
Submitted:
23 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Bacteria in the gut microbiome play an intrinsic part in immune activation, intestinal permeability, enteric reflex, and entero-endocrine signaling. Gut microbiota communicates with the central nervous system (CNS) through the production of bile acids, short-chain fatty acids (SCFAs), glutamate (Glu), γ-aminobutyric acid (GABA), dopamine (DA), norepinephrine (NE), serotonin (5-HT) and histamine. A vast number of signals generated in the gastrointestinal tract (GIT) reaches the brain via afferent fibers of the vagus nerve (VN). Signals from the CNS are returned to entero-epithelial cells (EES) via efferent VN fibers and communicate with 100 to 500 million neurons in the submucosa and myenteric plexus of the gut wall, which is referred to as the enteric nervous system (ENS). Intercommunications between the gut and CNS regulate mood, cognitive behavior, and neuropsychiatric disorders such as autism, depression, and schizophrenia. The modulation, development, and renewal of nerves in the ENS, and changes in the gut microbiome alter the synthesis and degradation of neurotransmitters, which ultimately influence our mental health. The more we decipher the gut microbiome and understand its effect on neurotransmission, the closer we may get to the developing of novel therapeutic and psychobiotic compounds to improve cognitive functions and prevent mental disorders. In this review, the intricate control of entero-endocrine signaling and immune responses that keep the gut microbiome in a balanced state, and the influence that changing gut bacteria have on neuropsychiatric disorders are discussed.
Keywords:
Gut microbiota
; enteric nervous system
; mental health
1. Introduction
Most neurological signals to and from the gut run through a bi-directional vagus nerve (VN) that exits the brain at the medulla oblongata. Signals from the central nervous system (CNS), passed along efferent VN fibers, reach 100 to 500 million neurons in the ENS [1] that surround the gastrointestinal tract (GIT). The enteric nervous system (ENS), often referred to as the “second brain”, is by far the largest of all nerve systems in the human body and functions independently from the VN [2]. Experiments on adult mice have shown that myenteric neurons have a short life span and are subjected to infrequent episodes of apoptosis [3]. According to Kulkarni et al. [4], more than 85% of myenteric neurons in the small intestine of mice are replaced within two weeks but are replaced at a constant rate. Newly formed neurons within myenteric ganglia express the neuroepithelial stem cell protein nestin, the nuclear protein Ki67, and the neurotrophin receptor p75NTR [4]. Nestin maintains the balance between neuronal apoptosis and neurogenesis [4,5], Ki67 is associated with rRNA transcription and p75NTR decreases the activation of Rho, thus favoring axonal elongation [6]. A decline in the health of neural stem cells (NSCs) will ultimately lead to a deficiency in the production of any of these proteins and the inability to renew damaged cells [7].
To fully understand the interaction between the GIT and the brain and how the gut-brain axis functions, it is important to understand the role of intrinsic and extrinsic sensory neurons in the ENS. Neurons in the ENS are interconnected via internodal strands with axons up to 13 cm in length to form two distinct ganglionated neuronal plexuses, referred to as the myenteric and submucosal plexuses [8]. Smooth muscle cells of the GIT are in close contact with excitatory and inhibitory motor neurons [9,10,11]. Motor neurons in the myenteric plexus coordinate muscle movements (peristalsis), while neurons in the submucosal plexus regulate secretion and absorption [8]. The major extrinsic neural pathways between the ENS and the CNS are shown in Figure 1. Vagal dorsomotor efferent fibers connect with parasympathetic enteric nerves of the stomach and upper part of the small intestine. Enteric nerves in the rest of the small intestine are connected to sympathetic nerves in the coeliac and superior mesenteric ganglia. Enteric nerves of the large intestine and distal part of the colon are connected to sympathetic efferent nerves in the inferior mesenteric ganglia and parasympathetic nerves in the pelvic ganglia. Motor and sensory neurons in the submucosa and myenteric plexus of the gut wall regulate muscle activity, gut wall motility, secretion of fluids, and blood flow [12]. The large intestine, specifically the distal colon, has afferent nerves linked to the spinal cord [13]. These signals reach enterochromaffin cells (ECs) and enteroendocrine cells (EECs) that are in direct contact with an estimated 4 trillion gut microorganisms. The celiac branch of the VN connects with the duodenum and the rest of the intestine to the distal part of the descending colon [14]. This intricate connection of neurons facilitates a rapid transfer of signals throughout the GIT [15,16]. The modulation, development, and renewal of neurons of the ENS are controlled by gut microbiota, especially those with the ability to produce and metabolize hormones (reviewed by Dicks [17]).
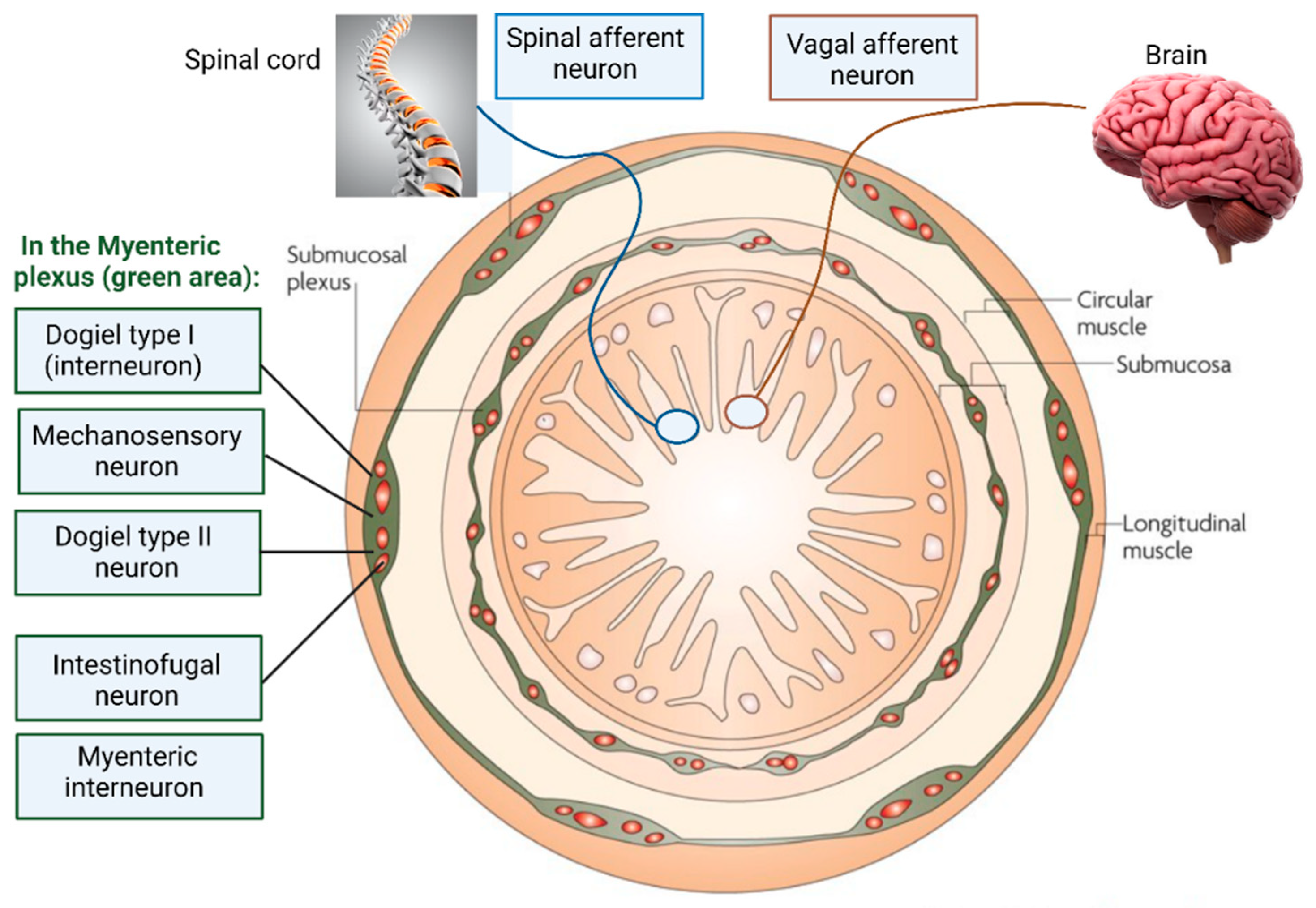
Dogiel type I neurons (Figure 2) are in the myenteric plexus of the human colon [18] and act as interneurons or motor neurons. Based on experiments conducted on the colon of guinea pigs, Dogiel type I neurons are short and are characterized by having one axon and up to 20 dendrites with broad or lamellar endings [19]. Dogiel type I neurons react to sensory stimuli from the GIT, but also synaptic signals received from other neurons [20,21]. Dogiel type II neurons (Figure 2) have a single axon with up to 16 long dendrites and react to chemical and mechanical stimuli, rendering them sensitive to changes in muscle tension [8,22]. Dogiel type II neurons may react on the release of substances such as serotonin from enterochromaffin cells (ECs) but there is no direct evidence supporting this [23].
At least 10 different types of EECs have been characterized. Apart from regulating the secretion of serotonin, they are involved in the secretion of chromogranin/secretogranin, neuropeptide Y (NPY), vasoactive intestinal peptide (VIP), cholecystokinin (CCK), somatostatin, glucagon-like peptide (GLP)-1/2, ghrelin, and substance P (SP) [24,25]. Receptors on these sensory cells are expressed by gut enteric neurons, but also vagal afferents, the brainstem, and the hypothalamus [26,27]. For more information on sensorial signals, neural circuits, and gut peristalsis regulated by the ENS, the reader is referred to the review by Spencer and Hu [28].
Signals generated by gut microbiota, either in response to signals received from the CNS or other organs, or generated because of gastrointestinal metabolic reactions, are returned to the brain via afferent VN fibers [14]. Afferent fibers outnumber efferent fibers by 9:1, which indicates the importance of signaling from the GIT to the CNS. Molecules produced by gut microbiota regulate the CNS, entero-endocrine pathways, and the immune system. These regulatory or signaling molecules range from simple compounds such as bile acids, short-chain fatty acids (SCFAs), glutamate (Glu), aspartate, D-serine, and histamine to more complex structures such as γ-aminobutyric acid (GABA), dopamine (DA), norepinephrine (NE, also called noradrenaline, NAd), and serotonin (5-HT) [13,29,30,31]. For more information on neurotransmitters and the modulation, development, and renewal of ENS neurons, the reader is referred to the review by Dicks [17]. Communication amongst gut microbiota via quorum sensing (QS) signals and the impact of these signals on the CNS, including mental health, is reviewed by Dicks [32] and will be briefly discussed. The CNS is not discussed in detail.
2. Maintenance of the ENS
The ENS stems from enteric neural crest cells (ENCCs) [33]. The proliferation and migration of ENCCs are regulated by the glial cell line-derived nerve growth factor (GDNF) protein that is predominantly expressed by neurons in the septum, striatum, and thalamus [33]. The expression of GDNF and other neurotrophic factors, such as neurturin (NTN), artemin (ART), and persephin (PSP) are regulated by Toll-like receptors TLR2, TLR4, TLR5, and TLR9 [34]. GDNF, NTN, ART, and PSP bind to growth factor receptors (GFR)α-1, GFRα-2, GFR α3, and GFRα-4, respectively, all attached to the transmembrane receptor tyrosine kinase (RET) located on the cell membrane. The GFRα1–GDNF complex changes to the active tyrosine-phosphorylated form which then signals to ENCCs to express ENS precursors and neurons [35].
Phosphatidylinositol 3-kinase (EC 2.7.1.137) and Akt (protein kinase B) in the PI3K/AKT signaling pathway stimulate neural survival and transmission. Phospholipase C gamma 1 (PLC-γ1; EC 3.1.4.3) is involved in cell growth, apoptosis, and transmission of neural cells. A ras/mitogen-activated protein kinase (RAS/MAPK) regulates neurogenesis [4,36]. ENS cells destroyed by apoptosis are replaced by newly formed cells [4]. However, little is known about the mechanisms that control ENS cell replenishment. Vicentini et al. [37] have shown that the small intestinal tract of antibiotic-induced mice (thus without gut microbiota) was longer than that observed in normal mice. This led to slower transit of gut contents, increased carbachol (carbamylcholine)-stimulated ion secretion, and an increase in gut wall permeability. Increased levels of carbachol activate acetylcholine (Ach) receptors [38] stimulate the formation of muscarinic and nicotinic receptors. Ach is the main neurotransmitter of the parasympathetic nervous system and plays a role in the contraction of smooth muscles, dilation of blood vessels, secretion of bodily fluids, and the downregulation of heart rate. Muscarinic Ach receptors form G protein-coupled receptor complexes in the membranes of parasympathetic nerve cells [38]. Polypeptide nicotinic acetylcholine receptors are present in the central and peripheral nervous system, muscle, and many other tissues [39]. Vicentini et al. [37] also noted a decrease of neurons in the submucosal and myenteric plexuses of the ileum and proximal colon of germ-free mice. The myenteric plexus of the ileum contained fewer glial cells (neuroglia) [37], which is an indication that neurons of the ENS had been deprived of nutrient and oxygen supply. Neuronal cells that were not insulated from each other did not destroy pathogens and could not remove dead neurons [40].
Research conducted on experimental animals has shown that Ach is stored in vesicles at the terminals of Ach-producing neurons. The secretion of Ach is most likely stimulated by certain Lactobacillus spp [41,42]. A decrease in the production of Ach was recorded when blood flow was restricted at the proximal cerebral artery, which altered the composition of gut microbiota [43,44]. Furthermore, a decrease in the production of Ach resulted in irregular peristalsis, dysbiosis, an increase in gut permeability [43,44], and inflammation of the GIT [45]. This, in turn, restricted the middle cerebral artery which led to a decline in goblet cells in the cecum and a lowering of mucin production [45]. The influence that a single neurotransmitter such as Ach has on the parasympathetic nervous system, and the gut microbiome, illustrates the sensitivity of ENS to changing conditions.
3. The influence of gut bacteria on the ENS
The human gut is host to 2766 microbial species, of which more than 90% belong to the phyla Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes [46,47,48,49,50,51,52]. The remaining 10% of the gut microbiome is composed of Fusobacteria and Verrucomicrobia [53]. Gut bacteria communicate with the CNS by using GABA, DA, NE, 5-HT, histamine [29], SCFAs [30], tryptophan [31], and secondary bile acids [13]. Signals generated by these neurotransmitters and molecules are transported to the CNS via afferent VN fibers. Signals from the brain reach ECCs and EECs in the gut wall, and the mucosal immune system via efferent VN fibers [54]. This bidirectional communication improves the integrity of the gut wall, reduces peripheral inflammation, and inhibits the release of pro-inflammatory cytokines [55]. On another level, signals generated by the hypothalamus reach the pituitary and adrenal glands and communicate with EECs via the hypothalamic pituitary adrenal axis (HPA) [56]. The intricate control of entero-endocrine signaling and immune responses keeps the gut microbiome in a balanced state and prevents dysbiosis or the development of major GI disorders such as diarrhoea, ulcerative colitis (UC), Crohn’s disease, and inflammable bowel diseases (IBDs) [57,58,59]. Toxins produced by pathogenic bacteria may alter the functioning of the intestinal- and blood-brain barrier (BBB), and lead to neurodegeneration [60,61,62,63].
The importance of gut microbiota, lipopolysaccharides (LPS), and SCFAs in the recovery of enteric neurons was illustrated by experiments conducted on mice [37]. It is, however, important to note that LPS stimulated the recovery of damaged neurons and the proliferation of gut microbiota but not the formation of new neurons. SCFAs, on the other hand, restored neuronal loss [37]. Concluded from these results, a decrease in SCFAs, as expected in patients with dysbiosis, may lead to a loss of enteric neurons and a weakened ENS. This emphasizes the importance of SCFA-producing gut microbiota. SCFAs, such as butyrate, acetate, lactate, and propionate, are largely produced in the colon by Bifodobacterium, Lactobacillus, Lachnospiraceae, Blautia, Coprococcus, Roseburia and Faecalibacterium [64]. SCFA receptors such as GPR43(FFAR2) and GPR41 (FFAR3) are located on the surface of IECs [65] and are also expressed in the ENS, portal nerve, and sensory ganglia [5]. GPR 41 in the ENS transfers signals induced by SCFAs directly to the CNS [66]. GPR43 communicates with SCFAs to stimulate energy expenditure in skeletal muscles and the liver [67].
Glutamate (Glu) released from presynaptic nerve terminals adheres to ionotropic glutamate receptors (iGluRs) located on postsynaptic terminals. This increases the transfer of Ca2+ through voltage-activated Ca channels (VACCs) located in the terminals and activates calcium calmodulin-dependent kinase (CaMK; EC 2.7.11.17), extracellular signal-regulated kinase (ERK, EC 2.7.11.24) and the cyclic AMP response element binding (CREB) protein [68]. Excess Glu is converted to glutamine, which is transported back to presynaptic nerve terminals and converted to Glu by glutaminase (EC 3.5.1.2). The newly formed Glu is then transported to vesicles in the presynaptic neuron by the vesicular glutamate transporters VGLUT1 and VGLUT2. This process is carefully controlled, as Glu excitotoxicity accelerates the progression of Alzheimer’s disease [69]. L-Glutamate (L-Glu) is also converted to D-Glu by Corynebacterium glutamicum, Lactobacillus plantarum, Lactococcus lactis, Lactobacillus paracasei, Brevibacterium avium, Mycobacterium smegmatis, Bacillus subtilis and Brevibacterium lactofermentum [49,70,71]. D-Glu is then decarboxylated by Glu decarboxylase (GAD; EC 4.1.1.15) to GABA [72,73]. The adhesion of GABA to GABA receptors (GABARs) on postsynaptic neurons prevents the transfer of Na+, K+, Ca2+ and Cl- [74]. Lactobacillus rhamnosus JB-1 altered the expression of GABARs in the brain, which led to a decrease in anxiety and depression [75]. Treatment with Lactobacillus elevated GABA levels in the hippocampal and prefrontal cortex [76].
GABA, also present in millimolar concentrations in the brain, is released into the synaptic cleft upon depolarization of presynaptic neurons [77,78]. In these neurons, GABA α-oxoglutarate transaminase (GABA-T, EC 2.6.1.19) converts α-ketoglutarate to L-glutamic acid, which is then decarboxylated to GABA by GAD. Glial cells do not express GAD and cannot decarboxylate Glu to GABA. Glial cells convert GABA to succinic semialdehyde and glutamine, which is then deaminated to glutamate before it re-enters the GABA shunt [77].
Acetate produced by microbiota in the colon is transferred across the BBB and enters GABA neuroglial cycling pathways [79]. Gut-derived GABA reaches the CNS via specific GABA transporters expressed in the BBB, of which at least six have been identified [80]. These GABA transporters regulate the uptake of GABA into presynaptic nerve terminals and surrounding glial cells [81]. The re-uptake of GABA by neurons results in a decreased uptake of Na+. Akkermansia muciniphila, Parabacteroides merdae and Parabacteroides distasonis alter GABA and glutamate ratios and increase glutamate levels in the brain [82].
Dopamine (DA) is produced in the substantia nigra, ventral tegmental area and hypothalamus. Although DA is generally referred to as the reward neurotransmitter, it also plays a role in behavior and cognition, voluntary movement, motivation, inhibition of prolactin production, sleep patterns, mood, attention, working memory, and learning [83]. Tyrosine is hydroxylated to L-dihydroxyphenylalanine (L-DOPA) and then decarboxylated to DA. In the presence of β-hydroxylase, DA is converted to NE and Epi (Ad) [84]. DA is also produced by certain Bacillus and Serratia species in the GIT [85]. Enterococcus faecalis decarboxylates L-DOPA to DA, but is then immediately dehydroxylated to m-tyramine by Eggerthella lenta [86].
Norepinephrine (NE) or Noradrenaline (NAd) is structurally similar to EPi (Ad). NE is produced during excitement but also plays a role in behavior, cognition [87], and inflammatory responses of the autonomic nervous system [88]. Low levels of NE (0.45 and 2.13 mM ) are produced by Bacillus mycoides, Bacillus subtilis, E. coli K12, Proteus vulgaris and Serratia marcescens [89]. An interesting finding was that NE stimulates the growth of Klebsiella pneumoniae, Shigella sonnei, Pseudomonas aeruginosa, Enterobacter cloacae, and Staphylococcus aureus [90].
Enteric serotonin levels are regulated by tryptophan hydroxylase TPH1 and serotonin produced from the ENS by tryptophan hydroxylase TPH2 [91]. Serotonin (5-HT) regulates appetite, gut motility, mood, cognition and sleeping patterns [92,93,94]. As much as 80% 5-HT is produced in the GIT by E. coli, Hafnia, Bacteroides, Streptococcus, Bifidobacterium, Lactococcus, Lactobacillus, Morganella, Klebsiella, Propionibacterium, Eubacterium, Roseburia and Prevotella [92,93]. Serotonin production by gut microbiota may have a greater effect on the CNS than originally anticipated, as ECs interact with 5-HT-receptive afferent fibers in vagal or dorsal root neurons [95]. Physiological concentrations of SCFAs can cause an eight- to ten-fold increase in serotonin production, as shown in an in vitro colonic mucosal system [58]. The production of serotonin is carefully controlled by transporting excess concentrations across the cell membrane by a serotonin reuptake transporter (SERT), followed by the conversion to an inactive form by monoamine oxidase (MAO) [91]. This is important, as high levels of serotonin may decrease the permeability of the gut wall [96]. Low levels of serotonin, on the other hand, lead to a decrease in the expression of occludin, which increases gut wall permeability. The latter was reported in patients diagnosed with IBS [96]. For further information on the effect of 5-HT on the ENS, the reader is referred to the review by Dicks [17].
Minor activation of the VN results in drastic changes in the production of neurotransmitters, which affects digestion, intestinal permeability, gastric motility, and immune regulation. This, in turn, alters the microbial composition in the GIT and may benefit the survival and proliferation of certain species. Neurohormones that may be affected by altering gut microbiota are oxytocin (Oxt) produced by the hypothalamus, and vasopressin, also known as antidiuretic hormone (ADH) [97]. As soon as a neurotransmitter binds to the receptor on the postsynaptic membrane, the rate at which Ca2+, Na+, K+, and Cl- are transferred across ligand-gated channels changes, which results in either a stimulatory- or an inhibitory response [97]. These intricate and highly interweaved signaling mechanisms help keep the gut microbiome in a balanced state. If unbalanced, the gut enters a state of dysbiosis, characterized by an increase in Enterobacteriaceae, especially Escherichia, Shigella, Proteus, and Klebsiella [60]. If left untreated, gastrointestinal disorders such as diarrhea, ulcerative colitis (UC), Crohn’s disease, and other inflammable bowel diseases (IBDs) may develop [57,58,59]. In severe cases, toxins produced by these bacteria may alter the functioning of the gut-blood barrier (GBB) and blood-brain barrier (BBB), ultimately leading to neurodegeneration [60,61,62,63].
4. Mental Health
Several research studies have shown that dysbiosis is a contributing factor to the development of neurological and psychiatric diseases. These include anxiety, depression, major depressive disorder (MDD), schizophrenia, bipolar disorder, autism, and obsessive-compulsive disorder (OCD) [98]. Acetate, lactate, butyrate, and propionate produced by anaerobic bacteria in the large intestine have a profound effect on reactions causing mental health issues. Experiments on animals have shown that butyrate inhibits histone deacetylase (HDAC) [56,99]. Overproduction of HDAC is associated with neurological disorders such as Parkinson’s disease, schizophrenia, and depression [99]. The inhibition of HDAC may thus be important in the treatment of brain trauma and dementia [99]. An increase in acetylated histones (ACHs), on the other hand, elevates the expression of the bdnf (brain-derived neurotrophic factor) gene in the frontal cortex and hippocampus and stimulates brain development [100,101]. Low levels of BDNF are associated with mood changes, depression, and anxiety [94,102,103,104]. Activation of G-protein-coupled receptors (GPCRs) by butyrate may lead to the development of multiple neurodegenerative disorders [56,99] and an increase in the production of inflammatory cytokines [105]. Butyrate (and other SCFAs) modifies the integrity of the blood-brain barrier (BBB) and the maturation of microglia [106,107]. In germ-free (GF) mice, the malfunctioning of microglia could be reversed by administering high levels of a combination of butyrate, propionate and acetate [108]. The function of acetate is different in that it crosses the BBB and accumulates in the hypothalamus where it controls appetite [79].
Oscillobacter spp. are known to produce valeric acid, a compound that closely resembles GABA and binds to GABA receptors [109]. With the binding of valeric acid to these receptors, GABA binding is inhibited, and the CNS signals are no longer blocked, resulting in anxiety. Of interest is that patients who suffer from anxiety and depression have lower Lactobacillus cell numbers. This is an important observation, as certain Lactobacillus spp. stimulates the secretion of GABA as well as the neurotransmitter acetylcholine [110,111].
Hormone production regulated by gut microbiota interacts with EECs [112,113] and generates chemical signals that, in turn, react with the ENS. Bodily functions affected by changes in hormone levels include digestion, salivation, lacrimation, urination, defecation, and sexual arousal [114]. A clear association exists between chronic stress and gut inflammation disorders, such as IBD and IBS [115].
Inflammatory factors produced by Alistipes spp. could play a role in depression, anxiety, IBD and chronic fatigue [116,117]. Prevotella spp. are often associated with pro-inflammatory responses) characteristics but low numbers of the species have also been associated with anxiety and depression. [118]. Faecalibacterium prausnitzii (ATCC 27766) relieved anxiety and depression, suggesting that the strain may have psychobiotic properties [119]. This may have to do with increased levels of SCFA in the cecum, elevated plasma IL-10 levels, and lower levels of corticosterone and IL-6 [120]. Ruminococcus flavefaciens upregulated genes involved in mitochondrial oxidative phosphorylation, whilst downregulating genes involved in synaptic signaling and neurogenesis [121]. Significantly lower levels of Proteobacteria, Haemophilus, Sutterella, and Clostridium spp. were reported in schizophrenic patients, whilst cell numbers of Anaerococcus spp. remained unchanged. Not all studies came to the same conclusion. In another study [122], high levels of Proteobacteria, Succinivibrio, Collinsella, Clostridium and Klebsiella spp., but low levels of Blautia, Coprococcus, and Roseburia spp. were reported in schizophrenic patients [123,124]. Concluded from the study conducted by Qing et al. [125], Firmicutes live in synergy with actinobacteria, fusobacteria, and Acidobacteria, especially during the early stages of schizophrenia. High cell numbers of Firmicutes were reported to be present in the saliva of patients with primary Sjögren’s syndrome [126]. High levels of lactic acid bacteria, especially Lactobacillus gasseri, were reported in schizophrenic patients. Bacteriodetes and Acinetobacteria, on the other hand, were in the minority [127]. In contrast to previous studies, the presence of Proteobacteria did not differ significantly between schizophrenic and non-schizophrenic patients.
Anaerococcus and Collinsella isolated from schizophrenic individuals are known to produce high levels of butyrate [123,124]. In bipolar and autistic patients butyrate-producing Faecalibacterium spp. was present in low numbers [128]. Patients with bipolar disorder are often diagnosed with an increase in gut wall permeability [129] and an increase in Flavonifractor [130]. Species in this genus cleaves the flavonoid quercetin [130]. The latter has anti-oxidative and anti-inflammatory properties [131] and changes in its levels could lead to bipolar disorder. Other changes in bacterial populations included a decrease in Faecalibacterium , and an increase in Actinobacteria and Coriobacteriaceae [132]. Evans et al. [133] reported a decrease in Faecalibacterium and Ruminococcaceae in bipolar patients. The decrease in Faecalibacterium resulted in a decline in anti-inflammatory reactions [134].
An increase in Clostridium spp. was reported in patients diagnosed with autism [135]. In a more detailed study on the complete microbiome of autistic patients, Finegold et al. [136] reported a significant increase in Bacteroidetes, Acintobacterium and Proteobacterium spp. but a decline of Firmicutes in autistic patients. High cell numbers of Clostridium defense, Clostridium hathewayi, and Clostridium orbiscindens were isolated from autistic patients. Faecalibacterium and Ruminococcus spp. were less abundant, which is an important observation given the anti-inflammatory properties of these species.
Obsessive-Compulsive Disorder (OCD), diagnosed in 2.3% of the population (mostly men), with a predominance in men [137], was originally classified as an anxiety disorder, similar to autism. Little is known about the gut microbial composition of individuals with OCD. Turna et al. [138] reported low numbers of Oscillospira, Odoribacter and Anaerostipes spp. in OCD patients. Odoribacter produces butyrate and low cell numbers may lead to an increase in inflammation, which may be the onset of OCD [139]. Experiments on mice showed a decrease in OCD when treated with Lactobacillus rhamnosus. In humans, similar findings were reported with the administration of Lactobacillus helveticus [140,141]. Findings such as these may provide valuable information in future research aimed at developing psychobiotics. For more information on probiotics and the effect on the nervous system, the reader is referred to the review published by Cryan et al. [142].
The production of trace amines β-phenylethylamine (PEA), p-tyramine (TYR) and tryptamine (TRP) by commensal gut microbiota is well documented [143,144,145], as many gut bacteria can produce aromatic L-amino acid decarboxylase (AADC; EC 4.1.1.28) [146]. Unlike DOPA, NE, EPI and 5-HT, , PEA, TYR and TRP are not stored and rapidly diffuse across membranes [147,148]. PEA diffuses across the blood-brain barrier and TYR passes through IECs [149]. Tyrosine is converted to l-3,4-dihydroxyphenylalanine (l-DOPA), the precursor of DOPA, NE and EPI [149]. A deficiency in L-tyrosine may thus lead to anxiety and low mood [149]. Treatments that increase monoamine neurotransmitter receptor activation leads to a decrease in PEA and TYR synthesis. Likewise, treatments that decrease receptor activation results in an increase in PEA and TYR synthesis. Reports on changes in AADC activity are almost exclusively based on L-DOPA as substrate. Binding of PEA, TYR, TRP and OCT to the trace amine-associated receptor TAAR1 in the brain regulates the release of neurotransmitters dopamine and serotonin [150]. Under- or overexpression of TAAR1 may lead to schizophrenia, depression and addiction [151].
5. Conclusions
Gut microbiota has a vast impact on the GBA and our overall mental health, as shown by drastic changes in the production of neurotransmitters when signals from the CNS reaches the ENS. These changes also affects digestion, intestinal permeability, gastric motility, and immune regulation. Many of the metabolic compounds produced by gut bacteria play an important role in anti-inflammatory responses and, because of this, also alleviate psychiatric disorders caused by inflammatory responses. Recent studies have shown that schizophrenia, bipolar disorder, autism and OCD may be prevented, and perhaps treated, by maintaining a healthy, balanced gut microbiome. Much more research is required to understand how gut microorganisms control cognitive behaviour, mood, and neuropsychiatric disorders. This entails the deciphering of the complex, everchanging network between cells and neurons. Further research is required to understand the interactions between gut microbiota. The identification of changes in the gut microbiome associated with psychological disorders may provide valuable information in the choice of treatment. Further in-depth studies need to be performed on the effect signals generated by the hypothalamus, and pituitary and adrenal glands have on the ENS. This is important, as the entero-endocrine signaling pathway and immunological reactions keeps the gut microbiome in a balanced state.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
None.
Conflicts of Interest
The author declare no conflict of interest.
References
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the interface of the microbiota-gut-brain axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.M.; Hofstra, R.; Burns, A.J. Building a brain in the gut: Development of the enteric nervous system. Clin. Genet. 2013, 83, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Corpening, J.C.; Cantrell, V.A.; Deal, K.K.; Southard-Smith, E.M. A Histone2BCerulean BAC transgene identifies differential expression of Phox2b in migrating enteric neural crest derivatives and enteric glia. Dev. Dyn. 2008, 237, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Micci, M-A.; Leser, J.; Shin, C.; Tang, S-C.; Fu, Y-Y.; et al. Adult enteric nervous system in health is maintained by a dynamic balance between neuronal apoptosis and neurogenesis. Proc. Natl Acad. Sci. USA 2017, 114, E3709–E3718. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Holm, L.M.; Karsenty, G.; MacPherson, A.J.; Olofsson, L.E.; et al. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Xiang, A.P.; Mao, F.F.; Zhang, L.; Di, C.-G.; Liu, X.-M.; et al. Nestin is required for the proper self-renewal of neural stem cells. Stem Cells 2010, 28, 2162–2171. [Google Scholar] [CrossRef]
- Furness, J.B. The enteric nervous system. 2006. Blackwell Publishing, Oxford, U.K.
- Bennett, M.R.; Burnstock, G.; Holman, M.E. Transmission from perivascular inhibitory nerves to the smooth muscle of the guinea-pig taenia coli. J. Physiol. 1966, 182, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Bulbring, E.; Tomita, T. Properties of the inhibitory potential of smooth muscle as observed in the response to field stimulation of the guinea-pig taenia coli. J. Physiol. 1967, 189, 299–315. [Google Scholar] [CrossRef]
- Furness, J.B. Types of neurons in the enteric nervous system. J. Auton. Nerv. Syst. 2001, 81, 87–96. [Google Scholar] [CrossRef]
- Berthoud, H.-R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Mahmoudian Dehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease—An emerging role for gut microbiome. Alzheimer’s Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Mukudai, S.; Sugiyama, Y.; Hisa, Y. Dorsal motor nucleus of the vagus. In Neuroanatomy and Neurophysiology of the Larynx; Springer: Tokyo, Japan, 2016; pp. 97–102. [Google Scholar]
- Brookes, S.J.; Song, Z.M.; Ramsay, G.A.; Costa, M. Long aboral projections of Dogiel type II, AH neurons within the myenteric plexus of the guinea pig small intestine. J. Neurosci. 1995, 15, 4013–4022. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Kunze, W.A.; Bertrand, P.P.; Clerc, N.; Bornstein, J.C. Intrinsic primary afferent neurons of the intestine. Prog. Neurobiol. 1998, 54, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.M.T. Gut Bacteria and neurotransmitters. Microorganisms 2022, 10, 1838. [Google Scholar] [CrossRef] [PubMed]
- Kugler, E.M.; Michel, K.; Zeller, F.; Demir, I.E.; Ceyhan, G.O.; Schemann, M. Mechanical stress activates neurites and somata of myenteric neurons. Front. Cell Neurosci. 2015, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Brehmer, A. Classification of human enteric neurons. Histochem. Cell Biol. 2021, 156(2), 95–108. [Google Scholar] [CrossRef]
- Spencer, N.J.; Smith, T.K. Mechanosensory S-neurons rather than AH-neurons appear to generate a rhythmic motor pattern in guinea-pig distal colon. J. Physiol. 2004, 558, 577–596. [Google Scholar] [CrossRef]
- Mazzuoli-Weber, G.; Schemann, M. Mechanosensitive enteric neurons in the guinea pig gastric corpus. Front Cell Neurosci. 2015, 9, 430. [Google Scholar] [CrossRef]
- Kunze, W.A.; Furness, J.B.; Bertrand, P.P.; Bornstein, J.C. Intracellular recording from myenteric neurons of the guinea-pig ileum that respond to stretch. J. Physiol. 1998, 506 (Pt 3) Pt 3, 827–842. [Google Scholar] [CrossRef]
- Bertrand, P.P.; Kunze, W.A.; Bornstein, J.C.; Furness, J.B.; Smith, M.L. Analysis of the responses of myenteric neurons in the small intestine to chemical stimulation of the mucosa. Am. J. Physiol. 1997, 273, G422–435. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, W.; Li, Y.; Cong, Y. Enteroendocrine cells: Sensing gut microbiota and regulating inflammatory bowel diseases. Inflamm. Bowel Dis. 2019, 26, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gribble, F.M.; Reimann, F. Enteroendocrine Cells: Chemosensors in the intestinal epithelium. Annu. Rev. Physiol. 2016, 78, 277–299. [Google Scholar] [CrossRef] [PubMed]
- De Silva, A.; Bloom, S.R. Gut Hormones and appetite control: A focus on PYY and GLP-1 as therapeutic targets in obesity. Gut Liver 2012, 6, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.; Parker, H.E.; Adriaenssens, A.E.; Hodgson, J.M.; Cork, S.C.; Trapp, S.; et al. Identification and characterization of GLP-1 receptor–expressing cells using a new transgenic mouse model. Diabetes 2014, 63, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Spencer, N.J.; Hu, H. Enteric nervous system: sensory transduction, neural circuits and gastrointestinal motility. Nat. Rev. Gastroenterol. Hepatol. 2020, 17(6), 338–351. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Van De Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; et al. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Planchais, J.; Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef]
- Dicks, L.M.T. How does quorum sensing of intestinal bacteria affect our health and mental status? Microorganisms 2022, 10, 1969. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Thuneberg, L.; Klűppel, M.; Malysz, J.; Mikkelsen, H.B.; Bernstein, A. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 1995, 373, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Gobbo, S.; Caputi, V.; Spagnol, L.; Schirato, G.; Pasqualin, M.; et al. Toll like receptor-2 regulates production of glial-derived neurotrophic factors in murine intestinal smooth muscle cells. Mol. Cell. Neurosci. 2015, 68, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.M.; Li, A.Y.; Nair, D.G.; Blennerhassett, M.G. Glial cell line-derived neurotrophic factor is a key neurotrophin in the postnatal enteric nervous system. Neurogastroenterol. Motil. 2010, 23, e44–e56. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Gershon, M.D. Neurogastroenterology: The dynamic cycle of life in the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 453–454. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, F.A.; Keenan, C.M.; Wallace, L.E.; Woods, C.; Cavin, J.-B.; Flockton, A.R. Intestinal microbiota shapes gut physiology and regulates enteric neurons and glia. Microbiome 2021, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Kruse, A.C.; Kobilka, B.K.; Gautam, D.; Sexton, P.; Christopoulos, A.; Wess, J. Muscarinic acetylcholine receptors: Novel opportunities for drug development. Nat. Rev. Drug Discov. 2014, 13, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Jie, W.U. Understanding of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 6, 653–655. [Google Scholar]
- Wolosker, H.; Dumin, E.; Balan, L.; Foltyn, V.N. d-Amino acids in the brain: D-serine in neurotransmission and neurodegeneration. FEBS J. 2008, 275, 3514–3526. [Google Scholar] [CrossRef] [PubMed]
- Ohgi, Y.; Futamura, T.; Kikuchi, T.; Hashimoto, K. Effects of antidepressants on alternations in serum cytokines and depressive-like behavior in mice after lipopolysaccharide administration. Pharmacol. Biochem. Behav. 2012, 103, 853–859. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, M.; Liu, S.; Chen, B.; Pan, C.; Yang, M.; et al. Suppressive immunoregulatory effects of three antidepressants via inhibiton of the nuclear factor-ƙB activation assessed using primary macrophages of carp (Cyprinus carpio). Toxicol. Appl. Pharmacol. 2017, 322, 1–8. [Google Scholar] [CrossRef]
- Singh, V.; Roth, S.; Llovera, G.; Sadler, R.; Garzetti, D.; Stecher, B.; et al. Microbiota dysbiosis controls the neuroinflammatory response after stroke. J. Neurosci. 2016, 36, 7428–7440. [Google Scholar] [CrossRef]
- Stanley, D.; Mason, L.J.; E Mackin, K.; Srikhanta, Y.N.; Lyras, D.; Prakash, M.D.; et al. Translocation and dissemination of commensal bacteria in post-stroke infection. Nat. Med. 2016, 22, 1277–1284. [Google Scholar] [CrossRef]
- Houlden, A.; Goldrick, M.; Brough, D.; Vizi, E.S.; Lénárt, N.; Martinecz, B.; et al. Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav. Immun. 2016, 57, 10–20. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T. Human microbiome project consortium: Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar]
- Nelson, W.C.; Weinstock, G.M.; Highlander, S.K.; Worley, K.C.; Creasy, H.H.; Wortman, J.R.; et al. The human microbiome jumpstart reference strains consortium. A catalog of reference genomes from the human microbiome. Science 2010, 328, 994–999. [Google Scholar] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Hugon, P.; Dufour, J.-C.; Colson, P.; Fournier, P.-E.; Sallah, K.; Raoult, D. A comprehensive repertoire of prokaryotic species identified in human beings. Lancet Infect. Dis. 2015, 15, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Bilen, M.; Dufour, J.-C.; Lagier, J.-C.; Cadoret, F.; Daoud, Z.; Dubourg, G. The contribution of culturomics to the repertoire of isolated human bacterial and archaeal species. Microbiome 2018, 6, 94. [Google Scholar] [CrossRef]
- Giovannini, M.; Lana, D.; Traini, C.; Vannucchi, M. The microbiota–gut–brain axis and Alzheimer disease. From dysbiosis to neurodegeneration: Focus on the central nervous system glial cells. J. Clin. Med. 2021, 10, 2358. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; et al. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Baj, A.; Moro, E.; Bistoletti, M.; Orlandi, V.; Crema, F.; Giaroni, C. Glutamatergic signaling along the microbiota-gut-brain axis. Int. J. Mol. Sci. 2019, 20, 1482. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J.; Chavan, S.S. Nerve stimulation for treatment of diseases and disorders. Patent application No 15/677,080, 25 Ene 2018.
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; et al. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.M.H.; Karlsson, H.; Crespo, J.G.; Johansson, M.E.V.; Eklund, L.; Sjövall, H.; et al. Altered O-glycosylation profile of MUC2 mucin occurs in active ulcerative colitis and is associated with increased inflammation. Inflamm. Bowel Dis. 2011, 17, 2299–2307. [Google Scholar] [CrossRef] [PubMed]
- Rausch, P.; Rehman, A.; Künzel, S.; Häsler, R.; Ott, S.J.; Schreiber, S.; et al. Colonic mucosa associated microbiota is influenced by an interaction of Crohn disease and FUT2 (Secretor) genotype. Proc. Natl. Acad. Sci. USA, 2011, 108, 19030–19035. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; McHardy, I.; Ruegger, P.; Goudarzi, M.; Kashyap, P.C.; Haritunians, T.; et al. Reprograming of gut microbiome energy metabolism by the FUT2 Crohn’s disease risk polymorphism. ISME J. 2014, 8, 2193–2206. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. The endotoxin hypothesis of neurodegeneration. J. Neuroinflammation 2019, 16, 180. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The central nervous system and the gut microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Friedland, R.P.; Chapman, M.R. The role of microbial amyloid in neurodegeneration. PLoS Pathog. 2017, 13, e1006654. [Google Scholar] [CrossRef]
- Willyard, C. How gut bacteria alter the brain. Nature 2021, 590, 22–25. [Google Scholar] [CrossRef]
- Cushing, K.; Alvarado, D.; A Ciorba, M. Butyrate and mucosal inflammation: New scientific evidence supports clinical observation. Clin. Transl. Gastroenterol. 2015, 6, e108. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Nohr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; et al. GPR41/FFAR3 and GPR43/FFAR2 as cosensors for shortchain fatty acids in enteroendocrine cells vs. FFAR3 in enteric neurons and FFAR2 in enteric leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef]
- Vyleta, N.P.; Smith, S.M. Spontaneous glutamate release is independent of calcium influx and tonically activated by the calcium-sensing receptor. J. Neurosci. 2011, 31, 4593–4606. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.; Tymianski, M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflugers Arch. 2010, 460, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Hashimoto, K.-I.; Sawada, Y.; Sokabe, M.; Kawasaki, H.; Martinac, B. Corynebacterium glutamicum mechanosensitive channels: Towards unpuzzling “glutamate efflux” for amino acid production. Biophys. Rev. 2018, 10, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Malathi, K.C.; Wachi, M.; Nagai, K. Isolation of the murI gene from Brevibacterium lactofermentum ATCC 13869 encoding D-glutamate racemase. FEMS Microbiol. Lett. 1999, 175, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-F.; Miller, C. Substrate selectivity in arginine-dependent acid resistance in enteric bacteria. Proc. Natl. Acad. Sci. USA, 2013, 110, 5893–5897. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Misgeld, U.; Bijak, M.; Jarolimek, W. A physiological role for GABAB receptors and the effects of baclofen in the mammalian central nervous system. Prog. Neurobiol. 1995, 46, 423–462. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; et al. . Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Janik, R.; Thomason, L.A.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic resonance spectroscopy reveals oral Lactobacillus promotion of increases in brain GABA, N-acetyl aspartate and glutamate. NeuroImage 2016, 125, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; DeLorey, T.M. Glycine receptor physiology and pharmacology. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; American Society for Neurochemistry: Philadelphia, PA, USA, 1999; pp. 335–346. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed]
- Takanaga, H.; Ohtsuki, S.; Hosoya, K.-I.; Terasaki, T. GAT2/BGT-1 as a system responsible for the transport of -aminobutyric acid at the mouse blood-brain barrier. J. Cereb. Blood Flow Metabol. 2001, 21, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; et al. The orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The gut microbiota mediates the anti-seizure effects of the ketogenic diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Strafella, A.P. Dopaminergic neurotransmission in the human brain: New lessons from perturbation and imaging. Neuroscientist 2012, 18, 149–168. [Google Scholar] [CrossRef]
- Olguín, H.J.; Guzmán, D.C.; García, E.H.; Mejía, G.B. The role of dopamine and its dysfunction as a consequence of oxidative stress. Oxidative Med. Cell. Longev. 2015, 2016, 1–13. [Google Scholar] [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; et al. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Liver Physiol. 2012, 303, G1288–G1295. [Google Scholar] [CrossRef]
- Rekdal, V.M.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and inhibition of an interspecies gut bacterial pathway for Levodopa metabolism. Science 2019, 364, 6323. [Google Scholar] [CrossRef] [PubMed]
- Borodovitsyna, O.; Flamini, M.; Chandler, D. Noradrenergic modulation of cognition in health and disease. Neural Plast. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.; Zeppenfeld, D.; McConnell, E.; Pena, S.; Nedergaard, M. Norepinephrine: A neuromodulator that boosts the function of multiple cell types to optimize CNS performance. Neurochem. Res. 2012, 37, 2496–2512. [Google Scholar] [CrossRef] [PubMed]
- Tsavkelova, E.A.; Botvinko, I.V.; Kudrin, V.S.; Oleskin, A.V. Detection of neurotransmitter amines in microorganisms with the use of high-performance liquid chromatography. Dokl. Biochem. 2000, 372, 115–117. [Google Scholar]
- O’Donnell, P.M.; Aviles, H.; Lyte, M.; Sonnenfeld, G. Enhancement of in vitro growth of pathogenic bacteria by norepinephrine: Importance of inoculum density and role of transferrin. Appl. Environ. Microbiol. 2006, 72, 5097–5099. [Google Scholar] [CrossRef] [PubMed]
- Gurda, G.T.; Guo, L.; Lee, S.-H.; Molkentin, J.D.; Williams, J.A. Cholecystokinin activates pancreatic calcineurin-NFAT signaling in vitro and in vivo. Mol. Biol. Cell 2008, 19, 198–206. [Google Scholar] [CrossRef]
- Stefano, G.B.; Pilonis, N.; Ptacek, R.; Raboch, J.; Vnukova, M.; Kream, R.M. Gut, microbiome, and brain regulatory axis: Relevance to neurodegenerative and psychiatric disorders. Cell. Mol. Neurobiol. 2018, 38, 1197–1206. [Google Scholar] [CrossRef]
- Kelly, J.R.; Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Hyland, N.P. Breaking down the barriers: The gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 392. [Google Scholar] [CrossRef]
- Kelly, J.R.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-gut-microbiota axis: Challenges for translation in psychiatry. Ann. Epidemiol. 2016, 26, 366–372. [Google Scholar] [CrossRef]
- Bellono, N.W.; Bayrer, J.R.; Leitch, D.B.; Castro, J.; Zhang, C.; O’Donnell, T.A.; et al. Enterochromaffin cells are gut chemosensors that couple to sensory neural pathways. Cell 2017, 170, 185–198.e16. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M. A role for the gut microbiota in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lou, J.; Shan, W.; Ding, J.; Jin, Z.; Hu, Y.; et al. Pathophysiologic role of neurotransmitters in digestive diseases. Front. Physiol. 2021, 12, 567–650. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.M.T.; Hurn, D.; Hermanus. Gut bacteria and neuropsychiatric disorders. Microorganisms 2021, 9, 2583. [CrossRef]
- Galland, L. The gut microbiome and the brain. J. Med. Food 2014, 17, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut microbes and the brain: Paradigm shift in neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. Available online: https://pubmed.ncbi.nlm.nih.gov/25830558/ (accessed on 2 November 2021). [PubMed]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.; Hu, X.; Wang, T.; Jin, F. Recognizing depression from the microbiota–gut–brain axis. Int. J. Mol. Sci. 2018, 19, 1592. [Google Scholar] [CrossRef]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; et al. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-F.; Shen, Y.-Q. Dysbiosis of gut microbiota and microbial metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Ong, I.M.; Gonzalez, J.G.; McIlwain, S.J.; Sawin, E.A.; Schoen, A.J.; Adluru, N.; et al. Gut microbiome populations are associated with structure-specific changes in white matter architecture. Transl. Psychiatry 2018, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabê de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Jewett, B.E.; Sharma, S. Physiology, GABA; StatPearls: Treasure Island, FL, USA, 2021; Available online: https://pubmed.ncbi.nlm.nih.gov/30020683/ (accessed on 14 October 2021).
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Roshchina, V.V. Evolutionary considerations of neurotransmitters in microbial, plant, and animal cells. In Microbial Endocrinology: Interkingdom Signaling in Infectious Disease and Health, 1st ed.; Lyte, M., Freestone, P.P.E., Eds.; Springer: New York, NY, USA, 2010; pp. 17–52. [Google Scholar]
- Sun, L.-J.; Li, J.-N.; Nie, Y.-Z. Gut hormones in microbiota-gut-brain cross-talk. Chin. Med. J. 2020, 133, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A. Gut feelings: The emerging biology of gut–brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef]
- Hyland, N.P.; Cryan, J.F. Microbe-host interactions: Influence of the gut microbiota on the enteric nervous system. Dev. Biol. 2016, 417, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, P.; Vollmer-Conna, U.; Hadzi-Pavlovic, D.; Grimm, M.C. A Review of inflammatory bowel disease: A model of microbial, immune and neuropsychological integration. Public Health Rev. 2021, 42, 1603990. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.; Park, S.-J. High-throughput 16S rRNA gene sequencing reveals alterations of mouse intestinal microbiota after radiotherapy. Anaerobe 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Saulnier, D.M.; Riehle, K.; Mistretta, T.-A.; Diaz, M.-A.; Mandal, D.; Raza, S.; et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterology 2011, 141, 1782–1791. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E. Gut microbiota in 2015: Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Wang, W.; Guo, R.; Liu, H. Faecalibacterium prausnitzii (ATCC 27766) has preventive and therapeutic effects on chronic unpredictable mild stress-induced depression-like and anxiety-like behavior in rats. Psychoneuroendocrinology 2019, 104, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Macchi, F.; Zecchillo, C.; Dell’Agli, M.; Colombo, E.; Calabrese, F.; et al. Modulation of the inflammatory response in rats chronically treated with the antidepressant agomelatine. Eur. Neuropsychopharmacol. 2013, 23, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Luki´c, I.; Getselter, D.; Ziv, O.; Oron, O.; Reuveni, E.; Koren, O.; et al. Antidepressants affect gut microbiota and Ruminococcus flavefaciens is able to abolish their effects on depressive-like behavior. Transl. Psychiatry 2019, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, J.; Li, Z.; Huang, Y.; Yuan, Y.; Wang, J.; et al. Analysis of gut microbiota diversity and auxiliary diagnosis as a biomarker in patients with schizophrenia: A cross-sectional study. Schizophr. Res. 2018, 197, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Zou, Y.; Dai, Y.; Luo, G.; Zhang, X.; Xiao, L. Characterization a novel butyric acid-producing bacterium Collinsella aerofaciens subsp. shenzhenensis subsp. nov. Microorganisms 2019, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, T.; Kawamura, Y.; Li, N.; Li, Z.Y.; Zhao, L.; Shu, S. Proposal of the genera Anaerococcus gen. nov., Peptoniphilus gen. nov. and Gallicola gen. nov. for members of the genus Peptostreptococcus. Int. J. Syst. Evol. Microbiol. 2001, 51, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Qing, Y.; Xu, L.; Cui, G.; Sun, L.; Hu, X.; Yang, X.; et al. Salivary microbiome profiling reveals a dysbiotic schizophrenia-associated microbiota. Npj Schizophr. 2021, 7, 51. [Google Scholar] [CrossRef]
- Zhou, S.; Cai, Y.; Wang, M.; Yang, W.D.; Duan, N. Oral microbial flora of patients with Sicca syndrome. Mol. Med. Rep. 2018, 18, 4895–4903. [Google Scholar] [CrossRef]
- Castro-Nallar, E.; Bendall, M.L.; Pérez-Losada, M.; Sabuncyan, S.; Severance, E.G.; Dickerson, F.B.; et al. Composition, taxonomy and functional diversity of the oropharynx microbiome in individuals with schizophrenia and controls. Peer J 2015, 3, e1140. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Severance, E.; Yolken, R. The microbiome, immunity, and schizophrenia and bipolar disorder. Brain Behav. Immun. 2017, 62, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Carlier, J.P.; Bedora-Faure, M.; K’ouas, G.; Alauzet, C.; Mory, F. Proposal to unify Clostridium orbiscindens Winter et al. 1991 and Eubacterium plautii (Séguin 1928) Hofstad and Aasjord 1982, with description of Flavonifractor plautii gen. nov., comb. nov. and reassignment of Bacteroides capillosus to Pseudoflavonifractor capiillosus gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 585–590. [Google Scholar] [PubMed]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Painold, A.; Mörkl, S.; Kashofer, K.; Halwachs, B.; Dalkner, N.; Bengesser, S.; et al. A step ahead: Exploring the gut microbiota in inpatients with bipolar disorder during a depressive episode. Bipolar Disord. 2019, 21, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.J.; Bassis, C.M.; Hein, R.; Assari, S.; Flowers, S.A.; Kelly, M.B.; et al. The gut microbiome composition associates with bipolar disorder and illness severity. J. Psychiatr. Res. 2017, 87, 23–29. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, M.; Yang, X.; Hong, N.; Yu, C. Faecalibacterium prausnitzii upregulates regulatory T cells and anti-inflammatory cytokines in treating TNBS-induced colitis. J. Crohns Colitis 2013, 7, e558–e568. [Google Scholar] [CrossRef]
- Mccartney, A.L.; Parracho, H.M.R.T.; Bingham, M.O.; Gibson, G.R. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J. Med. Microbiol. 2005, 54, 987–991. [Google Scholar] [CrossRef]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef]
- Ruscio, A.M.; Stein, D.J.; Chiu, W.T.; Kessler, R.C. The epidemiology of obsessive-compulsive disorder in the National Comorbidity Survey Replication. Mol. Psychiatry 2008, 15, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Turna, J.; Kaplan, K.G.; Anglin, R.; Van Ameringen, M. “What’s bugging the gut in OCD” A review of the gut microbiome in obsessive-compulsive disorder. Depress. Anxiety 2016, 33, 171–178. [Google Scholar] [CrossRef]
- Göker, M.; Gronow, S.; Zeytun, A.; Nolan, M.; Lucas, S.; Lapidus, A.; et al. Complete genome sequence of Odoribacter splanchnicus type strain (1651/6 T). Stand. Genom. Sci. 2011, 4, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, M.; Violle, N.; Bisson, J.-F.; Desor, D.; Javelot, H.; Rougeot, C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2011, 2, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Kantak, P.A.; Bobrow, D.N.; Nyby, J.G. Obsessive-compulsive-like behaviors in house mice are attenuated by a probiotic (Lactobacillus rhamnosus GG). Behav. Pharmacol. 2014, 25, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; de las Rivas, B.; Muñoz, R. First genetic characterization of a bacterial beta-phenylethylamine biosynthetic enzyme in Enterococcus faecium RM58. FEMS Microbiol. 2006, 258, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Irsfeld, M.; Spadafore, M.; Prüß, B.M. β-phenylethylamine, a small molecule with a large impact. Webmedcentral 2013, 4, 4409. [Google Scholar] [PubMed]
- Yang, Y.X.; Mu, C.L.; Luo, Z.; Zhu, W.Y. Bromochloromethane, a methane analogue, affects the microbiota and metabolic profiles of the rat gastrointestinal tract. Appl. Environ. Microbiol. 2015, 82, 778–787. [Google Scholar] [CrossRef]
- Dyck, L.E.; Yang, C.R.; Boulton, A.A. The biosynthesis of p-tyramine, m-tyramine, and beta-phenylethylamine by rat striatal slices. J. Neurosci. Res. 1983, 10, 211–220. [Google Scholar] [CrossRef]
- Berry, M.D.; Shitut, M.R.; Almousa, A.; Alcorn, J.; Tomberli, B. Membrane permeability of trace amines: Evidence for a regulated, activity-dependent, nonexocytotic, synaptic release. Synapse 2013, 67, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Mosnaim, A.D.; Callaghan, O.H.; Hudzik, T.; Wolf, M.E. Rat brain-uptake index for phenylethylamine and various monomethylated derivatives. Neurochem. Res. 2013, 38, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.B.; Raiteri, M.; Bertollini, A.; del Carmine, R. Interaction of betaphenethylamine with dopamine and noradrenaline in the central nervous system of the rat. J. Pharm. Pharmacol. 1976, 28, 456–457. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace amines and their receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef]
- Del Rio, J.A.; Soriano, E.; Ferrer, I. Development of GABA-immunoreactivity in the neocortex of the mouse. J. Comp. Neurol. 1992, 326, 501–526. [Google Scholar] [CrossRef]
Figure 1.
Major extrinsic neural pathways between the ENS, spinal cord and the brain. Extrinsic motor pathways are shown with blue lines. These pathways include the parasympathetic (vagal motor), and sympathetic nervous systems stemming from the thoracolumbar spinal cord and linking with the celiac, superior mesenteric ganglia, and inferior mesenteric ganglia (blue lines). In the oesophagus, and stomach the major extrinsic sensory nerves arise from the vagus nerve. In the colon, the major extrinsic sensory nerves stem from spinal afferent nerves, with cell bodies in dorsal root ganglia (DRG).
Figure 1.
Major extrinsic neural pathways between the ENS, spinal cord and the brain. Extrinsic motor pathways are shown with blue lines. These pathways include the parasympathetic (vagal motor), and sympathetic nervous systems stemming from the thoracolumbar spinal cord and linking with the celiac, superior mesenteric ganglia, and inferior mesenteric ganglia (blue lines). In the oesophagus, and stomach the major extrinsic sensory nerves arise from the vagus nerve. In the colon, the major extrinsic sensory nerves stem from spinal afferent nerves, with cell bodies in dorsal root ganglia (DRG).
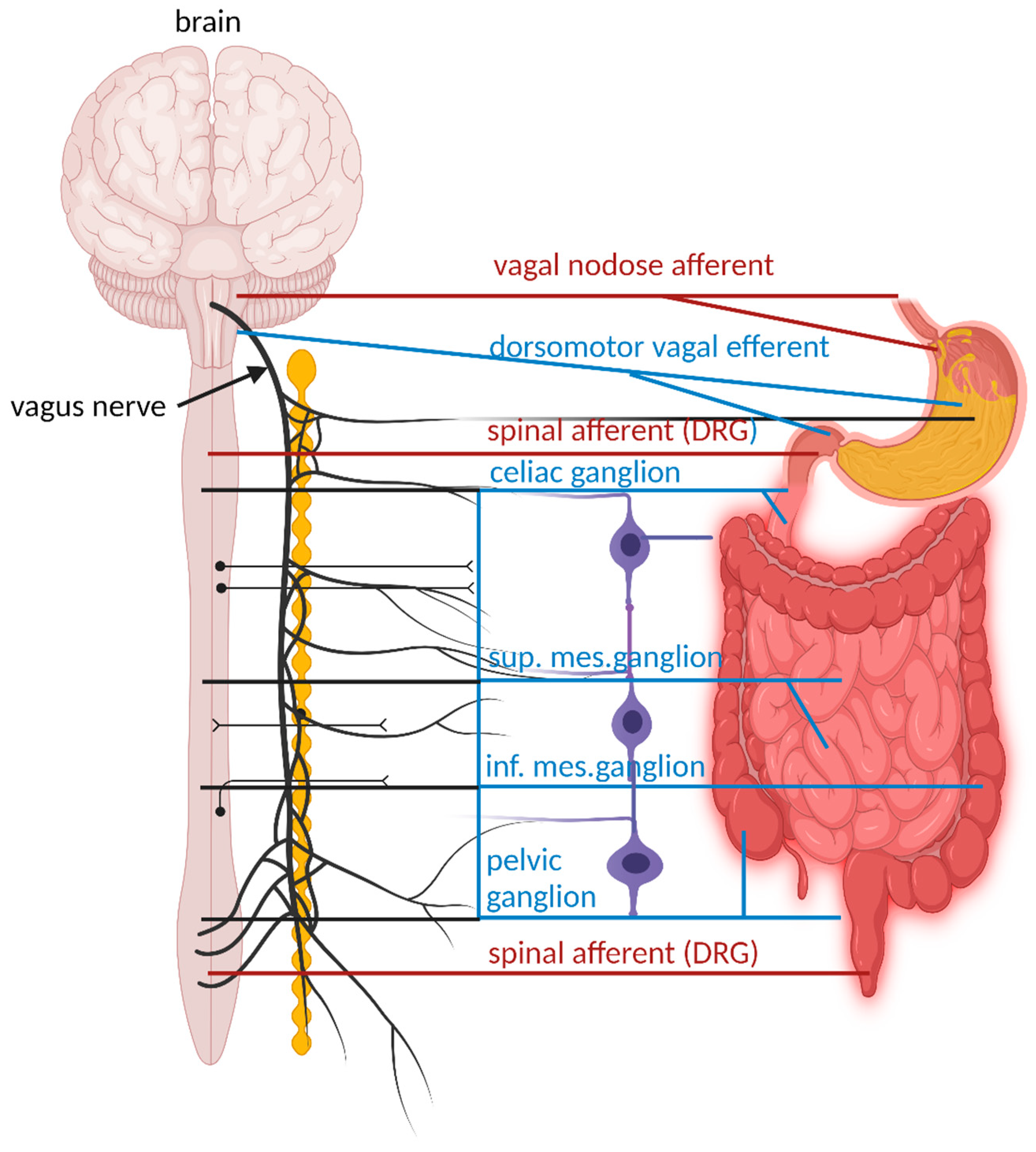
Figure 2.
Intrinsic sensory neurons and extrinsic sensory nerve endings in the ENS. Dogiel type I neurons, located in the myenteric plexus, are interneurons and are length-sensitive but tension insensitive. Mechanosensory neurons are rapidly adapting and excitatory. Dogiel type II neurons, also in the myenteric plexus, are chemosensory and mechanosensitive and react to rapid and slow synaptic inputs from other enteric neurons. Intestinofugal neurons are “second-order” neurons but react to direct mechanical stimuli. Extrinsic vagal afferent nerve endings supply signals to mostly the upper gut and act as slowly adapting tension receptors. Spinal afferent neurons send signals to the lower gut (distal colon) and are activated by changes in muscle tension.
Figure 2.
Intrinsic sensory neurons and extrinsic sensory nerve endings in the ENS. Dogiel type I neurons, located in the myenteric plexus, are interneurons and are length-sensitive but tension insensitive. Mechanosensory neurons are rapidly adapting and excitatory. Dogiel type II neurons, also in the myenteric plexus, are chemosensory and mechanosensitive and react to rapid and slow synaptic inputs from other enteric neurons. Intestinofugal neurons are “second-order” neurons but react to direct mechanical stimuli. Extrinsic vagal afferent nerve endings supply signals to mostly the upper gut and act as slowly adapting tension receptors. Spinal afferent neurons send signals to the lower gut (distal colon) and are activated by changes in muscle tension.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



