
Submitted:
18 November 2023
Posted:
23 November 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Severe COVID-19 is a major cause of morbidity and mortality worldwide, especially to those with co-morbidities, the elderly and the immunocompromised. However, the molecular determinants critical for severe COVID-19 progression remain to be fully elucidated. Meta-analysis of transcriptomic RNAseq and single-cell sequencing datasets comparing between severe and mild COVID-19 patients have demonstrated that the early expansion of myeloid-derived suppressor cells (MDSCs) could a key feature of severe COVID-19 progression. Besides serving as potential early prognostic biomarkers for severe COVID-19 progression, several studies have also indicated the functional roles of MDSCs in severe COVID-19 pathogenesis and possibly even long COVID. Given the potential links between MDSCs and severe COVID-19, we examined the existing literature summarizing the characteristics of MDSCs, the evidence of MDSCs in facilitating severe COVID-19 pathogenesis and discuss the potential therapeutic avenues that can be explored to reduce the risk and burden of severe COVID-19. We also provide a web app where users can visualise the temporal changes of specific genes or MDSC-related gene sets during severe COVID-19 progression and disease resolution at https://temporal-severe-covid.streamlit.app/, based on our previous study described by Ong et al., 2021.
Keywords:
SARS-CoV-2
; COVID-19
; myeloid-derived suppressor cells
; infection
; innate immunity
; T cells
1. Introduction
The COVID-19 pandemic is caused by SARS-CoV-2 infections, which has led to over 670 million cases and approximately 6.8 million deaths worldwide [1]. The virus genome encodes for 4 major structural proteins (nucleocapsid (N), spike (S), envelope (E) and membrane (M) proteins), 16 non-structural proteins, as well as, a number of accessory proteins [2]. SARS-CoV-2 S protein comprises 2 subunits: S1, which is responsible for binding to the angiotensin-converting enzyme 2 (ACE2) and S2, which facilitates membrane fusion between virus and the membrane of the target cell. SARS-CoV-2 infections are often asymptomatic, although symptomatic infections can lead to a broad range of clinical outcomes, which can range from mild to severe outcomes. The elderly and the immunocompromised [3] are at greatest risk of severe COVID-19, which is characterised by hyperinflammation and cytokine storm, where markedly elevated levels of circulating proinflammatory cytokines can potentially lead to multi-organ damage and acute respiratory distress syndrome (ARDS). Interestingly, based on our meta-analysis of RNAseq data across 7 independent cohorts, time-series analysis and single-cell RNA sequencing datasets, we previously demonstrated that the differential expression of MCEMP1 and HLA-DRA in CD14-positive cells could be detected as early as 4 days before nadir of respiratory function. As the upregulation of MCEMP1 and downregulation of HLA-DRA is characteristic of myeloid-derived suppressor cells (MDSCs), the results indicated that an early expansion of MDSCs could be involved in severe COVID-19 progression and pathogenesis [4]. Besides our study, emerging studies have also documented the contribution of MDSCs in severe COVID-19 pathogenesis. Therefore, besides serving as an early prognostic biomarker for severe COVID-19, MDSCs could potentially be targeted to reduce the burden of the disease. In light of these studies, we examined the current literature documenting the potential functional roles of MDSCs in severe COVID-19 and long-COVID pathogenesis, discuss strategies that can be employed to detect MDSCs and explore potential therapeutic avenues to target MDSCs reduce the risk of severe COVID-19.
2. MDSC development and functions
MDSCs was a term first introduced by Gabrilovich, D. I. et al. in 2007 [5] as an effort to describe a heterogeneous group of immature myeloid cells possessing potent immunosuppressive activities that are implicated in cancer pathology. MDSCs can be broadly subdivided into two main groups: granulocytic/polymorphonuclear MDSCs (PMN-MDSCs) and monocytic MDSCs (M-MDSCs) based on their lineage association. The third subgroup, termed as early MDSCs, make up the minority of MDSCs and are primarily comprised of myeloid precursors and progenitors.
MDSCs arise from hematopoietic stem cells (HSC) via granulo-monocytic precursors (GMP). GMP differentiate from the common myeloid progenitor (CMP). A two-signal model [6] is widely accepted for the development of MDSCs. The first signal involves the expansion of GMP by G-CSF, GM-CSF, M-CSF and transcriptional regulators such as IRF8, STAT3, STAT5, NOTCH and C/EBP-β [7]. This initial signal is thought to allow expansion of immature myeloid cells while preventing terminal differentiation [7,8,9]. Thereafter, the second signal involves cues from the extracellular environment such as damage associated molecular patterns (DAMP), S100A8/A9, and proinflammatory cytokines (e.g., IL-1β, tumor necrosis factor, interferon-γ (IFN-γ) , IL-4, IL-6, IL-13) in the peripheral tissues, which has been proposed to activate immature myeloid cells to gain immunosuppressive capabilities.
MDSC development occurs through the same differentiation pathways as neutrophils and monocytes [9]. Under normal physiological conditions, myelopoiesis is driven by GM-CSF while differentiation to granulocytes and macrophages is induced by G-CSF and M-CSF, respectively. Notably, while the differentiation to MDSCs, neutrophils and monocytes require GM-CSF, M- and G-CSF, the requirement for these factors are higher for MDSCs compared to neutrophils and monocytes. Thus, MDSCs usually do not accumulate under normal physiological conditions [10]. However, under cancer, chronic infections and other pathological conditions, normal myelopoiesis may be perturbed [7,8,9], as these conditions can provide the prolonged low-strength signals to persistently stimulate the myeloid compartment through the excessive production of signals such as GM-CSF, IL-6, IL-8, TNF-α, IL-1β, S100A8/9 and reactive oxygen species (ROS) [11] that are required for MDSC development. Emerging studies also support that these signals are greatly induced after infections by viruses such as SARS-CoV-2 [12], HIV [13], and hepatitis B virus [14], which can aid MDSC development. However, the underlying mechanisms for MDSC generation may differ in the different pathological conditions [15].
MDSCs are capable of suppressing various immune cell types such as B, NK and in particular, T cells [16]. MDSCs can exert their immunosuppressive activities [7,8,9,17] through depletion of arginine, tryptophan, and cystine/cysteine availability, induction of Treg cells, RNS/ROS, induction of T cell apoptosis via Fas/FasL interaction, production of inhibitory cytokines and PD-1/PD-L1 interaction [18] (Figure 1). Interestingly, many of these mechanisms involve immunometabolic regulations. For instance, arginine depletion by MDSC can be catalyzed by arginase-1 (Arg-1), which catabolizes arginine into L-ornithine and urea, reducing the availability of arginine and compromising T cell proliferation and function by reducing the CD3ζ chain [19]. In addition, MDSCs can deplete tryptophan availability for T cells by upregulating indoleamine 2,3-dioxygenase, catabolizing tryptophan to N-formylkynurenine, resulting in inhibition of effector T cell proliferation, while favouring the thriving of Foxp3+ Treg. Cysteine, another amino acid important to T cells, is also known to be sequestered by MDSCs by expressing SLC7A11 which helps in the uptake of cystine into the cell [20]. Because MDSCs do not express the alanine-serine-cysteine (ASC) transporter, the return of cysteine extracellularly is prevented [20].
Besides immunometabolic regulation, MDSCs can generate ROS to mediate immunosuppression [11]. The elevated production of superoxides can react with nitric oxide to form peroxynitrite, which leads to nitration of TCR-CD8, which has been shown to hamper the ability of CD8 T cells to bind and respond to peptide-major histocompatibility complex (pMHC) [21]. In addition, MDSC-like cells have been reported to release H2O2 to inhibit T cell activation and rely on superoxide generation to suppress T cell responses in a NOX2-dependent manner. ROS are also important mediators employed by MDSC for immunosuppression of NK and B cells [11]. Within MDSCs, some of these immunosuppressive strategies are preferentially used by PMN-MDSC subset (arginase-1, ROS, peroxynitrite) while others are used by M-MDSCs subset (nitric oxide, PD-L1 and immunosuppressive cytokines) [8].
3. MDSC identification and characterisation
While definitive identifiers for MDSCs are lacking, several combinations of cell markers have been suggested, namely CD11b+CD14−CD15+ or CD11b+CD14−CD66b+ for characterizing PMN-MDSCs and CD11b+CD14+HLA-DR−/loCD15− for the characterization of M-MDSCs in human peripheral blood mononuclear cells [22]. M-MDSCs can be distinguished from monocytes by detecting for low or undetectable HLA-DR protein expression. However, discriminating between PMN-MDSCs and neutrophils is more challenging, as the phenotypic markers and functions are highly overlapping. For instance, activated neutrophils can also secrete arginase-1 (Arg-1) and employ reactive oxygen species (ROS) to suppress T cell activity, akin to the function of PMN-MDSCs [23,24]. While more research will be required to better distinguish the PMN-MDSCs and neutrophils, a possible approach could be to separate them by density, where PMN-MDSC appear to be of a lower density compared to neutrophils [8,22].
Nonetheless, with the advent of single cell RNA sequencing, the PMN-MDSCs and M-MDSCs can be potentially characterised by transcriptomic signatures (Table 1). Although phenotypic cell markers specific for MDSCs are lacking, MDSCs have been found to have transcriptional profiles that differ from classical neutrophils or monocytes. M-MDSCs express genes involved in proinflammatory and immunosuppression, such as NOS2, IL10, TNF, WFDC17, STAT3, S100A8/9, IL6, ARG1/2, CD84 and TGFB1, VEGFA. On the other hand, gene signatures for PMN-MDSC include IL1B, IL4R, STAT1, STAT3, STAT6, OLR1, ARG1/2, PTGS2, S100A8/9, IRF1, LYZ2, TREM1, CXCL1, LILRA3, CSF1, IL6, ANXA1, which allows them to distinguished from classical neutrophils.
In both M- and PMN-MDSCs, STAT3 is upregulated, and is important for expansion while preventing further differentiation to other cell types. For instance, it has been demonstrated that inhibition of STAT3 expression abrogated MDSC expansion induced by tumour-derived factors [25]. Furthermore, binding of MDSC-promoting signals such as IL-6, G-CSF, GM-CSF and M-CSF to their respective cellular receptor activates JAK-STAT3 signalling can facilitate MDSC accumulation and expansion [26]. Activated (phosphorylated) STAT3 form homo or heterodimers that can subsequently translocate to the nucleus and modulate gene expression (e.g., downregulate IRF8 expression and promote S100A8/9 expression required for MDSC expansion and preventing their differentiation (reviewed by [27]). In addition to IRF8 downregulation, MDSC also exhibit increased expression of C/EBP-β (a member of the family of basic region leucine zipper transcriptional factors) which favours MDSC generation. IRF8 is a negative regulator of MDSC expansion while C/EBP-β, a master regulator of emergency myelopoiesis, also regulates arginase-1 and inducible nitric oxide synthase (iNOS) important for MDSC to exert its immunosuppressive effects [9]. Other gene signatures such as WFDC17, ARG2, IL1B and CD84 have also been identified to be overlapping between M- and PMN-MDSCs [7,8].
Table 1 summarises the gene sets that have been used by others to characterize MDSC. In addition, we annotated the transcripts (in bold) that were temporally regulated during severe COVID-19 progression and recovery, based on our study where temporal transcriptomic profiling was performed on 6 severe COVID-19 patients [28] (Table 1). As expected, majority of the genes characteristic of M-MDSC and PMN-MDSC were also differentially regulated during severe COVID-19 pathogenesis (Table 1). To allow users to query the temporal changes of genes involved in severe COVID-19 pathogenesis, we also created a web tool (https://temporal-severe-covid.streamlit.app/) where users can input a list of genes or a list of MDSC gene sets from Table 1 to plot the normalised expression values across different timepoints before and after respiratory nadir (Figure 2). Users can also run the web tool locally using the codes provided in GitHub (https://github.com/kuanrongchan/temporal_severe_covid).
4. Implications of MDSCs in COVID-19 pathogenesis
SARS-CoV-2, as with other viral infections, perturbs normal haematopoiesis, especially in severe COVID-19 patients [34]. Besides demonstrating that the early upregulation of MCEMP1 and downregulation of HLA-DRA expression occurred at least 4 days before respiratory of nadir function in severe COVID-19 patients [4], emerging studies have also shown increased accumulation of cells sharing features of M- [31] and PMN-MDSCs [33,35,36,37,38] in severe COVID-19 patients, suggesting the importance of these dysregulated myeloid cells in SARS-CoV-2 progression and pathogenesis [37]. The levels of M-MDSCs were detected more abundantly in the blood of severe COVID-19 patients rather than the nasopharyngeal and endotracheal aspirates [39], which is consistent with the findings showing that the MDSC gene signatures were detected more strongly in CD14-positive cells of the peripheral blood compared to the immune cells of the bronchoalveolar lavage fluid [4]. Since the MDSC signatures are prominent in peripheral blood of severe COVID-19 patients, several other groups have also explored the prognostic potential of MDSCs. For instance, in a study conducted on 41 COVID-19 patients who have yet to progress to peak severity, circulating M-MDSC levels within two weeks of symptom onset was able to predict disease severity [39]. In another study, CPT1a+VDAC1+HLA-DR- M-MDSC frequencies were found to be significantly higher in severe compared to mild COVID-19 patients [40], and the differences could be seen before the severe COVID-19 patients experience the respiratory nadir [4]. Other studies have also reported that M-MDSC levels were correlated with nasopharyngeal SARS-CoV-2 viral load [41] and associated with secondary infections in severe COVID-19 patients and mortality [42]. Interestingly, survivors from severe COVID-19 have lower PMN-MDSCs in their blood as compared to the deceased group [43] which is in agreement with other evidences showing the correlation of higher PMN-MDSC frequency with increased mortality/disease fatality [44].
Besides showing association of MDSC frequencies with disease severity, accumulating evidence now suggests that COVID-19 patient-derived MDSCs can exhibit immunosuppressive activities on T cells [44,45]. Specifically, M-MDSCs derived from COVID-19 patients have been shown to suppress proliferation and inhibit release of IFN-γ by T cells in an Arg-1-dependent manner [39]. In addition, ROS levels and the capacity of neutrophils to generate ROS is also significantly associated with disease severity [46,47], indicating the likely contribution of ROS in T cell suppression during severe COVID-19. Interestingly, in another study that performed co-culture of patient-derived MDSCs with autologous MDSC-depleted PBMCs, it was found that MDSCs can promote the expansion of Treg precursors while suppressing mature Treg proliferation [48]. Thus, T cell suppression could also be mediated by regulating Treg frequencies, which can consequently influence outcome of SARS-CoV-2 infection [49]. Given that the early induction of functional SARS-CoV-2 specific T cell responses correlates with rapid viral clearance and mild disease in COVID-19 patients [50], these studies collectively indicate that the early expansion of MDSCs serve as a pro-viral strategy to evade early T cell detection.
5. Role of inflammatory responses to MDSC development in severe COVID-19
Besides the contribution of arginine depletion, ROS generation and Treg in MDSC development and T cell suppression, increasing studies have also highlighted the role of inflammatory responses in shaping MDSC development. The viral proteins, such as the S protein, has been shown to induce release of proinflammatory cytokines in human macrophages and lung epithelial cells via TLR2-NF-κB pathway [51]. Other viral components such as E protein [52], N [53], ORF7a [54], has also been shown to induce production of inflammatory cytokines through activation of the inflammasome and NF-κB pathway [55,56]. Incubation of haematopoietic stem and progenitor cells (HSPC) with plasma from severe COVID-19 patients induced MDSC-like cells [29], indicating that the pro-inflammatory responses triggered by infection could directly influence myeloid cell development. Among the different pro-inflammatory responses triggered by SARS-CoV-2 infection, IL-6 is most well-known to be consistently elevated in the blood of severe COVID-19 patients [36,57,58], and has been shown to be able to induce HSPC [29] and human primary monocytes to become MDSC-like cells [59]. In other cases, blood levels of GM-CSF [60] and M-CSF [61] were significantly heightened in severe COVID-19 patients and combinatorial treatment of HSPC with IL-6, GM-CSF and M-CSF enhanced the generation of MDSC-like cells [59]. Other cytokines such as IL-8 [35,62], TNF-α, IL-1β were also observed to be markedly increased in the blood of severe COVID-19 patients [57] and found to be significantly correlated with MDSC levels, possibly recruiting these cells to peripheral blood.
In addition to viral proteins, the production of high levels of anti-spike IgG or afucosylated anti-spike IgG after SARS-CoV-2 infection, can trigger heightened proinflammatory response [63] in human alveolar macrophages, which can promote the release of cytokines such as IL-6 and TNF via the IFN-signalling pathways [64]. The addition of the Syk inhibitor, R406, was able to ameliorating the release of proinflammatory cytokines, highlighting the role of Syk-mediated signalling in inducing pro-inflammatory responses following SARS-CoV-2 infection. Furthermore, the ability of R406 to impede production of proinflammatory cytokines raises the question whether R406 may also impact MDSC development, as pro-inflammatory responses are known to promote MDSC development. Indeed, a separate study demonstrated that R406 can restore myeloid homeostasis by reducing PMN-MDSC levels, increasing monocytes with HLA-DRhi phenotype and inhibiting expression of antiviral genes [65]. However, more studies will need to be performed to establish the link between SARS-CoV-2 and Syk-signalling, and their interaction with MDSC development and severe disease outcome. The summary of the mechanisms involved are detailed in Figure 3.
6. Long-term changes in MDSCs after severe COVID-19 resolution
Given the short lifespan of MDSCs [8] and clearance of SARS-CoV-2 and inflammation after disease resolution, most individuals are expected to experience decline and normalisation of MDSC levels [66]. However, intriguingly, there have been studies reporting persisting or elevated levels of MDSC and myeloid progenitor cells [67,68,69,70,71], even in asymptomatic convalescents [72]. These observations may thus suggest that there may be epigenetic reprogramming in the myeloid cells that allow MDSC persistence in recovered severe COVID-19 patients. Indeed, in a study by Cheong and colleagues [67], ATAC-seq and single-nuclei RNA sequencing collectively uncovered that monocytes and HSPCs from severe COVID-19-recovered patients exhibited phenotype and the associated transcriptional programs relating to inflammation and myeloid activation. Notably, the increased myelopoiesis and epigenetic alterations can be observed as long as 12 months following acute infection [67]. While the precise mechanisms remain to be fully elucidated, these effects appear to be modulated by IL-6 as the effects were abrogated by IL-6 blockade [67]. Thus, in addition to suppressing hyper-inflammation responses, IL-6 inhibitors such as tocilizumab may be useful to restore myeloid dysregulation. Consistent with this hypothesis, a clinical study over 300 participants revealed that tocilizumab treatment was able to restore myeloid dysregulation and ameliorate inflammation as compared to the placebo patient group [32]. However, it is important to note that in another study involving 10 severe COVID-19 patients, tocilizumab did not affect MDSC [73], which may reflect that other factors may be crucial in governing MDSC levels. These may include other key regulators such as HIF1α [74] and PGE2 [75,76], which are known to be upregulated in severe COVID-19 patients.
As MDSC levels can persist following severe COVID-19 recovery, it is possible that MDSC may also contribute to long-COVID [77], where patients experience a myriad of lingering symptoms following recovery. Indeed, increased levels of immunoregulatory immune cells such as Treg cells [78] and MDSCs [69,70,72,79] have been observed in some long-COVID or COVID-19 recovered patients, and may last for as long as one year following recovery from SARS-CoV-2 infection [68]. Notably, some COVID-19 convalescents possess elevated blood levels of IL-6 [69], although the significance of these findings in MDSC development and long-COVID has not been completely resolved [73].
7. Potential therapeutic drugs for ameliorating MDSC levels in COVID-19 patients
Given the implications of MDSC in severe COVID-19, targeting MDSCs may be a rational option for COVID-19 and possibly other infectious diseases that rely on MDSC to facilitate pathogenesis. Induction of MDSC apoptosis and elimination of stimulus known to favour MDSC generation [80] are some of the proposed ways to limit MDSC effects on severe COVID-19 progression and pathogenesis. Some of these drugs have been shown by ex vivo or in vitro studies to abolish the excessive proinflammatory cytokine production or the increase in MDSC levels to noteworthy extents. For instance, direct treatment of monocytic/T cell co-culture with 5-fluorouracil can lead to lowered levels of cytokines such as IL-8 in the culture supernatant [73]. Removal or inhibition of stimuli known to favour MDSC generation may also be a potential therapeutic option. As S100A8/A9 is a stimuli favouring MDSC generation and is increased following SARS-CoV-2 infection, an inhibitor of S100A8/A9, paquinimod, has been proposed to reduce viral load and aberrant immature neutrophils in mouse models [81]. Finally, as MDSCs can exert its immunosuppressive effects through deprivation of amino acids important for T cell survival and function, the action of MDSCs may potentially be perturbed by immunometabolic regulation such as amino acid supplementation. For example, supplementation of amino acids such as L-arginine have been shown to be clinically beneficial for severe COVID-19 patients by reducing length of hospitalization and requirement for respiratory support [82]. Future studies that clarify whether these therapeutic strategies would be effective in reducing severe COVID-19 and long COVID burden will shed valuable insights on the contribution of MDSCs in severe COVID-19 pathogenesis.
8. Conclusion
The mounting evidence from multiple research groups have provided valuable hints on the mechanism of early MDSC expansion in severe COVID-19 patients, and support the role of MDSC in suppressing antiviral and T cell immune responses against SARS-CoV-2. However, several question remains unanswered, for instance, the underlying mechanisms involved in the persistence of MDSC in severe COVID-19 patients after recovery. As emphasized by Hegde et al. [7], it is also important that future efforts should emphasise on better characterisation of MDSC states, to allow easier detection of MDSC for early prognosis of severe COVID-19 and to enable a more in-depth understanding of the roles of MDSC in severe COVID-19 pathogenesis.
Supplementary Materials
Not applicable
Author Contributions
Conceptualization, J.S.L. and K.R.C.; software, C.W.T.; writing—original draft preparation, J.S.L.; writing—review and editing, J.S.L. and K.R.C.; supervision, K.R.C.; funding acquisition, K.R.C. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by the National Medical Research Council of Singapore (OF-IRG) Grant, grant number MOH-000610-00.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| MDSC | Myeloid derived suppressor cells |
| ACE2 | Angiotensin-converting enzyme 2 |
| Arg-1 | Arginase-1 |
| ASC | Alanine-serine-cysteine |
| CMP | Common myeloid progenitor |
| CSF | Colony-stimulating factor |
| DAMP | Damage associated molecular pattern |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| H2O2 | Hydrogen peroxide |
| HSC | Hematopoietic stem cells |
| LOX1 | Lectin-type oxidized LDL receptor 1 |
| M-CSF | Macrophage colony-stimulating factor |
| M-MDSC | Monocytic MDSC |
| PD-1 | Programmed cell death protein 1 |
| PD-L1 | Programmed death ligand 1 |
| PMN-MDSC | Polymorphonuclear mdscs |
| ROS | Reactive oxygen species |
| SLC7A11 | Solute carrier family 7 member 11 |
| Syk | Spleen tyrosine kinase |
| Treg | Regulatory T cells |
References
- Johns Hopkins University COVID-19 Map - Johns Hopkins Coronavirus Resource Center Available online: https://coronavirus.jhu.edu/map.html (accessed on 6 October 2023).
- Li, G.; Hilgenfeld, R.; Whitley, R.; De Clercq, E. Therapeutic Strategies for COVID-19: Progress and Lessons Learned. Nature Reviews Drug Discovery 2023, 22, 449–475. [CrossRef]
- Ioannidis, J.P.A. Global Perspective of COVID-19 Epidemiology for a Full-Cycle Pandemic. European Journal of Clinical Investigation 2020, 50, e13423. [CrossRef]
- Chan, K.R.; Koh, C.W.T.; Ng, D.H.L.; Qin, S.; Ooi, J.S.G.; Ong, E.Z.; Zhang, S.L.X.; Sam, H.; Kalimuddin, S.; Low, J.G.H.; et al. Early Peripheral Blood MCEMP1 and HLA-DRA Expression Predicts COVID-19 Prognosis. eBioMedicine 2023, 89. [CrossRef]
- Gabrilovich, D.I.; Bronte, V.; Chen, S.-H.; Colombo, M.P.; Ochoa, A.; Ostrand-Rosenberg, S.; Schreiber, H. The Terminology Issue for Myeloid-Derived Suppressor Cells. Cancer Research 2007, 67, 425–425. [CrossRef]
- Condamine, T.; Gabrilovich, D.I. Molecular Mechanisms Regulating Myeloid-Derived Suppressor Cell Differentiation and Function. Trends in Immunology 2011, 32, 19–25. [CrossRef]
- Hegde, S.; Leader, A.M.; Merad, M. MDSC: Markers, Development, States, and Unaddressed Complexity. Immunity 2021, 54, 875–884. [CrossRef]
- Veglia, F.; Sanseviero, E.; Gabrilovich, D.I. Myeloid-Derived Suppressor Cells in the Era of Increasing Myeloid Cell Diversity. Nature Reviews Immunology 2021, 21, 485–498. [CrossRef]
- Veglia, F.; Perego, M.; Gabrilovich, D. Myeloid-Derived Suppressor Cells Coming of Age. Nature Immunology 2018, 19, 108–119. [CrossRef]
- Ushach, I.; Zlotnik, A. Biological Role of Granulocyte Macrophage Colony-Stimulating Factor (GM-CSF) and Macrophage Colony-Stimulating Factor (M-CSF) on Cells of the Myeloid Lineage. Journal of Leukocyte Biology 2016, 100, 481–489. [CrossRef]
- Ohl, K.; Tenbrock, K. Reactive Oxygen Species as Regulators of MDSC-Mediated Immune Suppression. Frontiers in Immunology 2018, 9.
- Bordoni, V.; Sacchi, A.; Cimini, E.; Notari, S.; Grassi, G.; Tartaglia, E.; Casetti, R.; Giancola, M.L.; Bevilacqua, N.; Maeurer, M.; et al. An Inflammatory Profile Correlates With Decreased Frequency of Cytotoxic Cells in Coronavirus Disease 2019. Clinical Infectious Diseases 2020, 71, 2272–2275. [CrossRef]
- Garg, A.; Spector, S.A. HIV Type 1 Gp120–Induced Expansion of Myeloid Derived Suppressor Cells Is Dependent on Interleukin 6 and Suppresses Immunity. The Journal of Infectious Diseases 2014, 209, 441–451. [CrossRef]
- Yang, F.; Yu, X.; Zhou, C.; Mao, R.; Zhu, M.; Zhu, H.; Ma, Z.; Mitra, B.; Zhao, G.; Huang, Y.; et al. Hepatitis B e Antigen Induces the Expansion of Monocytic Myeloid-Derived Suppressor Cells to Dampen T-Cell Function in Chronic Hepatitis B Virus Infection. PLOS Pathogens 2019, 15, e1007690. [CrossRef]
- Tcyganov, E.N.; Hanabuchi, S.; Hashimoto, A.; Campbell, D.; Kar, G.; Slidel, T.W.F.; Cayatte, C.; Landry, A.; Pilataxi, F.; Hayes, S.; et al. Distinct Mechanisms Govern Populations of Myeloid-Derived Suppressor Cells in Chronic Viral Infection and Cancer. J Clin Invest 2021, 131. [CrossRef]
- Gabrilovich, D.I. Myeloid-Derived Suppressor Cells. Cancer Immunology Research 2017, 5, 3–8. [CrossRef]
- Ostrand-Rosenberg, S.; Lamb, T.J.; Pawelec, G. Here, There, and Everywhere: Myeloid-Derived Suppressor Cells in Immunology. The Journal of Immunology 2023, 210, 1183–1197. [CrossRef]
- Ballbach, M.; Dannert, A.; Singh, A.; Siegmund, D.M.; Handgretinger, R.; Piali, L.; Rieber, N.; Hartl, D. Expression of Checkpoint Molecules on Myeloid-Derived Suppressor Cells. Immunology Letters 2017, 192, 1–6. [CrossRef]
- Dean, M.J.; Ochoa, J.B.; Sanchez-Pino, M.D.; Zabaleta, J.; Garai, J.; Del Valle, L.; Wyczechowska, D.; Baiamonte, L.B.; Philbrook, P.; Majumder, R.; et al. Severe COVID-19 Is Characterized by an Impaired Type I Interferon Response and Elevated Levels of Arginase Producing Granulocytic Myeloid Derived Suppressor Cells. Frontiers in Immunology 2021, 12.
- Srivastava, M.K.; Sinha, P.; Clements, V.K.; Rodriguez, P.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells Inhibit T-Cell Activation by Depleting Cystine and Cysteine. Cancer Research 2010, 70, 68–77. [CrossRef]
- Nagaraj, S.; Gupta, K.; Pisarev, V.; Kinarsky, L.; Sherman, S.; Kang, L.; Herber, D.L.; Schneck, J.; Gabrilovich, D.I. Altered Recognition of Antigen Is a Mechanism of CD8+ T Cell Tolerance in Cancer. Nature Medicine 2007, 13, 828–835. [CrossRef]
- Bronte, V.; Brandau, S.; Chen, S.-H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for Myeloid-Derived Suppressor Cell Nomenclature and Characterization Standards. Nature Communications 2016, 7, 12150. [CrossRef]
- Aarts, C.E.M.; Hiemstra, I.H.; Béguin, E.P.; Hoogendijk, A.J.; Bouchmal, S.; van Houdt, M.; Tool, A.T.J.; Mul, E.; Jansen, M.H.; Janssen, H.; et al. Activated Neutrophils Exert Myeloid-Derived Suppressor Cell Activity Damaging T Cells beyond Repair. Blood Advances 2019, 3, 3562–3574. [CrossRef]
- Pillay, J.; Tak, T.; Kamp, V.M.; Koenderman, L. Immune Suppression by Neutrophils and Granulocytic Myeloid-Derived Suppressor Cells: Similarities and Differences. Cellular and Molecular Life Sciences 2013, 70, 3813–3827. [CrossRef]
- Nefedova, Y.; Nagaraj, S.; Rosenbauer, A.; Muro-Cacho, C.; Sebti, S.M.; Gabrilovich, D.I. Regulation of Dendritic Cell Differentiation and Antitumor Immune Response in Cancer by Pharmacologic-Selective Inhibition of the Janus-Activated Kinase 2/Signal Transducers and Activators of Transcription 3 Pathway. Cancer Research 2005, 65, 9525–9535. [CrossRef]
- Li, K.; Shi, H.; Zhang, B.; Ou, X.; Ma, Q.; Chen, Y.; Shu, P.; Li, D.; Wang, Y. Myeloid-Derived Suppressor Cells as Immunosuppressive Regulators and Therapeutic Targets in Cancer. Signal Transduction and Targeted Therapy 2021, 6, 362. [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-Derived Suppressor Cells as Regulators of the Immune System. Nature Reviews Immunology 2009, 9, 162–174. [CrossRef]
- Ong, E.Z.; Kalimuddin, S.; Chia, W.C.; Ooi, S.H.; Koh, C.W.; Tan, H.C.; Zhang, S.L.; Low, J.G.; Ooi, E.E.; Chan, K.R. Temporal Dynamics of the Host Molecular Responses Underlying Severe COVID-19 Progression and Disease Resolution. eBioMedicine 2021, 65. [CrossRef]
- Reyes, M.; Filbin, M.R.; Bhattacharyya, R.P.; Sonny, A.; Mehta, A.; Billman, K.; Kays, K.R.; Pinilla-Vera, M.; Benson, M.E.; Cosimi, L.A.; et al. Plasma from Patients with Bacterial Sepsis or Severe COVID-19 Induces Suppressive Myeloid Cell Production from Hematopoietic Progenitors in Vitro. Science Translational Medicine 2021, 13, eabe9599. [CrossRef]
- Wilk, A.J.; Lee, M.J.; Wei, B.; Parks, B.; Pi, R.; Martínez-Colón, G.J.; Ranganath, T.; Zhao, N.Q.; Taylor, S.; Becker, W.; et al. Multi-Omic Profiling Reveals Widespread Dysregulation of Innate Immunity and Hematopoiesis in COVID-19. Journal of Experimental Medicine 2021, 218, e20210582. [CrossRef]
- Kvedaraite, E.; Hertwig, L.; Sinha, I.; Ponzetta, A.; Hed Myrberg, I.; Lourda, M.; Dzidic, M.; Akber, M.; Klingström, J.; Folkesson, E.; et al. Major Alterations in the Mononuclear Phagocyte Landscape Associated with COVID-19 Severity. Proceedings of the National Academy of Sciences 2021, 118, e2018587118. [CrossRef]
- Shivram, H.; Hackney, J.A.; Rosenberger, C.M.; Teterina, A.; Qamra, A.; Onabajo, O.; McBride, J.; Cai, F.; Bao, M.; Tsai, L.; et al. Transcriptomic and Proteomic Assessment of Tocilizumab Response in a Randomized Controlled Trial of Patients Hospitalized with COVID-19. iScience 2023, 26. [CrossRef]
- LaSalle, T.J.; Gonye, A.L.K.; Freeman, S.S.; Kaplonek, P.; Gushterova, I.; Kays, K.R.; Manakongtreecheep, K.; Tantivit, J.; Rojas-Lopez, M.; Russo, B.C.; et al. Longitudinal Characterization of Circulating Neutrophils Uncovers Phenotypes Associated with Severity in Hospitalized COVID-19 Patients. Cell Reports Medicine 2022, 3, 100779. [CrossRef]
- Wang, X.; Wen, Y.; Xie, X.; Liu, Y.; Tan, X.; Cai, Q.; Zhang, Y.; Cheng, L.; Xu, G.; Zhang, S.; et al. Dysregulated Hematopoiesis in Bone Marrow Marks Severe COVID-19. Cell Discovery 2021, 7, 60. [CrossRef]
- Takano, T.; Matsumura, T.; Adachi, Y.; Terahara, K.; Moriyama, S.; Onodera, T.; Nishiyama, A.; Kawana-Tachikawa, A.; Miki, S.; Hosoya-Nakayama, K.; et al. Myeloid Cell Dynamics Correlating with Clinical Outcomes of Severe COVID-19 in Japan. International Immunology 2021, 33, 241–247. [CrossRef]
- Silvin, A.; Chapuis, N.; Dunsmore, G.; Goubet, A.-G.; Dubuisson, A.; Derosa, L.; Almire, C.; Hénon, C.; Kosmider, O.; Droin, N.; et al. Elevated Calprotectin and Abnormal Myeloid Cell Subsets Discriminate Severe from Mild COVID-19. Cell 2020, 182, 1401-1418.e18. [CrossRef]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419-1440.e23. [CrossRef]
- Agrati, C.; Sacchi, A.; Bordoni, V.; Cimini, E.; Notari, S.; Grassi, G.; Casetti, R.; Tartaglia, E.; Lalle, E.; D’Abramo, A.; et al. Expansion of Myeloid-Derived Suppressor Cells in Patients with Severe Coronavirus Disease (COVID-19). Cell Death & Differentiation 2020, 27, 3196–3207. [CrossRef]
- Falck-Jones, S.; Vangeti, S.; Yu, M.; Falck-Jones, R.; Cagigi, A.; Badolati, I.; Österberg, B.; Lautenbach, M.J.; Åhlberg, E.; Lin, A.; et al. Functional Monocytic Myeloid-Derived Suppressor Cells Increase in Blood but Not Airways and Predict COVID-19 Severity. J Clin Invest 2021, 131. [CrossRef]
- Thompson, E.A.; Cascino, K.; Ordonez, A.A.; Zhou, W.; Vaghasia, A.; Hamacher-Brady, A.; Brady, N.R.; Sun, I.-H.; Wang, R.; Rosenberg, A.Z.; et al. Metabolic Programs Define Dysfunctional Immune Responses in Severe COVID-19 Patients. Cell Reports 2021, 34, 108863. [CrossRef]
- Xue, G.; Jiang, M.; Zhao, R.; Le, A.; Li, J. Elevated Frequencies of CD14+HLA-DRlo/neg MDSCs in COVID-19 Patients. Aging (Albany NY) 2021, 13, 6236–6246. [CrossRef]
- Marais, C.; Claude, C.; Semaan, N.; Charbel, R.; Barreault, S.; Travert, B.; Piloquet, J.-E.; Demailly, Z.; Morin, L.; Merchaoui, Z.; et al. Myeloid Phenotypes in Severe COVID-19 Predict Secondary Infection and Mortality: A Pilot Study. Annals of Intensive Care 2021, 11, 111. [CrossRef]
- Jiménez-Cortegana, C.; Sánchez-Jiménez, F.; Pérez-Pérez, A.; Álvarez, N.; Sousa, A.; Cantón-Bulnes, L.; Vilariño-García, T.; Fuentes, S.; Martín, S.; Jiménez, M.; et al. Low Levels of Granulocytic Myeloid-Derived Suppressor Cells May Be a Good Marker of Survival in the Follow-Up of Patients With Severe COVID-19. Frontiers in Immunology 2022, 12.
- Sacchi, A.; Grassi, G.; Bordoni, V.; Lorenzini, P.; Cimini, E.; Casetti, R.; Tartaglia, E.; Marchioni, L.; Petrosillo, N.; Palmieri, F.; et al. Early Expansion of Myeloid-Derived Suppressor Cells Inhibits SARS-CoV-2 Specific T-Cell Response and May Predict Fatal COVID-19 Outcome. Cell Death & Disease 2020, 11, 921. [CrossRef]
- Xu, G.; Qi, F.; Li, H.; Yang, Q.; Wang, H.; Wang, X.; Liu, X.; Zhao, J.; Liao, X.; Liu, Y.; et al. The Differential Immune Responses to COVID-19 in Peripheral and Lung Revealed by Single-Cell RNA Sequencing. Cell Discovery 2020, 6, 73. [CrossRef]
- Veenith, T.; Martin, H.; Le Breuilly, M.; Whitehouse, T.; Gao-Smith, F.; Duggal, N.; Lord, J.M.; Mian, R.; Sarphie, D.; Moss, P. High Generation of Reactive Oxygen Species from Neutrophils in Patients with Severe COVID-19. Scientific Reports 2022, 12, 10484. [CrossRef]
- Badawy, M.A.; Yasseen, B.A.; El-Messiery, R.M.; Abdel-Rahman, E.A.; Elkhodiry, A.A.; Kamel, A.G.; El-sayed, H.; Shedra, A.M.; Hamdy, R.; Zidan, M.; et al. Neutrophil-Mediated Oxidative Stress and Albumin Structural Damage Predict COVID-19-Associated Mortality. eLife 2021, 10, e69417. [CrossRef]
- Kiaee, F.; Jamaati, H.; Shahi, H.; Roofchayee, N.D.; Varahram, M.; Folkerts, G.; Garssen, J.; Adcock, I.M.; Mortaz, E. Immunophenotype and Function of Circulating Myeloid Derived Suppressor Cells in COVID-19 Patients. Scientific Reports 2022, 12, 22570. [CrossRef]
- Jiménez-Cortegana, C.; Liró, J.; Palazón-Carrión, N.; Salamanca, E.; Sojo-Dorado, J.; de la Cruz-Merino, L.; Pascual, Á.; Rodríguez-Baño, J.; Sánchez-Margalet, V. Increased Blood Monocytic Myeloid Derived Suppressor Cells but Low Regulatory T Lymphocytes in Patients with Mild COVID-19. Viral Immunology 2021, 34, 639–645. [CrossRef]
- Tan, A.T.; Linster, M.; Tan, C.W.; Le Bert, N.; Chia, W.N.; Kunasegaran, K.; Zhuang, Y.; Tham, C.Y.L.; Chia, A.; Smith, G.J.D.; et al. Early Induction of Functional SARS-CoV-2-Specific T Cells Associates with Rapid Viral Clearance and Mild Disease in COVID-19 Patients. Cell Reports 2021, 34, 108728. [CrossRef]
- Khan, S.; Shafiei, M.S.; Longoria, C.; Schoggins, J.W.; Savani, R.C.; Zaki, H. SARS-CoV-2 Spike Protein Induces Inflammation via TLR2-Dependent Activation of the NF-κB Pathway. eLife 2021, 10, e68563. [CrossRef]
- DeDiego Marta L.; Nieto-Torres Jose L.; Regla-Nava Jose A.; Jimenez-Guardeño Jose M.; Fernandez-Delgado Raul; Fett Craig; Castaño-Rodriguez Carlos; Perlman Stanley; Enjuanes Luis Inhibition of NF-κB-Mediated Inflammation in Severe Acute Respiratory Syndrome Coronavirus-Infected Mice Increases Survival. Journal of Virology 2014, 88, 913–924. [CrossRef]
- Wu, Y.; Ma, L.; Cai, S.; Zhuang, Z.; Zhao, Z.; Jin, S.; Xie, W.; Zhou, L.; Zhang, L.; Zhao, J.; et al. RNA-Induced Liquid Phase Separation of SARS-CoV-2 Nucleocapsid Protein Facilitates NF-κB Hyper-Activation and Inflammation. Signal Transduction and Targeted Therapy 2021, 6, 167. [CrossRef]
- Su, C.-M.; Wang, L.; Yoo, D. Activation of NF-κB and Induction of Proinflammatory Cytokine Expressions Mediated by ORF7a Protein of SARS-CoV-2. Scientific Reports 2021, 11, 13464. [CrossRef]
- Nilsson-Payant Benjamin E.; Uhl Skyler; Grimont Adrien; Doane Ashley S.; Cohen Phillip; Patel Roosheel S.; Higgins Christina A.; Acklin Joshua A.; Bram Yaron; Chandar Vasuretha; et al. The NF-κB Transcriptional Footprint Is Essential for SARS-CoV-2 Replication. Journal of Virology 2021, 95, 10.1128/jvi.01257-21. [CrossRef]
- Robles, J.P.; Zamora, M.; Adan-Castro, E.; Siqueiros-Marquez, L.; Martinez de la Escalera, G.; Clapp, C. The Spike Protein of SARS-CoV-2 Induces Endothelial Inflammation through Integrin A5β1 and NF-κB Signaling. Journal of Biological Chemistry 2022, 298. [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Predicts COVID-19 Severity and Survival. Nature Medicine 2020, 26, 1636–1643. [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and Immunological Features of Severe and Moderate Coronavirus Disease 2019. J Clin Invest 2020, 130, 2620–2629. [CrossRef]
- Hackney, J.A.; Shivram, H.; Vander Heiden, J.; Overall, C.; Orozco, L.; Gao, X.; Kim, E.; West, N.; Qamra, A.; Chang, D.; et al. A Myeloid Program Associated with COVID-19 Severity Is Decreased by Therapeutic Blockade of IL-6 Signaling. iScience 2023, 26. [CrossRef]
- Thwaites, R.S.; Sanchez Sevilla Uruchurtu, A.; Siggins, M.K.; Liew, F.; Russell, C.D.; Moore, S.C.; Fairfield, C.; Carter, E.; Abrams, S.; Short, C.-E.; et al. Inflammatory Profiles across the Spectrum of Disease Reveal a Distinct Role for GM-CSF in Severe COVID-19. Science Immunology 2021, 6, eabg9873. [CrossRef]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum Cytokine and Chemokine Profile in Relation to the Severity of Coronavirus Disease 2019 in China. The Journal of Infectious Diseases 2020, 222, 746–754. [CrossRef]
- Cabrera, L.E.; Pekkarinen, P.T.; Alander, M.; Nowlan, K.H.A.; Nguyen, N.A.; Jokiranta, S.; Kuivanen, S.; Patjas, A.; Mero, S.; Pakkanen, S.H.; et al. Characterization of Low-Density Granulocytes in COVID-19. PLOS Pathogens 2021, 17, e1009721. [CrossRef]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M.; et al. Proinflammatory IgG Fc Structures in Patients with Severe COVID-19. Nature Immunology 2021, 22, 67–73. [CrossRef]
- Hoepel, W.; Chen, H.-J.; Geyer, C.E.; Allahverdiyeva, S.; Manz, X.D.; de Taeye, S.W.; Aman, J.; Mes, L.; Steenhuis, M.; Griffith, G.R.; et al. High Titers and Low Fucosylation of Early Human Anti–SARS-CoV-2 IgG Promote Inflammation by Alveolar Macrophages. Science Translational Medicine 2021, 13, eabf8654. [CrossRef]
- Wigerblad, G.; Warner, S.A.; Ramos-Benitez, M.J.; Kardava, L.; Tian, X.; Miao, R.; Reger, R.; Chakraborty, M.; Wong, S.; Kanthi, Y.; et al. Spleen Tyrosine Kinase Inhibition Restores Myeloid Homeostasis in COVID-19. Science Advances 2023, 9, eade8272. [CrossRef]
- Schrijver, I.T.; Théroude, C.; Antonakos, N.; Regina, J.; Le Roy, D.; Bart, P.-A.; Chiche, J.-D.; Perreau, M.; Pantaleo, G.; Calandra, T.; et al. COVID-19 Rapidly Increases MDSCs and Prolongs Innate Immune Dysfunctions. European Journal of Immunology 2022, 52, 1676–1679. [CrossRef]
- Cheong, J.-G.; Ravishankar, A.; Sharma, S.; Parkhurst, C.N.; Grassmann, S.A.; Wingert, C.K.; Laurent, P.; Ma, S.; Paddock, L.; Miranda, I.C.; et al. Epigenetic Memory of Coronavirus Infection in Innate Immune Cells and Their Progenitors. Cell 2023, 186, 3882-3902.e24. [CrossRef]
- Hopkins, F.R.; Govender, M.; Svanberg, C.; Nordgren, J.; Waller, H.; Nilsdotter-Augustinsson, Å.; Henningsson, A.J.; Hagbom, M.; Sjöwall, J.; Nyström, S.; et al. Major Alterations to Monocyte and Dendritic Cell Subsets Lasting More than 6 Months after Hospitalization for COVID-19. Frontiers in Immunology 2023, 13.
- Beliakova-Bethell, N.; Maruthai, K.; Xu, R.; Salvador, L.C.M.; Garg, A. Monocytic-Myeloid Derived Suppressor Cells Suppress T-Cell Responses in Recovered SARS CoV2-Infected Individuals. Frontiers in Immunology 2022, 13.
- Ruenjaiman, V.; Sodsai, P.; Kueanjinda, P.; Bunrasmee, W.; Klinchanhom, S.; Reantragoon, R.; Tunvirachaisakul, C.; Manothummetha, K.; Mejun, N.; Liengswangwong, K.; et al. Impact of SARS-CoV-2 Infection on the Profiles and Responses of Innate Immune Cells after Recovery. Journal of Microbiology, Immunology and Infection 2022, 55, 993–1004. [CrossRef]
- André F Rendeiro; Joseph Casano; Charles Kyriakos Vorkas; Harjot Singh; Ayana Morales; Robert A DeSimone; Grant B Ellsworth; Rosemary Soave; Shashi N Kapadia; Kohta Saito; et al. Profiling of Immune Dysfunction in COVID-19 Patients Allows Early Prediction of Disease Progression. Life Sci. Alliance 2021, 4, e202000955. [CrossRef]
- Siemińska, I.; Węglarczyk, K.; Surmiak, M.; Kurowska-Baran, D.; Sanak, M.; Siedlar, M.; Baran, J. Mild and Asymptomatic COVID-19 Convalescents Present Long-Term Endotype of Immunosuppression Associated With Neutrophil Subsets Possessing Regulatory Functions. Frontiers in Immunology 2021, 12.
- Tomić, S.; Đokić, J.; Stevanović, D.; Ilić, N.; Gruden-Movsesijan, A.; Dinić, M.; Radojević, D.; Bekić, M.; Mitrović, N.; Tomašević, R.; et al. Reduced Expression of Autophagy Markers and Expansion of Myeloid-Derived Suppressor Cells Correlate With Poor T Cell Response in Severe COVID-19 Patients. Frontiers in Immunology 2021, 12.
- Taniguchi-Ponciano, K.; Vadillo, E.; Mayani, H.; Gonzalez-Bonilla, C.R.; Torres, J.; Majluf, A.; Flores-Padilla, G.; Wacher-Rodarte, N.; Galan, J.C.; Ferat-Osorio, E.; et al. Increased Expression of Hypoxia-Induced Factor 1α mRNA and Its Related Genes in Myeloid Blood Cells from Critically Ill COVID-19 Patients. Annals of Medicine 2021, 53, 197–207. [CrossRef]
- Ricke-Hoch, M.; Stelling, E.; Lasswitz, L.; Gunesch, A.P.; Kasten, M.; Zapatero-Belinchón, F.J.; Brogden, G.; Gerold, G.; Pietschmann, T.; Montiel, V.; et al. Impaired Immune Response Mediated by Prostaglandin E2 Promotes Severe COVID-19 Disease. PLOS ONE 2021, 16, e0255335. [CrossRef]
- Mao, Y.; Sarhan, D.; Steven, A.; Seliger, B.; Kiessling, R.; Lundqvist, A. Inhibition of Tumor-Derived Prostaglandin-E2 Blocks the Induction of Myeloid-Derived Suppressor Cells and Recovers Natural Killer Cell Activity. Clinical Cancer Research 2014, 20, 4096–4106. [CrossRef]
- Altmann, D.M.; Whettlock, E.M.; Liu, S.; Arachchillage, D.J.; Boyton, R.J. The Immunology of Long COVID. Nature Reviews Immunology 2023, 23, 618–634. [CrossRef]
- Haunhorst, S.; Bloch, W.; Javelle, F.; Krüger, K.; Baumgart, S.; Drube, S.; Lemhöfer, C.; Reuken, P.; Stallmach, A.; Müller, M.; et al. A Scoping Review of Regulatory T Cell Dynamics in Convalescent COVID-19 Patients – Indications for Their Potential Involvement in the Development of Long COVID? Frontiers in Immunology 2022, 13.
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple Early Factors Anticipate Post-Acute COVID-19 Sequelae. Cell 2022, 185, 881-895.e20. [CrossRef]
- Park, S.-J.; Nam, D.; Seong, H.C.; Hahn, Y.S. New Discovery of Myeloid-Derived Suppressor Cell’s Tale on Viral Infection and COVID-19. Frontiers in Immunology 2022, 13.
- Guo, Q.; Zhao, Y.; Li, J.; Liu, J.; Yang, X.; Guo, X.; Kuang, M.; Xia, H.; Zhang, Z.; Cao, L.; et al. Induction of Alarmin S100A8/A9 Mediates Activation of Aberrant Neutrophils in the Pathogenesis of COVID-19. Cell Host & Microbe 2021, 29, 222-235.e4. [CrossRef]
- Fiorentino, G.; Coppola, A.; Izzo, R.; Annunziata, A.; Bernardo, M.; Lombardi, A.; Trimarco, V.; Santulli, G.; Trimarco, B. Effects of Adding L-Arginine Orally to Standard Therapy in Patients with COVID-19: A Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Trial. Results of the First Interim Analysis. eClinicalMedicine 2021, 40. [CrossRef]
Figure 1.
(A) MDSCs can produce reactive nitrogen species (RNS) such as nitric oxide through inducible nitric oxide synthase (iNOS). These NO• released by MDSC can react with superoxide (O2•–) to form peroxynitrite (ONOO-). Peroxynitrite can lead to nitration of chemokine (e.g., CCL2) and proteins involved in TCR signaling cascade (e.g., lymphocyte-specific protein tyrosine kinase) which hinders T cell recruitment and activation. Peroxynitrite can also reduce T cell binding and responsiveness to antigen-MHC complex through nitration of tyrosines (forming nitrotyrosine) in TCR-CD8. (B) MDSCs can produce reactive oxygen species (ROS) such as superoxide (O2•–) through NAPDH oxidase 2 (NOX2). Hydrogen peroxide (H2O2) from MDSCs have been shown to reduce T cell CD3ζ expression and inhibit effector function such as the production of IFN-γ by T cells. (C) PMN-MDSCs express Fas. Interaction between Fas on PMN-MDSC with FasL on T cell induces cytotoxic T cell apoptosis. M-MDSCs can also induce T cell apoptosis or anergy using PD-L1 to interact with PD-1 on T cells. (D) M-MDSC can release inhibitory cytokines such as IL-1β, IL-10, and TGF-β to inhibit T cell activation and functions. (E) MDSC can deplete amino acids essential for T cells. MDSCs can reduce cystine/cysteine availability. MDSCs can uptake cystine through SLC7A11 and cystine is converted to cysteine within the cell. The lack of expression of ASC transporter by MDSC prevents the exit of cysteine out of the cell. MDSC can deplete the availability of tryptophan for T cells through IDO, which catabolizes tryptophan to N-formylkynurenine. MDSC can deplete the availability of arginine for T cells through Arg-1, which catabolizes arginine to L-ornithine and urea. SLC7A11: solute carrier family 7 member 11. ASC: alanine-serine-cysteine transporter. IDO: indoleamine 2,3-dioxygenase. Arg-1: Arginase-1. Much of these MDSC immunosuppressive functions have been derived and uncovered from cancer studies.
Figure 1.
(A) MDSCs can produce reactive nitrogen species (RNS) such as nitric oxide through inducible nitric oxide synthase (iNOS). These NO• released by MDSC can react with superoxide (O2•–) to form peroxynitrite (ONOO-). Peroxynitrite can lead to nitration of chemokine (e.g., CCL2) and proteins involved in TCR signaling cascade (e.g., lymphocyte-specific protein tyrosine kinase) which hinders T cell recruitment and activation. Peroxynitrite can also reduce T cell binding and responsiveness to antigen-MHC complex through nitration of tyrosines (forming nitrotyrosine) in TCR-CD8. (B) MDSCs can produce reactive oxygen species (ROS) such as superoxide (O2•–) through NAPDH oxidase 2 (NOX2). Hydrogen peroxide (H2O2) from MDSCs have been shown to reduce T cell CD3ζ expression and inhibit effector function such as the production of IFN-γ by T cells. (C) PMN-MDSCs express Fas. Interaction between Fas on PMN-MDSC with FasL on T cell induces cytotoxic T cell apoptosis. M-MDSCs can also induce T cell apoptosis or anergy using PD-L1 to interact with PD-1 on T cells. (D) M-MDSC can release inhibitory cytokines such as IL-1β, IL-10, and TGF-β to inhibit T cell activation and functions. (E) MDSC can deplete amino acids essential for T cells. MDSCs can reduce cystine/cysteine availability. MDSCs can uptake cystine through SLC7A11 and cystine is converted to cysteine within the cell. The lack of expression of ASC transporter by MDSC prevents the exit of cysteine out of the cell. MDSC can deplete the availability of tryptophan for T cells through IDO, which catabolizes tryptophan to N-formylkynurenine. MDSC can deplete the availability of arginine for T cells through Arg-1, which catabolizes arginine to L-ornithine and urea. SLC7A11: solute carrier family 7 member 11. ASC: alanine-serine-cysteine transporter. IDO: indoleamine 2,3-dioxygenase. Arg-1: Arginase-1. Much of these MDSC immunosuppressive functions have been derived and uncovered from cancer studies.
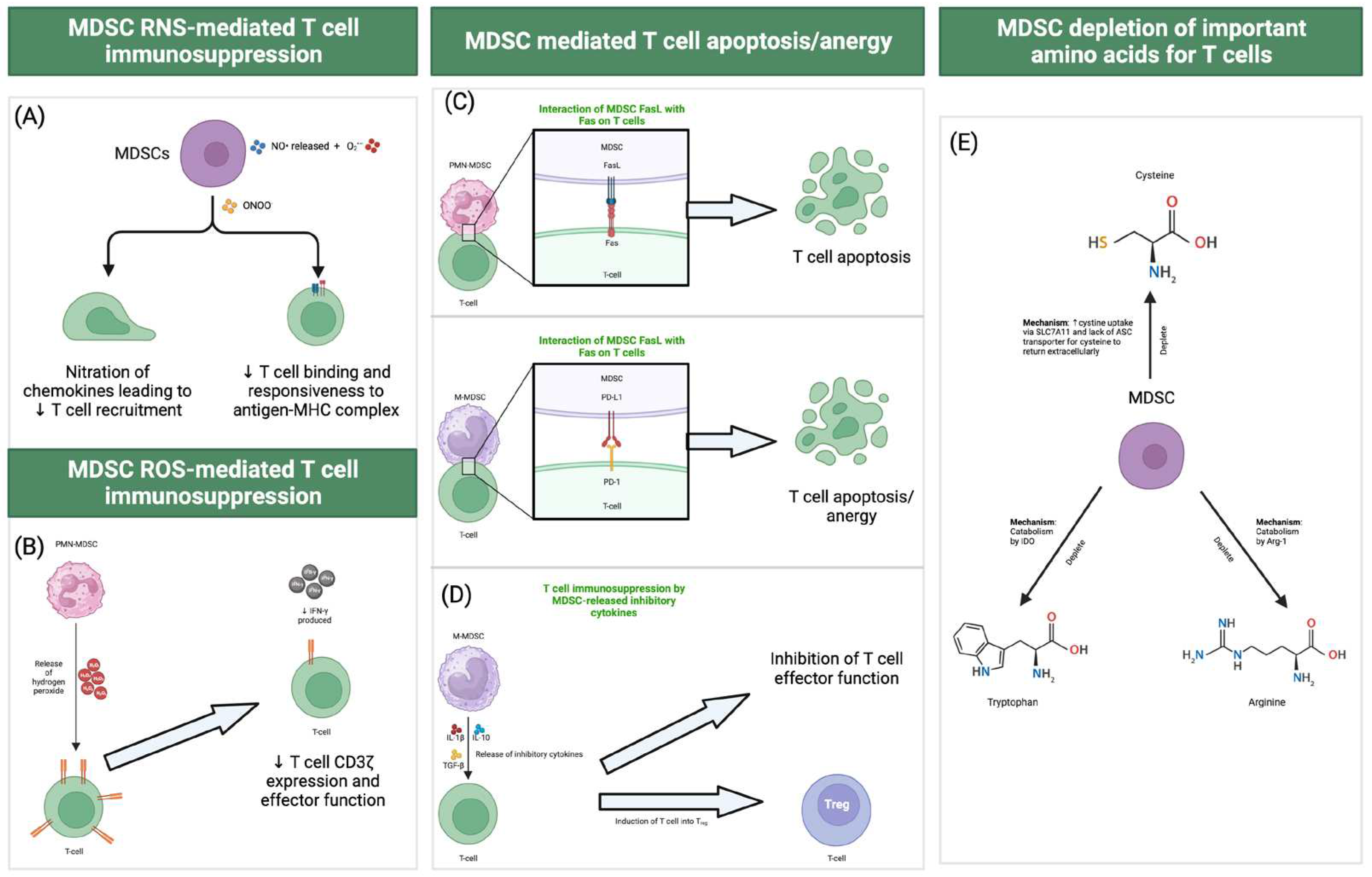
Figure 2.
Web tool demonstrating how users can query individual genes or MDSC gene sets (in this case the query gene is HLA-DRA) to examine their temporal dynamics across multiple timepoints before and after respiratory nadir. Data based on Ong et al., 2021. Web tool can be freely accessed at https://temporal-severe-covid.streamlit.app/.
Figure 2.
Web tool demonstrating how users can query individual genes or MDSC gene sets (in this case the query gene is HLA-DRA) to examine their temporal dynamics across multiple timepoints before and after respiratory nadir. Data based on Ong et al., 2021. Web tool can be freely accessed at https://temporal-severe-covid.streamlit.app/.
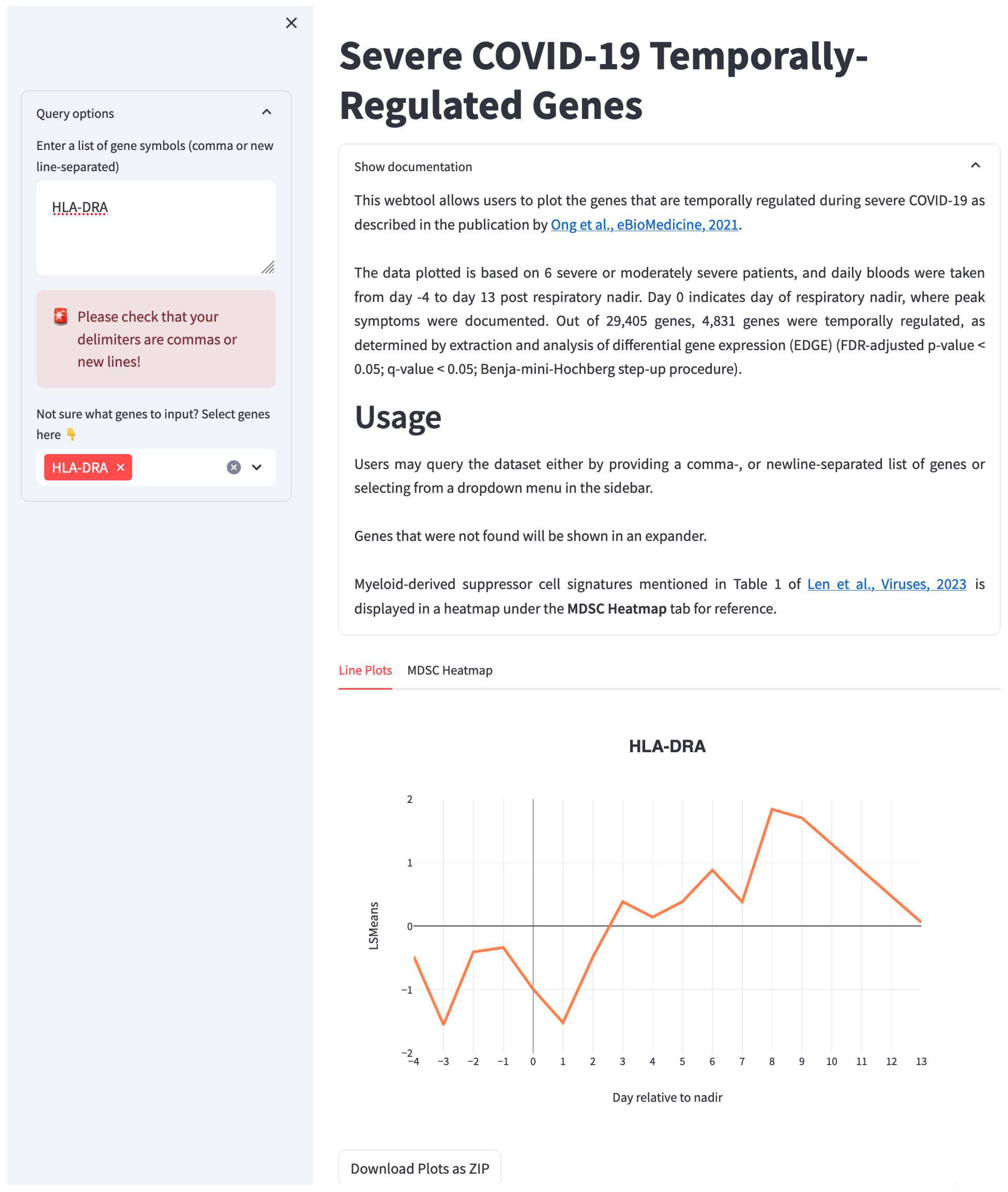
Figure 3.
Potential mechanism for elevated MDSC levels in COVID-19 patients as suggested by three separate studies. 1) Reyes et al. (2021) showed that incubation of human HSPCs with plasma from severe COVID-19 patients which contains high levels of the proinflammatory cytokine, IL-6, induced the generation of MDSC-like cells. 2) Hoepel et al. (2021) demonstrated that high titers of anti-spike IgG or afucosylated anti-spike IgG from severe COVID-19 patients could bind to FcγRIII and FcγRIIa which activates SYK which subsequently leads to enhanced transcription of proinflammatory cytokines such as IL-6. These effects were successfully abrogated by R406, the active component of fostamatinib, a SYK inhibitor. 3) Wigerblad et al. (2023) performed a clinical study which showed that R406 treatment decreased proinflammatory cytokine production and reduced generation of MDSCs, alongside other beneficial effects such as ameliorated interferon responses in severe COVID-19 patients. MDSC: Myeloid-derived suppressor cells. HSPC: hematopoietic stem and progenitor cells.
Figure 3.
Potential mechanism for elevated MDSC levels in COVID-19 patients as suggested by three separate studies. 1) Reyes et al. (2021) showed that incubation of human HSPCs with plasma from severe COVID-19 patients which contains high levels of the proinflammatory cytokine, IL-6, induced the generation of MDSC-like cells. 2) Hoepel et al. (2021) demonstrated that high titers of anti-spike IgG or afucosylated anti-spike IgG from severe COVID-19 patients could bind to FcγRIII and FcγRIIa which activates SYK which subsequently leads to enhanced transcription of proinflammatory cytokines such as IL-6. These effects were successfully abrogated by R406, the active component of fostamatinib, a SYK inhibitor. 3) Wigerblad et al. (2023) performed a clinical study which showed that R406 treatment decreased proinflammatory cytokine production and reduced generation of MDSCs, alongside other beneficial effects such as ameliorated interferon responses in severe COVID-19 patients. MDSC: Myeloid-derived suppressor cells. HSPC: hematopoietic stem and progenitor cells.
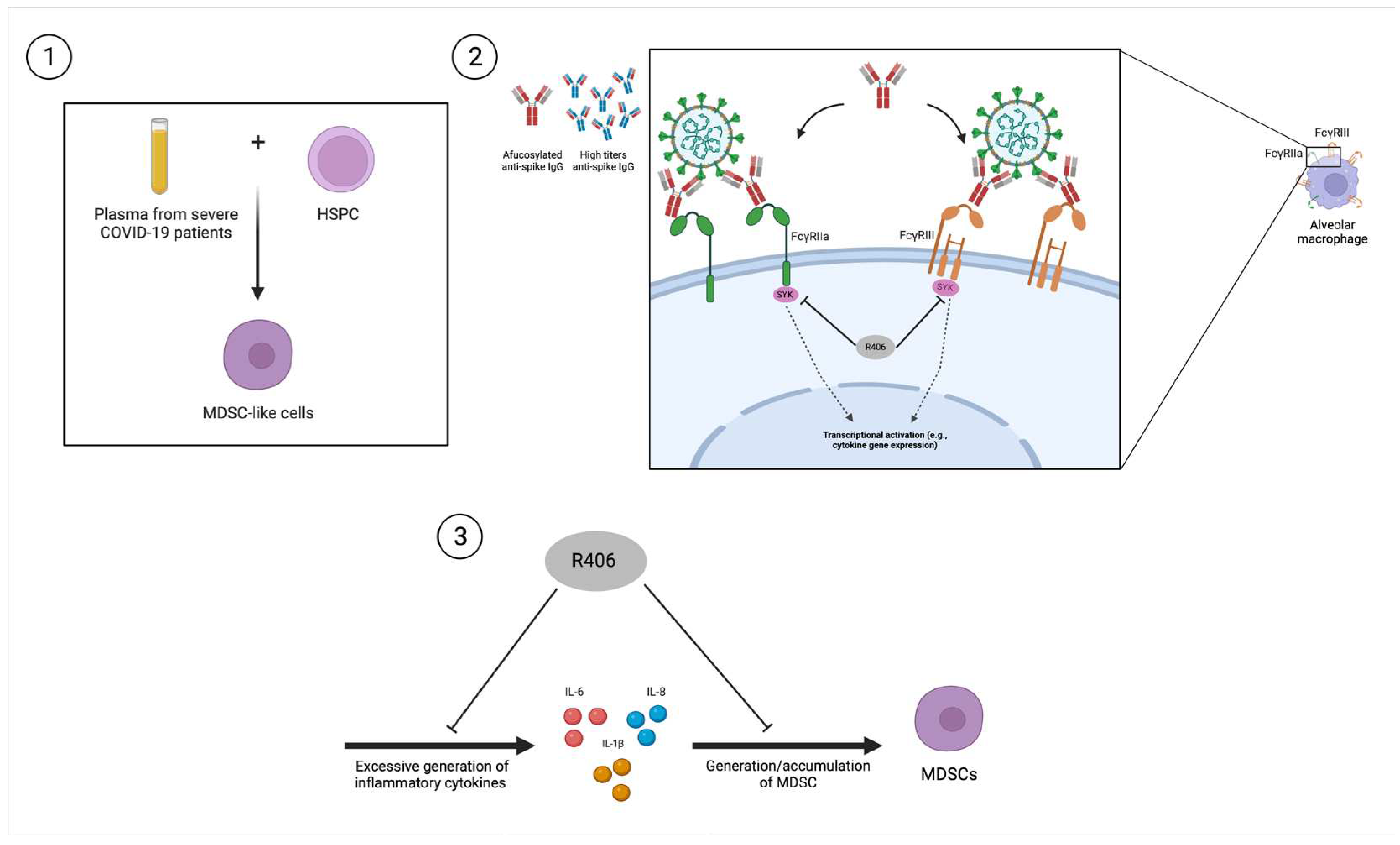

Table 1.
Gene sets or gene signatures identified/ used for identifying M- or PMN-MDSCs in the context of COVID-19. Genes in bold have expression temporally changed during COVID-19 progression. Users can query the temporal dynamics of specific genes or MDSC-related gene sets during severe COVID-19 progression and resolution using the web tool at https://temporal-severe-covid.streamlit.app/.
Table 1.
Gene sets or gene signatures identified/ used for identifying M- or PMN-MDSCs in the context of COVID-19. Genes in bold have expression temporally changed during COVID-19 progression. Users can query the temporal dynamics of specific genes or MDSC-related gene sets during severe COVID-19 progression and resolution using the web tool at https://temporal-severe-covid.streamlit.app/.
| M- or PMN-MDSC | Gene sets used/signature genes identified | Reference |
|---|---|---|
| M-MDSC | S100A8, S100A12, LGALS1, VCAN, RETN, LYZ, PLAC8, MNDA, CTSD, SELL, STXBP2, CYP1B1, VIM, CLU, NKG7, ALOX5AP, NCF1, MCEMP1, TIMP1, SOD2, CD163, NAMPT, FAM65B, ACSL1, VAMP5, LILRA5, VNN2, ANXA6, IL1R2, CALR | [29] |
| CSTA, IL1B, HBB, LRG1, CXCR2, PROK2, IGFBP6, HBA2, SOCS3, ASPRV1, C19orf12, GRINA, CSF3R, CCR1, IFITM2, HP, CTSD, TSPO, S100A11, JUNB, CYP4F2, CD84, CLEC4E, MAP1LC3B, GCNT2, GDA, DUSP1, HDC, C5AR1, MSRB1, SRGN, SELPLG, BTG1, MXD1, HIST1H2BI, STEAP4, SLC40A1, ARG2, SLPI, CLEC4D, TXN, FABP5, CDKN2D, UPP1, PRTN3, CHI3L1, TPD52, CXCL3, RNF149, GPCPD1, S100A6, GSR, LMNB1, TACSTD2, MTUS1, LITAF, NPL, C19orf38, SEPHS2, IL36G, GLIPR2, EIF4EBP1, F10, VCAN, PI16, SMPDL3A, GAPDH, ATP6V1G1, FCGR2A, PLA2G7, CD300LF, YPEL3, KIAA1551, ADIPOR1, UBA52, GLRX, SELL, ATG3, ABTB1, FGL2, ALOX5AP, GADD45A, CD14, SIGLEC9, FBXL5, MYD88, TGFBI, SNAP23, PILRA, FXYD5, BST1, FLOT1, LILRB4, PPT1, MRGPRX3, R3HDM4, PKM, PICALM, SORL1, ZYX, ATP11B, RGS3, CTSC, PTPN1, TALDO1, STK17B, ANKRD33B, GYG1, IL4R, IER3, NUDT4, SKAP2, IL13RA1, LILRB3, GATM, NCF4, F13A1, CDK2AP2, EMB, GCLM, RAB24, CD300LD, SERP1, CEBPB, ALDOA, UBE2H, TARM1, TSC22D3, CARHSP1, HEBP1, KLF13, RBM3, ELP1, PGD, ENO1, DHRS7, OGFRL1, GDPD3, SFXN5, PYGL, ANXA11, DCK, OSM, SDCBP, UBB, EDEM2, LDHA, UBALD2, CCRL2, FTH1, PKIB, PNPLA2, CAP1, KLF2, RND1, LCP1, ANXA2, HOPX, CAMK2D, SUPT4H1, GNG12, ARID5A, CD302, CSF2RB, PPP1R2, RAC2, SLC28A2, PIM1, MPP1, RNASEL, TONSL, SSU72, TBC1D14, ID2, SPI1, HIST1H1C, IER2, MMP8, RTP4, TMEM14C, UBE2B, GPR146, PGLYRP1, MBOAT7, LAMP2, BIN3, PLSCR1, EGR1, EMILIN2, C5orf30, TREM1, BIN2, CALM3, CBR3, SLFN12L | [30] | |
| IFITM1, IFITM2, IFITM3, GRINA, HLA-DRA, HLA-DQB1, HLA-DPA1, CD74 | [31] | |
| PMN-MDSC | SAMD9L, IL1RN, CSF3R, CSF1, GDA, HP, IFIT1, IFIT3, NUDT4, IFIT2, NDST1, IL18RAP, PGLYRP1, CD177, ZBP1, RSAD2, IL1R2, MCEMP1, TYROBP, SLPI, CRISPLD2, S100A6, CMPK2, PADI4, RAF1, S100A9, S100A8, SLC27A4, PFKFB4, C5AR1, FPR1, FPR2, ADIPOR1, RASGRP4, HCAR2, CD300LF, IP6K1, PAG1, CCR1, NBEAL2, ISG15, BST1, ABTB1, SELL, TREML2, TARM1, DMXL2, LCN2, FBXL5, MXD1, RTP4, TINAGL1, SLC2A3, PYGL, LYST, SLC2A6, TIMP2, FFAR2, UPP1, CD33, CERS6, SH2D3C, MMP8, MMP9, DUSP6, RHOB, IL17RA, FGR, MMP13, OAS2, IL1B, PECAM1, IRF7, CD44, TLR2, LILRA6, SLFN5, PTAFR, CXCR4, SAMHD1, THBS1, STX11, KCTD20, CXCR2, CCL3, EGLN3, GABBR1, SEMA4D, GSR, OSM, ASPRV1, KLF2, LRG1, PI16, SMAP2, AGRN, YPEL3, MSRB1, GRINA | [32] |
| S100A12, ARG1, CD177, MCEMP1, and GYG1 | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



