
Submitted:
22 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Campylobacter jejuni is a zoonotic bacterium, causing intestinal infection in humans. One characteristic feature of the pathogenicity process is the ability of this bacterium invading the epithelial cells. Several factors have been identified to contribute to this process and some recent analyses revealed changes in the bacterium cell on molecular level. However, our knowledge is still limited about responses of the host that accompany the bacterial internalisation and survival process. This study focused on molecular events on transcriptomic level detected between the 1st and the 3rd hour of internalisation in INT407 epithelial cells. From the 41.769 human genes tested, altogether 19,060 genes and 22,734 pseudogenes and introns were shown by whole transcriptome analysis to be affected in different extent. Most affected functions regulate transcription (28%), signal transduction (21%), apoptosis (15%), immune responses (9%), transmembrane transport (6%), cell-cell signaling (3%), cell-cell adhesions (3%) and carbohydrate metabolism (3%) and to a lesser extent other functions. Our results provide insight into a scenario where, the invaded cell focuses on survival, but, at the same time, flashing the possibility of pushing the process toward the death of the invaded and metabolically disrupted cell.
Keywords:
Campylobacter jejuni
; INT407 cell line infection
; WTA
1. Introduction
Campylobacter jejuni is one of the most important bacterial agents causing gastrointestinal infections with varying symptoms ranging from mild to bloody diarrhea. The infection is common also in developed and under developed countries. Outbreaks were reported in Eurasia, America, Africa and even in Oceania, which means that campylobacteriosis is a global problem. Epidemics caused by C. jejuni are not negligible, and these are registered more frequently in developing countries [1-5].
The typical source of infection is chicken meat, but besides, infection can also be transmitted to humans through the consumption of contaminated water, -milk, and -meats, through animal contact and in general by inadequate hygienic-sanitary conditions [6,7]. The prevalence of C. jejuni in India and Africa correlates with contaminated water that they drink and give to their animals [8,9].
Living near to and together with animals is also a risk factor [10,11], similarly to malnutrition, especially in case of children [12,13].
Although there is a big difference among the isolates in their pathogenic potential, virulence and genome organisation, the general pathogenicity process of C. jejuni can be described with the following stages. C. jejuni enters the host intestine through the gastric acid barrier of the stomach, and colonises the mucosa covering the distal ileum and colon. During the passage into the small intestine and the migration of the bacteria towards the mucus-filled crypts, C. jejuni reacts, presumably as an adaptive response, to the microenvironment of the current intestinal section, where it synthesizes a new set of proteins facilitating their subsequent interaction with the host’s target cells. It can penetrate enterocytes by paracellular or transcellular route [14,15]. The flagellae and the screw-shape of the bacterial cells play an important role in reaching the epithelial cells through the mucus layer [16]. A group of adhesion proteins support the binding of the bacterial cell to extracellular matrix proteins (ECMPs), including fibronectin and laminin,. This process is facilitated by several factors such as CadF, Peb1, Peb2, Peb3, Peb4, CapA, CjaA, FlpA, FbpA, JlpA, DocA [17].
As the first step of the invasion the pathogen interacts with the host through biochemical signals, such as Campylobacter invasion antigens (Cia) [18,19]. As a result, a signaling cascade triggers the rearrangement of the host's cytoskeleton, leading to the internalisation of the bacterium in a vacuole [20]. At this point a two-sided game begins in which the C. jejuni cell, in order to assure its survival, tries to maintain the vacuolised form by avoiding its fusion with lysosomes. On the other side, the eukaryotic cell attempts to eliminate the invader. Some recent studies have outlined molecular changes, such as the expression of capsule, lipooligosaccharide, and different membrane transport systems, and also the activation of stress related genes accompanying the invasion process and assuring the survival of the bacterial cell [17,21,22]. Other studies primarly focused on immunologic aspects such as the appearance of interleukines in the supernatant of invaded cells, showing increased expression of IL-8 due to infection [23] in a strain dependent manner, as well as minor changes in the expression of IL-1 and IL-4 in a time dependent manner with a slight increase at the 1st and 4th hour and a subsequent decrease by the 24th hour following infection [24]. Other studies demonstrated the massive expression of TNF-alpha suggesting that the immun response was shifted towards Th-1 type [25].
Transcriptomic methods have also been used to estimate activation of different interleukines, such as IL-4, IL-8, IL-10, also in the case of Guillan-Barre syndrome (GBS), one sequelae of C. jejuni infection [26-29]. However, during the invasion process, genes other than those with immunological function were not typically examined, so not much is known about the other side of the infectious process, which is how internalisation of C. jejuni affects the eukaryotic transcriptome of the invaded cell. This is an important issue, since the outcome of bacterial infections depends not only on the infectious agent, but also on the host itself, influencing in several cases the development of certain late onset complications following C. jejuni infection [30-32].
Aim of this study was to get a view about those molecular changes that occur between the 1st and 3rd hour following internalisation and to determine whether these changes help maintain or eliminate the internalised state of the C. jejuni cells. For this purpose a Whole Transcriptome Analysis (WTA) was performed by using the INT407 cell line and the recently isolated and partially characterised highly invasive C. jejuni strain CjTD-119 [17].
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
For this study the Campylobacter jejuni CjTD-199 strain was used. This strain was partially characterised in a recent study [17], is a representative of a 190 piece strain collection and among the seven strains isolated from a patient with bloody diarrhoea showing strong adhesion and invasion potentials. The strain was routinely grown on Charcoal Cefoperazone Deoxycholate Agar (CCDA) at 42°C under microaerophilic conditions.
2.2. Preparing the INT 407 Cell Line for the Invasion Assay
The analyses were performed on semi-confluent monolayer of INT407 human embryonic intestine cell line grown on 24-well culture plates. For monolayers, 3x105 cells were cultured in RPMI 1640 medium (BioWhittaker, Lonza, Switzerland) supplemented with 10% heat-inactivated (30 min for 56 °C) calf bovine serum (Sigma-Aldrich, USA), 10,000 U/ml of penicillin, 10 μg/ml of streptomycin and 0.5 mg/ml of neomycin, and incubated overnight at 37 °C in a humidified, 5% CO2 incubator.
2.3. Infection of INT 407 with the C. jejuni Strain CjTD-199
Plate culture of strain CjTD-199 was suspended in RPMI 1640 and the optical density was set to OD600=0.1. One ml from the suspension was added to wells containing the semi-confluent layers of INT407 cell line. Altogether 3 plates (72 wells) were used for the experiments. After addition, the bacterial cells (MOI 2), were centrifuged onto the monolayer, and the plates were incubated at 37 °C in a humidified incubator with 5% CO2 for one and three hours. At these timepoints total RNA was isolated from the wells.
2.4. Isolation of RNA from the INT 407
RNA isolations from the INT407 cells were performed as described earlier [17] with some modifications. Briefly, before collecting the cells wells were washed once with PBS. After removing not invading C. jejuni cells, 1 ml RNAprotect Bacteria Reagent (Qiagen, Hamburg, Germany) was applied to stabilize the RNA in each well. Following 5 minutes of incubation at room temperature, cells were trypsinized (Life Technologies, Carlsbad, California, USA) and detached cells were collected with centrifugation (2000 × g for 5 min). Collected cells were homogenized in RNAzol (Molecular Research Center, Cinncinatti, Ohio, USA) in a 1.5 ml microcentrifuge tube. The tubes were dropped three times into liquid nitrogen (-196 °C) for more effective extraction of the RNA. The total RNA concentration and purity was measured by an ND–1000 Spectrophotometer (Nanodrop, Thermo Scientific, Carlsbad, California, USA).
These isolation steps were performed on the relevant plates at the 1st and 3rd hour after infection.
2.5. Whole Transcriptome Analysis
WTA was performed as described earlier[17]. Qualitative and quantitative measurements of the isolated RNA were carried out with Bioanalyzer (Agilent Technologies, Santa Clara, California, USA) and Qubit (Life Technologies). High quality RNA samples were pooled for analysis. We used the SOLiD total RNA-Seq Kit (Life Technologies, USA) and RiboZero Prokaryotic rRNA Removal Kit (Epicentre, USA). Remaining RNAs were fragmented by using RNase III, and the 50-200 nucleotide long fragments were ligated to adaptors. Reverse transcription of these constructs was performed with ArrayScript RT kit (Thermo Fisher Scientific, USA). The Qiagen MinElute PCR Purification Kit (Qiagen, Germany) was used to purify the cDNA library and the SOLiD Library TaqMan Quantitation Kit (Life Technologies, USA) was used to determine library concentration. Emulsion PCR (ePCR) was used to amplify quality controlled libraries on SOLiD P1 DNA Beads, followed by enrichment for template-positive beads by hybridization with magnetic enrichment beads. Template-enriched beads were extended at the 3′ end in the presence of terminal transferase and a 3′ bead linker. Beads with the clonally amplified DNA were deposited onto sequencing slides and sequenced on a SOLiD5500XL Instrument (Thermo Fisher, Carlsbad, California, USA) using the 50-base sequencing chemistry.
2.6. Bioinformatics - Heatmap
The CLC Genomics Workbench (Qiagen) was used for data analyses. Low quality and short sequences were removed in a trimming step, and only sequences 45–50 nucleotides long were used for analysis. Sequences were mapped onto the human eukaryotic genome sequence. Results were manually cured to remove false positive hits, which showed highly skewed mapping of reads. Only genes, detected with at least a 2.0-fold increase or decrease in transcription level after normalization were considered for further analysis. For heatmap analysis, normalized expression values of genes for the two samples were plotted on a heat map using the pheatmap v1.0.12 package in the R language (v4.2.2). Genes with FDR p-value corrected Kal's Z-test p-values <0.001 were taken into acccount. The fifty most upregulated and fifty most downregulated genes, based on fold change of normalized values, were selected for visualization.
3. Results
3.1. Comparison of Expression Changes of Genes Associated with Immune Functions
The complete list of the 41.796 genes with their expression levels are listed in the Supplementary Table 1. Altogether 19.060 genes and 22.734 pseudogenes and introns were identified by whole transcriptome analysis by mapping them on to the human genom. From among the active genes 2.764 were upregulated (fold change ˃2.0), while 2.220 were downregulated (fold change ˃2.0) in the two hours’ time range between the 1st and 3rd hour after C. jejuni cells became internalized into the INT407 cell. Grouping of the genes with increased or decreased expression levels, based on their formerly confirmed or only hypothesized major physiological functions are summarized in Figure 1.
Genes associated with immunological functions showing an altered expression are presented in Table 1.
Data show that the infected human cells reacted to the C. jejuni infection with an upregulated immune response. Not only the genes responsible for innate, but also some others taking part in adaptive immune processes were upregulated. The gene showing the highest expression change was ulbP3 (ULBP3, Fc.: 10.10x), whose protein product ULBP3 is related to MHC class I proteins and is an important regulatory protein of the natural immune system. Another gene with strongly upregulated expression was cr1 (CR1; Fc.: 7.581x), coding for a transmembrane glycoprotein playing part in various processes in the body, including adhesion and phagocytosis of immune cells [33] and also associated with several diseases [34]. A gene responsible for macrophage activation, ifi44L, showed a characteristic increase (IFI44L, Fc.: 3.685x) which was among the highest in the group of genes affecting immune processes.
A slightly lower expression change (CD74, Fc.: 2.527x) was detected in the case of cd74, that performs many tasks in the eukaryotic cell. Its functions include (i) negative regulation of mature B cell apoptosis, (ii) positive regulation of neutrophil chemotaxis, (iii) positive regulation of T-helper type 2 immune response, and negative regulation of apoptosis [35]. Additionally, it is a positive regulator of B-cell proliferation, and the extracellular signal-regulated kinase 1/2 (ERK1 and ERK2) and mitogen-activated protein kinases (MAPKs), both of them taking part in a diverse array of cellular processes [36].
Expression levels of four genes cd36, procr, cd209, il10ra (CD36, Fc.: 2.73, PROC, Fc.: 2.527x; CD209, Fc.:2.52; IL10RA, Fc.:2.52), all responsible for antigen processing and presentation were also markedly increased. CD36 is an exogenous peptide and has a role in antigen presentation via MHC class I, and positive regulation of the MAPKKK cascade. The function of CD209 (CD209, Fc.: 2.53x) is: regulation of T-cell proliferation, antigen processing and presentation, and innate immune response [37]. IL10RA is the cell surface receptor for the cytokine IL-10 [38].
IL23R (IL23 receptor, Fc.: 6.31x) is the cell surface receptor for the cytokine, that regulates the activity of immune cells. When IL-23 binds to its receptor, it triggers a series of chemical signals inside the cell. These signals promote inflammation and help coordinate the immune system's response to foreign invaders such as bacteria and viruses.
3.2. Affected Genes Related to Metabolic Functions
Among the metabolism-related genes, we found four whose expression was changed more than 2 fold over the study period. ABCD2 (ABCD2, Fc.: 11.37x) has a key role in the fatty acid homeostasis of peroxisomes, the oxidative organelles of eukaryotic cells [49]. TRPM6 (TRPM6, Fc.: 11.37x) is a selective magnesium channel [50], while ENPP3 (ENPP3, Fc.: 8.84x) is an ectoenzyme, that hydrolyses extracellular nucleotides, such as ATP, preventing ATP induced apoptosis and deregulates cytokine production.
Several other metabolism-related genes were found to be up and downregulated (Table 2.) with changes less than 2.0 in expression level.
3.3. Comparison of Expression Changes of Genes Associated with Stress Responses
Concerning stress related genes, a drastic increase in the expression of vnn1 (VNN1, Fc.: 379.08x) was detected. This gene’s product contributes to tolerance to tissue damage and modulates the ability of the affected cell to cope with oxidative stresses. Additionally, another stress gene lpo, first described in connection to lipid peroxidation, and strongly related to oxidative stress conditions showed increased expression at the 3rd hour following infection (LPO, Fc.: 5.05x). Furthermore, a slight increase in the expression of 3 additional genes: chac1, adcyap1r1 and rgcc was detected (CHAC1, Fc.: 8.845x; ADCYAP1R1, Fc.: 3.791x; RGCC, Fc.: 3.791x). CHAC1, is thought to regulate the glutathion level and also oxidative balance in the cell. Lipid peroxidation, controlled by LPO is a fundamental constituent of oxidative stress and free radical production [54], while RGCC has an effect on stress fiber formation [55]. Expression of HSPA12B (HSPA12B, Fc.: -2.374x) a heatshock protein [56], and SCAMP5 (SCAMP5, Fc.: -3.165x) an inhibitor of endocytosis were downregulated during the investigated time period [57].

Table 3.
The expression changes of genes associated with stress conditions.
| Gene | Function | 1vs3 | Ref. |
|---|---|---|---|
| VNN1 | response to oxidative stress, pantothenate metabolic process | 379.08 | [58] Zhang et al. 2017 |
| CHAC1 | apoptosis in response to endoplasmic reticulum stress | 8.84537 | [59] Mungrue et al. 2009 |
| LPO | response to oxidative stress | 5.05 | [60] Kovács et al. 1996 |
| ADCYAP1R1 | multicellular organismal response to stress | 3.791 | [61] Ressler et al. 2011 |
| RGCC | positive regulation of stress fiber formation, cell cycle regulation | 3.791 | [62] Wang et al. 2011 |
| HSPA12B | response to stress | -2.374122 | [63] Zouein et al. 2013 |
| SCAMP5 | response to endoplasmic reticulum stress | -3.165496 | [57] Noh et al. 2009 |
3.4. Affected Genes Related to Apoptosis
At least 25 genes formerly hypothetised to have roles in the induction of apoptosis were also affected with more than 2 fold expression change. Among these, 22 were found to influence the apoptopic process positively.
Concerning the positive regulators of apoptosis, the expression of dcc (DCC, Fc.: 5.68x), dlc1 (DLC1, Fc.: 5.05x), cd27 (CD27, Fc.: 3.79x) has been documented in several different cells, while the expression of other genes, such as fndC1 (FNDC1, Fc.: 7.85), until now has only been detected in specific cell types, such as cardiac cells. FndC1 plays an important role in angiogenesis and is essential to hypoxia-triggered cardiomyocyte apoptosis [64]. Recent studies, however, demonstrated that aberrant expression of fndC1 is associated with tumorigenesis, for example, in the case of gastric cancer [65].
Other affected genes are inhibitors of apoptosis. It was reported, that serpinB9 (SERPINB9, Fc.: 11.37x) protects cells from the immune-killing effect of granzyme B (GRB) released by lymphocytes [66].
Table 4.
Genes that play a role in apoptosis.
| Genes. | 1vs3 | Funciton | References |
|---|---|---|---|
| SERPINB9 | 11.3726 | negative regulation of apoptosis by inhibiting granzyme B | [67] Bird et al. 2014 [68] Kaiserman et al. 2010 |
| ACVR1C | 10.1089 | regulation of apoptosis | [69] Asnaghi et al. 2019 |
| CHAC1 | 8.8454 | apoptosis in response to endoplasmic reticulum stress | [70] Zhou et al. 2023 |
| FNDC1 | 7.8517 | positive regulation of cardiac muscle cell apoptosis | [71] Das et al. 2017 [72] Yunwen et al. 2021 |
| G0S2 | 6.9499 | positive regulation of apoptosis | [73] Heckmann et al. 2013 |
| NFATC4 | 6.31812 | positive regulation of apoptosis | [74] Mognol et al. 2016 |
| HIC1 | 5.8969 | signal transduction resulting in induction of apoptosis | [75] Wang et al. 2017 |
| DCC | 5.6863 | regulation of apoptosis | [76] Mehlen et al. 1998 |
| DLC1 | 5.0545 | induction of apoptosis | [77] Zhang et Li 2020 [78] Ullmannova et al. 2007 |
| CD27 | 3.79087 | induction of apoptosis | [79] Prasad et al. 1997 |
| CASP3 | 2.7851 | nuclear fragmentation during apoptosis | [80] Porter et Jänicke 1999 |
3.5. Genes Involved in the Potential Development of Chronic Conditions
Expression change of several genes previously hypothesised or verified to have potential roles in the evolvement of post-infectious pathological conditions was revealed in our study. Four genes, formerly thought to influence the severity of Guillan-Barre syndrome (GBS) [81-83] were affected differently but in a moderate way (PTGS2, Fc.: 1.188x; ANXA3, Fc.: 1.31x; CREB1, Fc.: 1.73x) by the 3rd hour of infection (Table 5). Characteristic upregulation of the genes relb, birc3 and nfkbia (RELB, Fc.: 1.625x; BIRC3, Fc.:1.958x; NFKBIA, Fc.:-2.55x), associated with the progression of inflammatory reactions, was detected. However, a higher expression rate was seen in the case of ace (ACE, Fc.: 3.79x), a gene hypothesised to induce autoreactive TH1 and TH17 cells and suppress regulatory T cells, and thus beeing involved in autoimmune responses of the body[84].
Another characteristic group of genes whose expression was found to be altered in the investigated time range and which have previously been shown to be associated with pathological condition, are genes linked to tumorgenesis. The muc4 and muc6 (MUC4, Fc.: 3.15x; MUC6, Fc.: 3.36x) showed clear increases in their expressions, with values of 3.15x and 3.36x respectively. A more drastic expression was detected in the case of three genes serpinB9, fndC1, tacrD2 (SERPINB9, Fc.: 11.37x; FNDC1, Fc.: 7.58x; TACRD2, Fc.: 8.84x). SerpinB9 has been demonstrated to be significantly associated with the development of precancerous lesions [66]. In contrast, fndC1 promotes the invasiveness of gastric cancers and correlates with the appearance of peritoneal metastasis [65].
3.6. Markedly Affected Genes with Unknown Functions
Table 6.
| Gene name | Fold change |
|---|---|
| ST20-MTHFS | 18.95437 |
| PRR4 (NW_003571047 44554..48182) | 11.3726 |
| C12orf55 | 10.109 |
| NDUFA3(NW_003571054 77272..81394) | 10.109 |
| COL11A2 (NT_167245 4411775..4441552) | 7.581748 |
| ADCK5 (NT_037704 165142..185869) | 3.79087 |
3.7. Heat Map Analysis
Figure 1.
Visualisation of the relative expression ratios of the fifty most upregulated and fifty most downregulated genes based on the normalized fold change expression values.
Figure 1.
Visualisation of the relative expression ratios of the fifty most upregulated and fifty most downregulated genes based on the normalized fold change expression values.

4. Discussion
Up-, and downregulation of the 2.764 and 2.200 genes, respectively, from the 41.769 ORFs reflects intensive transcriptomic changes in the INT407 intestinal human cell line upon the invasion of C. jejuni. Our preliminary findings have revealed that by the first hour after infection, 85% of the C. jejuni cells became stably internalised. From that point the possible routes for elimination of the invader can be outlined.
Activation of immune functions was represented by the increased expression of several relevant gene products. Overexpression of ULBP3 (ULBP3, Fc.: 10.109x) (Table 1), encoding for an MHC I related cell surface protein, clearly indicates that the invaded cell prepares itself for antigene presentation, which is a crucial step in the immune recognition of cells infected with viruses or invasive bacteria [40]. Due to antigen presentation, cytotoxic T cells are able to recognise infected eukaryotic cells [100]. However, a proper processing of antigens is a prerequisite for this, in which the recently discussed [45-47] role of cd36 (CD36, Fc.:2,74x), procr (PROCR, Fc.:2.53x), cd209 (CD209, Fc.:2.523x) and il10ra (IL10RA, Fc.: 2.53x) is also supported by our results. Antigene presentation on the surface of the affected eukaryotic cell can not be effective without attracting immune cells, which makes the increased expression levels of cr1 (CR1, Fc.:7,59x) and ifi44L (IFI44L, Fc.: 3,317x) [40,43] not surprising, since these proteins make the infected cell more accessible to macrophages [33]. Cytokines and interleukins play an important role in the attraction of immune cells and coordination of immunological processes for which the expression of the proper receptors is inevitable. Increased expression rates of the IL10 (IL10RA, Fc.:2.53x) and IL23 (IL23R, Fc.:6.31x) (Table 1) receptors strongly suggests that these two interleukins contribute to the immune reaction evoked by the vacuolised pathogen. IL10RA activates the tyrosine phosphorylation of JAK1 and TYK2 kinases, two enzymes contributing to the alteration of the IFN-alpha/beta and gamma signaling pathways, thereby affecting the production of certain groups of cytokines [101]. The mild expression change of IL4 (IL4I1, Fc.: 1.25x) was in accordance with the findings of other authors [25], while the decreased expression of IL8 (IL8, Fc.:-2.74x) by the 3rd hour of infection is seemingly in contrast with an earlier finding [23]. A plausible explanation would be that our data were generated from the 3rd hour, while in the study of Hickey, samples were taken 24 hours following infection. Another possible explanation may be the strain dependent manner of the IL8 expression rate [23]. In a recent study the steep increase of the proinflammatoric citokines IL6, IL8 and IFN-γ and that of a regulator citokine IL10 was detected from the 5th hour following infection [102]. Our results are in partial accordance with these findings, showing moderate increases in the case of IL6 (IL6R, Fc.:1.7x), IL10 (IL10RA, Fc.:2.52x) and IFN-γ (IFNGR2, Fc.:1.4x), while the level of IL8 transcript dropped (IL8, Fc.:-2.74x) in the investigated time range (STable 1.)
Based on our data, campylobacter infection outlines two possibilities for the invaded eukaryotic cell. One is survival by activating the immune system, while the other option is to sacrifice the eukaryotic cell by apoptosis in order to eliminate the pathogen from the body [103]. In certain situations, for an infected cell it is much more rewarding to drive itself toward apoptosis, a notion supported by the upregulation of 12 genes listed in Table 4. In contrast, expression of serpinB9 and cd74 acts against apoptosis (Table 1), of which the latter one, as a positive regulator of the type-2 immune response [44], directs the process toward activation of adaptive immune processes.
Be it the activation of the immune system or apoptosis, both potential routes drastically reprogram metabolism and with that, require either the activation or the depression of transcription factors (STable 1). The coordinated increased expression of znf491 (ZNF491, Fc.:2.52x), znf560 (ZNF560, Fc.:2.52x), znf516 (ZNF516, Fc.:2.52x)[104], esrrg (ESRRG, Fc.:2.52x) [105] and the enhanced expression of translation and post-translation factors, such as GALNT5 (GALNT5 Fc.:8.84x), MUC3A (MUC3A, Fc.:6.31x) MUC6 (MUC6 Fc.:3.36x), MUC4 (MUC4 Fc.:3.15x) [106-108], PIWIL3 (PIWIL3 Fc.:3.79x), PIWIL4 (PIWIL4, Fc:3.79x, F9 (F9, Fc.:2,52x) and GFPT2 (GFPT2 Fc.:2.52x) support the existence of this highly accelerated metabolism. Downregulation of NDUFA13 (-4.11x), a negative regulator of translation [109], suggests that also the activity of genes or group of genes controlled by this regulator is crucial in the battle between the invader and the host. During this encounter, maintenance of the intracellular homeostasis is crucial that is represented by the increased expression of TRPM6 contributing to Mg homeostasis (Table 2) a key element of several enzymatic functions.
The markedly activated levels of CHCA1 (CHCA1, Fc.:8.84x)[59] and LPO (LPO, Fc.:5.05) [60] showed that the eukaryotic cell containing the vacuolised C. jejuni 1-3 hours after infection underwent a marked stress situation. Decreased expression of CHRNE, a regulator of membrane potential (CHRNE, Fc.:-4.74x), assumes the development of an osmotic shock. On the other hand, the drastically decreased expression of GAPDHS ( GAPDHS, Fc.:-4.43x), an enzyme taking part in glycolysis [110], clearly indicates a partial slowdown of some parts of the metabolic machinery of the eukaryotic cell.
Stress situations are characterised by the fact that the cell pauses its non-essential activities and focuses on saving energy. One feature of this may be the activation of the cell cycle arrest gene, sesn2 (SESN2, Fc.:3.23x) [111], thus stopping proliferation, an energy consuming process. Another example for energy saving is the decreased expressions of PARVG (PARVG, Fc.: -3.95x) [112] taking part in matrix protein synthesis and matrix protein processes. The reduced expression of matrix proteins could either facilitate the killing of the infected cell by making itself more accessible and at the same time more vulnerable to the damaging enzymes of macrophages. This hypothesis is supported by the powerful increase of EDN2 (EDN2, Fc.:3,68x) [42] a macrophage chemoattractant and the increased expression of the formerly mentioned ULBP3, responsible for natural killer cell activation.
The decreased expression of FAM132A, a negative regulator of inflammation [113], suggests an induced inflammation in the invaded INT407 cell. Activation of relb, birC3, nfkbia [114] further supports this observation. It is important to note, that as a consequence of bacterial infections inflammatory diseases and other pathological conditions, such as tumors can develop. In this context, C. jejuni infection has been implicated in the development of Guillan-Barre and Miller-Fisher syndromes (GBS and MFS). Reason of the more or less unaffected expression levels of genes coding for, ptgs2 (PTGS2, Fc.:1.2x), anxa3 (ANXA3, Fc.:1.32x) and cerb1 (CERB1, 1.73x) (Table 5.), four gene products considered to be associated with GBS, may be that the effects of the affected genes manifest themselves only after a long period of time or, as it was recently suggested, they are associated only with certain C. jejuni serotypes [81,82,115].
The slight increase in the expression levels of genes associated with tumor genesis supports recent assumptions and findings that certain bacterial infections increase the risk of developing malignant tumors in the colon [116], the biliary tract [117], and the esophagus [118]. Although tumor genesis is a complex process about which our knowledge is still limited, the high expression levels of serpinB9 (SERPINB9, Fc.: 11.37x) and tacr2 (TACR2, Fc.: 8.84x) [66,90], two proteins associated with tumor formation, supports the potential role of C. jejuni in the development of these pathological conditions.
5. Conclusions
Results of this study show that the expression of a wide repertoire of genes in the INT407 epithelial cell line changes during invasion of C. jejuni. Further studies can clarify the roles of concrete genes during the invasion process, and could help to reveal those till unknown molecular processes by that certain cell types, like macrophages, are able to defeat this zoonotic bacterium, in contrast epithelial cells where eliminiation of C. jejuni is inhibited.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization, György Schneider; methodology, József Kun and György Schneider; software, József Kun and Péter Urbán; validation, József Kun, Beáta Polgár; formal analysis, Anita Steinbach; investigation, Anita Steinbach; resources, György Schneider; data curation, József Kun and Beáta Polgár; writing—original draft preparation, Anita Steinbach; writing—review and editing, György Schneider. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sher, A.A.; Ashraf, M.A.; Mustafa, B.E.; Raza, M.M. Epidemiological trends of foodborne Campylobacter outbreaks in the United States of America, 1998–2016. Food Microbiology 2021, 97, 103751. [Google Scholar] [CrossRef] [PubMed]
- Melby, K.; Dahl, O.P.; Crisp, L.; Penner, J.L. Clinical and serological manifestations in patients during a waterborne epidemic due to Campylobacter jejuni. Journal of Infection 1990, 21, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.G.; Price, L.; Ahmed, R.; Woodward, D.L.; Melito, P.L.; Rodgers, F.G.; Jamieson, F.; Ciebin, B.; Li, A.; Ellis, A. Characterization of waterborne outbreak–associated Campylobacter jejuni, Walkerton, Ontario. Emerging infectious diseases 2003, 9, 1232. [Google Scholar] [CrossRef] [PubMed]
- Revez, J.; Llarena, A.-K.; Schott, T.; Kuusi, M.; Hakkinen, M.; Kivistö, R.; Hänninen, M.-L.; Rossi, M. Genome analysis of Campylobacter jejuni strains isolated from a waterborne outbreak. BMC genomics 2014, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gahamanyi, N.; Mboera, L.E.; Matee, M.I.; Mutangana, D.; Komba, E.V. Prevalence, risk factors, and antimicrobial resistance profiles of thermophilic Campylobacter species in humans and animals in sub-saharan Africa: A systematic review. International Journal of Microbiology 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Hlashwayo, D.F.; Sigauque, B.; Noormahomed, E.V.; Afonso, S.M.; Mandomando, I.M.; Bila, C.G. A systematic review and meta-analysis reveal that Campylobacter spp. and antibiotic resistance are widespread in humans in sub-Saharan Africa. PLoS One 2021, 16, e0245951. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.J. Microbial contamination of drinking water and disease outcomes in developing regions. Toxicology 2004, 198, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clinical microbiology reviews 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Samie, A.; Moropeng, R.C.; Tanih, N.F.; Dillingham, R.; Guerrant, R.; Bessong, P.O. Epidemiology of Campylobacter infections among children of 0–24 months of age in South Africa. Archives of Public Health 2022, 80, 107. [Google Scholar] [CrossRef]
- Damborg, P.; Olsen, K.E.; Møller Nielsen, E.; Guardabassi, L. Occurrence of Campylobacter jejuni in pets living with human patients infected with C. jejuni. Journal of Clinical Microbiology 2004, 42, 1363–1364. [Google Scholar] [CrossRef]
- Ghosh, R.; Uppal, B.; Aggarwal, P.; Chakravarti, A.; Jha, A.K. Increasing antimicrobial resistance of Campylobacter jejuni isolated from paediatric diarrhea cases in a tertiary care hospital of New Delhi, India. Journal of clinical and diagnostic research: JCDR 2013, 7, 247. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Ramamurthy, T.; Bhattacharya, M.K.; Rajendran, K.; Mukhopadhyay, A.K. Campylobacter jejuni in hospitalized patients with diarrhea, Kolkata, India. Emerging Infectious Diseases 2013, 19, 1155. [Google Scholar] [CrossRef] [PubMed]
- Budge, S.; Barnett, M.; Hutchings, P.; Parker, A.; Tyrrel, S.; Hassard, F.; Garbutt, C.; Moges, M.; Woldemedhin, F.; Jemal, M. Risk factors and transmission pathways associated with infant Campylobacter spp. prevalence and malnutrition: a formative study in rural Ethiopia. PLoS One 2020, 15, e0232541. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Mead, D.J.; Hayes, S.F.; Cieplak Jr, W. Translocation of Campylobacter jejuni across human polarized epithelial cell monolayer cultures. Journal of Infectious Diseases 1992, 166, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Ketley, J.M. Pathogenesis of enteric infection by Campylobacter. Microbiology 1997, 143, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Acheson, D.; Allos, B.M. Campylobacter jejuni Infections: Update on Emerging Issues and Trends. Clinical Infectious Diseases 2001, 32, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Kovács, J.K.; Cox, A.; Schweitzer, B.; Maróti, G.; Kovács, T.; Fenyvesi, H.; Emődy, L.; Schneider, G. Virulence traits of inpatient Campylobacter jejuni isolates, and a transcriptomic approach to identify potential genes maintaining intracellular survival. Microorganisms 2020, 8, 531. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Amill, V.; Konkel, M.E. Secretion of Campylobacter jejuni Cia proteins is contact dependent. In Mechanisms in the Pathogenesis of Enteric Diseases 2, Springer: 1999; pp. 225–229. [CrossRef]
- Rivera-Amill, V.; Kim, B.J.; Seshu, J.; Konkel, M.E. Secretion of the virulence-associated Campylobacter invasion antigens from Campylobacter jejuni requires a stimulatory signal. The Journal of infectious diseases 2001, 183, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Kopecko, D.J.; Hu, L.; Zaal, K.J. Campylobacter jejuni–microtubule-dependent invasion. TRENDS in Microbiology 2001, 9, 389–396. [Google Scholar] [CrossRef]
- Hendrixson, D.R.; DiRita, V.J. Identification of Campylobacter jejuni genes involved in commensal colonization of the chick gastrointestinal tract. Molecular microbiology 2004, 52, 471–484. [Google Scholar] [CrossRef]
- Reid, A.N.; Pandey, R.; Palyada, K.; Naikare, H.; Stintzi, A. Identification of Campylobacter jejuni genes involved in the response to acidic pH and stomach transit. Applied and environmental microbiology 2008, 74, 1583–1597. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.E.; Baqar, S.; Bourgeois, A.L.; Ewing, C.P.; Guerry, P. Campylobacter jejuni-stimulated secretion of interleukin-8 by INT407 cells. Infection and immunity 1999, 67, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-P.; Ingmer, H.; Madsen, M.; Bang, D.D. Cytokine responses in primary chicken embryo intestinal cells infected with Campylobacter jejuni strains of human and chicken origin and the expression of bacterial virulence-associated genes. BMC microbiology 2008, 8, 1–10. [Google Scholar] [CrossRef]
- Al-Salloom, F.S.; Al Mahmeed, A.; Ismaeel, A.; Botta, G.A.; Bakhiet, M. Campylobacter-stimulated INT407 cells produce dissociated cytokine profiles. Journal of Infection 2003, 47, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Borrmann, E.; Berndt, A.; Hänel, I.; Köhler, H. Campylobacter-induced interleukin-8 responses in human intestinal epithelial cells and primary intestinal chick cells. Veterinary microbiology 2007, 124, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Chaloner, G.; Kemmett, K.; Davidson, N.; Williams, N.; Kipar, A.; Humphrey, T.; Wigley, P. Campylobacter jejuni is not merely a commensal in commercial broiler chickens and affects bird welfare. MBio 2014, 5, e01364-14. [Google Scholar] [CrossRef]
- Al-Amri, A.I.; Botta, G.A.; Tabbara, K.S.; Ismaeel, A.Y.; Al-Mahmeed, A.E.; Qareeballa, A.Y.; Dayna, K.M.B.; Bakhiet, M.O. Campylobacter jejuni induces diverse kinetics and profiles of cytokine genes in INT-407 cells. Saudi medical journal 2008, 29, 514. [Google Scholar]
- Nyati, K.K.; Prasad, K.N.; Rizwan, A.; Verma, A.; Paliwal, V.K. TH1 and TH2 response to Campylobacter jejuni antigen in Guillain-Barre syndrome. Archives of neurology 2011, 68, 445–452. [Google Scholar] [CrossRef]
- Sun, X.; Threadgill, D.; Jobin, C. Campylobacter jejuni induces colitis through activation of mammalian target of rapamycin signaling. Gastroenterology 2012, 142, 86–95.e5. [Google Scholar] [CrossRef]
- Nyati, K.K.; Nyati, R. Role of Campylobacter jejuni infection in the pathogenesis of Guillain-Barré syndrome: an update. BioMed research international 2013, 2013. [Google Scholar] [CrossRef]
- Lo, Y. Clinical and immunological spectrum of the Miller Fisher syndrome. Muscle & nerve 2007, 36, 615–627. [Google Scholar]
- Java, A.; Liszewski, M.K.; Hourcade, D.E.; Zhang, F.; Atkinson, J.P. Role of complement receptor 1 (CR1; CD35) on epithelial cells: A model for understanding complement-mediated damage in the kidney. Molecular immunology 2015, 67, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Khera, R.; Das, N. Complement Receptor 1: disease associations and therapeutic implications. Molecular immunology 2009, 46, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Na, N.; Zhang, X.; Zhao, Y. The biological function and significance of CD74 in immune diseases. Inflammation Research 2017, 66, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.; Liu, M.; Ji, B.; Bai, B.; Cheng, B.; Wang, C. Role of the extracellular signal-regulated kinase 1/2 signaling pathway in ischemia-reperfusion injury. Frontiers in physiology 2019, 10, 1038. [Google Scholar] [CrossRef] [PubMed]
- Klimov, E.; Novitskaya, E.; Koval’chuk, S. CD209 (DC-SIGN) – role in the work of the innate immunity and pathogen penetration. Veterinariya, Zootekhniya i Biotekhnologiya 2020, 1, 64–71. [Google Scholar] [CrossRef]
- Shouval, D.S.; Biswas, A.; Goettel, J.A.; McCann, K.; Conaway, E.; Redhu, N.S.; Mascanfroni, I.D.; Al Adham, Z.; Lavoie, S.; Ibourk, M. Interleukin-10 receptor signaling in innate immune cells regulates mucosal immune tolerance and anti-inflammatory macrophage function. Immunity 2014, 40, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.D. Structure and function of natural-killer-cell receptors. Immunologic research 2003, 27, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Fällman, M.; Andersson, R.; Andersson, T. Signaling properties of CR3 (CD11b/CD18) and CR1 (CD35) in relation to phagocytosis of complement-opsonized particles. The Journal of Immunology 1993, 151, 330–338. [Google Scholar] [CrossRef]
- Lupardus, P.J.; Garcia, K.C. The structure of interleukin-23 reveals the molecular basis of p40 subunit sharing with interleukin-12. Journal of molecular biology 2008, 382, 931–941. [Google Scholar] [CrossRef]
- Grimshaw, M.J.; Wilson, J.L.; Balkwill, F.R. Endothelin-2 is a macrophage chemoattractant: implications for macrophage distribution in tumors. European journal of immunology 2002, 32, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- DeDiego, M.L.; Martinez-Sobrido, L.; Topham, D.J. Novel functions of IFI44L as a feedback regulator of host antiviral responses. Journal of Virology 2019, 93. [Google Scholar] [CrossRef]
- Starlets, D.; Gore, Y.; Binsky, I.; Haran, M.; Harpaz, N.; Shvidel, L.; Becker-Herman, S.; Berrebi, A.; Shachar, I. Cell-surface CD74 initiates a signaling cascade leading to cell proliferation and survival. Blood 2006, 107, 4807–4816. [Google Scholar] [CrossRef]
- Urban, B.C.; Willcox, N.; Roberts, D.J. A role for CD36 in the regulation of dendritic cell function. Proceedings of the National Academy of Sciences 2001, 98, 8750–8755. [Google Scholar] [CrossRef]
- Kishi, Y.; Kondo, T.; Xiao, S.; Yosef, N.; Gaublomme, J.; Wu, C.; Wang, C.; Chihara, N.; Regev, A.; Joller, N. Protein C receptor (PROCR) is a negative regulator of Th17 pathogenicity. Journal of Experimental Medicine 2016, 213, 2489–2501. [Google Scholar] [CrossRef]
- Preza, G.C.; Tanner, K.; Elliott, J.; Yang, O.O.; Anton, P.A.; Ochoa, M.-T. Antigen-presenting cell candidates for HIV-1 transmission in human distal colonic mucosa defined by CD207 dendritic cells and CD209 macrophages. AIDS research and human retroviruses 2014, 30, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, S.; Ho, A.; de Waal Malefyt, R.; Moore, K.W. Expression cloning and characterization of a human IL-10 receptor. Journal of Immunology (Baltimore, Md.: 1950) 1994, 152, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Fourcade, S.; Ruiz, M.; Camps, C.; Schluter, A.; Houten, S.M.; Mooyer, P.A.; Pampols, T.; Dacremont, G.; Wanders, R.J.; Giros, M. A key role for the peroxisomal ABCD2 transporter in fatty acid homeostasis. American Journal of Physiology-Endocrinology and Metabolism 2009, 296, E211–E221. [Google Scholar] [CrossRef]
- Chubanov, V.; Gudermann, T. Trpm6. Mammalian Transient Receptor Potential (TRP) Cation Channels: Volume I 2014, 503-520. [CrossRef]
- van der Wijst, J.; Bindels, R.J.; Hoenderop, J.G. Mg2+ homeostasis: the balancing act of TRPM6. Current opinion in nephrology and hypertension 2014, 23, 361–369. [Google Scholar] [CrossRef]
- Tsai, S.H.; Kinoshita, M.; Kusu, T.; Kayama, H.; Okumura, R.; Ikeda, K.; Shimada, Y.; Takeda, A.; Yoshikawa, S.; Obata-Ninomiya, K. The ectoenzyme E-NPP3 negatively regulates ATP-dependent chronic allergic responses by basophils and mast cells. Immunity 2015, 42, 279–293. [Google Scholar] [CrossRef]
- Wang, Q.; Karvelsson, S.T.; Kotronoulas, A.; Gudjonsson, T.; Halldorsson, S.; Rolfsson, O. Glutamine-fructose-6-phosphate transaminase 2 (GFPT2) Is upregulated in breast epithelial–mesenchymal transition and responds to oxidative stress. Molecular & Cellular Proteomics 2022, 21. [Google Scholar]
- Signorini, C.; De Felice, C.; Durand, T.; Oger, C.; Galano, J.-M.; Leoncini, S.; Pecorelli, A.; Valacchi, G.; Ciccoli, L.; Hayek, J. Isoprostanes and 4-hydroxy-2-nonenal: markers or mediators of disease? Focus on Rett syndrome as a model of autism spectrum disorder. Oxidative Medicine and Cellular Longevity 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers–assembly, dynamics and biological roles. Journal of cell science 2012, 125, 1855–1864. [Google Scholar] [CrossRef]
- Hu, G.; Tang, J.; Zhang, B.; Lin, Y.; Hanai, J.-i.; Galloway, J.; Bedell, V.; Bahary, N.; Han, Z.; Ramchandran, R. A novel endothelial-specific heat shock protein HspA12B is required in both zebrafish development and endothelial functions in vitro. Journal of cell science 2006, 119, 4117–4126. [Google Scholar] [CrossRef]
- Noh, J.-Y.; Lee, H.; Song, S.; Kim, N.S.; Im, W.; Kim, M.; Seo, H.; Chung, C.-W.; Chang, J.-W.; Ferrante, R.J. SCAMP5 links endoplasmic reticulum stress to the accumulation of expanded polyglutamine protein aggregates via endocytosis inhibition. Journal of Biological Chemistry 2009, 284, 11318–11325. [Google Scholar] [CrossRef]
- Zhang, L.; Li, L.; Gao, G.; Wei, G.; Zheng, Y.; Wang, C.; Gao, N.; Zhao, Y.; Deng, J.; Chen, H. Elevation of GPRC5A expression in colorectal cancer promotes tumor progression through VNN-1 induced oxidative stress. International journal of cancer 2017, 140, 2734–2747. [Google Scholar] [CrossRef]
- Mungrue, I.N.; Pagnon, J.; Kohannim, O.; Gargalovic, P.S.; Lusis, A.J. CHAC1/MGC4504 is a novel proapoptotic component of the unfolded protein response, downstream of the ATF4-ATF3-CHOP cascade. The Journal of Immunology 2009, 182, 466–476. [Google Scholar] [CrossRef]
- Kovacs, P.; Juranek, I.; Stankovicova, T.; Svec, P. Lipid peroxidation during acute stress. Die Pharmazie 1996, 51, 51–53. [Google Scholar] [PubMed]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef]
- Wang, J.-N.; Shi, N.; Xie, W.-B.; Guo, X.; Chen, S.-Y. Response gene to complement 32 promotes vascular lesion formation through stimulation of smooth muscle cell proliferation and migration. Arteriosclerosis, thrombosis, and vascular biology 2011, 31, e19–e26. [Google Scholar] [CrossRef]
- Zouein, F.A.; Kurdi, M.; Booz, G.W. HSPA12B and repairing the heart: beauty in simplicity. Cardiovascular Research 2013, 99, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Wuensch, T.; Wizenty, J.; Quint, J.; Spitz, W.; Bosma, M.; Becker, O.; Adler, A.; Veltzke-Schlieker, W.; Stockmann, M.; Weiss, S. Expression analysis of fibronectin type III domain-containing (FNDC) genes in inflammatory bowel disease and colorectal cancer. Gastroenterology research and practice 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Gao, W.; Lin, S.; Chen, H.; Du, B.; Liu, Q.; Lin, X.; Chen, Q. FNDC1 promotes the invasiveness of gastric cancer via Wnt/β-catenin signaling pathway and correlates with peritoneal metastasis and prognosis. Frontiers in Oncology 2020, 10, 590492. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-J.; Wang, J.; Ouyang, C.; Chen, C.; Xu, X.-F.; Ye, X.-Q. Overview of serpin B9 and its roles in cancer. Oncology Reports 2021, 46, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.H.; Christensen, M.E.; Mangan, M.; Prakash, M.D.; Sedelies, K.A.; Smyth, M.; Harper, I.; Waterhouse, N.J.; Bird, P.I. The granzyme B-Serpinb9 axis controls the fate of lymphocytes after lysosomal stress. Cell Death & Differentiation 2014, 21, 876–887. [Google Scholar]
- Kaiserman, D.; Bird, P.I. Control of granzymes by serpins. Cell Death & Differentiation 2010, 17, 586–595. [Google Scholar]
- Asnaghi, L.; White, D.T.; Key, N.; Choi, J.; Mahale, A.; Alkatan, H.; Edward, D.P.; Elkhamary, S.M.; Al-Mesfer, S.; Maktabi, A. ACVR1C/SMAD2 signaling promotes invasion and growth in retinoblastoma. Oncogene 2019, 38, 2056–2075. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, H. CHAC1 exacerbates LPS-induced ferroptosis and apoptosis in HK-2 cells by promoting oxidative stress. Allergologia et Immunopathologia 2023, 51, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Das, D.K.; Ogunwobi, O.O. A novel microRNA-1207-3p/FNDC1/FN1/AR regulatory pathway in prostate cancer. RNA & disease (Houston, Tex.) 2017, 4. [Google Scholar]
- Yunwen, C.; Shanshan, G.; Zhifei, B.; Saijun, C.; Hua, Y. The silencing of FNDC1 inhibits the tumorigenesis of breast cancer cells via modulation of the PI3K/Akt signaling pathway. Molecular Medicine Reports 2021, 23, 1–8. [Google Scholar] [CrossRef]
- Heckmann, B.L.; Zhang, X.; Xie, X.; Liu, J. The G0/G1 switch gene 2 (G0S2): regulating metabolism and beyond. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids 2013, 1831, 276–281. [Google Scholar] [CrossRef]
- Mognol, G.P.; Carneiro, F.R.G.; Robbs, B.K.; Faget, D.V.; Viola, J.P.d.B. Cell cycle and apoptosis regulation by NFAT transcription factors: new roles for an old player. Cell death & disease 2016, 7, e2199–e2199. [Google Scholar]
- Wang, Y.; Liang, H.; Zhou, G.; Hu, X.; Liu, Z.; Jin, F.; Yu, M.; Sang, J.; Zhou, Y.; Fu, Z. HIC1 and miR-23~ 27~ 24 clusters form a double-negative feedback loop in breast cancer. Cell Death & Differentiation 2017, 24, 421–432. [Google Scholar]
- Mehlen, P.; Rabizadeh, S.; Snipas, S.J.; Assa-Munt, N.; Salvesen, G.S.; Bredesen, D.E. The DCC gene product induces apoptosis by a mechanism requiring receptor proteolysis. Nature 1998, 395, 801–804. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, G. A tumor suppressor DLC1: The functions and signal pathways. Journal of Cellular Physiology 2020, 235, 4999–5007. [Google Scholar] [CrossRef]
- Ullmannova, V.; Popescu, N.C. Inhibition of cell proliferation, induction of apoptosis, reactivation of DLC1, and modulation of other gene expression by dietary flavone in breast cancer cell lines. Cancer detection and prevention 2007, 31, 110–118. [Google Scholar] [CrossRef]
- Prasad, K.; Ao, Z.; Yoon, Y.; Wu, M.X.; Rizk, M.; Jacquot, S.; Schlossman, S.F. CD27, a member of the tumor necrosis factor receptor family, induces apoptosis and binds to Siva, a proapoptotic protein. Proceedings of the National Academy of Sciences 1997, 94, 6346–6351. [Google Scholar] [CrossRef]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell death & differentiation 1999, 6, 99–104. [Google Scholar]
- Kieseier, B.; Clements, J.; Pischel, H.; Wells, G.; Miller, K.; Gearing, A.; Hartung, H.P. Matrix metalloproteinases MMP-9 and MMP-7 are expressed in experimental autoimmune neuritis and the Guillain-Barre syndrome. Annals of neurology 1998, 43, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.; Newsom-Davis, J.; Perkin, G.; Pierce, J. Controlled trial of prednisolone in acute polyneuropathy. The Lancet 1978, 312, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Gu, W.; McLandsborough, L. Low concentration of ethylenediaminetetraacetic acid (EDTA) affects biofilm formation of Listeria monocytogenes by inhibiting its initial adherence. Food microbiology 2012, 29, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Youssef, S.; Hur, E.M.; Ho, P.P.; Han, M.H.; Lanz, T.V.; Phillips, L.K.; Goldstein, M.J.; Bhat, R.; Raine, C.S. Blocking angiotensin-converting enzyme induces potent regulatory T cells and modulates TH1-and TH17-mediated autoimmunity. Proceedings of the National Academy of Sciences 2009, 106, 14948–14953. [Google Scholar] [CrossRef] [PubMed]
- Breuer, K.; Foroushani, A.K.; Laird, M.R.; Chen, C.; Sribnaia, A.; Lo, R.; Winsor, G.L.; Hancock, R.E.; Brinkman, F.S.; Lynn, D.J. InnateDB: systems biology of innate immunity and beyond—recent updates and continuing curation. Nucleic acids research 2013, 41, D1228–D1233. [Google Scholar] [CrossRef]
- Connell, S.; Meade, K.G.; Allan, B.; Lloyd, A.T.; Kenny, E.; Cormican, P.; Morris, D.W.; Bradley, D.G.; O'Farrelly, C. Avian resistance to Campylobacter jejuni colonization is associated with an intestinal immunogene expression signature identified by mRNA sequencing. 2012. [CrossRef]
- Wang, B.G.; Yi, D.H.; Liu, Y.F. TLR3 gene polymorphisms in cancer: a systematic review and meta-analysis. Cancer Communications 2015, 34, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y. CD36 tango in cancer: signaling pathways and functions. Theranostics 2019, 9, 4893. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Pan, Y.; Jin, M.; Zhang, M.; Zhang, S.; Li, Q.; Jiang, X.; Liu, H.; Guo, J.; Liu, H. Association of genetic variants in tachykinins pathway genes with colorectal cancer risk. International journal of colorectal disease 2012, 27, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Jianfeng, W.; Yutao, W.; Jianbin, B. TACR2 is associated with the immune microenvironment and inhibits migration and proliferation via the Wnt/β-catenin signaling pathway in prostate cancer. Cancer Cell International 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, L.; Shi, B.; Bao, J.; Zheng, D.; Zhou, B.; Shi, J. GALNT5 uaRNA promotes gastric cancer progression through its interaction with HSP90. Oncogene 2018, 37, 4505–4517. [Google Scholar] [CrossRef] [PubMed]
- Marín, F.; Bonet, C.; Munoz, X.; García, N.; Pardo, M.L.; Ruiz-Liso, J.M.; Alonso, P.; Capella, G.; Sanz-Anquela, J.M.; González, C.A. Genetic variation in MUC1, MUC2 and MUC6 genes and evolution of gastric cancer precursor lesions in a long-term follow-up in a high-risk area in Spain. Carcinogenesis 2012, 33, 1072–1080. [Google Scholar] [CrossRef]
- Chang, Y.J.; Bae, J.; Zhao, Y.; Lee, G.; Han, J.; Lee, Y.H.; Koo, O.J.; Seo, S.; Choi, Y.-K.; Yeom, S.C. In vivo multiplex gene targeting with Streptococcus pyogens and Campylobacter jejuni Cas9 for pancreatic cancer modeling in wild-type animal. Journal of Veterinary Science 2020, 21. [Google Scholar] [CrossRef]
- Xia, X.; Wu, W.; Huang, C.; Cen, G.; Jiang, T.; Cao, J.; Huang, K.; Qiu, Z. SMAD4 and its role in pancreatic cancer. Tumor Biology 2015, 36, 111–119. [Google Scholar] [CrossRef]
- Naderi, A.; Couch, F.J. BRCA2 and pancreatic cancer. International journal of gastrointestinal cancer 2002, 31, 99–106. [Google Scholar] [CrossRef]
- Olakowski, M.; Tyszkiewicz, T.; Jarzab, M.; Król, R.; Oczko-Wojciechowska, M.; Kowalska, M.; Kowal, M.; Gala, G.M.; Kajor, M.; Lange, D. NBL1 and anillin (ANLN) genes over-expression in pancreatic carcinoma. Folia histochemica et cytobiologica 2009, 47, 249–255. [Google Scholar] [CrossRef]
- Singh, A.P.; Chaturvedi, P.; Batra, S.K. Emerging roles of MUC4 in cancer: a novel target for diagnosis and therapy. Cancer research 2007, 67, 433–436. [Google Scholar] [CrossRef]
- Pinho, R.M.; Garas, L.C.; Huang, B.C.; Weimer, B.C.; Maga, E.A. Malnourishment affects gene expression along the length of the small intestine. Frontiers in Nutrition 2022, 9, 894640. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Qin, W.; Buya, M.; Dong, X.; Zheng, W.; Lu, W.; Chen, J.; Guo, Q.; Wu, Y. VNN1, a potential biomarker for pancreatic cancer-associated new-onset diabetes, aggravates paraneoplastic islet dysfunction by increasing oxidative stress. Cancer Letters 2016, 373, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhou, W.; Wu, X.; Hu, Z.; Qiu, L.; Zhang, H.; Chen, X.; Zhang, S.; Lu, Z. CTLs: Killers of intracellular bacteria. Frontiers in Cellular and Infection Microbiology 2022, 1627. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.; Mangan, N.E.; Cumming, H.; Horvat, J.C.; Mayall, J.R.; Stifter, S.A.; De Weerd, N.; Roisman, L.C.; Rossjohn, J.; Robertson, S.A. Interferon-ε protects the female reproductive tract from viral and bacterial infection. Science 2013, 339, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Hamza, E.; Kittl, S.; Kuhnert, P. Temporal induction of pro-inflammatory and regulatory cytokines in human peripheral blood mononuclear cells by Campylobacter jejuni and Campylobacter coli. PloS one 2017, 12, e0171350. [Google Scholar] [CrossRef]
- Kvansakul, M. Viral infection and apoptosis. MDPI: 2017; p 356. [CrossRef]
- Shannon, M.; Kim, J.; Ashworth, L.; Branscomb, E.; Stubbs, L. Tandem zinc-finger gene families in mammals: insights and unanswered questions. DNA Sequence 1998, 8, 303–315. [Google Scholar] [CrossRef]
- Li, W.; Gong, M.; Park, Y.P.; Elshikha, A.S.; Choi, S.-C.; Brown, J.; Kanda, N.; Yeh, W.-I.; Peters, L.; Titov, A.A. Lupus susceptibility gene Esrrg modulates regulatory T cells through mitochondrial metabolism. JCI insight 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Kitamoto, S.; Yamada, N.; Yokoyama, S.; Houjou, I.; Higashi, M.; Yonezawa, S. Promoter hypomethylation contributes to the expression of MUC3A in cancer cells. Biochemical and biophysical research communications 2010, 397, 333–339. [Google Scholar] [CrossRef]
- Sheng, Y.H.; Hasnain, S.Z.; Florin, T.H.; McGuckin, M.A. Mucins in inflammatory bowel diseases and colorectal cancer. Journal of gastroenterology and hepatology 2012, 27, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.K.; Praharaj, A.B.; Dehury, B.; Negi, S. Exploring the role and diversity of mucins in health and disease with special insight into non-communicable diseases. Glycoconjugate journal 2015, 32, 575–613. [Google Scholar] [CrossRef]
- Pinto, M.; Máximo, V. NDUFA13 (NADH: ubiquinone oxidoreductase subunit A13). Atlas of Genetics and Cytogenetics in Oncology and Haematology 2018, 8. [Google Scholar]
- Taugbøl, A.; et al. Small changes in gene expression of targeted osmoregulatory genes when exposing marine and freshwater threespine stickleback (Gasterosteus aculeatus) to abrupt salinity transfers. PLoS One 2014, 9, e106894. [Google Scholar] [CrossRef]
- Kornberg, M.D.; Bhargava, P.; Kim, P.M.; Putluri, V.; Snowman, A.M.; Putluri, N.; Calabresi, P.A.; Snyder, S.H. Dimethyl fumarate targets GAPDH and aerobic glycolysis to modulate immunity. Science 2018, 360, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Kowalsky, A.H.; Namkoong, S.; Mettetal, E.; Park, H.-W.; Kazyken, D.; Fingar, D.C.; Lee, J.H. The GATOR2–mTORC2 axis mediates Sestrin2-induced AKT Ser/Thr kinase activation. Journal of Biological Chemistry 2020, 295, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Korenbaum, E.; Olski, T.M.; Noegel, A.A. Genomic organization and expression profile of the parvin family of focal adhesion proteins in mice and humans. Gene 2001, 279, 69–79. [Google Scholar] [CrossRef]
- Sargolzaei, J.; Chamani, E.; Kazemi, T.; Fallah, S.; Soori, H. The role of adiponectin and adipolin as anti-inflammatory adipokines in the formation of macrophage foam cells and their association with cardiovascular diseases. Clinical Biochemistry 2018, 54, 1–10. [Google Scholar] [CrossRef]
- Finsterer, J. Triggers of Guillain–Barré syndrome: campylobacter jejuni predominates. International Journal of Molecular Sciences 2022, 23, 14222. [Google Scholar] [CrossRef]
- Chang, K.-H.; Chuang, T.-J.; Lyu, R.-K.; Ro, L.-S.; Wu, Y.-R.; Chang, H.-S.; Huang, C.-C.; Kuo, H.-C.; Hsu, W.-C.; Chu, C.-C. Identification of gene networks and pathways associated with Guillain-Barre syndrome. PloS one 2012, 7, e29506. [Google Scholar] [CrossRef]
- He, Z.; Gharaibeh, R.Z.; Newsome, R.C.; Pope, J.L.; Dougherty, M.W.; Tomkovich, S.; Pons, B.; Mirey, G.; Vignard, J.; Hendrixson, D.R. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut 2019, 68, 289–300. [Google Scholar] [CrossRef]
- de Savornin Lohman, E.; Duijster, J.; Groot Koerkamp, B.; van der Post, R.; Franz, E.; Mughini Gras, L.; de Reuver, P. Severe Salmonella spp. Or Campylobacter spp. infection and the risk of biliary tract cancer: a population-based study. Cancers 2020, 12, 3348. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Man, S.M.; Mitchell, H.M. Is Campylobacter to esophageal adenocarcinoma as Helicobacter is to gastric adenocarcinoma? Trends in microbiology 2015, 23, 455–462. [Google Scholar] [CrossRef]
Figure 1.
Genes of the Campylobacter jejuni strain CjTD-199 affected between the 1st and 3rd hour of the invasion process. Other functions are the following: response to toxin, response to oxidative stress, receptor-mediated endocytosis, stress-activated MAPK cascade, peptidyl-tyrosine phosphorylation, cell-cell junction organization.
Figure 1.
Genes of the Campylobacter jejuni strain CjTD-199 affected between the 1st and 3rd hour of the invasion process. Other functions are the following: response to toxin, response to oxidative stress, receptor-mediated endocytosis, stress-activated MAPK cascade, peptidyl-tyrosine phosphorylation, cell-cell junction organization.
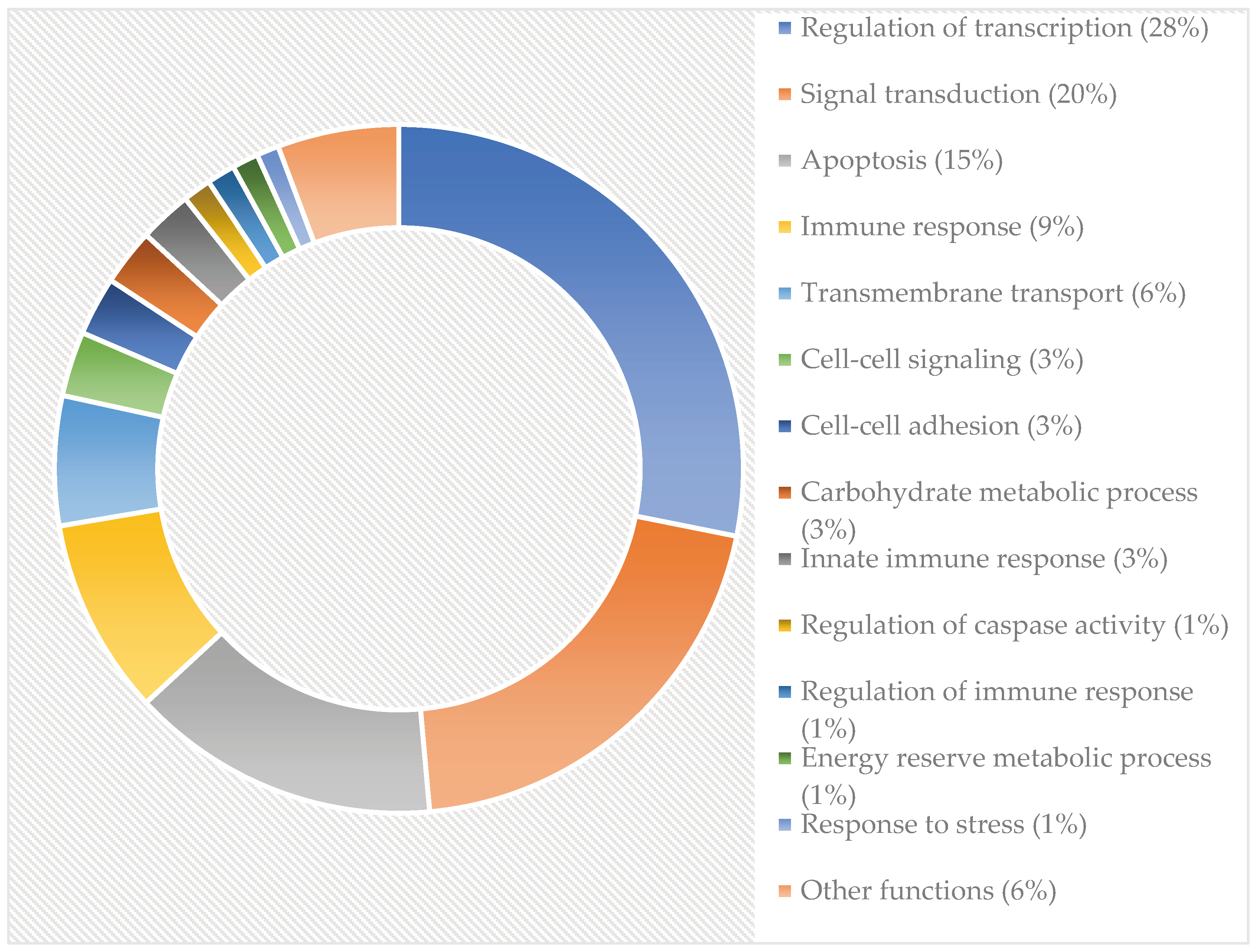
Table 1.
Affected genes associated with immunfunctions.
| Gene Name | Function | 1vs3 Experiment - Fold Change (normalized values) | Refer. |
|---|---|---|---|
| ULBP3 | regulation of immune response, natural killer cell activation | 10.1090 | [39] Sun 2013 |
| CR1 | innate immune response phagocytosis |
7.581748 | [40] Fällman et al. 1996 |
| IL23R | positive regulation of defense response to virus by host, inflammatory response | 6.31 | [41] Lupardus et Garcia 2008 |
| EDN2 | macrophage activation, signaling patway | 3.68557 | [42] Grimshaw et al 2002 |
| IFI44L | defense response to virus, immune response | 3.317 | [43] DeDiego et al.2019 |
| CD74 | negative regulation of mature B cell apoptosis, positive regulation of neutrophil chemotaxis, positive regulation of T-helper 2 type immune response, T cell selection, negative regulation of apoptosis, positive regulation of B cell proliferation, positive regulation of ERK1 and ERK2 cascade, antigen processing and presentation |
2.5272 | [35] Su et al.2017 [44] Starlets et al.2006 |
| CD36 | antigen processing and presentation of exogenous, peptide antigen via MHC class I, antigen processing and presentation of peptide antigen via MHC class I |
2.73785 | [45] Urban et al. 2001 |
| PROCR | antigen processing and presentation, PROCR acted as a negative regulator of Th17 pathogenicity | 2.5272 | [46] Kishi et al. 2016 |
| CD209 | regulation of T cell proliferation, antigen processing and presentation, innate immune response |
2.5272 | [47] Preza et al. 2014 |
| IL10RA | inhibits the synthesis of proinflammatoric cytokines | 2.5272 | [48] Liu et at. 1994 |
Table 2.
Genes with metabolic functions.
| Gene name | Function | Fold change | References |
|---|---|---|---|
| ABCD2 | very-long-chain fatty acid metabolic process | 13.89 | [49] Fourcade et al.2009 |
| TRPM6 | Mg2+ channel, and uptake regulator | 11.37 | [51] van der Wijst et al.2014 |
| ENPP3 | phosphate metabolic process, nucleoside triphosphate catabolic process | 8.84537 | [52] Tsai et al. 2015 |
| GFPT2 | glutamine metabolic process, fructose 6-phosphate metabolic process | 2.52 | [53] Wang et al.2022 |
Table 5.
Genes involved in the potential development of chronic conditions.
| Gene name | 1vs3 | referenc. | |
|---|---|---|---|
| Guillan- Barré s. disease severity (GBS) | PTGS2 | 1.188331 | [83] Chang et al.(2012) |
| ANXA3 | 1.315609 | [82] Hughes et al. (1978) |
|
| CREB1 | 1.732517 | [82] Hughes et al. (1978) |
|
| Inflammatory | RELB | 1.624660 | [85] Breuer et al. 2013 |
| BIRC3 | 1.958618 | [85] Breuer et al. 2013 |
|
| NFKBIA | -2.553647 | [85] Breuer et al. 2013 |
|
| Autoimmune inflammation | ACE | 3.79087 |
[86] Connell et al. 2012 |
| General cancer markers | TLR3 | 3.79087 | [87] Wang et al. 2015 |
| CD36 | 2.73785 | [88] Wang et Li 2019 |
|
| Tumorgenesis | SERPINB9 | 11.37262 | Wang et al. 2021 |
| FNDC1 | 7.581749 | Jiang et al. 2020 | |
| TACR2 | 8.845373 | [89] Yu et al. 2012 [90] Jianfeng et al. 2021 |
|
| Gastric cancer | GALNT5 | 8.84537 | [65,91] Jiang et al.2020 Guo et al.2018 |
| MUC6 | 3.36 | [92] Marín et al. 2012 |
|
| Pancreatic cancer |
KRAS | 2.011985 | [93] Chang et al. 2020 |
| SMAD4 | 1.34157 | [94] Xia et al. 2015 |
|
| BRCA2 | 1.231316 | [95] Naderi et Couch. 2002 |
|
| NBL1 | 5.054499 | [96] Olakowski et al. 2009 |
|
| MUC4 | 3.15 | [97] Singh et al. 2007 |
|
| Oxidative stress in the intestine | VNN1 | 379.087 | [98] Pinho et al. 2022 [99] Kang et al. 2016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



