
Submitted:
22 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Fusarium spp. are commonly associated with the root rot complex of soybean (Glycine max). Previous surveys identified six common Fusarium species from Manitoba, including F. oxysporum, F. redolens, F. graminearum, F. solani, F. avenaceum and F. acuminatum. This study aimed to determine their pathogenicity, assess host resistance and evaluate the genetic diversity of Fusarium spp. isolated from Canada. Pathogenicity was tested on two cultivars, ‘Akras’ (moderate resistance) and ‘B150Y1’ (susceptible), under greenhouse conditions. The virulence of the fungal isolates varied, with root rot severities ranging from 1.5 to 3.3 on a 0-4 scale. The six species were used to screen a panel of 20 Canadian soybean cultivars for resistance in a greenhouse. Cluster and principal component analyses were conducted based on the same traits as for the pathogenicity study. Two of the cultivars, ‘P15T46R2’ and ‘B150Y1’, were consistently found to be tolerant to F. oxysporum, F. redolens, F. graminearum and F. solani. To investigate the incidence and prevalence of Fusarium spp. in Canada, fungi were isolated from 106 soybean fields surveyed across Manitoba, Saskatchewan, Ontario and Quebec. Two-hundred twenty-one Fusarium isolates were identified, with phylogenetic analyses indicating diversity of the isolates in the major soybean production regions of Canada.
Keywords:
Fusarium spp.
; genetic diversity
; pathogenicity
; phylogenetic analyses
; resistance
; root rot
; Sanger sequencing
; soybean
1. Introduction
Soybean (Glycine max L.) is one of the most important legume crops worldwide, known as one of the few ‘complete protein’ sources with all nine human-essential amino acids. Cultivation is mainly centered in North and South America, comprising 80% of total world production [1,2,3]. In Canada, soybean production has increased over the past decades, and it has become a crucial crop for human and livestock consumption as well as global exportation. Starting in Ontario and expanding to the Prairie provinces, soybean ranks behind only wheat, canola and barley in acreage [4,5]. In 2022, 1.6 million hectares of soybeans were grown in eastern Canada (mainly in Ontario and Quebec), yielding 5.1 million tonnes, with another 0.5 million hectares (1.4 million tonnes) grown in western Canada (primarily Manitoba and Saskatchewan). The value of the crop in Canada was estimated at more than $33 billion USD [5,6].
In spite of this rapid expansion, several biotic constraints continue to be a major limitation to soybean production. To date, over 300 diseases have been reported on soybean, causing yield losses averaging 11% [7]. A soybean survey conducted in the USA and Ontario found 23 specific diseases from 2015 to 2019, resulting in yield losses of 6-11% [8]. Addressing these biotic constraints is essential for enhancing soybean production, ensuring food security and supporting farmers worldwide.
Fusarium spp. are significant soybean pathogens, causing root rot, wilt, sudden death syndrome (SDS), seed decay and seedling blight [9,10,11]. In some cases, soybean diseases are caused by a single Fusarium species, for instance, Fusarium wilt caused by F. oxysporum Schltdl [12,13,14]. Others involve multiple species, constituting a pathogen ‘complex’. The soybean root rot complex, widespread in North America in recent years, results from infection by multiple Fusarium spp. [9,10,15,16,17]. These species include F. solani, F. oxysporum, F. acuminatum, F. avenaceum, F. cerealis, F. culmorum, F. equiseti, F. graminearum, F. poae, F. proliferatum, F. pseudograminearum, F. redolens, F. sporotrichioides, F. fujikuroi, F. incarnatum-equiseti, F. tricinctum, F. semitectum, F. armeniacum, F. commune and F. verticillioides [9,10,15,16,18,19,20,21,22,23]. While F. solani and F. oxysporum were initially considered as the dominant species in the soybean root rot complex, the composition of Fusarium spp. and their prevalence varied across different geographic regions [9,24,25]. Zhao et al. [19] reported that F. proliferatum is the most virulent species on soybean in Hubei, China. In contrast, in Sichuan province, the most aggressive species were F. oxysporum, F. equiseti and F. graminearum [26]. At the reproductive stage of the crop, F. acuminatum, F. graminearum, and F. solani were more prevalent than other Fusarium spp. in Iowa [10]. Zhang et al. [27] reported eight Fusarium spp. were associated with soybean root rot in Ontario, with the most aggressive species identified as F. graminearum. In western Canada, F. oxysporum and F. acuminatum were the dominant species in surveys of soybean in Manitoba and Alberta, respectively [9,14].
Generally, Fusarium spp. are classified based on various morphological characters, including colony features and pigmentation on different media, as well as the appearance, size and presence of the three spore types: microconidia, macroconidia, and chlamydospores [28]. However, the morphology, ecology, physiology and even genetic traits of Fusarium spp. often exhibit variations across different studies [14,28,29,30]. Currently, the most common and effective approach for identifying Fusarium spp. in soybean relies on DNA Sanger sequencing of PCR amplification products obtained with primers for specific genes or genomic regions. This method offers rapid, precise, sensitive, and convenient results [9,31]. The internal transcribed spacer (ITS) region of nuclear-encoded ribosomal DNA (ITS rDNA) is commonly used to generate primers to identify plant pathogens [32,33]. Both the ITS1/ITS4 and ITS5/ITS4 primer sets are popular for the identification of Fusarium spp. in various crops [14,19,34,35,36]. Additionally, numerous other primers targeting genes within the Fusarium genus have been widely applied in DNA sequence analysis. These alternate targets include the translation elongation factor 1 alpha (TEF1a) genes, mating type locus genes, phosphate permease gene, beta-tubulin gene, as well as the largest and second largest subunit nucleotide sequences of RNA polymerase II, RPB1 and RPB2, respectively [9,37,38]. Furthermore, the most up to date Fusarium database, FUSARIUM-ID v.3.0, primarily identifies Fusarium spp. based on TEF1a sequences, complemented with some other loci, including RPB1, RPB2 and ITS [39]. At present, molecular methods with multiple primers, combined with evaluation of morphological characters, are necessary to differentiate Fusarium spp. and analyze their phylogenetic relationships [19,26,40,41,42].
Fusarium spp. have a wide host range, infecting various crops such as cereals, soybean, other legume crops, canola and corn [43,44,45,46]. Notably, F. graminearum, known for causing severe Fusarium head blight (FHB) in cereal crops, also exhibits high aggressiveness towards soybean [9,47,48]. Cross-pathogenicity among different crops can limit the efficacy of crop rotation in controlling Fusarium diseases [9,44]. As such, seed treatments are widely applied in North America to enhance seedling emergence and provide protection against soilborne pathogens of soybean [49,50,51]. Studies of fungicides and biocontrol agents as soybean seed treatments have also reported potential efficacy against single species of Fusarium under controlled conditions. The effectiveness of these treatments, however, remains limited in field trials conducted in natural environments [25,52,53].
The most promising strategy to manage soybean diseases caused by Fusarium spp. lies in the selection and breeding of resistant cultivars. However, globally available commercial cultivars with complete resistance are yet to be developed [27,54,55]. Incomplete resistance, controlled by polygenetic loci [56,57,58,59], is influenced by the interaction between the cultivar and the environment [60,61,62]. Cultivar screening and detection of resistance to SDS caused by F. virguliforme in soybean has been reported in the United States [58]. Similarly, tolerance or incomplete resistance to virulent isolates of Fusarium also has been identified in Alberta and Ontario [27,52,63]. However, resistance to Fusarium spp. has not been evaluated in Manitoba or other regions of Canada.
To improve management of Fusarium diseases in Canadian soybean cultivation, it is essential to understand the species diversity, distribution, and pathogenicity of Fusarium spp., as well as to assess soybean cultivar resistance in the major production areas. This study aims has three primary objectives: 1) to evaluate the pathogenicity of six common Fusarium spp. on soybean under controlled conditions; 2) to assess resistance/tolerance to these six common Fusarium spp. in a selection of 20 soybean cultivars; and 3) to investigate the genetic diversity and distribution of Fusarium spp. across the major soybean production regions of Canada.
2. Materials and Methods
2.1. Fungal Material and Inoculum Preparation
Twelve isolates representing the six most common Fusarium species (two isolates per species) in Manitoba as reported by Kim et al. [64], consisting of F. oxysporum, F. redolens, F. graminearum, F. solani, F. avenaceum and F. acuminatum, were obtained from the culture collection of the Agriculture and Agri-Food Canada, Brandon Research and Development Centre, Brandon, Manitoba (Table 1). Grain inoculum of each isolate was prepared according to a procedure modified from Chang et al. [15]. Briefly, each of the 12 isolates was cultured on Difco™ potato-dextrose agar (PDA) (Becton, Dickinson and Company, Sparks, MD USA) for one week at 25 °C in the dark. Approximately 10 L of wheat grain was soaked in distilled water overnight, and 600 mL of the soaked grain was transferred to an autoclave bag and autoclaved at 121°C for 90 min. After cooling, the grain was inoculated with 10 pieces of 5-day-old mycelial plugs excised from the PDA colonies of each fungus, and incubated at 25 °C in the dark for 2 weeks. The infested wheat grain was dried and ground into a powder (particle sizes between 0.25 and 1 mm) for use as inoculum.
2.2. Pathogenicity Test
Two soybean cultivars, ‘Akras’ and ‘B150Y1′ were selected to evaluate the pathogenicity of the 12 fungal isolates. Plastic cups (473 mL volume) were filled with soil mix (Sun Gro Horticulture Canada Ltd., Seba Beach, AB), and a layer of grain inoculum powder (10 mL) was applied to each cup and covered with 1 cm of soil mix. Seven seeds of each soybean cultivar were sown into each cup and covered with an additional 1 cm of soil mix. The cups were then placed in a greenhouse maintained at 25 °C with natural light supplemented by artificial lighting (light/dark cycle of 18 h/6 h). Non-inoculated controls of each soybean cultivar received wheat grain powder that had not been inoculated with any of the Fusarium spp. The pathogenicity test was arranged in a randomized complete block design (RCBD) with five replicates and repeated twice.
2.3. Evaluation of Resistance
The most virulent isolate of each Fusarium spp. was selected based on the results of the pathogenicity test and applied to screen a germplasm collection of 20 soybean commercial cultivars, including ‘HS11Ry07′, ‘S15B4′, ‘P15T46R2′, ‘B150Y1′, ‘Williams’, ‘Akras’, ‘TH32004R2Y’, ‘AC Proteus’, ‘AAC Edward’, ‘AAC Springfield’, ‘OAC Prudence’, ‘Misty’, ‘NSC Reston’, ‘OAC Ayton’, ‘Bloomfield’, ‘AC Harmony’, ‘OAC Petrel’, ‘Mandor’, ‘OT15-02′, and ‘NSC Dauphin’. Seeds were provided by the University of Alberta, and Agriculture and Agri-Food Canada (Harrow Research and Development Centre, Morden Research and Development Centre, and Brandon Research and Development Centre). The same seeding and inoculation methods were used as described above for the pathogenicity test. All 20 cultivars were inoculated with each of the six Fusarium spp. Non-inoculated controls, prepared as above, were also included for each cultivar. The experiment was arranged in a RCBD with five replicates and repeated twice.
2.4. Greenhouse Data Collection
Emergence counts were conducted on the 7th, 14th and 21st day after seeding. The plant height from stem to the top leaf was measured on the 14th day. Root rot severity was evaluated on a 0–4 scale as previously described by Chang et al. [15] and Zhou et al. [14] with minor modifications, where: 0, no symptoms; 1, taproot slightly discolored and fibrous roots healthy, aboveground growth appears normal; 2, taproot mostly discolored and/or fibrous roots with visible lesions, aboveground growth slightly reduced; 3, taproot completely discolored and fibrous roots with obvious lesions, aboveground growth severely reduced; and 4, taproot dark, degraded, few and reddish-brown fibrous roots, plants dead or dying. The 0-4 disease scale was applied to rate symptoms caused by each of the six Fusarium spp. evaluated (Figure 1). The 21-day-old seedlings from each replicate were collected after disease rating; their shoots and roots were separated, dried at 35 °C for 48 h, and weighed.
2.5. Fungal Isolation from Field Samples
Soybean root samples were collected from 106 fields in Manitoba (55 fields), Saskatchewan (18 fields), Ontario (30 fields) and Quebec (3 fields) in the 2022 growing season. Fifteen roots with symptoms of root rot were selected from each field for pathogen isolation. Two symptomatic root pieces (3-5 mm long) were cut from the root tip and crown of each root, surface-sterilized in 1% NaOCl for 60 s, and then rinsed three times with sterilized water. The surface-sterilized root pieces were transferred to PDA containing 0.2 mg/mL streptomycin and incubated for 7 days at 24 °C. The cultures were examined and any colonies of Fusarium were transferred to water agar and incubated for 5 days, at which time a single hyphal tip was cut and transferred to PDA in Petri dishes for purification. The purified cultures were grouped and sub-grouped based on their morphological characteristics, including color, mycelium type, released pigment on both sides of the Petri dishes [28].
2.6. DNA Extraction, PCR Amplification and Sanger Sequencing
The purified cultures were grouped based on their morphology on PDA and one isolate was randomly selected from each group for molecular identification. A total of 336 isolates from 89 fields and six reference isolates of Fusarium spp. were used to extract genomic DNA using a modified CTAB method following O’Donnell et al. [65]. Approximately 100 mg of the mycelium was scraped from the surface of a 10-day-old colony grown on PDA and transferred to a 1.5 ml microcentrifuge tube. The samples were flash-frozen in liquid nitrogen and mechanically homogenized using a TissueLyser II (Qiagen, Hilden, Germany) at 25 Hz with 3 mm beads for 1 min. Seven-hundred µl of CTAB buffer (Teknova, Hollister, CA, United States) was added to each 1.5 mL microcentrifuge tube and incubated at 65°C for 30 min. A 500 µl aliquot of phenol:chloroform:isoamylalcohol (25:24:1) (Invitrogen, Waltham, MA, United States) was added to each extraction tube and centrifuged at 1,107 g (3,000 rpm) for 30 min. The aqueous phase was transferred to a new microcentrifuge tube and mixed with an equal volume of isopropanol, incubated at -20 °C for 1 h and then centrifuged for 30 min at 1,107 g (3,000 rpm). The supernatant was decanted and the DNA pellet was washed twice with 70% ethanol and allowed to dry overnight at room temperature. The DNA pellet was dissolved in 40 µl TE buffer (10 mM Tris-HCl, 1 mM EDTA), the DNA concentration was estimated using a Nanodrop ND2000 (Thermo Fisher Scientific, Wilmington, USA), and diluted to a final concentration of 50 ng DNA/µl in distilled water.
The primer sets ITS4/5 (5′GGAAGTAAAAGTCGTAACAAGG 3′/5′TCCTCCGCTTATTGATATGC 3′; ITS region)[33] and T12 (5′AACATGCGTGAGATTGTAAGT 3′/5′TAGTGACCCTTGGCCCAGTTG 3′; beta-tubulin gene) [35,65] were used for PCR amplification and sequencing of each DNA sample. Amplifications were conducted in a 50 µl reaction volume, which included 1 ng/μl DNA template (5 µl), 10× Go Taq buffer (10 µl), MgCl2 buffer (5 µl), sterile distilled water (26.75 µl), 2 mM dNTPs (1 µl), 1 μl each of forward and reverse primers (2.5 μM), and 0.25 μl of DNA Taq polymerase (Titanium Taq, Promega, Madison, USA). The PCR conditions were set as follows: initial denaturation at 95°C for 120 s; followed by 40 cycles at 95°C for 45 s, annealing for 45 s, and extension at 72°C for 1 min; and a final extension at 72°C for 10 min followed by cooling at 4 °C until recovery of the samples. The annealing temperatures were 55 °C and 52 °C for the ITS4/5 and T12 beta-tubulin genes, respectively. A 2 µl aliquot from each PCR was examined for presence/absence of amplicons by electrophoresis in 2% agarose. The amplicons in the remaining 48 µl volume were purified using a QIAquick PCR Purification Kit (Qiagen, Hilden, Germany) and sent for Sanger sequencing at the Molecular Biology Services Unit (MBSU) at the University of Alberta in Edmonton, AB. The sequences obtained were used in BLAST searches of the National Center for Biotechnology Information (NCBI) databases (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The BLAST comparison was conducted in Standard databases (nr etc.) with organism of Nucleotide collection (nr/nt) using highly similar sequences (megablast). The BLAST results were sorted by percent identity of which the first result was used to identify the species of DNA sample.
2.7. Phylogenetic Analysis
All 336 sequences from the isolates obtained from the soybean root samples collected in 2022 and the reference sequences from the six Fusarium spp. were edited using Bio-Edit software v. 7.2.5 [66] with manual adjustment. The six reference sequences that best matched F. oxysporum, F. redolens, F. graminearum, F. solani, F. avenaceum and F. acuminatum were downloaded from GenBank. All of the sequences identified as Fusarium spp. and the six reference sequences were included in a phylogenetic analysis by the maximum likelihood (ML) method using the Jukes Cantor model with default parameters in the CLC main Workbench v.23.0.3 (QIAGEN, Aarhus, Denmark). Bootstrap values (BV) (%) were calculated with 1,000 replicates, and a phylogenetic tree was constructed with BV>70.
2.8. Data Analysis
Analysis of variance (ANOVA) was conducted for all traits analyzed in the pathogenicity test and cultivar resistance evaluation trials in greenhouse using R v. 4.2.0 [67]. The average mean of plants in each cup was calculated for the three seedling germination counts named “count1”, “count2”, “count3”, plant height (cm) named “height”, root rot severity on scale of 0-4 named “DS1”, as well as dry shoot and root weight (g) named “shoot” and “root”, respectively. An additional estimated mean for disease severity that considered the non-germinated seeds as dead plants with a score of 4 was calculated as “DS2”. A least significant difference (LSD) at p< 0.05 was applied to compare the estimated means of all the traits among different treatments using the R package “agricolae”. The reduction percentage was calculated for cluster analysis using R v. 4.2.0 [67]. To demonstrate the level of high tolerance, moderate tolerance, moderate susceptibility and high susceptibility, the cluster group number was set as four. A principal components analysis (PCA) was carried out estimating the reduction percentage of the three germination counts, height, shoot weight and root weight, as well as the increase of two types of disease severity (DS1 and DS2) in R using the package “ggbiplot”. For the Fusarium spp. isolation, the incidence of each species was calculated using the formula: Incidence % = (n/N) ×100, of which n is number of fields where the species was detected in each province and N is the total field number in each province.
3. Results
3.1. Pathogenicity Test
Symptoms caused by the six Fusarium spp. varied, encompassing rotting, girdling and the development of brown sunken lesions (Figure 1). Analysis of variance revealed significant interactions between Fusarium spp. and the soybean cultivars for all the traits, while the repeat effect of two greenhouse experiments was not significant (Table 1). Consequently, data from the two repeated experiments were combined for all traits. Mean and LSD estimates for the six Fusarium spp. were calculated using ‘Akras’ and ‘B150Y1’ for all traits. Fusarium avenaceum isolate 1 exhibited the most adverse effects, resulting in a three-fold reduction in germination counts on both ‘Akras’ and ‘B150Y1′ (Figure 2a).
All Fusarium spp. infections significantly reduced plant height except for F. oxysporum on both cultivars and F. redolens on ‘Akars’ (Figure 2b). For the parameter DS2 (root rot severity), all 12 isolates caused significant severity compared with the non-inoculated control for both ‘Akras’ and ‘B150Y1′, with scores ranging from 1.5 to 3.3 on the 0-4 scale (Figure 2c). The dry shoot weight of infected plants were significantly lower than the non-inoculated controls for both cultivars following inoculation with each of the fungal isolates. A similar trend was observed for dry root weight, although the reductions caused by F. solani isolate 1 on ‘B150Y1,’ F. solani isolate 2 on ‘Akaras,’ F. avenaceum isolate 2 on both cultivars, and both isolates of F. redolens on ‘Akaras’ were not statistically significant relative to the control (Figure 2d).
In the pathogenicity study, the virulence of each isolate of each Fusarium spp. was reflected consistently across all traits. The most virulent isolates of each species were selected based on higher disease severity for the cultivar resistance evaluation trials, including F. oxysporum isolate 2, F. redolens isolate 2, F. solani 1, F. graminearum isolate 2, F. acuminatum isolate 2 and F. avenaceum isolate 1.
3.2. Cultivar Resistance Evaluation
Both greenhouse experiments yielded corresponding results for all variables, with a non-significant repeat effect (p>0.05) (Table 2). Cultivar effect, Fusarium spp. effect, and their interaction were all found to be significant for all of the traits (p<0.05) (Table 2). As such, the reaction of each of the 20 soybean cultivars was evaluated in response to each of the six Fusarium spp. Notably, significant differences among the soybean cultivars were observed for all traits in the non-inoculated control, with ranges of 3.9 - 7.0 for the three germination counts (Count1, Count2, Count3), 8.6 cm - 16.5 cm for plant height, 0.9 g - 1.5 g for dry shoot weight, and 0.2g - 0.4 g for dry root weight. As expected, however, the non-inoculated controls of all 20 cultivars remained completely healthy, with a score of 0 for both DS1 and DS2 (Supplementary Table 1). Consequently, the disease reactions for the six Fusarium spp. were evaluated using percentage reduction for Count1, Count2, Count3, Height, Shoot, and Root, as well as disease severity increase.
Fusarium oxysporum caused the greatest average reduction in germination (80%), while the maximum average height reduction was observed for F. avenaceum (64%). For shoot and root loss, F. oxysporum had the most pronounced effects, with reductions of 81% and 83%, respectively. Regarding disease severity, inoculation with Fusarium oxysporum and F. avenaceum caused greatest severity scores for DS1 (2.8) and DS2 (3.4), respectively. In contrast, F. solani had the lowest impact on all of the traits compared with the other five Fusarium spp.
Principal components analysis with four clusters revealed a positive correlation among variables and cultivar reactions following inoculation with F. graminearum, F. avenaceum, F. acuminatum, F. oxysporum, F. redolens and F. solani (Figure 3). Notably, four cultivars, ‘B150Y1′, ‘P15T46R2′, ‘Misty’ and ‘Mandor’, clustered in the high-tolerance group against F. graminearum. Against F. avenaceum, the most tolerant cultivars were ‘OAC Prudence’, ‘Misty’, ‘NSC Reston’, ‘OAC Ayton’ and ‘AC Harmony’. The cultivars ‘S15B4′, ‘OT15-2′ and ‘Mandor’ displayed the greatest tolerance to F. acuminatum, and ‘P15T46R2′, ‘B150Y1′, ‘Williams’, ‘TH32004R2Y’, ‘AAC Edward’, ‘OAC Prudence’ and ‘OT15-02′ were most tolerant to F. oxysporum. Additionally, the most tolerant cultivars against F. redolens were ‘P15T46R2′, ‘B150Y1′, ‘TH32004R2Y’, ‘AC Proteus’ and ‘OAC Petrel’. The cultivars, ‘HS11Ry07′, ‘S15B4′, ‘P15T46R2′, ‘B150Y1′, ‘Williams’, ‘TH32004R2Y’ and ‘OAC Prudence’ showed the greatest tolerance to F. solani. Overall, the soybean cultivars ‘P15T46R2′ and ‘B150Y1′ displayed suppression against F. graminearum, F. oxysporum, F. redolens and F. solani.
3.3. Fusarium Spp. Identification
A total of 983 purified isolates were obtained from the symptomatic root samples and separated into three morphological groups and nine subgroups (Figure 4). After filtering and grouping based on colony morphology, 336 isolates were selected for molecular identification. The primer sets ITS4/5 (ITS region) and T12 (beta-tubulin gene) produced single bands of ~500 bp [36] and ~580 bp [35], respectively, from the six reference isolates (Figure 5) and 336 isolates collected in this study. Following removal of non-Fusarium species and isolates with poor sequence quality, 221 isolates were confirmed as Fusarium spp., primarily based on the ITS4/5-amplified gene sequences complemented with the T12-amplified sequences (Supplementary Table 2).
The distribution and prevalence of identified Fusarium spp. varied across Canada (Table 3). Based on the ITS sequences, the most prevalent species was F. avenaceum (26%), followed by F. oxysporum (19%), F. acuminatum (18%), F. redolens (9%) and F. equiseti (8%). All of the species identified in this study were grouped into four Fusarium spp. complexes except for the isolates classified as F. solani and F. redolens, which belonged to the F. tricinctum species complex (FTSC), F. oxysporum species complex (FOSC), F. incarnatum-equiseti species complex (FIESC), and F. sambucinum species complex (FSSC). The frequencies of FTSC, FOSC, FIESC and FSSC were 47%, 20%, 12% and 6% of all Fusarium spp. isolates recovered, respectively. Additionally, F. graminearum, and F. solani were exclusively isolated from samples collected in Saskatchewan and Ontario, respectively. Fusarium acuminatum and F. redolens appeared unique to the Manitoba and Saskatchewan. Notably, 91% of FIESC isolates were obtained from western Canada.
Moreover, 36% of the 221 isolates were identified as Fusarium spp. based on the beta-tubulin sequences. Among these, there were 56 isolates of F. acuminatum, four isolates of F. avenaceum, six isolates of F. equiseti, seven isolates of F. flagelliforme, two isolates of F. solani, one isolate each of F. compactum and F. sporotrichioides.
3.4. Phylogenetic Analysis
The ML phylogenetic analyses, based on the ITS4/5-amplified sequences, provided clear separation of the 221 isolates of Fusarium spp. into six distinct groups (Supplementary Figure 1). Group 1 consisted of 118 sequences belonging to FTSC [9,68,69], with 58, 39, 1, 2, 2 and 1 isolates identified as F. avenaceum, F. acuminatum, F. tricinctum, F. reticulatum, F. compactum and F. flocciferum. The remaining 14 sequences were unknown Fusarium spp. and reference sequences from the current study or GeneBank. Group 2 clustered three isolates of F. graminearum, two isolates of F. sporotrichioides, five isolates of F. cerealis and three isolates of F. culmorum into FSSC [9,70]. Group 3 comprised 17 isolates of F. equiseti, eight isolates of F. incarnatum and one of F. longipes, forming FIESC, with a strong bootstrap value (BV) of 91 [9,71]. Group 4 and Group 6 included 42 isolates of F. oxysporum, along with three F. verticillioides isolates and two F. oxysporum reference sequences, all known to be associated with FOSC [9,72]. Group 5 encompassed all three F. solani isolates and F. falciforme from Ontario, along with two reference sequences, with a BV of 100. Group 7 consisted of 19 isolates of F. redolens, one unknown fungal species, one reference isolate and one reference sequence from GeneBank, supported by a BV of 90. However, the grouping separation among FTSC, FSSC and FOSC-2 was not supported strongly (BV<70).
The sequences that were identified as species of FTSC and FIESC by amplification of the beta-tubulin gene were also analyzed for their phylogenetic relationships (Figure 6a and 6b, respectively). Clade 1 included four F. avenaceum isolates, one reference sequence and one reference isolate. Clade 2 comprised 56 isolates of F. acuminatum, one reference sequence and one reference isolate. For FIESC, Clades 3 and 4 consisted of seven isolates of F. flagelliforme and six isolates of F. equiseti, respectively. Additionally, the trees were rooted with one isolate of F. compactum (NSRR22_218) serving as an outgroup.
4. Discussion
Soybean production in western Canada, particularly Manitoba, has been increasing significantly in recent decades [4,5,6]. Root rot of soybean is a global concern involving numerous soilborne pathogens, with Fusarium spp. found to be predominant in disease surveys conducted in eastern [63] and western Canada [9,14,16]. However, studies investigating the pathogenicity of Fusarium spp. in Canada, especially in western Canada, are limited.
Abdelmagid et al. [16] evaluated the pathogenicity of five F. sporotrichioides isolates from Manitoba, which caused up to 70% root rot severity and significant reductions in root and shoot length in soybean. The cross-pathogenicity of five Fusarium spp., including isolates of F. cerealis, F. culmorum, F. graminearum and F. sporotrichioides collected in Manitoba, was also examined on soybean and wheat; the resulting in root rot severity on soybean ranged from 1.89 to 3.33 on a 0-4 scale [9]. In Alberta, F. proliferatum was reported as the most aggressive species on soybean based on greenhouse trials, while other tested Fusarium spp. caused mild to moderate levels of disease [14]. In this study, the pathogenicity of six Fusarium species was compared on soybean, using isolates previously collected from Manitoba. These species included F. oxysporum, F. graminearum, F. solani, F. acuminatum, which are predominant in North America, as well as F. avenaceum, which is also prevalent in Canada, and F. redolens, which has previously reported in the country [9,14]. The severity of root rot caused by these six species varied, with F. oxysporum, F. avenaceum and F. graminearum causing the most severe root rot on the soybean cultivar ‘Akras’. Despite this variability, root rot severity was generally greater to previous studies [14,52,53], suggesting that Fusarium spp. have become more aggressive in recent decades. This trend of increasing virulence should be emphasized to farmers when implementing control measures against Fusarium root rot in soybean.
Several studies have detected horizontal resistance in soybean controlled by polygenetic loci against Fusarium spp. Acharya et al.[57] identified one major and one minor QTL on soybean chromosomes 8 and 6 controlling partial resistance to F. graminearum. Quantitative resistance was also detected against SDS on all 20 soybean chromosomes [56,58]. In this study, 20 commercial soybean cultivars were screened for resistance to virulent isolates of F. oxysporum, F. graminearum, F. solani, F. acuminatum, F. avenaceum and F. redolens selected from the earlier pathogenicity test. The host reactions revealed varying degrees of tolerance. Fusarium oxysporum and F. avenaceum were identified as the most virulent species, while F. solani caused the lowest root rot severity, consistent with the findings of the pathogenicity testing. Complete resistance (root rot severity ≤ 1) was not observed on any of the 20 soybean cultivars evaluated. Moderate tolerance was detected in one, one, five, 15 and nine cultivars tested against F. acuminatum, F. avenaceum, F. graminearum, F. redolens and F. solani, respectively (1 ≤ DS1 ≤2). However, when considering non-germinated seeds as dead plants due to pathogen infection, moderate resistance was only observed against F. redolens and F. solani (1 ≤DS2 ≤2) (Supplementary Table 1). In this context, the level of tolerance was taken into consideration, evaluating all of the traits investigated in this study. The six Fusarium spp. tested also had varying effects on germination counts, plant height, dry shoot weight and dry root weight. In another study, 57 commercial soybean cultivars were evaluated against F. oxysporum, F. graminearum, F. avenaceum, and F. tricinctum, with resistance against all four species identified in the soybean cultivar ‘Maple Amber’, based on root rot severity, rather than emergence, plant height or dry root weight [63]. However, in that study, the root rot severity ranged from 0.5 to 2.7 on a 0-5 scale, and reductions in emergence, plant height and dry root weight were generally < 50%. Nyandoro et al. [52] evaluated the resistance of 12 soybean cultivars against F. avenaceum in greenhouse trials and found high root rot severity among cultivars (ranging from 2.6-3.4) and considerable reductions in emergence (26.7-75.5%). Given the variable performance of soybean cultivars across different trails, PCA was conducted to evaluate tolerance to Fusarium spp., taking into account root rot severity, germination counts, plant height, dry shoot weight and dry root weight. Some tolerant cultivars were identified against each Fusarium spp., with broad-spectrum resistance detected in ‘P15T46R2′ and ‘B150Y1′. The cultivars were tolerant/partially resistant to F. graminearum, F. oxysporum, F. redolens and F. solani.
Numerous studies have highlighted the variability of Fusarium spp. associated with the soybean root rot complex across different environments, locations and years [11,73,74,75]. In the current study, 18 Fusarium spp. were identified from a large collection of symptomatic roots using a combination of morphological and molecular methods (Supplementary Table 2). All the six species included in the pathogenicity test and cultivar evaluation trials were also recovered in the fungal isolation study, with F. avenaceum, F. oxysporum, F. acuminatum and F. redolens among the major groups. Another significant group was F. equiseti, which has been commonly reported in north California and South Dakota in the USA [76,77] as well as in Ontario, Alberta and Manitoba, Canada [9,14,27]. However, species such as F. poae, F. graminearum, F. solani, F. sporotrichioides, F. tricinctum, F. torulosum, F. commune and F. proliferatum , which were reported as predominant in Canada [9,14,27], were only sparsely identified or absent in the current study. Notably, isolates of F. acuminatum, F. solani, and F. redolens displayed location-specific characteristics, consistent with previous reports [9,14].
Colony morphology played a crucial role for the primary grouping and species selection of isolates within the same field, allowing us to narrow down the initial 983 isolates to 336 isolates for molecular identification. The isolates grouped into “White2” were identified as F. redolens except for “NSRR22_186” and “NSRR22_187, which were identified as F. solani. Similarly, all the “White4” isolates and a few “White1” isolates were identified as F. incarnatum-equiseti. Fusarium oxysporum had three morphology types, including “White1”, “Purple1” and “Slimy1”. The similarity in morphology between F. oxysporum and F. equiseti has also been reported in tomato in northeast India [78]. Additionally, three isolates in the “Purple1” subgroup were identified as F. verticillioides. Within the “Red” group, however, the four subgroups (“Red2,” “Red2,” “Red3,” and “Red4”) failed to distinguish whether the isolates belonged to FTSC or FSSC.
Furthermore, sequencing of DNA fragments amplified by PCR based on the ITS region and beta-tubulin gene identified 225 and 79 isolates that belonged to Fusarium spp., respectively. Inconsistencies were frequently observed among species identified within the same Fusarium species complex. For example, while five isolates were classified as F. incarnatum or F. equiseti with the primer set ITS4/5, they were recognized as F. flagelliforme (also a member of FIESC) with the T12 primer set (beta-tubulin gene). Notably, F. flagelliforme was not detected using ITS4/5 in our current study. Chang et al. [26] demonstrated that variable identification of isolates across species complexes or among species within the Fusarium genus was common in studies with multiple primers, including those targeting the beta-tubulin and ITS regions. For instance, ITS sequences have been reported as unsuitable for distinguishing F. equiseti or F. incarnatum [34] . Similarly, the beta-tubulin sequence has been applied to identify species in the FIESC and F. chlamydosporum Species Complexes (FCSC)[79], but failed to distinguish F. armeniacum, F. acuminatum, F. sportrichioides and F. langsethiae [80]. While the TEF1a gene has been widely used to define species and reveal phylogenetic relationships within the genus Fusarium in the past, multiple primers are more frequently employed in recent studies [9,19,41,81,82]. The results of this study underscore the need to use a combination of colony morphology and multiple primers to identify Fusarium spp. accurately.
Phylogenetic analysis successfully grouped most of the Fusarium spp. identified in this study into distinct complexes, such as FTSC, FOSC, FIESC, FSSC, F. redolens and F. solani. However, only FIESC, F. redolens and F. solani had strong bootstrapping support (BV≥70) and displayed location-specific identification. Within groups like FTSC, FOSC-2, and FSSC, the bootstrap values varied, indicating significant genetic variation. For F. oxysporum, the isolates in the morphological subgroups “Slimy1”, “White1” and “Purple1” were not well distinguished by ITS4/5. Meanwhile, isolate overlap was observed in FTSC between F. avenaceum and F. acuminatum, as well as in FIESC between F. equiseti and F. incarnatum. On the other hand, beta-tubulin sequences clearly distinguished F. avenaceum and F. acuminatum in FTSC as well as F. equiseti and F. flagelliforme. O’Donnell et al. [82] concluded that the beta-tubulin gene is not universally informative within Fusarium and is only suitable to distinguish Fusarium spp. forming part of the F. solani and F. incarnatum-equiseti species complexes. This study was the first to use the T12 primer set specific for the beta-tubulin gene to separate Fusarium spp. in FTSC and FIESC. Hafez et al. [9] also generated a phylogenetic tree that clearly distinguished Fusarium spp. of FTSC, FSSC, FOSC, FIESC, F. redolens and F. solani collected from Carman and Melita, Manitoba. In a study investigating Fusarium isolates from central and southern Alberta, overlapping species were found in phylogenetic trees based on both the ITS and TEF1a sequences [14]. Fusarium oxysporum was reported to have extensive genetic variation [10] and morphological variability [83]. A whole genome sequence study demonstrated the large diversity in FOSC in Australia among the identified clades [75]. Overall, phylogenetic analysis based solely on ITS4/5 was insufficient to classify the different Fusarium species complexes or a single species, while beta-tubulin clearly distinguished Fusarium spp. in FTSC and FIESC.
5. Conclusions
Evaluation of the pathogenicity of six Fusarium spp. on soybean indicated that F. avenaceum and F. oxysporum were the most strongly virulent, while F. graminearum, F. acuminatum, and F. redolens also caused significant levels of disease. In contrast, F. solani was weakly virulent, causing mild symptoms of Fusarium root rot on soybean. An assessment of the reaction of a suite of 20 soybean cultivars to inoculation with each of the Fusarium spp., based on root rot severity, emergence, plant height, and dry shoot and root weight, indicated that while no hosts were completely resistant, some cultivars showed partial resistance or tolerance to disease. The soybean cultivars ‘P15T46R2′ and ‘B150Y1′ in particular were consistently tolerant to F. graminearum, F. oxysporum, F. redolens and F. solani, making them promising candidates for farmers to minimize the risk of Fusarium root rot. Furthermore, this study provided valuable insights into the distribution and composition of Fusarium spp. in the major soybean production areas of Canada, updating and complementing existing information on the Fusarium root rot complex in soybean cultivation. These findings may help to guide the development of effective measures to mitigate the risk of Fusarium root rot of soybean.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Phylogenetic tree of 221 Fusarium isolates identified; Table S1: Root rot severity, emergence, plant height, and dry shoot and root weight in response to inoculation with each of six Fusarium spp.; Table S2: Full list of fungal isolates and associated details.
Author Contributions
Conceptualization, S.E.S, Y.M.K and S.F.H; methodology, R.F.A. and L.F.W.; validation, Y.M.K and S.E.S.; formal analysis, L.F.W. and S.H.O; writing—original draft preparation, L.F.W.; writing—review and editing, S.E.S., O.W., R.R.B. and Y.M.K.; visualization, L.F.W. and S.H.O; supervision, S.E.S. and Y.M.K; project administration, S.F.H., Y.M.K and S.E.S; funding acquisition, S.F.H., Y.M.K and S.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agriculture and Agri-Food Canada AgriScience Program through the Canadian Agricultural Partnership, with industry support from the Canadian Field Crop Research Alliance (CFCRA) whose members include: Atlantic Grains Council; Producteurs de grains du Quebec; Grain Farmers of Ontario; Manitoba Crop Alliance; Manitoba Pulse & Soybean Growers; Saskatchewan Pulse Growers; Prairie Oat Growers Association; SeCan; and FP Genetics.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Analysis of variance, least significant difference analysis of estimated mean in pathogenicity test and cultivar evaluation in greenhouse trials, sequence identification in genetic diversity evaluation are available in the main manuscript or as supplementary data.
Acknowledgments
The authors acknowledge the technical support of Ashley Wragg, Vanessa Tremblay, Waldo Penner, Dennis Stoesz, Shelby Zatylny, and Tom Henderson. The authors thank Manitoba Agriculture, Manitoba Pulse and Soybean Growers, Saskatchewan Ministry of Agriculture, and Saskatchewan Pulse Growers for contributing soybean samples. The in-kind support of the University of Alberta and Agriculture and Agri-Food Canada is also gratefully acknowledged.
Conflicts of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
References
- Michelfelder, A.J. Soy: A Complete Source of Protein. afp 2009, 79, 43–47. [Google Scholar]
- Voora, V.; Larrea, C.; Bermúdez, S. Global Market Report: Soybeans. 2020.
- Shahbandeh, M. Production of Soybeans in Leading Countries Worldwide, 2012-2023. Available online: https://www.statista.com/statistics/263926/soybean-production-in-selected-countries-since-1980/ (accessed on 25 April 2023).
- Government of Canada, S.C. Estimated Areas, Yield, Production, Average Farm Price and Total Farm Value of Principal Field Crops, in Metric and Imperial Units. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210035901 (accessed on 17 April 2023).
- Barthet, V.J.; Puvirajah, A. Quality of Canadian Oilseed-Type Soybeans 2022. 2022.
- Canada, A. and A.-F. Canada: Outlook for Principal Field Crops, 2022-11-18. Available online: https://agriculture.canada.ca/en/sector/crops/reports-statistics/canada-outlook-principal-field-crops-2022-11-18 (accessed on 25 April 2023).
- Hartman, G.L.; Sinclair, J.B.; Rupe, J.C. Compendium of Soybean Diseases, 4th Edn, Ed. G. L. HARTMAN, J. B. SINCLAIR & J. C. RUPE. Vi+100 Pp. St. Paul, Minnesota: APS Press (1999). $37.00 (Paperback). ISBN 0 89054 238 4. The Journal of Agricultural Science 2000, 135, 95–100. [Google Scholar] [CrossRef]
- Bradley, C.A.; Allen, T.W.; Sisson, A.J.; Bergstrom, G.C.; Bissonnette, K.M.; Bond, J.; Byamukama, E.; Chilvers, M.I.; Collins, A.A.; Damicone, J.P.; et al. Soybean Yield Loss Estimates Due to Diseases in the United States and Ontario, Canada, from 2015 to 2019. Plant Health Progress 2021, 22, 483–495. [Google Scholar] [CrossRef]
- Hafez, M.; Abdelmagid, A.; Aboukhaddour, R.; Adam, L.R.; Daayf, F. Fusarium Root Rot Complex in Soybean: Molecular Characterization, Trichothecene Formation, and Cross-Pathogenicity. Phytopathology® 2021, 111, 2287–2302. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.M.D.; Munkvold, G.P.; Ellis, M.L.; Leandro, L.F.S. Distribution and Frequency of Fusarium Species Associated with Soybean Roots in Iowa. Plant Disease 2013, 97, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Leandro, L.; Munkvold, G. Frequency of Isolation, Aggressiveness, and Impact on Yield of Fusarium Root Rot Species in Soybean in Iowa; 2012; p. 30.
- Michielse, C.B.; Rep, M. Pathogen Profile Update: Fusarium Oxysporum. Molecular Plant Pathology 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.M.; Armstrong, J.K. Biological races of the Fusarium causing wilt of Cowpea and Soybeans. Phytopathology 1950, 40. [Google Scholar]
- Zhou, Q.; Li, N.; Chang, K.-F.; Hwang, S.-F.; Strelkov, S.E.; Conner, R.L.; McLaren, D.L.; Fu, H.; Harding, M.W.; Turnbull, G.D. Genetic Diversity and Aggressiveness of Fusarium Species Isolated from Soybean in Alberta, Canada. Crop Protection 2018, 105, 49–58. [Google Scholar] [CrossRef]
- Chang, K.F.; Hwang, S.F.; Conner, R.L.; Ahmed, H.U.; Zhou, Q.; Turnbull, G.D.; Strelkov, S.E.; McLaren, D.L.; Gossen, B.D. First Report of Fusarium Proliferatum Causing Root Rot in Soybean (Glycine Max L.) in Canada. Crop Protection 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Abdelmagid, A.; Hafez, M.; Soliman, A.; Adam, L.R.; Daayf, F. First Report of Fusarium Sporotrichioides Causing Root Rot of Soybean in Canada and Detection of the Pathogen in Host Tissues by PCR. Canadian Journal of Plant Pathology 2021, 43, 527–536. [Google Scholar] [CrossRef]
- Yang, X.B.; Feng, F. Ranges and Diversity of Soybean Fungal Diseases in North America. Phytopathology® 2001, 91, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Pioli, R.N.; Mozzoni, L.; Morandi, E.N. First Report of Pathogenic Association Between Fusarium Graminearum and Soybean. Plant Disease 2004, 88, 220–220. [Google Scholar] [CrossRef]
- Zhao, L.; Wei, X.; Zheng, T.; Gou, Y.-N.; Wang, J.; Deng, J.-X.; Li, M. Evaluation of Pathogenic Fusarium Spp. Associated with Soybean Seed (Glycine Max L.) in Hubei Province, China. Plant Dis 2022. [Google Scholar] [CrossRef]
- Abdelmagid, A.; Hafez, M.; Lawley, Y.; Adam, L.R.; Daayf, F. First Report of Fusarium Cerealis Causing Root Rot on Soybean. Plant Disease 2018, 102, 2638. [Google Scholar] [CrossRef]
- Hartman, G.L. Compendium of Soybean Diseases and Pests; Fifth edition.; APS Press, The American Phytopathological Society, 2015. ISBN 978-0-89054-473-0.
- Ellis, M.L.; Arias, M.M.D.; Leandro, L.F.; Munkvold, G.P. First Report of Fusarium Armeniacum Causing Seed Rot and Root Rot on Soybean (Glycine Max) in the United States. Plant Disease 2012, 96, 1693–1693. [Google Scholar] [CrossRef]
- Ellis, M.L.; Arias, M.M.D.; Jimenez, D.R.C.; Munkvold, G.P.; Leandro, L.F. First Report of Fusarium Commune Causing Damping-off, Seed Rot, and Seedling Root Rot on Soybean (Glycine Max) in the United States. Plant Dis 2013, 97, 284. [Google Scholar] [CrossRef]
- Killebrew, J.F. Greenhouse and Field Evaluation of Fusarium Solani Pathogenicity to Soybean Seedlings. Plant Dis. 1988, 72, 1067. [Google Scholar] [CrossRef]
- Cruz Jimenez, D.R.; Ellis, M.L.; Munkvold, G.P.; Leandro, L.F.S. Isolate–Cultivar Interactions, In Vitro Growth, and Fungicide Sensitivity of Fusarium Oxysporum Isolates Causing Seedling Disease on Soybean. Plant Disease 2018, 102, 1928–1937. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Dai, H.; Wang, D.; Zhou, H.; He, W.; Fu, Y.; Ibrahim, F.; Zhou, Y.; Gong, G.; Shang, J.; et al. Identification of Fusarium Species Associated with Soybean Root Rot in Sichuan Province, China. Eur J Plant Pathol 2018, 151, 563–577. [Google Scholar] [CrossRef]
- Zhang, J.X.; Xue, A.G.; Cober, E.R.; Morrison, M.J.; Zhang, H.J.; Zhang, S.Z.; Gregorich, E. Prevalence, Pathogenicity and Cultivar Resistance of Fusarium and Rhizoctonia Species Causing Soybean Root Rot. cjps 2013, 93, 221–236. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons, 2008. ISBN 978-0-470-27646-4.
- Vujanovic, V.; Hamel, C.; Yergeau, E.; St-Arnaud, M. Biodiversity and Biogeography of Fusarium Species from Northeastern North American Asparagus Fields Based on Microbiological and Molecular Approaches. Microb Ecol 2006, 51, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Hwang, R.; Chang, K.F.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; Conner, R.L.; Turnbull, G.D. Genetic Variation in Fusarium Avenaceum Causing Root Rot on Field Pea. Plant Pathology 2010, 59, 845–852. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; Brankovics, B.; Iltes, J.; van der Lee, T.A.J.; Waalwijk, C. Diagnosis of Fusarium Infections: Approaches to Identification by the Clinical Mycology Laboratory. Curr Fungal Infect Rep 2015, 9, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.M.; Brian Golding, G. Are Similarity- or Phylogeny-Based Methods More Appropriate for Classifying Internal Transcribed Spacer (ITS) Metagenomic Amplicons? New Phytologist 2011, 192, 775–782. [Google Scholar] [CrossRef] [PubMed]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In Pcr Protocols: a Guide to Methods and Applications,; 1990; Vol. 31, pp. 315–322.
- Wang, M.; Chen, Q.; Diao, Y.; Duan, W.; Cai, L. Fusarium Incarnatum-Equiseti Complex from China. Persoonia 2019, 43, 70–89. [Google Scholar] [CrossRef]
- Kalman, B.; Abraham, D.; Graph, S.; Perl-Treves, R.; Meller Harel, Y.; Degani, O. Isolation and Identification of Fusarium Spp., the Causal Agents of Onion (Allium Cepa) Basal Rot in Northeastern Israel. Biology (Basel) 2020, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Q.; Strelkov, S.E.; Hwang, S.-F. Genetic Diversity and Aggressiveness of Fusarium Spp. Isolated from Canola in Alberta, Canada. Plant Disease 2014, 98, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.L.; Rai, S.; Kumar, S.; Srivastava, A.K.; Anandaraj, M.; Sharma, A.K. Mating Type Genes and Genetic Markers to Decipher Intraspecific Variability among Fusarium Udum Isolates from Pigeonpea: Diversity Analysis of Fusarium Udum Isolates. J. Basic Microbiol. 2015, 55, 846–856. [Google Scholar] [CrossRef]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic Analyses of RPB1 and RPB2 Support a Middle Cretaceous Origin for a Clade Comprising All Agriculturally and Medically Important Fusaria. Fungal Genetics and Biology 2013, 52, 20–31. [Google Scholar] [CrossRef]
- Torres-Cruz, T.J.; Whitaker, B.K.; Proctor, R.H.; Broders, K.; Laraba, I.; Kim, H.-S.; Brown, D.W.; O’Donnell, K.; Estrada-Rodríguez, T.L.; Lee, Y.-H.; et al. FUSARIUM-ID v.3.0: An Updated, Downloadable Resource for Fusarium Species Identification. Plant Disease 2022, 106, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Dash, A.; Gurdaswani, V.; D’Souza, J.; Ghag, S. Functional Characterization of an Inducible Bidirectional Promoter from Fusarium Oxysporum f. Sp. Cubense. Scientific Reports 2020, 10. [Google Scholar] [CrossRef]
- Khuna, S.; Kumla, J.; Thitla, T.; Nuangmek, W.; Lumyong, S.; Suwannarach, N. Morphology, Molecular Identification, and Pathogenicity of Two Novel Fusarium Species Associated with Postharvest Fruit Rot of Cucurbits in Northern Thailand. J Fungi (Basel) 2022, 8, 1135. [Google Scholar] [CrossRef] [PubMed]
- Spanic, V.; Lemmens, M.; Drezner, G. Morphological and Molecular Identification of Fusarium Species Associated with Head Blight on Wheat in East Croatia. Eur J Plant Pathol 2010, 128, 511–516. [Google Scholar] [CrossRef]
- Parikh, L.; Kodati, S.; Eskelson, M.J.; Adesemoye, A.O. Identification and Pathogenicity of Fusarium Spp. in Row Crops in Nebraska. Crop Protection 2018, 108, 120–127. [Google Scholar] [CrossRef]
- Moparthi, S.; Burrows, M.; Mgbechi-Ezeri, J.; Agindotan, B. Fusarium Spp. Associated With Root Rot of Pulse Crops and Their Cross-Pathogenicity to Cereal Crops in Montana. Plant Disease 2021, 105, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Shabeer, S.; Tahira, R.; Jamal, A. Fusarium Spp. Mycotoxin Production, Diseases and Their Management: An Overview. PJAR 2021, 34. [Google Scholar] [CrossRef]
- Safarieskandari, S.; Chatterton, S.; Hall, L.M. Pathogenicity and Host Range of Fusarium Species Associated with Pea Root Rot in Alberta, Canada. Canadian Journal of Plant Pathology 2021, 43, 162–171. [Google Scholar] [CrossRef]
- Ellis, M.L.; Munkvold, G.P. Trichothecene Genotype of Fusarium Graminearum Isolates from Soybean (Glycine Max) Seedling and Root Diseases in the United States. Plant Dis 2014, 98, 1012. [Google Scholar] [CrossRef] [PubMed]
- Cruz, D.R.; Leandro, L.F.S.; Mayfield, D.A.; Meng, Y.; Munkvold, G.P. Effects of Soil Conditions on Root Rot of Soybean Caused by Fusarium Graminearum. Phytopathology® 2020, 110, 1693–1703. [Google Scholar] [CrossRef]
- Gaspar, A.P.; Marburger, D.A.; Mourtzinis, S.; Conley, S.P. Soybean Seed Yield Response to Multiple Seed Treatment Components across Diverse Environments. Agronomy Journal 2014, 106, 1955–1962. [Google Scholar] [CrossRef]
- Esker, P.D.; Conley, S.P. Probability of Yield Response and Breaking Even for Soybean Seed Treatments. Crop Science 2012, 52, 351–359. [Google Scholar] [CrossRef]
- McKenzie-Gopsill, A.; Beaton, A.; Foster, A.J. Investigating the Effects of Seed Treatments on the Economically Optimal Seeding Rate of Conventional Soybean in Atlantic Canada. cjps 2022, 103, 201–213. [Google Scholar] [CrossRef]
- Nyandoro, R.; Chang, K.F.; Hwang, S.F.; Ahmed, H.U.; Turnbull, G.D.; Strelkov, S.E. Management of Root Rot of Soybean in Alberta with Fungicide Seed Treatments and Genetic Resistance. Can. J. Plant Sci. 2019, 99, 499–509. [Google Scholar] [CrossRef]
- Zhang, J.X.; Xue, A.G.; Tambong, J.T. Evaluation of Seed and Soil Treatments with Novel Bacillus Subtilis Strains for Control of Soybean Root Rot Caused by Fusarium Oxysporum and F. Graminearum. Plant Disease 2009, 93, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jacobs, J.L.; Roth, M.G.; Chilvers, M.I. Temporal Dynamics of Fusarium Virguliforme Colonization of Soybean Roots. Plant Disease 2019, 103, 19–27. [Google Scholar] [CrossRef]
- Herman, T.K.; Bowen, R.; Mahan, A.L.; Hartman, G.L. Evaluation of Soybean Germplasm for Resistance to Fusarium Virguliforme, the Major Pathogen Causing Sudden Death Syndrome of Soybean in the United States. Crop Science 2023, 63, 1344–1353. [Google Scholar] [CrossRef]
- Kazi, S.; Shultz, J.; Afzal, J.; Johnson, J.; Njiti, V.N.; Lightfoot, D.A. Separate Loci Underlie Resistance to Root Infection and Leaf Scorch during Soybean Sudden Death Syndrome. Theor Appl Genet 2008, 116, 967–977. [Google Scholar] [CrossRef]
- Acharya, B.; Lee, S.; Rouf Mian, M.A.; Jun, T.-H.; McHale, L.K.; Michel, A.P.; Dorrance, A.E. Identification and Mapping of Quantitative Trait Loci (QTL) Conferring Resistance to Fusarium Graminearum from Soybean PI 567301B. Theor Appl Genet 2015, 128, 827–838. [Google Scholar] [CrossRef]
- Swaminathan, S.; Abeysekara, N.S.; Liu, M.; Cianzio, S.R.; Bhattacharyya, M.K. Quantitative Trait Loci Underlying Host Responses of Soybean to Fusarium Virguliforme Toxins That Cause Foliar Sudden Death Syndrome. Theor Appl Genet 2016, 129, 495–506. [Google Scholar] [CrossRef]
- Almeida-Silva, F.; Venancio, T.M. Integration of Genome-Wide Association Studies and Gene Coexpression Networks Unveils Promising Soybean Resistance Genes against Five Common Fungal Pathogens. Sci Rep 2021, 11, 24453. [Google Scholar] [CrossRef] [PubMed]
- Weinig, C.; Schmitt, J. Environmental Effects on the Expression of Quantitative Trait Loci and Implications for Phenotypic Evolution. BioScience 2004, 54, 627–635. [Google Scholar] [CrossRef]
- Li, S.; Wang, J.; Zhang, L. Inclusive Composite Interval Mapping of QTL by Environment Interactions in Biparental Populations. PLOS ONE 2015, 10, e0132414. [Google Scholar] [CrossRef]
- Luckew, A.S.; Cianzio, S.R.; Leandro, L.F. Screening Method for Distinguishing Soybean Resistance to Fusarium Virguliforme in Resistant × Resistant Crosses. Crop Science 2012, 52, 2215–2223. [Google Scholar] [CrossRef]
- Zhang, J.X.; Xue, A.G.; Zhang, H.J.; Nagasawa, A.E.; Tambong, J.T. Response of Soybean Cultivars to Root Rot Caused by Fusarium Species. Can. J. Plant Sci. 2010, 90, 767–776. [Google Scholar] [CrossRef]
- Kim, Y.M.; McLaren, D.; Conner, R.; Henriquez, M.A.; Abdelmagid, A.; Wu, L.; Hwang, S.; Strelkov, S.; Chatterton, S.; Wally, O. The Fusarium Root Rot Complex of Soybean, Dry Bean and Field Pea in Manitoba, Canada. In book of Abstracts, The 12th International congress of Plant Pathology. Lyon, France, August 20-25, 2023. pp. 1124-1125.
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic rDNA ITS2 Types within a Monophyletic Lineage of the FungusFusariumAre Nonorthologous. Molecular Phylogenetics and Evolution 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symposium Series 1999, 41, 95–98. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2022.
- Moreira, G.M.; Machado, F.J.; Pereira, C.B.; Neves, D.L.; Tessmann, D.J.; Ward, T.J.; Del Ponte, E.M. First Report of the Fusarium Tricinctum Species Complex Causing Fusarium Head Blight of Wheat in Brazil. Plant Disease 2020, 104, 586–586. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; van Iperen, A.L.; Starink-Willemse, M.; Sandoval-Denis, M.; Groenewald, J.Z. Citizen Science Project Reveals Novel Fusarioid Fungi (Nectriaceae, Sordariomycetes) from Urban Soils. Fungal Syst Evol 2021, 8, 101–127. [Google Scholar] [CrossRef] [PubMed]
- Paugh, K.R.; Del Castillo Múnera, J.; Swett, C.L. First Report of Fusarium Falciforme (FSSC 3 + 4) Causing Rot of Industrial Hemp (Cannabis Sativa) in California. Plant Disease 2022, 106, 1753. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Yang, X.; Shen, G.; Wang, S.; Teng, H.; Yang, C.; Liu, X.; Wang, X.; Zhao, J.; et al. Fusarium Species Associated with Maize Leaf Blight in Heilongjiang Province, China. Journal of Fungi 2022, 8, 1170. [Google Scholar] [CrossRef] [PubMed]
- Dongzhen, F.; Xilin, L.; Xiaorong, C.; Wenwu, Y.; Yunlu, H.; Yi, C.; Jia, C.; Zhimin, L.; Litao, G.; Tuhong, W.; et al. Fusarium Species and Fusarium Oxysporum Species Complex Genotypes Associated With Yam Wilt in South-Central China. Front Microbiol 2020, 11, 1964. [Google Scholar] [CrossRef]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of Environmental Conditions and Agronomic Practices on the Prevalence of Fusarium Species Associated with Ear- and Stalk Rot in Maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Pramunadipta, S.; Widiastuti, A.; Wibowo, A.; Suga, H.; Priyatmojo, A. Identification and Pathogenicity of Fusarium Spp. Associated with the Sheath Rot Disease of Rice (Oryza Sativa) in Indonesia. J Plant Pathol 2022, 104, 251–267. [Google Scholar] [CrossRef]
- Achari, S.R.; Kaur, J.; Dinh, Q.; Mann, R.; Sawbridge, T.; Summerell, B.A.; Edwards, J. Phylogenetic Relationship between Australian Fusarium Oxysporum Isolates and Resolving the Species Complex Using the Multispecies Coalescent Model. BMC Genomics 2020, 21, 248. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, D.; Rosso, M.L.; Zhang, B. Identification of Fungi Associated with Soybeans and Effective Seed Disinfection Treatments. Food Sci Nutr 2019, 7, 3194–3205. [Google Scholar] [CrossRef] [PubMed]
- Okello, P.N.; Petrović, K.; Kontz, B.; Mathew, F.M. Eight Species of Fusarium Cause Root Rot of Corn (Zea Mays) in South Dakota. Plant Health Progress 2019, 20, 38–43. [Google Scholar] [CrossRef]
- Singha, I.M.; Kakoty, Y.; Unni, B.G.; Das, J.; Kalita, M.C. Identification and Characterization of Fusarium Sp. Using ITS and RAPD Causing Fusarium Wilt of Tomato Isolated from Assam, North East India. Journal of Genetic Engineering and Biotechnology 2016, 14, 99–105. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Gueidan, C.; Crous, P.W.; Geiser, D.M. Novel Multilocus Sequence Typing Scheme Reveals High Genetic Diversity of Human Pathogenic Members of the Fusarium Incarnatum-F. Equiseti and F. Chlamydosporum Species Complexes within the United States. J Clin Microbiol 2009, 47, 3851–3861. [Google Scholar] [CrossRef]
- Nosratabadi, M.; Kachuei, R.; Rezaie, S.; Harchegani, A.B. Beta-Tubulin Gene in the Differentiation of Fusarium Species by PCR-RFLP Analysis. Infez Med 2018, 26, 52–60. [Google Scholar]
- O’Donnell, K.; Gräfenhan, T.; Laraba, I.; Busman, M.; Proctor, R.H.; Kim, H.-S.; Wiederhold, N.P.; Geiser, D.M.; Seifert, K.A. Fusarium Abutilonis and F. Guadeloupense, Two Novel Species in the Fusarium Buharicum Clade Supported by Multilocus Molecular Phylogenetic Analyses. Mycologia 2022, 114, 682–696. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Kim, H.-S.; McCormick, S.P.; Busman, M.; Aoki, T.; et al. DNA Sequence-Based Identification of Fusarium: A Work in Progress. Plant Disease 2022, 106, 1597–1609. [Google Scholar] [CrossRef]
- Tian, Y.H.; Hou, Y.Y.; Peng, C.Y.; Wang, Y.Y.; He, B.L.; Gao, K.X. [Genetic diversity and phylogenetic analysis of Fusarium oxysporium strains isolated from the Cucurbitaceae hosts revealed by SRAPs]. Ying Yong Sheng Tai Xue Bao 2017, 28, 947–956. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Root rot severity following inoculation of soybean with (a) Fusarium oxysporum, (b) Fusarium redolens, (c) Fusarium graminearum, (d) Fusarium solani, (e) Fusarium avenaceum, and (f) Fusarium acuminatum as evaluated on a 0-4 scale based on Chang et al. [17] and Zhou et al. [14].

Figure 2.
Effect of inoculation with each of 12 Fusarium isolates on (a) germination count, (b) plant height; (c) root rot severity; and (d) dry shoot and root weight of two soybean cultivars, ‘Akras’ and ‘B150Y1′. The orange and blue lines in (b-d) denote the estimated mean of the non-inoculated controls of ‘Akras’ and ‘B150Y1′, respectively. The bars indicate the values in response to the different fungal isolates.
Figure 2.
Effect of inoculation with each of 12 Fusarium isolates on (a) germination count, (b) plant height; (c) root rot severity; and (d) dry shoot and root weight of two soybean cultivars, ‘Akras’ and ‘B150Y1′. The orange and blue lines in (b-d) denote the estimated mean of the non-inoculated controls of ‘Akras’ and ‘B150Y1′, respectively. The bars indicate the values in response to the different fungal isolates.
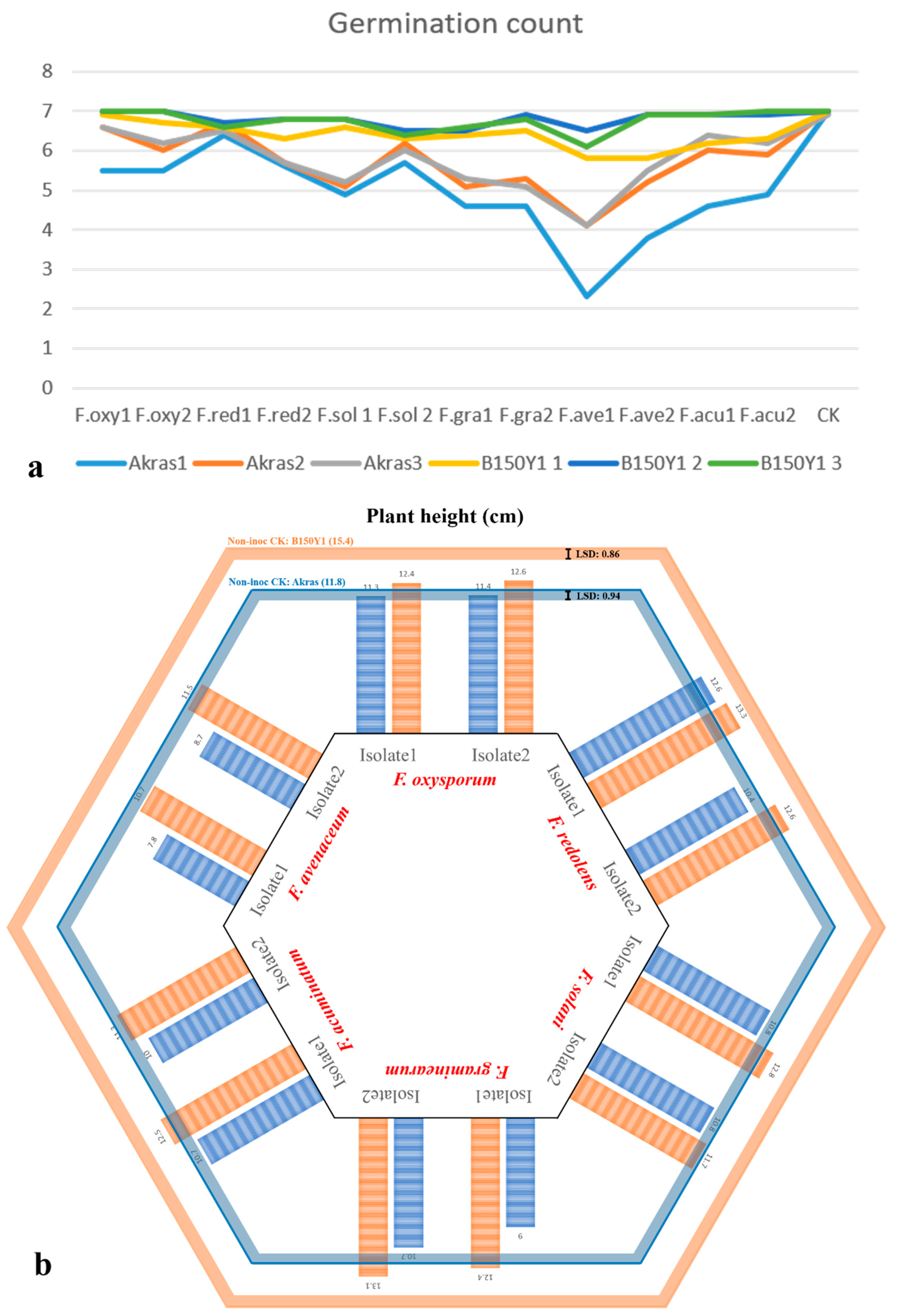
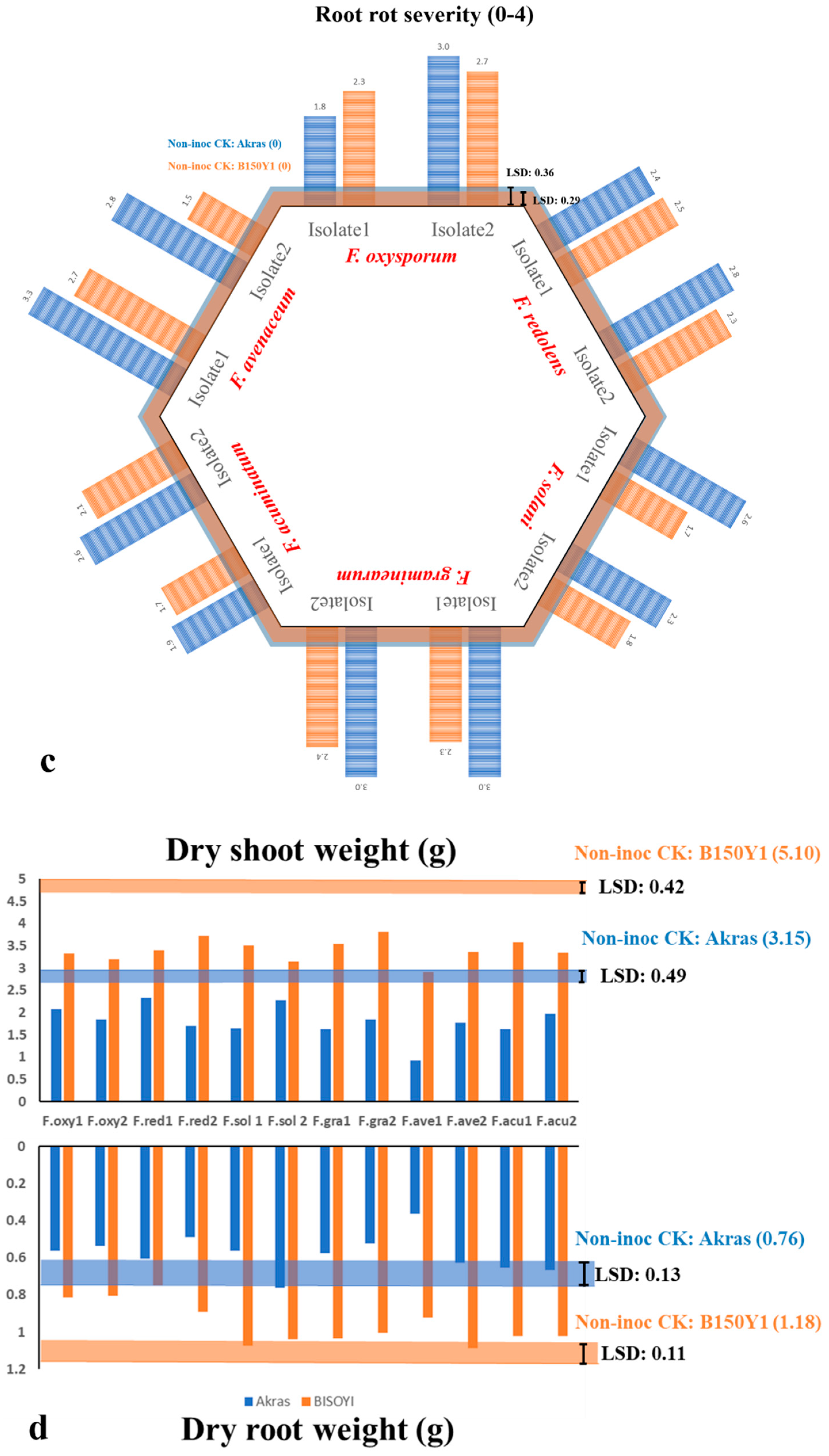
Figure 3.
Cluster and principal component analyses of 20 soybean cultivars evaluated against six Fusarium spp., including (a) Fusarium oxysporum, (b) Fusarium redolens, (c) Fusarium graminearum, (d) Fusarium solani, (e) Fusarium avenaceum and (f) Fusarium acuminatum. Hosts were divided into four groups (denoted by circles; high tolerance, moderate tolerance, moderate susceptibility, and high susceptibility) based on cluster analysis.
Figure 3.
Cluster and principal component analyses of 20 soybean cultivars evaluated against six Fusarium spp., including (a) Fusarium oxysporum, (b) Fusarium redolens, (c) Fusarium graminearum, (d) Fusarium solani, (e) Fusarium avenaceum and (f) Fusarium acuminatum. Hosts were divided into four groups (denoted by circles; high tolerance, moderate tolerance, moderate susceptibility, and high susceptibility) based on cluster analysis.
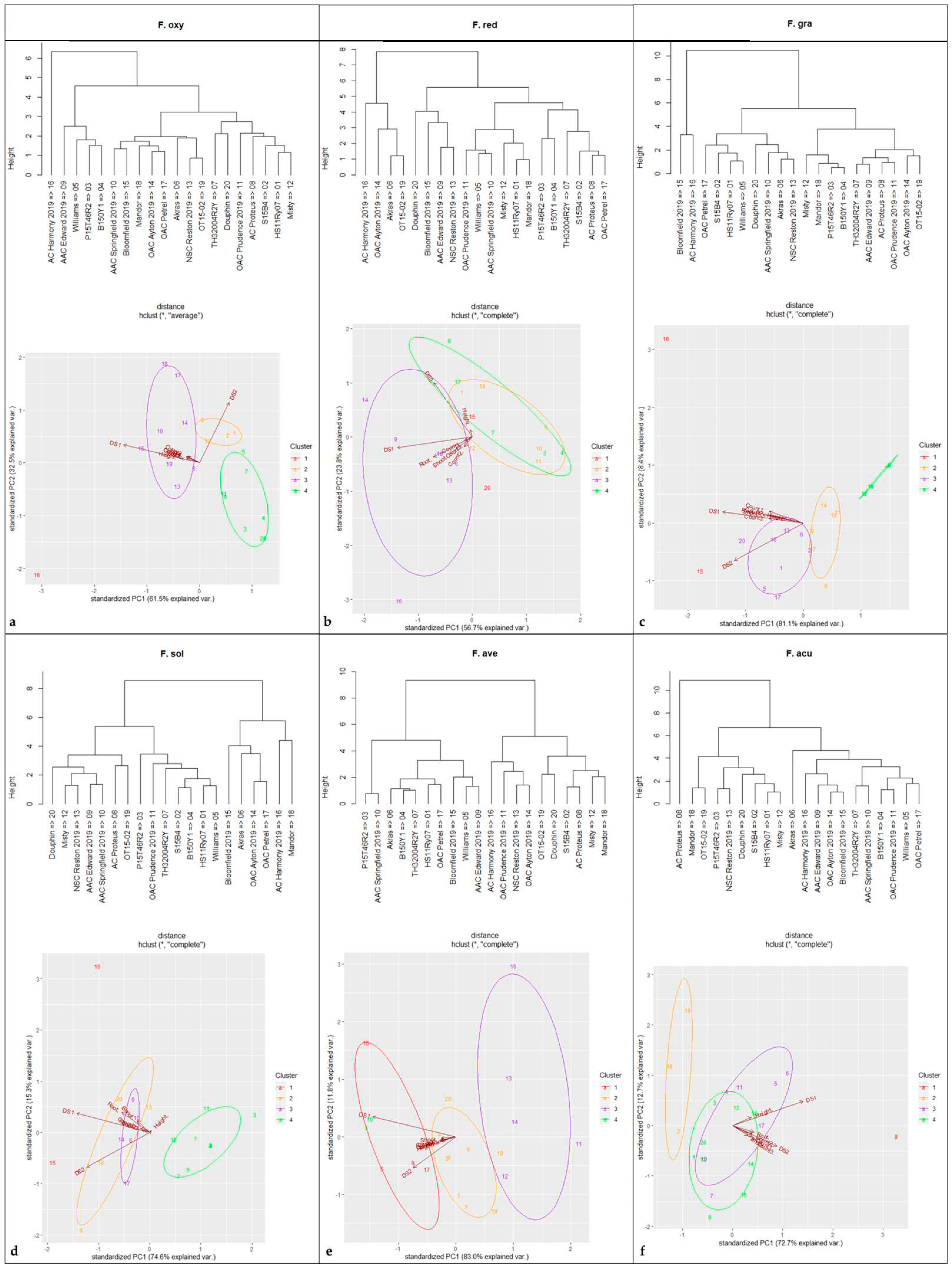
Figure 4.
Colony growth on potato dextrose agar of the Fusarium isolates (a) NSRR22_029, (b) NSRR22_078, (c) NSRR22_113, (d) NSRR22_128, (e) NSRR22_139, (f) NSRR22_175, (j) NSRR22_213, (h) NSRR22_245, and (i) NSRR22_314, representing the colony morphology subgroups “Red1”, “Red2”, “Red3”, “Red4”, “White1”, “White2”, “White4”, “Purple1” and “Slimy1”, respectively. The bar in each panel = 1 cm.
Figure 4.
Colony growth on potato dextrose agar of the Fusarium isolates (a) NSRR22_029, (b) NSRR22_078, (c) NSRR22_113, (d) NSRR22_128, (e) NSRR22_139, (f) NSRR22_175, (j) NSRR22_213, (h) NSRR22_245, and (i) NSRR22_314, representing the colony morphology subgroups “Red1”, “Red2”, “Red3”, “Red4”, “White1”, “White2”, “White4”, “Purple1” and “Slimy1”, respectively. The bar in each panel = 1 cm.
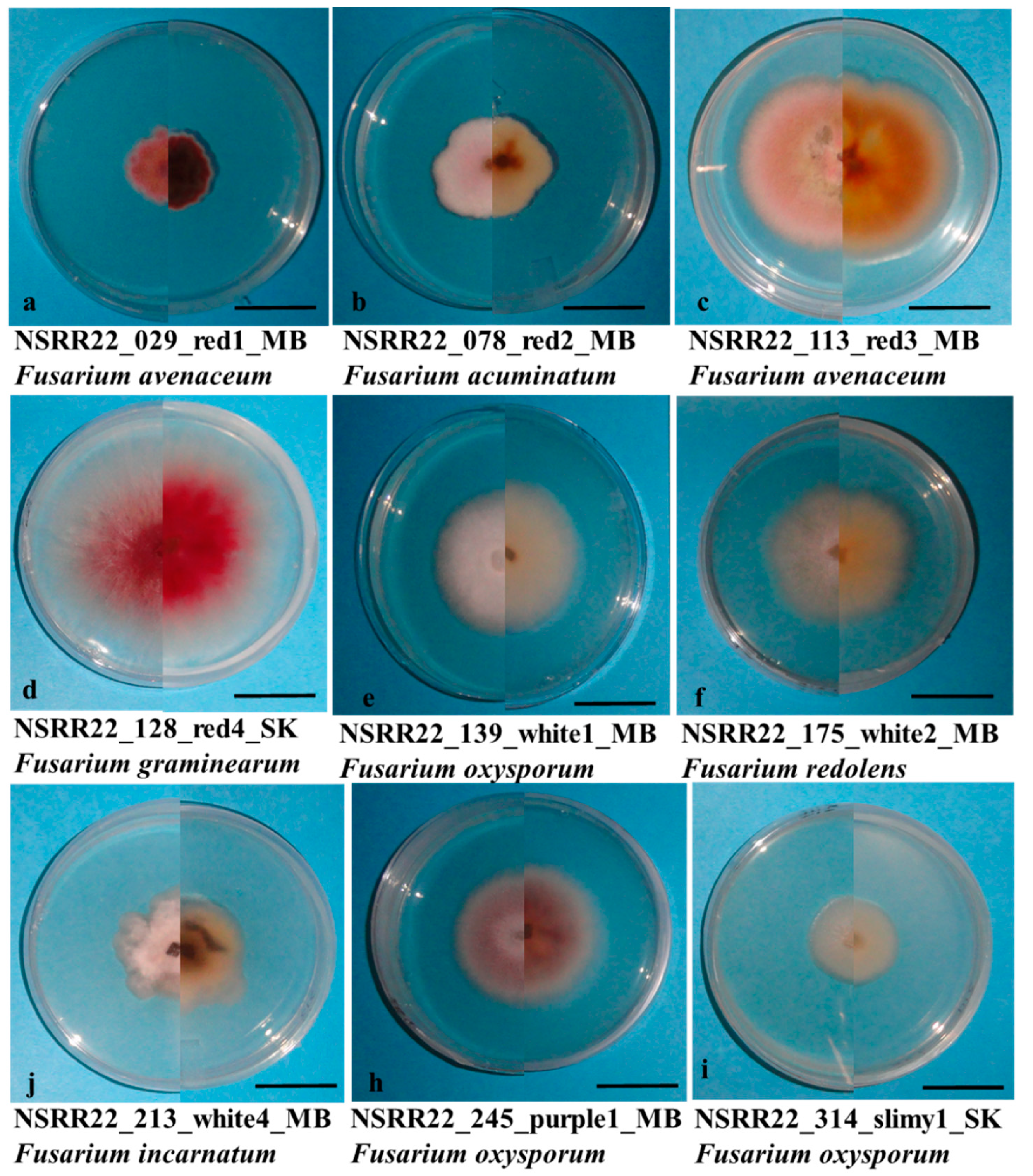
Figure 5.
Molecular identification of reference isolates of Fusarium oxysporum (F. oxy), Fusarium redolens (F. red), Fusarium graminearum (F. gra), Fusarium solani (F. sol), Fusarium avenaceum (F. ave), and Fusarium acuminatum (F. acu). Genomic DNA of each isolate was amplified with each of two primer sets: (a) ITS4/5 (targeting the ITS region) and (b) T12 (targeting the beta-tubulin gene), and the products resolved by electrophoresis on 2% agarose. A band (500 bp – 600 bp) is visible for each isolate. A DNA ladder is included on the left of each panel.
Figure 5.
Molecular identification of reference isolates of Fusarium oxysporum (F. oxy), Fusarium redolens (F. red), Fusarium graminearum (F. gra), Fusarium solani (F. sol), Fusarium avenaceum (F. ave), and Fusarium acuminatum (F. acu). Genomic DNA of each isolate was amplified with each of two primer sets: (a) ITS4/5 (targeting the ITS region) and (b) T12 (targeting the beta-tubulin gene), and the products resolved by electrophoresis on 2% agarose. A band (500 bp – 600 bp) is visible for each isolate. A DNA ladder is included on the left of each panel.
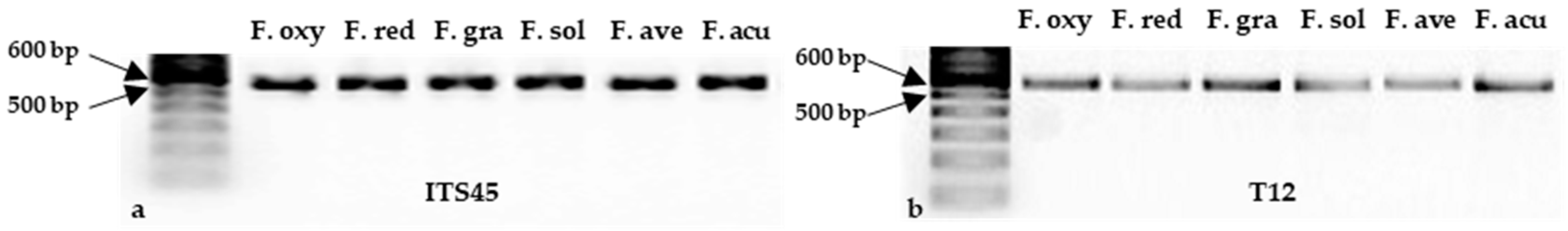
Figure 6.
Phylogenetic tree of Fusarium isolates in (a) F. tricinctum species complex (FTSC) and (b) F. incarnatum-equiseti species complex (FIESC) from Manitoba (MB), Saskatchewan (SK), Ontario (ON) and Quebec (QC) based on neighbour-joining analysis of the beta-tubulin sequence. Bootstrap values, which are indicated on the branches, were based on 1000 replicates.
Figure 6.
Phylogenetic tree of Fusarium isolates in (a) F. tricinctum species complex (FTSC) and (b) F. incarnatum-equiseti species complex (FIESC) from Manitoba (MB), Saskatchewan (SK), Ontario (ON) and Quebec (QC) based on neighbour-joining analysis of the beta-tubulin sequence. Bootstrap values, which are indicated on the branches, were based on 1000 replicates.
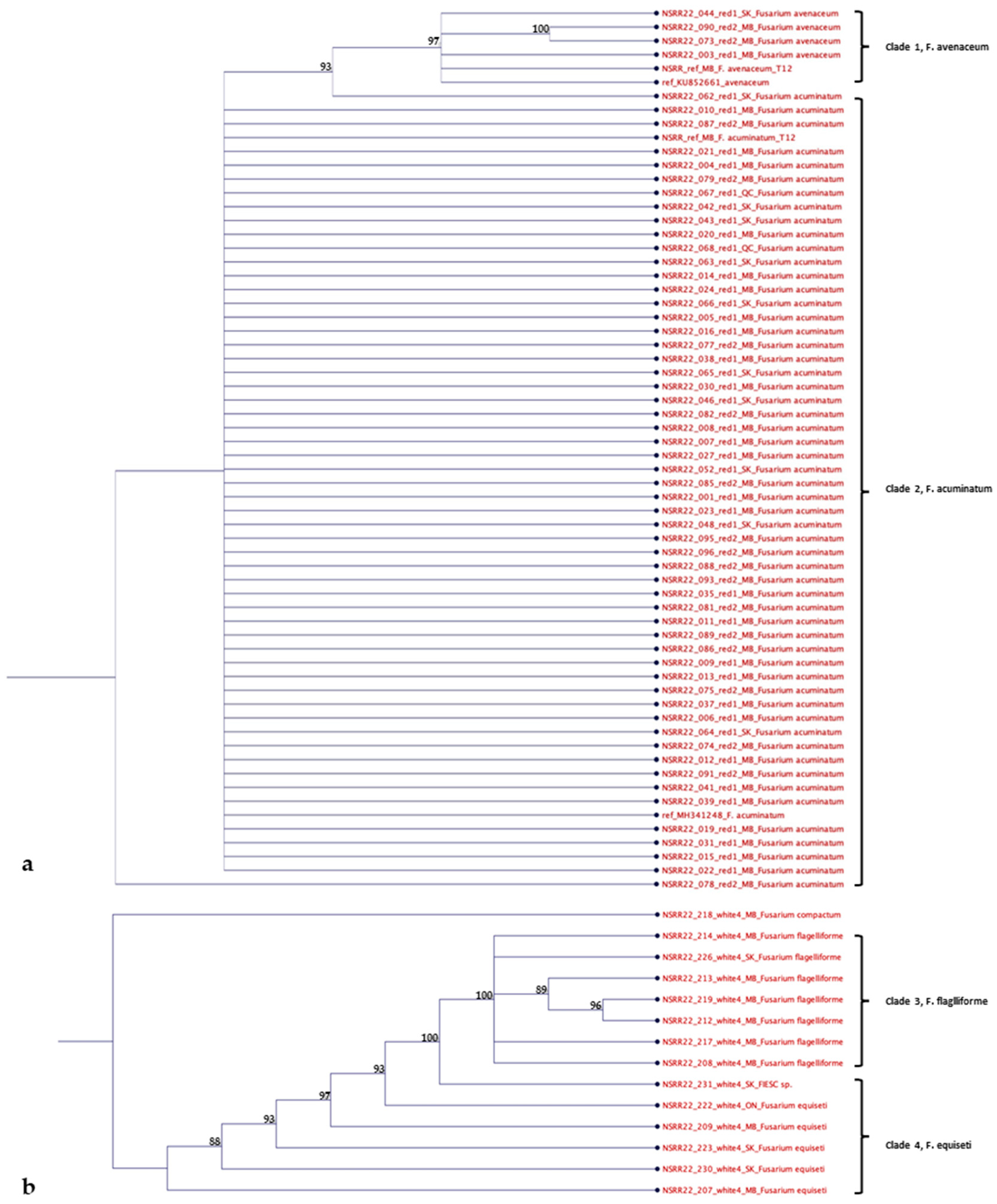

Table 1.
ANOVA table for three germination counts (Count1, Count2 and Count3) taken at 7, 14 and 21 days after seeding, plant height (Height), disease severity with two calculation methods (DS1 and DS2), dry shoot weight (Shoot) and dry root weight (Root) of two soybean cultivars, ‘Akras’ and ‘B150Y1′, inoculated under greenhouse conditions with each of 12 fungal isolates representing F. oxysporum, F. redolens, F. graminearum, F. solani, F. avenaceum and F. acuminatum.
Table 1.
ANOVA table for three germination counts (Count1, Count2 and Count3) taken at 7, 14 and 21 days after seeding, plant height (Height), disease severity with two calculation methods (DS1 and DS2), dry shoot weight (Shoot) and dry root weight (Root) of two soybean cultivars, ‘Akras’ and ‘B150Y1′, inoculated under greenhouse conditions with each of 12 fungal isolates representing F. oxysporum, F. redolens, F. graminearum, F. solani, F. avenaceum and F. acuminatum.
| Source of Variance | Df | Mean square | |||||||
| Count1 | Count2 | Count3 | Height | DS1 | DS2 | Shoot | Root | ||
| F.spp1 | 12 | 10.4*2 | 4.1* | 4.7* | 25.5* | 9.7* | 11* | 4.3* | 0.2* |
| CV1 | 1 | 128.8* | 70.1* | 57.3* | 261.4* | 2.9* | 10.7* | 175.9* | 9.5* |
| Repeat1 | 1 | 0.2 | 1.4 | 0.3 | 0.3 | 0.0 | 0.0 | 0.1 | 0.1 |
| F.spp:CV | 12 | 3.8* | 2.6* | 2.0* | 4.3* | 0.7* | 1.0* | 0.9* | 0.1* |
| F.spp:Repeat | 12 | 1.5 | 0.8 | 1.0 | 0.8 | 0.1 | 0.2 | 0.3 | 0.0 |
| CV:Repeat | 1 | 1.7 | 0.9 | 0.4 | 0.0 | 0.0 | 0.0 | 0.5 | 0.1 |
| F.spp:CV:Repeat | 12 | 0.8 | 0.9 | 1.0 | 1.0 | 0.1 | 0.2 | 0.2 | 0.0 |
| Residuals | 208 | 1.0 | 0.6 | 0.6 | 1.1 | 0.1 | 0.1 | 0.2 | 0.0 |
1“F.spp”, “CV” and “Repeat” refer to the variance from 12 Fusarium isolates and non-inoculated controls, two soybean cultivars, and two repeated greenhouse experiments, respectively. 2A bold mean squares denoted with an asterisk (*) indicates that the treatment effect was significant (p <0.01).
Table 2.
ANOVA table of 20 soybean cultivars evaluated for reaction to Fusarium oxysporum, Fusarium redolens, Fusarium graminearum, Fusarium solani, Fusarium avenaceum and Fusarium acuminatum in greenhouse studies. The estimated traits included three germination counts (Count1, Count2 and Count3) taken at 7, 14 and 21 days after seeding, plant height (Height), disease severity with two calculation methods (DS1 and DS2), dry shoot weight (Shoot) and dry root weight (Root).
Table 2.
ANOVA table of 20 soybean cultivars evaluated for reaction to Fusarium oxysporum, Fusarium redolens, Fusarium graminearum, Fusarium solani, Fusarium avenaceum and Fusarium acuminatum in greenhouse studies. The estimated traits included three germination counts (Count1, Count2 and Count3) taken at 7, 14 and 21 days after seeding, plant height (Height), disease severity with two calculation methods (DS1 and DS2), dry shoot weight (Shoot) and dry root weight (Root).
| Source of Variance | Df | Mean square | |||||||
| Count1 | Count2 | Count3 | Height | DS1 | DS2 | Shoot | Root | ||
| F.spp1 | 6 | 643.2*2 | 657.1* | 709* | 1597.9* | 139.4* | 227.1* | 28.9* | 2.12* |
| CV1 | 19 | 28.0* | 26.8* | 25.8* | 106.4* | 2.3* | 2.0* | 2.2* | 0.18* |
| Repeat1 | 1 | 0.1 | 0.1 | 0.0 | 10.3 | 0.1 | 0.3 | 0.0 | 0.00 |
| CV:F.spp | 114 | 3.1* | 2.8* | 3.1* | 15* | 0.9* | 0.5* | 0.2* | 0.02* |
| CV:Repeat | 19 | 0.6 | 0.6 | 0.6 | 5.3 | 0.5 | 0.2 | 0.0 | 0.00 |
| F.spp:Repeat | 6 | 1.0 | 1.7 | 1.3 | 0.7 | 0.1 | 0.5 | 0.0 | 0.00 |
| CV:F.spp:Repeat | 114 | 0.8 | 0.7 | 0.7 | 4.3 | 0.4 | 0.2 | 0.0 | 0.00 |
| Residuals | 839 | 1.6 | 1.6 | 1.5 | 7.7 | 0.6 | 0.3 | 0.1 | 0.01 |
1“F.spp”, “CV” and “Repeat” refer to the variance from six Fusarium species and non-inoculated controls, 20 soybean cultivars, and two repeated greenhouse experiments, respectively. 2A bold mean squares denoted with an asterisk (*) indicates that the treatment effect was significant (p <0.01).
Table 3.
The frequency (%) of identification of Fusarium acuminatum (F. acu), Fusarium avenaceum (F. ave), Fusarium oxysporum (F. oxy), Fusarium graminearum (F, gra), Fusarium equiseti (F. equ), Fusarium solani (F. sol),and Fusarium redolens (F. red), and four Fusarium species complexes (Fusarium tricinctum species complex (FTSC), Fusarium oxysporum species complex (FOSC), Fusarium incarnatum-equiseti species complex (FIESC), and Fusarium sambucinum species complex (FSSC)) in samples collected from Manitoba (MB), Saskatchewan (SK), Ontario (ON) and Quebec (QC).
Table 3.
The frequency (%) of identification of Fusarium acuminatum (F. acu), Fusarium avenaceum (F. ave), Fusarium oxysporum (F. oxy), Fusarium graminearum (F, gra), Fusarium equiseti (F. equ), Fusarium solani (F. sol),and Fusarium redolens (F. red), and four Fusarium species complexes (Fusarium tricinctum species complex (FTSC), Fusarium oxysporum species complex (FOSC), Fusarium incarnatum-equiseti species complex (FIESC), and Fusarium sambucinum species complex (FSSC)) in samples collected from Manitoba (MB), Saskatchewan (SK), Ontario (ON) and Quebec (QC).
| F. acu | F. ave | FTSC | F. oxy | FOSC | F. gra | FSSC | F. equ | FIESC | F. sol | F. red | |
| MB | 13.6 | 15.8 | 31.2 | 11.3 | 12.2 | 0.0 | 1.8 | 5.0 | 6.3 | 0.0 | 4.5 |
| SK | 4.1 | 7.7 | 12.7 | 2.7 | 2.7 | 1.4 | 3.2 | 2.3 | 4.5 | 0.0 | 4.1 |
| Western | 17.6 | 23.5 | 43.9 | 14.0 | 14.9 | 1.4 | 5.0 | 7.2 | 10.9 | 0.0 | 8.6 |
| ON | 0.0 | 0.0 | 0.0 | 3.6 | 4.1 | 0.0 | 0.0 | 0.5 | 0.9 | 1.4 | 0.0 |
| QC | 0.0 | 2.7 | 2.7 | 1.4 | 1.4 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 |
| Eastern | 0.0 | 2.7 | 2.7 | 5.0 | 5.4 | 0.0 | 0.9 | 0.5 | 0.9 | 0.0 | 0.0 |
| Total | 17.6 | 26.2 | 46.6 | 19.0 | 20.4 | 1.4 | 5.9 | 7.7 | 11.8 | 1.4 | 8.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



