to increase phloem loading and contributes to the translocation of Cl- to petioles, younger leaves, or roots) [157].
3.5.2. Nitrate Transporter 1/Peptide Transporter Family (NFP)
The plasma-membrane localized NPF transports nitrite, glucosinolates, phytohormones, and Cl- and is involved in the influx of 2H+:NO3- [158]. AtNPF2.5 encodes a Cl-permeable transporter that regulates root-to-soil Cl- efflux and enhances shoot Cl- exclusion and salt tolerance in Arabidopsis [141]. The Zm-NPF6.6 in maize, a NO3- preferring transporter, also transports Cl-. In maize the plasma membrane localized Zm-NPF6.4 and Zm-NPF6.6 were identified as Cl--preferring transporters [159]. It was demonstrated in Arabidopsis that Cl- translocation from root to shoot was accelerated by the rapid downregulation of AtNPF2.4, a Cl- transporter located in the plasma membrane of root stelar cells [142].
Accumulation of Cl− in wheat is inhibited by silicon
The Cl− absorption in salt-stressed wheat cultivar Vinjett was decreased by silicon supplementation, revealing that Cl-transport from roots to shoots is regulated. Additionally, it was noted that the wheat cultivar Vinjett expresses two Cl-transporter genes, TaCLC1 and TaNPF2.4/2.5, in both the roots and the shoots [160]. Since TaCLC1 was more highly expressed in roots than in shoots, it is most likely that more Cl− is accumulated in the root cell vacuoles and less is transferred to the shoots as a result of Si action. Moreover, the cytosol of leaf mesophyll protoplasts significantly absorbed Cl− under salinity. If Si was supplied before NaCl, this absorption was greatly reduced. This outcome supports the findings of the TaCLC1 expression analysis, which showed that seedlings treated with Si and NaCl expressed more TaCLC1 than those treated with NaCl alone. This should improve the sequestration of Cl− into the vacuoles, particularly in shoots. The effects of Si on TaCLC1 and TaNPF2.4/2.5 may be the cause of the decreased concentration of Cl− in the cytosol of leaf mesophyll cells after treatment, although other Cl− transporters might also be involved. It is hypothesized that this makes the wheat plant more resistant to high Cl− concentrations when Si is added. While vacuolar chloride is not harmful, high quantities of chloride present in the cytosol is toxic [160].
3.6. Cl- Compartmentalizing transporters under salinity
3.6.1. Cation chloride cotransporters (CCCs)
The coordinated symport of K+, Na+ and Cl-, is catalyzed by cation Cl- cotransporter proteins. These transporters play a direct or indirect role in controlling root Na+ and Cl- absorption in both glycophytes and halophytes.
CCCs in rice, soybean, grapevine and Arabidopsis
In rice CCCs are controlling the ion homeostasis. K+ and Cl- concentrations were lowered when OsCCC1 was knocked out [161]. OsCCC1 in rice is a plasma membrane-localized transporter and participates in cell elongation.
In Arabidopsis, the CCC1 is localized to the Golgi and trans-Golgi network (TGN) [162]. Under salt stress, the atccc mutants have higher shoot Cl- [163,164]. VviCCC in grapevine (Vitis vinifera) and AtCCC in Arabidopsis, have been postulated to be involved in long-distance ion transport [162]. Inflorescence stems, roots, leaves, and siliques all had shorter organs when AtCCC was knocked out [42], demonstrating that AtCCC is involved in development and Cl- homeostasis. Grapevine VviCCC gene complementation of the atccc mutant lowered shoot Cl- and Na+ concentration to wild-type levels under moderate salinity.
4. K+ concentrations and signaling under salinity
To maintain K+ homeostasis, plants need efficient K+ absorption pathways, especially under salt stress during which a spike in Na+ content is always accompanied by K+ loss from roots and shoot tissues, which disrupts the cytosolic Na+/ K+ ratio, a crucial property of plant salt tolerance [165]. Plant cell K+ concentration represents K+ uptake and efflux. Cell and tissue-specific K+ channels and transporters mediate these activities. This review covers salinity-induced alterations in cellular K+ content, uptake, transport, efflux, and signaling cascades.
4.1. Cytosolic K+ retention is higher in salt-tolerant plants
Plants' ability to store cytosolic K+ during salt stress is crucial to salt tolerance. Smethurst et al. [166] found that salt-tolerant lucerne genotypes retained more K+ in their roots. Salt-tolerant types of barley, bread wheat, brassica, poplar and cotton had higher cytosolic K+ retention in mesophyll tissues than their salt-sensitive counterparts [167].
4.2. K+ channels under salinity
The identified 77 K+ permeable channel genes in Arabidopsis were classified into two major groups: 15-gene K+ selective channels (9 Shaker and 6 Tandem-Pore K+ channels) and 62-gene non-selective cation channels (NSCCs) which include 1 two-pore channel (TPC), 20 cyclic nucleotide gated channels (CNGCs), 20 ionotropic glutamate receptors (iGLRs), 10 mechanosensitive-like channels (MSLs), 2 "Mid1-Complementing Activity" channels (MCAs), 1 mechanosensitive Piezo channel and 8 annexins [168,169,170]. This review examines how salt stress affects the functions of each K+ channel.
4.2.1. Shaker K+ channels under salinity
Shaker K+ channels allow enormous, passive K+ fluxes through cell membranes. K+ channels maintain electrical and osmotic balance. Cellular signaling and metabolic regulation in response to oxidant and salt stressors depend on shaker K+ channels [171]. Shaker channels dominate membrane conductivity in most cells.
Shaker K+ channels in Arabidopsis
Nine genes in Arabidopsis thaliana encode four functionally distinct plant Shaker K+ channels. (i) Kin channels: the AKT1, KAT1, KAT2, AKT5, and SPIK (Shaker Pollen Inward K+ channel) genes encode inwardly-rectifying K+ influx channels which transport K+ into the cell when the plasma membrane is electrically hyperpolarized (ii) Kout channels: the depolarization releases K+ from Kout channels. GORK (Gated Outwardly Rectifying K+ channel) and SKOR (Stelar K+ Outward Rectifier) encode Kout channels. (iii) Kweak channels: weakly rectifying AKT2/3 channels mediate Kin-like influx or bidirectional influx/efflux K+ channels and (iv) Ksilent channels: these channels do not conduct K+ currents, but when hetero tetramers with Kin channel subunits, they regulate K+ homeostasis. Only KC1 encodes the Ksilent channels [172,173]. Plant Shaker K+ channels appear to play role in stress-related responses, specifically osmotic adjustment by maintaining high cytosolic concentrations [174].
Shaker K+ channels in rice and soybean
It has been demonstrated that the Shaker potassium channels OsKAT1 and OsAKT2 in rice promote salt tolerance and protect yield losses from salinity stress [175,176]. Wang et al. [177] reported that GmAKT1 expression was induced by salinity. GmAKT1 overexpression in Arabidopsis promoted plant growth and enhanced K+ concentrations, which decreased Na+/ K+ ratios. GmSKOR, GmsSOS1, GmHKT1, and GmNHX revealed increased expression in the transgenic Arabidopsis expressing GmAKT1. Additionally, Feng et al. [178] demonstrated that three K+ channel genes (GmKAT2.1, GmSKOR.1, and GmGORK.2) in roots and five genes (GmAKT6.2, GmAKT1.1, GmKAT2.1, GmSKOR.1, and GmGORK.2) in leaves were elevated in response to the salt stress treatment. GmAKT1 was markedly elevated in both leaves and roots in response to both salt and drought stress.
At a high concentration of NaCl, ROS increased GORK-mediated K+ efflux in rice [179], while NaCl treatment reduces root K+ transporters and channel-associated genes (OsGORK, OsAKT1, OsHAK1, and OsHAK5). OsRbohA knockout mutant disrupted K+ homeostasis. Overexpressing OsRbohA boosted K+ transporter and channel gene expression, reducing root K+ loss [180].
4.2.3. Tandem-pore K+ channels
During stress, three K+ channels: slowly activating channel (SV), fast vacuolar channels and vacuolar K+ channel (VK) transport K+ between the cytoplasm and vacuole [181]. Maathuis et al. [182] found higher SV-channel activity in leaf vacuoles from the extreme halophyte Suaeda maritima cultivated in highly salinized circumstances. K+ transport blockage may cause osmoregulation issues in salt-stressed plant cells [26]. A rise in luminal Na+/ K+ ratio reflected Na+ accumulation in vacuoles under salt stress and changed the threshold for SV activation to positive potentials, reducing the SV channel open probability under saline circumstances. Under salt stress, plants regulate SV channel activity differently to limit Na+ leakage into the cytoplasm [183]. Choi et al. [184] were the first to show that TPC1 affects the velocity of the salt-induced Ca2+ wave in Arabidopsis and salt stress induced Ca2+ waves in the tpc1-2 knockout mutant, although they were weaker. Whereas the TPC1 overexpression accelerated the wave.
4.2.4. Non-selective cation channels (NSCCs) under salinity
The fundamental entry points for potassium ion extrusion and sodium ion uptake in roots are NSCCs [185]. Plant NSCCs are widespread at the plasma and tonoplast membranes.
With regard to K+ efflux in response to salt stress, depolarization-activated non-selective activation channels (DA-NSCCs) play a significant role [46]. Wu et al. [186] demonstrated a salt-induced K+ efflux from wheat mesophyll cells similar to that in Arabidopsis. When roots are exposed to salinity, the strong inwardly directed electrochemical Na+ gradient drives a major flux of Na+ into root cells, possibly via NSCCs, and, this results in a significant plasma-membrane depolarization [187,188,189]. The depolarization-activated GORK channels react to this alteration quickly and facilitate the enhanced K+ efflux. The ROS-induced NSCC for K+ exclusion in pea roots was demonstrated by Bose et al. [190]. ROS, created when cytosolic K+ decreases and cytosolic Ca2+ increases further activate ROS-producing NADPH oxidases and amplifies GORK-mediated K+ efflux under high salinity stress. This is the molecular mechanism enabling the K+ efflux mediated by ROS-activated NSCCs [191].
4.2.5. Cyclic nucleotide-gated channels (CNGCs) and glutamate-like receptors (GLRs)
The CNGCs and GLRs are two major NSCCs in plants. Under salinity, at least some CNGCs exhibit equal permeability for K+ and Na+, which may have an effect on cytosolic K+/ Na+ ratios [192]. Mutants such as atcngc3 and atcngc10 of Arabidopsis and oscngc1 of rice showed better growth by preventing the excessive inflow of Na+ [193,194,195].
Plant GLRs function as NSCCs, much like the ionotropic glutamate receptors (iGluRs) in mammalian cells and are ligand-gated channels permeable to Ca2+, K+, and Na+. Under salt stress, the glutamic acid gradient in the root tips is enhanced to high level which function as an activator of GLRs [46]. When exposed to salt stress, glutamate generated in the root tips stimulates GLRs, which then activate the plasma membrane's NADPH oxidase to produce too much hydrogen peroxide (H2O2). This further activates outward-rectified K+ channels and results in programmed cell death [196].
Transgenic plants which overexpressed AtGLR2 are more sensitive to K+ and Na+ salt. All 20 AtGLR genes have significant levels of expression in Arabidopsis roots, suggesting that AtGLRs are crucial for controlling ion intake from the soil, especially K+ uptake [197]. The calcium-dependent protein kinase (CDPK) phosphorylates AtGLR3.7 and influences calcium signaling, and the atglr3.7-2 mutant was hypersensitive to salinity [198]. AtGLR3.4 mediates Na+ accumulation in germinating seeds under salt stress because the ratio of Na+ to K+ was higher in atgl3.4-1 seedlings than in wild-type seeds under salt stress [199].
4.2.6. Two-pore K+ channels under salinity
Vacuolar TPK channels are both mechanosensitive and osmo-sensitive. They may serve as cellular osmo-sensors during abiotic stresses [200]. Under salt stress, VK channels provide two beneficial functions: they export K+ from the vacuole to increase the cytosolic K+/ Na+ ratio and act as a shunt conductance for H+-pumping [201]. Arabidopsis TPK1, maintains K+ homoeostasis under salinity stress by closing stomata via KIN7 kinase and assisting plants in resisting salt tolerance [202]. Significant salt-induced variations in the transcripts of TPK (VK) were observed [203].
When exposed to salt stress, calcium-dependent protein kinases (CDPKs) phosphorylate the TPK1 vacuolar K+ channel, and both the cpk3 and tpk1 mutants showed salt-sensitive phenotypes. TPK1 and CPK3 are constitutively co-expressed, and calcium activates them both. Salt stress greatly induces VK/TPK1, and interactions between the cytoplasmic N-terminal region and 14-3-3 proteins increase the probability of VK channel opening [204]. Since NtTPK1 showed considerable selectivity for K+ over Na+, the fact that tobacco TPK1 expression increased under salinity suggests that this protein may be involved in delivering K+ into the cytosol [203].
4.2.6.1. TPKs in rice
4.3. K+ transporters under salinity
High Affinity K+ (HAK) / K+ Uptake Permeases (KUP) / K+ Transporter (KT) family, K+ and/or Na+ selective High-affinity K+ Transporter (HKT) and cation: proton antiporter (CPA) family are the three major K+ transporter families in plants [196,207].
4.3.1. HAK transport systems under salinity
During the early stages of salinity, the HAK/KUP/KT transport systems play a major role in K+ absorption [167]. Numerous plants such as corn [208], barley [209], rice [210], tomato [211], peach [212], pear trees [213], Ipomoea [214], tea [215], cotton [180] and Arabidopsis [201], have been shown to contain HAK/KUP/KT transporters. In addition to their crucial roles in K+ absorption and transport, the HAK/KUP/KT family is shown to confer tolerance to both salt and drought stresses.
HAKs in rice, maize, Medicago and pepper
When rice OsHAK5 was expressed in tobacco BY2 cells, it was shown that the cells accumulated a significant quantity of K+ rather than Na+ when exposed to salt stress. This finding suggests that OsHAK5 is a salt-sensitive high-affinity K+ transporter that imparts enhanced salt tolerance [216]. The K+ to Na+ ratio and salt tolerance are decreased by rice OsHAK21 knockout mutation [217]. ZmHAK4 is selectively expressed in the root stele in maize and involves in removing Na+ from xylem sap. Furthermore, ZmHAK4 exerts specific roles in increasing shoot Na+ exclusion and salt tolerance when used in combination with ZmHKT1 [218]. Song et al. [219] in rice showed that an ER-localized OsCYB5-2, a cytochrome protein can bind to OsHAK21, enhances OsHAK21-mediated K+ absorption and improving salt tolerance. MtHK2/7/12 from Medicago truncatula and MsHAK2/6/7 from Medicago sativa were significantly induced by salt stress [220]. Pepper CaHAK3 and CaHAK7 transcript abundance was strongly and specifically up-regulated in pepper roots under low K+ or high salinity conditions, suggesting the crucial roles for conserving the Na+/ K+ balance during salt stress in pepper [221].
KUP-transport systems under salinity
In several plants varying number of KUPs were identified [222]. Ions notably Cl- and Na+ that accumulate excessively hamper certain transport systems, including the KUP transporters [223]. The abundance of McKUP1 and KUP4 homologous transcripts increased in common ice plants with salt exposure and K+ deficiency [224]. KUP2 transcription is downregulated in the shoots of salt-stressed plants [225]. During salt stress, the expression levels of both KUP6 and KUP11 transcripts in root tissue are upregulated [226]. The KUP7 transporters [227], or depolarization activated K+ channels or transporters like SKOR [228] should mediate the xylem K+ loading when hyperpolarization activated AKT channels are suppressed during the onset of salinity.
4.3.1. HKT-transport systems under salinity
Both K+ starvation and NaCl stress regulated plant HKTs [226]. Many plant HKT transporters mediate influx of Na+, and some are Na+/ K+ symporters [229].
HKTs in wheat, rice, barley, soybean and tomato
Wheat TaHKT2;1 mediates efflux of Na+ in root tissue like rice OsHKT2;1 [230]. Salt and K+ deprivation trigger TaHKT2;1 [229]. Furthermore, HKTs such as TmHKT1;5-A in einkorn wheat [228], HvHKT1;5 in barley [231], SlHKT1;1 and SlHKT1;2 in tomato [232], GmHKT1;4 in soybean [233] and TaHKT1:5-D in bread wheat [234] function diversely and improve plant K+ use efficiency.
An enhanced action of OsHKT1;5 leading to Na+ unloading from the xylem stream was observed [37]. Suppressing rice OsHKT2;1 gene may make plants tolerate salt [235] and salt sensitivity results from root-specific OsHKT1;5 deletion [236]. OsHKT2;4 is likely a K+ transporter rather than a Na+/ K+ symporter and helps maintain a greater K+ level even in salinity [237]. Transcriptional proteins OsSUVH7, OsMYB106, and OsBag4 work together to upregulate OsHKT1;5 in roots during salt stress [238]. Disrupting OsSUVH7 or OsMYB106 binding sites in the OsHKT1;5 promoter or knocking down these genes affects the expression of OsHKT1;5 in roots and ultimately rice salt tolerance [236]. After salt stress, osbhlh044 CRISPR/Cas9 mutants had more Na+ and less K+, resulting in a higher Na+/ K+ ratio [239].
Salinity-induced higher expression of barley HvHKT1;5 and HvHKT2;1 [240,241] and maize ZmHKT2 [242] result in enhanced plant growth and K+ concentration, lowering salt stress by increased Na+/ K+ ratios and ultimately salt tolerance. Transgenic Arabidopsis expressing soybean K+ transporters such as GmHKT1, GmSKOR, GmsSOS1, and GmNHX1 and rice OsHKT2;1 also exhibited higher salt tolerance by higher K+/ Na+ ratios.
4.3. K++exchangers and antiporters under salinity
Plant monovalent cation-proton antiporters (CPAs) are shown to regulate K+ homeostasis and are classified into two types: NHAP/SOS and NHE/NHX transporters (CPA1) and K+ efflux antiporters, KEAs (CPA2). CHXs - cation/H+-exchangers (CHX) are also important. [45,63,243].
4.3.1. CPA1 transporters in transgenic Arabidopsis, sugar beet, rice and cotton with increased salt tolerance
The H+-linked K+ transport at the tonoplast and the K+ distribution between the vacuole and cytosol are processes that are mediated by NHX proteins [244]. AtNHX1-carrying transgenic plants clearly displayed salt tolerance [245]. Transgenic Arabidopsis expressing wheat NHX1 and date palm PdNHX6 displayed a balanced Na+/ K+ ratio and transgenic sugar beet expressing AtNHX3 have been demonstrated to promote salt tolerance [246]. Additionally, increased salt tolerance has been observed by the transgenic expression of the NHX1 gene from one genus in another [247]. When AtNHX1 is overexpressed, plasma membrane localized GhSOS1 expression levels in cotton roots are considerably elevated [248]. It was discovered that NHXs from different plants have varying expression patterns in various plant organs due to salt stress [249]. The expression of OsNHX1, OsNHX2, OsNHX3, and OsNHX5 in rice is triggered by salt stress in different tissues plays a significant role in conferring salinity tolerance [250].
4.4.2. K+ efflux antiporters, KEAs (CPA2) mediate both K+ influx and efflux
Arabidopsis K+ efflux antiporters (KEAs) are comparable to Escherichia coli KefB and KefC. The chloroplasts of a kea1 kea2 double mutant, as well as a kea1 kea2 kea3 triple mutant, are swollen and shorter, which make mutants susceptible to salt stress [251]. Escherichia coli cells expressing KEA1, KEA2, and KEA3 are hypersensitive to even mild salt stress. It is suggested that KEA4-KEA6 may improve salinity tolerance by regulating the pH of the endomembrane network [252]. Overexpression of KEA4, KEA5, or KEA6 in the kea4 kea5 kea6 mutant abolished salt susceptibility. Under salinity, the kea4 kea5 kea6 expressing NHX1 or NHX2 grew better than the triple mutant. Coordinated actions of KEAs and NHXx in specific combinations result in enhanced salt tolerance [67].
4.4.3. CHX transporters in Arabidopsis
Plant CHX transporters mediate transports of K+, Na+, H+, and Cl- [45]. CHX transporters are essential for cytosolic K+ level maintenance and producing K+ signatures during salinity stress [191]. In the cortical cells of adult root zones and epidermal cells, Arabidopsis AtCHX17 was expressed preferentially and is induced by salt stress, indicating that AtCHX17 may also be involved in K+ uptake and homeostasis during salinity. AtCHX17 knockout mutants accumulated less K+ in roots than the wild type [253]. Arabidopsis transgenic lines overexpressing soybean GsCHX19.3 displayed reduced Na+ concentration and higher K+/Na+ ratios during salt and alkaline and produced plant resistance to both stressors [254].
4.4. K+ signaling under salinity
Besides being an essential nutrient, K+ has multiple roles namely ion signaling, enzyme catalysis, water osmoregulation, and stress tolerance [255]. To establish salt resistance mechanisms, plants are compelled to consume the maximum fraction of the available ATP [256]. Salinity-induced transient cytosolic K+ efflux activates a metabolic modification which limits energy-draining anabolic reactions and conserves energy for a strategic advantage under the salinity-induced energy-limiting conditions [171]. Importantly, K+ efflux caused by salt does not compete with the nutrient requirement of plants for K+. The transient nature and substantial tissue specificity of stress-induced K+ efflux finely regulates this [46].
Various plants adopt various patterns of K+ signaling that exhibit commonalities with cytosolic Ca2+ signaling [35]. Transient Ca2+ surges in pants under salinity are processed by a variety of protein kinases and calcium-binding proteins [257]. Activation of the Ca2+ signaling cascade, which includes the CBL-CIPK complex and CDPKs, is one of the key regulatory mechanisms for K+ uptake and transport in plants [258]. Salt tolerance of transgenic rice was improved by the overexpression of bermudagrass CdtCBL4 (calcineurin B-like protein 5) and CdtCIPK5 (CBL-interacting protein kinase 5). Under salt stress, Na+/ K+ homeostasis was regulated by CBL1 in conjunction with CIPK24, CIPK25, and CIPK26 in poplar [259]. Hence, the CBL-CIPK signaling network regulates K+ signaling to increase plant salt tolerance.
An et al. [260] further showed that alfalfa MsCBL4 not only amplified the activities of antioxidant enzymes during salt stress, but also accumulated Ca2+ and decreased the Na+/ K+ ratio in roots, as well as the level of ROS, to overcome the oxidative damage. As a result, crosstalk between Ca2+, K+ and ROS could improve plant salt resistance. By enhancing stomatal conductance and raising the CO2 content in leaf tissue, the accumulation of K+ may restrict the production of ROS during salt stress, lowering lipid peroxidation and promoting tomato growth [261,262].
The transcription and post-translation of plasma membrane H+ ATPase to increase K+ uptake in pumpkin root tips was mediated by RBOH-dependent H2O2 signaling, which improved salt tolerance [39]. The mangrove tree, Kandelia obovate, is better able to adapt to high salinity because nitric oxide (NO) increases its capacity for K+ uptake, lessen the cytosolic K+ loss, and activates AKT1 K+ channel and NHX antiporters to maintain K+/ Na+ equilibrium [263].
Under salinity, a lower Na+/ K+ ratio and increased ROS production in the roots of mung bean, an increase in net K+ and Ca2+ fluxes caused by H2O2 in the roots of barley, a higher K+/ Na+ ratio in the leaves of tomato, a decrease in Na+/ K+ homeostasis and root K+ loss in the leaves and roots of brome grass overexpressing BdCIPK31, and ROS-induced K+ efflux and Ca2+ absorption in barley reflect the cross talk among K+, Ca2+ and ROS signaling cascades which collectively enhance plant salt tolerance [264].
5. cytosolic calcium signaling
Calcium is an important second messenger under both abiotic and biotic stresses, and is also involved in cell division, polarity, growth and development [265]. Salinity causes both osmotic stress and ionic toxicity in plants, which induce various Ca2+ “signatures”, free cytosolic concentration changes, leading to different downstream reactions. Salt-stress activated Ca2+ elevation is connected with an increase of reactive oxygen species, ROS, and ABA concentrations [266,267].
5.1. Calcium transport system
The influx and efflux of calcium are mediated by a complex system of proteins, such as channels, antiporters and ATPase-mediated pumps [268,269]. Calcium transport in a plant cell is shown in Figure 4.
The increase of cytosolic free Ca2+ concentration, [Ca2+] cyt, depends on calcium transport from the apoplast or from internal stores, such as vacuole, ER, or mitochondria [270,271]. Under salinity plants transport calcium into the cytosol by channels or protein transporters, in Arabidopsis cyclic nucleotide-gated channels, AtCNGCs, and glutamate receptor-like receptors, AtGLRs mediate calcium influx [272]. Calcium changes under salt stress also can be due to transport via two-pore channels, TPCs, in the tonoplast, reduced hyperosmolarity-induced [Ca2+]cyt increase channels, OSCAs, which is transporting calcium under osmotic stress, and three types of mechanosensitive-like channels, MSLs [273,274]. Calcium is transported out from the cytosol mainly by pumps or Ca2+/H+ antiporters in an active and secondary active way, respectively [268,269].
5.1.1. Calcium transport by channels.
Calcium is taken up into the cytosol by many different channels: HACCs, hyperpolarization-activated cation channels, DACCs, depolarization -activated cation channels, and other NSCCs, nonselective cation channels, such as CNGCs, cyclic nucleotide gated channels and GLRs, glutamate receptor-like channels, and also MLRs, mechanosensitive-like channels.
Several Ca2+channels also are located in the endomembranes [268]. Ca2+ is transported out from the vacuole by 3 channels: SV, slow-activating vacuolar channel; RyR, cyclic ADP-ribose-activator ryanodine receptor-like channel and InsP3R, inositol 1,4,5-trisphosphate receptor-like channel. Calcium is transported out from the ER by the channels NAADP, InsP3R and RyR, but by FACCs, fast-activating cation channels, into the chloroplast. In the nucleus Ca2+ is transported by NSCCs. Within the mitochondria Ca2+ is moved by an antiporter and a uniporter.
Most investigated of the calcium-influx channels are the CNGC channels (Figure 5). They are regulated by binding of cGMP and cAMP and phosphorylation [269]. Reports show that CNGCs are selective for calcium uptake and are located mainly in PM and tonoplast of root epidermis and leaf mesophyll cells. Experiments showed that some of them are activated when co-expressed with protein kinase CPKs.
5.1.2. Calcium transport from the cytosol and chloroplast and into ER, Golgi and vacuole
Calcium is actively transported from cytosol by the Ca2+ATPase ACA and by secondary active Ca2+/H+ antiporters, CAXs. ACAs also transport Ca2+ into the ER, vacuole, and Golgi, but out from the chloroplast. ECAs, ER-type Ca2+ATPases are only localized in the endomembranes, such as the ER. It pumps Ca2+ into the ER. The Ca2+ATPases are high affinity Ca2+ transporters that shape the Ca2+ concentration to a resting value (50 -100 nM) after a calcium elevation. The Ca2+/H+ antiporters are low affinity Ca2+ transporters that transport higher concentrations of Ca2+.
In the tonoplast, Ca2+/H+ antiporters, the calcium exchangers, CAXs 1-6, with lower affinity for calcium than the Ca2+ATPase, mediate calcium transport [275]. CAX1 and CAX3 are important for keeping calcium homeostasis. It was reported that in Arabidopsis CAX1 is higher expressed in shoot and CAX3 higher expressed in roots. It is proposed that co-expression of CAX1-CAX3 complex is involved in salt tolerance [276].
5.2. Calcium signals depends on type of stress, transporter location and type, and duration of stress.
Measurements of free [Ca2+cyt] in single protoplasts of rice, wheat and beans by use of dual wavelength flourescence microscopy and the calcium indicator Fura2, AM 297, [277,278,279]. show that the rise in calcium usually is lower than in intact plants, such as transgene Arabidopsis [280,281]. This may depend on the lack of cell walls in protoplasts, as cell walls also can be involved in calcium signaling [281]. The extracellular domain of the protein FERONIA is connected with cell-wall pectin and supposed to take part in Ca2+ signaling under salt stress [178]. Excess of ions in the cell walls might cause structure changes that can be sensed by glycoproteins and plasma-membrane-localized receptor kinases and cell wall associated kinases, WAKs. WAKs also can bind both pectin and Ca2+ and excess of Na+ can affect the binding and induce calcium signaling [24]. In addition to the cytosol and apoplast, cell organelles and the endomembrane system also might participate in calcium signaling. In some of the organelles, calcium fluxes are connected with the cytosol, in others the signaling is independent of the cytosol. We will not discuss organellar signaling in this review as it is still scarce information on this topic.
5.2.1. Dynamics of the cytosolic calcium signals
Calcium signals under salinity are usually transient, or sustained, and in the latter case Ca2+cyt can be oscillating or remain for long time [64,271,279]. Other types of abiotic stress show different calcium signals and different channels are involved in the Ca2+cyt increase [49]. Oxygen-deficiency stress induces Ca2+cyt elevation during long time, 10 min or more [282]. Under chilling stress in tomato quite different [Ca2+cyt] signals were monitored depending on the cells’ resting level of calcium under non-stress condition [283]. In Arabidopsis, salt stress in roots induced transient Ca2+ waves through the whole plant with a speed of 400 µm/s [184]. The dynamics of the Ca2+cyt signaling depend on both influx and efflux mechanisms of Ca2+and stress duration.
The calcium signal often depends on the external Ca2+ concentration. In the presence of 0.1 mM external Ca2+ rice cultivars with different salt-tolerance demonstrated a higher increase of [Ca2+cyt] in mesophyll protoplasts of the salt-tolerant cv. Pokkali, than in mesophyll protoplasts of sensitive cv. BRRI Dhan29 [277]. Salt addition in the presence of 1.0. mM Ca2+, did not change [Ca2+cyt] in the tolerant cultivar, but increased it in the sensitive one. Since 1.0 mM Ca2+ is known to block NSCCs, it was concluded that this type of channel was present in cv. Pokkali. Pharmacological experiments showed that the apoplast was the main store for Ca2+ elevation in the sensitive cultivar, but in the tolerant cultivar, Ca2+ was mainly transported from internal stores. Contrary to ionic stress, which causes an increase in [Ca2+cyt], osmotic stress in rice caused a decrease.
5.3. Possible Na+ Sensors
5.3.1. The Na+/H+ antiporter SOS1 is critical for cytosolic calcium elevation, as shown in Arabidopsis, soybean and rice
Few investigations deal with Na+cyt influx into Arabidopsis mesophyll cells. A recent study compared cytosolic Na+ influx in Arabidopsis Wt, sos1;1 and nhx1 mutants and the induced cytosolic calcium changes [64]. By epi-fluorescence microscopy and fluorescent probes, specifically binding to Na+ and Ca2+, SBFI, AM and Fura 2, AM (Figure 2), a higher Na+cyt influx was demonstrated in both mutants compared with Wt when 100 mM NaCl was added. In Wt the influx was low and mainly transient, but in the other genotypes the influx continued for a longer time showing different dynamics. Addition of NaCl-induced Ca2+cyt elevation in the Wt, but not in the sos1;1. A small increase in Ca2+ was also obtained in nhx1. Pretreatment of protoplasts with LiCl before NaCl addition caused a strong inhibition of Ca2+ elevation. The results suggest that the salt-induced calcium elevation depends on influx from both internal and external stores and is obtained in the presence of an intact Na+/H+ antiporter.
In soybean, Glycine max, the GmSOS1 in roots is critical for salt-stress tolerance [221]. The mutant gmsos1, constructed by CRISPR-Cas9 system, accumulated more Na+ in the roots of the soybean than the Wt and increased K+ efflux under salt stress. Moreover, the SOS1 in rice was also considered important for salt tolerance [284].
5.3.2. GIPC can bind Na+
It is still uncertain how plants sense Na+. Recently, it was reported that the monocation-induced [Ca2+] increases1, MOCA1, encoding a glucuronosyl transferase, involved in biosynthesis of a sphingolipid, glycosyl inositol phosphorylceramide, GIPC, was required for calcium signaling under ionic stress. The findings suggest that Na+ can bind to the plasma membrane located GIPC, and that Na+ is transported into the cytosol and activates an unknown calcium channel [11,285] (Figure 6).
5.3.3. A proposed structure and function of SOS1.
SOS1 is a protein with 12 transmembrane domains and is the longest ion transporter in the plant plasma membrane with 1146 amino acids [286]. It is proposed to be important for salt stress tolerance as it can transport Na+ out of the cytosol by its Na+/H+ antiporter. It was recently demonstrated that Na ions can bind to GIPC, a glycosyl inositol phosphoryl ceramide sphingolipid in the plasma membrane [11] The model is described by Xie et al. [286] suggesting that Ca2+ can be transported by the same lipids. In our model (Figure 6) we suggest that Na+ both is bound to the GIPC and transported into the cytosol by this sphingolipid, and that Ca2+ instead is transported via a Ca2+ channel close to GIPC, maybe induced by a membrane potential change or conformational changes in the protein. One reason for this is that our previous results from rice indicated that Na+ is transported into the cytosol before a calcium elevation is induced [287]. In Arabidopsis, the annexin channel AtANN4 was shown to transfer Ca2+under salt stress, but other channels may also be involved. Some reports show that annexins are cytosolic proteins that are associated with PM phospholipids [288,289]. The CNGCs, cyclic nucleotide-gated channel was reported to induce selective calcium transport under salt stress and might also be a candidate channel in the salt-stress signaling [290].
Calcium in sensed by two proteins with similar function in the cytosol, the CBL10/SOS3, a calcineurin B-like protein10, and SCABP8, a calcium-binding protein 8, activated by calcium binding and phosphorylation by binding to the protein kinase SOS2, SOS2/CIPK24 or CIPK8 [286]. The SOS3/SOS2 and SCABP8/SOS2 can transfer SOS2 to the 1136-1138 inhibitory domain of SOS1 protein and release the inhibition of the SOS1. Na+ can then be transported to the apoplast by the Na+/H+ antiport. For easier reading only SOS3, SCABP8 and SOS2 are shown in the Figure 6.
SOS2 can also activate the K, Na+/H+ antiporter NHX in the tonoplast, and CAX, Ca2+/H+ antiporter in PM [291].
5.3.4. ANN1, KEAs and FERONIA and other calcium transporters
Other suggested sensors of high salt that might cause calcium signaling are the plasma membrane localized annexin1, ANN1, plastid-K exchange antiporters, KEAs, and FERONIA (FER), proteins [251,288,292]. At high external Na+ annexin1 mediates a ROS-activated Ca2+ influx into the cytosol. A mutation in ANN1 gene causes an increase of Na+ and diminishes plant growth. KEAs might function as sensors of osmotic stress, but also mechanosensitive-like proteins, MSLs, can react to changes in the turgor pressure, which is connected with osmotic stress. FER belongs to a receptor-like kinase-like (RLKiL) protein family that binds to pectin. Loss of function of this protein results in inhibition of salt-triggered Ca2+ signaling. In plasma membrane of root cells, mechanosensitive-like channels, MSLs, but also protein kinases, calcium channels, e.g., NSCCs, cyclic nucleotide-gated channels, CNGCs, and receptor-like kinases, RLKs, have been suggested to be sensors of salinity.
6. Calcium signal transmission into intracellular downstream reactions
Not only calcium, but also NO, ROS, phophatidic acid and cyclic nucleotides might serve as secondary messengers in stress reactions and make them very complex [292]. Calcium signaling is often connected with pH changes, but also with K+ signaling [293]. Plants respond to stress by activating various signaling pathways that allow them to escape from the stress, or acclimate to it. The specific stress-induced calcium elevation is recognized by different calcium-binding proteins, which transmit the signal into a cascade of reactions that involve protein phosphorylation, protein translocation and gene expression [257,294].
The calcium signals are decoded by many types of sensors, such as calmodulin, CaM, CaM-like proteins, CMLs, calcineurin B-like proteins, CBLs, and calcium-dependent protein kinases, CIPKs [266]. Calcium can bind to a domain on the sensor called EF hand motif, Elongating Factor-hand motif. Sensor proteins can both function as sensor relays and sensor responders. The major calcium sensors are the calcineurin B-like proteins, CBLs. These proteins interact with protein-kinases, CIPKs, to a complex CBL-CIPK network in order to combat the specific type of stress by different mechanisms (Figure 7). Stimuli can rapidly produce an increase in cytosolic Ca2+, [Ca2+]cyt, but also production of ABA, abscisic acid, and ROS, reactive oxygen species. An increase in Ca2+]cyt might also occur from ROS and ABA formation. ABA is necessary for growth and development, and also for surviving salt, drought and cold stresses. At ABA-depended stress, calcium is suggested to be sensed by both CBL1 and CBL9. These proteins bind to CIPK15 and CIPK3, respectively, which activate ABI1/2, ABA-insensitive ½ and ABR1, Abscisic acid repressor1, which are involved in ABA signaling.
7. pH changes and signaling under salinity
It is well known since the nineties, that stress in higher plants induces changes in pHcyt that often occur together with [Ca2+cyt] changes [296,295]. The pH changes in the cytosol of cells or protoplasts can be measured by dual-wavelength fluorescence microscopy and the pH sensitive dye BCECF, AM, an acetoxymethyl ester of carboxyfluorescein, while vacuolar pH can by monitored with confocal microscopy and 6-carboxyfluorescein or Origon Green Dextran dyes [277,279].
7.1. pH signaling under salinity
Okazaki and coworkers [297] demonstrated that NaCl stress triggers fast changes in [Ca2+cyt], pHcyt and vacuolar pH, pHvac in a charophyte alga. The pH changes in rice were connected with Ca2+cyt signaling in a complex way depending on concentration of stressor, duration of stress and sensitivity of species, organ or cell type [57]. In some cases, the pH changes also occur independently of calcium changes. It is expected that protons have a role in signaling as there are steep concentration differences both of calcium and protons within the plant cell. Under normal condition pH is around 7.5 in the cytosol, but in the vacuole lumen and apoplast, pH is much lower, around 5.5 [298]. pH regulation in a cell is depending on the proton pump and antiporter activities, but also other mechanisms are needed. Recent findings suggest that the CCC1, a cation-chloride cotransporter in Arabidopsis is necessary for regulation of pH in in Golgi-network early endosome [299]. If this regulation does not function the plant cannot develop normally, it will get reduced size, chlorophyll defects and stem necrosis and can’t respond to abiotic stress [42].
7.2. Salinity induces different cytosolic and vacuolar pH changes in salt- sensitive and salt-tolerant species or cultivars.
In the salt-tolerant halophytes quince and quinoa and in tolerant rice, salt stress caused an increase in cytosolic pH, but a decrease in vacuolar pH [52,57,58,277]. In the halophyte quinoa pHcyt increased from 7.5 to 8.5 upon salt addition and was then stable at 7.7. No reaction occurred upon addition of salt to protoplasts from salinity-cultivated quinoa, but these protoplasts had a somewhat higher resting pHcyt than the ones cultivated without salinity. In root-epidermis of quinoa, salinity treatment activated the PM H+-ATPase [76].
Since the increase in pHcyt in tolerant rice was inhibited by NH4NO3, an inhibitor of V+ATPase in the tonoplast, is was suggested that protons were transported from cytosol into the vacuole in tolerant plants. This was also confirmed by expression analysis of VHA, which showed an increased expression in rice after salt-stress cultivation [33].
In sensitive plants, like pea, field beans and sensitive rice, the opposite reactions occurred, pHcyt decreased and pHvac increased by salt treatments [58,82,277]. In this case the pHcyt decrease was inhibited by NH4 VO3, an inhibitor of H+ATPase at the plasma membrane and could be caused by activation of the Na+/H+ antiporter SOS1 in the plasma membrane, which is energized by the PM H+ATPase [300,301]. Experiments with leaves from salt-sensitive tobacco and potato plants that were transformed by a gene for a pH-sensitive protein, Pt-GFP, also reacted in the same way: salt treatment caused a decrease of pHcyt, and depended on the time the plants were subjected to stressor [302].
The different reaction in quinoa when salt was added might depend on some kind of acclimation, probably by activation of the H+ATPase, which was absent in pea [56].
Transient pH changes likely occur by Na+/H+ antiporter or H+/Cl- symporter activities. The salinity-induced acidification in pea might result from salt-stress inhibition of the H+-ATPase activity by calcium-activated protein kinases [303], but also from a Cl-/H+ co-transport.
Reports show that halophytes can maintain a high negative membrane-potential difference under salinity, necessary for keeping a stable cell [K+cyt] and to exclude Na+. [69] demonstrated that the high salt-tolerance in halophytes, compared with glycophytes, do not depend on a higher AHA transcript level, but their ability to rapidly activate the PM H+-ATPase upon salt treatment.
7.3. Extra addition of calcium or potassium under cultivation of wheat and field beans in saline medium affects the cellular pH and H+ATPase activity.
Besides being an important second messenger under stress, calcium is necessary for normal growth and development of plants [79]. Potassium is an important nutrient too, since this element is a co-factor for many enzymes and used for osmotic regulation.
Salinity can cause both Ca2+ and K+ deficiencies in plants under cultivation. When the soil K+ concentration is low, a high Na+ level will compete with K+ for the same uptake sites as these ions have a similar chemical structure. A strategy is to add extra CaSO4 and/or K2SO4 to the root media.
An essential premise for plant growth is a low apoplastic pH and depends on an active H+-ATPase that pumps proton from the cytosol into the apoplast [304,305]. A high external pH triggers a cytosolic calcium increase, where a kinase protein, PKS5, is involved in the signaling and regulates the H+ATPase activity [80].
When extra CaSO4 (5 mM) was added to the cultivation medium of wheat cultivars with different salt tolerance, the cytosolic calcium increased in both cultivars and decreased the overall concentration of Na+. On the other hand, the cytosolic pH increased only in the tolerant cultivar and the cytosolic Na+ increased only in the sensitive cultivar. It is likely that both higher cytosolic [Ca2+] and pH reflect tolerance mechanisms [81].
In experiments with field beans cultivated under salinity, both extra K2SO4 (10 mM) and extra CaSO4 (5 mM) caused similar increases of FW and DW [82]. Analysis using Oregon Green dextran dye and ratio imaging of the leaves demonstrated that extra calcium in vivo under cultivation acidified pHapo, but extra potassium had no effect, neither after 7d nor after 21d. After 21 d both extra calcium and extra potassium reduced the cytosolic Na+, but after 7 d only calcium reduced Na+ concentration. These changes were correlated with changes of the PM H+-ATPase hydrolyzing activity. Extra calcium addition induced an activation of the PM H+ATPase that resulted in a higher cytosolic pH.
It was concluded that for wheat and field beans, extra Ca2+ is better than K+ to decrease [Na+cyt], but for cultivation during longer time, 21 d, even extra K+ is advantageous. Potassium treatment in vitro increases the H+-ATPase activity by binding to the phosphorylation domain of the ATPase. This reaction causes a dephosphorylation, which leads to activation of the proton pump [306]. Moreover, both K+ and Ca2+ might bind to pockets at the phosphorylation and nucleotide-binding domains of the H+-ATPase and modulate its activity [307].
Kinetic studies of ATPase activity in field beans showed that addition of extra calcium did not affect the apparent Km, but increased Vmax of the H+-ATPase in a noncompetitive way both after 7 d and 21 d with and without salinity [82]. The increase in Vmax by calcium supply is likely depending on that more enzyme units are expressed. It is well known that calcium has an important role in regulating gene expression and improve transcriptional level of proteins under stress [308]. However, without extra calcium, salinity decreased Vmax both after 7 d and 21 d. Extra potassium supply diminished both the apparent Km and Vmax in controls without salinity. Only after 21 d with salinity K increased Vmax and apparent Km [82]. The competition between potassium and sodium may depend on that protons are used both for Na+/H+ antiport and K++H+ symport. The increase in Km suggests that the affinity between enzyme and substrate of the H+ATPase, MgATP, decreases. Probably less competition exists
These results explain why extra calcium during the cultivation of field bean will improve growth of shoot and leaves. The increase of ATPase activity caused acidification of apoplast, alkalization of the cytosol and a lower Na+cyt level in the leaves.
8. Systemic calcium signaling
8.1. Electrical signals were first proposed in Mimosa pudica
Long time ago it was demonstrated that plants own a system for long-distance transmission of stimuli, such as wounding and other types of stress. When the plant Mimosa pudica was subjected to a drop of cold water, less than 10°C, an active current was obtained that propagated through the stem, petioles, pinnae and pulvini [309]. The rate of the systemic signal was dependent on temperature. It was also demonstrated that wounding triggered a signal that could pass through dead tissue and indicated that the signal was passively transported in the xylem by an apoplastic messenger, called a “Ricca factor” [309,310]. Later research has accentuated a role for electric and hydraulic signals together with calcium and ROS in systemic signaling at wounding ad high-light stresses [311,312].
8.2. Translocation of amino acids by diffusion and bulk flow activates calcium channels
By use of voltage- and ion selective microprobes [313] could show that a mild salt solution (30-50 mM KCl) or amino acids (1 mM glutamic acid or 5 mM GABA) additions to leaves of barley induced action potentials (APs), rapid voltage changes, that propagated basipetally and acropetally from leaf to leaf and from root to leaf. Salt additions caused biphasic voltage changes at a speed 20-30 cm/min, starting with a strong depolarization causing calcium influx, and then anion efflux. The calcium-influx was reflected by a small depolarization, but the anion efflux caused a voltage break-through. A K+ -efflux started after the depolarizing voltage had passed the K+ equilibrium potential.
Amino acid addition did not induce APs through depolarization, instead it was suggested that APs was caused by binding the amino acids to receptors or ligand-gated Ca2+-conducting channels. Thereafter, calcium activated anion efflux.
Recent results show that long-distance calcium waves induced at wounding and salt stresses, are mediated by diffusion and bulk flow of the amino-acid messenger glutamate [314]. The authors propose that stress triggers a release of amino acids that diffuse in the apoplast from the local-stimuli perception to a distal tissue. During their translocation they activate glutamate receptor-like channels GLR3.3 and GLR3.6, which enable calcium transport by bulk flow in the vascular tissue. Analysis made by quantitative imaging of wild type and mutants expressing calcium and glutamate fluorescent tracers shows that salinity and several other stimuli might induce calcium waves with similar dynamics. In this case the APs were connected with transient apoplastic pH increases (1 unit) and cytoplasmic pH decrease (0.5 units).
8.3. Electrical signals and glutamate may be involved
An investigation on systemic signaling in Arabidopsis triggered by wounding also indicates the involvement of electrical signals and calcium [315]. The signals were dependent on glutamate and pH-regulated Ca2+ channels. Both the glutamate receptor-like proteins, GLR3.3 and GLR3.6, had a role in the signal transmission from leaf to leaf. The same proteins were involved in Ca2+ and electrical signaling from root to shoot, but the process required an inactivated proton pump AHA1. The later systemic signaling also was triggered by glutamate in the absence of stress. Moreover, the finding showed that a leakage of glutamate from phloem into the apoplast occurred, as well as an increase in apoplastic pH, leading to an activation of the GLRs and a systemic signaling.
8.4. Ca2+-ROS interactions
Propagation of stress-induced signals in barley leaves were proposed to be mediated by different regulators, such as by changes in membrane potentials and ion fluxes [313] and by involvement of reactive oxygen species, ROS, as suggested [316]. Choi et al. [184] reported that salt stress induced systemic cytosolic-calcium waves that were propagated within the root cortical and endodermal cells at a speed of approximately 400 µM/s.
Evans and co-workers [317] emphasized that salt stress triggers calcium waves propagating through the whole plant, and are assisted by ROS. From experiments with Arabidopsis, the authors proposed that calcium was transported from the vacuole into cytosol by two-pore channels, AtTPC1s. The plasma membrane NADPH oxidase, AtRBOHD can be activated by cytosolic Ca2+ increase. The authors proposed a self-propagating calcium -- > ROS --> calcium model, as the salt-induced cytosolic calcium increase can activate the AtRBOHD to release ROS by phosphorylation and calcium binding to its E- hand motifs, and that ROS then can trigger Ca release into the cytosol by ROS-regulated plasma membrane channels. This model is plausible, since inhibition of AtRBOHD, either by the by ascorbate or diphenyliodonium, inhibition of calcium channels by La or Ruthenium Red, or by knockout of the TPC1 gene, very much reduced the wave transmission speed that was obtained under the control measurements. As expected, overexpression of the gene led to an increased speed.
9. Conclusion
We have shown that salt-tolerant and sensitive species or cultivars exhibit different mechanisms for influx and efflux of Na+, K+, Cl- and pH changes under salt stress. Most of the important crops like wheat, rice, barley and beans are sensitive to salinity. Halophytes and other salt-tolerant plants display a rapid transient uptake of Na+ into the cytosol, to prevent toxic Na+, while sensitive plants take up and retain Na+ for a longer time in the cytosol [56]. The SOS1 antiporter in the plasma membrane is important for salt tolerance as it transports Na+ from the cytosol into the apoplast or out from the cell. Recent findings suggest that CCX1, a calcium cation exchanger in the plasma membrane, also is important for salt tolerance [71]. The Na+, K+/H+ antiporter NHX in the tonoplast transports both Na+ and K+ into the vacuole and is important for pH regulation [68]. The efflux way for Na+ from the cytosol depends on species and H+ATPase activities at each membrane. Some plants can prevent long-distance Na+ transport from root to shoot as high Na+ is very harmful for photosynthesis and this transport can be regulated at the xylem/parenchyma cell border, where Na+ either is transported into xylem for further transport by the transpiration stream or is recirculated into the root.
Calcium is established as an important second messenger for abiotic and biotic stresses, and for growth and development [258,291]. Calcium changes are often connected with pH changes, and pH is considered an important secondary messenger under salt stress too. Potassium takes part in signaling together with calcium in the CBL-CIPK network that leads to ion and pH homeostasis and ROS signaling [266]. Salt stress induces a cytosolic Ca2+increase, which is sensed by the SOS system, Salt Overly System, and signals are transmitted to the CBL/CIPK system [286,318] (Figure 7).
Since long time researchers have tried to find the sensor for salt, probably localized either in the plasma membrane or in the cytosol. Some researchers suggested that the long C-terminal tail of the SOS1 protein could be the sensor [57,318]. It was recently proposed that the PM sphingolipid GIPC acts as a Na+ sensor, and that the Na-GIPC complex might activate an unknown calcium channel [11]. However, in the model suggested by [286], Na+ is bound to GIPC and calcium is transported by the same lipid. In the Figure 6, we instead suggest that Na+ both binds to GIPC and is transported by this lipid, and that another channel transports calcium across the membrane. The reason for this is that an earlier investigation with salt-tolerant rice, shows that Na+ enters the cytosol before a calcium elevation is induced [287].
Research on calcium signaling has focused on cellular cytosolic signaling, but calcium signals are traveling within the whole plants. A rapid communication between cell to cell might occur by ROS producing the NADPH oxidase RBOHD, by the two-pore ion channel, TPC1, or by glutamate receptor-like channels GLR3.3 and GLR3.6 [311]. Recent results from [314] suggest that glutamate is a messenger involved in systemic-calcium signaling in plants. Translocation by glutamate, and other amino acids, are supposed to activate glutamate-receptor-like channels for calcium, causing calcium waves from the local stimulus to distal tissues and cells under salinity and wounding.
The new results concerning systemic-calcium signaling with glutamate as a messenger, and that GIPC might be a sensor for salt are interesting and have improved our knowledge on salt-stress signaling. However, there is a gap in the knowledge between local cytosolic signals and systemic signaling waves in the whole plant. There are still many questions to answer. Is GIPC the only sensor of Na+, or are many different sensors working? Little is known about the role of glutamate and its receptor proteins in different cells, tissue and organs and if the calcium waves show the same dynamics within the whole plant. How the systemic calcium waves are connected with organellar signals are still enigmatic and needs further investigations.
Author Contributions
The authors contributed equally to this work. S.L and A.P wrote and edited the text and approved the submitted version.
Funding
No funding was received.
Data available
Not applicable
Conflict of interest
The authors declare no conflict of interest.
References
- Munns, R.; Gilliham, M. Salinity tolerance of crops – What is the cost? New Phytol. 2015, 208, 668-673. [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025-1043. [CrossRef]
- Maathuis, F.J.M. Sodium in plants: perception, signaling and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849-858. [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12.667458. Sec. Plant Abiotic Stress. [CrossRef]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250-258. [CrossRef]
- Colmeneros-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peindo-Torrubia, P.; Rosales, M.A. Chloride as a beneficial macronutrient in higher plants: New Roles and regulation. Inter. J. of Mol. Sci. 2019, 20, 4686. [CrossRef]
- Wu, H.; Li, Z. Vacuolar Cl- sequestration: Revisiting the role of Cl- transport in plant salt tolerance. Front. Plant Sci. 2019, 10, 1418. [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentration of Na+ and Cl- ions in soil solution has simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449-4459. [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66-71. [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integrat. Plant Biol. 2018, 60, 796-804. [CrossRef]
- Jiang, Z.; Zhou, X.; Pei, Z-M. Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx. Nature 2019, 572, 341-346. [CrossRef]
- Lu, Y.; Fricke, W. Changes in root hydraulic conductivity in wheat (Triticum aestivum L.) in response to salt stress and day/night can best be explained through altered activity of aquaporins. Plant Cell Environ. 2023, 46, 747-763. [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Res. 2015, 95, 1321-1358. [CrossRef]
- Flowers, T.; Yeo, A. The driving force for water and solute movement. In Yeo, A. and Flowers, T. Eds. Plant Solute Transport, 1992, pp. 29-45.
- Kronzucker, H.J.; Britto, D.T. Sodium transport in plants: a critical review. New Phytol. 2011, 189, 54-81. [CrossRef]
- Shi, Y.; Wang, Y.; Flowers, T.J.; Gong, H. Silicon decreases chloride transport in rice (Oryza sativa L.). J. Plant Physiol. 2013, 170, 847-853. [CrossRef]
- Amtmann, A.; Sanders, D. Mechanisms of Na+ uptake by plant roots. Adv. Bot. Res. 1999, 29, 75-112. [CrossRef]
- Zhang, J.L.; Flowers, T.J.;Wang, S.M. Mechanisms of sodium uptake by roots of higher plants. Plant Soil 2010, 326, 45-60. [CrossRef]
- Haro, R.; Banuelos, M.A.; Rodriguez-Navarro, A. High affinity uptake of sodium in land plants. Plant Cell Physiol. 2010, 51, 68-79. [CrossRef]
- Davenport, R.J.; Tester, M. A weakly voltage-dependent, nonselective cation channel mediates toxic sodium influx in wheat. Plant Physiol. 2000, 12, 823-834. [CrossRef]
- Demidschik, V.; Maathuis, F.J.M. Physiological roles of nonselective cation channels in plants: from salt stress to signaling and development. New Phytol. 2007, 175, 387-404. [CrossRef]
- Leng, Q.; Mercier, R.W.; Hua, B.; Fromm, H.; Berkowitz, G.A. Electrophysiological analysis of cloned cyclic nucleotide-gated ion channels. Plant Physiol. 2002, 128, 400-410. [CrossRef]
- Keisham, M.; Mukherjee, S.; Bhatla, S. Mechanisms of sodium transport in plants - progresses and challenges. Int. J. Mol. Sci. 2018, 19, 647. [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanism of plant responses and adaptation to soil salinity. Inno. 2020, 1, 100017. [CrossRef]
- Golldack, D.; Su, H.; Quigley, F.; Kamasani, U.R.; Munoz-Garay, C.; Balderas, E.; Popova, O.V.; Bennett, J.; Hans J. Bohnert, H.J.; Pantoja, O. Characterization of an HKT-type transporter in rice as a general alkali cation transporter. Plant J. 2002, 31, 529-542. [CrossRef]
- Tester, M.; Davenport, R.J. Na+ transport and Na+ tolerance in higher plants. Ann. Bot. 2003, 91, 503-527. [CrossRef]
- Greger, M.; Ahmad H. Kabir, A.H.; Landberg, T.; Maity, P.J.; Lindberg, S. Silicate reduces cadmium uptake into cells of wheat. Environ. Poll. 2016, 211, 90-97. [CrossRef]
- Mian, A.; Oomen, R.J.; Isayenkov, S.; Sentenac, H.; Maathuis, F.J.; Véry, A.A. Over-expression of a Na+ and K+ permeable HKT transporter in barley improves salt tolerance. Plant J. 2011, 68, 468-479. [CrossRef]
- Han, Y.; Yin, S.; Huang, l.; Wu, X.; Zeng, J.; Liu, X.; Qiu, L.; Munns, R.; Chen, Z-H.; Zhang, G. A sodium transporter HvHKT1 confers salt tolerance in barley via regulating tissue cell ion homeostasis. Plant Cell Physiol. 2018, 59, 1976-1989. [CrossRef]
- Essah, P.A.; Davenport, R.; Tester, M. Sodium influx and accumulation in Arabidopsis. Plant Physiol. 2003, 133, 307-318. [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [CrossRef]
- Byrt, C.S.; Zhao, M.; Koughi, M.; Bose, J.; Henderson, S.W.; Qiu, J.; et al. Non-selective cation channel activity of aquaporin AtPIP2;1 regulated by Ca2+ and pH. Plant Cell Environ. 2017, 40, 802-815. [CrossRef]
- Kader, Md.A.; Seidel, T.; Golldack, D.; Lindberg, S. Expressions of OsHKT1, OsHKT2 and OsVHA are differently regulated under NaCl stress in salt-sensitive and salt-tolerant rice (Oryza sativa L.) cultivars. J. Exp. Bot. 2006, 57, 4257-4268. [CrossRef]
- Horie, T.; Yoshida, K.; Nakayama, H.; Yamada, K.; Oiki, S.; Shinmyo, A. Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa. Plant J. 2001, 27, 129-138. [CrossRef]
- Rubio, F.; Nieves-Cordones, M.; Horie, T.; Shabala, S. Doing ‘business as usual’ comes with a cost: evaluating energy cost of maintaining plant intracellular K+ homeostasis under saline conditions. New Phytol. 2019, 225, 1097-1104. [CrossRef]
- Berthomieu, P.; Cone, G.; Nublat, A.; Brackenbury, W.J.; Lambert, C.; Savio, C.; Uozumi, N.; Oiki, S.; Yamada, K.; Cellier, F.; et al. Functional analysis of AtHKT1 in Arabidopsis shows that Na recirculation by the phloem is crucial for salt tolerance. EMBO J. 2003, 22, 2004-2014. [CrossRef]
- Ren, H.; Gao, J.P.; Li, L.G. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Gen. 2005, 37, 1141-1146. [CrossRef]
- Brini, F.; Masmoudi, K. Ion transporters and abiotic stress tolerant plants. ISRN Mol. Biol. 2012, 927436. [CrossRef]
- Huang, L.; Kuang, L.; Wu, L.; Wu, D.; Zhang, G. Comparison in functions of HKT1;5 transporters between Hordeum marinum and Hordeum vulgare in responses to salt stress. Plant Growth Reg. 2019a, 89, 309-319. [CrossRef]
- Møller, I.S.; Gilliham, M.; Jha, D.; Mayo, G.M.; Roy, S.J.; Coates, J.C.; Haseloff, J.; Tester, M. Shoot Na+ exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na+ transport in Arabidopsis. Plant Cell 2009, 21, 2163-2178. [CrossRef]
- Davenport, R.; Munoz-Mayer, A.; Jha, D., Essah, P.A.; Rus, A.; Tester, M. The Na+ transporter AtHKT1;1 controls retrieval of Na+ from the xylem in Arabidopsis. Plant Cell Environ. 2007, 30, 497-507. [CrossRef]
- Colmeneros-Flores, J.M.; Martinez, G.; Gamba, G.; Vazquez, N.; Iglesias, D.J.; Brumos, J.; Talon, M. Identification and functional characterization of cation-chloride cotransporters in plants. Plant J. 2007, 50, 278-292. [CrossRef]
- Henderson, S.W.; Wege, S.; Gilliham, M. Plant cation-chloride cotransporters (CCC): Evolutionary origin and functional insights. Int. J. Mol. Sci. 2018, 19, 492. [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651-681. [CrossRef]
- Isayenkov, S.V.; Dabravolski, S.A.; Pan, T.; Shabala, S. Phylogenetic diversity and physiological roles of plant monovalent cation/H+ antiporters. Front. Plant Sci. 2020, 11, 573564. [CrossRef]
- Shabala, S.; Demidchik, V.; Shabala, L.; Cuin, T.A.; Smith, S.J.; Miller, A.J.; Davies, J.M.; Newman, I.A. Extracellular Ca2+ ameliorates NaCl induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 2006, 141, 1653-1665. [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2014, 115, 419-431. [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008. 179, 945-963. [CrossRef]
- Wang, C.M.; Zhang, J.L.; Liu, X.S.; Li, Z.; Wu, G.Q.; Cai, J.Y.; Wang, S.M. Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+. Plant Cell Environ. 2009, 32, 486-496. [CrossRef]
- Schachtman, D.P.; Schroeder, J.I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants. Nature 1994, 370, 655-658. [CrossRef]
- Schachtman, D.P.; Liu, W.H. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends Plant Sci. 1999, 4, 281-287. [CrossRef]
- Wang, S.M.; Zhang, J.L.; Flowers, T.J. Low-affinity Na+ uptake in the halophyte Suaeda maritima. Plant Physiol. 2007, 145, 559-571. [CrossRef]
- Zepeda-Jazo, I.; Shabala, S.; Chen, Z.; Pottosin, I.I. Na+-K+ transport. in roots under salt stress. Plant Signal. Behav. 2008, 3, 401-403. [CrossRef]
- Roberts, S.K.; Tester, M. Permeation of Ca2+ and monovalent cations through an outwardly rectifying channel in maize root stelar cells. J Exp. Bot. 1997, 48, 839-846. [CrossRef]
- Adem, G.D.; Chen, G.; Shabala, L.; Chen, Z.H. GORK channel: A master switch of plant metabolism. Trends Plant Sci. 2020, 25, 434- 445. [CrossRef]
- Sun, Y.; Lindberg, S.; Shabala, L.; Morgan, S.; Shabala, S.; Jacobsen, S. E. A comparative analysis of cytosolic Na+ changes under salinity between halophyte quinoa (Chenopodium quinoa) and glycophyte pea (Pisum sativum). Environ. Exp. Bot. 2017, 141, 154-160. [CrossRef]
- Shabala, S.; Shabala, L.; Cuin, T.A.; Pang, J.; Percey, W.; Chen, Z.; Wegner, L.H. Xylem ionic relations and salinity tolerance in barley. Plant J. 2010, 61, 839-853. [CrossRef]
- Shabala, S. (2017). Signaling by potassium: another second messenger to add to the list? J. Exp. Bot. 2017, 68, 4003-4007. [CrossRef]
- Shabala, S.; Hariadi, Y.; Jacobsen, S.E.; Genotypic difference in salinity tolerance in quinoa is determined by differential control of xylem Na+ loading and stomatal density. J. Plant Physiol. 2013, 170, 906-914. [http://dx.doi.org/10.1016/j.jplph.2013.01. 014.
- Jia, W.; Wang, Y.; Zhang, S.; Zhang, J. Salt-stress-induced ABA accumulation is more sensitively triggered in roots than in shoots. J. Exp. Bot. 2002, 53, 2201-2206. [http://dx.doi.org/10.1093/jxb/erf079.
- Shi, H.Z.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 2002, 14, 465-477. [http://dx.doi.org/10.1105/tpc.010371.
- Zhu, M.; Zhou, M.; Shabala, L.; Shabala, S. Physiological and molecular mechanisms mediating xylem Na+ loading in the context of salinity stress tolerance. Plant Cell Environ. 2016, 1-12. [http://dx.doi.org/10.1111/pce.12727.
- Huang, L.; Kuang, L.; Li, X.; Wu, L.;Wu, D.; Zhang, G. (2018). Metabolomics and transcriptomic analyses reveal the reason why Hordeum marinum has higher salt tolerance than Hordeum vulgare. Env. Exp. Bot. 2018, 156, 48-61. [CrossRef]
- Ngoc, N.T.; Tri, P.N.; Le Hong, T.; Quoc, C.D. Biomolecular evaluation of three contrasting rice cultivars (Oryza sativa L.) in salt stress response at seedling stage. Plant Sci. Today 2022, 9, 491-503. [CrossRef]
- Foster, K.J.; Miklavcic, S.J. A comprehensive biophysical model of ion and water transport in plant roots. II. Clarifying the rules of SOS1 in the salt-stress response Arabibopsis. Front. Plant Sci. 2019, 10, 1121. [CrossRef]
- Halperin, S.J.; Lynch, J.P. Effect of salinity on cytosolic Na+ and K+ in root hairs of Arabidopsis thaliana: in vivo measurement using the fluorescent dyes SBFI and PBFI. J. Exp. Bot. 2003, 54, 2035-2043. [CrossRef]
- Morgan, S.H.; Kader, Md.A.; Lindberg, S. Cytosolic sodium influx in mesophyll protoplasts of Arabidopsis thaliana, Wt, sos1;1, and nhx1 differs and induces different calcium changes. Plants 2022, 11, 3439. [CrossRef]
- Bassil, E.B.; Zhang, S.; Gong, H.; Tajima, H.; Blumwald, E. Cation specificity of vacuolar NHX type Cation/H+ antiporters. Plant Physiol. 2019, 179, 616-629. [CrossRef]
- Chen, Z.; Wu, Y.; Di, L.; Shen, Y.; Wang, G. AtCCX1 transports Na+ and K+ in Pitch pastoris. Afr. J. Biotechnol. 2011, 10, 9743-9750. [DOI: 10.5897/AJB11.455.
- Zhu, X.; Pan, T.; Zhang, X.; Fan, L.; Quintero, F.J.; Zhao, H.; Su, X.; Li, X.; Villalta, I.; Mendoza ,I.; et al. (2018). K+ efflux antiporters 4, 5, and 6 mediate pH and K+ homeostasis in endomembrane compartments. Plant Physiol. 2018, 178, 1657-1678. [CrossRef]
- Yang, J.; Li, W.; Guo, X.; Chen, P.; Cheng, Y.; Mao, K.; Ma, F. Cation/Ca2+ exchanger1 (MdCCX1), a plasma membrane-localized Na+ transporter, enhances plant salt tolerance by inhibiting excessive accumulation of Na+ and reactive oxygen species. Front. Plant Sci. 2021, 12, 746189. [CrossRef]
- D'Onofrio, C.; Kader, A.; Lindberg, S. Uptake of sodium in quince, wheat and sugar beet protoplasts determined by the fluorescent sodium-binding benzofuran isophthalate dye. J. Plant Physiol. 2005, 162, 421-428. [CrossRef]
- Kader, A.; Lindberg, S. Uptake of sodium in protoplasts of salt-sensitive and salt-tolerant cultivars of rice, Oryza sativa L. J. Exp. Bot. 2005, 56, 3149-3158. [CrossRef]
- Negulescu, P.A.; Harootunian, A.; Tsien, R.Y.; and Machen, T.E. Fluorescence measurement of cytosolic free Na concentration influx and efflux in gastri cells. Cell Regul. 1990, 1, 259-268. [CrossRef]
- Demidschik, V.; Tester, M. Sodium fluxes through nonselective cation channels in plasma membrane of protoplasts from Arabidopsis roots. Plant Physiol. 2002, 128, 379-387. [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Lai, D.; Xie, Y.; Shen, W.; Shabala, S. Rapid regulation of plasma membrane H+ATPase activity is essential to salinity tolerance in two halophyte species, Atriplex lentiformis and Chenopodium quinoa. Ann. Bot. 2015, 115, 481-494. [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F., Jacobsen, S.E. Quinoa − a model crop for understanding salt tolerance mechanisms in halophytes. Plant Biosyst. 2016, 150, 357-371. [http://dx.doi.org/10.1080/11263504.2015.1027317.
- Anil, V.S.; Krishnamurthy, H.; Mathew, M. Limiting cytosolic Na confers salt tolerance in rice cell in culture: a two-photon microscopy study of SBFI-loaded cells. Physiol. Plant. 2007, 129, 607-621. [CrossRef]
- Blumwald, E.; Poole, R.J. The Na+/H+ antiport in isolated tonoplast vesicles from storage tissue of Beta vulgaris. Plant Physiol. 1985, 78, 163-167. [CrossRef]
- Hassidim, M.; Braun, Y.; Lerner, H.R.; Reinhold, L. Na+/H+ and K+/H+ antiport in root membrane vesicles isolated from the halophyte Atriplex and the glycophyte cotton. Plant Physiol. 1990, 94, 1795-1801. [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441-445. [CrossRef]
- Perez-Hormaeche, J.; Villalta, I.; Espartero, J.M.; Gámez-Arjona, F.M. A critical role of sodium flux via the plasma membrane Na+/H+ Exchanger SOS1 in the salt tolerance of rice. Plant Physiol. 2018, 180, 1046-1065. [CrossRef]
- Sze, H.; Schumacher, K.; Muller, M.; Padmanaban, S.; Taiz, L. A simple nomenclature for a complex proton pump: VHA genes encode the vacuolar H+ATPase. Trends Plant Sci. 2002, 7, 157-161. [CrossRef]
- Teakle, N.L.; Tyerman, S.D. Mechanisms of Cl- transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566-589. [CrossRef]
- Lorenzen, I.; Aberle, T.; Plieth, C. Salt stress-induced chloride flux: a study using transgenic Arabidopsis expressing a fluorescent anion probe. Plant J. 2004, 38, 539-544. [CrossRef]
- Geilfus, C.M. Review on the significance of chlorine for crop yield and quality. Plant Sci. 2018, 270, 114-122. [CrossRef]
- Cao, Y.; Zhou, X.; Song, H.; Zhang, M.; Jiang, C. Advances in deciphering salt tolerance mechanism in maize. Crop J. 2023, 11, 1001-1010. [CrossRef]
- Rajendran, K.; Tester, M.; Roy, S.J. Quantifying the three main components of salinity tolerance in cereals. Plant Cell Environ. 2009, 32, 237-249. [CrossRef]
- Guan, Z.; Chen, S.; Chen, F.; Liu, Z.; Fang, W.; Tang, J. Comparison of stress effect of NaCl, Na+ and Cl- on two Chrysanthemum species. Acta Hortic. 2012, 937, 369-375. [CrossRef]
- Abdelgadir, E. M.; Oka, M.; Fujiyama, H. Characteristics of nitrate uptake by plants under salinity. J. Plant Nutr. 2005, 28, 33-46. [CrossRef]
- Hu, Y.; Schmidhalter, U. Spatial distribution sand net deposition rates of mineral elements in the elongating wheat (Triticum aestivum L.) leaf under saline soil conditions. Planta 1998, 204, 212-219. [CrossRef]
- Massa, D.; Mattson, N. S.; Lieth, H. J. Effects of saline root environment (NaCl) on nitrate and potassium uptake kinetics for rose plants: a Michaelis–Menten modelling approach. Plant Soil 2009, 318, 101-115. [CrossRef]
- Grewal, H. S. Water uptake, water use efficiency, plant growth and ionic balance of wheat, barley, canola and chickpea plants on a sodic vertosol with variable subsoil NaCl salinity. Agric. Water Manag. 2010, 97, 148-156. [CrossRef]
- Tregeagle, J.M.; Tisdall, J.M.; Tester, M.; Walker, R.R. Cl- uptake, transport and accumulation in grapevine rootstocks of differing capacity for Cl- exclusion. Funct. Plant Biol. 2010, 37, 665-673. [CrossRef]
- Yamashita, K.; Kasai, M.; Yamamoto, Y.; Matsumoto, H. Stimulation of plasma membrane H+-transport activity in barley roots by salt stress. Possible role of increase in chloride permeability. Soil Sci. Plant Nutr. 1994, 40, 555-563. [CrossRef]
- Skerrett, M.; Tyerman, S.D. A channel that allows inwardly-directed fluxes of anions in protoplasts derived from wheat roots. Planta. 1994, 192, 295-305. [CrossRef]
- Wegner, L.H.; Raschke, K. Ion channels in the xylem parenchyma of barley roots. Plant Physiol. 1994, 105, 799-813. [CrossRef]
- White, P.J. Separation of K+ and Cl−-selective ion channels from rye roots on a continuous sucrose density gradient. J. Exp. Bot. 1995, 46, 361-376. [CrossRef]
- Ryan, P.R.; Skerrett, M.; Findlay, G.P.; Tyerman, S.D. Aluminum activates an anion channel in the apical cells of wheat roots. Proc. Natl. Acad. Sci. USA 1997, 94, 6547-6552. [CrossRef]
- Kiegle, E.; Gilliham, M.; Haseloff, J.; Tester, M. Hyperpolarisation-activated calcium currents found only in cells from the elongation zone of Arabidopsis thaliana roots. Plant J. 2000, 21, 225-229. [CrossRef]
- Diatloff, E.; Roberts, M.; Sanders, D.; Roberts, S.K. Characterization of anion channels in the plasma membrane of Arabidopsis epidermal root cells and the identification of a citrate permeable channel induced by phosphate starvation. Plant Physiol. 2004, 136, 4136-4149. [CrossRef]
- Ouerghi, Z.; Cornic, G.; Roudani, M.; Ayadi, A.; Brulfert, J. Effect of NaCI on photosynthesis of two wheat species (Triticum durum and T. aestivum) differing in their sensitivity to salt stress. Plant Physiol. 2000, 156, 335-340. [CrossRef]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil, 2003, 253, 201-218. [http://dx.doi.org/10.1023/A:1024553303144.
- Skerrett, M.; Tyerman, S.D. (1994). A channel that allows inwardly directed fluxes of anions in protoplasts derived from wheat roots. Planta 1994, 192, 295-305. [CrossRef]
- Tyerman, S.D.; Skerrett, M.; Garrill, A.; Findlay, G.P.; Leigh, R.A. Pathways for the permeation of Na+ and Cl− into protoplasts derived from the cortex of wheat roots. J. Exp. Bot. 1997, 48, 459-480. [CrossRef]
- Diédhiou, C.J.; Golldack, D. Salt-dependent regulation of chloride channel transcripts in rice. Plant Sci. 2006, 170, 793-800. [CrossRef]
- Nakamura, A.; Fukuda, A.; Sakai, S.; Tanaka, Y. Molecular cloning, functional expression and subcellular localization of two putative vacuolar voltage-gated chloride channels in rice (Oryza sativa L.). Plant Cell Physiol. 2006, 47, 32- 42. [CrossRef]
- Wei, P.; Che, B.; Shen, L.; Cui, Y.; Wu, S.; Cheng, C.; Liu, F.; Li, M.A.; Bingjun Yu, B.; Lam, H.M. Identification and functional characterization of the chloride channel gene, GsCLC-c2 from wild soybean. BMC Plant Biol. 2019, 19, 121. [CrossRef]
- Nguyen, C.T.; Agorio, A.; Jossier, M.; Depré, S.; Thomine, S.; Filleur, S. Characterization of the chloride channel-like, AtCLCg, involved in chloride tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 764-775. [CrossRef]
- Liu, W.; Feng, J.; Ma, W.; Zhou, Y.; Ma, Z. GhCLCg-1, a vacuolar chloride channel, contributes to salt tolerance by regulating ion accumulation in upland cotton. Front. Plant Sci. 2021, 12, 765173. [CrossRef]
- Jossier, M.; Kroniewicz, L.; Dalmas, F.; Le Thiec, D.; Ephritikhine, G.; Thomine, S.; Barbier-Brygoo, H.; Vavasseur, A.; Filleur, S.; Leonhardt, N. The Arabidopsis vacuolar anion transporter, AtCLCc, is involved in the regulation of stomatal movements and contributes to salt tolerance. Plant J. 2010, 64, 563-576. [CrossRef]
- Kohler, B.; Raschke, K. The delivery of salts to the xylem. Three types of anion conductance in the plasmalemma of the xylem parenchyma of roots of barley. Plant Physiol. 2000, 122, 243-254. [CrossRef]
- Gilliham, M.; Tester, M. The regulation of anion loading to the maize root xylem. Plant Physiol. 2005, 137, 819-828. [CrossRef]
- Roberts, S.K. Plasma membrane anion channels in higher plants and their putative functions in roots. New Phytol. 2006, 169, 647-66. [CrossRef]
- Hedrich, R.; Becker, D. Green circuits - The potential of plant specific ion channels. Plant Mol. Biol. 1994, 26, 1637-1650. [CrossRef]
- Dietrich, P.; Hedrich, R. Anions permeate and gate GCAC1, a voltage-dependent guard cell anion channel. 1998, Plant J. 15, 479-487. [CrossRef]
- Wu, H.; Li, Z. The importance of Cl- exclusion and vacuolar Cl- sequestration: Revisiting the role of Cl- transport in plant salt tolerance. Front. Plant Sci. 2019, 10, 418. [CrossRef]
- Negi, J.; Matsuda, O.; Nagasawa, T.; Oba, Y.; Takahashi, H.; Kawai-Yamada, M.; Uchimiya, H.; Hashimoto, M.; Iba, K. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature 2008, 452, 483-486. [CrossRef]
- Vahisalu, T.; Kollist, H.; Wang, Y.F.; Nishimura, N.; Chan, W.Y.; Valerio, G.; amminmäki, A.; Brosché, M.; Moldau, H.; Desikan, R.; Schroeder, J.I.; Kangasjärvi, J. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature 2008, 452, 487-491. [CrossRef]
- Planes, M.D.; Niñoles, R.; Rubio, L.; Bissoli, G.; Bueso, E; García-Sánchez, M.J.; Alejandro, S.; Gonzalez-Guzmán, M.; Hedrich, R.; Rodriguez, P.L.; et al. A mechanism of growth inhibition by abscisic acid in germinating seeds of Arabidopsis thaliana based on inhibition of plasma membrane H+-ATPase and decreased cytosolic pH, K+, and anions. J. Exp. Bot. 2015, 66, 813-825. [CrossRef]
- Cubero-Font, P.; Maierhofer, T.; Jaslan, J.; Rosales, M.A.; Espartero, J.; Díaz-Rueda, P., Müller, H.M.; Hürter, A.L.; Al-Rasheid, K.A.; Marten, I.; et al. Silent S-type anion channel subunit SLAH1 gates SLAH3 open for chloride root-to-shoot translocation. Curr. Biol. 2016, 26, 2213-2220. [CrossRef]
- Flowers, T.J.; Hajibagheri, M.A. Salinity tolerance in Hordeum vulgare: ion concentrations in root cells of cultivars differing in salt tolerance. Plant Soil 2001, 231, 1-9. [CrossRef]
- Brumos, J.; Talon, M.; Bouhal, R.; Colmenero-Flores, J.M. Cl- homeostasis in includer and excluder citrus rootstocks: transport mechanisms and identification of candidate genes. Plant Cell Environ. 2010, 33, 2012-2027. [CrossRef]
- Boursier, P.; Läuchli, A. Mechanisms of chloride partitioning in the leaves of salt-stressed Sorghum bicolor L. Physiol. Plant. 1989,77, 537-544. [CrossRef]
- Thomine, S.; Zimmermann, S.; Guern, J.; Barbier-Brygoo, H. ATP-dependent regulation of an anion channel at the plasma membrane of epidermal cells of Arabidopsis hypocotyls. Plant Cell 1995, 7, 2091-2100. [CrossRef]
- Allen, G.J.; Sanders, D. The Plant Vacuole. In: Advances in Botanical Research. Vol. 25, Edited by Leigh, R.A. and Sanders, D. 1997, pp. 218–252. Academic Press London, New York. [CrossRef]
- Herdean, A.; Teardo, E.; Nilsson, A.; Pfeil, B.E.; Johansson, O.N.; Ünnep, R.; Nagy, G.; Zsiros, O.; Dana, S.; Solymosi, K.; et al. A voltage-dependent chloride channel fine-tunes photosynthesis in plants. Nat Comm. 2016, 7, 11654. [CrossRef]
- Keller, B.; Hedrich, R.; Raschke, K. Voltage-dependent anion channels in the plasma membrane of guard cells. Nature 1989, 341, 450-453. [CrossRef]
- Speer, M.; Kaiser, W.M. Ionic relations of symplastic and apoplastic space in leaves from Spinacia oleracea L. and Pisum sativum L. under salinity. Plant Physiol. 1991, 97, 990-997. [CrossRef]
- Cosgrove, D.J.; Hedrich, R. Stretch-activated Cl-, K+ and Ca2+ channels coexisting in plasma membrane of guard cells of Vicia faba L. Planta 1991, 186, 143-153. [CrossRef]
- Ward, J.M.; Maser, P.; Schroeder, J.I. Plant ion channels: gene families, physiology, and functional genomics analyses. Annu. Rev. Physiol. 2009, 71, 59-82. [CrossRef]
- Veley, K.M.; Haswell, E.S. Plastids and pathogens: mechanosensitive channels and survival in a hypoosmotic world. Plant Signal Behav. 2012, 7, 668-671. [CrossRef]
- Maksaev, G.; Haswell, E.S. MscS-Like10 is a stretch-activated ion channel from Arabidopsis thaliana with a preference for anions. Proc. Natl. Acad. Sci. USA 2012, 109, 19015-19020. [CrossRef]
- Deger, A.G.; Scherzer, S.; Nuhkat, M.; Kedzierska, J.; Kollist, H.; Brosch, M.; Unyayar, S.; Boudsocq, M.; Hedrich, R.; Roelfsema M.R.G. Guard cell SLAC1-type anion channels mediate flagellin-induced stomatal closure. New Phytol. 2015, 208, 162-173. [CrossRef]
- Michard, E.; Simon, A.A.; Tavares, B.; Wudick, M.M.; Feijó, J.A. Signaling with ions: The keystone for apical cell growth and morphogenesis in pollen tubes. Plant Physiol. 2017, 173, 91-111. [CrossRef]
- De Angeli, A.; Zhang, J.; Meyer, S.; Martinoia, E. AtALMT9 is a malate-activated vacuolar chloride channel required for stomatal opening in Arabidopsis. Nat Comm. 2013, 4, 1804. [CrossRef]
- Husain, S.; Susanne von Caemmerer, S.V.; Munns, R. Control of salt transport from roots to shoots of wheat in saline soil. Funct. Plant Biol. 2004, 31, 1115-1126. [CrossRef]
- Yermiyahu, U.; Ben-Gal, A.; Keren, R.; Reid, R.J. Combined effect of salinity and excess boron on plant growth and yield. Plant Soil 2008, 304, 73-87. [CrossRef]
- Masood, S.; Wimmer, M.A.; Witzel, K.; Zörb, C.; Mühling, K.H. Interactive effects of high boron and NaCl stresses on subcellular localization of chloride and boron in wheat leaves. J. Agron. Crop Sci. 2012, 198, 227-235. [CrossRef]
- Pineros, M.A.; Cancado, G.M.A.; Kochian, L.V. Novel properties of the wheat aluminum tolerance organic acid transporter (TaALMT1) revealed by electrophysiological characterization in Xenopus oocytes: functional and structural implications. Plant Physiol. 2008, 147, 2131-2146. [CrossRef]
- Li, B.; Qiu, J.; Jayakannan, M.; Xu, B.; Li, Y.; Mayo, G.M.; et al. AtNPF2.5 modulates chloride (Cl-) efflux from roots of Arabidopsis thaliana. Front. Plant Sci. 2017, 7, 02013. [CrossRef]
- Li, B.; Byrt, C.; Qiu, J.; Baumann, U.; Hrmova, M.; Evrard, A.; Johnson, A.A.T.; Birnbaum, K.D.; Mayo, G.M.; Jha, D.; et al. Identification of a stelar-localized transport protein that facilitates root-to-shoot transfer of chloride in Arabidopsis. Plant Physiol. 2016, 170, 1014-1029. [CrossRef]
- Baetz, U.; Eisenach, C.; Tohge, T.; Martinoia, E.; Angeli, A.D. Vacuolar chloride fluxes impact ion content and distribution during early salinity stress. Plant Physiol. 2016, 172, 1167-1181. [CrossRef]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the Move. Trends Plant Sci. 2017, 22, 236-248. [CrossRef]
- Wege, S.; Gilliham, M.; Henderson, S.W. Chloride: Not simply a ‘cheap osmoticum’, but a beneficial plant macronutrient. J. Exp. Bot. 2017, 68, 3057-3069. [CrossRef]
- Wang, S.; Su, S. Z.; Wu, Y.; Li, S. P.; Shan, X. H.; Liu, H. K.; Wang, S.; Yuan, Y.P. Overexpression of maize chloride channel gene ZmCLC-d in Arabidopsis thaliana improved its stress resistance. Biol. Plant. 2015, 59, 55-64. [CrossRef]
- Luo, M.; Zhang, Y.; Li, J.; Zhang, P.; Chen, K.; Song, W.; Wang, X.; Yang, J.; Lu, X.; Lu, B.; Zhao, Y.; Zhao, J. Molecular dissection of maize seedling salt tolerance using a genome-wide association analysis method. Plant Biotech. 2021, 19, 1937-1951. [CrossRef]
- Li, W.Y.F.; Wong, F.L.; Tsai, S.N.; Phang T.H.; Shao, G.H.; Lam, H.M. Tonoplast-located GmCLC1 and GmNHX1 from soybean enhance NaCl tolerance in transgenic bright yellow (BY)-2 cells. Plant Cell Environ. 2006, 29, 1122-1137. [CrossRef]
- Wong, T.H.; Li, M.W.; Yao, X.Q.; Lam, H.M. The GmCLC1 protein from soybean functions as a chloride ion transporter. J. Plant Physiol. 2013, 170, 101-104. [CrossRef]
- Wei, P.; Wang, L.; Liu, A. Yu, B.; Lam, H.M. GmCLC1 confers enhanced salt tolerance through regulating chloride accumulation in soybean. Front. Plant Sci. 2016, 7, 1082. [CrossRef]
- Guan, R.; Qu, Y.; Guo, Y.; Yu, L.; Liu, Y.; Jiang, J.; Chen, J.; Ren, Y.; Liu, G.; Tian, L.; et al. Salinity tolerance in soybean is modulated by natural variation in GmSALT3. Plant J. 2014, 80, 937-950. [CrossRef]
- Do, T.D.; Chen, H.; Hien, V.T.T.; Hamwieh, A.; Yamada, T.; Sato, T.; Yan, Y.; Cong, H.; Shono, M.; Suenaga, K.; Xu, D. Ncl synchronously regulates Na+, K+ and Cl− in soybean and greatly increases the grain yield in saline field conditions. Sci. Rep. 2016, 6, 19147. [CrossRef]
- Liu, Y.; Yu, L.; Qu, Y.; Chen, J.; Liu, X.; Hong, H.; Liu, Z.; Chang, R.; Gilliham, M.; Qiu, L.; Guan, R.. GmSALT3, which confers improved soybean salt tolerance in the field, increases leaf Cl- exclusion prior to Na+ exclusion but does not improve early vigor under salinity. Front. Plant Sci. 2016, 7, 1485. [CrossRef]
- Zifarelli, G.; Pusch, M. CLC transport proteins in plants. FEBS Lett. 2010, 584, 2122-2127. [CrossRef]
- Lv, Q.; Tang, R.; Liu, H.; Gao, X.; Li, Y.; Zheng, H.; Zhang, H. Cloning and molecular analyses of the Arabidopsis thaliana chloride channel gene family. Plant Sci. 2009, 176, 650-661. [CrossRef]
- Flowers, T.J.; Hajibagheri, M.A.; Yeo, A.R. Ion accumulation in the cell-walls of rice plants growing under saline conditions: evidence for the Oertli hypothesis. Plant Cell Environ. 1991, 14, 319-325. [CrossRef]
- Maathuis, F.J.M. Sodium in plants: perception, signalling and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849-858. [CrossRef]
- Corratgé-Faillie, C.; Jabnoune, M.; Zimmermann, S.; Véry, A.A.; Fizames, C.; Sentenac, H. Potassium and sodium transport in non-animal cells: the Trk/Ktr/HKT transporter family. Cell Mol. Life Sci. 2010, 67, 2511-2532. [CrossRef]
- Wen, Z.; Tyerman, S.D.; Dechorgnat, J.; Ovchinnikova, E.; Dhugga, K.S.; Kaiser, B.N. Maize NPF6 proteins are homologs of Arabidopsis CHL1 that are selective for both nitrate and chloride. Plant Cell 2017, 29, 2581-2596. [CrossRef]
- Premkumar, A.; Javed, M.T.; Pawlowski, K.; Lindberg, S.M. Silicate inhibits the cytosolic influx of chloride in protoplasts of wheat and affects the chloride transporters, TaCLC1 and TaNPF2.4/2.5. Plants 2022, 11, 1162 . [CrossRef]
- Chen, Z.C.; Yamaji, N.; Fujii-Kashino, M.; Ma, J.F. A cation-chloride cotransporter gene is required for cell elongation and osmoregulation in rice. Plant Physiol. 2016, 171, 494-507. [CrossRef]
- Nikolovski, N.; Rubtsov, D.; Segura, M.P.; Miles, G.P.; Stevens, T.J.; Dunkley, T.P.J.; Munro, S.; Lilley, K.S.; Dupree, P. Putative glycosyltransferases and other plant Golgi apparatus proteins are revealed by LOPIT proteomics. Plant Physiol. 2012, 160, 1037-1051. [CrossRef]
- Groen, A.J.; Sancho-Andrés, G.; Breckels, L.M.; Gatto, L.; Aniento, F.; Lilley, K.S. Identification of trans-golgi network proteins in Arabidopsis thaliana root tissue. J. Prote. Res. 2014, 13, 763-776. [CrossRef]
- Henderson, S.W.; Wege, S.; Qiu, J.; Blackmore, D.H.; Walker, A.R.; Tyerman, S.D. Walker, R.R.; Gilliham, M. Grapevine and Arabidopsis cation-chloride cotransporters localise to the golgi and trans-golgi network and indirectly influence long-distance ion transport and plant salt tolerance. Plant Physiol. 2015, 169, 2215-2229. [CrossRef]
- Hauser, F.; Horie, T. A conserved primary salt tolerance mechanism mediated by HKT transporters: a mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant Cell Environ. 2010, 33, 552-565. [CrossRef]
- Smethurst, C.F.; Rix, K.; Garnett, T.; Auricht, G.; Bayart, A.; Lane, P.; Wilson, S.J.; Shabala, S. Multiple traits associated with salt tolerance in lucerne: revealing the underlying cellular mechanisms. Funct. Plant Biol. 2008, 35, 640-650. [CrossRef]
- Wu, H.; Zhang, X,; Giraldo, J.P.; and Shabala, S. It is not all about sodium: revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1-17. [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: ecological and agricultural implications. Plants 2021, 10, 419. [CrossRef]
- Srivastava, A.K.; Shankar, A.; Chandran, A.K.N.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608-619. [CrossRef]
- Adhikari, S.; Anuragi, H.; Chandra, K.; Tarte, S.H.; Dhaka, S.R.; Jatav, H.S.; Hingonia, K. Molecular basis of plant nutrient use efficiency - concepts and challenges for its improvement. In Sustainable Plant Nutrition: Molecular Interventions and Advancements for Crop Improvement. 2023, 107-151. [CrossRef]
- Britto, D.T.; Coskun, D.; Kronzucker, H.J. Potassium physiology from Archean to Holocene: A higher-plant perspective. J. Plant Physiol. 2021, 262, 153432. [CrossRef]
- Sharma, T.; Dreyer, I.; Riedelsberger, J.; The role of K+ channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 224. [CrossRef]
- Sánchez-Barrena, M.J.; Chaves-Sanjuan, A.; Raddatz, N.; Mendoza, I.; Cortés, Á.; Gago, F.; González-Rubio, J.M.; Benavente, J.L.; Quintero, F.J.; Pardo, J.M.; Albert, A. Recognition and activation of the plant AKT1 potassium channel by the kinase CIPK23. Plant Physiol. 2020, 182, 2143-2153. [CrossRef]
- Durdagi, S.; Noskov, S.Y. Mechanism of K+/Na+ selectivity in potassium channels from the perspective of the non-selective bacterial channel NaK. Channels 2011, 5, 198-200. [CrossRef]
- Obata, T.; Kitamoto, H.K.; Nakamura, A.; Fukuda, A.; Tanaka, Y. Rice shaker potassium channel OsKAT1 confers tolerance to salinity stress on yeast and rice cells. Plant Physiol. 2007, 144, 1978-1985. [CrossRef]
- Tian, Q.; Shen, L.; Luan, J.; Zhou, Z.; Guo, D.; Shen, Y.; Jing, W.; Zhang, B.; Zhang, A.; Zhang, W. Rice shaker potassium channel OsAKT2 positively regulates salt tolerance and grain yield by mediating K+ redistribution. Plant Cell Environ. 2021, 44, 2951-2965. [CrossRef]
- Wang, X.; Zhao, J.; Fang, Q.; Chang, X.; Sun, M.; Li, W.; Li, Y. GmAKT1 is involved in K+ uptake and Na+/K+ homeostasis in Arabidopsis and soybean plants. Plant Sci. 2021, 304, 110736. [CrossRef]
- Feng, C.; He, C.; Wang, Y.; Xu, H.; Xu, K.; Zhao, Y.; Yao, B.; Zhang, Y.; Zhao, Y.; Carther K.F.I.; et al. Genome-wide identification of soybean Shaker K+ channel gene family and functional characterization of GmAKT1 in transgenic Arabidopsis thaliana under salt and drought stress. J. Plant Physiol. 2021, 266, 153529. [CrossRef]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: single-channel properties, genetic basis and involvement in stress-induced cell death. J Cell Sci. 2010, 123, 1468-1479. [CrossRef]
- Wang, Q.; Ni, L.; Cui, Z.; Jiang, J.; Chen, C.; Jiang M. The NADPH oxidase OsRbohA increases salt tolerance by modulating K+ homeostasis in rice. Crop J. 2022, 10, 1611-1622. [CrossRef]
- Gobert, A.; Isayenkov, S.; Voelker, C.; Maathuis, F.J.M. The two-pore channel TPK1 gene encodes the vacuolar K+ conductance and plays a role in K+ homeostasis. Proc. Nat. Acad. Sci. USA, 2007, 104, 10726-10731. [CrossRef]
- Maathuis, F. J. M.; Flowers, T.J. Sodium chloride compartmentation leaf vacuoles of the halophyte Suaeda maritima (L.) Dum. and its relation to tonoplast permeability. J. Exp. Bot. 1992, 43, 1219-1223, . [CrossRef]
- Bonales-Alatorre, E.; Pottosin, I.; Shabala, L.; Chen, Z.H.; Zeng F.; Jacobsen, S.E.; Shabala, S. Differential activity of plasma and vacuolar membrane transporters contributes to genotypic differences in salinity tolerance in a halophyte species, Chenopodium quinoa. Int. J. Mol. Sci. 2013, 14, 9267-9285. [CrossRef]
- Choi, W.G.; Toyota, M.; Kim, S.H.; Gilroy, S. Salt stress-induced Ca2+ waves are associated with rapid, long-distance root-to-shoot signaling in plants. Proc. Nat. Acad. Sci. USA 2014, 111, 6497-6502. [CrossRef]
- Pottosin, I.; Dobrovinskaya, O. Non-selective cation channels in plasma and vacuolar membranes and their contribution to K+ transport. J. Plant Physiol. 2014, 171, 732-742. [CrossRef]
- Wu, H.; Shabala, L.; Zhou, M.; Shabala, S. Durum and bread wheat differ in their ability to retain potassium in leaf mesophyll: implications for salinity stress tolerance. Plant Cell Physiol. 2014, 55, 1749-1762. [CrossRef]
- Apsea, M.P.; Blumwaldb, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247-2254. [CrossRef]
- Hamam, A.M.; Coskun, D.; Britto D.; Plett, D.T.; Kronzucker, H.J. Plasma-membrane electrical responses to salt and osmotic gradients contradict radiotracer kinetics, and reveal Na+-transport dynamics in rice (Oryza sativa L.). Planta 2019, 249, 1037-1051. [CrossRef]
- Su, N.; Wu, Q.; Chen, J.; Shabala, L.; Mithöfer, A.; Wang, H.; Qu, M.; Yu, M.; Cui, J.; Shabala, S. GABA operates upstream of H+-ATPase and improves salinity tolerance in Arabidopsis by enabling cytosolic K+ retention and Na+ exclusion. J. Exp. Bot. 2019, 70, 6349-6361. [CrossRef]
- Bose, J.; Shabala, L.; Pottosin, I.; Zeng, F.; Velarde-Buendi, A.; Massart, A.; Poschenrieder, C.; Haridi, Y.; Shabala, S. Kinetics of xylem loading, membrane potential maintenance, and sensitivity of K+-permeable channels to reactive oxygen species: physiological traits that differentiate salinity tolerance between pea and barley. Plant Cell Environ. 2014, 37, 589-600. [CrossRef]
- Rubio, F.; Nieves-Cordones, M.; Horie, T.; Shabala, S. Doing ‘business as usual’ comes with a cost: evaluating energy cost of maintaining plant intracellular K+ homeostasis under saline conditions. New Phytol. 2020, 225, 1097-1104. [CrossRef]
- Bridges, D.; Fraser, M.E.; Moorhead, G.B. Cyclic nucleotide binding proteins in the Arabidopsis thaliana and Oryza sativa genomes. BMC Bioinform. 2005, 6, 6. [CrossRef]
- Gobert, A.; Park, G.; Amtmann, A.; Sanders, D.; Maathuis, F.J.M. Arabidopsis thaliana cyclic nucleotide gated channel 3 forms a non-selective ion transporter involved in germination and cation transport. J. Exp. Bot. 2006, 57, 791-800. [CrossRef]
- Senadheera, P.; Singh, R.K.; Maathuis, F.J.M. Differentially expressed membrane transporters in rice roots may contribute to cultivar dependent salt tolerance. J. Exp. Bot. 2009, 60, 2553-2563. [CrossRef]
- Jin, Y.K.; Jing, W.; Zhang, Q.; Zhang, W.H. Cyclic nucleotide gated channel 10 negatively regulates salt tolerance by mediating Na+ transport in Arabidopsis. J. Plant Res. 2015, 128, 211-220. [CrossRef]
- Lecourieux, D.; Mazars, C.; Pauly, N.; Ranjeva, R.; Pugin, A. Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. Plant Cell 2002, 14, 2627-2641. [CrossRef]
- Chiu, J.C.; Brenner, E.D.; DeSalle, R.; Nitabach, M.N.; Holmes, T.C.; Coruzzi, G.M. Phylogenetic and expression analysis of the glutamate-receptor-like gene family in Arabidopsis thaliana. Mol. Biol. Evol. 2002, 19, 1066-1082. [CrossRef]
- Wang, P.H.; Lee, C.E.; Lin, Y.S.; Lee, M.H.; Chen, P.Y.; Chang, H.C.; Chang, I.F. The glutamate receptor-like protein GLR3.7 interacts with 14-3-3ω and participates in salt stress response in Arabidopsis thaliana. Front Plant Sci. 2019, 10, 1169. [CrossRef]
- Cheng, Y.; Zhang X.; Sun, T.; Tian, Q.; Zhang, W.H. Glutamate receptor homolog 3.4 is involved in regulation of Seed germination under salt stress in Arabidopsis. Plant Cell Physiol. 2018, 59, 978-988. [CrossRef]
- Maathuis, F.J.M. Vacuolar two-pore K+ channels act as vacuolar osmosensors. New Phytol. 2011, 191, 84-91. [CrossRef]
- Pottosin, I. I.; Martínez-Estévez, M.; Dobrovinskaya, O. R.; Muñiz, J. Potassium-selective channel in the red beet vacuolar membrane. J. Exp. Bot. 2003, 54, 663-667. [CrossRef]
- Isner, J.C.; Begum, A.; Nuehse, T.; Hetherington, A.M.; Maathuis, F.J.M. KIN7 kinase regulates the vacuolar TPK1 K+ channel during stomatal closure. Curr. Biol. 2018, 28, 466-472. [CrossRef]
- Hamamoto, S.; Marui, J.; Matsuoka, K.; Maeshima, M.; Yabe, I.; Uozumi, N. Characterization of a tobacco TPK-type K+ channel as a novel tonoplast K+ channel using yeast tonoplasts. Mem. Trans. Stru. Funct. Biogen. 2008, 283, 1911-1920. [CrossRef]
- Latz, A.; Mehlmer, N.; Zapf, S.; Mueller, T.D.; Wurzinger, B.; Pfister, B.; Csaszar, E.; Hedrich, R., Teige, M.; Becker, D. Salt stress triggers phosphorylation of the Arabidopsis vacuolar K+ channel TPK1 by calcium-dependent protein kinases (CDPKs). Mol. Plant. 2013, 6, 274-1289. [CrossRef]
- Isayenkov, S.; Isner, J.C.; Maathuis, F.J. Membrane localisation diversity of TPK channels and their physiological role. Plant Signal Behav. 2011, 6, 1201-1204. [CrossRef]
- Ahmad, I.; Devonshire, J.; Mohamed, R.; Schultze, M.; Maathuis, F.J.M. Overexpression of the potassium channel TPKb in small vacuoles confers osmotic and drought tolerance to rice. New Phytol. 2016, 209, 1040-1048. [CrossRef]
- Demidchik, V.; Maathuis, F.J.M. Physiological roles of nonselective cation channels in plants: from salt stress to signaling and development. New Phytol. 2007, 175, 387-404. [CrossRef]
- Zhang, Z.; Zhang, J.; Chen, Y.; Li, R.; Wang, H.; Weim J.Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.). Mol. Biol. Rep. 2012, 39, 8465-8473. [CrossRef]
- Vallejo, A.J.; Peralta, M.L.; Santa-Maria, G.E. Expression of potassium-transporter coding genes, and kinetics of rubidium uptake, along a longitudinal root axis. Plant, Cell Environ. 2005, 28, 850-886. [CrossRef]
- Bañuelos, M.A..; Garciadeblas, B.; Cubero, B.; Rodriguez-Navarro, A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 2002, 130, 784-795. [CrossRef]
- Hyun, T.K.; Rim, Y.; Kim, E.; Kim, J.S. Genome-wide and molecular evolution analyses of the KT/HAK/KUP family in tomato (Solanum lycopersicum L.). Genes Genom. 2014, 36, 365-374. [CrossRef]
- Song, Z.Z.; Ma, R.J., Yu, M.L. Genome-wide analysis and identification of KT/HAK/KUP potassium transporter gene family in peach (Prunus persica). Genet. Mol. Res. 2015, 14, 774-787. [CrossRef]
- Li, Y.; Peng, L.; Xie, C.; Shi, X.; Dong, C.; Shen, Q.; Xu, Y. Genome-wide identification, characterization, and expression analyses of the HAK/KUP/KT potassium transporter gene family reveals their involvement in K+ deficient and abiotic stress responses in pear rootstock seedlings. Plant Growth Regul. 2018, 85, 187-198. [CrossRef]
- Jin, R.; Jiang, W.; Yan, M. Zhang, A.; Liu, M.; Zhao, P.; Chen, X.; Tang, Z. Genome-wide characterization and expression analysis of HAK K+ transport family in Ipomoea. Biotech. 2021, 11, 3. [CrossRef]
- Yang, T.; Lu, X.; Wang, Y. et al. HAK/KUP/KT family potassium transporter genes are involved in potassium deficiency and stress responses in tea plants (Camellia sinensis L.): expression and functional analysis. BMC Genomics 2020, 21, 556 . [CrossRef]
- Horie, T.; Sugawara, M.; Okada, T.; Taira, T.; Kaothien-Nakayama, P.; Katsuhara, M.; Shinmyo, A.; Nakayama, H. Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells. J. Biosci. Bioeng. 2011, 111, 346-356. [CrossRef]
- Shen, Y.; Shen, L.; Shen, Z.; Jing, W.; Ge, H.; Zhao, J.; Zhang, W. The potassium transporter OsHAK21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell Environ. 2015, 38, 2766-2779. [CrossRef]
- Zhang, X.; Liu, L.; Chen, B.; Qin, Z.; Xiao, Y.; Zhang, Y.; Yao, R.; Liu, H.; Yang, H. Progress in understanding the physiological and molecular responses of Populus to salt stress. Int. J. Mol. Sci. 2019, 20, 1-17. [CrossRef]
- Song, T.; Shi, Y.; Shen, L.; Cao, C.; Shen, Y.; Jing, W.; Tian, Q.; Lin, F.; Li, W.; Zhang, W. An endoplasmic reticulum-localized cytochrome b5 regulates high-affinity K+ transport in response to salt stress in rice. Proc. Nat. Acad. Sci. USA. 2021, 118, e2114347118. [CrossRef]
- Li, A.; Liu, A.; Wu, S.; Qu, K, Hu, H.; Yang, J.; Shrestha, N.; Liu, J.; Ren, G. Comparison of structural variants in the whole genome sequences of two Medicago truncatula ecotypes: Jemalong A17 and R108. BMC Plant Biol. 2022, 22, 77. [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23,104-119. [CrossRef]
- Acharya, B.R.; Sandhu, D.; Dueñas, C.; Dueñas, M.; Pudussery, M., Kaundal, A.; Ferreira, J.F.S.; Suarez, D.L.; Skaggs, T.H. Morphological, physiological, biochemical, and transcriptome studies reveal the importance of transporters and stress signaling pathways during salinity stress in Prunus. Sci. Rep. 2022, 12, 274. [CrossRef]
- Santa-María, G.E.; Rubio, F.; Dubcovsky, J.; Rodríguez-Navarro, A. The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell 1997, 9, 2281-2289. [CrossRef]
- Su, H.; Golldack, D.; Katsuhara, M.; Zhao, C.S.; Bohnert, H.J. Expression and stress-dependent induction of potassium channel transcripts in the common ice plant. Plant Physiol. 2001, 125, 604-614. [CrossRef]
- Elumalai, R.P.; Nagpal, P.; Reed, J. A mutation in the Arabidopsis KT2/KUP2 potassium transporter gene affects shoot cell expansion. Plant Cell. 2002, 14, 119-131. [CrossRef]
- Frans, J.M.; Maathuis, F.J.M. The role of monovalent cation transporters in plant responses to salinity. J. Exp. Bot. 2006, 57, 1137-1147. [CrossRef]
- Han, M.; Wu, W.; Wu, W.H.; Wang, Y. Potassium transporter KUP7 is involved in K+ acquisition and translocation in Arabidopsis root under K+-limited conditions. Mol. Plant 2016, 9, 437-446. [CrossRef]
- Ródenas, R.; García-Legaz, M.F.; López-Gómez, E.; Martínez, V.; Rubio, F.; Botella, M.A. NO3-, PO43− and SO42− deprivation reduced LKT1-mediated low-affinity K+ uptake and SKOR-mediated K+ translocation in tomato and Arabidopsis plants. Physiol. Plant. 2017, 160, 410-424. [CrossRef]
- Schachtman, D.P.; Schroeder, J.I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants. Nature 1994, 370, 655-658. [CrossRef]
- Horie, Y.; Hauser, F.; Schroeder, J.I. HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends in Plant Sci. 2009, 14, 660-668. [CrossRef]
- Zhu, J.; Guo, Y.; Zhou, H.; Hong, Y.; Zhao, C.; Shabalak, S.; Lv, C.; Guo, B.; Zhou, M.; Xu, Z. Genome wide association study and haplotype analysis reveals the role of HvHKT1;5 in potassium retention but not Na+ exclusion in barley (Hordeum vulgare L.). Environ. Exp. Bot. 2022, 201, 104973. [CrossRef]
- Asins, M.J.; Villalta, I.; Aly, M.M.; Olías, R.; Morales P.A.; Huertas, R.; Belver, A. Two closely linked tomato HKT coding genes are positional candidates for the major tomato QTL involved in Na+/K+ homeostasis. Plant Cell Environ. 2013, 36, 1171-1191. [CrossRef]
- Chen, H.; Chen, X; Gu, H.; Wu, B.; Zhang, H.; Yuan, X.; Cui, X. GmHKT1;4, a novel soybean gene regulating Na+/K+ ratio in roots enhances salt tolerance in transgenic plants. Plant Growth Regul. 2014, 73, 299-308 . [CrossRef]
- Venkataraman, V.; Shabala, S.; Véry, A.A.; Hariharan, G.N.; Somasundaram, S.; Pulipati, S.; Sellamuthu, G.; Harikrishnan, M.; Kumari, K.; Shabala, L.; Zhou, M.; Chen, Z, To exclude or to accumulate? Revealing the role of the sodium HKT1;5 transporter in plant adaptive responses to varying soil salinity. Plant Physiol. Biochem. 2021, 169, 333-342. [CrossRef]
- Suzuki, K.; Costa, A.; Nakayama, H.; Katsuhara, M.; Shinmyo, A.; Horie, T. OsHKT2;2/1-mediated Na+ influx over K+ uptake in roots potentially increases toxic Na+ accumulation in a salt-tolerant landrace of rice Nona Bokra upon salinity stress. J. Plant Res. 2016, 129, 67-77. [CrossRef]
- Kobayashi, N.I.; Yamaji; N.; Yamamoto H.; Okubo, K.; Ueno, H.; Costa, A.; Tanoi, K.; Matsumura, H.; Fujii-Kashino, M.; Horiuchi, T.; et al. OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice. Physiol. Plant. 2017, 91, 657-670. [CrossRef]
- Mohanty, A.; Chakraborty, K.; Mondal, S.; Jena, P.; Panda, R.K.; Samal, K.C.; Chattopadhyay, K. Relative contribution of ion exclusion and tissue tolerance traits govern the differential response of rice towards salt stress at seedling and reproductive stages. Environ. Exp. Bot. 2023, 206, 105131. [CrossRef]
- Wang, J.; Nan, N.; Li, N.; Liu, Y.; Wang, T. J.; Hwang, I.; Liu, B.; Xu Z.Y. A DNA methylation reader-chaperone regulator-transcription factor complex activates OsHKT1;5 expression during salinity stress. Plant Cell 2020, 32, 3535-3558. [CrossRef]
- Alam, M.S.; Yang, Z.; Li, C.; Yan, Y.; Liu, Z.; Nazir, M.M.; Xu, J. Loss-of-function mutations of OsbHLH044 transcription factor lead to salinity sensitivity and a greater chalkiness in rice (Oryza sativa L.). Plant Physiol. Biochem. 2022, 193, 110-123. [CrossRef]
- Hazzouri, K.M.; Khraiwesh, B.; Amiri, K.M.A.; Pauli, D.; Blake, T.; Shahid, M.; Mullath, S.K.; Nelson, D.; Mansour, A.L.; Salehi-Ashtiani, K.; Purugganan, M.; Masmoudi, K. Mapping of HKT1;5 Gene in barley using GWAS approach and its implication in salt tolerance mechanism. Front. Plant Sci. 2018, 9, 156. [CrossRef]
- Hmidi, D.; Messedi, D.; Corratgé-Faillie, C.; Marhuenda, T.; Fizames, C.; Zorrig, W.; Abdelly, C.; Sentenac, H.; Véry, A. Investigation of Na+ and K+ transport in halophytes: Functional analysis of the HmHKT2;1 transporter from Hordeum maritimum and expression under saline conditions. Plant Cell Physiol. 2019, 60, 2423-2435. [CrossRef]
- Cao, Y.; Liang, X.; Yin, P.; Zhang, M.; Jiang, C. A domestication-associated reduction in K+ -preferring HKT transporter activity underlies maize shoot K+ accumulation and salt tolerance. New Phytol. 2019, 222, 301-317. [CrossRef]
- Leidi, E.O.; Barragánm V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, M.T.; Cubero, B.; Fernández, J.A.; Bressan, R.A.; Hasegawa, P.M.; Quintero, F.J.; Pardo, J.M. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495-506. [CrossRef]
- Kumari, P.H.; Kumar, S.A.; Sivan, P.; Katam, R.; Suravajhala, P.; Rao, K.S.; Varshney, R.K.; Kishor, P.B.K. Overexpression of a plasma membrane bound Na+/H+ antiporter-like protein (SbNHXLP) confers salt tolerance and improves fruit yield in tomato by maintaining ion homeostasis. Front. Plant Sci. 2017, 7, 2016. [CrossRef]
- Al-Harrasi, I.; Jana, G.A.; Patankar, H.V.; Al-Yahyai, R.; Rajappa, S.; Kumar, P.P.; Yaish, M.W. A novel tonoplast Na+/H+ antiporter gene from date palm (PdNHX6) confers enhanced salt tolerance response in Arabidopsis. Plant Cell Rep. 2020, 39, 1079-1093. [CrossRef]
- Li, W.; Wang, D.; Jin, T.; Chang, Q.; Yin, D.; Xu, S.; Liu, B.; Liu, L. The vacuolar Na+/H+ antiporter gene SsNHX1 from the halophyte Salsola soda confers salt tolerance in transgenic alfalfa (Medicago sativa L.). Plant Mol. Biol. Rep. 2011, 29, 278-290. [CrossRef]
- Joshi, S.; Kaur, K.; Khare, T.; Srivastava, A.K.; Suprasanna, P.; Kumar, V. Genome-wide identification, characterization and transcriptional profiling of NHX-type (Na+/H+) antiporters under salinity stress in soybean. Biotech. 2021, 11, 16. [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107-115. [CrossRef]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2011, 233, 175-188. [CrossRef]
- Stephan, A.B.; Kunz, H.; Yang, E.; Schroeder, J. I. Rapid hyperosmotic-induced Ca2+ responses in Arabidopsis thaliana exhibit sensory potentiation and involvement of plastidial KEA transporters. Proc. Nat. Acad. Sci. USA. 2018, 113, E5242-E5249. [CrossRef]
- Tsujii, M.; Kera, K.; Hamamoto, S.; Kuromori, T.; Shikanai, T.; Uozumi, N. Evidence for potassium transport activity of Arabidopsis KEA1-KEA6. Sci. Rep. 2019, 9, 10040. [CrossRef]
- Cellier, F.; Conéjéro, G.; Ricaud, L.; Luu, D.T.; Lepetit, M.; Gosti, F.; Casse, F. Characterization of AtCHX17, a member of the cation/H+ exchangers, CHX family, from Arabidopsis thaliana suggests a role in K+ homeostasis. Plant J. 2004, 39, 834-846. [CrossRef]
- Jia, B.; Sun, M.; Duan-Mu, H.; Ding, X.; Liu, B.; Zhu, Y.; Sun, X. GsCHX19.3, a member of cation/H+ exchanger superfamily from wild soybean contributes to high salinity and carbonate alkaline tolerance. Sci. Rep. 2017, 7, 9423. [CrossRef]
- Shabala, S. Signaling by potassium: another second messenger to add to the list? J. Exp. Bot. 2017, 68, 4003-4007. [CrossRef]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.; Foster, K.J. et al. Energy costs of salt tolerance in crop plants. New Phytol. 2019, 225, 1072-1090. [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Ramiske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414-431. [CrossRef]
- Xiang, J.; Zhou, X.; Zhang, X.; Liu, A.; Xiang, Y.; Yan, M. The Arabidopsis AtUNC-93 acts as a positive regulator of abiotic stress tolerance and plant growth via modulation of ABA signaling and K+ homeostasis. Front Plant Sci. 2018, 9, 1-15. [CrossRef]
- Huang, S.; Chen, M.; Zhao, Y.; Wen, X.; Guo, Z.; Lu, S. CBL4-CIPK5 pathway confers salt but not drought and chilling tolerance by regulating ion homeostasis. Environ. Exp. Bot. 2020, 179, 104230. [CrossRef]
- Zhang, H.; Lv, F.; Han, X.; Xia, X.; Yin, W. The calcium sensor PeCBL1, interacting with PeCIPK24/25 and PeCIPK26, regulates Na+/K+ homeostasis in Populus euphratica. Plant Cell Rep. 2013, 32, 611-621. [CrossRef]
- An, Y.; Yang, X.X.; Zhang, L.; Zhang, J.; Du, B.; Yao, L.; Li, X.T.; Guo, C. Alfalfa MsCBL4 enhances calcium metabolism but not sodium transport in transgenic tobacco under salt and saline-alkali stress. Plant Cell Rep. 2020, 39, 997-1011. [CrossRef]
- Amjad, M.; Akhtar, J.; Murtaza, B.; Abbas, G.; Jawad, H. Differential accumulation of potassium results in varied salt-tolerance response in tomato (Solanum lycopersicum L.) cultivars. Hortic. Environ. Biotechnol. 2016, 57, 248-258. [CrossRef]
- Che, Y.; Yao, T.; Wang, H.; Wang, Z.; Zhang, H.; Sun, G.; Zhang, H. Potassium ion regulates hormone, Ca2+ and H2O2 signal transduction and antioxidant activities to improve salt stress resistance in tobacco. Plant Physiol. Biochem. 2022, 186, 40-51. [CrossRef]
- Luo, Q.; Wei, Q.; Wang, R.; Zhang, Y.; Zhang, F.; He, Y.; Zhou, S.; Feng, J.; Yang, G.; He, G. BdCIPK31, a calcineurin B-like protein-interacting protein kinase, regulates plant response to drought and salt stress. Front Plant Sci. 2017, 8, 1184. [CrossRef]
- Khan, Y.; Xiong, Z.; Zhang, H.; Liu, S.; Yaseen, T.; Hui, T. Expression and roles of GRAS gene family in plant growth, signal transduction, biotic and abiotic stress resistance and symbiosis formation. Plant Biol. 2021, 24, 404-416. [CrossRef]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541-563. [CrossRef]
- Ma, X.; Li, Q-H.; Yu, Y.N.; Qiao, Y-M.; ul Haq, S.; Gong, Z.-H. The CBL-CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [CrossRef]
- Tomar, R. S.; Katarina, S.; Jajoo, A. Behind the scene: critical role of reactive oxygen species and reactive nitrogen species in salt stress tolerance. J. Agron. Crop Sci. 2021, 207, 577-588. [CrossRef]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium efflux system in signaling and adaptation in plants. Front. Plant Sci. 2011, 2, 85. [CrossRef]
- Demidchik, V.; Shabala,S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: mechanisms and functions. New Phytol. 2018, 220, 49-69. [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in Plants. Ann. Bot. 2003, 92, 487-511. [CrossRef]
- Dodd, A.N., Kudla, J. and Sanders, D. The language of calcium signalling. Annu. Rev. Plant Biol. 2010, 61, 593-620. [CrossRef]
- Park, C.J.; Shin, R. Calcium channels and transporters: Roles in response to biotic and abiotic stresses. Front. Plant Sci. Sec. Plant Membrane Traffic and Transport. 2022, 13, 964059. [CrossRef]
- Pirayesh, N.; Giridhar, M.; Khedher, A.B.; Vothknecht, U.C.; Chigri, F. Organellar calcium signaling in plants: An update. BBA Mol. Cell Res. 2021. 4, 118948. [CrossRef]
- Jiang, W.; Tong, T.; Chen, X.; Deng, F.; Zeng, F.; Pan, R.; Zhang, W.; Chen, G.; Chen, Z.H. Molecular response and evolution of plant anion transport systems to abiotic stress. Plant Mol. Biol. 2022, 110, 397-412. [CrossRef]
- Shigaki, T.; Hirschi, K.D. Diverse functions and molecular properties emerging for CAX cation/H+ exchangers in plants. Plant Biol. 2006, 8, 419-429. [CrossRef]
- Zhao, J.; Shigaki,T.; Mei, H., Guo, X.G.; Cheng, N-H.; Hirschi, K.D. Interaction between Arabidopsis Ca2+/H+ exchanger CAX1 and CAX3. J. Biol. Chem. 2009, 284, 4605-4615. [CrossRef]
- Schulz, P.; Herde, M.; Romeis, T. Calcium-dependent protein kinases: Hubs in plant stress signaling and development. Plant Physiol. 2013, 163, 523-530. [CrossRef]
- Yemelyanov, V.; Shishova, M.; Chirkova, T.V.; Lindberg, S. Anoxia-induced elevation of cytosolic Ca2+ concentration in rice and wheat protoplasts. Planta 2011, 234, 271-280. [CrossRef]
- Morgan, S.H.; Lindberg, S.; Maity, P.J.; Geilfus, C.M.; Plieth, C.; Mühling, K.H. Apoplastic and cytosolic Ca2+ and pH dynamics in salt-stressed Vicia faba leaves change in response to calcium. Func. Plant Biol. 2017, 44, 515-524. [CrossRef]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Calcium signaling in Arabidopsis thaliana responsing to drought and salinity. Plant J. 1997, 12, 1067-1078. [CrossRef]
- Gao, D.; Knight, M.R.; Trewavas, A.J.; Sattelmacher, B.; Plieth, C. Self-reporting Arabidopsis expressing pH and [Ca2+] indicators unveil ion dynamics in the cytoplasm and in the apoplast under abiotic stress. Plant Physiol. 2004, 134, 898-908. [CrossRef]
- Premkumar, A.; Lindberg, S.; Lager, I., Rasmussen, U.; Schulz, A. Phospholipases D with C2-domains are involved in hypoxia signal transduction in Arabidopsis. Physiol. Plant. 2018, 167, 90-110. [CrossRef]
- Sebastiani, L.; Lindberg, S.; Vitagliano, C. Cytoplasmic free Ca2+ dynamics in single tomato (Lycopersicon esculentum) protoplasts subjected to chilling temperatures. Physiol. Plant. 1999, 105, 239-245. [CrossRef]
- Mahi, H. E.; Pérez-Hormaec he, J.; Luca, A. D.; Villalta, I.; Espartero, J.; Gámez-Arjona, F.; Fernández, J.L; Bundó, M.; Mendoza, I.; Mieulet, D.; et al. A critical role of sodium flux via the plasma membrane Na+/H+ exchanger SOS1 in the salt tolerance of rice. Plant Physiol. 2019, 180, 1046-1065. [CrossRef]
- Lamers, J.; Van Der Meer, T.; Testerink, C. How plants sense and respond to stressful environments. Plant Physiol. 2020, 182, 1624-1635. [CrossRef]
- Xie, Q.; Zhou, Y.; Jiang, X. Structure, function, and regulation of the plasma membrane Na+/H+ antiporter Salt Overly Sensitive 1 in plants. Front. Plant Sci. 2022, 13, 866265. [CrossRef]
- Kader, M.A.; Lindberg, S.; Seidel, T.; Golldack, D.; Yemelyanov, V. Sodium sensing induces different changes in free cytosolic calcium concentration and pH in salt-tolerant and -sensitive rice (Oryza sativa) cultivars. Physiol. Plant. 2007, 130, 99-111. [CrossRef]
- Laohavisit, A.; Richards, S.L.; Shabala, L.; Chen, C.; Colaço, R.D.D.A.; Swarbreck, S.M.; Shaw, E.; Dark, A.; Shabala, S.; Shang, Z. et al. Salinity-induced calcium signaling and root adaptation in Arabidopsis require the calcium regulatory protein annexin1. Plant Physiol. 2013, 163, 253-262. [CrossRef]
- Konopka-Postupolka, D.; Clark, G. Annexins overlooked regulators of membrane trafficking in plant cells. Int. J. Sci. 2017, 18, 863. [CrossRef]
- Demidschik, V.; Shabala, S. Mechanisms of cytosolic calcium elevation in plants: the role of ion channels, calcium extrusion systems and NADPH oxidase-mediated ‘ROS-Ca2+ Hub’. Func. Plant Biol. 2018, 45, 9-27.
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium signaling and salt tolerance are diversely entwined in plants. Plant Sig. Behav. 2019, 14, 1665455. [CrossRef]
- McCarty, D.R.; Chory, J. Conservation and innovation in plant signaling pathways. Cell 2000, 3, 201-209. [CrossRef]
- Rubio, F.; Fon, M.; Ródenas, R.; Nieves-Cordones, M.; Alemán, F.; Rivero, R.M.; Martínez, V. A low K+ signal is required for functional high-affinity K+ uptake through HAK5 transporters. Physiol Plant. 2014, 152, 558-570. [CrossRef]
- Pandey, G.K. Emergence of a novel calcium-signaling pathway in plants: CBL-CIPK signaling network. Phys. Mol. Biol. Plants 2008, 14, 51-68. [CrossRef]
- Behera, S.; Xu, Z.; Luoni, L.; Bonza, M.G.; Doccula, F.G.; De Michelis, M.I.; Morris, R.J.; Schwarzländer, M. Cellular Ca2+ signals generate defined pH signatures in plants. Plant Cell 2018, 30, 2704-2719. [CrossRef]
- Ward, J.M.; Schroeder, J.I. Roles of ion channels in initiation of signal transduction in higher plants. In: Aducci, P. (eds) Signal Transduction in Plants. 1997, MCBU Molecular and Cell Biology Updates. Birkhäuser Basel. [CrossRef]
- Okazaki, Y.; Kikuyama, M.; Hiramoto, Y.; Iwasaki, N. Short-term regulation of cytosolic Ca2+, cytosolic pH and vacuolar pH under NaCl stress in the charophyte alga Nitellopsis obtuse. Plant Cell Environ. 1996, 19, 569-576. [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiol. Fourth Ed., Sinauer Associates, Inc., Publisher, Sunderland, Massachusetts, 2006, p. 694.
- McKay, D.W.; McFarlane, H.E.; Qu, Y.; Situmorang, A.; Gilliham, M.; Wege, S. Plant trans-Golgi network/early endosome pH regulation requires cation chloride cotransporter (CCC1). Life 2022, 11, e70701. [CrossRef]
- Kader, Md.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal. Behav. 2010, 3, 1-7. [CrossRef]
- Silva, P.; Geros, H. Regulation by salt of vacuolar H+ATPase and H+pyrophosphatase activities and Na+/H+ exchange. Plant Signal. Behav. 2009, 4, 718-726. [CrossRef]
- Pecherina, A.; Grinberg, M.; Ageyeva, M.; Zanegina, D.; Akinchits, E.; Brilkina, A.; Vodeneev, V. Salt-induced changes in cytosolic pH and photosynthesis in tobacco and potato leaves. Int. J. Mol. Sci. 2022, 24, 491. [CrossRef]
- Chung, C.; Mader, C.; Schmitz, J.; Atladottir, J.; Fitchev, P.; Cornwell, M.L.; Koleske, A.J.; Crawford, S.E.; Gorelick, F. The vacuolar-ATPase modulates matrix metalloproteinase isoforms in human pancreatic cancer. Lab Invest. 2011, 91, 732-743. [CrossRef]
- Hager, A. Role of plasma membrane H+-ATPase in auxin-induced elongation growth: historical and new aspects. J. Plant Res. 2003, 116, 4483-505. [CrossRef]
- Pitann, B.; Schubert, S.; Mühling, K.H. Decline in leaf growth under salt stress due to an inhibition of H+-pumping activity and increase in apoplastic pH of maize leaves. J. Plant Nutr. Soil Sci. 2009, 172, 535-543. [CrossRef]
- Buch-Pedersen, M.J.; Rudashevskaya, E.L.; Berner, T.S.; Venema K, Palmgren MG. Potassium as an intrinsic uncoupler of the plasma membrane H+-ATPase. J. Biol. Chem. 2006, 15, 38285-38292. [CrossRef]
- Ekberg, K.; Pedersen, B.P.; Sørensen, D.M.; Pedersen, M.J. Structural identification of cation binding pockets in the plasma membrane proton pump. Proc. Nat. Acad. Sci. USA 2012, 107, 21400-21405. [CrossRef]
- Reddy, A.S.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with stresses. Role of calcium and calcium/calmodulin-regulated gene expression. Plant Cell 2011, 23, 2010-2032. [CrossRef]
- Hauwink, A.L. The conduction of exitation in Mimosa pudica. Rcueil des travaux neerlandais 1935, 32, 51-91. https://natuurtijdschriften.nl/pub/552269.
- Ricca, U. Transmission of stimuli in plants. Nature 1926, 117, 654-655. [CrossRef]
- Choi, W.G.; Miller, G.; Wallace, I.; Harper, J.; Mittler, R.; Gilroy, S. Orchestrating rapid long-distance signaling in plants with Ca2+, ROS and electrical signals. Plant J. 2017, 90, 698-707. [CrossRef]
- Fichman, Y.; Mittler, R. Integration of electric, calcium, reactive oxygen species and hydraulic signals during rapyid sstemic signaling in plants. Plant J. 2021, 107, 7-20. [CrossRef]
- Felle, H.H.; Zimmermann, M.R. Systemic signalling in barley through action potentials. Planta 2007, 226, 203-214. [CrossRef]
- Bellandi, A.; Papp, D.; Breakspear, A.; Joyce, A.J.; Johnston, M.G.; de Keljzer, J.; Raven, E.C.; Ohtsu, M.; Vincent, T.R.; Miller, A.J.; Sanders, D.; Hogenhout, S.A.; Morris, R.J.; Faulkner, C. Diffusion and bulk flow of amino acids mediate calcium waves in plants. Sci. Adv. 2022, 8, eabo6693. https://www.science.org/doi/10.1126/sciadv.abo6693.
- Shao, Q.; Gao, Q.; Lhamo, D.; Zhang, H., Luan, S. The glutamate- and pH-regulated Ca2+ channels are required for systemic wound signaling in Arabidopsis. Sci. Signal, 2020, 14, 453. https://www.science.org/doi/10.1126/scisignal.aba1453.
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dang, J.L.; Mittler, R. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci Signal. 2009, 18, 45. [CrossRef]
- Evans, M.J.; Choi, W.G.; Gilroy, S.; Morris, R.J. A ROS-assisted calcium wave dependent on the AtRBOHD NADPH oxidase and TPC1 cation channel propagates the systemic response to salt stress. Plant Physiol. 2016, 171, 1771-1784. [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441-445. [CrossRef]
Figure 1.
Na+ influx into a root cell or mesophyll cell in the leaf causes an efflux of K+ from the cell by GORKs or NSCCs by Na+-induced membrane depolarization. The depolarization might activate the H+ATPase in the plasma membrane, pumping out protons that can be used for the Na+/H+ antiporter (SOS1). Na+ can also leak into the vacuole by SVs or FVs. Under high salinity stress most Na+ is transported via NSCCs and HKTs into cells, and also by HAKs. GORK, outwards-rectifying K+ channel; HAK, high affinity K+ channel; NSCC, non-selective cation channel; SV, slow vacuolar channel; FV, fast vacuolar channel.
Figure 1.
Na+ influx into a root cell or mesophyll cell in the leaf causes an efflux of K+ from the cell by GORKs or NSCCs by Na+-induced membrane depolarization. The depolarization might activate the H+ATPase in the plasma membrane, pumping out protons that can be used for the Na+/H+ antiporter (SOS1). Na+ can also leak into the vacuole by SVs or FVs. Under high salinity stress most Na+ is transported via NSCCs and HKTs into cells, and also by HAKs. GORK, outwards-rectifying K+ channel; HAK, high affinity K+ channel; NSCC, non-selective cation channel; SV, slow vacuolar channel; FV, fast vacuolar channel.
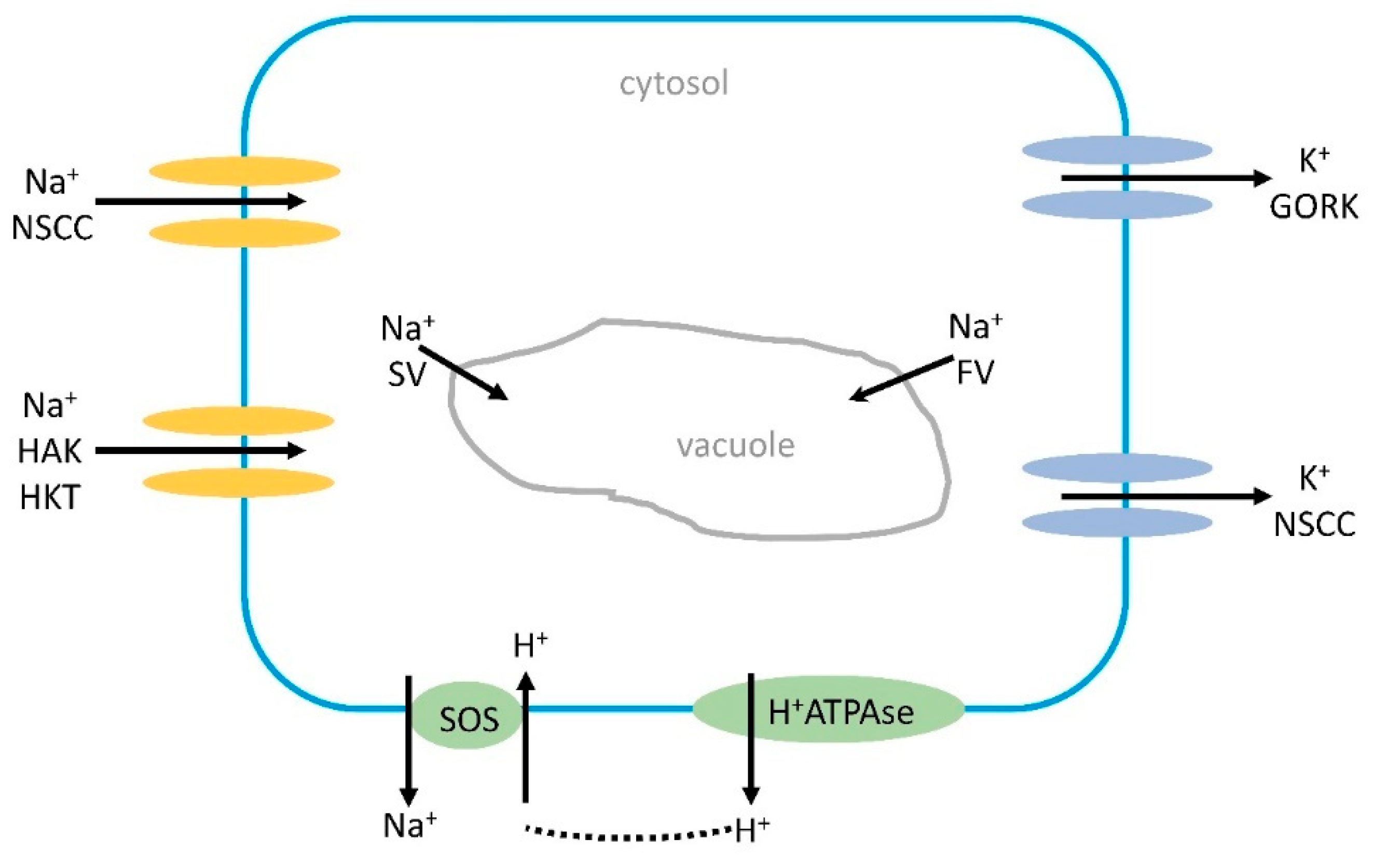
Figure 2.
Protoplasts from rice mesophyll in transmitted light (a), and labelled with Fura 2 (b), labelled with SBFI (c), protoplasts from rice root labelled with SBFI (d), from wheat root in transmitted light (e) and labelled with SBFI (f). Fluorescence emission measured at 530-550 nm.
Figure 2.
Protoplasts from rice mesophyll in transmitted light (a), and labelled with Fura 2 (b), labelled with SBFI (c), protoplasts from rice root labelled with SBFI (d), from wheat root in transmitted light (e) and labelled with SBFI (f). Fluorescence emission measured at 530-550 nm.
Figure 3.
Chloride transport in root, leaf and stem of a plant via channels and transporters. SLAH, anion channel associated homolog; ALMT, aluminum-activated malate transporter; CCC, cation/chloride cotransporter; CLC, chloride channels; NRT, nitrate transporter; NPF, nitrate transporter 1/peptide transporter; SALT3, salt Tolerance-associated gene on chromosome 3; SLAC, slow anion channel; NAXT1, nitrate excretion transporter1, MSL10, mechano-sensitive ion channel 10; X-IRAC, inwardly-rectifying anion channel. For Cl-, various transporters, including NRT, NPF, SLAH, ALMT, and CCC, are involved in uptake and transport over long distances. Cl- influx in the vacuole also involves ALMT and CLC.
Figure 3.
Chloride transport in root, leaf and stem of a plant via channels and transporters. SLAH, anion channel associated homolog; ALMT, aluminum-activated malate transporter; CCC, cation/chloride cotransporter; CLC, chloride channels; NRT, nitrate transporter; NPF, nitrate transporter 1/peptide transporter; SALT3, salt Tolerance-associated gene on chromosome 3; SLAC, slow anion channel; NAXT1, nitrate excretion transporter1, MSL10, mechano-sensitive ion channel 10; X-IRAC, inwardly-rectifying anion channel. For Cl-, various transporters, including NRT, NPF, SLAH, ALMT, and CCC, are involved in uptake and transport over long distances. Cl- influx in the vacuole also involves ALMT and CLC.
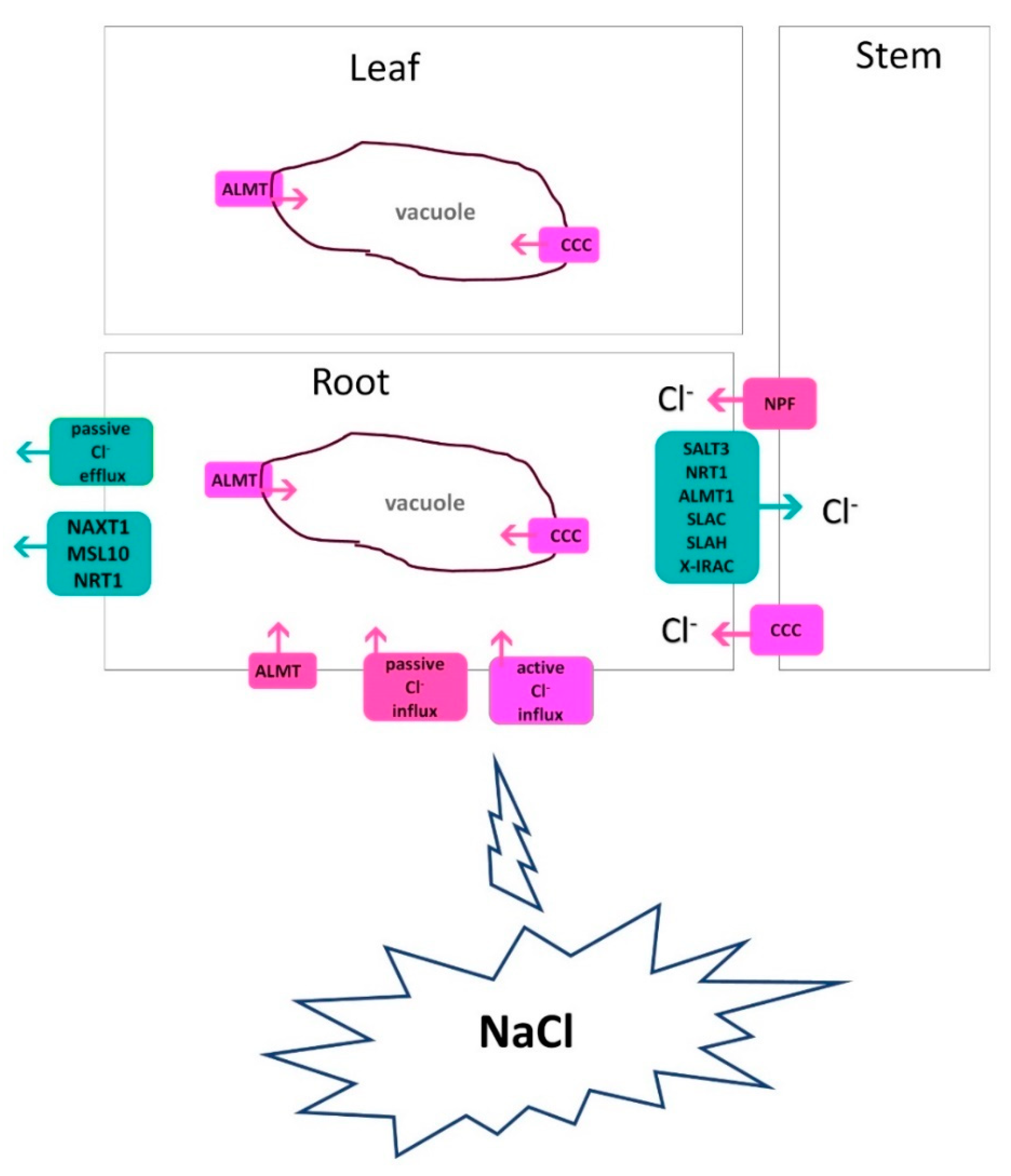
Figure 4.
Calcium transport by channels, antiporters and ATPase-mediated pumps in a plant cell. ACA, autoinhibited Ca2+ATPase; CAX, calcium exchanger; CNGC, cyclic nucleotide-gated channel; DACC, depolarization-activated cation channel; ECA, ER-type Ca2+ATPase; FACC, fast-activating cation channel; GLR, glutamate receptor-like channel; HACC, hyperpolarization-activated cation channel; InsP3R, inositol 1,4,5-trisphosphate receptor-like channel; MLC, mechanosensitive-like channel; NSCC, non-selective cation channel; RyR, cyclic ADP-ribose (cADPR)-activator ryanodine receptor-like channel; SV, slow-activating vacuolar channel.
Figure 4.
Calcium transport by channels, antiporters and ATPase-mediated pumps in a plant cell. ACA, autoinhibited Ca2+ATPase; CAX, calcium exchanger; CNGC, cyclic nucleotide-gated channel; DACC, depolarization-activated cation channel; ECA, ER-type Ca2+ATPase; FACC, fast-activating cation channel; GLR, glutamate receptor-like channel; HACC, hyperpolarization-activated cation channel; InsP3R, inositol 1,4,5-trisphosphate receptor-like channel; MLC, mechanosensitive-like channel; NSCC, non-selective cation channel; RyR, cyclic ADP-ribose (cADPR)-activator ryanodine receptor-like channel; SV, slow-activating vacuolar channel.
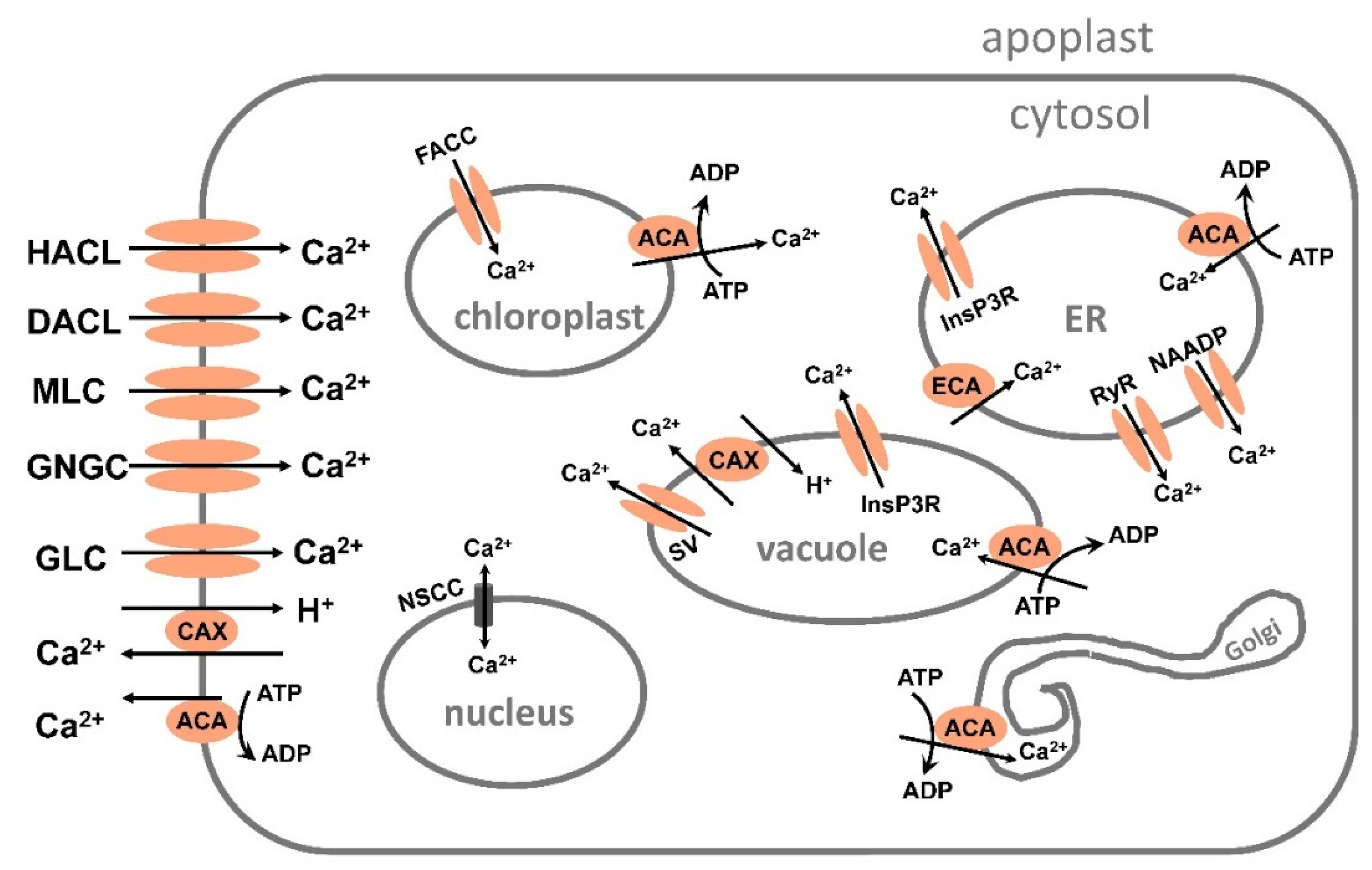
Figure 5.
Simplified model of a cyclic nucleotide-gated channel (adapted from Demidchik et al. [269], with modification). The figure shows one subunit of the tetrameric channel with three CaM-Ca2+ (calmodulin-Ca2+)-binding sites. NT and CT, N- and C-terminal CaM-binding sites; IQ, isoleucine-glutamine domain, where CaM is bound; CNBD, cyclic nucleotide-binding domain; P, part of pore domain.
Figure 5.
Simplified model of a cyclic nucleotide-gated channel (adapted from Demidchik et al. [269], with modification). The figure shows one subunit of the tetrameric channel with three CaM-Ca2+ (calmodulin-Ca2+)-binding sites. NT and CT, N- and C-terminal CaM-binding sites; IQ, isoleucine-glutamine domain, where CaM is bound; CNBD, cyclic nucleotide-binding domain; P, part of pore domain.
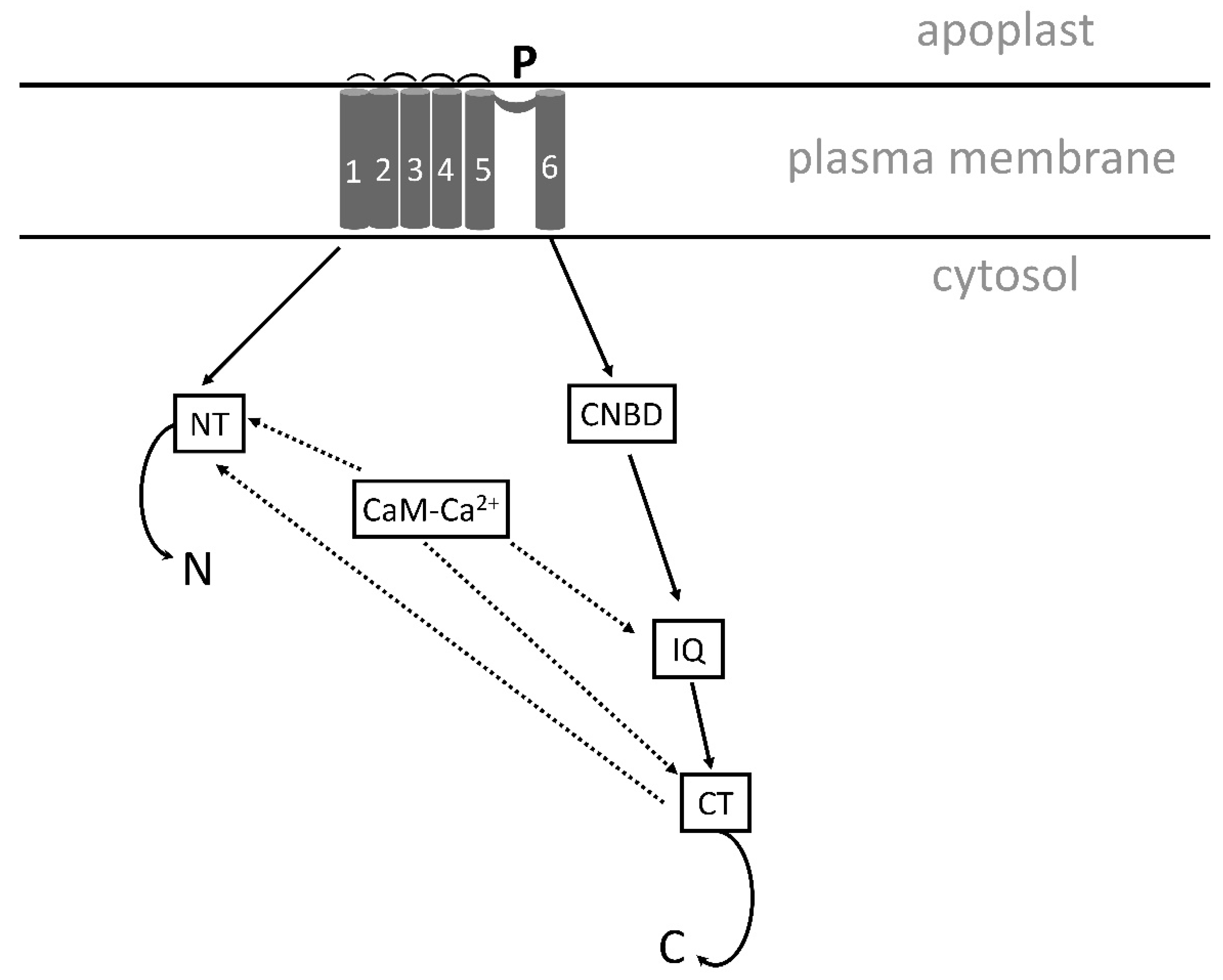
Figure 6.
A possible model of SOS1 structure and its role in Na+-stress tolerance (adopted from Xie et al. [286]. SOS1, the Salt Overly Sensitive 1 Na+/H+ antiporter, is composed of 12 transmembrane domains and transports Na+ out of the cell cytosol and protons into the cytosol. Na+ is proposed to bind to GIPC, a glycosyl inositol phosphoryl ceramide, a sphingolipid in the PM and might also be transported into the cytosol by this lipid. Binding of Na+ to the lipid induces a transient efflux of calcium into the cytosol, either via the sphingolipid molecule or by unknown calcium channels. Calcium can be sensed by SOS3, a calcineurin B-like protein 10, and calcium-binding protein 8, SCABP8. These proteins can bind to and activate SOS2 and transport SOS2 to the serine phosphoryl domain 1136-1138 of SOS1 and activate this antiporter by phosphorylation. 749-925 activation area, 998-1146 C-terminal, 925-997 non-conservative area.
Figure 6.
A possible model of SOS1 structure and its role in Na+-stress tolerance (adopted from Xie et al. [286]. SOS1, the Salt Overly Sensitive 1 Na+/H+ antiporter, is composed of 12 transmembrane domains and transports Na+ out of the cell cytosol and protons into the cytosol. Na+ is proposed to bind to GIPC, a glycosyl inositol phosphoryl ceramide, a sphingolipid in the PM and might also be transported into the cytosol by this lipid. Binding of Na+ to the lipid induces a transient efflux of calcium into the cytosol, either via the sphingolipid molecule or by unknown calcium channels. Calcium can be sensed by SOS3, a calcineurin B-like protein 10, and calcium-binding protein 8, SCABP8. These proteins can bind to and activate SOS2 and transport SOS2 to the serine phosphoryl domain 1136-1138 of SOS1 and activate this antiporter by phosphorylation. 749-925 activation area, 998-1146 C-terminal, 925-997 non-conservative area.
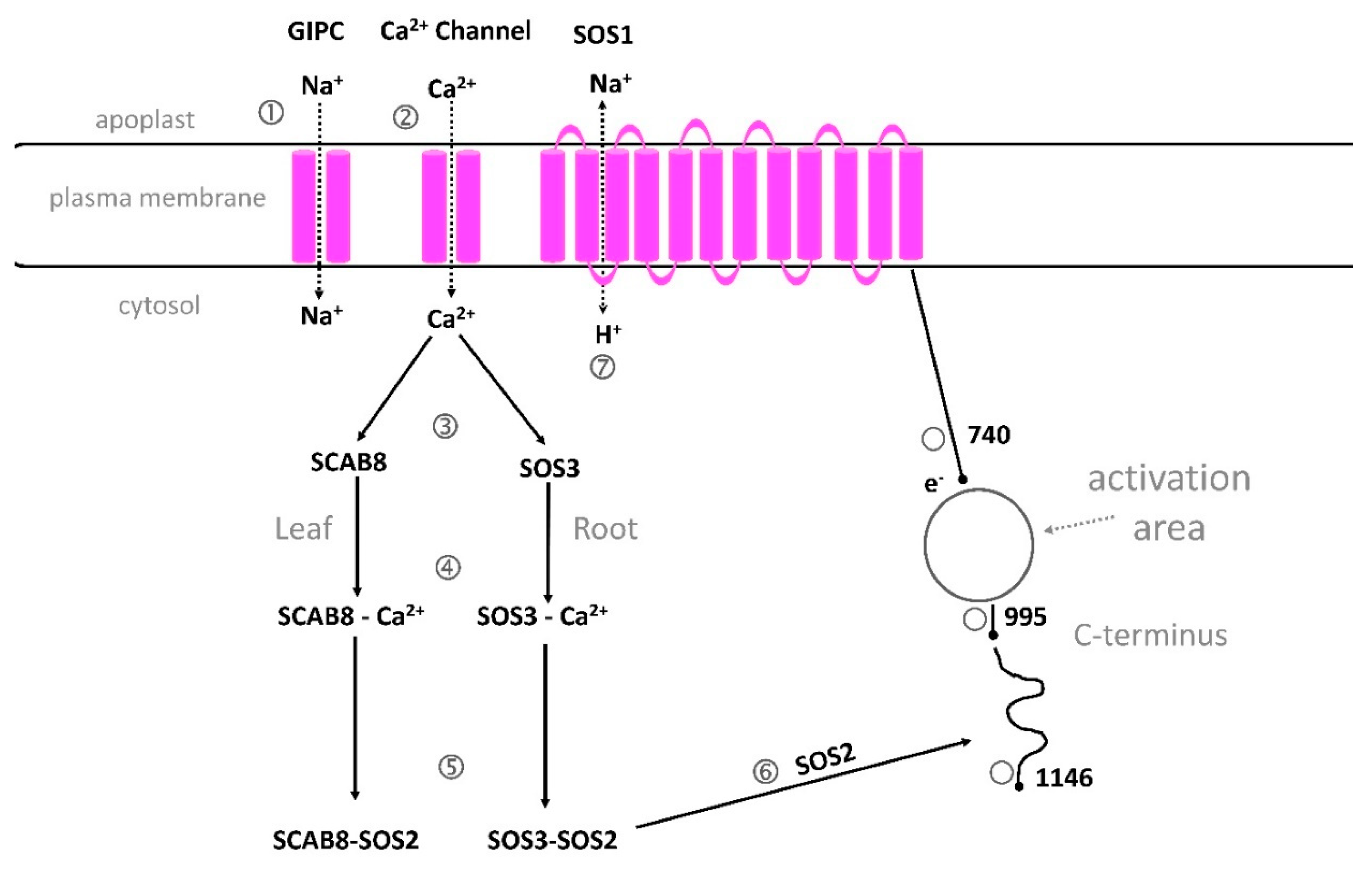
Figure 7.
A simplified model of the CBL-CIPK network leading to ionic and pH homeostasis and ROS, reactive oxygen species, signaling. i/Salt-stress induced calcium signals are sensed by CBL4 and CBL10, which combine with CIPK24 and CIPK8 to CBL4/CIPK24 and CBL10/CIPK8, respectively, which activate the Na+/H + antiporter in the PM and Na+, K+/H+ antiporter in the tonoplast, respectively. This leads to less Na+ in the cytosol. ii/ROS-induced calcium signals are sensed by CBL1, which combines with CIPK26 and activates RBOHF, the respiratory burst oxidase homolog F, which produces ROS by calcium binding to its EF-hand motif. iii/K+ homeostasis is obtained by two mechanisms: In Arabidopsis CBL1 combines with CIPK23 and activates AKT1 and HAK5, Arabidopsis K transporter1 and High affinity K transporter, respectively, and CBL4 combines with CIPK6 and activates AKT2, K transporter2. Iv /The pH homeostasis (by H+) is obtained by binding between CBL7 and CIPK11, which activates AHA2, the PM H+ATPase, responsible for a cytosolic pH homeostasis around 7.5 under normoxia.
Figure 7.
A simplified model of the CBL-CIPK network leading to ionic and pH homeostasis and ROS, reactive oxygen species, signaling. i/Salt-stress induced calcium signals are sensed by CBL4 and CBL10, which combine with CIPK24 and CIPK8 to CBL4/CIPK24 and CBL10/CIPK8, respectively, which activate the Na+/H + antiporter in the PM and Na+, K+/H+ antiporter in the tonoplast, respectively. This leads to less Na+ in the cytosol. ii/ROS-induced calcium signals are sensed by CBL1, which combines with CIPK26 and activates RBOHF, the respiratory burst oxidase homolog F, which produces ROS by calcium binding to its EF-hand motif. iii/K+ homeostasis is obtained by two mechanisms: In Arabidopsis CBL1 combines with CIPK23 and activates AKT1 and HAK5, Arabidopsis K transporter1 and High affinity K transporter, respectively, and CBL4 combines with CIPK6 and activates AKT2, K transporter2. Iv /The pH homeostasis (by H+) is obtained by binding between CBL7 and CIPK11, which activates AHA2, the PM H+ATPase, responsible for a cytosolic pH homeostasis around 7.5 under normoxia.
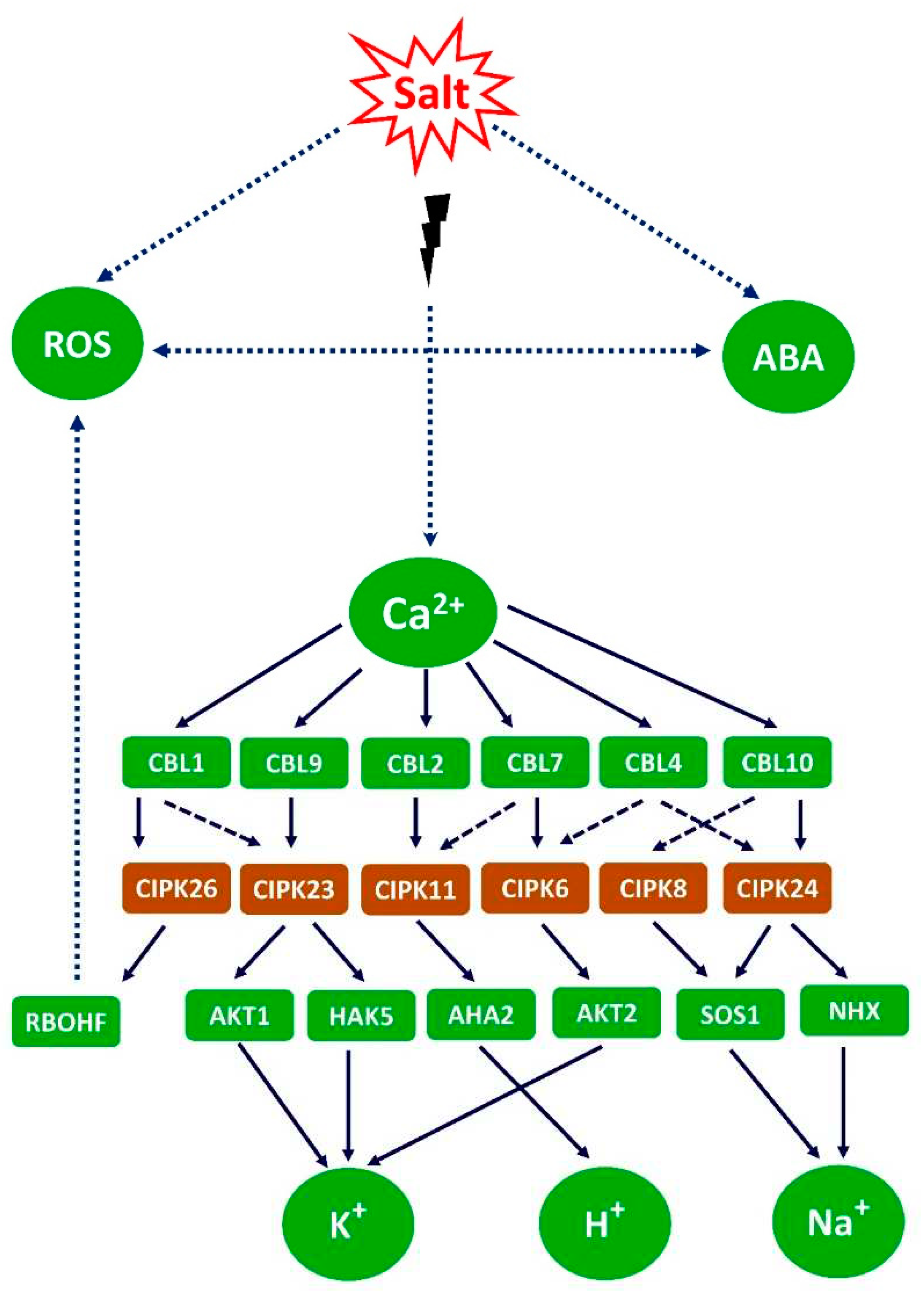
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



