
Submitted:
20 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
In our work, using the analysis of DNA barcodes and morphology (wing color, male genitalia, sphragis shape in females), we show that the group of species close to P. mnemosyne is represented by two phylogenetic lineages, the western and eastern ones. The eastern lineage includes P. stubbendorfii, P. glacialis and P. hoenei. The western lineage includes three morphologically similar species, P. mnemosyne (Western Eurasia), P. turatii (southwestern Europe) and P. nubilosus stat. nov. (Turkmenistan and NE Iran), as well as P. ariadne (Altai). The latter species differs from the rest of the group in the presence of red spots on the wings. Parnassius mnemosyne s.s. is represented by four differentiated mitochondrial clusters that show clear association with specific geographic regions. We propose to interpret them as subspecies: P. mnemosyne mnemosyne (Central and Eastern Europe, N Caucasus, N Turkey), P. mnemosyne adolphi (the Middle East), P. mnemosyne orientalis (Tian Shan) and P. mnemosyne gigantea (Gissar-Alai in Central Asia). We demonstrate that in P. ariadne the red spots on the wing evolved as a reversion to the ancestral wing pattern. This reversion is observed in Altai, where the distribution areas of the western lineage, represented by P. ariadne, and the eastern lineage, represented by P. stubbendorfii, overlap. These two species hybridize in Altai, and we hypothesize that the color change in P. ariadne is the result of reinforcement of prezygotic isolation in the contact zone. Lectotype of Parnassius mnemosyne var. nubilosus Christoph, 1873 is designated.
Keywords:
biodiversity
; COI
; DNA barcoding
; insects
; Lepidoptera
; Parnassius
; Driopa
; taxonomy
; mitochondrial DNA
; sphragis
; reinforcement
1. Introduction
Butterflies of the genus Parnassius Latreille, 1804 are characterized by a unique type of wing pattern consisting of a milky white background, a combination of black spots and translucent areas, and large bright red eye-shaped spots [1,2]. Another morphological feature of the genus is that fertilized females carry a mating plug [2], known in the literature as sphragis (plural: sphragides) [3]. It is a firm waxy structure originating from male accessory glands, fixed to ventral side of female’s abdomen following insemination, where it blocks the ostium bursae [3,4]. Within the genus Parnassius, sphragides are extremely diverse in size and shape [5]. Despite significant indidividual variation [6,7], the sphragides are species-specific and represent a reliable character for species identification [2,5].
The genus attracts the attention of taxonomists due to the complexity of its evolutionary history and classification [1,2,8,9,10,11,12,13,14,15,16,17]. It includes a large number of species protected by international, national and regional laws [18,19,20,21]. It is a model system for studying issues of ecology [22,23,24], biogeography [11,12,25] and insect - plant relationships [26]. It is not surprising, therefore, that significant progress has been made in the study of evolutionary history and taxonomy of the genus, particularly, in the phylogeny reconstruction at the level of subgenera and major species groups [2,8,9,10,11,12,13,15,16,17,27]. There is much less clarity on issues relating to the finer taxonomic structure of the genus at the species level. Especially many unresolved questions remain in the taxonomy of species complexes distributed in the little-studied regions of Central Asia, for example in species that are close to P. delphius Eversmann, 1843 and P. staudingeri Bang-Haas, 1882 [28].
Oddly enough, similar problems exist for the species living in the much more studied regions of Western Europe and the Asian Far East. An example of such a poorly studied complex is a group of taxa close to P. mnemosyne (Linnaeus, 1758). This group is included in the subgenus Driopa Korshunov, 1988 of the genus Parnassius [13,17]. A morphological feature of the P. mnemosyne species complex is the complete absence of red ocelli on the wings [29]. This complex is represented by two phylogenetic lineages, the western and the eastern ones. The western lineage, whose range occupies the western part of Eurasia from the Pyrenees to the Trans-Urals and Central Asia, is known to be represented by the species P. mnemosyne and P. turatii Fruhstorfer, 1908 [30,31]. The eastern lineage, whose range occupies the eastern part of Eurasia from Altai to the Japanese Islands, is represented by the species P. stubbendorfii Ménétriés, 1849, P. glacialis Butler, 1866 and P. hoenei Schweitzer, 1912 [15]. It has recently been shown that this complex should also include the local Altai species P. ariadne (Lederer, 1853) [13,15], which differs sharply from other species of the group by the presence of red ocelli on the hind wings, thus representing a condition characteristic of the genus Parnassius as a whole.
The Western European and Far Eastern populations of the P. mnemosyne group have been the subject of several phylogeographic studies based on the analysis of molecular markers [8,13,21,24,32,33]. However, populations of this group from Eastern Europe, Central Asia and Altai remain almost completely unstudied.
In our study, we close this gap by presenting data on mitochondrial DNA barcodes, wing patterns, and sphragis shape for 189 specimens of the western lineage of the P. mnemosyne species complex from eastern Europe, the Caucasus, Transcaucasia, the Levant, Iran, and Central Asia, and 11 specimens of P. ariadne from Kazakhstan and Russia.
As a result of the research, we
(1) demonstrate that the western lineage of the P. mnemosyne species complex is a group consisting of three morphologically similar species, P. mnemosyne s.s. (Western Eurasia), P. turatii (southwestern Europe) and P. nubilosus Christoph, 1873 stat. nov. (Turkmenistan, NE Iran) and the morphologically highly differentiated species P. ariadne (Altai);
(2) designate the lectotype of the taxon P. mnemosyne var. nubilosus Christoph, 1873;
(3) show that P. mnemosyne s.s. is represented by four differentiated mitochondrial clusters that demonstrate clear connections to certain geographical regions. We propose to interpret them as subspecies: P. mnemosyne mnemosyne (Central and Eastern Europe, Northern Caucasus and Northern Turkey), P. mnemosyne adolphi Bryk, 1911 (the Middle East, Transcaucasus), P. mnemosyne orientalis Verity, 1911 (Tian Shan) and P mnemosyne gigantea Staudinger, 1886 (Gissaro-Alai in Central Asia);
(4) show that the red ocelli on the wings of P. ariadne arose secondarily as a reversion to the ancient wing color pattern characteristic of the genus Parnassius as a whole, and
(5) hypothesize that the red spots arose in P. ariadne as a result of reinforcement of prezygotic reproductive isolation in the zone of secondary contact between the western and eastern lineages of P. mnemosyne species complex.
2. Materials and Methods
Samples
Standard mitochondrial DNA barcodes (658 bp fragments of the cytochrome c oxidase subunit I gene) were obtained for 189 samples of the western lineage of the P. mnemosyne species complex of P. mnemosyne, eleven samples of P. ariadne, and three samples of P. nordmanni (Table 1). The specimens of P. mnemosyne were collected in European part of Russia, Turkey, Iran, Israel, Georgia, Armenia, Azerbaijan, Kazakhstan, Kyrgyzstan, Uzbekistan and Tajikistan (Table 1). The collecting sites cover nearly all the known distribution regions of P. mnemosyne in the eastern half of its distribution range. Specimens examined are deposited in the Zoological Institute of the Russian Academy of Sciences and the Research collection of B. Khramov (St. Petersburg, Russia). Full details of the voucher samples are presented in Table S1 in Supplementary Material 1.
COI Amplification and sequencing
DNA was extracted with standard protocols [34] from single legs removed from dried voucher specimens prior to their re-hydration for spreading. All the voucher specimens are now identified with labels that include the butterfly field numbers (Table S1 in Supplementary Material 1). Additionally, the Barcode labels automatically generated by BOLD system have been added to each specimen. For the majority of samples (175 specimens), the primers LepF (5_-ATTCAACCAATCATAAAGATATTGG-3_) and LepR (5_-TAAACTTCTGGATGTCCAAAAAATCA-3_) amplified the target 658-bp fragment of COI. For the rest of samples, most of which were 20 or more years old, we amplified shorter fragments by using the primer combination MF1 (5_-GCTTTCCCACGAATAAATAATA- 3_)-LepR (407-bp amplicon) and MH-MR1 (5_-CCTGTTCCAGCTCCATTTTC-3_)-LepF (311-bp amplicon). Sequences were obtained by using either ABI 377 or ABI 3730 sequencers (Applied Biosystems).
Sequence analysis
Sequences were edited to remove ambiguous base calls and primer sequences and were assembled by using SEQUENCHER (Gene Codes, Ann Arbor, MI) (https://www.genecodes.com/sequencher-features, accessed on 18 October 2023). Sequences were then aligned by using CLUSTALW [35] software and manually edited. Sequence information was entered in the Barcode of LifeDatabase (BOLD, www.barcodinglife.org) along with an image and collateral information for each voucher specimen. The detailed specimen records and sequence information, including trace files, are available on the BOLD LOWAM project file. All sequences have been submitted to GenBank (Table 1).
Comparison of the obtained COI barcodes revealed 38 unique haplotypes within the five studied species (Table 1). The data matrix for subsequent phylogenetic analysis also included the mitochondrial haplotypes known for P. mnemosyne and P. turatii from Western Europe and Asia Minor, as well as the known haplotypes of other species of the subgenus Driopa [13,32,36,37,38,39,40,41]. Parnassius orleans Oberthür, 1890 is known to be the sister group to all other species of the subgenus Driopa [13,15,17], so it was chosen as the outgroup for rooting the tree. The final alignment of the analyzed samples (file in the FASTA format) is presented in Supplementary Material 2.
The Bayesian analysis of the matrix was performed using the program MrBayes3.2 [42] as previously described [43,44]. A GTR substitution model with gamma-distributed rate variation across sites and a proportion of invariable sites was specified before running the program for 10,000,000 generations with default settings. The first 2500 trees (out of 10000) were discarded prior to computing a consensus phylogeny and posterior probabilities. The consensus of the obtained trees was visualized using FigTree 1.4.4 (http://tree.bio.ed.ac.uk/software/) (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 18 October 2023).
The minimum COI p-distances (%) between the taxa of the subgenus Driopa were calculated using the MEGA 11 program [45].
Morphology analysis
All samples that were used for barcode analysis were also used for morphological studies. Additionally, the samples from the collection of the Zoological Institute (St. Petersburg, Russia) were inspected. This collection includes several thousand individuals of the subgenus Driopa, including two syntypes of P. nubilosus and the samples of P. nubilosus, collected relatively recently by V. Dubatolov in Kopetdagh (Turkmenistan). Photographs of butterflies and sphragides were taken with Nikon D810 digital camera equipped with Nikon AF-S Micro Nikkor 105 mm lens.
For genitalia preparation adult abdomens were soaked in hot (90 °C) 10% KOH for 3–10 min. Then they were transferred to water, the genitalia were carefully extracted and macerated under a stereomicroscope with the help of a pair of preparation needles or with the help of a needle and a watchmaker’s tweezer. Once cleansed of all unwanted elements, they were transferred and stored in glycerine. Cleansed genital armatures were handled, studied and photographed while immersed in glycerine, free from pressure due to mounting and, therefore, free from the ensuing distortion. Photographs of genitalia were taken with a Leica M205C binocular microscope equipped with a Leica DFC495 digital camera, and processed using the Leica Application Suite, v.4.5.0 software.
To reconstruct the probabilities of ancestral states, a Bayesian approach was used as implemented in the program MrBayes3.2 [42]. The states studied were coded as 0 (absence of red spots) and 1 (presence of red spots). These states were implemented into a matrix of molecular features. The probability of ancestral states for each node was calculated separately. The command block used to analyze ancestral states is given in Supplementary Material 3.
3. Results
Phylogenetic analysis
Within the genus Driopa (excluding P. orleans), Bayesian analysis identified three highly supported (in all cases posterior probability = 1) major clades (Figure 1). The first clade is represented by the species P. nordmanni Ménétriés, [1850]. The second clade is represented by the species P. eversmanni Ménétriés, [1850] + P. clodius Ménétriés, 1855. The third clade is represented by species of the P. mnemosyne group, that is, species lacking red spots on the wings. The third clade also included P. ariadne, a species with red spots. The third clade is divided into two highly supported subclades, which can be designated as the western and eastern subclades (Figure 1).
The red-spotted species P. ariadne was found to be deeply nested within the western subclade. Within the western subclade, four main lineages were identified. These are the lineages of (1) P. mnemosyne, (2) P. nubilosus stat. nov., (3) P. ariadne, and (4) P. turatii. The phylogenetic relationships between these lineages were not solved because insufficiently high resolution level of the phylogenetic analysis. Within the eastern subclade, three main lineages were identified, which correspond to the species P. stubbendorfii, P. glacialis and P. hoenei. Within the species P. glacialis, two sublineages were found, which correspond to mainland (China) and island (Japan) populations.
Within the species P. mnemosyne (Fig 2), three highly supported sublineages were identified, which are designated as P. mnemosyne adolphi (the Middle East), P. mnemosyne orientalis (Tian Shan) and P. mnemosyne gigantea (Gissar-Alai in Central Asia). The haplotypes of P. mnemosyne mnemosyne from East Europe, NW Turkey and Caucasus were found to form a basal polytomy on the tree.
All the detected lineages, both major (Figure 1) and secondary lineages (Figure 2), were found to show a clear connection with certain geographical regions (Figure 3): P. turatii with southwestern Europe, P. mnemosyne with Western Eurasia, P. nubilosus stat. nov. with Turkmenistan and NE Iran, P. ariadne with Altai and Saur-Tarbagatai Mts, P. mnemosyne mnemosyne with Central and Eastern Europe, N Caucasus and N Turkey, P. mnemosyne adolphi with the Middle East, P. mnemosyne orientalis with Tian Shan and P. mnemosyne gigantea with Gissar-Alai region in Central Asia.
Morphology
Wing pattern. The wing pattern of butterflies of the P. mnemosyne group is very variable (Figure 4 and Figure 5). However, it is possible to identify elements that are fixed or almost fixed for individual populations. Firstly, it is necessary to note the complete absence of red spots in all species except P. ariadne. In P. ariadne, red spots are present, with the exception of rare aberrant specimens (Figure 5H). A feature of butterflies from the Tian Shan (subspecies orientalis), Gissaro-Alai (subspecies gigantea) and the Middle East (the taxa nubilosus and adolphi) is a white band in the apical transparent part of the fore wings. The males of the taxon nubilosus are characterized by the complete absence of black spots on the hind wings, reduced black spots on the fore wings and an additional black streak on the discal. The last three characters, although they create a characteristic appearance for P. nubilosus, are not absolutely specific to this taxon. Sporadically, the males without black spots on the hind wings and with a reduced pattern on the fore wings are found in populations classified as P. mnemosyne gigantea, P. mnemosyne adolphi, P. mnemosyne mnemosyne and P. turatii (but not P. mnemosyne orientalis).
Male genitalia. In P. nubilosus (Figure 6A,B) as well as in other species of the subgenus Driopa (Figure 6C,D, see also figures in ref. [46]), uncus paired; branches of the gnathos are straight, rod-shaped; saccus is conical with a rounded apex; juxta is strongly sclerotized, V-shaped; valvae are massive; the caudal (lower) process of the valva is separated from the lobe-shaped costal (upper) process by a semicircular notch; aedeagus is thin, long, almost cylindrical. The specific features of the taxon nubilosus are compact, small-sized valvae with square outlines from the lateral view (Figure 6B), and a massive saccus (Figure 6A).
Ancestral state reconstruction
Reconstruction of ancestral states using the MrBayes 3.2 program showed that with probability of 92%, the common ancestor of the P. mnemosyne clade did not have red ocelli on the wings (Figure 1). The common ancestor of the eastern subclade did not have red ocelli with a 99% probability. The common ancestor of the western subclade, which includes species closely related to P. mnemosyne and P. ariadne, did not have red ocelli with a 96% probability. Thus, with a 96% probability, the red ocelli of P. ariadne arose secondarily, probably as a reversion to the condition observed in species of the subgenus occupying a more basal position on the phylogenetic tree.
Nomenclature and Lectotype designation of P. nubilosus
The type series of Parnassius mnemosyne var. nubilosus was collected by famous Russian (of German origin) entomologist Hugo Christoph in 1870 and 1871 from the area of Hadschyabad and Tasch in northeastern Iran [47], a region for which the name Hyrcania was used in the zoological literature of the time [48]. Judging by the original description, the type series included several specimens. This is evidenced by the fact that the taxon was not uncommon in the type-locality (“nicht selten”), the plural is used several times in the description of butterflies, and descriptions of both sexes are given [47].
It is known that one part of Christoph’s collections from Iran (Persia) first ended up in the collection of Grand Duke Nikolai Mikhailovich Romanov, and then in the Zoological Institute in St. Petersburg, and the other part of these collections came through the H.J. El-wes’s collection to the Natural History Museum in London [49]. One syntype (male) of P. mnemosyne var. nubilosus was discovered by Verity [50] in London, where it remains today [51]. This syntype is depicted in the works of Verity [50] and Tshikolovets with coauthors [52]. This male was mistakenly called the holotype in the work of Tshikolovets with coauthors [52]. According to the Codex of Zoological Nomenclature (Article 74.5) [53], this action does not constitute a valid lectotype designation.
In the collection of the Zoological Institute we found two other syntypes of P. mnemosyne var. nubilosus
The name nubilosus was repeatedly used in the taxonomic literature not only for butterflies from NE Iran and S Turkmenistan, but also for butterflies from other regions, including Europe [48,50,54], which is obviously erroneous, based on the data we received. Therefore, to maintain the stability of Zoological nomenclature, we designate the lectotype of Parnassius mnemosyne var. nubilosus.
As a lectotype, we select the male specimen shown in Figure 8 and bearing the following labels: “Hyrcania | v. Nebulosus” [sic]” (in upperside, handwritten), “Alph”[eraky] (in underside, printed); “Кoлл. Вел. Князя | Никoлая | Михаилoвича.” [in Russian. Translation: “coll. Grand Duke Nikolai Mikhailovich]” (printed); “♂ Lectotype | P. mnemosyne | var. nubilosus | Christoph, 1873 | Lukhtanov des. | 24 Oct 2023” (handwritten). The lectotype is preserved in the collection of the Zoological Institute, Russian Academy of Sciences, St. Petersburg.
The paralectotype (female) in the collection of the Zoological Institute, Russian Academy of Sciences, St. Petersburg has the following labels: “Hyrcania | v. Nebulosus” [sic]” (in upperside, handwritten), “Alph”[eraky] (in underside, printed); “Кoлл. Вел. Князя | Никoлая | Михаилoвича.” [in Russian. Translation: “coll. Grand Duke Nikolai Mikhailovich]” (printed); “Paralectotype ♀| P. mnemosyne | var. nubilosus | Christoph, 1873 | Lukhtanov des. | 24 Oct 2023” (handwritten). The male from the collection of the Natural History Museum in London, figured by Verity [50] and Tshikolovets et al. [52] also becomes the paralectotype.
Interestingly, the lectotype has the identification “Nebulosus” (the word is misspelled). Judging by the label, it was made by Sergei Alpheraky, who was the curator of the Grand Duke’s collection. It is interesting that the same spelling of this name (“Nebulosus”) is also found in the monograph by Grum-Grshimailo [55]. Thus, this spelling is more likely to be an unjustified correction of the original spelling than a misprint.
Taxonomy and nomenclature of P. ariadne
Eversmann (ref. [56], pages 539–540, Tab. IX, fig. 1, a-c) described and figured a new species named Doritis clarius, mentioning that it derived from “promontoriis Altaicis australibus” (“mountain spurs of south Altai). However, the name Doritis clarius Eversmann, 1843, is invalid as it is a junior secondary homonym of Papilio clarius Hübner, 1805 (currently both taxa belong to the genus Parnassius) and was replaced by the name Doritis ariadne Lederer (ref. 57, page 354) by Hemming (ref. 58, page 198). The name Doritis ariadne was first published by Lederer [57] in synonymy with Doritis clarius Eversmann, 1843, and is available according to Article 11.6.1 [53] as it was used before 1961; first by Hemming (ref. [58], page 198).
The lectotype of Doritis clarius Eversmann, 1843 was designated by Lukhtanov et al. (2019) [59]. It is preserved in the collection of the Zoological Institute, Russian Academy of Sciences, St. Petersburg. This is a male, labelled: “coll Eversmann” (printed), “clarius ♂” (handwritten), “clarius Eversmann | 1843 Lectotypus ♂ | Kreuzberg design. | 12.09.1989” (red paper, handwritten), “Tarbagatei” (broken label with a handwritten inscription), “Zoological Institute | St. Petersburg | INS_LEP_0000666” (printed) (Figure 9). The lectotype of Doritis clarius Eversmann, 1843 also becomes lectotype of Doritis ariadne Lederer, 1853 according to Code Article 72.7 [53].
The designated lectotype is virtually identical with the male figured by Eversmann (ref. [56], Tab. IX, fig. 1, a-b); most likely, it is this specimen that is depicted by Eversmann on the plate (Tab. IX, fig. 1, a-b). According to the label, the lectotype originated from the “Tarbagatai” Mountains in eastern Kazakhstan. These mountains are located to the south of the Altai Mountains. In old literature the Tarbagatai Mountains were often considered a part of the Altai (e.g., [60]), but currently they are treated as a part of the Saur-Tarbagatai mountain system, which is located between Altai and Tian Shan.
P. ariadne is a local species known from the Kazakhstani, Russian and Chinese parts of Altai and from the Saur-Tarbagatai mountain system in Kazakhstan and China. Populations of this species comprise three geographical groups: (1) the Saur-Tarbagatai group, (2) the group inhabiting the Altai mountains in the Irtysh river basin (separated from the first group by the Zaisan depression), and (3) the group inhabiting the Altai mountains in the basin the Ob River (separated from the Irtysh group by the high main drainage divide of the Altai). We did not find any differentiation between butterflies from these three groups at the COI gene, which is in stark contrast to the situation found within P. mnemosyne.
As for morphology, there are also no significant differences between butterflies from these three population groups. An exception is the population of P. ariadne, described as P. ariadne erlik Yakovlev, 2009 [61] from the uppermost part of the Chuya River basin in the eastern part of the Russian Altai, where butterflies have small red eyes, and individuals without red eyes, as in Figure 5H, are relatively common. However, these characteristics are neither fixed nor unique. In addition, as a distinctive feature of the subspecies P. ariadne erlik, its association with the plant Corydalis stricta Steph. ex DC. (Papaveraceae: Fumarioideae) (the probable food plant of the caterpillars) was noted [61], while the remaining populations of the species are associated with Corydalis nobilis (L.) Pers. We obtained a DNA barcode from one individual of P. ariadne erlik, with characteristic reduced red eyes. The individual was collected by V. Lukhtanov 15 km north of Kosh-Agach on a mountain slope overgrown with Corydalis stricta, not far from the subspecies type locality. Its DNA barcode differed from the barcodes of individuals from other populations by one nucleotide substitution, which cannot be considered a serious distinction. We therefore consider this population, associated with Corydalis stricta, to be a highland form of P. ariadne rather than a separate subspecies.
For this reason, we consider all of the following names proposed as subspecific names as synonyms of P. ariadne ariadne:
clarus Bryk et Eisner, 1932 (Parnassina 2:91) (TL: Saur-Gebirge);
dentatus Austaut, 1889 (Les Parnassiens de la Faune Paléarcticae, p.149 (TL: “…les montagnes de Saisan…”;
erlik Yakovlev, 2009 Atalanta, 40 (1/2): 201; pl. 23, figs. 1–6. TL: «Altai Rep., Chikhacheva Mts., Tabduair [Talduair] Mt., 2500 m»
jiadengyuensis Huang et Murayama, 1992 (Tyo Ga 43: 3) (TL: Altai, Xinjiang Province, China) (TL: Jiadengyu, Altai Mts, Xinjiang, China).
4. Discussion
At the level of species and species groups, the topology of our mitochondrial tree (Figure 1) is fully compatible with the topology obtained for the subgenus Driopa using multigene mitochondrial [15] and nuclear phylogenomic data [17]. Thus, there is no conflicting signals in mitochondrial and nuclear DNA and no reason to assume the influence of Wolbachia or introgressions [62,63] that would lead to mitochondrial discordance in the subgenus Driopa. At the level of terminal lineages, the resulting topology is compatible with the pattern that would be expected if diversification was allopatric, that is, strictly confined to specific geographic regions. The P. turatii lineage is limited to the mountainous regions of southwestern Europe. This type of habitat is not unusual for many species [64]. The lineage of P. mnemosyne mnemosyne is limited to central and eastern Europe, including the adjacent territories of the Urals, Trans-Urals, North Caucasus and Northern Turkey. The P. mnemosyne adolphi lineage is found in the highlands of Western Asia. The P. mnemosyne orientalis lineage is limited to the Tian Shan. The lineage of P. mnemosyne gigantea is limited to the mountain systems of the Gissaro-Alai and western Pamirs. The P. nubilosus lineage is found in the mountain systems of southeastern Iran and Turkmenistan (Kopet Dag). The P. ariadne lineage is found in Altai and the Saur-Tarbagatai mountain system. Each of these regions has numerous endemics at the species and subspecies level, and thus represents areas of independent speciation and subspeciation [64,65,66]. The absence of mitonuclear discordance and the pattern “one lineage - one geographic region” indicate that the mitochondrial phylogenies [13, 16, our data) correctly reflects the processes of phylogenesis and geographic differentiation in the subgenus Driopa.
The level of mitochondrial differentiation between the studied lineages ranges from 0.95 to 2.42% for the subspecies of P. mnemosyne and from 2.29 to 7.8% for the species of the subgenus Driopa. Taking into account these data and the known range rates of mitochondrial evolution in insects [67,68,69], we come to the conclusion that the age of these lineages is in the range of 0.5 - 5 million years. This range fits entirely within the Pliocene and Pleistocene periods. These age estimations for the subgenus Driopa corresponds to the dates in the works of previous authors [13,17]. In the work of Michel et al. [13], P. mnemosyne orientalis and P. mnemosyne gigantea are presented as poorly differentiated taxa, which is a consequence of an error in the subspecies identification. As P. mnemosyne gigantea, this work presents a specimen from the Chatkal Range (Tian Shan), which actually belongs to P. mnemosyne orientalis.
Therefore, it can be assumed that the early divergence of the above-mentioned taxa occurred in Pliocene and Pleistocene refugia in the Pyrenees and/or Apennines (turatii), Altai (ariadne), Kopetdagh (nubilosus), South Anatolia and Transcaucasus (adolphi), the mountains of southeastern Europe and Caucasus (mnemosyne s.s.), Tian Shan (orientalis) and Gissaro-Alai (gigantea). Evolution in these refugia led to deep differentiation, and in four cases to speciation.
Previously, an analysis of molecular markers revealed the taxonomic heterogeneity of populations attributed to P. mnemosyne [13]. On this basis, P. turatii was isolated from this complex as a separate species [30,31]. Our data show that the structure of this group is even more complex and includes another deeply differentiated lineage, namely P. nubilosus. The study of morphology shows that this lineage has a set of almost fixed differences in the pattern of the wings, the different shape of valva in males and a fixed difference in the shape of the sphragis in females. The latter feature is traditionally considered species-specific [1,2]. It is interesting that the sphragis, although it appears after copulation, is not a cast of the male genital structures, since in the genus Driopa slight differentiation in the structure of the male genitalia (e.g., see [46]) is accompanied by noticeable changes in the size and shape of the sphragis. We conclude that this difference is a consequence of the different reproductive behavior of butterflies. Thus, sphragis is a morphological structure, most likely encoded not by genes responsible for anatomical characters, but by genes responsible for behavior.
Theoretically, the complete allopatry of the taxa P. mnemosyne and P. nubilosus and the parapatry of P. mnemosyne and P. turatii allow them to be interpreted as subspecies of a single species. However, from the point of view of the criteria we proposed earlier [28] and taking into account the fact that the COI threshold between 2.1% and 2.6% p-distance produces molecular groupings largely consistent with traditional, morphologically defined species [70], they should be interpreted as species.
The most intriguing question in the evolution of the P. mnemosyne species complex is the origin and loss of a specialized reduced wing pattern without red ocelli on the hindwings. Although the red-spotted wing pattern undoubtedly predominates in the genus Parnassius as a whole [1,2], our data show that the white-black pattern without red ocelli is ancestral for the P. mnemosyne species group and that the red ocelli evolved in P. ariadne secondary as a reversion to the wild type. The secondary appearance of the red spots is observed in the secondary contact zone in Altai, where the western branch of the complex, represented by the species P. ariadne, overlaps with the eastern branch, represented by the species P. stubbendorfii. These two species sporadically hybridize in Altai [71], and we hypothesize that the reversion to the red-ocelli of in P. ariadne is the result of increased prezygotic isolation in order to avoid the maladaptive hybridization [72].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Material 1: List of the studied voucher samples; Supplementary Material 2: The alignment of the analyzed COI sequences; Supplementary Material 2: Reconstruction of ancestral states.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, V.L. and E.Z.; methodology, V.L. and E.Z.; software, V.L. and E.Z.; formal analysis, V.L. and E.Z.; data curation, V.L. and E.Z.; writing—original draft preparation, V.L.; writing—review and editing, V.L. and E.Z. Both authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Russian Science Foundation Grant No 19-14-00202 (Continuation) awarded to the Zoological Institute of the Russian Academy of Sciences (St. Petersburg, Russian Federation) and by the state research project 122031100272-3.
Data Availability Statement
All the analyzed DNA sequences are available via the GenBank links provided.
Acknowledgments
The authors are sincerely grateful to Dr. S.Yu.Sinev for help in working with the collection of the Zoological Institure (St. Petersburg), A.V.Gagarina for the help in sequencing the sample of P. ariadne erlik, B.A.Khramov for the materials provided for examination, A.Novikov (Jerusalem) and E.A.Pazhenkova (Ljubljana) for help in field studies, Ju.Ja.Sokolova and V.V. Dubatolov for providing the samples of P. nubilosus from Turkmenistan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bryk, F. Lepidoptera, Parnassidae pars II. (Subfam. Parnassiinae); W. de Gruyter: Berlin-Leipzig, Germany, 1935; pp. 1–790. [Google Scholar]
- Ackery, P.R. A guide to the genera and species of Parnassiinae (Lepidoptera: Papilionidae). Bull. Br. Mus. 1975, 31, 71–105. [Google Scholar] [CrossRef]
- Carvalho, A.P.S.; Orr, A.G.; Kawahara, A.Y. A review of the occurrence and diversity of the sphragis in butterflies (Lepidoptera, Papilionoidea). Zookeys 2017, 694, 41–70. [Google Scholar] [CrossRef] [PubMed]
- Vlasanek, P.; Konvicka, M. Sphragis in Parnassius mnemosyne (Lepidoptera: Papilionidae): Male-derived insemination plugs loose efficiencywith progress of female flight. Biologia 2009, 64, 1206–1211. [Google Scholar] [CrossRef]
- Bryk, F. Geographische und individuelle Variabilität der Sphragisbildung (Lepidoptera: Parnassiidae). Entomol. tidskr. 1950, 71, 230–234. [Google Scholar]
- Gór, Á.; Lang, Z.; Pásztor, K.; Szigeti, V.; Vajna, F.; Kis, J. Mate-guarding success depends on male investment in a butterfly. Ecol Evol. 2023, 13, e10533. [Google Scholar] [CrossRef] [PubMed]
- Gór, Á.; Fónagy, A.; Pasztor, K.; Szigeti, V.; Lang, Z.; Kis, J. Facultative male investment in prolonged mate-guarding in a butterfly. Behaviour 2023, 160, 515–557. [Google Scholar] [CrossRef]
- Yagi, T.; Katoh, T.; Chichvarkhin, A.; Shinkawa, T.; Omoto, K. Molecular phylogeny of butterflies Parnassius glacialis and P. stubbendorfii at various localities in East Asia. Genes Genet. Syst. 2001, 76, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Omoto, K.; Katoh, T.; Chichvarkhin, A.; Yagi, T. Molecular systematics and evolution of the “Apollo” butterflies of the genus Parnassius (lepidoptera: Papilionidae) based on mitochondrial DNA sequence data. Gene 2004, 326, 141–147. [Google Scholar] [CrossRef]
- Omoto, K.; Yonezawa, T.; Shinkawa, T. Molecular systematics and evolution of the recently discovered “Parnassian” butterfly (Parnassius davydovi Churkin, 2006) and its allied species (Lepidoptera, Papilionidae). Gene 2009, 441, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Sperling, F.A.H.; Nazari, V. Mitochondrial DNA divergence and phylogeography in western Palaearctic Parnassiinae (lepidoptera: Papilionidae): How many species are there? Insect Syst. Evol. 2007, 38, 121–138. [Google Scholar] [CrossRef]
- Nazari, V.; Zakharov, E.V.; Sperling, F.A.H. Phylogeny, historical biogeography, and taxonomic ranking of Parnassiinae (lepidoptera, Papilionidae) based on morphology and seven genes. Mol. Phylogenet. Evol. 2007, 42, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Rebourg, C.; Cosson, E.; Descimon, H. Molecular phylogeny of Parnassiinae butterflies (lepidoptera: Papilionidae) based on the sequences of four mitochondrial DNA segments. Ann. Soc. Entomol. Fr. 2008, 44, 1–36. [Google Scholar] [CrossRef]
- Rose, K.; Weiss, J.C. The Parnassiinae of the world. Part 5; Goecke & Evers: Keltern, Germany, 2011; 120p. [Google Scholar]
- Condamine, F.L.; Rolland, J.; Höhna, S.; Sperling, F.A.H.; Sanmartín, I. Testing the role of the red queen and court jester as drivers of the macroevolution of Apollo butterflies. Syst. Biol. 2018, 67, 940–964. [Google Scholar] [CrossRef]
- Zhao, Y.; He, B.; Tao, R.; Su, C.; Ma, J.; Hao, J.; Yang, Q. Phylogeny and biogeographic history of Parnassius butterflies (Papilionidae: Parnassiinae) reveal their origin and deep diversification in West China. Insects 2022, 13, 406. [Google Scholar] [CrossRef]
- Tian, X.; Mo, S.; Liang, D.; Wang, H.; Zhang, P. Amplicon capture phylogenomics provides new insights into the phylogeny and evolution of alpine Parnassius butterflies (Lepidoptera: Papilionidae). Syst. Entomol. 2023, 48, 571–584. [Google Scholar] [CrossRef]
- Todisco, V.; Gratton, P.; Cesaroni, D.; Sbordoni, V. Phylogeography of Parnassius apollo: Hints on taxonomy and conservation of a vulnerable glacial butterfly invader. Biol. J. Linn. Soc. 2010, 101, 169–183. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F. Anthropogenic threats to high-altitude parnassian diversity. News of The Lepidopterists’ Society 2018, 60, 94–99. Available online: https://hal.science/hal-02323624/.
- The IUCN Red List. Available online: https://www.iucnredlist.org/search?query=Parnassius&searchType=species (accessed on 18 October 2023).
- Talla, V.; Mrazek, V.; Höglund, J.; Backström, N. Whole genome re-sequencing uncovers significant population structure and low genetic diversity in the endangered clouded Apollo (Parnasssius mnemosyne) in Sweden. Conserv. Genet. 2023, 24, 305–314. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.H.; Wahlberg, N.; Rasplus, J.; Kergoat, G.J. What causes latitudinal gradients in species diversity? Evolutionary processes and ecological constraints on swallowtail biodiversity. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zaman, K.; Hubert, M.K.; Schoville, S.D. Testing the role of ecological selection on color pattern variation in the butterfly Parnassius clodius. Mol Ecol. 2019, 28, 5086–5102. [Google Scholar] [CrossRef]
- Tao, R.; Xu, C.; Wang, Y.; Sun, X.; Li, C.; Ma, J.; Hao, J.; Yang, Q. Spatiotemporal differentiation of alpine butterfly Parnassius glacialis (Papilionidae: Parnassiinae) in China: Evidence from mitochondrial DNA and nuclear single nucleotide polymorphisms. Genes 2020, 11, 188. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Xie, T.; Wang, Y.; Si, C.; Li, L.; Ma, J.; Li, C.; Sun, X.; Hao, J.; Yang, Q. Miocene diversification and high-altitude adaptation of Parnassius butterflies (lepidoptera: Papilionidae) in Qinghai–Tibet plateau revealed by large-scale transcriptomic data. Insects 2020, 11, 754. [Google Scholar] [CrossRef] [PubMed]
- Allio, R.; Nabholz, B.; Wanke, S.; Chomicki, G.; Pérez-Escobar, O.A.; Cotton, A.M.; Clamens, A.L.; Kergoat, G.J.; Sperling, F.A.H.; Condamine, F.L. Genome-wide macroevolutionary signatures of key innovations in butterflies colonizing new host plants. Nat Commun. 2021, 12, 354. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhao, Y.; Su, C.; Lin, G.; Wang, Y.; Li, L.; Ma, J.; Yan, Q.; Hao, J. Phylogenomics reveal extensive phylogenetic discordance due to incomplete lineage sorting following the rapid radiation of alpine butterflies (Papilionidae: Parnassius). Syst. Entomol. 2023, 48, 585–599. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Sourakov, A.; Zakharov, E.V. DNA barcodes as a tool in biodiversity research: Testing pre-existing taxonomic hypotheses in Delphic Apollo butterflies (Lepidoptera, Papilionidae). Syst. Biodivers. 2016, 14, 599–613. [Google Scholar] [CrossRef]
- Weiss, J.C. The Parnassiinae of the world. Part 3. Venette, France: Sciences Nat. 1999. Canterbury: Hillside Books 98 pp.
- Bolotov, I.N.; Gofarov, M.Y.; Gorbach, V.V.; Kolosova, Y.S.; Zheludkova, A.A.; Kondakov, A.V.; Spitsyn, V.M. Parnassius nebrodensis: A threatened but neglected Apollo butterfly species from Southern Europe (Lepidoptera: Papilionidae). Ecol. Montenegrina 2021, 40, 140–163. [Google Scholar] [CrossRef]
- Cotton, A.M.; Bolotov, I.N.; Gofarov, M.Y.; Gorbach, V.V.; Kolosova, Y.S.; Zheludkova, A.A.; Kondakov, A.V.; Spitsyn, V.M. The correct name for the South Western European species recently separated from Parnassius mnemosyne (Linnaeus, 1758) (Lepidoptera: Papilionidae). Ecol. Montenegrina 2021, 43, 56–58. [Google Scholar] [CrossRef]
- Gratton, P.; Konopiński, M.K.; Sbordoni, V. Pleistocene evolutionary history of the Clouded Apollo (Parnassius mnemosyne): Genetic signatures of climate cycles and a ‘time-dependent’ mitochondrial substitution rate. Mol Ecol. 2008, 17, 4248–4262. [Google Scholar] [CrossRef] [PubMed]
- Scalercio, S.; Cini, A.; Menchetti, M.; Vodă, R.; Bonelli, S.; Bordoni, A.; Casacci, L.P.; Dincă, V.; Balletto, E.; Vila, R.; et al. How long is 3 km for a butterfly? Ecological constraints and functional traits explain high mitochondrial genetic diversity between Sicily and the Italian Peninsula. J. Anim. Ecol. 2020, 89, 2013–2026. [Google Scholar] [CrossRef]
- Hajibabaei, M.; deWaard, J.R.; Ivanova, N.V.; Ratnasingham, S.; Dooh, R.T.; Kirk, S.L.; Mackie, P.M.; Hebert, P.D. Critical factors for assembling a high volume of DNA barcodes. Philos Trans R Soc Lond B Biol Sci. 2005, 360, 1959–1967. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Dapporto, L.; Cini, A.; Vodă, R.; Dincă, V.; Wiemers, M.; Menchetti, M.; Magini, G.; Talavera, G.; Shreeve, T.; Bonelli, S.; et al. Integrating three comprehensive data sets shows that mitochondrial DNA variation is linked to species traits and paleogeographic events in European butterflies. Mol Ecol Resour. 2019, 19, 1623–1636. [Google Scholar] [CrossRef] [PubMed]
- Dincă, V.; Zakharov, E.V.; Hebert, P.D.; Vila, R. Complete DNA barcode reference library for a country’s butterfly fauna reveals high performance for temperate Europe. Proc. Biol Sci. 2011, 278, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Dincă, V.; Dapporto, L.; Somervuo, P.; Vodă, R.; Cuvelier, S.; Gascoigne-Pees, M.; Huemer, P.; Mutanen, M.; Hebert, P.D.N.; Vila, R. High resolution DNA barcode library for European butterflies reveals continental patterns of mitochondrial genetic diversity. Commun Biol. 2021, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Espeland, M.; Breinholt, J.; Willmott, K.R.; Warren, A.D.; Vila, R.; Toussaint, E.F.A.; Maunsell, S.C.; Aduse-Poku, K.; Talavera, G.; Eastwood, R.; et al. A comprehensive and dated phylogenomic analysis of butterflies. Curr. Biol. 2018, 28, 770–778.e5. [Google Scholar] [CrossRef] [PubMed]
- Litman, J.; Chittaro, Y.; Birrer, S.; Praz, C.; Wermeille, E.; Fluri, M.; Stalling, T.; Schmid, S.; Wyler, S.; Gonsethet, Y. A DNA barcode reference library for Swiss butterflies and forester moths as a tool for species identification, systematics and conservation. PLoS ONE 2018, 13, e0208639. [Google Scholar] [CrossRef] [PubMed]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Huemer, P.; Dincă, V.; van Nieukerken, E.J.; Lopez-Vaamonde, C.; et al. Species-level para- and polyphyly in DNA barcode gene trees: Strong operational bias in European Lepidoptera. Syst Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Vershinina, A.O.; Lukhtanov, V.A. Geographical distribution of the cryptic species Agrodiaetus alcestis alcestis, A. alcestis karacetinae and A. demavendi (Lepidoptera, Lycaenidae) revealed by cytogenetic analysis. Comp. Cytogen. 2010, 4, 1–11. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Gagarina, A.V. Molecular phylogeny and taxonomy of the butterfly subtribe Scolitantidina with special focus on the genera Pseudophilotes, Glaucopsyche and Iolana (Lepidoptera, Lycaenidae). Insects 2022, 13, 1110. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gorbunov, P.J. The butterflies of Russia: Classification, genitalia, keys for identification (Lepidoptera: Hesperioidea and Pappilionoidea). Thesis: Ekaterinburg, Russia, 2001, 320 pp.
- Christoph, H. Weiterer Beitrag zum Verzeichnisse der in Nord-Persien einheimischen Schmetterlinge. Horae Soc. Ent. Ross. 1873, 10, 3–55. [Google Scholar]
- Staudinger, O. Macrolepidoptera. In Catalog der Lepidopteren des palaearctischen Faunengebites; Staudnger, O., Rebel, H., Eds.; Friedländer: Berlin, Germany, 1901; pp. 1–411. [Google Scholar]
- Horn, W.; Kahle, I.; Friese, G.; Gaedike, R. Collectiones entomologicae. Ein Kompendium über den Verbleib entomologischer Sammlungen der Welt bis 1960. Akademie der Landwirtschaftswissenschaften der Deutschen Demokratischen Republik: Berlin, 1990. Teil 1: A bis K, pp. 1−220. Teil 2: L bis Z, pp. 221−573.
- Verity, R. Rhopalocera palaearctica: Iconographie et description des papilions diurnes de la region paléarctique. Papilionidae et Pieridae. Landi Florence, Italy, 1905-1911. 368p.
- Ackery, P.R. A list of the type-specimens of Parnassius (Lepidoptera: Papilionidae) in the British Museum (Natural History). Bull. Br. Mus. Nat. 1973, 29, 3–35. [Google Scholar]
- Tshikolovets, V.; Naderi, A.; Eckweiler, W. The butterflies of Iran and Iraq. Series: Butterflies of Palaearctic Asia, Volume: 10; Tshikolovets Publications: Pardubice, Czech Republic, 2014; 366p. [Google Scholar]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature; The International Trust for Zoological Nomenclature: London, UK, 1999; Available online: https://www.iczn.org/the-code/the-code-online/ (accessed on 15 November 2023).
- Nekrutenko, Y.P. The butterflies of the Caucasus. The Guide. Paplionidae, Pieridae, Satyridae, Danaidae. Naukova Dumka: Kiev, Ukraine, 1990; pp. 1–216. (in Russian).
- Grum-Grshimailo, G.E. Le Pamir et sa faune lépidoptérologiae. In Mémoires sur les Lépidoptères; Rimonoff, N.M., Ed.; St.-Pétersbourg Imprimerie de M.M. Stassuléwitch: St.-Petersburg, 1890; Volume 4, 575p. [Google Scholar]
- Eversmann, E. Quedam Lepidopterorum species novae, in montibus Uralensibus et Altaicis habitantes, nunc descriptae et depictae. Bull. Soc. Nat. Moscou 1843, 16, 535–555, + pls. vii–x. [Google Scholar]
- Lederer, J. Lepidopterologisches aus Sibirien. Verh. Zool.-Bot. Ver. Wien 1853, 3, 351–386. [Google Scholar]
- Hemming, F. Revisional notes on some species of Rhopalocera (Lepidoptera). Stylops 1934, 3, 193–200. [Google Scholar]
- Lukhtanov, V.A.; Pelham, J.P.; Cotton, A.M.; Calhoun, J.V. Case 3767 – Papilio phoebus Fabricius, 1793 (currently Parnassius phoebus; Insecta, Lepidoptera): Proposed conservation of prevailing usage of the specific name and that of Doritis ariadne Lederer, 1853 (currently Parnassius ariadne) by the designation of a neotype. Bull. zool. nomencl. 2019, 78, 14–22. [Google Scholar] [CrossRef]
- Elwes, H. On the Lepidoptera of the Altai mountains. Trans. ent. Soc. Lond. 1899, 47, 295–367. [Google Scholar] [CrossRef]
- Yakovlev, R.V. A new subspecies of Parnassius ariadne (Lederer, 1853) from the South-Eastern part of Russian Altai (Lepidoptera, Papilionidae). Atalanta 2009, 40, 201. [Google Scholar]
- Ritter, S.; Michalski, S.G.; Settele, J.; Wiemers, M.; Fric, Z.F.; Sielezniew, M.; Šašić, M.; Rozier, Y.; Durka, W. Wolbachia infections mimic cryptic speciation in two parasitic butterfly species, Phengaris teleius and P. nausithous (Lepidoptera: Lycaenidae). PLoS ONE 2013, 8, e78107. [Google Scholar] [CrossRef]
- Sucháčková Bartoňová, A.; Konvička, M.; Marešová, J.; Wiemers, M.; Ignatev, N.; Wahlberg, N.; Schmitt, T.; Faltýnek Fric, Z. Wolbachia affects mitochondrial population structure in two systems of closely related Palaearctic blue butterflies. Sci. Rep. 2021, 11, 3019. [Google Scholar] [CrossRef] [PubMed]
- Dapporto, L. Speciation in Mediterranean refugia and post-glacial expansion of Zerynthia polyxena (Lepidoptera, Papilionidae). J. Zool. Syst. Evol. Res. 2010, 48, 229–237. [Google Scholar]
- Kryzhanovsky, O.L. Composition and Distribution of Insect Faunas of the World; KMK Press: Moscow, Russia, 2002; 237p, [in Russian]. [Google Scholar]
- Kolesnichenko, K.A.; Kotlobay, A.A. Review of the fritillary species systematically close to Melitaea lutko Evans, 1932 (Lepidoptera: Nymphalidae) with analysis of their geographic distribution and interrelations with host plants. Eur. J. Taxon. 2022, 830, 1–60. [Google Scholar] [CrossRef]
- Brower, A.V.Z. Rapid morphological radiation and convergence among races of the butterfly Heliconius erato inferred from patterns of mitochondrial DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6491–6495. [Google Scholar] [CrossRef] [PubMed]
- Farrell, B.D. Evolutionary assembly of the milkweed fauna: Cytochrome oxidase I and the age of Tetraopes beetles. Mol. Phylogenet. Evol. 2001, 18, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Quek, S.P.; Stuart, J.D.; Itino, T.; Pierce, N.E. Codiversification in an ant –plant mutualism: Stem texture and the evolution of host use in Crematogaster (Formicidae: Myrmicinae) inhabitants of Macaranga (Euphorbiaceae). Evolution 2004, 58, 554–570. [Google Scholar] [CrossRef] [PubMed]
- Young, R.G.; Abbott, C.L.; Therriault, T.W.; Adamowicz, S.J. Barcode-based species delimitation in the marine realm: A test using Hexanauplia (Multicrustacea: Thecostraca and Copepoda). Genome 2017, 60, 169–182. [Google Scholar] [CrossRef]
- Yakovlev, R.V. New information about Parnassius stubbendorfii (Ménétriés, 1948) and Parnassius clarius (Eversman, 1843) (Lepidoptera, Papilionidae) from South-West and Central Altai. Entomol. News Russ. 1998, 1, 20–25. [Google Scholar]
- Lukhtanov, V.A.; Kandul, N.P.; Plotkin, J.B.; Dantchenko, A.V.; Haig, D.; Pierce, N.E. Reinforcement of pre-zygotic isolation and karyotype evolution in Agrodiaetus butterflies. Nature 2005, 436, 385–389. [Google Scholar] [CrossRef]
Figure 1.
The Bayesian tree of the Parnassius (Driopa) species based on analysis of the mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (BPP) (higher than 0.5). Black/red values in the rectangles show the probabilities of the ancestral states “absence of red spots”/”presence of red spots”. Parnassius ariadne erlik is shown in bold.
Figure 1.
The Bayesian tree of the Parnassius (Driopa) species based on analysis of the mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (BPP) (higher than 0.5). Black/red values in the rectangles show the probabilities of the ancestral states “absence of red spots”/”presence of red spots”. Parnassius ariadne erlik is shown in bold.
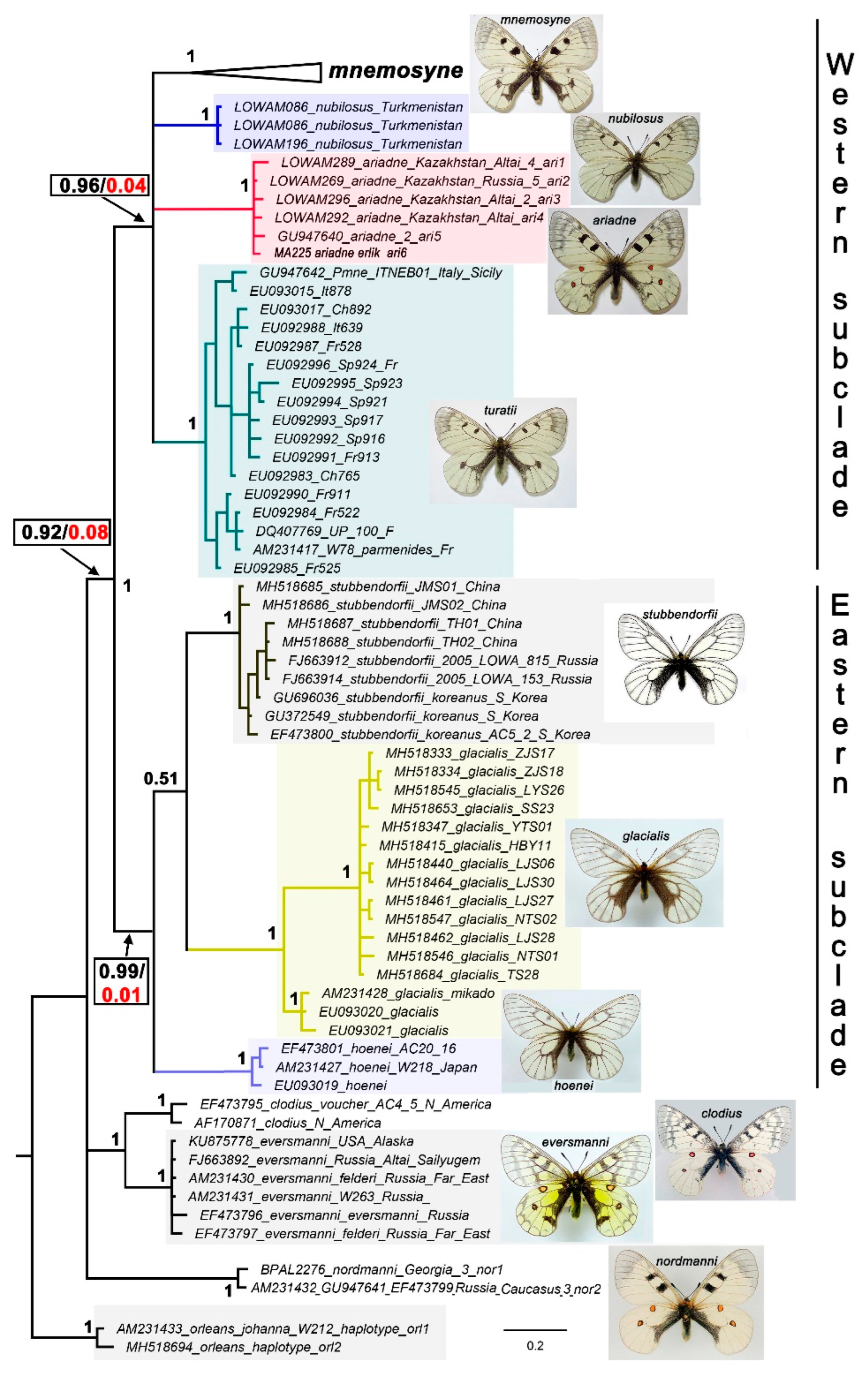
Figure 2.
A part or the Bayesian tree of the Parnassius (Driopa) species (Figure 1) showing the structure of P. mnemosyne sensu stricto. The tree is based on analysis of the mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (BPP) (higher than 0.5).
Figure 2.
A part or the Bayesian tree of the Parnassius (Driopa) species (Figure 1) showing the structure of P. mnemosyne sensu stricto. The tree is based on analysis of the mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (BPP) (higher than 0.5).
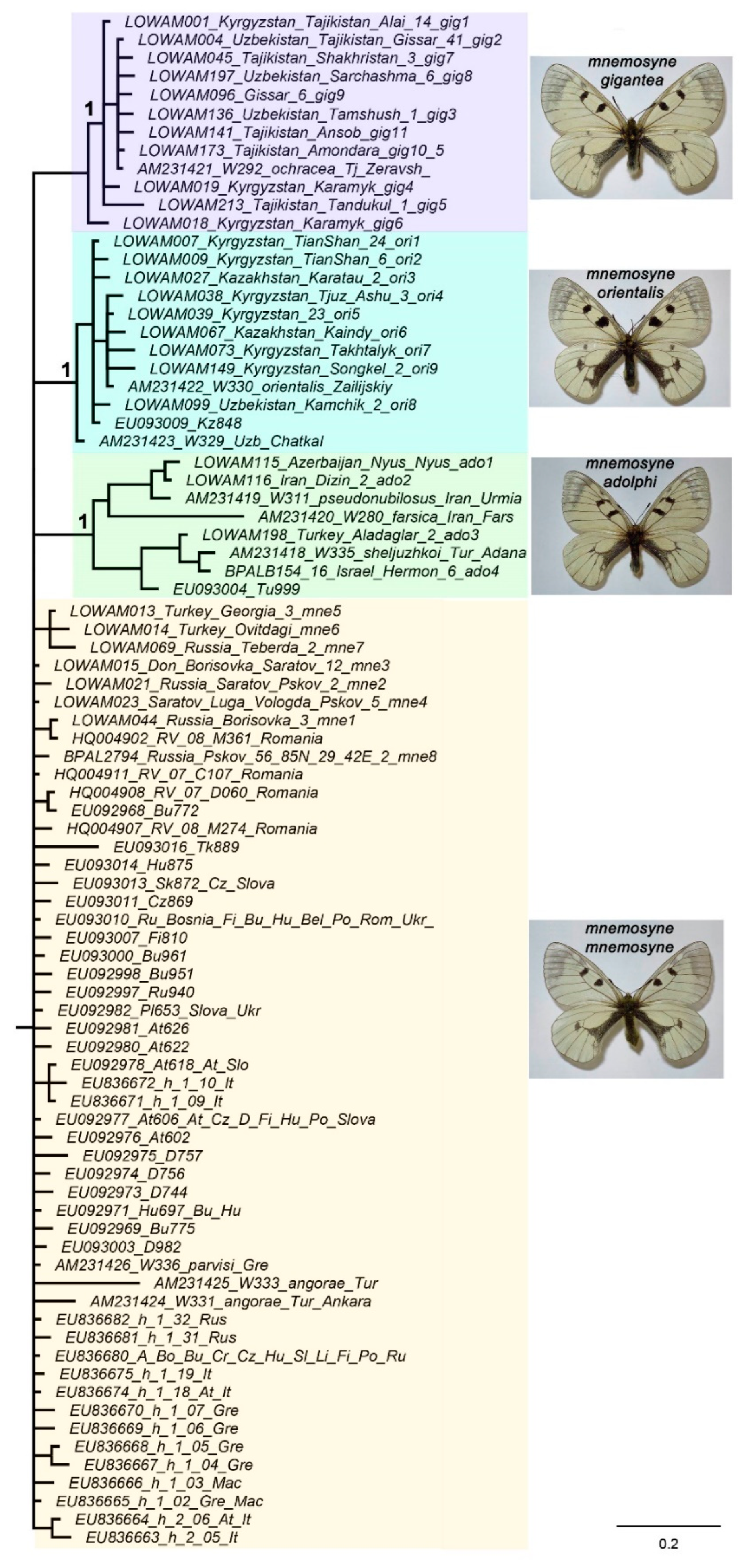
Figure 3.
Schematic representation of the ranges of P. mnemosyne, P. turatii, P. nubilosus stat. nov., and P. ariadne. Number 1 indicates geographic isolates of P. mnemosyne mnemosyne. Number 2 indicates geographic isolates of P. turatii. Number 3 indicates geographic isolates of P. mnemosyne adolphi.
Figure 3.
Schematic representation of the ranges of P. mnemosyne, P. turatii, P. nubilosus stat. nov., and P. ariadne. Number 1 indicates geographic isolates of P. mnemosyne mnemosyne. Number 2 indicates geographic isolates of P. turatii. Number 3 indicates geographic isolates of P. mnemosyne adolphi.
Figure 4.
Parnassius nubilosus (A-D), P. turatii (E, F) and P. mnemosyne mnemosyne (G,H). All specimens in coll. Zoological Institute, St. Petersburg. Scale = 1cm. A and B, P. nubilosus, males, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. C and D, P. nubilosus, females, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. E, P. turatii, male, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. F, P. turatii, female, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. G, P. mnemosyne mnemosyne, male, sample J178, Russia, Kaluga region, 27 May 1979, leg. I. Sokolov. H, P. mnemosyne mnemosyne, female, Russia, Penza region, Serdobsk, 20 May 2008, leg. Polumordvinov.
Figure 4.
Parnassius nubilosus (A-D), P. turatii (E, F) and P. mnemosyne mnemosyne (G,H). All specimens in coll. Zoological Institute, St. Petersburg. Scale = 1cm. A and B, P. nubilosus, males, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. C and D, P. nubilosus, females, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. E, P. turatii, male, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. F, P. turatii, female, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. G, P. mnemosyne mnemosyne, male, sample J178, Russia, Kaluga region, 27 May 1979, leg. I. Sokolov. H, P. mnemosyne mnemosyne, female, Russia, Penza region, Serdobsk, 20 May 2008, leg. Polumordvinov.

Figure 5.
Parnassius mnemosyne adolphi (A, B), P. mnemosyne gigantea (C,D), P. mnemosyne orientalis (E, F) and P. ariadne (G-I). All specimens in coll. Zoological Institute, St. Petersburg. Scale = 1cm. A, P. mnemosyne adolphi, male, sample J115, Azerbaijan, Nakhichevan, Njus-Njus vill., Sary-Dara Mt, 2300 m, 7 July 1984, V.A.Lukhtanov leg. B, P. mnemosyne adolphi, female, Transcaucasus, distr. Zangezur, loc. Litshk (prope Megri), 6 June 1910, E.Miller leg. C, P. mnemosyne gigantea, male, Uzbekistan, Gissar Mts, 50 km east of Shakhrisyabz, 1700-2300 m, 19 May 1994, N.Kandul leg. D, P. mnemosyne gigantea, female, [Kyrgyzstan], Alai Mts, Kok-Su, Kosh-Tjube, 3300 m, 28 July 1964, Bundel leg. E, P. mnemosyne orientalis, male, sample J112, Kyrgyzstan, Naryn-Too Range, 29-30 July 1995, V.Shchurov leg. F, P. mnemosyne orientalis, female, Kyrgyzstan, Kyrgyz Ala-Too Range, Uzyngyr, 2500 m, 13 July 1974, V.V. Dubatolov leg. G, P. ariadne, male, Kazakhstan, Saur Mts, Zhanaturmys, 1100 m, 4 June 1987, V.A.Lukhtanov leg. H, P. ariadne, male, Kazakhstan, S Altai, Kurtchum Mts, Kalinino, 22 June 1985, V.A.Lukhtanov leg. H, P. ariadne, female, Kazakhstan, S Altai, Markakol Lake, Urunkhaika, 1700 m, 24 June 1983, V.A.Lukhtanov leg.
Figure 5.
Parnassius mnemosyne adolphi (A, B), P. mnemosyne gigantea (C,D), P. mnemosyne orientalis (E, F) and P. ariadne (G-I). All specimens in coll. Zoological Institute, St. Petersburg. Scale = 1cm. A, P. mnemosyne adolphi, male, sample J115, Azerbaijan, Nakhichevan, Njus-Njus vill., Sary-Dara Mt, 2300 m, 7 July 1984, V.A.Lukhtanov leg. B, P. mnemosyne adolphi, female, Transcaucasus, distr. Zangezur, loc. Litshk (prope Megri), 6 June 1910, E.Miller leg. C, P. mnemosyne gigantea, male, Uzbekistan, Gissar Mts, 50 km east of Shakhrisyabz, 1700-2300 m, 19 May 1994, N.Kandul leg. D, P. mnemosyne gigantea, female, [Kyrgyzstan], Alai Mts, Kok-Su, Kosh-Tjube, 3300 m, 28 July 1964, Bundel leg. E, P. mnemosyne orientalis, male, sample J112, Kyrgyzstan, Naryn-Too Range, 29-30 July 1995, V.Shchurov leg. F, P. mnemosyne orientalis, female, Kyrgyzstan, Kyrgyz Ala-Too Range, Uzyngyr, 2500 m, 13 July 1974, V.V. Dubatolov leg. G, P. ariadne, male, Kazakhstan, Saur Mts, Zhanaturmys, 1100 m, 4 June 1987, V.A.Lukhtanov leg. H, P. ariadne, male, Kazakhstan, S Altai, Kurtchum Mts, Kalinino, 22 June 1985, V.A.Lukhtanov leg. H, P. ariadne, female, Kazakhstan, S Altai, Markakol Lake, Urunkhaika, 1700 m, 24 June 1983, V.A.Lukhtanov leg.

Figure 6.
Male genitalia of P. nubilosus (A, B, sample J086) and P. mnemosyne adolphi (A, C, sample J115). A, C, lateral view, left valve removed. B, D, left valva. A,B, Turkmenistan, Kopet-Dagh, Ai-Dere, 1 April 1979, Ju.Ya.Sokolova leg. C, D, Azerbaijan, Nakhichevan, Njus-Njus vill., Sary-Dara Mt, 2300 m, 7 July 1984, V.A.Lukhtanov leg.
Figure 6.
Male genitalia of P. nubilosus (A, B, sample J086) and P. mnemosyne adolphi (A, C, sample J115). A, C, lateral view, left valve removed. B, D, left valva. A,B, Turkmenistan, Kopet-Dagh, Ai-Dere, 1 April 1979, Ju.Ya.Sokolova leg. C, D, Azerbaijan, Nakhichevan, Njus-Njus vill., Sary-Dara Mt, 2300 m, 7 July 1984, V.A.Lukhtanov leg.
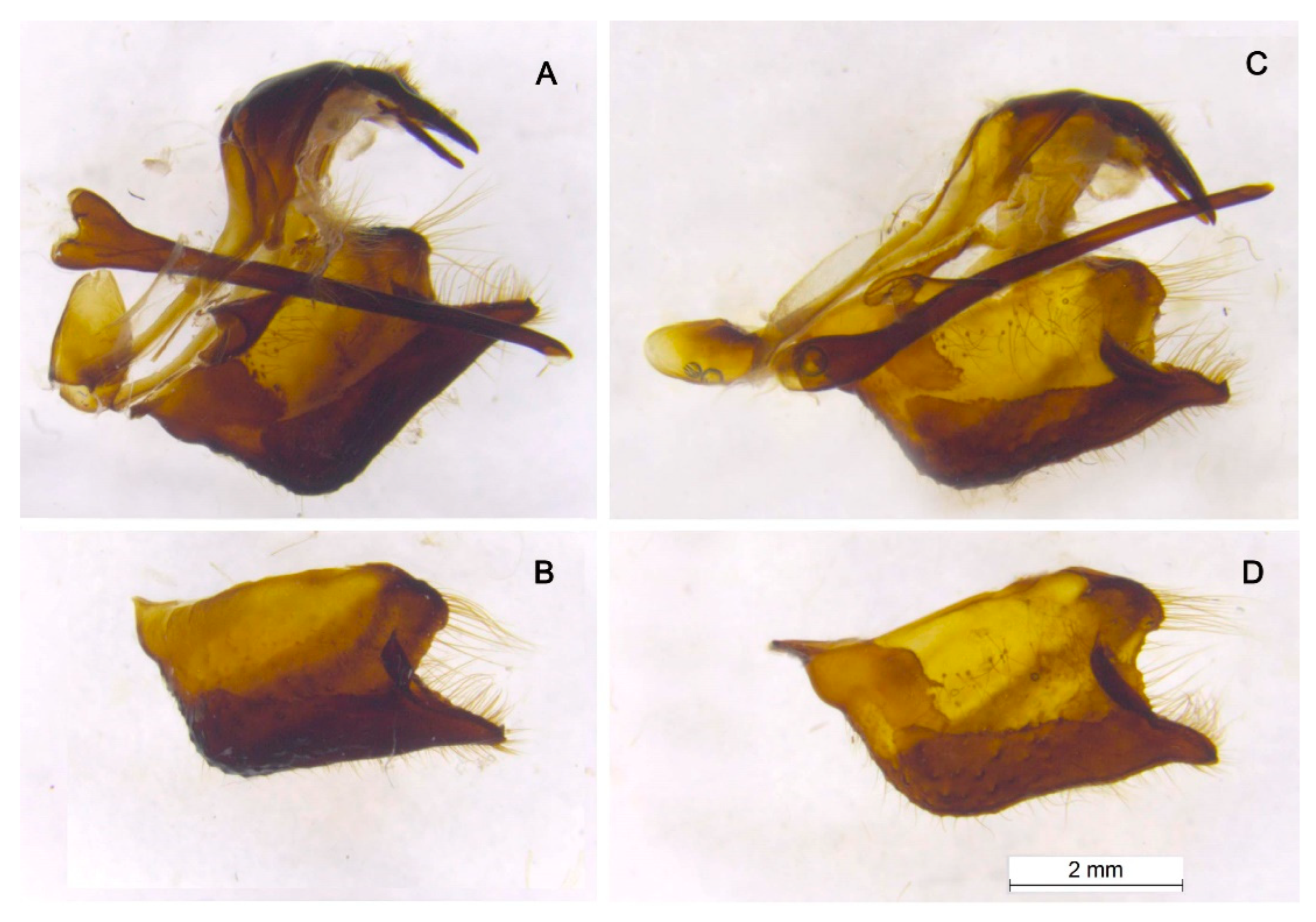
Figure 7.
Sphragis is females of P. nubilosus (A, B), P. mnemosyne orientalis (C), P. mnemosyne adolphi (D), P. mnemosyne gigantea (E), P. mnemosyne mnemosyne (F), P. turatii (G) and P. ariadne (H). All specimens in coll. Zoological Institute, St. Petersburg. A, B, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. C, Kyrgyzstan, Kyrgyz Ala-Too Range, Uzyngyr, 2500 m, 13 July 1974, V.V. Dubatolov leg. D, Transcaucasus, distr. Zangezur, loc. Litshk (prope Megri), 6 June 1910, E.Miller leg. E, [Kyrgyzstan], Alai Mts, Kok-Su, Kosh-Tjube, 3300 m, 28 July 1964, Bundel leg. F, Russia, Penza region, Serdobsk, 20 May 2008, leg. Polumordvinov. G, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. H, Kazakhstan, S Altai, Kurtchum Mts, Kalinino, 7 June 1986, V.A.Lukhtanov leg.
Figure 7.
Sphragis is females of P. nubilosus (A, B), P. mnemosyne orientalis (C), P. mnemosyne adolphi (D), P. mnemosyne gigantea (E), P. mnemosyne mnemosyne (F), P. turatii (G) and P. ariadne (H). All specimens in coll. Zoological Institute, St. Petersburg. A, B, Turkmenistan, Kopetdagh, Dushak Mt, 6 June 1986, V.Dubatolov leg. C, Kyrgyzstan, Kyrgyz Ala-Too Range, Uzyngyr, 2500 m, 13 July 1974, V.V. Dubatolov leg. D, Transcaucasus, distr. Zangezur, loc. Litshk (prope Megri), 6 June 1910, E.Miller leg. E, [Kyrgyzstan], Alai Mts, Kok-Su, Kosh-Tjube, 3300 m, 28 July 1964, Bundel leg. F, Russia, Penza region, Serdobsk, 20 May 2008, leg. Polumordvinov. G, Italy, Sicilia, Madonie, 1200’, 15 July 1910, coll. Krüger, mus. Turati E, coll. Avinov. H, Kazakhstan, S Altai, Kurtchum Mts, Kalinino, 7 June 1986, V.A.Lukhtanov leg.
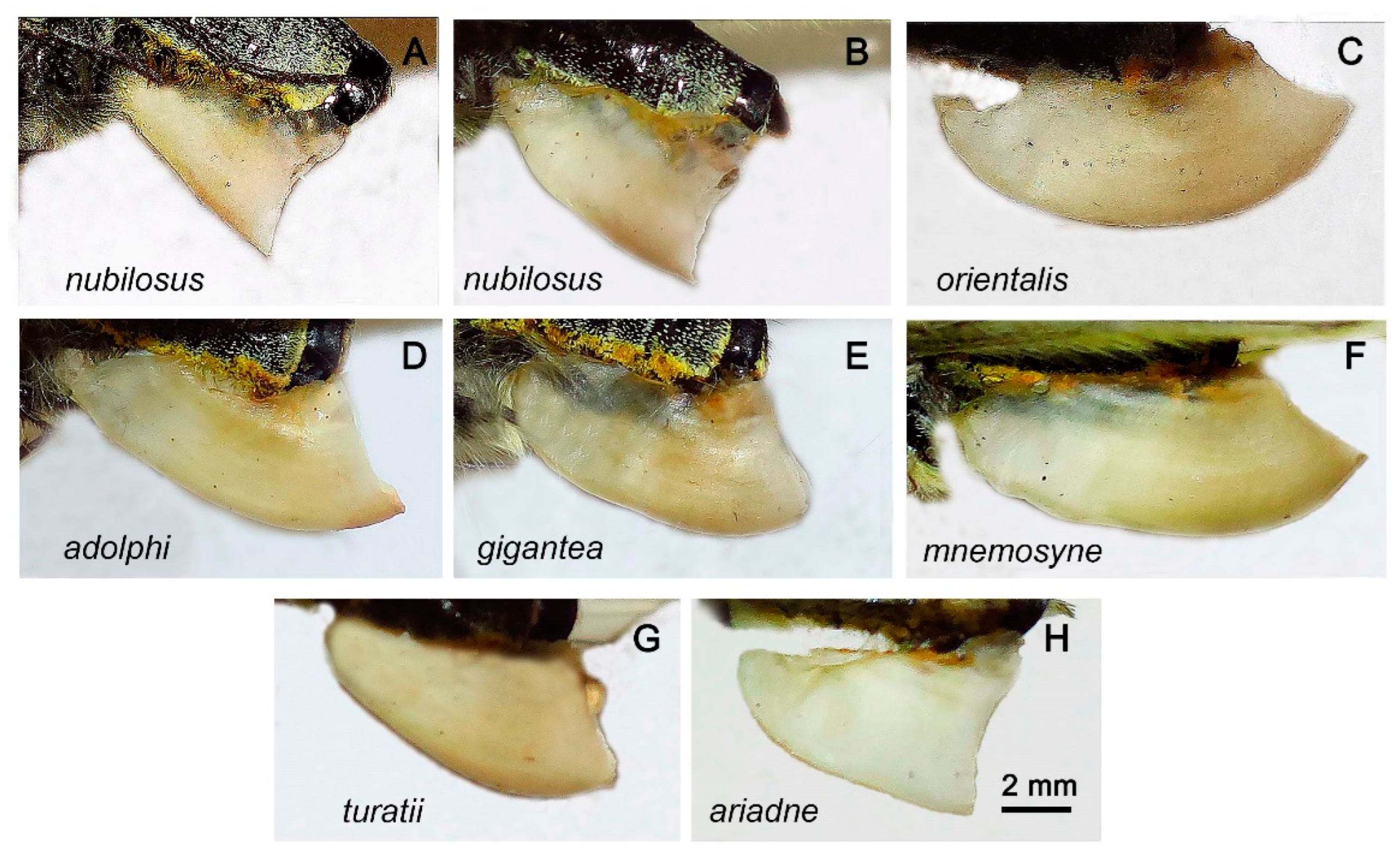
Figure 8.
Lectotype of Parnassius mnemosyne var. nubilosus Christoph, 1873 (Zoological Institute, Russian Academy of Sciences, St. Petersburg. Scale = 1 cm.
Figure 8.
Lectotype of Parnassius mnemosyne var. nubilosus Christoph, 1873 (Zoological Institute, Russian Academy of Sciences, St. Petersburg. Scale = 1 cm.

Figure 9.
Lectotype of Doritis clarius Eversmann, 1843 [= Parnassius ariadne (Lederer, 1853] (Zoological Institute, Russian Academy of Sciences, St. Petersburg, catalogue number: INS_LEP_0000666). Scale = 1 cm.
Figure 9.
Lectotype of Doritis clarius Eversmann, 1843 [= Parnassius ariadne (Lederer, 1853] (Zoological Institute, Russian Academy of Sciences, St. Petersburg, catalogue number: INS_LEP_0000666). Scale = 1 cm.


Table 1.
List of the studied samples and obtained COI sequences.
| BOLD ID | GenBank ID | Haplotype | Identification | Country | Region |
| LOWAM001-08 | OR822545 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM002-08 | OR822537 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM003-08 | OR822507 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM004-08 | OR822401 | gig2 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM005-08 | OR822544 | gig2 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM006-08 | OR822581 | gig2 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM007-08 | OR822585 | ori1 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM008-08 | OR822475 | ori1 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM009-08 | OR822518 | ori2 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM010-08 | OR822548 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM011-08 | OR822588 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM012-08 | OR822423 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM013-08 | OR822478 | mne5 | mnemosyne | Turkey | Ovitdagi Gecidi |
| LOWAM014-08 | OR822546 | mne6 | mnemosyne | Turkey | Ovitdagi Gecidi |
| LOWAM015-08 | OR822577 | mne3 | mnemosyne | Russia | Don River |
| LOWAM016-08 | OR822394 | mne3 | mnemosyne | Russia | Don River |
| LOWAM017-08 | OR822413 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM018-08 | OR822525 | gig6 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM019-08 | OR822524 | gig4 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM020-08 | OR822403 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM021-08 | OR822586 | mne2 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM022-08 | OR822563 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM023-08 | OR822554 | mne4 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM024-08 | OR822454 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM025-08 | OR822543 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM026-08 | OR822532 | gig2 | mnemosyne | Uzbekistan | W Gissar Mts |
| LOWAM027-08 | OR822515 | ori3 | mnemosyne | Kazakhstan | Karatau Mts |
| LOWAM028-08 | OR822520 | ori3 | mnemosyne | Kazakhstan | Karatau Mts |
| LOWAM029-08 | OR822502 | gig2 | mnemosyne | Tajikistan | Taribak |
| LOWAM030-08 | OR822471 | gig2 | mnemosyne | Tajikistan | Taribak |
| LOWAM031-08 | OR822490 | gig2 | mnemosyne | Tajikistan | Taribak |
| LOWAM032-08 | OR822523 | gig2 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM033-08 | OR822421 | gig2 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM034-08 | OR822419 | gig2 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM036-08 | OR822448 | mne2 | mnemosyne | Russia | Pskovskaya Oblast |
| LOWAM037-08 | OR822459 | mne4 | mnemosyne | Russia | Pskovskaya Oblast |
| LOWAM038-08 | OR822452 | ori4 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM039-08 | OR822428 | ori5 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM040-08 | OR822506 | ori4 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM041-08 | OR822406 | ori1 | mnemosyne | Kyrgyzstan | Alabel Pass |
| LOWAM042-08 | OR822465 | mne3 | mnemosyne | Russia | Belgorodskaya oblast |
| LOWAM043-08 | OR822426 | mne3 | mnemosyne | Russia | Belgorodskaya oblast |
| LOWAM044-08 | OR822503 | mne1 | mnemosyne | Russia | Belgorodskaya oblast |
| LOWAM045-08 | OR822481 | gig7 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM046-08 | OR822493 | gig7 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM047-08 | OR822538 | gig7 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM048-08 | OR822444 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM049-08 | OR822411 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM050-08 | OR822408 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM051-08 | OR822590 | gig8 | mnemosyne | Tajikistan | Revad |
| LOWAM052-08 | OR822477 | gig2 | mnemosyne | Tajikistan | |
| LOWAM055-08 | OR822420 | gig8 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM056-08 | OR822404 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM057-08 | OR822480 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM058-08 | OR822508 | ori1 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM059-08 | OR822460 | ori1 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM060-08 | OR822418 | ori1 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM061-08 | OR822425 | ori1 | mnemosyne | Kazakhstan | Saryaigyr |
| LOWAM062-08 | OR822587 | gig2 | mnemosyne | Tajikistan | Gissar Mts (West) |
| LOWAM063-08 | OR822469 | gig8 | mnemosyne | Tajikistan | Gissar Mts (West) |
| LOWAM064-08 | OR822447 | ori9 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM065-08 | OR822517 | ori9 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM066-08 | OR822552 | ori9 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM067-08 | OR822415 | ori6 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM069-08 | OR822551 | mne7 | mnemosyne | Russia | W Caucasus |
| LOWAM070-08 | OR822533 | mne5 | mnemosyne | Russia | W Caucasus |
| LOWAM072-08 | OR822571 | ori1 | mnemosyne | Kyrgyzstan | Chatkalsky Khrebet |
| LOWAM073-08 | OR822453 | ori7 | mnemosyne | Kyrgyzstan | Takhtalyk |
| LOWAM074-08 | OR822491 | ori5 | mnemosyne | Kyrgyzstan | Naryn-Too Mts |
| LOWAM075-08 | OR822570 | ori1 | mnemosyne | Kyrgyzstan | Naryn-Too Mts |
| LOWAM076-08 | OR822498 | ori5 | mnemosyne | Kyrgyzstan | Ak-Muz |
| LOWAM077-08 | OR822557 | ori2 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM078-08 | OR822516 | ori5 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM079-08 | OR822439 | ori9 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM080-08 | OR822500 | ori5 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM081-08 | OR822463 | ori5 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM082-08 | OR822436 | ori5 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM086-08 | OR822567 | nub1 | nubilosus | Turkmenistan | Kopetdagh |
| LOWAM087-08 | OR822476 | gig8 | mnemosyne | Uzbekistan | Gissar Mts (West) |
| LOWAM088-08 | OR822432 | gig2 | mnemosyne | Uzbekistan | Gissar Mts (West) |
| LOWAM089-08 | OR822505 | gig2 | mnemosyne | Uzbekistan | Gissar Mts (West) |
| LOWAM090-08 | OR822528 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM091-08 | OR822501 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM092-08 | OR822510 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM093-08 | OR822565 | ori5 | mnemosyne | Kyrgyzstan | between Alabel and Tjuz-Ashu passes |
| LOWAM094-08 | OR822391 | ori5 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM095-08 | OR822527 | ori5 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM096-08 | OR822499 | gig9 | mnemosyne | Tajikistan | Gissar Mts (SW) |
| LOWAM097-08 | OR822574 | gig9 | mnemosyne | Tajikistan | Gissar Mts (SW) |
| LOWAM098-08 | OR822399 | gig9 | mnemosyne | Tajikistan | Gissar Mts (SW) |
| LOWAM099-08 | OR822461 | ori8 | mnemosyne | Uzbekistan | Kuraminsky Khrebet |
| LOWAM100-08 | OR822560 | ori5 | mnemosyne | Kyrgyzstan | Ferganski Khrebet |
| LOWAM101-08 | OR822450 | ori5 | mnemosyne | Kyrgyzstan | Ferganski Khrebet |
| LOWAM102-08 | OR822579 | ori5 | mnemosyne | Kyrgyzstan | Ferganski Khrebet |
| LOWAM103-08 | OR822550 | ori1 | mnemosyne | Kazakhstan | Turaigyr |
| LOWAM104-08 | OR822529 | ori1 | mnemosyne | Kazakhstan | Turaigyr |
| LOWAM105-08 | OR822561 | ori1 | mnemosyne | Kazakhstan | Turaigyr |
| LOWAM106-08 | OR822485 | ori8 | mnemosyne | Uzbekistan | S of Tashkent |
| LOWAM107-08 | OR822564 | gig2 | mnemosyne | Uzbekistan | Seravshaski Khrebet |
| LOWAM111-08 | OR822431 | ori5 | mnemosyne | Kyrgyzstan | Naryn-Too Mts |
| LOWAM112-08 | OR822430 | ori5 | mnemosyne | Kyrgyzstan | Naryn-Too Mts |
| LOWAM115-08 | OR822592 | ado1 | mnemosyne | Azerbaijan | Nakhichevan |
| LOWAM116-08 | OR822566 | ado2 | mnemosyne | Iran | Elburs Mts |
| LOWAM117-08 | OR822397 | ado2 | mnemosyne | Iran | Elburs Mts |
| LOWAM118-08 | OR822396 | mne1 | mnemosyne | Russia | Belgorod Region |
| LOWAM119-08 | OR822526 | mne1 | mnemosyne | Russia | Belgorod Region |
| LOWAM120-08 | OR822486 | mne3 | mnemosyne | Russia | Belgorod Region |
| LOWAM121-08 | OR822416 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM122-08 | OR822542 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM123-08 | OR822553 | mne3 | mnemosyne | Russia | Saratovskaya Oblast |
| LOWAM124-08 | OR822479 | ori5 | mnemosyne | Kyrgyzstan | Ak-Muz |
| LOWAM125-08 | OR822445 | ori5 | mnemosyne | Kyrgyzstan | Ak-Muz |
| LOWAM126-08 | OR822539 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM127-08 | OR822511 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM128-08 | OR822578 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM129-08 | OR822457 | gig8 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM130-08 | OR822575 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM131-08 | OR822562 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM132-08 | OR822496 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM133-08 | OR822488 | gig2 | mnemosyne | Uzbekistan | Gissar Mts West |
| LOWAM134-08 | OR822559 | gig2 | mnemosyne | Uzbekistan | Gissar Mts West |
| LOWAM135-08 | OR822462 | gig9 | mnemosyne | Uzbekistan | Gissar Mts West |
| LOWAM136-08 | OR822451 | gig3 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM137-08 | OR822449 | gig2 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM138-08 | OR822556 | gig9 | mnemosyne | Uzbekistan | Gissar Mts |
| LOWAM139-08 | OR822434 | gig9 | mnemosyne | Tajikistan | Gissar Mts SW |
| LOWAM140-08 | OR822473 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM141-08 | OR822521 | gig11 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM142-08 | OR822417 | ori2 | mnemosyne | Kyrgyzstan | Modatoo Mts |
| LOWAM143-08 | OR822402 | ori1 | mnemosyne | Kyrgyzstan | Modatoo Mts |
| LOWAM144-08 | OR822582 | ori1 | mnemosyne | Kyrgyzstan | Modatoo Mts |
| LOWAM145-08 | OR822458 | ori2 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM146-08 | OR822536 | ori2 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM147-08 | OR822519 | ori2 | mnemosyne | Kyrgyzstan | Moldatoo Mts |
| LOWAM148-08 | OR822456 | ori5 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM149-08 | OR822464 | ori9 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM150-08 | OR822422 | ori5 | mnemosyne | Kyrgyzstan | Songkel Lake |
| LOWAM151-08 | OR822433 | ori4 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM152-08 | OR822409 | ori1 | mnemosyne | Kyrgyzstan | Kirgizsky Khrebet |
| LOWAM153-08 | OR822443 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM154-08 | OR822535 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM155-08 | OR822541 | gig1 | mnemosyne | Kyrgyzstan | Alai Mts |
| LOWAM156-08 | OR822522 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM157-08 | OR822410 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM158-08 | OR822470 | ori1 | mnemosyne | Kyrgyzstan | Chychkan |
| LOWAM159-08 | OR822509 | ori5 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM160-08 | OR822455 | ori5 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM161-08 | OR822580 | ori5 | mnemosyne | Kazakhstan | Kirgizsky Khrebet |
| LOWAM162-08 | OR822555 | ori2 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM163-08 | OR822483 | ori1 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM164-08 | OR822393 | ori1 | mnemosyne | Kazakhstan | Karzhantau Mts |
| LOWAM165-08 | OR822395 | gig2 | mnemosyne | Tajikistan | Taribak |
| LOWAM166-08 | OR822583 | gig2 | mnemosyne | Tajikistan | Taribak |
| LOWAM167-08 | OR822474 | gig10 | mnemosyne | Tajikistan | Taribak |
| LOWAM168-08 | OR822487 | gig2 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM169-08 | OR822576 | gig2 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM170-08 | OR822497 | gig10 | mnemosyne | Uzbekistan | Nuratau Mts |
| LOWAM171-08 | OR822424 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM172-08 | OR822429 | gig2 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM173-08 | OR822530 | gig10 | mnemosyne | Tajikistan | Turkestanski Mts |
| LOWAM196-09 | OR822494 | nub1 | nubilosus | Turkmenistan | West Kopet-Dagh |
| LOWAM197-09 | OR822435 | nub1 | nubilosus | Turkmenistan | West Kopet-Dagh |
| LOWAM198-09 | OR822540 | ado3 | mnemosyne | Turkey | Taurus (gory Tavr) |
| LOWAM199-09 | OR822513 | ado3 | mnemosyne | Turkey | Taurus (gory Tavr) |
| LOWAM200-09 | OR822584 | mne5 | mnemosyne | Georgia | Mestia |
| LOWAM201-09 | OR822531 | mne5 | mnemosyne | Georgia | Mestia |
| LOWAM202-09 | OR822467 | mne4 | mnemosyne | Russia | St. Petersburg region |
| LOWAM203-09 | OR822414 | mne4 | mnemosyne | Russia | St. Petersburg region |
| LOWAM205-09 | OR822512 | ori1 | mnemosyne | Kyrgyzstan | Kungey-Alatoo Mts |
| LOWAM206-09 | OR822441 | mne4 | mnemosyne | Russia | Vologodskaya oblast |
| LOWAM207-09 | OR822534 | gig10 | mnemosyne | Tajikistan | Obihingou |
| LOWAM208-09 | OR822504 | gig8 | mnemosyne | Tajikistan | Peter The Great Mts |
| LOWAM209-09 | OR822412 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM210-09 | OR822440 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM212-09 | OR822427 | gig2 | mnemosyne | Tajikistan | Gissar Mts |
| LOWAM213-09 | OR822495 | gig5 | mnemosyne | Tajikistan | Alai Mts |
| BPALB154-16 | OR822438 | ado4 | mnemosyne | Israel | Hermon |
| BPALB230-17 | OR822392 | gig2 | mnemosyne | Tajikistan | |
| BPALB245-17 | OR822591 | gig2 | mnemosyne | Tajikistan | |
| BPALB258-17 | OR822442 | gig1 | mnemosyne | Tajikistan | |
| BPALB369-17 | OR822468 | gig1 | mnemosyne | Tajikistan | |
| BPALB389-17 | OR822484 | gig1 | mnemosyne | Tajikistan | |
| BPAL2225-13 | OR822589 | ado4 | mnemosyne | Israel | N. Israel |
| BPAL2226-13 | OR822405 | ado4 | mnemosyne | Israel | N. Israel |
| BPAL2794-15 | OR822572 | mne8 | mnemosyne | Russia | Pskov |
| BPAL2795-15 | OR822568 | mne8 | mnemosyne | Russia | Pskov |
| BPAL3195-16 | OR822437 | ado4 | mnemosyne | Israel | Hermon |
| BPAL3196-16 | OR822489 | ado4 | mnemosyne | Israel | Hermon |
| BPAL3358-16 | OR822547 | ado4 | mnemosyne | Israel | Hermon Mt |
| BPAL2276-13 | OR822466 | nor1 | nordmanni | Georgia | Adzharia |
| BPAL2277-13 | OR822573 | nor1 | nordmanni | Georgia | Adzharia |
| BPAL2278-13 | OR822398 | nor1 | nordmanni | Georgia | Adzharia |
| LOWAM269-11 | OR822482 | ari2 | ariadne | Kazakhstan | West Altai |
| LOWAM273-11 | OR822514 | ari2 | ariadne | Kazakhstan | West Altai |
| LOWAM287-11 | OR822407 | ari1 | ariadne | Kazakhstan | Altai |
| LOWAM288-11 | OR822446 | ari1 | ariadne | Kazakhstan | Altai |
| LOWAM289-11 | OR822472 | ari1 | ariadne | Kazakhstan | Altai |
| LOWAM290-11 | OR822400 | ari1 | ariadne | Kazakhstan | Altai |
| LOWAM292-11 | OR822558 | ari4 | ariadne | Kazakhstan | Altai |
| LOWAM295-11 | OR822492 | ari3 | ariadne | Kazakhstan | Altai |
| LOWAM296-11 | OR822569 | ari3 | ariadne | Kazakhstan | Altai |
| LOWAM301-11 | OR822549 | ari2 | ariadne clarus | Kazakhstan | Saur Mts |
| MA225 | ari6 | ariadne erlik | Russia | N of Kosh-Agach |
Table 2.
Minimum COI p-distances (%) between the taxa of the subgenus Driopa.
| ori | gig | mne | ado | nub | tur | ari | stu | hoe | gla | clo | eve | nor | |
| orientalis | 0 | ||||||||||||
| gigantea | 1.22 | ||||||||||||
| mnemosyne | 0.92 | 0.92 | |||||||||||
| adolphi | 2.45 | 2.45 | 2.14 | ||||||||||
| nubilosus | 2.91 | 2.29 | 2.60 | 3.52 | |||||||||
| turatii | 3.24 | 2.93 | 2.78 | 3.70 | 2.78 | ||||||||
| ariadne | 3.37 | 3.67 | 3.36 | 3.06 | 3.52 | 3.55 | |||||||
| stubbendorfii | 4.74 | 4.43 | 4.74 | 5.35 | 4.28 | 4.17 | 5.05 | ||||||
| hoenei | 5.09 | 5.09 | 4.78 | 5.25 | 3.70 | 4.32 | 5.25 | 4.01 | |||||
| glacialis | 6.29 | 5.98 | 6.29 | 6.76 | 5.66 | 6.51 | 7.08 | 4.38 | 6.03 | ||||
| clodius | 3.52 | 3.52 | 3.82 | 3.82 | 3.98 | 3.86 | 3.82 | 3.81 | 4.78 | 5.94 | |||
| eversmanni | 3.98 | 3.67 | 3.67 | 3.98 | 3.52 | 3.86 | 3.98 | 3.65 | 4.46 | 5.48 | 1.98 | ||
| nordmanni | 5.58 | 5.89 | 5.58 | 6.36 | 4.81 | 5.43 | 5.43 | 5.12 | 5.74 | 7.18 | 4.34 | 4.65 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



