
Submitted:
22 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Precision cancer medicine primarily aims to identify individual patient genomic variations and exploit vulnerabilities in cancer cells to select suitable patients for specific drugs. These genomic features are commonly determined by gene sequencing prior to therapy, to identify individuals who would be most responsive. This precision approach in cancer therapeutics remains a powerful tool that benefits a smaller pool of patients, sparing others from unnecessary treatments. A limitation of this approach is that proteins, not genes, are the ultimate effectors of biological functions, and therefore the targets of therapeutics. An additional dimension in precision medicine that considers an individual’s cytokine response to cancer therapeutics is proposed. Cytokine responses to therapy are multifactorial and vary among individuals. Thus, precision is dictated by the nature and magnitude of cytokine responses in the tumor microenvironment exposed to therapy. This review highlights cytokine responses as modules for precision medicine in cancer therapy, including potential challenges. For solid tumors, both detectability of cytokines in tissue fluids and their being amenable to routine sensitive analyses could address the difficulty of specimen collection for diagnosis and monitoring. Therefore, in precision cancer medicine, cytokines offer rational targets that can be utilized to enhance the efficacy of cancer therapy.
Keywords:
Precision medicine
; pharmacogenomics
; cytokines
; chemotherapy
; radiotherapy
1. Precision Cancer Medicine
At the center of precision medicine, often interchangeably termed as ‘personalized medicine’ or ‘individualized medicine,’ is the need to deliver ‘the right drug, with the right dose at the right time to the right patient’ [1,2]. Irrespective of the different interpretations of these terminologies, the overarching premise remains the recognition of individual patient’s molecular differences as the basis to guide disease management, and consideration of individual patients as independent biological units to avoid ‘group’-prescription of therapy and subsequent side effects or non-responsiveness.
Currently, the most common application of precision cancer medicine involves pharmacogenomics, for which genomic determinants of drug metabolism (pharmacokinetics and pharmacodynamics) can be predicted from the polymorphisms and expressions of the enzymes that are involved [3,4,5]. Specifically, genomic information from normal or malignant cells is used to select an appropriate drug or patient, with the intent to avoid unnecessary treatment (Figure 1). Unlike proteomic, metabolomic and other elements, genomic data is the least variable, making it the most stable and widely comparable across multiple platforms.
It is practical and convenient to make clinical ‘individualization’ of patients based on the identification of genomic (DNA) or RNA signatures, and as such, to sort patients into ‘most or least likely to respond,’ or ‘most likely to experience side-effects’ groups. However, precision medicine centered on DNA/RNA-based tools leaves a translational uncertainty, since proteins are the actionable targets and ultimate effectors of biological functions. This uncertainty becomes obvious when the proportions of patients ‘expected’ to respond are compared to patients who clinically ‘respond.’ Understandably, this uncertainty is multifactorial in origin and remains a challenge to identify and address the reasons for non-responsiveness, causing doubts about the entire precision approach [6,7].
2. Cytokine pathways as therapeutic targets
Cytokines, cytokine receptors, and associated pathways offer opportunities to target signaling pathways associated with various disease conditions. Given their roles, inflammatory cytokines and receptors, as potential therapeutic targets, have been the focus of researchers, clinicians, and pharmaceutical companies. The most clinically advanced of these include antibody-based anti-psoriasis drugs targeting interleukin 23 (IL23, tildrakizumab), IL17A (secukinumab, ixekizumab) or IL17R (broadalumab). Additionally, anti-TNF antibodies have been deployed for the treatment of rheumatoid arthritis and psoriasis [8,9,10]. Among the most widely used drugs are an anti-TNFα antibody drug for arthritis marketed for humans as Humira, and an anti-IL-31 antibody for canine atopic or allergic dermatitis used under the trade name Cytopoint [11]. An IL6R-targeting mAb (tocilizumab) has been approved to treat a few inflammatory conditions, including rheumatoid arthritis and cytokine release syndrome after therapies [12]. However, the utility of tocilizumab against COVID-19 could not be established unequivocally, although IL6 levels correlated with severe disease [13,14]. IL6R antagonists tocilizumab and sarilumab are currently recommended for severely or critically ill COVID-19 patients (Therapeutics and COVID-19: living guideline. Geneva: World Health Organization [15]). Two cytokines, IL2 and IFNα, exhibited moderate therapeutic effects for the treatment of a variety of malignant conditions and were thus approved by the Food and Drug Administration (FDA). IL2 is used for advanced metastatic renal cell cancer and melanoma [16]; IFNα is indicated for hairy cell leukemia, follicular non-Hodgkins's lymphoma, melanoma, and AIDS-related Kaposi’s sarcoma. In 2015, IL15 was approved for treatment of solid tumors [17].
Chemokines form a sub-group of cytokines characterized as molecules mediating the trafficking of immune cells [18]. Although several drugs targeting chemokine pathways have been developed, only a few have been approved for a disease condition. As an example, maraviroc, a CCR5 co-receptor antagonist that interferes with HIV entry into CD4 T cells, was approved in 2007 for treatment of HIV patients, in combination with other antiviral medications [19,20].
Currently, the primary goal in cytokine-directed therapy is to block the interaction of these molecules with the corresponding receptors, or to dampen the pathways downstream of the receptors. The impetus to advance the inhibitory approaches is because of the role of most of the targeted cytokines in enhancing disease processes, commonly inflammation. Unlike inhibition, an approach to enhance the expression or functions of cytokines for cancer therapy is still experimental, or is expected to be more challenging than inhibition, because of the need to fine-tune the activity of cytokines. For example, IL2, a potent T-cell mitogen, enhances anti-tumor responses of the immune system, but high doses can be detrimental to patients because of excessive cytokine release and activity [21].
3. Cells of cytokine origin
Most cell types within the tumor microenvironment, including tumor cells, immune cells such as macrophages, dendritic cells, and T cells, as well as stromal cells such as fibroblasts and endothelial cells produce cytokines in response to chemotherapy or radiation. Thus, tumor cells are involved in their own response to treatment. Macrophages, which are involved in the immune response, also produce cytokines in response to chemotherapy and radiation. Dendritic cells, another type of immune cells, also produce cytokines [22,23]; their role in cytokine production may involve the presentation of tumor antigens to T-cells, leading to an immune response. Antigen-stimulated CD4+ T-cells [24], along with natural killer T cells [25], CD8+ T-cells [26], mast cells [27], and dendritic cells [28] are the primary producers of IL2. Endogenous IL2 therapy increases the expression of CD25 and IL2 receptor, with subsequent proliferation of CD8+ T-cells [29]. IL2 enhances the expression of LAMP-1 [30] on the surface of CD8+ T-cells while decreasing the expression of PD-1 [31], an inhibitory receptor, thereby facilitating CD8+ T-cell cytotoxicity.
4. Cytokine responses in solid-tumors therapy: an emerging theme in cancer precision medicine
As mentioned above, the concept of targeting cytokines or cytokine pathways to control specific diseases or processes is not new. After the role of receptors in viral entry was established, these pathways have even been targeted for infectious diseases such as HIV (CXCR4 and CCR5) [32,33,34]. Although most primary solid or intraepithelial tumors can be treated with several types of chemotherapy and radiation, adaptive resistance of tumor cells and recurrence of cancer have hampered durable success of these therapies. The central theme of cancer precision medicine has been to identify patients who will favorably respond to a current therapy module, avoiding the side-effects of therapy on those who are least likely to respond [7,35].
For colorectal cancers, the clinical trials and applications of precision medicine have been narrowly focused on microsatellite status and the MAPK pathway proteins downstream of EGFR and HER2 receptors [36]; these are used for targeted therapeutics. Although the results and ultimate impact of these genomics-based approaches are still to be determined, to benefit patients, additional targets for precision medicine need to be identified. Microsatellite instability (MSI), defined by its genomic signatures and pro-mutagenic characteristics, is an appropriate tumor characteristic to identify colon cancer patients better suited for certain chemotherapy regimens, and especially for immunotherapy. However, a substantial proportion of the MSI tumors do not respond to current immunotherapy regimens [37]. Within the multifactorial background of ‘non-responsiveness,’ which may involve added gene alterations, the role for signaling proteins may be substantial. After initiation of standard cancer therapy, a panel of targetable proteins unique to a patient could constitute a framework for precision medicine. Since the type and intensity of protein-response by malignant and normal cells in the tumor microenvironment will vary among individuals, the development and inclusion of ‘therapy-induced cytokine signature’ is emerging as a new dimension of precision medicine [38,39,40].
In malignant cells, cancer therapy using drugs or radiation elicits ‘danger’ signaling pathways, which have, primarily, a protective nature. These signals could be mediated by cytokines, and some cytokine pathways, such as transforming growth factor-beta (TGF-β), may have dual roles of inhibition or acceleration of tumor growth [41,42]. Other cytokine pathways may drive cell death, attract, or deflect immune cells, or induce a senescence-like state to remain dormant through the stress period [43,44,45,46]. Therefore, an analysis of active cytokines in an individual’s cancer microenvironment or tissues during chemotherapy, especially in comparison to the pre-intervention levels or activities, could reveal information about the biological mechanisms operating in the tumor under therapy stress. Such cytokine readouts may correlate with patient clinical outcomes [47,48,49,50]. Since cytokines are readily detectable in tissue fluids, this approach to precision medicine has an added value for clinical monitoring and decision making.
5. Cytokine responses to chemotherapy and radiation
Cytokines are small proteins (peptides or polypeptides or glycoproteins with 5 – 30 kDa molecular weight) that are involved in modulation of the immune response. They are produced by various cells and act as signaling molecules, allowing for communication between different cell types. In the context of cancer, cytokines in the tumor microenvironment may be produced by both neoplastic and non-neoplastic cells that are involved in the regulation of tumor growth, proliferation, survival, and response to therapy. In recent years, there has been growing interest in understanding the role of cytokines in tumor response to chemotherapy and radiation [51,52].
Chemotherapy and radiotherapy induce various types of cytokines. These include pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), IL1, and IL6, as well as anti-inflammatory cytokines, such as IL10 and TGFβ [53]. Other cytokines, such as IFNs, chemokines, and growth factors, are also involved in tumor cytokine responses to chemotherapy and radiation. For example, IFNs enhance the anti-tumor immune response, and chemokines attract immune cells to the tumor site. Furthermore, growth factors, such as epidermal growth factor and vascular endothelial growth factor, are involved in angiogenesis, which is the formation of new blood vessels to support tumor growth [54]. Below, we provide examples of studies that assessed cytokine response after clinical or in vitro exposure of cells to chemotherapy or radiation.
A study comparing treated and untreated cell lines [55] evaluated the effects of clinical chemotherapeutics oxaliplatin, cisplatin, 5-fluorouracil (5-FU), doxorubicin, paclitaxel, docetaxel, and carboplatin, using a panel of 52 cytokines in MCF7 breast cancer (BC) and HCT116 colorectal cancer (CRC) cells. The results showed that chemotherapy-inducible cytokine transcripts have varying potential for prognostic significance in CRC versus BC. For the BC cell line, the expression of TRAIL-R2 and chitinase 3-like 1 were upregulated by most of the drugs; TRAIL and IFN-β were downregulated. Beyond that, the two cell lines did not respond similarly, which suggested drug-specific and tissue-specific cytokine regulation after chemotherapy. Similarly, a comparison of chemotherapy drugs on cytokine production in CRC cell lines showed drug- and p53-dependent signatures, suggesting a personalized approach that considers the p53-status [56]. We also previously reported that topoisomerase inhibitor treatment of CRC cell lines induced cytokines that were clinically associated with patient overall survival [57].
In another study, the cytokine signature of conditioned media produced by the non-tumorigenic mammary epithelial cell line MCF10A, as well as by MCF7 and MDA-MB-231 BC cell lines, following single high doses of irradiation was examined [58]. The results showed that both 9 Gy and 23 Gy of ionizing radiation triggered the release of cytokines and growth factors capable of influencing tumor outcome in a dose-independent and cell-line dependent pattern.
In an investigation that used both in vitro and in vivo CRC models, Wang et al. found CCL20 to be a major cytokine induced after 5-FU chemotherapy and a mediator of regulatory T cell (Treg) recruitment and drug resistance. Blockade of the cytokine by specific antibodies blocked tumor growth in vivo [59]. In another study with non-tumor bearing mice treated to measure the effects of 5-FU on cytokine expression in the brain, Groves TR et al. 2017 found that chemotherapy altered cytokine abundance in the hippocampus, with IL-1, -2, -3, -4, -5, -17, GMCSF (granulocyte macrophage colony stimulating factor), and RANTES expressed at higher levels in treated mice compared to controls, suggesting a mechanism for potential toxic side-effects of the drug in the brain. Oxaliplatin increased the splenic populations of CD4, CD8, and Treg cells, and altered splenic cytokine expression, despite decreasing the splenic mass [60]. Anthracyclines, used in chemotherapy for certain types of solid cancers, induced a type-I interferon response that upregulated CXCL10, resulting in an antitumor outcome [61].
Like chemotherapy, radiotherapy of solid cancers alters local or systemic cytokines [62], and radiotherapy may be combined with cytokine treatments for a beneficial outcome [63]. Nevertheless, unlike the antitumor mechanism of radiotherapy mediated by cell death, the contributions of cytokine-mediated functions of radiotherapy are not clearly understood. For example, although CXCL9, CXCL10, and CXCL16, in response to radiotherapy, mediate anticancer effects, other molecules, such as TGF-β, CCL2, CSF1, CXCL12 and insulin-like growth factor 1, which are also induced by radiotherapy, may create an immunosuppressive microenvironment [63]. After a sub-lethal dose of irradiation, enhanced levels of multifunctional cytokines IL1β, IL6, IL8, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage-colony stimulating factor (GM-CSF), and TNF-α were recorded [48].
In a study aimed to characterize the predictive value of cytokines/chemokines for rectal cancer patients receiving chemoradiation therapy (CRT), platelets, immune system, and cancer cells, cross-linked through various cytokines/chemokines, were examined. The pre-CRT levels of soluble CD40-ligand (sCD40L) and the post-CRT levels of chemokine ligand-5 (CCL5) were associated with the malignant tumor behaviors of depth and invasion, and higher post-CRT IL6 was associated with a poor response [64].
In a separate study, serum cytokines (IL1β, IL2, IL6, and TNFα) were examined for a small number of patients undergoing chemotherapy for malignant bone and soft tissue tumors [65]. After intravenous infusion of antitumor drugs, increases of IL6 and TNFα production and decreases in neutrophil counts were evident. Similarly, when 13 cytokines were monitored after trans arterial chemoembolization (TACE) for patients with hepatocellular carcinoma, IL-6 reached a peak on day 3 before decreasing on and after day 7. In contrast, IL4, IL5, and IL10 increased at 2 months after TACE [66]. When the effect of CRT on IFN-γ, IL6, and TNFα was investigated for patients with locally advanced non-small-cell lung cancer (NSCLC) or glioblastoma multiforme (GBM), TNF-α levels in NSCLC and IFN-γ levels in GBM decreased [67].
The role of IL18 in response to gamma radiation-induced injury was reviewed [68]. The same authors also reported increases of IL1β, IL18, and IL33 expression in various organs, after exposure of mice, minipigs, and nonhuman primates to gamma radiation; they found IL18 as a potential biomarker to check radiation injury [69].
Elgstrom et al. [70] assessed the cytokine profile in the tumor microenvironment during tumor regression induced by radioimmunotherapy using an antibody conjugated to the beta emitter 177Lu. Although most cytokines in the tumor microenvironment were associated with immune cell infiltration, the differences in cytokine levels between treated and controlled tumors were small. The authors concluded that, for 177Lu radioimmunotherapy, tumor regression was not accompanied by a significant increase in cytokines that attract immune cells. In contrast, ablative local radiotherapy-mediated tumor control involved the production of IFN-β, and the antitumor effect of radiotherapy was diminished in mice deficient in type I IFN response [71]. The authors concluded that local high-dose radiotherapy initiated innate and adaptive immune attack cascades on the tumor through the type I IFN pathway.
6. Role of cytokines post chemo- or radio-therapy
In tumors, the cytokine response to chemotherapy or radiation serves several functions. First, pro-inflammatory cytokines such as TNF-α, IL1, and IL6 promote tumor cell death and inhibit tumor growth [72]. In tumor cells, these cytokines induce apoptosis, or programmed cell death, leading to a decrease in tumor mass. Second, pro-inflammatory cytokines attract immune cells to the tumor site and enhance their anti-tumor activity [73]. Third, cytokines modulate the immune response by promoting the differentiation and activation of immune cells [74]. Fourth, cytokines promote angiogenesis by stimulating the production of growth factors that induce the formation of new blood vessels in the tumor microenvironment [75]. Taken together, these cytokines can have pro-inflammatory effects, leading to tumor cell death, inhibition of tumor growth, attraction of immune cells to the tumor site, enhancement.
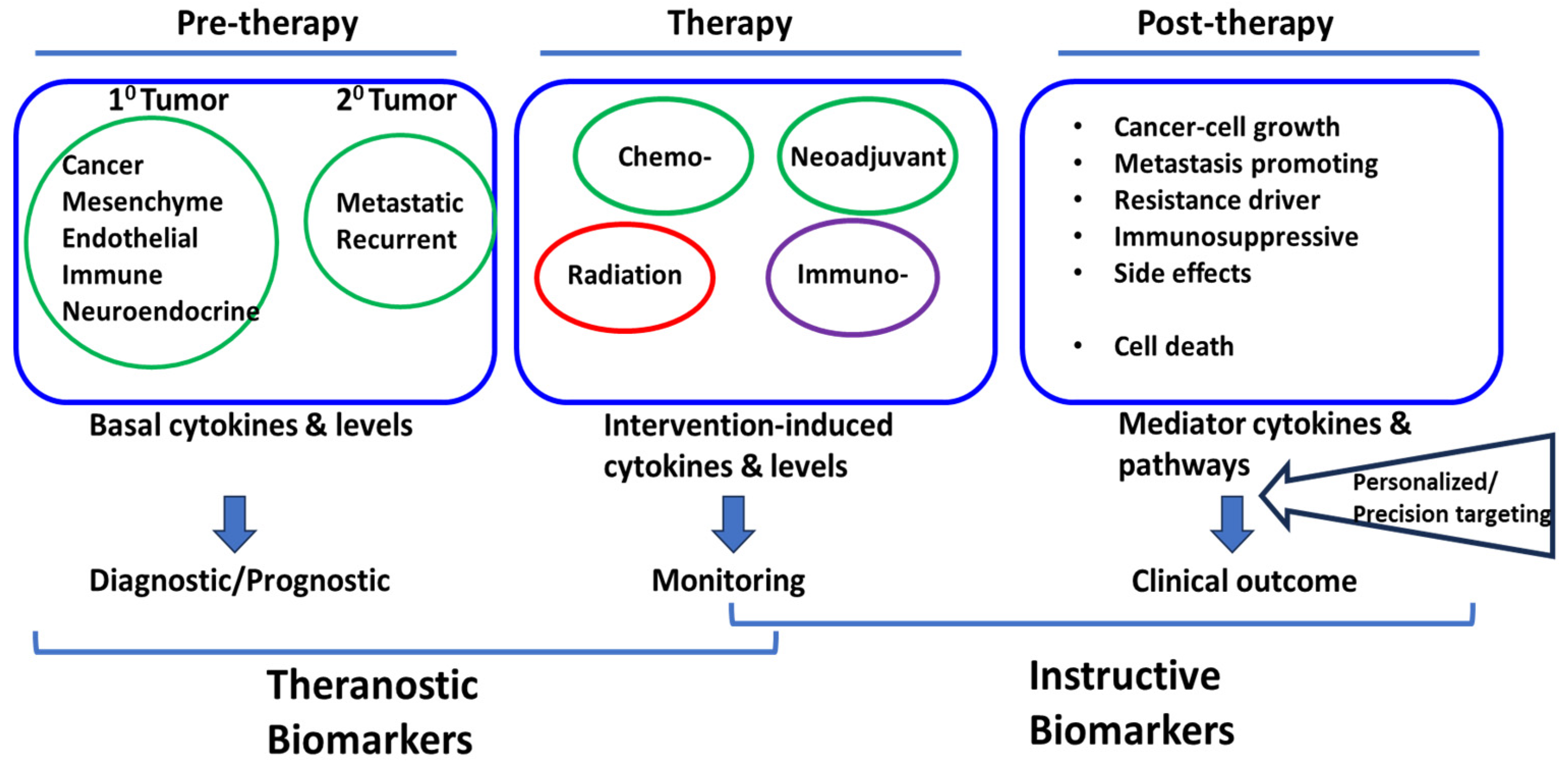
The reports briefly presented above point to the complex cytokine responses induced by chemotherapy and radiotherapy of diverse types of cancer. Elevated levels of specific cytokines may be associated with poor or favorable clinical outcomes; others may serve as potential theranostic biomarkers for treatment responses (Figure 2). Understanding the interplay between cytokines and receptors, cells of origin, and cancer therapy could lead to the development of new therapeutic approaches and personalized treatment strategies to improve patient outcomes. Further research in this field will uncover additional cytokine-related mechanisms and targets to enhance the efficacy of cancer treatment.
7. Challenges in cytokine-directed precision approach
Unlike a pharmacogenomics approach to precision medicine, in which primarily genomic information is used to establish the choice of patients, cytokine response to any cancer drug may not be directly predictable from the patient genomic sequence. However, in a precision medicine approach, a specific cytokine response or pathway may be selectively or non-selectively triggered, enhanced, or suppressed, independently of the gene or transcript. Although cytokines or similarly functioning molecules induced by chemo- or radiotherapy may be considered for precision or personalized medicine, challenges inherent to cytokine biology and functions remain. Most cytokines are produced in small amounts, most act locally, and some of their functions may be transient [62]. Moreover, the ultimate biological outcomes of cytokine response may depend on the net additive or competitive functions of factors within the solid-tumor microenvironment space and time. Since the protocols and sensitivities of the methods to measure cytokines vary, across-the-board standardization or interpretation of results will pose challenges. Further, cytokines measured after therapy need to be calibrated against the downstream pathways activated by the trigger (therapy), to show their functional relevance as actionable biomarkers. Due to the overlaps and redundancies in cytokine-receptor interactions, as well as to conditional tumor promoting or suppressive dual roles of some cytokines, challenges will remain in predicting which outcome would dominate after therapeutic stress is induced in tumors. With these in mind, studies on cytokines as ‘instructive biomarkers’ to enhance cancer therapy need to be designed. Fortunately, advances in multi-omics and sensitive analytical methods could enable us to overcome such challenges.
Conclusion
With the advent of muti-omics platforms and technologies, the use of genomics-based cancer precision medicine can now be complemented with cytokine- or related protein-based clinical decision strategies to find which patients could benefit from a secondary intervention after initiation of cancer therapy. This approach could open new avenues for the treatment of solid tumors for which options are limited, or for which low response rates or recurrences after chemotherapy or radiation hamper success. Maps of cytokine profiles in an individual’s tumor specimen or liquid biopsy, with serial comparisons pre- and post-therapy, could guide rational and targeted intervention, which is the central tenet of precision medicine.
Author Contributions
Conceptualization TS, MH, DG; draft preparation, DG, MH, TS; review, editing and TS, DB, PD, UM.
Funding
Research in TS, DB, PD, and UM labs is supported by NIH grant #U54CA118623.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
The authors thank Dr Donald Hill for editorial assistance.
Conflicts of Interest
Nothing to declare.
References
- Mathur, S.; Sutton, J. Personalized medicine could transform healthcare. Biomed Rep 2017, 7, 3–5. [Google Scholar] [CrossRef]
- Sadee, W.; Dai, Z. Pharmacogenetics/genomics and personalized medicine. Hum Mol Genet 2005, 14, R207–R214. [Google Scholar] [CrossRef]
- Agúndez, J.A.G.; García-Martín, E. Editorial: Insights in Pharmacogenetics and Pharmacogenomics: 2021. Front Pharmacol 2022, 13, 907131. [Google Scholar] [CrossRef] [PubMed]
- Wilke, R.A.; Reif, D.M.; Moore, J.H. Combinatorial pharmacogenetics. Nat Rev Drug Discov 2005, 4, 911–918. [Google Scholar] [CrossRef]
- Shastry, B.S. Pharmacogenetics and the concept of individualized medicine. Pharmacogenomics J 2006, 6, 16–21. [Google Scholar] [CrossRef]
- Le Tourneau, C.; et al. Molecularly targeted therapy based on tumour molecular profiling versus conventional therapy for advanced cancer (SHIVA): a multicentre, open-label, proof-of-concept, randomised, controlled phase 2 trial. Lancet Oncol 2015, 16, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.L. Giving Up on Precision Oncology? Not So Fast! Clin Transl Sci 2017, 10, 128–129. [Google Scholar] [CrossRef]
- Ghoreschi, K.; et al. Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet 2021, 397, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Noack, M.; Miossec, P. Selected cytokine pathways in rheumatoid arthritis. Semin Immunopathol 2017, 39, 365–383. [Google Scholar] [CrossRef]
- Aguero, R.; et al. Interleukin (IL)-17 Versus IL-23 Inhibitors: Which Is Better to Treat Patients With Moderate-to-Severe Psoriasis and Mild Psoriatic Arthritis in Dermatology Clinics? J Rheumatol 2023. [CrossRef]
- Moyaert, H.; et al. A blinded, randomized clinical trial evaluating the efficacy and safety of lokivetmab compared to ciclosporin in client-owned dogs with atopic dermatitis. Vet Dermatol 2017, 28, 593–e145. [Google Scholar] [CrossRef] [PubMed]
- Maini, R.N.; et al. Double-blind randomized controlled clinical trial of the interleukin-6 receptor antagonist, tocilizumab, in European patients with rheumatoid arthritis who had an incomplete response to methotrexate. Arthritis Rheum 2006, 54, 2817–2829. [Google Scholar] [CrossRef] [PubMed]
- Cardona-Pascual, I.; et al. Effect of tocilizumab versus standard of care in adults hospitalized with moderate-severe COVID-19 pneumonia. Med Clin (Engl Ed) 2022, 158, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Antwi-Amoabeng, D.; et al. Clinical outcomes in COVID-19 patients treated with tocilizumab: An individual patient data systematic review. J Med Virol 2020, 92, 2516–2522. [Google Scholar] [CrossRef] [PubMed]
- WHO. A living WHO guideline on drugs to prevent covid-19. in British Medical Journal 2021, n526. [Google Scholar] [CrossRef] [PubMed]
- West, W.H. Continuous infusion recombinant interleukin-2 (rIL-2) in adoptive cellular therapy of renal carcinoma and other malignancies. Cancer Treat Rev 1989, 16 (Suppl A), 83–89. [Google Scholar] [CrossRef] [PubMed]
- Berraondo, P.; et al. Cytokines in clinical cancer immunotherapy. Br J Cancer 2019, 120, 6–15. [Google Scholar] [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat Rev Immunol 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Sayana, S.; Khanlou, H. Maraviroc: a new CCR5 antagonist. Expert Rev Anti Infect Ther 2009, 7, 9–19. [Google Scholar] [CrossRef]
- Ndegwa, S. Maraviroc (Celsentri) for multidrug-resistant human immunodeficiency virus (HIV)-1. Issues Emerg Health Technol 2007, 1–8. [Google Scholar]
- Rallis, K.S.; et al. Cytokine-based Cancer Immunotherapy: Challenges and Opportunities for IL-10. Anticancer Res 2021, 41, 3247–3252. [Google Scholar] [CrossRef] [PubMed]
- Roufas, C. The Expression and Prognostic Impact of Immune Cytolytic Activity-Related Markers in Human Malignancies: A Comprehensive Meta-analysis. Front Oncol 2018, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; et al. Targeting macrophages: a novel treatment strategy in solid tumors. J Transl Med 2022, 20, 586. [Google Scholar] [CrossRef]
- Choudhry, H.; et al. Prospects of IL-2 in Cancer Immunotherapy. Biomed Res Int 2018, 2018, 9056173. [Google Scholar] [CrossRef] [PubMed]
- Yui, M.A.; et al. Preferential activation of an IL-2 regulatory sequence transgene in TCR gamma delta and NKT cells: subset-specific differences in IL-2 regulation. J Immunol 2004, 172, 4691–4699. [Google Scholar] [CrossRef] [PubMed]
- Paliard, X.; et al. Simultaneous production of IL-2, IL-4, and IFN-gamma by activated human CD4+ and CD8+ T cell clones. J Immunol 1988, 141, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.Y.; et al. Mast cell interleukin-2 production contributes to suppression of chronic allergic dermatitis. Immunity 2011, 35, 562–571. [Google Scholar] [CrossRef]
- Granucci, F.; et al. Inducible IL-2 production by dendritic cells revealed by global gene expression analysis. Nat Immunol 2001, 2, 882–888. [Google Scholar] [CrossRef]
- Hinrichs, C.S.; et al. IL-2 and IL-21 confer opposing differentiation programs to CD8+ T cells for adoptive immunotherapy. Blood 2008, 111, 5326–5333. [Google Scholar] [CrossRef]
- Hromadnikova, I.; et al. Influence of In Vitro IL-2 or IL-15 Alone or in Combination with Hsp 70 Derived 14-Mer Peptide (TKD) on the Expression of NK Cell Activatory and Inhibitory Receptors on Peripheral Blood T Cells, B Cells and NKT Cells. PLoS One 2016, 11, e0151535. [Google Scholar] [CrossRef]
- Thommen, D.S.; et al. A transcriptionally and functionally distinct PD-1(+) CD8(+) T cell pool with predictive potential in non-small-cell lung cancer treated with PD-1 blockade. Nat Med 2018, 24, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; et al. Identification of a major co-receptor for primary isolates of HIV-1. Nature 1996, 381, 661–666. [Google Scholar] [CrossRef]
- Simmons, G.; et al. Primary, syncytium-inducing human immunodeficiency virus type 1 isolates are dual-tropic and most can use either Lestr or CCR5 as coreceptors for virus entry. J Virol 1996, 70, 8355–8360. [Google Scholar] [CrossRef]
- Wu, L.; et al. CD4-induced interaction of primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5. Nature 1996, 384, 179–183. [Google Scholar] [CrossRef]
- Tsimberidou, A.M.; et al. Review of precision cancer medicine: Evolution of the treatment paradigm. Cancer Treat Rev 2020, 86, 102019. [Google Scholar] [CrossRef] [PubMed]
- Riedesser, J.E.; Ebert, M.; Betge, J. Precision medicine for metastatic colorectal cancer in clinical practice. Ther Adv Med Oncol 2022, 14, 17588359211072703. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K. Optimizing immunotherapy for colorectal cancer. Nat Rev Gastroenterol Hepatol 2022, 19, 93–94. [Google Scholar] [CrossRef]
- Zhao, C.; et al. Identification of immune checkpoint and cytokine signatures associated with the response to immune checkpoint blockade in gastrointestinal cancers. Cancer Immunol Immunother 2021, 70, 2669–2679. [Google Scholar] [CrossRef]
- Botticelli, A.; et al. The role of immune profile in predicting outcomes in cancer patients treated with immunotherapy. Front Immunol 2022, 13, 974087. [Google Scholar] [CrossRef]
- Lim, J.U.; Yoon, H.K. Potential predictive value of change in inflammatory cytokines levels subsequent to initiation of immune checkpoint inhibitor in patients with advanced non-small cell lung cancer. Cytokine 2021, 138, 155363. [Google Scholar] [CrossRef]
- Achyut, B.R.; Yang, L. Transforming growth factor-beta in the gastrointestinal and hepatic tumor microenvironment. Gastroenterology 2011, 141, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Desoteux, M.; et al. Transcriptomic evidence for tumor-specific beneficial or adverse effects of TGFbeta pathway inhibition on the prognosis of patients with liver cancer. FEBS Open Bio 2023, 13, 1278–1290. [Google Scholar] [CrossRef] [PubMed]
- Dranoff, G. Cytokines in cancer pathogenesis and cancer therapy. Nat Rev Cancer 2004, 4, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Demaria, M.; et al. Cellular Senescence Promotes Adverse Effects of Chemotherapy and Cancer Relapse. Cancer Discov 2017, 7, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Guillon, J.; et al. Chemotherapy-induced senescence, an adaptive mechanism driving resistance and tumor heterogeneity. Cell Cycle 2019, 18, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Sriram, G.; et al. The injury response to DNA damage in live tumor cells promotes antitumor immunity. Sci Signal 2021, 14, eabc4764. [Google Scholar] [CrossRef]
- Lotti, F.; et al. Chemotherapy activates cancer-associated fibroblasts to maintain colorectal cancer-initiating cells by IL-17A. J Exp Med 2013, 210, 2851–2872. [Google Scholar] [CrossRef] [PubMed]
- Reers, S.; et al. Cytokine changes in response to radio-/chemotherapeutic treatment in head and neck cancer. Anticancer Res 2013, 33, 2481–2489. [Google Scholar]
- Edwardson, D.W.; Parissenti, A.M.; Kovala, A.T. Chemotherapy and Inflammatory Cytokine Signalling in Cancer Cells and the Tumour Microenvironment. Adv Exp Med Biol 2019, 1152, 173–215. [Google Scholar] [CrossRef] [PubMed]
- van der Sijde, F.; et al. Serum cytokine levels are associated with tumor progression during FOLFIRINOX chemotherapy and overall survival in pancreatic cancer patients. Front Immunol 2022, 13, 898498. [Google Scholar] [CrossRef]
- Gu, C.; Xiong, X.; Liu, W. Prognostic Significance of the CXCLs and Its Impact on the Immune Microenvironment in Ovarian Cancer. Dis Markers 2023, 2023, 5223657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fu, Y. Comprehensive Analysis and Identification of an Immune-Related Gene Signature with Prognostic Value for Prostate Cancer. Int J Gen Med 2021, 14, 2931–2942. [Google Scholar] [CrossRef] [PubMed]
- Hincu, M.A.; et al. Relevance of Biomarkers Currently in Use or Research for Practical Diagnosis Approach of Neonatal Early-Onset Sepsis. Children 2020, 7, 309. [Google Scholar] [CrossRef] [PubMed]
- Pernot, S.; Evrard, S.; Khatib, A.M. The Give-and-Take Interaction Between the Tumor Microenvironment and Immune Cells Regulating Tumor Progression and Repression. Front Immunol 2022, 13, 850856. [Google Scholar] [CrossRef] [PubMed]
- Groysman, L.; et al. Chemotherapy-induced cytokines and prognostic gene signatures vary across breast and colorectal cancer. Am J Cancer Res 2021, 11, 6086–6106. [Google Scholar]
- Carlsen, L.; et al. Pan-drug and drug-specific mechanisms of 5-FU, irinotecan (CPT-11), oxaliplatin, and cisplatin identified by comparison of transcriptomic and cytokine responses of colorectal cancer cells. Oncotarget 2021, 12, 2006–2021. [Google Scholar] [CrossRef] [PubMed]
- Bedi, D.; et al. Camptothecin Induces PD-L1 and Immunomodulatory Cytokines in Colon Cancer Cells. Medicines 2019, 6, 51. [Google Scholar] [CrossRef]
- Bravata, V.; et al. Cytokine profile of breast cell lines after different radiation doses. Int J Radiat Biol 2017, 93, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; et al. Colorectal cancer cell-derived CCL20 recruits regulatory T cells to promote chemoresistance via FOXO1/CEBPB/NF-kappaB signaling. J Immunother Cancer 2019, 7, 215. [Google Scholar] [CrossRef]
- Stojanovska, V.; et al. Oxaliplatin Treatment Alters Systemic Immune Responses. Biomed Res Int 2019, 2019, 4650695. [Google Scholar] [CrossRef]
- Sistigu, A.; et al. Cancer cell-autonomous contribution of type I interferon signaling to the efficacy of chemotherapy. Nat Med 2014, 20, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Schaue, D.; Kachikwu, E.L.; McBride, W.H. Cytokines in radiobiological responses: a review. Radiat Res 2012, 178, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Palata, O.; et al. Radiotherapy in Combination With Cytokine Treatment. Front Oncol 2019, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Tada, N.; et al. Changes in the plasma levels of cytokines/chemokines for predicting the response to chemoradiation therapy in rectal cancer patients. Oncol Rep 2014, 31, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Sato, M. [Changes in serum cytokine levels in patients with malignant bone and soft tissue tumors in the course of chemotherapy]. Gan To Kagaku Ryoho 1992, 19 (Suppl 10), 1449–1452. [Google Scholar] [PubMed]
- Kim, M.J.; et al. Change in inflammatory cytokine profiles after transarterial chemotherapy in patients with hepatocellular carcinoma. Cytokine 2013, 64, 516–522. [Google Scholar] [CrossRef]
- Deniz, C.D.; Gurbilek, M.; Koc, M. Prognostic value of interferon-gamma, interleukin-6, and tumor necrosis factor-alpha in the radiation response of patients diagnosed with locally advanced non-small-cell lung cancer and glioblastoma multiforme. Turk J Med Sci 2018, 48, 117–123. [Google Scholar] [CrossRef]
- Xiao, M. The Role of Proinflammatory Cytokine Interleukin-18 in Radiation Injury. Health Phys 2016, 111, 212–217. [Google Scholar] [CrossRef]
- Ha, C.T.; et al. Circulating interleukin-18 as a biomarker of total-body radiation exposure in mice, minipigs, and nonhuman primates (NHP). PLoS One 2014, 9, e109249. [Google Scholar] [CrossRef]
- Elgstrom, E.; Ohlsson, T.G.; Eriksson, S.E. Cytokine evaluation in untreated and radioimmunotherapy-treated tumors in an immunocompetent rat model. Tumour Biol 2017, 39, 1010428317697550. [Google Scholar] [CrossRef]
- Burnette, B.C.; et al. The efficacy of radiotherapy relies upon induction of type i interferon-dependent innate and adaptive immunity. Cancer Res 2011, 71, 2488–2496. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.H.; et al. The Inflammatory Cytokine Profile of Patients with Malignant Pleural Effusion Treated with Pleurodesis. J Clin Med 2020, 9, 4010. [Google Scholar] [CrossRef] [PubMed]
- Chiamulera, M.M.A.; et al. Salivary cytokines as biomarkers of oral cancer: a systematic review and meta-analysis. BMC Cancer 2021, 21, 205. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; et al. Nanomedicine therapies modulating Macrophage Dysfunction: a potential strategy to attenuate Cytokine Storms in severe infections. Theranostics 2020, 10, 9591–9600. [Google Scholar] [CrossRef]
- Di Caro, G.; et al. Circulating Inflammatory Mediators as Potential Prognostic Markers of Human Colorectal Cancer. PLoS One 2016, 11, e0148186. [Google Scholar] [CrossRef]
Figure 1.
Current precision targets in cancer medicine as a subset of the broader elements of precision medicine.
Figure 1.
Current precision targets in cancer medicine as a subset of the broader elements of precision medicine.
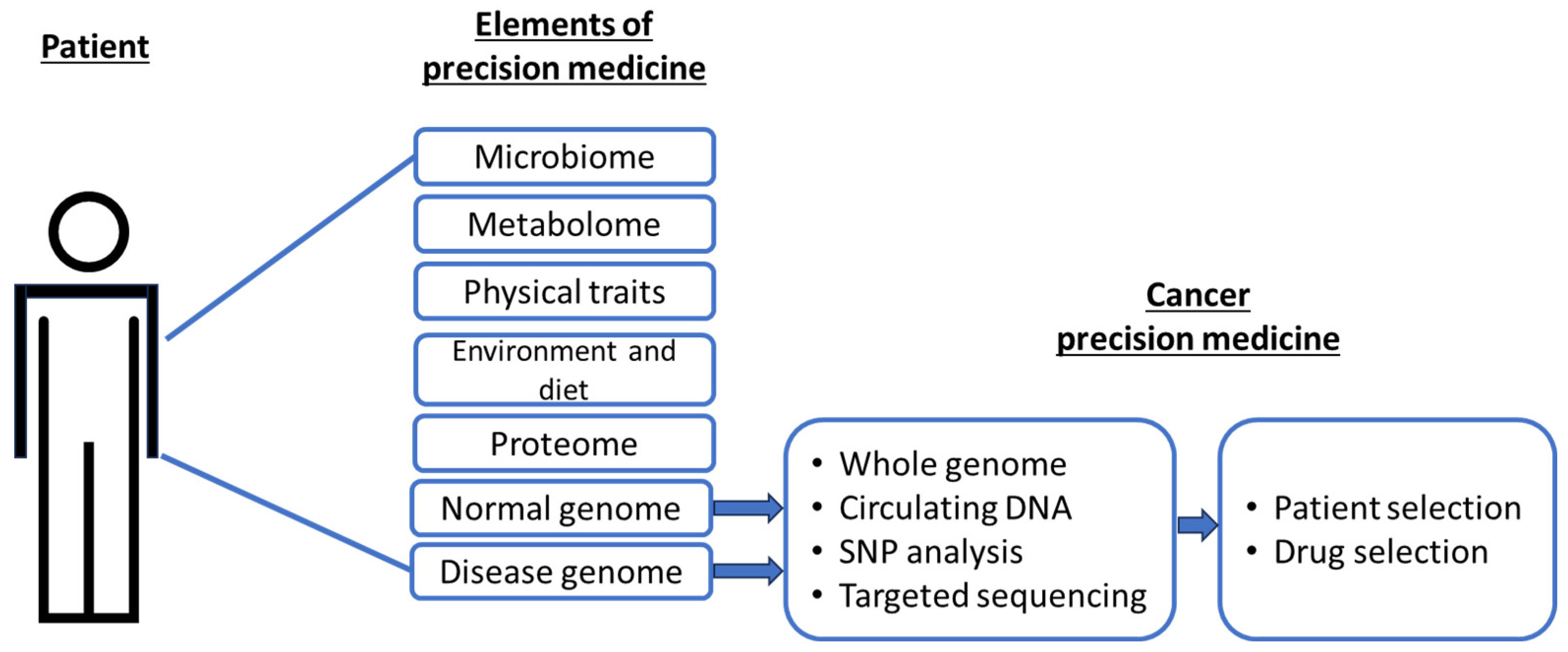
Figure 2.
Post-therapy Instructive Biomarkers as potential targets for personalized precision approach.
Figure 2.
Post-therapy Instructive Biomarkers as potential targets for personalized precision approach.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



