
Submitted:
22 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
A three-year survey was conducted to estimate the incidence of grapevine trunk diseases (GTDs) in Greece and identify fungi associated with the disease complex. In total, 310 vineyards in different geographical regions in northern, central, and southern Greece were surveyed, and 533 fungal strains were isolated from diseased vines. Morphological, physiological and molecular (5.8S rRNA gene-ITS sequencing) analyses revealed that isolates belonged to 35 distinct fungal genera, including well-known (e.g. Botryosphaeria sp, Diaporthe spp., Eutypa sp., Diplodia sp., Fomitiporia sp., Phaeoacremonium spp., Phaeomoniella sp.) and lesser-known (e.g. Neosetophoma sp., Seimatosporium sp., Didymosphaeria sp., Kalmusia sp.) grapevine wood inhabitants. GTDs-inducing population structure differed significantly among the discrete geographical zones. Phaeomoniella chlamydospora (26.62%, n=70), Diaporthe spp. (18.25%, n=48) and F. mediterranea (10.27%, n=27) were the most prevalent in Heraklion, whereas D. seriata, Alternaria spp., P. chlamydospora and Fusarium spp. were predominant in Nemea (central Greece). In Amyntaio and Kavala (northern Greece), D. seriata was the most frequently isolated species (>50% frequency). Multi-genes (rDNA-ITS, LSU, tef1-α, tub2, act) sequencing of selected isolates, followed by pathogenicity tests revealed that Neosetophoma italica, Seimatosporium vitis, Didymosphaeria variabile and Kalmusia variispora caused wood infection with the former being the most virulent. To the best of our knowledge, this is the first report of N. italica associated with GTDs worldwide. This is also the first record of K. variispora, S. vitis and D. variabile associated with wood infection of grapevine in Greece. The potential associations of disease indices with vine age, cultivar, GTD-associated population structure and the prevailed meteorological conditions in different viticultural zones in Greece are presented and discussed.
Keywords:
Kalmusia variispora
; Neosetophoma italica
; Paraconiothyrium variabile
; Seimatosporium vitis
; meteorological conditions
; wood inhabitant
1. Introduction
Grapevine (Vitis vinifera L.) is one of the most commonly cultivated plant species in several countries like Australia, France, Greece, Italy, South Africa, Spain, and USA, and has a significant social, economic, and environmental impact [1]. Greece is belonging to the top 20th leading grapevine-growing countries of the world in terms of production quantity [2], while viticulture is essential for the economy and rural development of the country, covering an area of 89.230 ha with an annual yield of 818.860 tons in 2021 [3]
In recent decades, grapevine trunk diseases (GTDs) have emerged as the most severe threat to viticulture sustainability worldwide [4]. GTDs are a cluster of diseases incited by a vast number of fungal pathogens (mostly ascomycetes) that colonize and infect wood tissues, causing internal symptoms such as wood streaking and necrosis, vascular browning, and white rot [5,6]. Consequential external symptoms include leaf chlorosis, yellowing and wilting, wood cankers, and dieback of spurs, canes, and cordons, leading to stunted plant growth, decline, reduced longevity and productivity, and occasionally plant death.
Increased incidence of GTDs globally has been attributed to several factors like the withdrawal of effective chemicals (i.e. sodium arsenate, benzimidazole fungicides), alterations in cultivation practices, globalization in the dissemination of plant material contaminated with latent pathogens, and climate change [7-10]. In particular, climate change and global warming can increase plant stress and make vines vulnerable to various biotic stresses, including GTD-inciting pathogens, in a synergistically interactive manner [10-11]. On the other hand, rapid alterations of wet and dry cycles along with extremely low and/or high temperatures, as a consequence of climate change, may favor pathogen dispersal from their original hosts and adaption to Vitis, resulting in the emergence of new GTD-associated fungal species as it has been speculated for other woody hosts [8,12,13]. This possibly explains the increased number of fungal pathogens identified as causal agents of GTDs nowadays compared to the past. Additionally, the availability of high-resolution molecular tools has also provided a deeper insight into the GTD-associated microbiome in recent years compared to previous ones and has enriched, so far, our knowledge of microbial species that contribute to the GTD complex [14-18].
To date, more than 140 fungal species belonging to 35 genera have been involved in GTDs complex worldwide, although the pathogenic potential has not been confirmed for all of them in pathogenicity trials [7,19-21]. Interestingly, recent surveys conducted in marginal temperate regions in Cyprus and southern Italy revealed several lesser-known and/or novel species associated with GTDs, like Cadophora luteo-olivacaea, Colletotrichum fioriniae, Seimatosporium vitis-vinifera, Seim. cyprium sp. nov., Sporocadus kurdistanicus, Spo. rosigena and Truncatella angustata, and proved their pathogenic potential [20-21].
Spatial determination of the phytopathogenic status of GTDs in grapevine-growing countries following a regional-scale mapping is important for planning and implementing effective management practices targeting specific diseases. Furthermore, accurate estimation of disease parameters in distinct viticultural zones may provide valuable information on the susceptibility level for widely used genotypes and thus, improved disease management through risk assessment [13].
Although GTDs incidence and implicated microorganisms have extensively been investigated for most grapevine-growing countries, only crude estimations have been conducted in Greece. In particular, Rumbos and Rumbou [22], determined specific fungal genera (i.e. Phaeomoniella, Fomitiporia, Stereum, Phaeoacremonium, Cylindrocarpon, and Botryosphaeria) isolated from different-aged vine samples in Greece following a culture-depended and morphological identification approach. Due to their low isolation frequency, authors suggested incorrectly that these fungi could not be the cause of young grapevine decline. In a recent study, Bekris et al. [14] determined the fungal and bacterial microbiome in symptomatic and asymptomatic grapevines of three important cultivars grown in geographically distinct viticultural zones in Greece following a metagenomic approach and revealed several lesser-known fungal genera related to GTD affected vines. Both studies are inadequately informative about the incidence and severity of GTDs as well as the phytopathogenic role of certain fungal species identified in the GTD-associated microbiome in the country. Therefore, the main objectives of the present study were to: i) estimate disease parameters and identify fungi associated with GTDs in Greece following a culture-dependent approach, ii) determine the population structure of GTDs-inciting fungi in distinct viticultural zones, iii) investigate the phytopathogenic potential of lesser-known fungal species on grapevine, and iv) identify the potential associations of disease indices with vine age, genotype and the prevailed meteorological conditions in discrete viticultural zones.
2. Materials and Methods
2.1. Field Surveys and Disease Incidence Assessment
In total, 310 vineyards in three (3) geographically distinct viticultural zones in northern (n=234), central (n=28), and southern (n=48) Greece were surveyed during July and August of the years 2018-2020. The number of vineyards surveyed for the assessment of GTDs incidence in each grapevine-growing region and their age scale are shown in Table 1. The geographical distribution of the surveyed locations is shown in Supplementary Figure S1. For each vineyard, either all or at least 100 vines in randomly selected lines in the vineyard were surveyed, and the percentage of diseased vines (including dead and non-dead vines), symptomatic but non-dead vines, and dead/apoplectic vines, was measured. Additional information about each vineyard's age, genotype (cultivar/rootstock), training-pruning system, and geographical position was also recorded.
2.2. Sampling and fungal isolation
Wood samples from 41 vineyards in southern (Heraklion), 16 vineyards in central (Nemea), and 19 vineyards in northern Greece (9 vineyards in Amyntaio and 10 vineyards in Kavala) were collected and analyzed. In each vineyard, 3 to 9 representative grapevines with typical GTDs symptoms were sampled. Overall, 252 vines were sampled from northern (n=60), central (n=69), and southern (n=129) Greece. Wood samples from the trunk and the cordon of each vine (approximately 30-cm-length each) were cut and visually inspected to verify the occurrence of internal wood symptoms. Samples were transferred to the laboratory and cut crosswise and lengthwise, the phloem was removed, and wood fragments (approx. 10-cm-length) were surface sterilized by soaking them in 93% ethanol and passing through a flame, thrice. Xylem chips taken from symptomatic wood tissue were aseptically placed onto acidified potato dextrose agar (PDA, Merck, Germany). Plates were incubated at 24±0.2 °C in the dark and observed daily for at least two weeks. The emerging fungal colonies that grew out of tissue excisions were examined visually and under a light microscope, and the fungi were transferred onto new PDA plates. For short-term storage, all fungal isolates were maintained on PDA at 4 oC, whereas for long-term they were stored at -80 oC in a 25% (v/v) aqueous glycerol solution.
2.3. Morphological and Cultural Characterization of Isolates
To estimate mycelial growth of fungal isolates, mycelial-colonized PDA agar discs (5 mm in diameter) was transferred into the center of new 92 mm-diameter PDA plates (one disc per plate, three per isolate). Plates were incubated at 24 oC in the dark, and the colonies’ diameter was measured periodically for up to three weeks or stopped earlier when the fungus completely covered the plate surface. The growth rate of fungal isolates was expressed in mm/day. At the end of the incubation period, colony characteristics (colour, mycelium, colony texture, and shape) were observed. Dimensions of available conidia (30 readings per isolate), carpophores, and hyphal features (color, shape, presence or absence of septum, clump, and chlamydospores) were also recorded.
2.4. DNA Extraction, PCR Amplification and Sequencing
Molecular identification of fungal isolates was carried out by extracting fungal DNA and sequencing their internal transcribed spacer regions of ribosomal DNA (rDNA-ITS) gene. In addition, rDNA large subunit (LSU), translation elongation factor 1-alpha (tef1-α), β-tubulin (tub2) and actin (act) genes of isolates HOURD2.1AVR1, PEROG2.1YP2, SAROG1.3AVR10, SAROG1.3AVR13, SAROG1.2AKO1, and SAROG1.3AVR7 were sequenced. Fungal DNA was extracted from 100 mg mycelium (fresh weight), scraped from the surface of 2 to 3-week-old PDA cultures by using a sterilized scalpel, according to Cary et al. [23]. The quantity and quality of the obtained DNA’s were determined using a Q5000 UV-Vis Spectrophotometer (Quawell, San Jose, CA, USA). Final DNA concentration of each isolate was adjusted to 20 ng ml-1 and stored at -20 °C until use. The primer pairs used were ITS1/ITS4 for the rDNA-ITS region, LROR/Un-Lo28S1220 for LSU [24,25], EF1-728F/EF1-986R for tef1-α [26], T1/Bt2b-R for tub2 [26,27] and ACT-512F/ACT-783R for act [26] regions of the six isolates mentioned above. All PCR assays were carried out in an FG-TC01 FastGene® Gradient thermocycler (NIPPON Genetics EUROPE), by the use of BK 1003 KAPA Taq PCR kit (KAPABIOSYSTEMS, Wilmington, Massachusetts, USA). PCR assay for rDNA-ITS included an initial denaturation at 95 oC for 3 min; followed by 35 cycles of 30 sec of denaturation at 94 oC, 30 sec of annealing at 54 oC, and 50 sec of extension at 72 oC; and a final extension step at 72 oC for 5 min. LSU amplification included initial denaturation at 95 oC for 3 min; followed by 35 cycles of 30 sec of denaturation at 94 oC, 30 sec of annealing at 50 oC, and 1 min of extension at 72 oC; and a final extension step at 72 oC for 10 min. Cycling conditions for tef1-α, tub2 and act were similar to those of rDNA-ITS but with annealing temperatures of 53 oC, 51 oC, and 57 oC, respectively. Amplified products were purified with the “NuncleoSpin® Gel and PCR Clean-up” kit (MACHEREY-NAGEL, Düren, Germany) and sequenced by both forward and reverse primers at Macrogen Europe B.V., Amsterdam, the Netherlands. The “BioEdit 7.0.1” software was used to edit the raw sequencing data [28]. For isolates HOURD2.1AVR1, PEROG2.1YP2, SAROG1.3AVR10, SAROG1.3AVR13, SAROG1.2AKO1, and SAROG1.3AVR7 the assembled sequences of their rDNA-ITS, LSU, tef1-α, tub2, and act regions were deposited in GenBank under the accession numbers shown in Supplementary Table S1.
2.5. Identification and Characterization of Isolates
The fungal isolates obtained from diseased wood tissues were characterized using morphological and physiological features, along with rDNA-ITS gene sequences. Molecular identification of the wood-inhabiting fungi was conducted by utilizing the "blastn" option at NCBI and comparing their gene sequences with those in the GenBank database. Additionally, for six (6) selected lesser-known fungal isolates (HOURD2.1AVR1, PEROG2.1YP2, SAROG1.3AVR10, SAROG1.3AVR13, SAROG1.2AKO1, and SAROG1.3AVR7), their LSU, tef1-α, tub2, and act gene sequences were employed. Phylogenetic analysis was performed to assess the relationships of these isolates within the genera Neosetophoma, Didymosphaeria, Seimatosporium, and Kalmusia (Supplementary Table S2). Phylogenetic analysis was carried out for each respective genus using two or more genomic regions: Seimatosporium (rDNA-ITS, LSU, tef1-α, and tub2), Didymosphaeria (rDNA-ITS, LSU, tub2), Kalmusia, and Neosetophoma (rDNA-ITS, LSU). A combined dataset of aligned multi-locus sequences was constructed using "ClustalW," and evolutionary analyses was conducted in Geneious Prime® 2022.1.1 software by employing the Neighbor-Joining method (NJ) and Tamura-Nei model [29]. The NJ consensus trees were based on 1000 bootstrap replications.
2.6. Correlation between Meteorological Data and GTDs Pathogens` Frequency
The potential associations of GTD-incited population structure with the prevailed meteorological conditions in different viticultural zones in Greece were investigated by inquiring and obtaining meteorological data over the last decade (2010-2020) in the regions of interest from the Hellenic National Meteorological Service. In detail, a Principle Component Analysis (PCA) was conducted to assess the correlation between the frequency of GTD pathogens and endophytic fungi isolated from each region and the average values of air temperature, relative humidity, and rainfall prevailed. PCA analysis was performed using GraphPad Prism Software Version 10.0.0 for Windows, Boston, Massachusetts, USA.
2.7. Pathogenicity Tests
In April 2022, a field trial was set up to confirm pathogenicity of four isolates, representative of Seimatosporium vitis (isolate SARO1.3AVR10), Didymosphaeria variabile (isolate SARO1.2AKO1), Kalmusia variispora (isolate HOURD2.1AVR1) and Neosetophoma italica (isolate SARO1.3AVR7), identified by morphological and molecular analyses. The former three species have not been identified previously as causal agents of GTDs in Greece; whereas the latter species has never been reported infecting grapevine worldwide. Isolates were used to inoculate 2-year-old canes of the vine cv. Soultanina grafted onto the 110 Richter rootstock. Pathogenicity trials were conducted in a 15-year-old vineyard located in Hellenic Mediterranean University, Crete, southern Greece. Only vines that remained visibly healthy for at least the last 3 years were included in the experiment.
Artificial inoculation was performed, according to Markakis et al. [12,13]. In brief, a 6.0-mm-diameter and 10.0-mm-length hole was opened into the cane using a Black & Decker EPC 12 drill, and one 5-mm diameter mycelial disc taken from a 2-week-old PDA culture was inserted into the hole. Then, the hole was sealed with cellophane membrane and covered with adhesive paper tape to protect the inoculum. In total, twenty canes (10 vines having had two canes each) were inoculated with each isolate. Additionally, a set of twenty canes (ten vines with two canes each) were similarly treated with sterilized PDA discs and served as controls. Treated canes remained under ambient conditions and were inspected periodically for foliar symptom development.
All the inoculated and control canes were collected in October 2022 (6 months post-inoculation), their leaves were removed, and longitudinal and transverse sections were performed to estimate the extension of wood tissue symptoms above and below the inoculation point. Five representative canes per inoculated fungus were randomly selected to verify the presence of the applied fungi in wood tissues, and pathogen re-isolations onto APDA were conducted as mentioned above.
2.8. Statistical Analysis
Data were checked for normality of distribution using the Kolmogorov-Smirnov and Shapiro-Wilk tests, and for homogeneity using the Levene’s test. Data on disease indices in the field surveys were analyzed using the Kruskal-Wallis (non-parametric) test, followed by Dunn’s posthoc multiple comparison test of average rankings (P ≤ 0.05). The percentage of vines and vineyards infected by individual fungal genera in three viticultural zones were analyzed using the Chi-square test by pair-wise comparisons (at P ≤ 0.05). In pathogenicity trials, analysis of variance (ANOVA) was employed to evaluate the pathogenic effects of the fungal strains on wood discoloration length in canes. When a significant F test was obtained for treatments (P ≤ 0.05), the data were subjected to means separation by Tukey’s HSD test. Standards errors of means were also calculated.
3. Results
3.1. Disease Incidence in Respect to Geographical Region, Vine Age and Cultivar
Vineyards in central and southern Greece showed significantly higher (p<0.05) frequencies of diseased and symptomatic-non dead vines than those in northern Greece (Figure 1). In contrast, a significantly higher (p<0.05) percentage of dead/apoplectic vines was recorded in northern than in southern Greece; whereas no significant difference (p>0.05) was observed between central and northern and between central and southern Greece in terms of dead/apoplectic vines (Figure 1).
In respect of the vineyard age, elder vines (≥20 year-old) showed significantly higher (p<0.05) disease incidence than the middle-aged (10-19 year-old) vines, whereas younger vines demonstrated significantly lower (p<0.05) disease incidence compared to the ones belonging to the other two age categories (Figure 2). The frequencies of symptomatic non-dead plants in vineyards of Heraklion region (southern Greece) and Nemea (central Greece) were significantly higher (p<0.05) than those of Amyntaio, Kavala, Lemnos, Naousa and Nea Mesimvria (northern Greece) (Figure 3). However, no significant difference (p>0.05) in the percentage of dead-apoplectic vines was found among the studied regions (Figure 3).
Different grapevine cultivars varied significantly in terms of diseased (df=23, x2=64.092, p<0.001), symptomatic-non dead (df=23, x2=70.420, p<0.001), and dead/apoplectic vines (df=23, x2=44.575, p≤0.001) frequencies. In detail, the frequency of diseased vines of cvs. Moschofilero, Tzortzina, Nebiollo, Liatiko and Asyrtiko was significantly lower (p<0.05) compared to Cabernet Sauvignon, Traminer, Vidiano, Tempranillo, Soultanina, Agiorgitiko, Cisnaut, and Kotsifali. At the same time, most of the rest cultivars did not differ significantly (p>0.05) from the highly or the lowly affected cultivars mentioned above (Figure 4A). Moreover, the frequency of symptomatic-non dead vines in Soultanina, Agiorgitiko and Kotsifali vineyards was significantly higher (p<0.05) than those of most of the other cultivars tested (Figure 4B). Furthermore, the frequency of dead-apoplectic vines of Moschofilero was significantly lower (p<0.05) compared to that of Syrah, Tempranilo, Agiorgitiko, and Cinsaut, whereas most of the other cultivars did not differ significantly (p>0.05) from the above cultivar groups in terms of dead/apoplectic vines frequency (Figure 4C).
3.2. Fungal Genera Frequencies
In total, 533 isolates were obtained from northern (n= 124), central (n= 146) and southern (n= 263) Greece. Determination of fungal genera based on morphological and molecular (5.8S rRNA gene-ITS sequencing) analyses revealed that isolates belonged to 35 distinct fungal genera (Figure 5). Amongst them, 18 fungal genera (including 407 out of 533 isolates) have been commonly associated with GTDs worldwide including well-known (i.e. Botryosphaeria dothidea, Cytospora spp., Diaporthe spp., Diplodia sp., Dothiorella sp., Eutypa sp., Eutypella sp., Fomitiporia mediterranea, Ilyonectria sp., Neofusicoccum sp., Phaeoacremonium spp., Phaeomoniella clamydospora, Phellinus sp.) and lesser-known (i.e. Didymosphaeria sp., Fusarium sp., Seimatosporium sp., Kalmusia sp.) grapevine wood inhabitants; whereas the rest 17 determined genera (e.g. Alternaria sp., Aspergillus sp., Clonostachys sp., Neosetophoma sp., Trichoderma sp. etc.) have never/rarely been associated with GTDs.
GTDs-inducing population structure differed significantly (p<0.05) among the discrete geographical zones. Phaeomoniella chlamydospora (26.62%, n=70/263), Diaporthe spp. (18.25%, n= 48/263) and F. mediterranea (10.27%, n=27/263) were the most prevalent in Heraklion, whereas D. seriata (28.77%, n=42/146), P. chlamydospora and Fusarium spp. (10.27%, n=15/146) were predominant in Nemea (central Greece). In Amyntaio and Kavala (northern Greece), D. seriata was the most frequently isolated species (51.61%, n= 64/124) followed by Lasiodiplodia sp. (8.87%, n=11/124). Interestingly, P. chlamydospora was not isolated from vines in northern Greece, nor Lasiodiplodia sp. from southern or central Greece, while Diaporthe sp. was not found in central Greece (Figure 5).
Overall, increased frequencies of isolates belonging to GTD-associated genera originated from elder (39.80%, n=162/407) and middle-aged (49.39%, n=201/407) vines compared to the younger ones (10.81%, n=44/407). Relative frequencies of isolates per fungal genus and vine age category are shown in Figure 6. Apart from Botryosphaeria sp. and Fusarium sp., isolates of the most frequently occurring genera were obtained mainly from middle-aged and elder vines. Although at low frequencies, 11 out of 18 most commonly known GTD-associated genera were also found in younger vines.
3.3. Incidence of GTD-Associated Fungal Genera in Vineyards and Vines in Discrete Viticultural Zones
The incidence of specific fungal genera associated with GTDs in vineyards and vines in discrete viticultural zones differed significantly (p<0.05). The percentage of vineyards in northern (Amyntaio and Kavala), central (Nemea), and southern (Heraklion) Greece, where each of the 18 commonly known GTD-associated genera was detected, is shown in Figure 7. Ten out of 18 genera were found at significantly different (p<0.05) frequencies among vineyards of the discrete zones. Furthermore, nine (9) genera were isolated at significantly different (p<0.05) frequencies among vines of discrete zones (Figure 8). Indicatively, the percentage of diseased vines that were infected by Diplodia spp. (mostly D. seriata) in northern Greece (58.33%) was significantly higher (p<0.05) than that in central Greece (42.03%), while the percentage of Diplodia-infected vines in southern Greece (4.88%) was significantly lower (p<0.05) compared to those in the other two zones. Contrariwise, the percentage of vines that were infected by Phaeomoniella sp. (P. chlamydospora) in southern Greece (28.46%) was significantly higher (p<0.05) than that in central Greece (8.70%), whereas no Phaeomoniella-infected vines were found in northern Greece (0.00%) (Figure 8). Furthermore, 26.32% of the studied vineyards and 10.00% of symptomatic vines were infected by Lasiodiplodia spp. (mostly L. viticola and L. theobromae) in northern Greece, exclusively (Figure 7 and Figure 8).
3.4. Correlation between Meteorological Data and GTDs Pathogens` Frequency
PCA analysis allowed us to investigate the possible correlation of the past decade's meteorological data (air temperature, rainfall, and relative humidity) with the frequencies of the prevalent fungal genera isolated from symptomatic grapevines of Southern, Central, and Northern Greece (Figure 9). PCA analysis was performed on the correlation matrix of 23 variables, and the first two components explained 100% of the total variation of the data (68.85% for PC1, and 31.15% for PC2) (Figure 9). The arrangement of variables on the graph reveals how they are clustered together across the two dimensions. More specifically, the three examined viticultural zones were differentiated into three clusters; PC2 separated the viticultural zones of northern and central Greece, whereas PC1 only differentiate the datasets representing the southern zone (Figure 9). The correlation between two variables is determined by the angle formed by the arrows pointing towards them, thus acute angle indicates a (strong) positive correlation, a right angle represents no correlation, and an obtuse angle signifies a (strong) negative correlation. Furthermore, arrows pointing in the same direction indicate a positive correlation, and the length of the arrow reflects the strength of the correlation pattern. Conversely, arrows pointing in opposite directions reveal a negative correlation. A positive correlation was noticed among southern zone of Greece, air temperature, relative humidity (RH), and several endophytic fungal genera, such as Phaemoniella sp., Phaeoacremonium sp., Fomitiporia sp., Seimatosporium sp., Botryosphaeria sp., Kalmusia sp., Diaporthe sp., and Didymosphaeria sp. In contrast, the fungal genera of Eutypa sp., Neofusicoccum sp., Diplodia sp., and Lasiodiplodia sp., were positively correlated with the northern zones of Greece and negatively correlated with the temperature (Figure 9). Additionally, central zone of Greece was positively correlated with rainfall and the fungal genera of Phellinus sp., Aspergillus sp., Cytospora sp., Fusarium sp., and Alternaria sp. (Figure 9).
3.5. Morphological, Physiological, and Molecular Identification of Lesser-Known Fungal Species
Morphological and physiological characteristics of the six (6) selected isolates (HOURD2.1AVR1, PEROG2.1YP2, SAROG1.3AVR10, SAROG1.3AVR13, SAROG1.2AKO1, and SAROG1.3AVR7), along with their GenBank accession numbers and identities based on the analysis of their rDNA-ITS, LSU, tef1-α, tub2 and act sequences, are presented in Supplementary Table S1. These isolates were selected and analyzed thoroughly since they belonged to lesser-known GTD-associated fungal genera that have not been reported previously in Greece (i.e. Seimatosporium, Didymosphaeria and Kalmusia) or genera that have not been reported as GTD-inciting agents worldwide (i.e. Neosetophoma). Apart from the tef1-α sequence of the isolate PEROG2.1YP2 (K. variispora), all isolates showed over 99.50% similarity with already published rDNA-ITS, LSU, tef1-α and tub2 sequences that are deposited in NCBI. Moreover, gene sequences of the fungal species that were not available in NCBI, were deposited (i.e. act sequences of K. variispora and S. vitis, LSU sequence of D. variabile, and LSU, tef1-α, tub2 and act sequences of N. italica).
In this study, we conducted a comprehensive phylogenetic analysis using combined alignments of rDNA-ITS, LSU, tub2, and tef1-a sequences from two S. vitis isolates (SAROG1.3AVR10 and SAROG1.3AVR13). The Neighbor-Joining (NJ) method was employed and a phylogenetic tree with four (4) distinct clades was generated. Our isolates were found to be grouped in the same clade as the reference isolates of S. vitis (Napa764, L189, CBS 123004, P210) (Supplementary Table S2) with a strong bootstrap support of 100% (Supplementary Figure S2). Furthermore, NJ analysis based on the combined sequences of rDNA-ITS and LSU showed that our N. italica isolate (SAROG1.3AVR7), along with the reference isolates (MFLU 14-0809 and 108) (Supplementary Table S2), formed a well-supported clade with a bootstrap value of 100% (Supplementary Figure S3). Similar results were observed for the K. variispora isolates, as NJ analysis of the assembled sequences of rDNA-ITS and LSU demonstrated a well-supported clustering (93.4%) with the reference strains KV-9, CBS 121517, and KV-13 (Supplementary Table S1, Supplementary Figure S4). Finally, we utilized combined sequences of rDNA-ITS, LSU, and tub2 to construct a concatenated phylogenetic tree that included our isolate of D. variabile (SAROG1.2AKO1) along with other species of the genus Didymospaeria. The NJ analysis revealed a phylogenetic tree with two (2) well-supported clades, where our isolate (SAROG1.2AKO1) clustered together with the reference isolate D. variabile (CBS 120014) (Supplementary Table S2) with a bootstrap support of 100% (Supplementary Figure S5).
Through Blast and phylogenetic analysis, we successfully identified N. italica (SAROG1.3AVR7), S. vitis (SAROG1.3AVR10 and SAROG1.3AVR13), D. variabile (SAROG1.2AKO1), and K. variispora (HOURD2.1AVR1 and PEROG2.1YP2) isolates, while their colony color, growth rate, and microscopic features were indicative of their respective holotypes among the isolates.
3.6. Pathogenicity of Fungal Isolates
After a 6-month incubation period, longitudinal and transverse sections revealed severe wood tissue discoloration in inoculated canes treated with S. vitis (isolate SARO1.3AVR10), D. variabile (isolate SARO1.2AKO1), K. variispora (isolate HOURD2.1AVR1) and N. italica (isolate SARO1.3AVR7), compared to controls (Figure 10, Supplementary Figure S6). Wood discoloration length caused by N. italica was significantly higher (p<0.05) than the other fungal species tested. All these fungal species were successfully re-isolated from the artificially inoculated canes, whereas no fungus was obtained from control canes, thus confirming pathogenicity. No external leaf symptoms were observed in control- and fungal-treated canes within the 6-month period of bioassays.
4. Discussion
In the last two decades, increased incidence of grapevine trunk diseases (GTDs) has been reported in numerous studies, and their detrimental effect on grapevine industry has raised awareness globally [4,7,10]. In the present study, a three-year survey (from 2018 to 2020) was carried out to investigate the incidence and identify fungi associated with GTDs in Greece. In total, 310 vineyards in different geographical regions with variable pedoclimatic conditions in northern, central, and southern Greece were surveyed, and 533 fungal strains were isolated from diseased vines and identified.
Measurements of GTDs incidence in geographically distinct regions in Greece, revealed higher frequency of symptomatic plants in vineyards in the warm and dry region of Heraklion (southern Greece) compared to those in cold and wet regions in northern Greece. Vineyards in central Greece (Nemea), characterized by moderate rainfall levels during the period of increased grapevine water requirements (in spring and summer months), indicated intermediate disease incidence. Differences in GTD symptom expression between vineyard regions can be attributed to variable climatic conditions [30]. Although correlation between climatic or environmental conditions and GTD symptom severity is not always clear, the combined effect of heat and water stresses on grapevine has been speculated to favor initiation and progress of GTDs [10,11].
In addition, differences in the disease incidence between regions may be correlated with differences in the level of susceptibility to GTDs of the vine cultivars, grown in each region. Knowledge of susceptibility for widely used plant genotypes is essential for estimating disease risk and planning effective management strategies [13]. The present study's data suggests that grapevine cultivars grown in Greece are differentially affected by GTDs, thus demonstrating variable resistance. Considerably increased disease incidence (in terms of diseased vines and symptomatic-non dead vines) was recorded on ‘Agiorgitiko’ grown primarily in central, and ‘Kotsifali’ and ‘Soultanina’ grown primarily in southern Greece. Apart from Kotsifali, these cultivars have been mentioned among the most susceptible to various GTD-pathogens in several studies as defined by artificial inoculation experiments and field observations [31]. On the other hand, recent unpublished data of our group suggest that cultivars, such as Xinomavro or Limnio, which are among the most commonly used cultivars in northern Greece are highly resistant to GTDs (Testempasis, Unpublished data) and this could explain the lower disease incidence measured in northern Greece.
The occurrence of GTDs exhibits a positive correlation with the age of vines, displaying an exponential-related pattern. Our research findings indicate a heightened prevalence of GTDs in older vines (≥20 years old), followed by intermediate and lower incidence rates in middle-aged (10-19 years old) and younger (0-9 years old) vines, respectively. These outcomes align with earlier studies that observed increased frequencies of diseased vines in middle-aged and older categories compared to younger ones [32,33]. The variations in disease incidence across age groups were, to some extent, linked to the frequencies of GTD-inducing fungal genera isolated from each vine age category. Notably, the majority of GTD-inciting fungi were predominantly harbored in middle-aged and elder vines. Previous research has similarly documented a correlation between vine's age and the presence of specific GTDs, including White rot, Botryosphaeria and Eutypa dieback [7,34,35].
Climatic conditions, including heat and water stress, influence the prevalence and distribution of GTDs fungi [11]. Indeed, in the case of geographically distinct viticultural regions studied here, the population structure of GTD-related fungi differed significantly. Indicatively, the most predominant genera/species in regions of southern Greece (e.g. P. chlamydospora and Diaporthe spp.) were completely absent or found at very low frequencies in northern and/or central Greece and vice versa. These findings are strongly supported by a recent grapevine wood microbiome study conducted in Greece [14] pointing out that certain GTD-associated pathogens like P. chlamydospora and Diaporthe spp. were consistently present in cultivar ‘Vidiano’ collected from vineyards in southern (Heraklion, Crete) but not in ‘Xinomavro’ collected from vineyards in northern Greece (Amyntaio). On the contrary, Diplodia spp. (mostly D. seriata) were the most frequently isolated species in northern and central Greece but considerably less frequent in southern Greece. Furthermore, Lasiodiplodia spp. (mostly L. viticola and L. theobromae) were found in northern Greece, exclusively. Likewise, Hernandez and Alchala [32], reported higher abundancies of Botryosphaeria dieback pathogens in the northern regions of Oregon compared to the southern ones. Additionally, the elevated incidence of Botryosphaeria dieback complex pathogens in the northern regions of Greece might be associated with the prevalence of these pathogens in temperate regions or those with cold winters [36].
Out of 35 fungal genera identified in total, 25 were found in the warm and dry region in southern Greece, whereas 15 and 19 out of 35 genera were found in the comparatively cooler regions in northern and central Greece, respectively. Amongst them, 18 fungal genera are well-known (e.g. Botryosphaeria dothidea, Diaporthe spp., Diplodia seriata, Eutypa sp., Fomitiporia mediterranea, Phaeoacremonium spp., Phaeomoniella clamydospora) or lesser-known (i.e. Fusarium sp., Seimatosporium sp., Didymosphaeria sp., Kalmusia sp.) GTDs-inducing agents [7,16,19,20,21]. The rest 17 determined genera include wood canker agents or saprophytes (e.g. Alternaria sp., Aspergillus sp., Penicillium sp., Rhizopus sp.), endophytic symbionts with potential biocontrol activity (e.g. Clonostachys sp., Epicoccum sp., Trichoderma sp.), or genera that have not been reported previously as wood tissue colonizers (e.g. Lopadostroma sp., Monocillium sp., Neosetophoma sp.) [14, 37-41]. The increased number of fungal genera originated from GTD-affected vines in southern Greece could be partially attributed to the thermophilic nature (within a certain limit) of most GTD-associated fungi [11]. Besides, the higher number of samples collected and analyzed from southern Greece may also play a role in the increased fungal diversity identified there.
Sequencing of rDNA-ITS, LSU, tef1-α, tub2, and act genes of selected isolates, coupled with pathogenicity tests, revealed that N. italica, S. vitis, D. variabile and K. variispora caused wood infection on grapevine canes, with the former species being the most virulent. Although these fungi were identified (at the genus level) in a grapevine wood microbiome study conducted recently, their pathogenic potential was not examined [14]. Therefore, this is the first record of K. variispora, S. vitis and D. variabile associated with GTDs in Greece. Seimatosporium vitis has never been isolated in Greece previously, whereas K. variispora and D. variabile have been reported as causal agents of fruit rot on apples [42] and leaf spot on Phoenix theophrasti [43], respectively. Τhis is also the first report of N. italica associated with GTDs worldwide. Neosetophoma italica was first found as saprobic on dead leaves of Iris germanica L. in Italy and typified in 2015 [44]. Its close relative, N. samararum has been reported as a pathogen causing leaf spots of various hosts [45], but up to now, N. italica has never been shown to infect plant hosts. The increased virulence of N. italica revealed here, may suggest the predominant role of this species in GTD-associated microbiome, and should be taken into consideration in future surveys in Greece and elsewhere.
Given all the above, the data from the present study suggest an overall increased incidence of GTDs in Greece. Differences in disease incidence and population structure of the GTD-associated microbiome were observed among the discrete viticultural zones. The increased incidence of GTDs and the high variability of fungal genera implicated with the disease complex, recorded in the driest and warmest region of Heraklion (southern Greece) may portend even more ominous scenarios for viticulture sustainability globally, against climate change. Apart from the different climatological conditions, these variances may be reasonably attributed to the differential susceptibility level of grapevine genotypes cultivated in geographically distinct viticultural regions. Herein, we record for the first time K. variispora, S. vitis and D. variabile as GTD-associated pathogens in Greece, and N. italica as GTDs-inducing pathogen worldwide. This highly virulent species may have a predominant role in GTDs complex, and its biological cycle and epidemiology should be further investigated. Regional-scale mapping conducted here will provide valuable support in planning and implementing effective management practices targeting specific diseases.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Map illustrating the geographical distribution of the surveyed regions across Greece; Table S1: Morphological and physiological features of the fungal isolates analyzed in the present study, along with their geographic origin, the plant tissue they were isolated and the GenBank accession numbers; Table S2: Sequences used in the phylogenetic analysis; Figure S2: Neighbor-joining (NJ) phylogram of multi locus allignment of Seimatosporium species; Figure S3: Neighbor-joining (NJ) phylogram of multi locus allignment of Neosetophoma species; Figure S4: Neighbor-joining (NJ) phylogram of multi locus allignment of Kalmusia species; Figure S5: Neighbor-joining (NJ) phylogram of multi locus allignment of Didymosphaeria and Paraconiothiorum species; Figure S6: Wood discoloration of artificially inoculated canes of cv. Soultanina with Seimatosporium vitis, Didymosphaeria variabile, Kalmusia variispora and Neosetophoma italica. .
Author Contributions
Conceptualization, E.A.M, G.S.K., E.P., A.K.T.; methodology, E.A.M, G.S.K, E.P, S.I.T, D.G, A.K.T.; software, E.A.M., S.I.T; validation, E.A.M., G.S.K., A.K.T., E.P., S.I.T.; investigation, S.I.T., E.A.M., G.I.T., S.K.S., C.T., D.G., A.K.T., E.P., G.S.K.; resources, E.A.M., S.I.T., G.S.K.; writing—original draft preparation, E.A.M., S.I.T.; writing—review and editing, E.A.M, S.I.T., G.S.K., E.P., A.K.T., D.G.; visualization, E.A.M., S.I.T; supervision, E.A.M, G.S.K., E.P., A.K.T.
Funding
This work was funded by Greek national funds through the Public Investments Program (PIP) of the General Secretariat for Research & Technology (GSRT), under the Emblematic Action “Routes of Vineyards”, Grant No. 6070.03.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
This work was funded by Greek national funds through the Public Investments Program (PIP) of the General Secretariat for Research & Technology (GSRT), under the Emblematic Action “Routes of Vineyards”, Grant No. 6070.03. Authors are also grateful to grapevine growers for providing access and information for the studied vineyards.
Conflicts of Interest
The authors declare no conflict of interest.
References
- OIV. Distribution of the World’s Grapevine Varieties. Int. Organ. Vine Wine.
- OIV. Country Statistics. Int. Organ. Vine Wine 2021. [Google Scholar]
- FAOSTAT. Statistics “Crops and Livestock products”. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 September 2023).
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases: A Review; OIV Publications, 2016.
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine Trunk Diseases: Complex and Still Poorly Understood. Plant Pathol. 2012, 62, 243–265. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and Brown Wood-Streaking: Two Old and Elusive Diseases of Grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk Diseases with Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Kraus, C.; Markakis, E.; Alves, E.; Armengol, J.; Eichmeier, A.; Compant, S.; Gramaje, D. Phytopathologia Mediterranea. Phytopathol. Mediterr. 2022, 61, 563–599. [Google Scholar] [CrossRef]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Urbez-Torres, J.R. Phytopathologia Mediterranea. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef]
- Songy, A.; Fernandez, O.; Clement, C.; Larignon, P.; Fontaine, F. Grapevine Trunk Diseases under Thermal and Water Stresses. Planta 2019, 249, 1655–1679. [Google Scholar] [CrossRef]
- Markakis, E.A.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Evaluation of Greek Grapevine Cultivars for Resistance to Phaeomoniella Chlamydospora. Eur. J. Plant Pathol. 2017, 149, 277–283. [Google Scholar] [CrossRef]
- Markakis, E.A.; Ligoxigakis, E.K.; Roussos, P.A.; Chrysi Sergentani; Nektarios Kavroulakis; Emmanouil Roditakis; Georgios Koubouris. Differential Susceptibility Responses of Greek Olive Cultivars to Fomitiporia Mediterranea. Eur. J. Plant Pathol. 2018, 153, 1055–1066. [Google Scholar] [CrossRef]
- Bekris, F.; Vasileiadis, S.; Papadopoulou, E.; Samaras, A.; Testempasis, S.; Gkizi, D.; Tavlaki, G.; Tzima, A.; Paplomatas, E.; Markakis, E.; et al. Correction To: Grapevine Wood Microbiome Analysis Identifies Key Fungal Pathogens and Potential Interactions with the Bacterial Community Implicated in Grapevine Trunk Disease Appearance. Environ. Microbiome 2021, 17. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gautier, A.; Laval, V.; Compant, S.; Maurer, W.; Sessitsch, A.; Lebrun, M.; Rey, P. Major Changes in Grapevine Wood Microbiota Are Associated with the Onset of Esca, a Devastating Trunk Disease. Environ. Microbiol. 2020, 22, 5189–5206. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Zhang, W.; Zhang, J.; Wang, H.; Peng, J.; Wang, X.; Yan, J. Belowground Microbiota Analysis Indicates That Fusarium Spp. Exacerbate Grapevine Trunk Disease. Environ. Microbiome 2023, 18. [Google Scholar] [CrossRef]
- Martín, L.; García-García, B.; Alguacil, M. del M. Interactions of the Fungal Community in the Complex Patho-System of Esca, a Grapevine Trunk Disease. Int. J. Mol. Sci. 2022, 23, 14726. [Google Scholar] [CrossRef]
- Patanita, M.; Albuquerque, A.; Campos, M.D.; Materatski, P.; Varanda, C.M.R.; Ribeiro, J.A.; Félix, M. do R. Metagenomic Assessment Unravels Fungal Microbiota Associated to Grapevine Trunk Diseases. Horticulturae 2022, 8, 288. [Google Scholar] [CrossRef]
- Bustamante, M.I.; Elfar, K.; Smith, R.J.; Bettiga, L.J.; Tian, T.; Torres, G.A.; Eskalen, A. First Report of Fusarium Annulatum Associated with Young Vine Decline in California. Plant Dis. 2022, 106, 2752. [Google Scholar] [CrossRef]
- Kanetis, L.; Taliadoros, D.; Makris, G.; Christoforou, M. A Novel Seimatosporium and Other Sporocadaceae Species Associated with Grapevine Trunk Diseases in Cyprus. Plants 2022, 11, 2733–2733. [Google Scholar] [CrossRef]
- Raimondo, M.L.; Carlucci, A.; Ciccarone, C.; Sadallah, A.; Lops, F. Identification and Pathogenicity of Lignicolous Fungi Associated with Grapevine Trunk Diseases in Southern Italy. Phytopathol. Mediterr. 2019, 58, 639–662. [Google Scholar]
- Rumbos, I.; Rumbou, A. Fungi Associated with Esca and Young Grapevine Decline in Greece. Phytopathol. Mediterr. 2001, 40, S330–S335. [Google Scholar] [CrossRef]
- Cary, J.W.; Ehrlich, K.C.; Beltz, S.B.; Harris-Coward, P.; Klich, M.A. Characterization of the Aspergillus Ochraceoroseus Aflatoxin/Sterigmatocystin Biosynthetic Gene Cluster. Mycologia 2009, 101, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Bala, K.; Robideau, G.P.; Désaulniers, N.; de Cock, A.W.A.M.; Lévesque, C.A. Taxonomy, DNA Barcoding and Phylogeny of Three New Species of Pythium from Canada. Persoonia - Mol. Phylogeny Evol. Fungi 2010, 25, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the FungusFusariumAre Nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and Phylogeny of Gliocladium Analysed from Nuclear Large Subunit Ribosomal DNA Sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10. [Google Scholar] [CrossRef]
- Fussler, L.; Kobes, N.; Bertrand, F.; Maumy, M.; Grosman, J.; Savary, S. A Characterization of Grapevine Trunk Diseases in France from Data Generated by the National Grapevine Wood Diseases Survey. Phytopathol. ® 2008, 98, 571–579. [Google Scholar] [CrossRef]
- Markakis, E.A.; Kavroulakis, N.; Ntougias, S.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Characterization of Fungi Associated with Wood Decay of Tree Species and Grapevine in Greece. Plant Dis. 2017, 101, 1929–1940. [Google Scholar] [CrossRef]
- Hernandez, M.; KC, A. A Systematic Survey on Prevalence of Grapevine Trunk Disease Pathogens in Oregon Vineyards. Plant Dis. 2022, 107, 1355–1364. [Google Scholar] [CrossRef]
- White, C.-L.; Halleen, F.; Mostert, L. Symptoms and Fungi Associated with Esca in South African Vineyards. Phytopathol. Mediterr. 2011, 50, S236–S246. [Google Scholar]
- Moretti, S.; Pacetti, A.; Pierron, R.; Hanns-Heinz Kassemeyer; Fischer, M. ; Jean-Pierre Péros; Pérez-González, G.; Bieler, E.; Schilling, M.; Stefano Di Marco; Éric Gelhaye; Mugnai, L.; Bertsch, C.; Farine, S. Fomitiporia Mediterranea M. Fisch., the Historical Esca Agent: A Comprehensive Review on the Main Grapevine Wood Rot Agent in Europe. Phytopathol. Mediterr. 2021, 60, 351–379. [Google Scholar] [CrossRef]
- Urbez-Torres, J.R.; Peduto, F.; Striegler, R.K.; Urrea-Romero, K.E.; Rupe, J.C.; Cartwright, R.D.; Gubler, W.D. Characterization of Fungal Pathogens Associated with Grapevine Trunk Diseases in Arkansas and Missouri. Fungal Divers. 2011, 52, 169–189. [Google Scholar] [CrossRef]
- Larach, A.; Vega-Celedón, P.; Salgado, E.; Salinas, A.; Riquelme, N.; Castillo-Novales, D.; Sanhueza, P.; Seeger, M.; Besoain, X. Higher Virulence of Diplodia Seriata Isolates on Vines of Cv. Cabernet Sauvignon Associated with 10-Year-Old Wood Compared to Young Tissue. Plants 2023, 12, 2984. [Google Scholar] [CrossRef]
- Cobos, R.; Ibañez, A.; Diez-Galán, A.; Calvo-Peña, C.; Ghoreshizadeh, S.; Coque, J.J.R. The Grapevine Microbiome to the Rescue: Implications for the Biocontrol of Trunk Diseases. Plants 2022, 11, 840. [Google Scholar] [CrossRef]
- Del Frari, G.; Cabral, A.; Nascimento, T.; Boavida Ferreira, R.; Oliveira, H. Epicoccum Layuense a Potential Biological Control Agent of Esca-Associated Fungi in Grapevine. PLOS ONE 2019, 14, e0213273. [Google Scholar] [CrossRef]
- Michailides, T.J.; Peacock, W.; Christensen, P.; Morgan, D.P.; Felts, D. First Report of Aspergillus Vine Canker of Table Grapes Caused by Aspergillus Niger. Plant Dis. 2002, 86, 75–75. [Google Scholar] [CrossRef]
- Silva-Valderrama, I.; Toapanta, D.; Miccono, M. de los A.; Lolas, M.; Díaz, G.A.; Cantu, D.; Castro, A. Biocontrol Potential of Grapevine Endophytic and Rhizospheric Fungi against Trunk Pathogens. Front. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Vitale, A.; Cirvilleri, G.; Panebianco, A.; Epifani, F.; Perrone, G.; Polizzi, G. Molecular Characterisation and Pathogenicity of Aspergillus Sect. Nigri Causing Aspergillus Vine Canker of Table Grapes in Italy. Eur. J. Plant Pathol. 2011, 132, 483–487. [Google Scholar] [CrossRef]
- Ntasiou, P.; Samaras, A.; Karaoglanidis, G. Apple Fruit Core Rot Agents in Greece and Control with Succinate Dehydrogenase Inhibitor Fungicides. Plant Dis. 2021, 105, 3072–3081. [Google Scholar] [CrossRef]
- Ligoxigakis, E.K.; Papaioannou, I.A.; Markakis, E.A.; Typas, M.A. First Report of Leaf Spot of Phoenix Theophrasti Caused by Paraconiothyrium Variabile in Greece. Plant Dis. 2013, 97, 1250–1250. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal Diversity Notes 1–110: Taxonomic and Phylogenetic Contributions to Fungal Species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Phookamsak, R.; Liu, J.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.A.; Thambugala, K.M.; Dai, D.-Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar] [CrossRef]
Figure 1.
Grapevine Trunk Diseases incidence (symptomatic-non dead and dead-apoplectic vines) in vineyards located in northern, central and southern Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
Figure 1.
Grapevine Trunk Diseases incidence (symptomatic-non dead and dead-apoplectic vines) in vineyards located in northern, central and southern Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
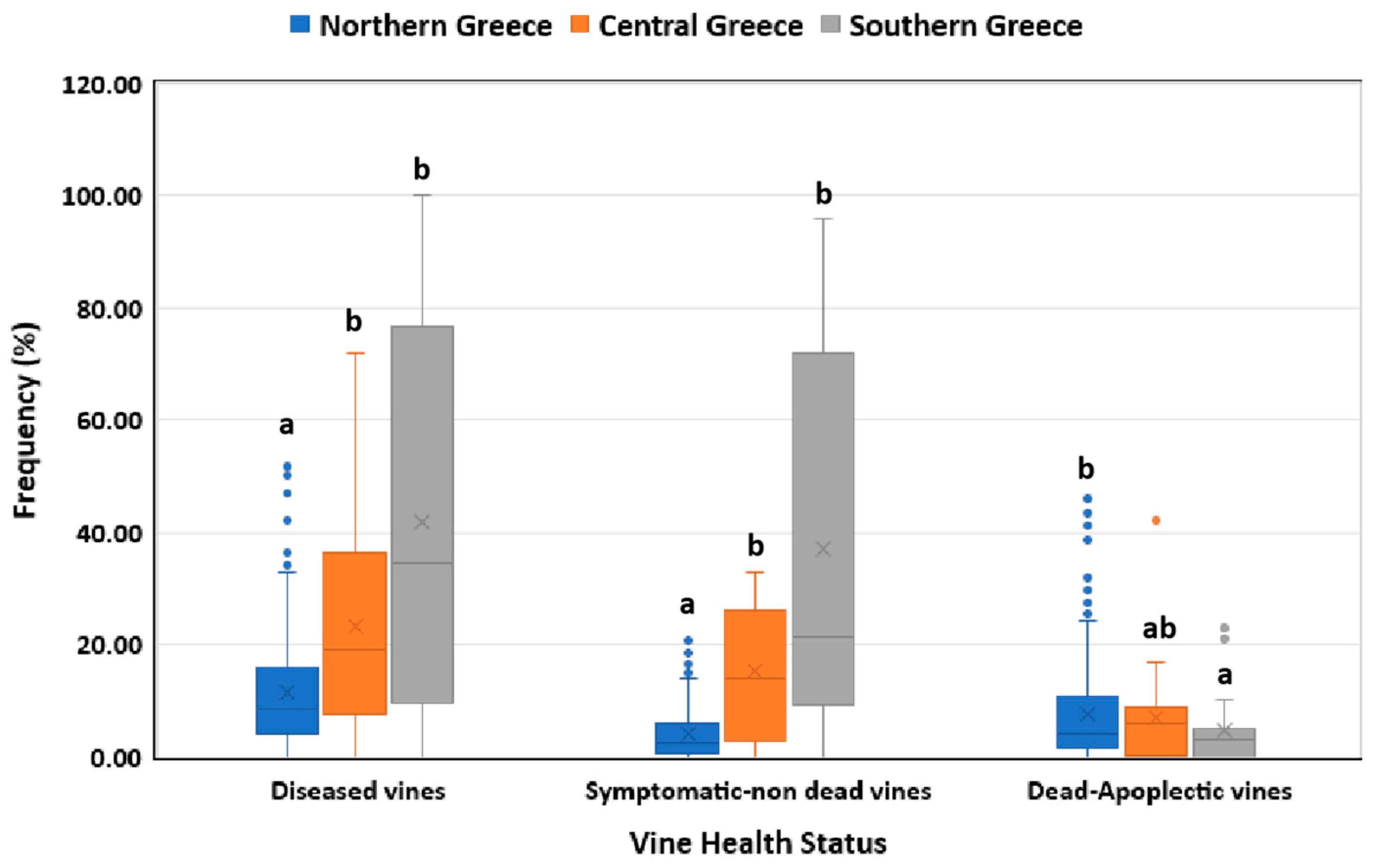
Figure 2.
Grapevine Trunk Diseases incidence (symptomatic-non dead and dead-apoplectic vines) for young (0-9 years old), middle-aged (10-19) and elder (≥20 years old) vineyards. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
Figure 2.
Grapevine Trunk Diseases incidence (symptomatic-non dead and dead-apoplectic vines) for young (0-9 years old), middle-aged (10-19) and elder (≥20 years old) vineyards. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
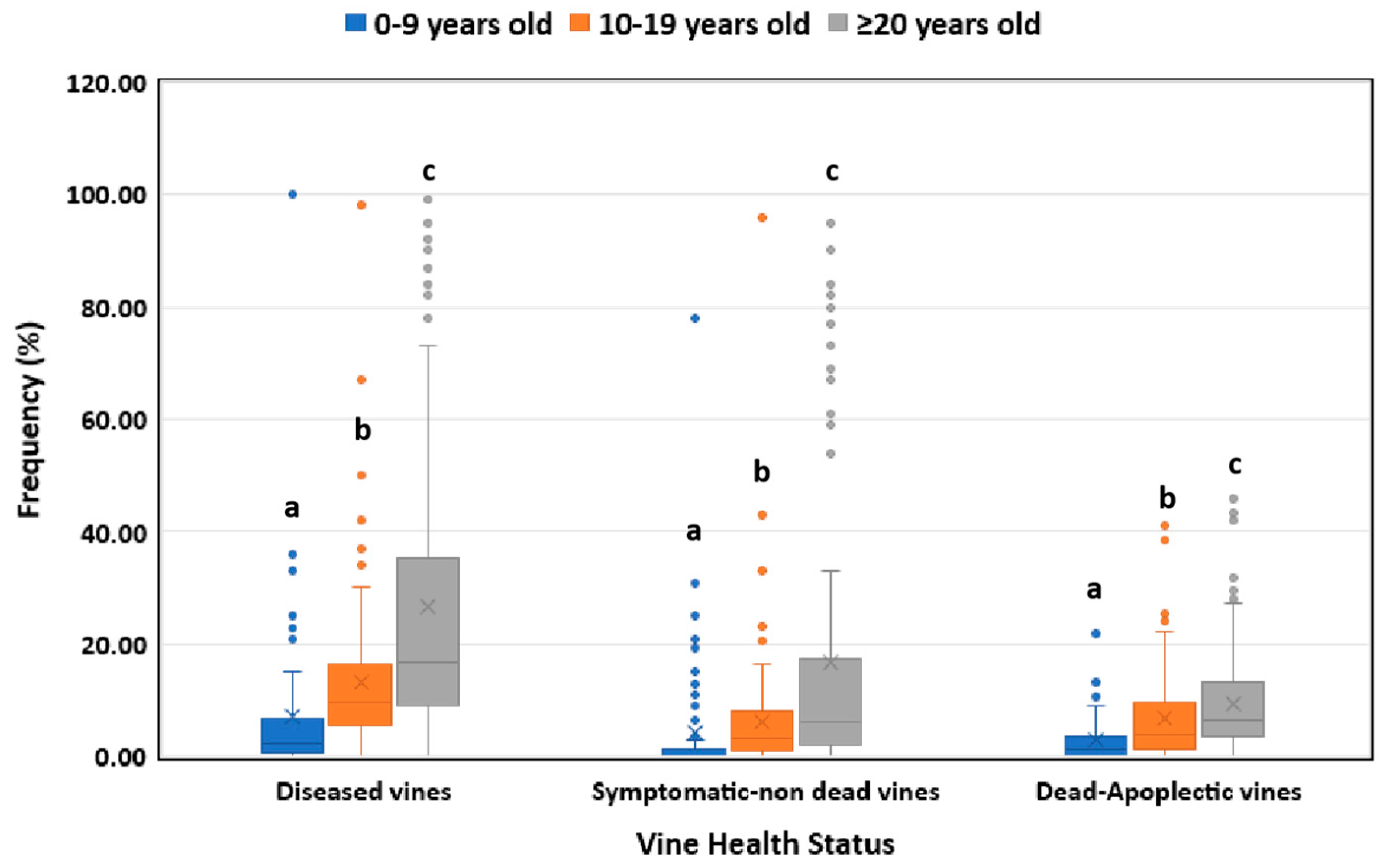
Figure 3.
Grapevine Trunk Diseases incidence (diseased, symptomatic-non dead and dead/apoplectic vines) for vineyards located in different regions in Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
Figure 3.
Grapevine Trunk Diseases incidence (diseased, symptomatic-non dead and dead/apoplectic vines) for vineyards located in different regions in Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Within each group of plants, columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
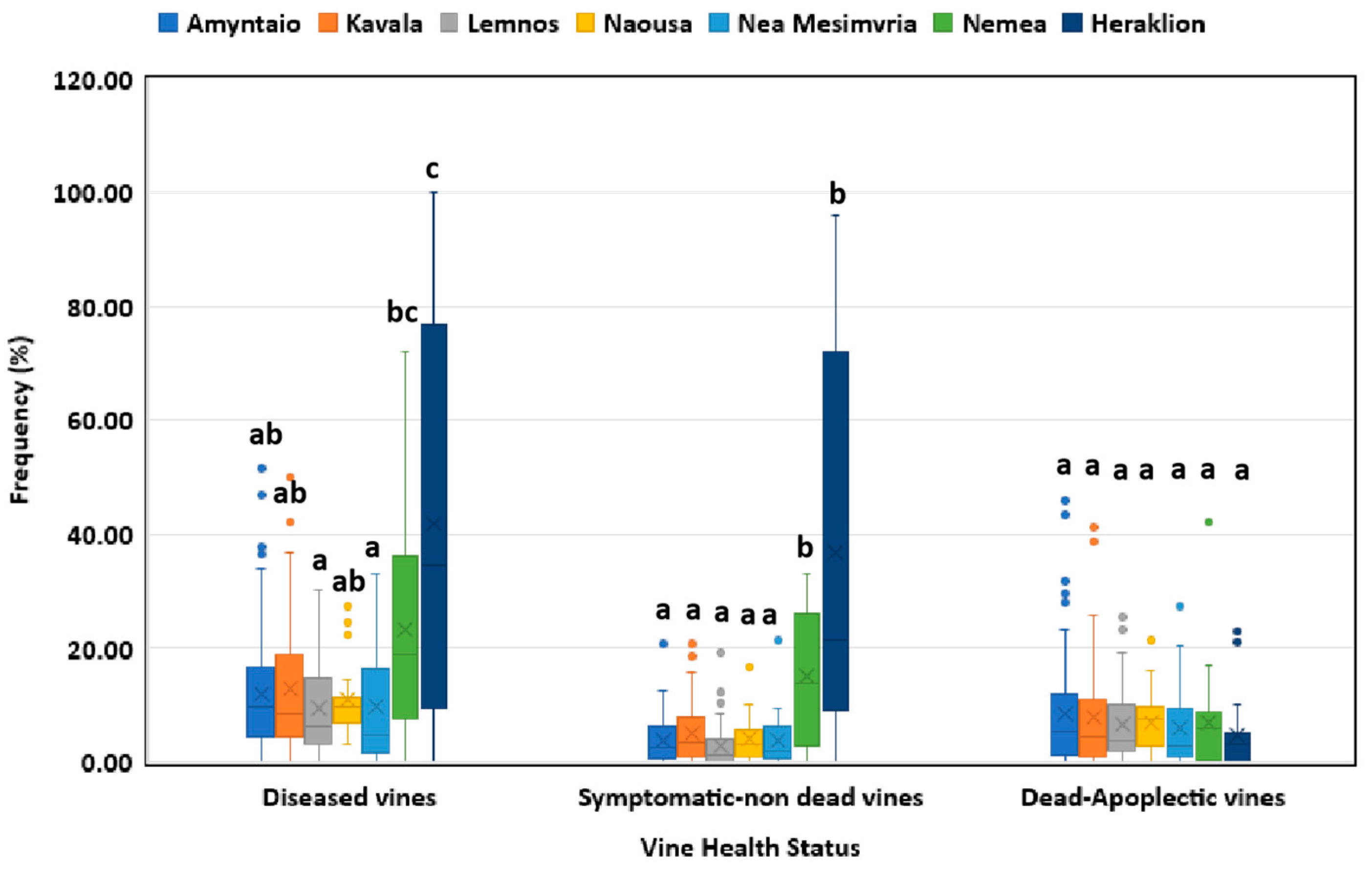
Figure 4.
Frequency (%) of diseased (A), symptomatic-non dead (B) and dead/apoplectic (C) vines on 24 grapevine cultivars grown in Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
Figure 4.
Frequency (%) of diseased (A), symptomatic-non dead (B) and dead/apoplectic (C) vines on 24 grapevine cultivars grown in Greece. Within the box, the median and the mean are represented by the solid line and the ‘X’, respectively. Top and bottom lines of the box correspond to the 25th and 75th percentiles of the data, respectively. Error bars represent the 10th and 90th percentiles. Columns with the same letter do not differ significantly according to Kruskal-Wallis and the Dunn’s test for multiple comparisons (p ≤ 0.05).
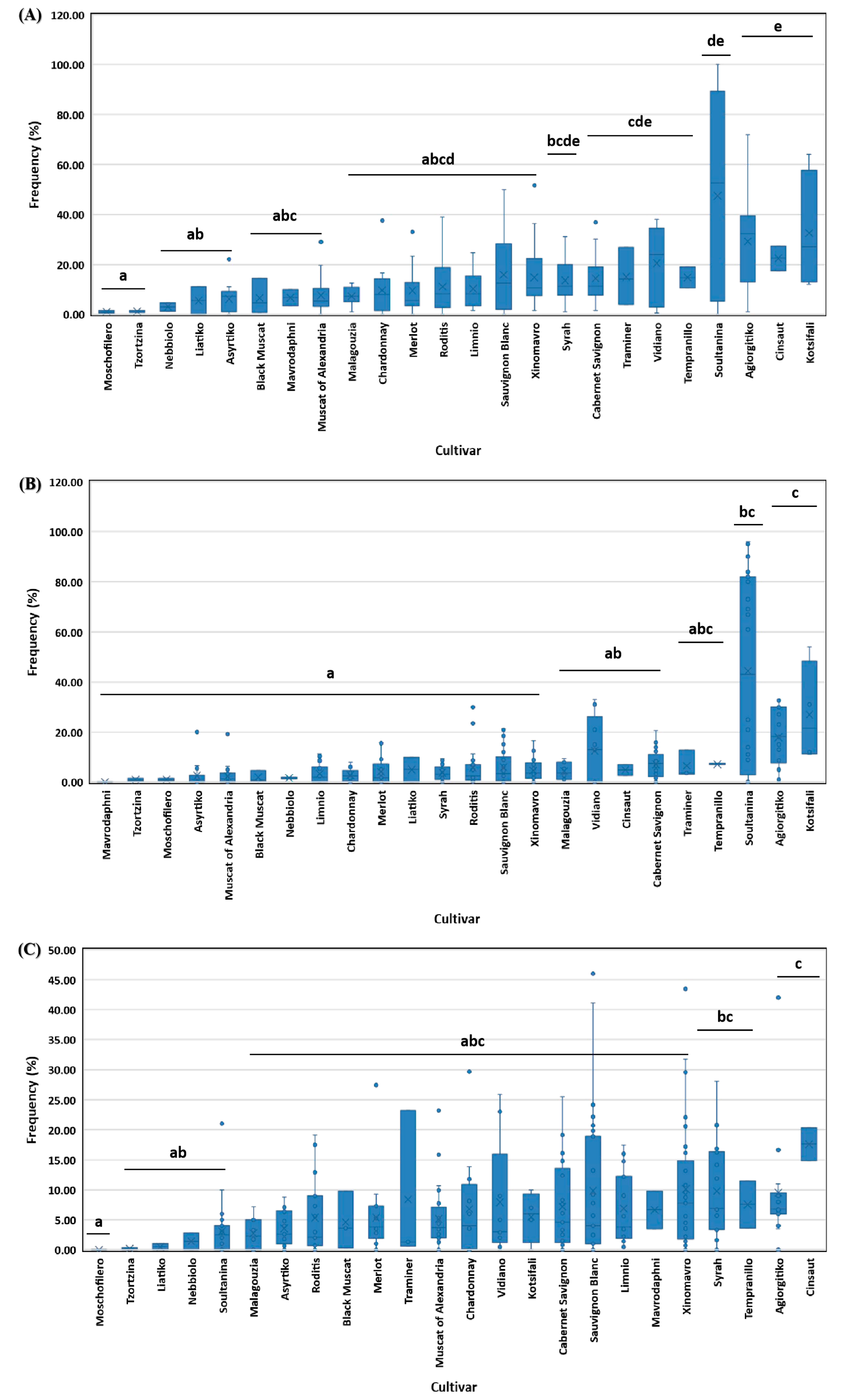
Figure 5.
Frequencies of isolates belonging to discrete fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece.
Figure 5.
Frequencies of isolates belonging to discrete fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece.
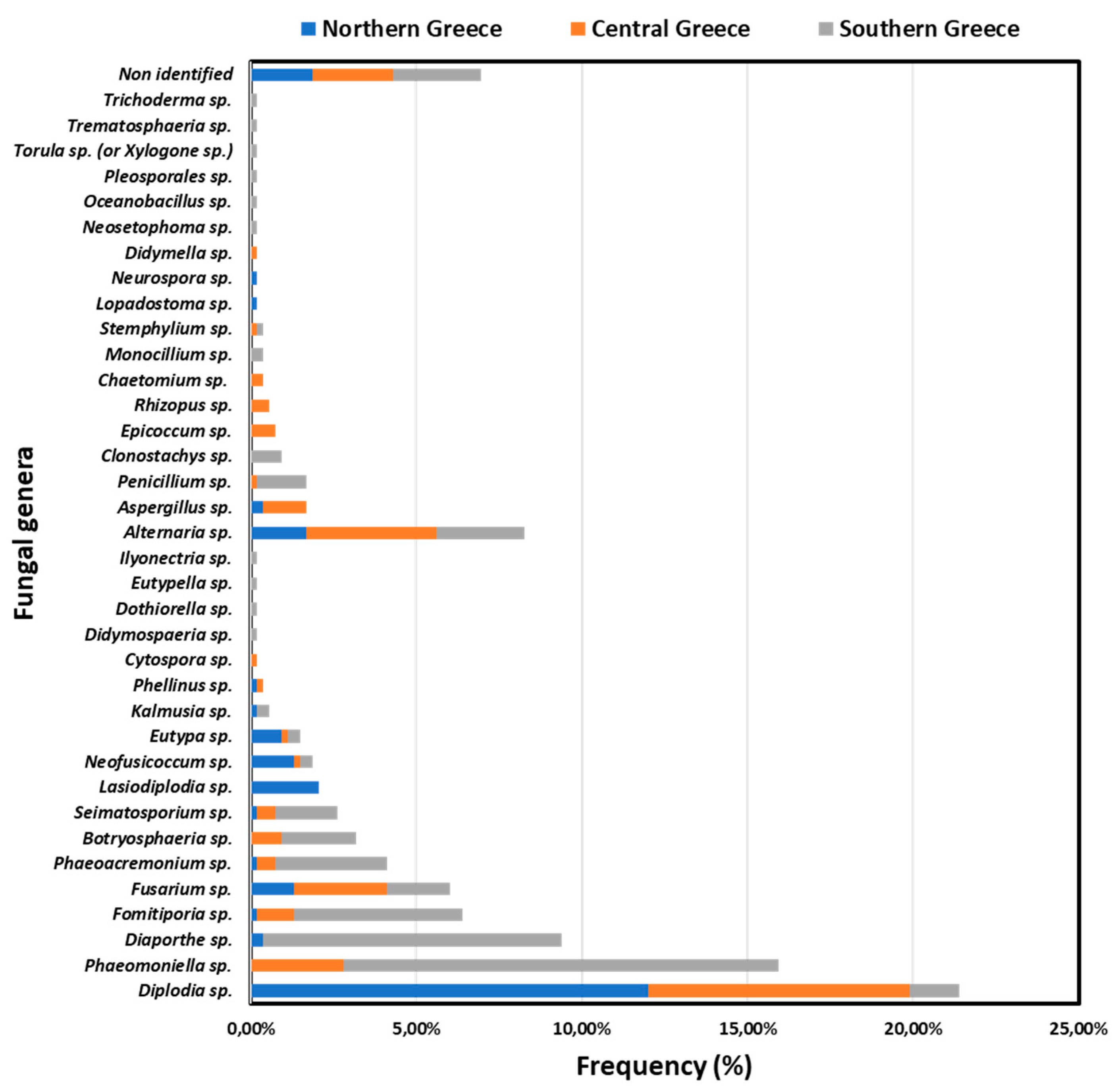
Figure 6.
Frequencies of isolates belonging to 18 discrete fungal genera associated with grapevine trunk diseases, originated from young (0-9 years old), middle-aged (10-19) and elder (≥20 years old) vineyards in Greece.
Figure 6.
Frequencies of isolates belonging to 18 discrete fungal genera associated with grapevine trunk diseases, originated from young (0-9 years old), middle-aged (10-19) and elder (≥20 years old) vineyards in Greece.
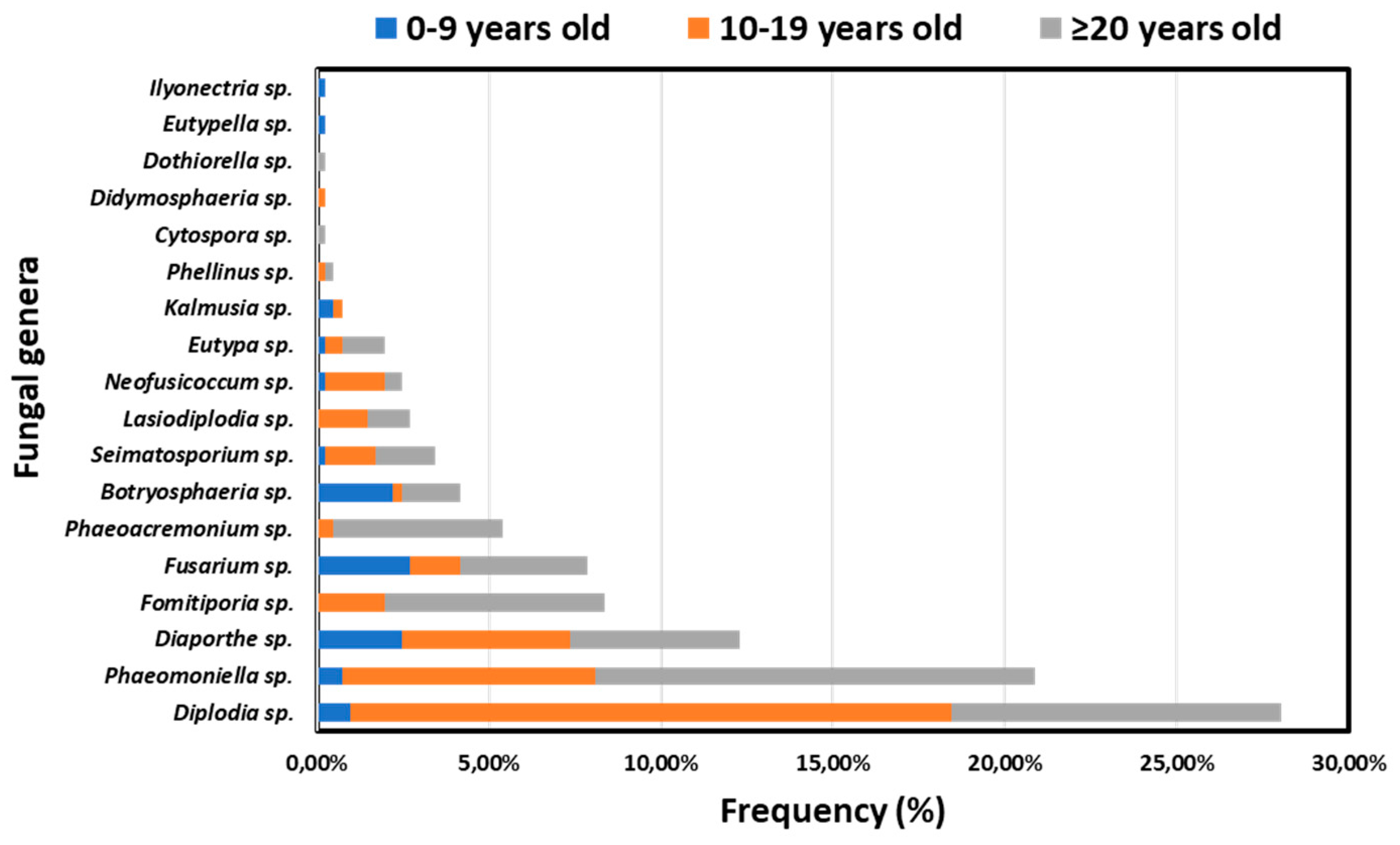
Figure 7.
Percentage of vineyards infested by each fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece. Within each genus, columns with the same letter do not differ significantly according to Chi-square test and a pair-wise comparison, whereas asterisks (*, ** and ***) indicate significance at p ≤ 0.05 , 0.01 and 0.001, respectively.
Figure 7.
Percentage of vineyards infested by each fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece. Within each genus, columns with the same letter do not differ significantly according to Chi-square test and a pair-wise comparison, whereas asterisks (*, ** and ***) indicate significance at p ≤ 0.05 , 0.01 and 0.001, respectively.
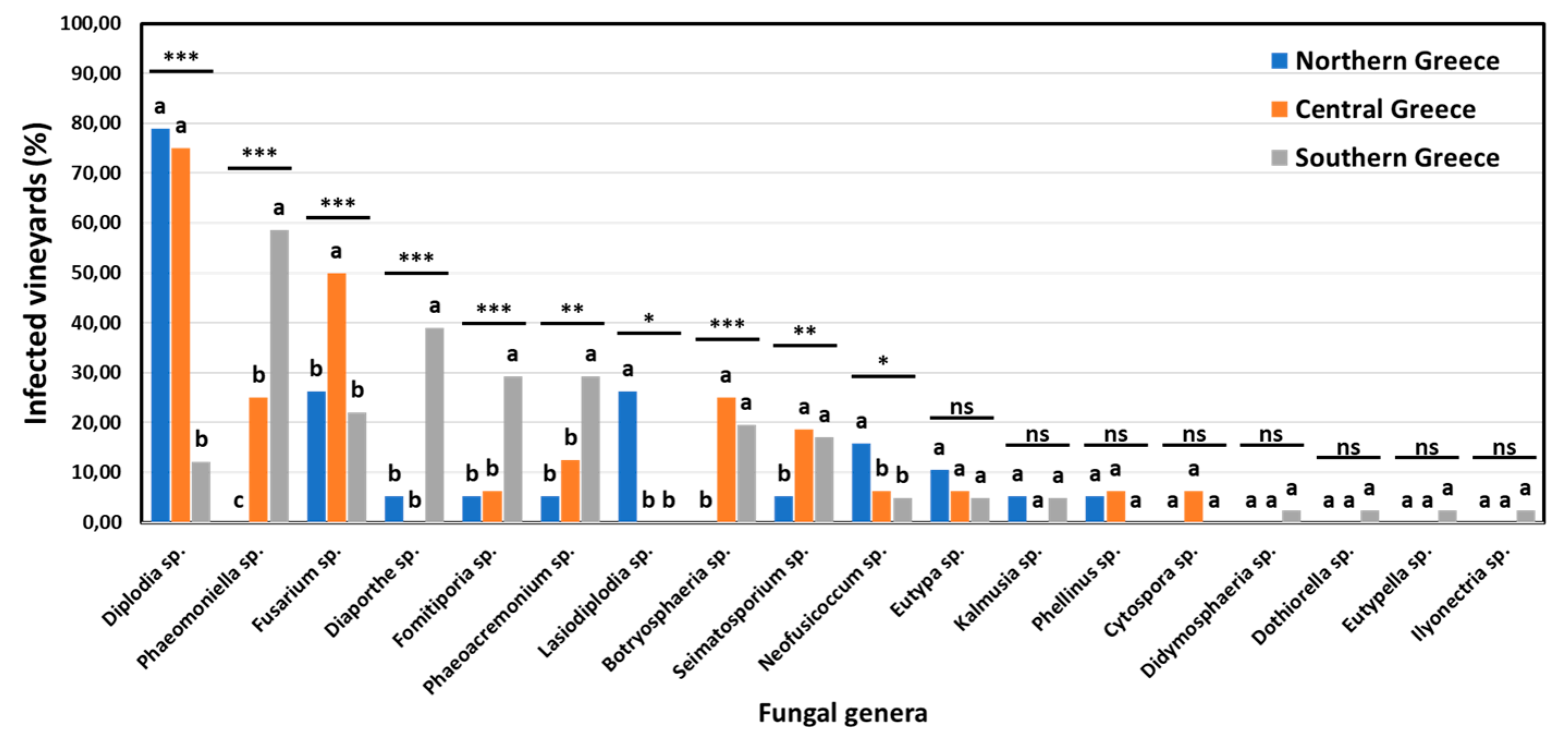
Figure 8.
Percentage of vines infested by each fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece. Within each genus, columns with the same letter do not differ significantly according to Chi-square test and a pair-wise comparison, whereas asterisks (*, ** and ***) indicate significance at P ≤ 0.05, 0.01 and 0.001, respectively.
Figure 8.
Percentage of vines infested by each fungal genera in northern (Amyntaio and Kavala), central (Nemea) and southern (Heraklion) Greece. Within each genus, columns with the same letter do not differ significantly according to Chi-square test and a pair-wise comparison, whereas asterisks (*, ** and ***) indicate significance at P ≤ 0.05, 0.01 and 0.001, respectively.
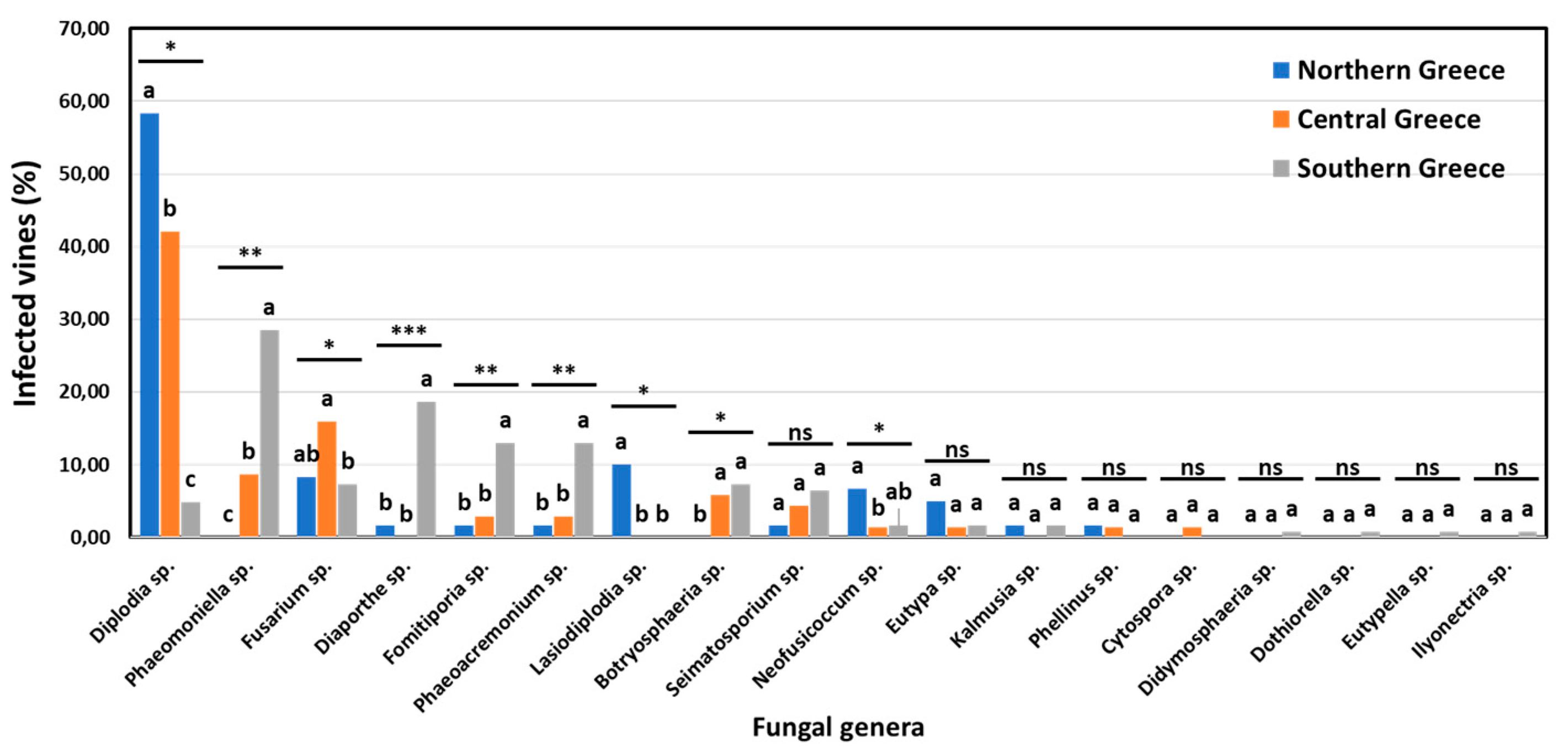
Figure 9.
Principal component analysis (PCA) biplot illustrating the correlation between meteorological data (air temperature, RH, and rainfall) from past decades (2010-2020) and the identified fungal genera isolated from symptomatic grapevines in the viticultural zones of southern, central, and northern Greece.
Figure 9.
Principal component analysis (PCA) biplot illustrating the correlation between meteorological data (air temperature, RH, and rainfall) from past decades (2010-2020) and the identified fungal genera isolated from symptomatic grapevines in the viticultural zones of southern, central, and northern Greece.
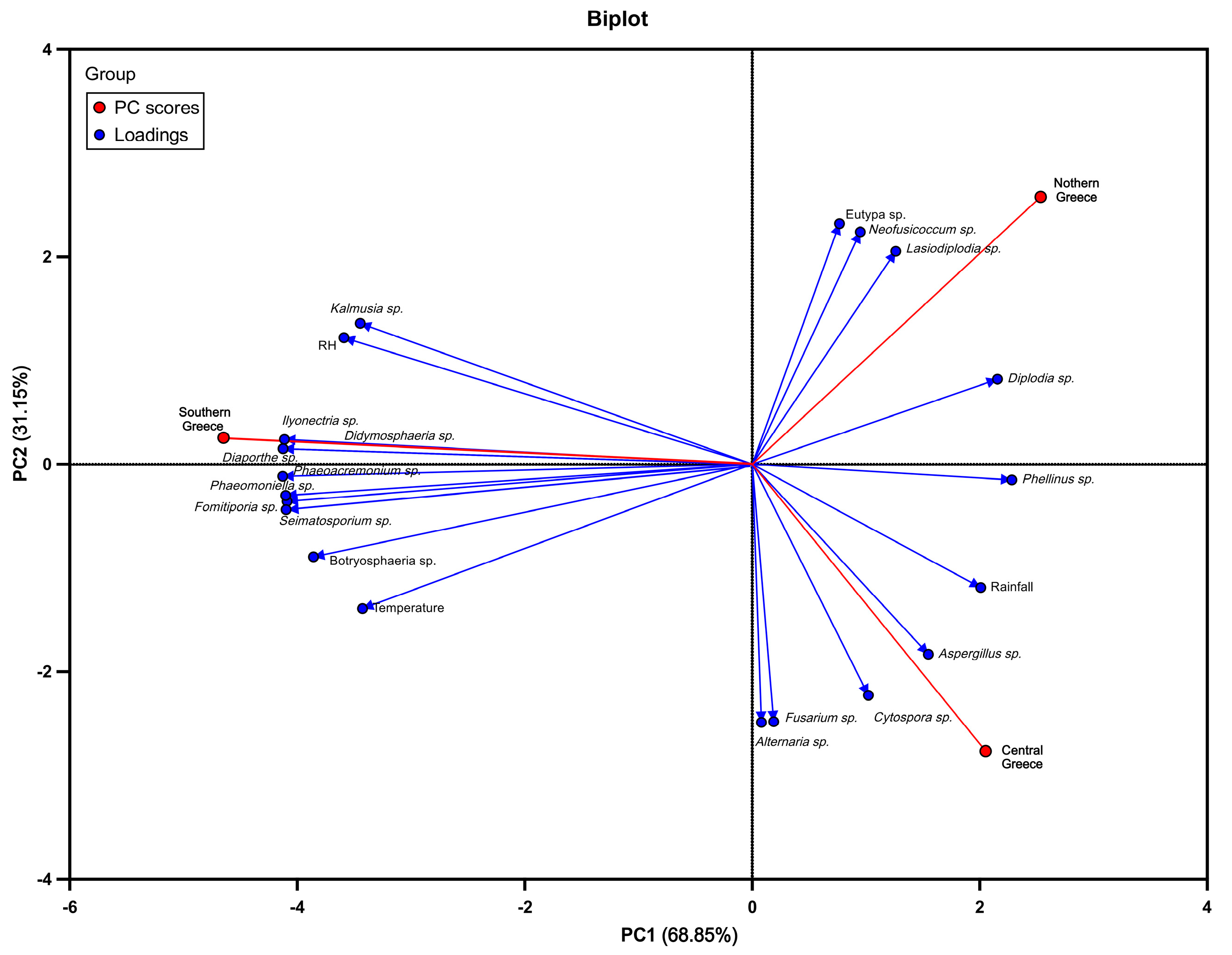
Figure 10.
Length of vascular tissue discoloration in 2-year-old canes of cv. Soultanina, 6 months after artificial inoculation with Seimatosporium vitis (isolate SARO1.3AVR10), Didymosphaeria variabile (isolate SARO1.2AKO1), Kalmusia variispora (isolate HOURD2.1AVR1) and Neosetophoma italica (isolate SARO1.3AVR7). Control canes (C-) were mock-inoculated with sterilized PDA discs. Columns followed by the same letter do not differ significantly according to Tukey honestly significant difference test (P ≤ 0.05). Each column represents the mean of 20 canes and vertical bars indicate standard errors.
Figure 10.
Length of vascular tissue discoloration in 2-year-old canes of cv. Soultanina, 6 months after artificial inoculation with Seimatosporium vitis (isolate SARO1.3AVR10), Didymosphaeria variabile (isolate SARO1.2AKO1), Kalmusia variispora (isolate HOURD2.1AVR1) and Neosetophoma italica (isolate SARO1.3AVR7). Control canes (C-) were mock-inoculated with sterilized PDA discs. Columns followed by the same letter do not differ significantly according to Tukey honestly significant difference test (P ≤ 0.05). Each column represents the mean of 20 canes and vertical bars indicate standard errors.
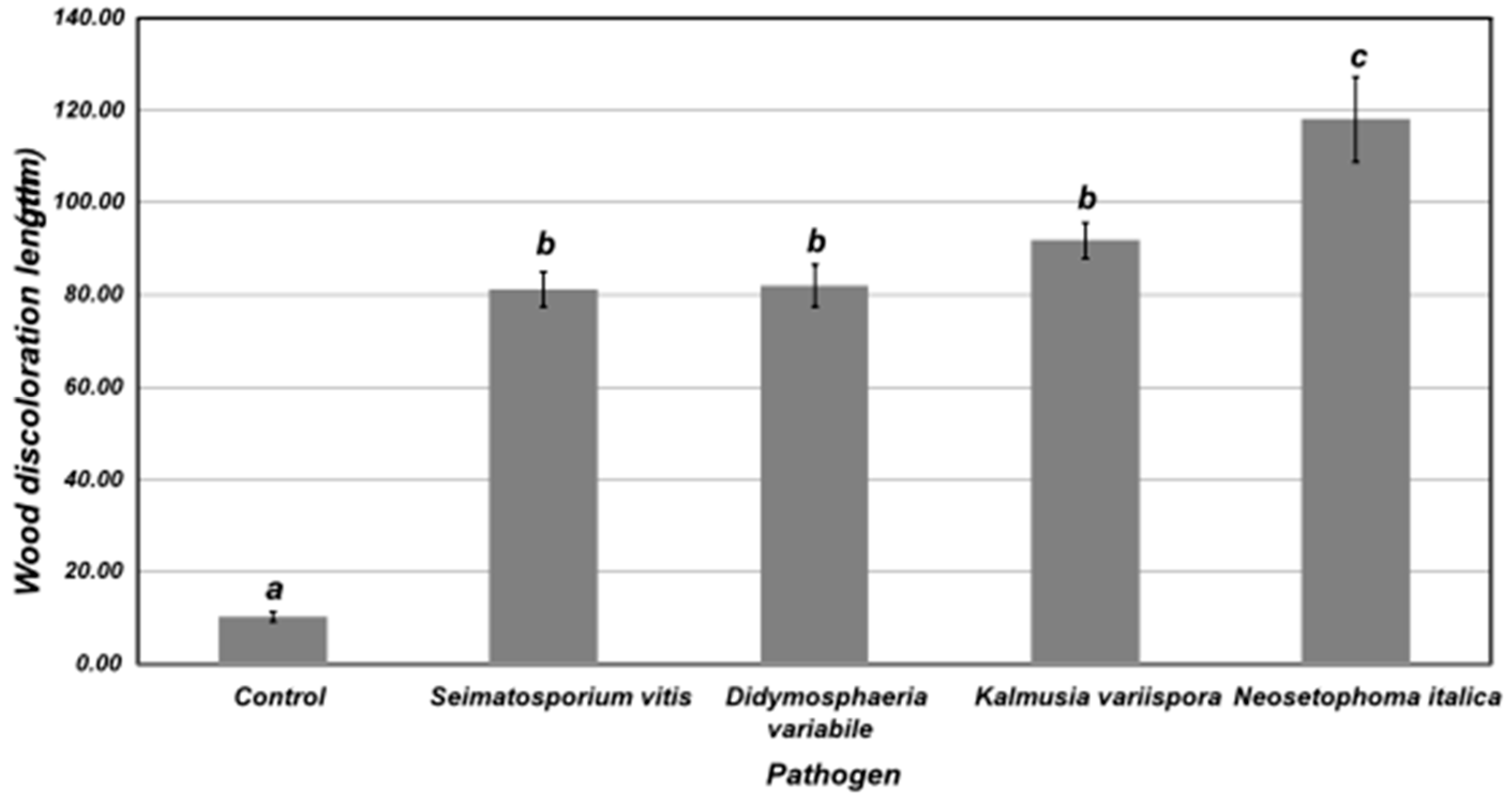

Table 1.
Numerical distribution of vineyards located in seven (7) important grapevine-growing regions in three geographically distinct viticultural zones (northern, central and southern Greece) falling into three (3) age-scales, surveyed for the assessment of GTD disease parameters.
Table 1.
Numerical distribution of vineyards located in seven (7) important grapevine-growing regions in three geographically distinct viticultural zones (northern, central and southern Greece) falling into three (3) age-scales, surveyed for the assessment of GTD disease parameters.
| Viticultural zone | Region | Number of vineyards | Vineyard age (years) | ||
|---|---|---|---|---|---|
| <10 | 10-19 | ≥20 | |||
| Northern Greece | Amyntaio | 78 | 10 | 39 | 29 |
| Kavala | 60 | 8 | 27 | 25 | |
| Lemnos | 51 | 21 | 14 | 16 | |
| Naousa | 20 | 0 | 17 | 3 | |
| Nea Mesimvria | 25 | 9 | 7 | 9 | |
| Central Greece | Nemea | 28 | 3 | 8 | 17 |
| Southern Greece | Heraklion | 48 | 19 | 4 | 25 |
| Total | 310 | 70 | 116 | 124 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



