
Submitted:
23 November 2023
Posted:
24 November 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Post-viral fatigue syndrome (PVFS) encompasses a wide range of complex neuroimmune disorders of unknown cause characterized by disabling post-exertional fatigue, myalgia and joint pain, cognitive impairments, unrefreshing sleep, autonomic dysfunction, and neuropsychiatric symptoms. It includes myalgic encephalomyelitis, also known as chronic fatigue syndrome (ME/CFS), fibromyalgia (FM), and more recently post-COVID-19 condition (Long COVID). To date, there are no definitive clinical case criteria and no FDA-approved pharmacological therapies for PVFS. Given the current lack of effective treatments, there is a need to develop novel therapeutic strategies for these disorders. Mitochondria, the cellular organelles responsible for tissue energy production, have recently garnered attention in research into PVFS due to their crucial role in cellular bioenergetic metabolism in these conditions. Accumulating literature has identified a link between mitochondrial dysfunction and low-grade systemic inflammation in ME/CFS, FM, and Long COVID. To address this issue, this article aimed to critically review the evidence relating to mitochondrial dysfunction in the pathogenesis of these disorders; in particular, to evaluate the effectiveness of coenzyme Q10 supplementation on chronic fatigue and pain symptoms as a novel therapeutic strategy for the treatment of PVFS.
Keywords:
chronic fatigue syndrome
; coenzyme Q10
; fibromyalgia
; Long COVID
; mitochondrial dysfunction
; myalgic encephalomyelitis
; post-viral fatigue syndrome
1. Introduction
Post-viral fatigue syndrome (PVFS) comprises common neuroimmune conditions of unknown etiology based on the updated WHO International Classification of Diseases for Mortality and Morbidity Statistics (https://icd.who.int/browse11/l-m/en; accessed on 15 September 2023). PVFS includes myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS), fibromyalgia (FM), and recently post-COVID-19 condition (Long COVID). PVFS is characterized by prolonged post-exertional fatigue as hallmark symptom which worsens by minimal physical and mental exertions, myalgia and joint pain, cognitive impairments, unrefreshing sleep, dysautonomia and neuropsychiatric symptoms such as emotional lability, anxiety/depression and apathy that typically occur following repeated viral infections. Although PVFS is not a generally recognized disorder, it has recently become increasingly associated with the post-COVID-19 condition [1,2].
There are over 65 million people worldwide suffering from chronic disabling disorders who are diagnosed without a clear elucidation of pathophysiologic mechanisms. Adding to this massive disorder burden the CDC now estimate that the largest increase in ME/CFS prevalence after COVID-19 is expected to affect more than 150 million of people by 2050 worldwide, a challenge still unresolved by global healthcare system [3].
To date, there are no definitive diagnostic case criteria, no accurate diagnostic test, and there is no FDA-approved pharmacological treatment for PVFS. Because the exact aetiology of this disorder is not fully understood, together with the unpredictable nature of symptoms, can make the diagnosis and management of the disease challenging. The onset of disease is multifactorial (e.g., a combination of immunogenetic and environmental factors), can occur suddenly or develop gradually over time, and is typically associated with triggering events; in addition to viral infections, these may include physical and emotional trauma [4]. PVFS is often triggered by common viral infections such as Epstein-Barr virus (EBV), human herpesvirus (HHVs), cytomegalovirus (CMV), SARS-CoV-2 (COVID-19), amongst others [5,6].
ME/CFS and FM are multifaceted post-viral syndromes characterized by overwhelming fatigue and widespread musculoskeletal pain that is not alleviated by rest and cannot be explained by any underlying medical condition. In addition to fatigue and chronic pain as “prime” symptoms, individuals may experience an array of symptoms such as post-exertional malaise, unrefreshing sleep, cognitive impairments commonly known as “brain fog”, orthostatic intolerance and gastrointestinal complaints [7,8,9].
These conditions affect millions of people of any age, gender (predominantly women), or socio-economic burden worldwide, and the widely varying impact on individuals’ daily functioning and quality of life can be significant [10,11]. The disease severity can fluctuate in frequency and intensity over time among patients, ranging from mild to moderate symptoms, while others can be severely affected, with around 25% housebound or bedridden [12,13]. The above conditions are established disorders and their background will not be described further. Long COVID is a more recently recognized disorder, and is further described below.
Post-acute sequelae of SARS-CoV-2 infection (PASC), also known as post-COVID-19 condition or Long COVID is an emerging umbrella condition, defined as a constellation of ongoing, relapsing, or new symptoms experienced by people following acute COVID-19 infection that continue for months and even years [14,15]. While most individuals recover from COVID-19 infection within a few weeks, some of them continue experiencing prolonged symptoms that can significantly impact their daily functioning and quality of life. People who experience Long COVID sometimes refer to themselves as “long-haulers” [16]. The term “Long COVID” will be used throughout this review.
A significant portion of convalescent COVID-19 patients, estimated at 10–30% (over 30 million people in the U.S., 20 million in Europe, and up to 180 million worldwide) may experience long-COVID [17]. The wide variation in the estimated prevalence of long-COVID within and between countries may result from a number of determinants, including age and sex of subjects, comorbid health conditions, timing of assessment, sociodemographic factors, and self-reported questionnaire variability, etc [18]. There is emerging evidence suggesting that some individuals with Long COVID exhibit symptoms common to patients with ME/CFS and FM, indicating potential overlapping biological pathomechanisms still unclear [1].
The precise pathomechanism underlying Long COVID is unknown, and is the subject of ongoing research. Risk factors for long-COVID include increasing age, obesity, pre-existing respiratory disorders, and sociodemographic factors [18]. However, it is not yet clear why some individuals develop Long COVID while others recover fully after acute COVID-19 infection. Long-COVID can affect individuals varying in the severity of initial infection, including asymptomatic children/adolescents [19]. Long-term longitudinal clinical and -omics studies are needed to determine if Long COVID can lead to distinct subgroups using cluster analysis in some individuals with ME/CFS and FM [20,21].
ME/CFS, FM and Long COVID can occur independently or coexist in some individuals with other comorbid health conditions, such as irritable bowel syndrome (IBS) and mood disorders such as anxiety/depression, which can further complicate the understanding and management of these conditions [22]. There is currently no cure for these disorders; a multidisciplinary approach that focuses on symptom relief, pacing activities, and improving overall well-being for affected people is often employed [23,24,25,26]. While the exact cause of these illnesses remains unknown, ongoing research is crucial to unravel the complexities of the connection between these conditions, and to develop a prominent hypothesis to deepen the aetiology and underlying pathomechanisms, risk factors, develop more effective diagnostic tools, and identify more effective treatment strategies.
This article aimed to evaluate the evidence relating to mitochondrial dysfunction in the pathogenesis of PVFS; in particular, to review issues relating to the efficacy of coenzyme Q10 (CoQ10) supplementation as a novel therapeutic strategy for treatment of post-viral fatigue syndrome.
2. Evidence of Mitochondrial Dysfunction in Post-Viral Fatigue Syndrome
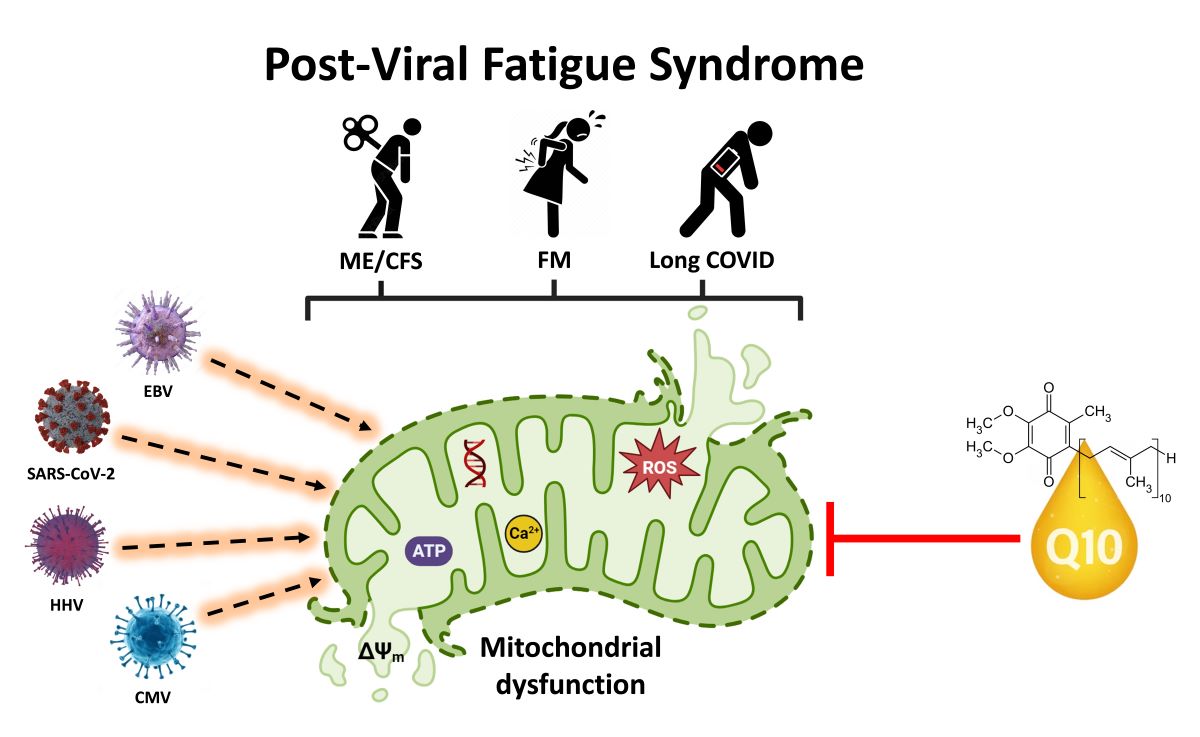
Mitochondria, the cellular powerhouses responsible for energy production, have recently garnered attention in research into PVFS due to their crucial role in cellular energy metabolism. In addition to their role in energy production, mitochondria have key roles in many other aspects of cell metabolism, including free radical metabolism, calcium homeostasis, pyrimidine and lipid synthesis, and apoptosis [27,28,29]. Accumulating literature has identified a link between mitochondrial dysfunction (including oxidative stress, redox imbalance, altered mitochondrial membrane potential/permeability, disrupted calcium homeostasis, and impaired ATP production) and low-grade systemic inflammation in ME/CFS, FM and Long COVID [30,31].
Research has indicated that patients with ME/CFS, FM and Long COVID often exhibit abnormalities in mitochondrial function. Studies have shown decreased ATP production, impaired mitochondrial respiration, abnormal mtDNA levels, immune dysregulation, increased oxidative stress, imbalance redox metabolism, and chronic systemic inflammation perpetuating symptoms in these patients [32,33,34]. Additionally, abnormalities in mitochondrial structure and function have been also observed in muscle biopsies, indicating a systemic impact on energy production. These findings suggest that mitochondrial dysfunction could contribute to the energy depletion and fatigue experienced in these disorders [35,36,37]. In addition to fatigue, these diseases are frequently accompanied by a variety of overlapping symptoms, such as cognitive impairments, sleep disturbances, brain fog, concentration/memory impairments and muscle pain. These symptoms can be attributed, at least partially, to mitochondrial dysfunction, which can have an impact on brain energy metabolism [38,39] (Add here Figure 1 )
Several studies have reported mitochondrial abnormalities in skeletal muscle cells of ME/CFS patients compared to healthy controls. Another study reported lower mitochondrial respiration rates and increased mitochondrial reactive oxygen species production in immune cells of ME/CFS patients. These findings suggest potential mitochondrial impairments in ME/CFS, contributing to energy production deficits and oxidative stress [40,41]. Similarly, in FM, some studies have observed evidence of mitochondrial dysfunction. Research has shown reduced mitochondrial ATP production, impaired oxidative phosphorylation, and increased oxidative stress biomarkers in muscle cells of FM patients. However, more studies are needed to validate and expand upon these findings [34,42].
Regarding post-COVID-19 syndrome, emerging research has indicated potential evidence of mitochondrial dysfunction in blood immune cells from COVID-19 patients. When the COVID-19 virus first enters the host respiratory tract, infection is initiated via the binding of the spike protein with angiotensin-converting enzyme 2 (ACE2) receptors, with subsequent utilization of the transmembrane protease-serine 2 (TMPRSS2) to enter host cells; the virus then hijacks the host cellular machinery for viral RNA replication and protein production [43]. In a similar manner, there is evidence that the SARS-CoV-2 is able also to hijack the host cells’ mitochondria for viral advantage, for example to evade host immune response [44]. It is of note that many other viral, bacterial, fungal or parasitic pathogens also modulate host mitochondrial function to evade host immune response and promote infection [45]. The subject of mitochondrial hijacking in fatigue related disorders is further described in a subsequent section of this article.
COVID-19 can cause systemic inflammation and oxidative stress, which may impact mitochondrial function. Preliminary studies have shown mitochondrial abnormalities in tissues and cells affected by COVID-19, including lung epithelial cells and immune cells. However, more research is needed to understand the specific relationship between mitochondrial dysfunction and post-COVID condition [46]. Several mechanisms may underlie mitochondrial dysfunction in PVFS. These include viral-induced mitochondrial damage, dysregulation of mitochondrial biogenesis and dynamics, immune-mediated mitochondrial dysfunction, and increased oxidative stress. Additionally, dysregulated mitochondrial calcium handling and impaired mitochondrial membrane potential may further contribute to the pathogenesis of PVFS [47,48,49].
One theory proposes that mitochondrial dysfunction in these diseases could be caused by a combination of genetic predisposition and environmental factors. Genetic variations in genes involved in mitochondrial function and energy metabolism, such as those related to mitochondrial DNA, electron transport chain components, and oxidative stress response, may increase the susceptibility to mitochondrial dysfunction in individuals with PVFS. Moreover, various environmental triggers, including infectious agents, toxins, and physical or emotional stressors, have been proposed as potential triggers that can induce or exacerbate mitochondrial dysfunction in susceptible individuals. These triggers may lead to mitochondrial damage, oxidative stress, and inflammation, further impairing mitochondrial function and perpetuating the cycle of fatigue and other core symptoms in people with PVFS [50,51,52,53,54,55].
Understanding the connection between mitochondrial dysfunction and cardinal symptoms in PVFS has important clinical implications. Biomarkers of mitochondrial dysfunction, such as markers of oxidative stress and mitochondrial DNA damage, may aid in the diagnosis and subtyping of PVFS. Furthermore, targeting mitochondrial dysfunction through therapeutic interventions aimed at improving mitochondrial function and reducing oxidative stress and imbalance redox holds promise for the management of PVFS. For example, supplementation with CoQ10 has shown promise in alleviating symptoms in some PVFS patients. However, further research is crucial to unravel the complexities of this connection and develop targeted interventions to improve the quality of life for these individuals and to explore the effectiveness and safety of such interventions [56,57,58,59,60,61,62,63,64]. The rationale for CoQ10 supplementation with regard to mitochondrial hijacking is considered in the following section.
3. Hijacking of Host Mitochondria in Post-Viral Fatigue Related Disorders
In addition to their role in cellular energy provision, mitochondria also have a key role in host innate immune response in the first line defence against RNA viruses. Viral infection results in the activation of mitochondrial antiviral signalling proteins (MAVS), which in turn results in the release of cytokines/chemokines and growth factors by the infected cell; this induces a further immune response which kills the infected host cell, facilitating clearance of the infecting virus [65,66]. Certain types of virus such as SARS-CoV-2 have adapted to promote viral survival and replication by suppressing the host immune response, by forming double-membrane vesicles (DMV) around its RNA, thus shielding the latter from detection, and by inhibiting MAVS in the antiviral innate immune response [67]. While these DMVs are generally believed to be formed via viruses manipulating the endoplasmic reticulum (ER) membrane, the mechanism for importing and packaging proteins and RNA into these miniature organelles is not clearly understood [68]. One possible mechanism for importing viral RNA involves the virus exploiting the RNA localization mechanisms that the cell already possesses for endogenous double-membrane organelles, namely, the mitochondria.
In addition, the COVID-19 virus can directly impair mitochondrial energy metabolism via targeted action on oxygen availability and utilization, and an effective host immune response will be impaired when the available mitochondrial energy is reduced [69]. COVID-19 viruses produce accessory proteins called open reading frames (ORF) which interact with mitochondrial outer membrane receptors. One particular interaction involves ORF-9 interaction with MAVS, in their role as mitochondrial import receptors and cytoplasmic viral recognition receptors [70]. Thus ORF has shown to suppress MAVS activity, thus limiting the initial host cell, innate immune, interferon, and antiviral response. Interactions of the SARS-CoV-2 proteins such as ORF and NSP with host cell mitochondrial proteins lead to loss of membrane integrity and also cause dysfunction in the bioenergetics of the mitochondria.
The hijacking of mitochondria by intracellular viral RNA and protein components also occurs during infections with the Ebola, Zika, and influenza A virus [71]. It follows that drugs which help to prevent mitochondrial hijacking or restore mitochondrial function (of which CoQ10 would be an example) may provide novel therapeutic strategies to help prevent or treat virus infection [72]. As noted in the section of this article on COVID-19 infection, to date variable outcomes have been reported from clinical studies supplementing CoQ10 in COVID-19 patients. Thus an open study by Barletta et al. reported supplementation with 200 mg/day CoQ10 and 200 mg/day alpha lipoic acid for 2 months improved fatigue in chronic COVID-19 patients [73]. However, a randomised controlled trial supplementing CoQ10 (500 mg/day for 6 weeks) found no significant benefit on reducing the number or severity of symptoms in patients with post-COVID-19 condition [74]. The development of novel therapies based on countering the effects of viral hijacking of host mitochondria in post-infectious fatigue disorders therefore remains an area for more intensive future research.
4. Effects of Coenzyme Q10 Supplementation in Post-Viral Fatigue Related Conditions
All of the clinical studies described below supplemented the oxidised form of CoQ10 (ubiquinone) unless otherwise specified (Table 1).
4.1. Coenzyme Q10 Supplementation in Fibromyalgia
With regard to interventions on nutritional supplementation in fibromyalgia, CoQ10 (alone or in nutrient combinations) has been shown to be effective in clinical studies for the symptomatic relief of this disorder. As noted in earlier sections of this article, fibromyalgia patients have depleted CoQ10 levels in tissues (typically 40-50% of normal), together with increased levels of mitochondrial dysfunction, oxidative stress and inflammation, both in adults and children/adolescents [75,76].
A randomised controlled clinical study by Cordero et al. [77] conducted in 20 FM patients found supplementation with CoQ10 (Pharma Nord Bio-Quinone, 300 mg/day for 40 days) significantly reduced (by more than 50%) chronic pain and fatigue; there was a corresponding improvement in mitochondrial energy generation, and reduced oxidative stress and inflammation. In this study, psychopathological symptoms (including anxiety/depression) were also significantly improved; this was linked to the effect of supplemental CoQ10 in reducing oxidative stress and inflammation, as well as increased levels of serotonin [78,79]. In addition, Cordero et al. [80] correlated headache symptoms with reduced CoQ10 levels and increased oxidative stress in FM patients, with headache symptoms and oxidative stress levels significantly improved following CoQ10 supplementation (300 mg/day for 3 months). In adolescent FM patients (aged 8-17 years), Miyamae et al. [76] reported CoQ10 supplementation (100 mg/day for 3 months) significantly improved fatigue. In an open label cross-over study, supplementation with a water-soluble form of CoQ10 (400 mg/day for 6 months) resulted in significant reductions (by 20-40%) in chronic fatigue and pain [81].
A clinical study by Sawaddiruk et al. [82] described the role of supplementary CoQ10 in further reducing chronic pain in FM patients treated with pregabalin. In a double blind randomised controlled study, FM patients were treated either with pregabalin plus CoQ10 or pregabalin plus placebo for 6 weeks. After a two week wash-out period, patients in the pregabalin plus CoQ10 group were switched to pregabalin plus placebo, and vice-versa, for a further 6 weeks. A number of parameters were monitored during the study, including pain pressure threshold, pain score, and anxiety/depression level, as well as biomarkers of antioxidant activity, inflammation and mitochondrial dysfunction. Whilst pregabalin alone reduced pain and anxiety/depression, there was no effect on inflammation and mitochondrial function. However, treatment with pregabalin plus CoQ10 resulted in a significantly greater reduction in pain and anxiety/depression, together with a reduction in oxidative stress, inflammation and mitochondrial dysfunction. The results from this study provide evidence that supplementary CoQ10 can provide further pain relief in FM patients treated with pregabalin, by reducing oxidative stress imbalance and inflammation, and improving mitochondrial function.
With regard to nutrient combinations, an open uncontrolled study of 23 FM patients reported supplementation with 200 mg CoQ10 and 200 mg ginkgo biloba resulted in a significant improvement in health-related quality of life [83]. In a randomised controlled trial conducted in 21 FM patients supplemented with CoQ10, vitamin D, alpha-lipoic acid, magnesium, and tryptophan for 3 months, there was a significant reduction in pain [84].
4.2. Coenzyme Q10 Supplementation in Acute COVID-19 Infection
In a clinical study by Israel et al. [85], intake of CoQ10 was associated with a significantly reduced risk of hospitalisation from COVID-19 patients. Fernandez-Ayala et al. [86] reviewed evidence for mitochondrial dysfunction as a key factor determining the severity of SARS-CoV-2 infection; in particular, the authors noted the increased susceptibility to COVID-19 infection in individuals over 65 years of age, the same age by which endogenous CoQ10 levels have become substantially depleted. Similarly, Gvozdjakova et al. [72] consider one of the main consequences of COVID-19 infection to be virus-induced oxidative stress causing mutations in one or more of the genes responsible for CoQ10 biosynthesis, in turn resulting in mitochondrial dysfunction. Also of note is the computational study by Caruso et al. [87], in which the authors identified CoQ10 as a compound capable of inhibiting the SARS-CoV-2, via binding to the active site of the main viral protease.
Clinical studies supplementing CoQ10 in COVID-19 patients (and also Long COVID) have reported mixed outcomes. In a prospective observational study, 116 patients with chronic COVID-19 were supplemented with 200 mg CoQ10 and 200 mg alpha-lipoic acid for 2 months versus 58 COVID-19 patients who received no treatment; the severity of fatigue was substantially reduced in the treated patients compared to the placebo group [73]. However, a randomised controlled intervention study comprising 121 chronic COVID-19 patients supplemented with 500 mg CoQ10 per day for 6 weeks reported no significant benefit on chronic COVID-19 symptoms [74].
In patients severely affected by COVID-19 infection, in addition to the “cytokine storm” and hyperinflammation status affecting lung tissue, cardiac injury biomarkers may also be elevated, characteristic of myocarditis and heart failure. Whilst the precise mechanism underlying the damaging effect of COVID-19 infection on heart function is not completely understood, attention has been focussed on ACE-2 enzyme, which has roles both in cardiovascular function and the penetration of host cells by the SARS-CoV-2 infection.
It is well known that people with pre-existing medical conditions such as cardiovascular disease are at greater risk of adverse outcomes following COVID-19 infection. The acute effects of COVID-19 infection in terms of cardiovascular function are well established as noted above. What is less well known is the risk of longer term cardiovascular dysfunction following COVID-19 infection. Individuals who have been hospitalised because of COVID-19 infection have a much higher risk of subsequently developing heart problems, including myocarditis, atrial fibrillation, myocardial infarction, heart failure and stroke. However, it is not just people who had suffered more serious COVID-19 infection who are prone to developing heart problems; even people who were not hospitalised and seemed to have recovered from mild infection were subsequently found to be at risk of developing serious heart complications.
These conclusions were based on a number of clinical studies published in the medical literature, including studies led by Dr Ziad Al-Aly (Washington University Medical School, St Louis, USA), Dr Mouaz Al-Mallah (Debakey Heart Centre, Houston, USA), and Prof. Colin Berry (Glasgow University). Dr Al-Aly’s study comprising some 150,000 patients found both hospitalised and non-hospitalised individuals were at substantially increased risk of developing a number of heart conditions, including myocarditis and heart failure [88]. The study by Dr Al-Mallah of 100 COVID-19 patients assessed via positron emission tomography/myocardial blood flow found infection doubled the risk of developing unhealthy endothelial cells lining the heart and blood vessels, increasing the likelihood of heart failure [89]. Additionally, Prof Berry’s study, comprising more than 1000 patients, found that one in eight individuals who had been hospitalised due to COVID infection were later diagnosed with myocarditis [90]. The pathophysiological mechanisms by which SARS-CoV-2 infection causes longer term cardiovascular dysfunction is not completely understood. Possible contributory factors include direct cellular damage resulting from viral invasion of cardiomyocytes and subsequent cell death, endothelial cell infection and endothelitis, persistent hyperactivation of the immune response, and persistence of the virus in tissues, possibly lying dormant for months or even years to be reactivated under conditions of stress/insults.
The above data provide a rationale for the use of supplemental CoQ10 to try and prevent the development of heart problems following COVID-19 infection; in addition, there is evidence that supplemental selenium may also be of benefit, as outlined below. There are a number of reasons why the nutritional supplements CoQ10 and selenium could be of benefit regarding COVID infection. Firstly, both CoQ10 and selenium have important roles in immune function and could help to prevent COVID infection from taking place. Secondly, the combination of CoQ10 and selenium can reduce the excessive inflammation associated with virus infection, as well as inflammation in individuals without virus infection. Thirdly, CoQ10 and selenium have important roles in normal heart function, and clinical trials supplementing CoQ10 and selenium have shown significant benefit in reducing the risk of developing heart disease in normal individuals as well as reducing mortality risk in heart failure patients [91,92,93,94].
4.3. Coenzyme Q10 Supplementation in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome
A randomised controlled trial comprising 207 ME/CFS patients were supplemented with 200 mg CoQ10 and 20 mg NADH per day for 3 months, or placebo; there was a significant improvement in fatigue perception (assessed by FIS-40 scale), health related quality of life (36-item short-form health survey) and sleep quality (assessed by PSQI questionnaire) [63]. In an open label study, 27 ME/CFS patients were supplemented with 400 mg CoQ10 and 200 mcg of selenium per day for 8 weeks; there was a significant improvement in overall fatigue severity and global quality of life compared to baseline among participants [64]. Supplementation with CoQ10 for symptoms such as fatigue, pain and cognitive dysfunction in ME/CFS is suggested in an agreed report from the European Network on ME/CFS (EUROMENE consortium) [95]. In a randomised controlled trial, 43 ME/CFS patients were supplemented with the reduced CoQ10 form (ubiquinol) (150 mg/day for 3 months) or placebo. Although there was no improvement in perceived fatigue (assessed by Chandler's fatigue scale), supplementation improved several other illness symptoms such as night-time awakenings [96].
A potential issue regarding the success or otherwise of clinical trials is the quality and bioavailability of the CoQ10 supplement used, and this is addressed in the following section.
5. Importance of CoQ10 Supplement Quality and Bioavailability
All prescription type drugs require a marketing authorisation. To obtain a marketing authorisation, manufacturers must submit to the relevant regulatory authorities an extensive dossier of data relating to product manufacturing quality, together with proof of efficacy and safety [97]. Once marketing authorisation has been approved, products must be subject to a continual process of pharmacovigilance [98]. By contrast, products classed as food supplements are not subject to the same regulatory standards; there is no mandatory requirement relating to product quality or proof of efficacy and safety. As an example, surveys of Ginkgo biloba supplements available on the European and the US markets have reported issues with formulation discrepancies and adulteration problems. There is similarly no regulatory requirement for the manufacturers of most CoQ10 supplements to guarantee the quality, efficacy and safety of their products [99,100,101].
To date, there is only one CoQ10 product produced to pharmaceutical standards with a marketing authorisation; this is Myoqinon manufactured by Pharma Nord, with a marketing authorisation in Hungary for the adjunctive treatment of heart failure. Marketing authorisation approval acts as a guarantee relating to accuracy of the stated dosage, absence of adulterants, and bioavailability. Products manufactured according to food supplement standards do not have this guarantee, which in turn may have adverse effects on the outcome of clinical studies utilising such products. In this regard, a recent review by Drs Mantle & Hargreaves identified 38 clinical studies in which supplemental CoQ10 had been used to treat patients with primary CoQ10 deficiency. Only two of these studies provided information about the manufacturer of the supplement used. Based on this limited information; it becomes difficult to evaluate the outcomes of such studies in which poor quality supplements may have been used [102].
The other key issue relevant to the success or otherwise of clinical studies is that of bioavailability. Bioavailability is defined as the proportion of an ingested substance that reaches the bloodstream. CoQ10 is a lipid-type substance, and as such is absorbed from the digestive tract in the same general manner as other lipid substances. The process by which this takes place has been described in detail in the review by Mantle & Dybring [103] from which the following information has been summarised. Because of the particular chemical structure of CoQ10 (one of the most hydrophobic naturally occurring substances), the bioavailability of supplemental CoQ10 is low, estimated to be around 5% at most. The single most effective method to date for optimising CoQ10 bioavailability is arguably the patented CoQ10 crystal modification process used by Pharma Nord ApS in the manufacture of their ubiquinone form CoQ10 supplements.
Coenzyme Q10 is produced via a yeast fermentation process in the form of polymorphic crystals, which cannot be absorbed from the digestive tract. CoQ10 can be absorbed only as individual molecules, as noted above. To be effective as a supplement, the CoQ10 crystals must therefore be dissociated first into individual CoQ10 molecules prior to absorption. The above process involves changing the shape of the CoQ10 crystals in such a way as to increase the ratio of the crystals’ surface area to the volume, thus making it easier for the crystals to dissolve into single molecules at body temperature. This modification to the CoQ10 crystalline form should remain in place throughout the shelf life of the CoQ10 preparation.
Supplement manufacturers may make extravagant claims about the bioavailability of their respective CoQ10 supplements, but the only definitive measure of bioavailability is that determined in human subjects, based on clinical studies published in the peer-reviewed medical literature. A good example in this regard is the bioavailability study by Lopez-Lluch et al. [104]. In this randomised controlled clinical trial, seven CoQ10 supplements differing in formulation (CoQ10 crystal modification status, type of carrier oil, composition of other excipients, and CoQ10 oxidation state) were administered in a single 100 mg dose to the same series of 14 healthy individuals, using a crossover/washout protocol. The bioavailability of the different formulations was quantified as the area under the curve at 48 hours. The supplement that had been subject to the crystal modification process (Myoqinon) had the highest level of bioavailability, whilst the bioavailability of the same CoQ10 material that had not been subject to this process was reduced by 75%. It is of note that the Myoqinon formulation received a marketing authorisation within the E.U., as noted above, demonstrating the importance of utilising a CoQ10 product manufactured to pharmaceutical standards, rather than food supplement standards.
A number of studies have been carried out with the objective of improving CoQ10 bioavailability using a variety of agents; examples include polyethylene glycol, phosphorylated tocopherols, poloxamer/polyvinyl pyrrolidine, and hydrolysed proteins. However, again, the bioavailability of most of these formulations has not been directly compared with ubiquinone that has undergone crystal modification, the importance of which is demonstrated in the above Lopez-Lluch et al. study [104]. In addition, none of the modified forms of CoQ10 described above have been subject to an extensive evaluation of efficacy and safety in randomised controlled trials. In comparison, the efficacy and safety of the crystal-modified form of CoQ10 have been confirmed in a number of such clinical studies, an example being the Q-SYMBIO study in which supplemental CoQ10 was shown to substantially reduce mortality risk in heart failure patients [94].
Notwithstanding the issues outlined above, it should be noted that there are other factors that may influence the success of clinical studies involving CoQ10 supplementation, as reviewed by Mantle et al. [105]. Examples include the problem that some individuals appear to have an inherently low capacity to absorb supplemental CoQ10 into the bloodstream, even with high bioavailability formulations - the reason for which is currently unknown. Additionally, the question of whether supplemental CoQ10 can effectively cross the blood-brain barrier in human subjects remains unclear.
6. Conclusions, Unresolved Issues and Future Perspectives
Disorders constituting PVFS, namely ME/CFS, FM and Long COVID, currently have no effective treatments. As part of a strategy to identify new treatment methods, we have identified considerable evidence for the involvement of mitochondrial dysfunction in the pathogenesis of these disorders. This in turn suggests a possible role for CoQ10 supplementation in the treatment of the above disorders, given the key role of CoQ10 in normal mitochondrial function. Randomised controlled clinical trials have reported significant symptomatic benefit in the treatment of these disorders, particularly for FM; further randomised controlled trials are required to confirm the efficacy of CoQ10 supplementation in patients with ME/CFS and Long COVID.
There are a number of unresolved issues relating to CoQ10 intervention studies and mitochondrial dysfunction in PVFS which require further investigation; these include whether the CoQ10 bioavailability could be improved by utilising alternative administration routes (e.g. intravenous, intraperitoneal, intramuscular, etc), whether supplemental CoQ10 is able to cross the blood-brain barrier, and how CoQ10 is transported into and within cells, among others. With regard to future work, the question arises whether additional symptomatic benefit may be obtained by co-supplementation with other substances which have important roles in mitochondrial function, as suggested by Castro-Marrero et al. and Mantle & Hargreaves [63,106]. The possibility of designing dietary interventions based on CoQ10 co-supplementation targeted specifically at boosting mitochondrial function to improve neuroimmune and inflammatory health outcomes in ME/CFS, FM and Long COVID appears well reach within the next half decade. Mitochondria-dependent pathways may thereby represent an attractive therapeutic target for ameliorating PVFS.
Author Contributions
Conceptualization and design, D.M., J.C.D. and J.C.-M.; formal analysis and investigation, D.M., I.P.H. and J.C.-M.; writing—original draft preparation, D.M., J.C.D, and J.C.-M.; writing—review and editing, D.M., J.C.D. and J.C.-M.; visualization, D.M. and J.C.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. Dr Mantle is medical adviser to Pharma Nord (UK) Ltd.
References
- Komaroff, A.L.; Lipkin, W.I. ME/CFS and Long COVID share similar symptoms and biological abnormalities: road map to the literature. Front Med (Lausanne) 2023, 10, 1187163. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L. Chronic "post-infectious" fatigue syndrome. Trans Am Acad Insur Med 1993, 76, 82–95. [Google Scholar] [PubMed]
- Poenaru, S.; Abdallah, S.J.; Corrales-Medina, V.; Cowan, J. COVID-19 and post-infectious myalgic encephalomyelitis/chronic fatigue syndrome: a narrative review. Ther Adv Infect Dis 2021, 8, 20499361211009385. [Google Scholar] [CrossRef] [PubMed]
- Mahroum, N.; Shoenfeld, Y. Autoimmune Autonomic Dysfunction Syndromes: Potential Involvement and Pathophysiology Related to Complex Regional Pain Syndrome, Fibromyalgia, Chronic Fatigue Syndrome, Silicone Breast Implant-Related Symptoms and Post-COVID Syndrome. Pathophysiology 2022, 29(3), 414–425. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W.T.; Gallagher, A.M.; Thomas, J.M.; White, P.D. The prognosis of different fatigue diagnostic labels: a longitudinal survey. Fam Pract 2005, 22(4), 383–8. [Google Scholar] [CrossRef] [PubMed]
- Behan, P.O.; Behan, W.M.; Gow, J.W.; Cavanagh, H.; Gillespie, S. Enteroviruses and postviral fatigue syndrome. Ciba Found Symp 1993, 173, 146-54, discussion 154-9. [Google Scholar] [CrossRef]
- Murga, I.; Aranburu, L.; Gargiulo, P.A.; Gomez Esteban, J.C.; Lafuente, J.V. Clinical Heterogeneity in ME/CFS. A Way to Understand Long-COVID19 Fatigue. Front Psychiatry, 2021; 12, 735784. [Google Scholar] [CrossRef]
- Castro-Marrero, J.; Saez-Francas, N.; Santillo, D.; Alegre, J. Treatment and management of chronic fatigue syndrome/myalgic encephalomyelitis: all roads lead to Rome. Br J Pharmacol 2017, 174(5), 345–369. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Gradisnik, S.; Eaton-Fitch, N. Understanding myalgic encephalomyelitis. Science 2022, 377(6611), 1150–1151. [Google Scholar] [CrossRef] [PubMed]
- Mazurkiewicz, I.; Chatys-Bogacka, Z.; Slowik, J.; Klich-Raczka, A.; Fedyk-Lukasik, M.; Slowik, A.; Wnuk, M.; Drabik, L. Fatigue after COVID-19 in non-hospitalized patients according to sex. Brain Behav 2023, 13(2), e2849. [Google Scholar] [CrossRef]
- Faro, M.; Saez-Francas, N.; Castro-Marrero, J.; Aliste, L.; Fernandez de Sevilla, T.; Alegre, J. Gender differences in chronic fatigue syndrome. Reumatol Clin 2016, 12(2), 72–7. [Google Scholar] [CrossRef]
- Bonilla, H.; Quach, T.C.; Tiwari, A.; Bonilla, A.E.; Miglis, M.; Yang, P.C.; Eggert, L.E.; Sharifi, H.; Horomanski, A.; Subramanian, A.; Smirnoff, L.; Simpson, N.; Halawi, H.; Sum-Ping, O.; Kalinowski, A.; Patel, Z.M.; Shafer, R.W.; Geng, L.N. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome is common in post-acute sequelae of SARS-CoV-2 infection (PASC): Results from a post-COVID-19 multidisciplinary clinic. Front Neurol 2023, 14, 1090747. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, C.; Kirchner, E.; Calabrese, L.H. Long COVID and rheumatology: Clinical, diagnostic, and therapeutic implications. Best Pract Res Clin Rheumatol 2022, 36(4), 101794. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.B.; Javanmard, S.H. Post-COVID-19 Syndrome Mechanisms, Prevention and Management. Int J Prev Med 2023, 14, 59. [Google Scholar] [PubMed]
- Natarajan, A.; Shetty, A.; Delanerolle, G.; Zeng, Y.; Zhang, Y.; Raymont, V.; Rathod, S.; Halabi, S.; Elliot, K.; Shi, J.Q.; Phiri, P. A systematic review and meta-analysis of long COVID symptoms. Syst Rev 2023, 12(1), 88. [Google Scholar] [CrossRef] [PubMed]
- Haslam, A.; Olivier, T.; Prasad, V. The definition of long COVID used in interventional studies. Eur J Clin Invest 2023, e13989. [Google Scholar] [CrossRef] [PubMed]
- Arabi, M.; Al-Najjar, Y.; Sharma, O.; Kamal, I.; Javed, A.; Gohil, H.S.; Paul, P.; Al-Khalifa, A.M.; Laws, S.; Zakaria, D. Role of previous infection with SARS-CoV-2 in protecting against omicron reinfections and severe complications of COVID-19 compared to pre-omicron variants: a systematic review. BMC Infect Dis 2023, 23(1), 432. [Google Scholar] [CrossRef] [PubMed]
- Raman, B.; Bluemke, D.A.; Luscher, T.F.; Neubauer, S. Long COVID: post-acute sequelae of COVID-19 with a cardiovascular focus. Eur Heart J 2022, 43(11), 1157–1172. [Google Scholar] [CrossRef]
- Ludvigsson, J.F. Case report and systematic review suggest that children may experience similar long-term effects to adults after clinical COVID-19. Acta Paediatr 2021, 110(3), 914–921. [Google Scholar] [CrossRef]
- Tate, W.P.; Walker, M.O.M.; Peppercorn, K.; Blair, A.L.H.; Edgar, C.D. Towards a Better Understanding of the Complexities of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and Long COVID. Int J Mol Sci 2023, 24(6). [Google Scholar] [CrossRef]
- Yong, S.J.; Liu, S. Proposed subtypes of post-COVID-19 syndrome (or long-COVID) and their respective potential therapies. Rev Med Virol 2022, 32(4), e2315. [Google Scholar] [CrossRef]
- Barhorst, E.E.; Boruch, A.E.; Cook, D.B.; Lindheimer, J.B. Pain-Related Post-Exertional Malaise in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) and Fibromyalgia: A Systematic Review and Three-Level Meta-Analysis. Pain Med 2022, 23(6), 1144–1157. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Brown, G.M.; Pandi-Perumal, S.R. Possible Application of Melatonin in Long COVID. Biomolecules 2022, 12(11). [Google Scholar] [CrossRef] [PubMed]
- Porter, N.S.; Jason, L.A.; Boulton, A.; Bothne, N.; Coleman, B. Alternative medical interventions used in the treatment and management of myalgic encephalomyelitis/chronic fatigue syndrome and fibromyalgia. J Altern Complement Med 2010, 16(3), 235–49. [Google Scholar] [CrossRef] [PubMed]
- Skilbeck, L. Patient-led integrated cognitive behavioural therapy for management of long COVID with comorbid depression and anxiety in primary care - A case study. Chronic Illn 2022, 18(3), 691–701. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.; Sawant, H.B.; Flannery, T.; Tarrant, R.; Shardha, J.; Bannister, R.; Ross, D.; Halpin, S.; Greenwood, D.C.; Sivan, M. Effect of using a structured pacing protocol on post-exertional symptom exacerbation and health status in a longitudinal cohort with the post-COVID-19 syndrome. J Med Virol 2023, 95(1), e28373. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Mitochondria in Health and Disease. Cells 2019, 8(7). [Google Scholar] [CrossRef] [PubMed]
- San-Millan, I. The Key Role of Mitochondrial Function in Health and Disease. Antioxidants (Basel) 2023, 12(4). [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A mitochondrial bioenergetic etiology of disease. J Clin Invest 2013, 123(4), 1405–12. [Google Scholar] [CrossRef]
- Komaroff, A.L.; Bateman, L. Will COVID-19 Lead to Myalgic Encephalomyelitis/Chronic Fatigue Syndrome? Front Med (Lausanne) 2020, 7, 606824. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L.; Lipkin, W.I. Insights from myalgic encephalomyelitis/chronic fatigue syndrome may help unravel the pathogenesis of postacute COVID-19 syndrome. Trends Mol Med 2021, 27(9), 895–906. [Google Scholar] [CrossRef] [PubMed]
- Holden, S.; Maksoud, R.; Eaton-Fitch, N.; Cabanas, H.; Staines, D.; Marshall-Gradisnik, S. A systematic review of mitochondrial abnormalities in myalgic encephalomyelitis/chronic fatigue syndrome/systemic exertion intolerance disease. J Transl Med 2020, 18(1), 290. [Google Scholar] [CrossRef] [PubMed]
- Myhill, S.; Booth, N.E.; McLaren-Howard, J. Chronic fatigue syndrome and mitochondrial dysfunction. Int J Clin Exp Med 2009, 2(1), 1–16. [Google Scholar] [PubMed]
- Castro-Marrero, J.; Cordero, M.D.; Saez-Francas, N.; Jimenez-Gutierrez, C.; Aguilar-Montilla, F.J.; Aliste, L.; Alegre-Martin, J. Could mitochondrial dysfunction be a differentiating marker between chronic fatigue syndrome and fibromyalgia? Antioxid Redox Signal 2013, 19(15), 1855–60. [Google Scholar] [CrossRef] [PubMed]
- Tomas, C.; Brown, A.E.; Newton, J.L.; Elson, J.L. Mitochondrial complex activity in permeabilised cells of chronic fatigue syndrome patients using two cell types. PeerJ 2019, 7, e6500. [Google Scholar] [CrossRef] [PubMed]
- Arnold, D.L.; Bore, P.J.; Radda, G.K.; Styles, P.; Taylor, D.J. Excessive intracellular acidosis of skeletal muscle on exercise in a patient with a post-viral exhaustion/fatigue syndrome. A 31P nuclear magnetic resonance study. Lancet 1984, 1(8391), 1367–9. [Google Scholar] [CrossRef] [PubMed]
- Plioplys, A.V.; Plioplys, S. Electron-microscopic investigation of muscle mitochondria in chronic fatigue syndrome. Neuropsychobiology 1995, 32(4), 175–81. [Google Scholar] [CrossRef] [PubMed]
- Pintos-Pascual, I.; Moreno-Torres, V.; Ibanez-Estellez, F.; Corrales-Rodriguez, P.; Trevino, A.; Corpas, M.; Corral, O.; Soriano, V.; de Mendoza, C. Is SARS-CoV-2 the only cause of long-COVID? AIDS Rev 2022, 24(4), 183–196. [Google Scholar] [CrossRef] [PubMed]
- Thapaliya, K.; Marshall-Gradisnik, S.; Barth, M.; Eaton-Fitch, N.; Barnden, L. Brainstem volume changes in myalgic encephalomyelitis/chronic fatigue syndrome and long COVID patients. Front Neurosci 2023, 17, 1125208. [Google Scholar] [CrossRef] [PubMed]
- Tomas, C.; Brown, A.; Strassheim, V.; Elson, J.L.; Newton, J.; Manning, P. Cellular bioenergetics is impaired in patients with chronic fatigue syndrome. PLoS One 2017, 12(10), e0186802. [Google Scholar] [CrossRef] [PubMed]
- Smits, B.; van den Heuvel, L.; Knoop, H.; Kusters, B.; Janssen, A.; Borm, G.; Bleijenberg, G.; Rodenburg, R.; van Engelen, B. Mitochondrial enzymes discriminate between mitochondrial disorders and chronic fatigue syndrome. Mitochondrion 2011, 11(5), 735–8. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.D.; de Miguel, M.; Carmona-Lopez, I.; Bonal, P.; Campa, F.; Moreno-Fernandez, A.M. Oxidative stress and mitochondrial dysfunction in fibromyalgia. Neuro Endocrinol Lett 2010, 31(2), 169–73. [Google Scholar] [PubMed]
- Bourgonje, A.R.; Abdulle, A.E.; Timens, W.; Hillebrands, J.L.; Navis, G.J.; Gordijn, S.J.; Bolling, M.C.; Dijkstra, G.; Voors, A.A.; Osterhaus, A.D.; van der Voort, P.H.; Mulder, D.J.; van Goor, H. Angiotensin-converting enzyme 2 (ACE2), SARS-CoV-2 and the pathophysiology of coronavirus disease 2019 (COVID-19). J Pathol 2020, 251(3), 228–248. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Chaubey, G.; Chen, J.Y.; Suravajhala, P. Decoding SARS-CoV-2 hijacking of host mitochondria in COVID-19 pathogenesis. Am J Physiol Cell Physiol 2020, 319(2), C258–C267. [Google Scholar] [CrossRef] [PubMed]
- Tiku, V.; Tan, M.W.; Dikic, I. Mitochondrial Functions in Infection and Immunity. Trends Cell Biol 2020, 30(4), 263–275. [Google Scholar] [CrossRef] [PubMed]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Munch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nature 2020, 583(7816), 469–472. [Google Scholar] [CrossRef] [PubMed]
- Cabanas, H.; Muraki, K.; Eaton, N.; Balinas, C.; Staines, D.; Marshall-Gradisnik, S. Loss of Transient Receptor Potential Melastatin 3 ion channel function in natural killer cells from Chronic Fatigue Syndrome/Myalgic Encephalomyelitis patients. Mol Med 2018, 24(1), 44. [Google Scholar] [CrossRef] [PubMed]
- Cabanas, H.; Muraki, K.; Balinas, C.; Eaton-Fitch, N.; Staines, D.; Marshall-Gradisnik, S. Validation of impaired Transient Receptor Potential Melastatin 3 ion channel activity in natural killer cells from Chronic Fatigue Syndrome/ Myalgic Encephalomyelitis patients. Mol Med 2019, 25(1), 14. [Google Scholar] [CrossRef] [PubMed]
- Cabanas, H.; Muraki, K.; Eaton-Fitch, N.; Staines, D.R.; Marshall-Gradisnik, S. Potential Therapeutic Benefit of Low Dose Naltrexone in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: Role of Transient Receptor Potential Melastatin 3 Ion Channels in Pathophysiology and Treatment. Front Immunol 2021, 12, 687806. [Google Scholar] [CrossRef] [PubMed]
- Cabanas, H.; Muraki, K.; Staines, D.; Marshall-Gradisnik, S. Naltrexone Restores Impaired Transient Receptor Potential Melastatin 3 Ion Channel Function in Natural Killer Cells From Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients. Front Immunol 2019, 10, 2545. [Google Scholar] [CrossRef] [PubMed]
- Missailidis, D.; Annesley, S.J.; Fisher, P.R. Pathological Mechanisms Underlying Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Diagnostics (Basel) 2019, 9(3). [Google Scholar] [CrossRef]
- Morris, G.; Berk, M.; Galecki, P.; Maes, M. The emerging role of autoimmunity in myalgic encephalomyelitis/chronic fatigue syndrome (ME/cfs). Mol Neurobiol 2014, 49(2), 741–56. [Google Scholar] [CrossRef] [PubMed]
- van Tilburg, M.A.L.; Parisien, M.; Boles, R.G.; Drury, G.L.; Smith-Voudouris, J.; Verma, V.; Khoury, S.; Chabot-Dore, A.J.; Nackley, A.G.; Smith, S.B.; Whitehead, W.E.; Zolnoun, D.A.; Slade, G.D.; Tchivileva, I.; Maixner, W.; Diatchenko, L. A genetic polymorphism that is associated with mitochondrial energy metabolism increases risk of fibromyalgia. Pain 2020, 161(12), 2860–2871. [Google Scholar] [CrossRef] [PubMed]
- Ovrom, E.A.; Mostert, K.A.; Khakhkhar, S.; McKee, D.P.; Yang, P.; Her, Y.F. A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia. Biomedicines 2023, 11(4). [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Taylor, K.; Kozubek, J.; Sardell, J.; Gardner, S. Genetic risk factors for ME/CFS identified using combinatorial analysis. J Transl Med 2022, 20(1), 598. [Google Scholar] [CrossRef] [PubMed]
- Campisi, L.; La Motta, C. The Use of the Coenzyme Q10 as a Food Supplement in the Management of Fibromyalgia: A Critical Review. Antioxidants (Basel) 2022, 11(10). [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Araque, A.; Verde, Z.; Torres-Ortega, C.; Sainz-Gil, M.; Velasco-Gonzalez, V.; Gonzalez-Bernal, J.J.; Mielgo-Ayuso, J. Effects of Antioxidants on Pain Perception in Patients with Fibromyalgia-A Systematic Review. J Clin Med 2022, 11(9). [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, I.P.; Mantle, D. Targeted Treatment of Age-Related Fibromyalgia with Supplemental Coenzyme Q10. Adv Exp Med Biol 2021, 1286, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Lowry, E.; Marley, J.; McVeigh, J.G.; McSorley, E.; Allsopp, P.; Kerr, D. Dietary Interventions in the Management of Fibromyalgia: A Systematic Review and Best-Evidence Synthesis. Nutrients 2020, 12(9). [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, S.; Askari, G.; Miraghajani, M.; Tavakoly, R.; Arab, A. Effect of coenzyme Q10 supplementation on fatigue: A systematic review of interventional studies. Complement Ther Med 2019, 43, 181–187. [Google Scholar] [CrossRef]
- Castro-Marrero, J.; Cordero, M.D.; Segundo, M.J.; Saez-Francas, N.; Calvo, N.; Roman-Malo, L.; Aliste, L.; Fernandez de Sevilla, T.; Alegre, J. Does oral coenzyme Q10 plus NADH supplementation improve fatigue and biochemical parameters in chronic fatigue syndrome? Antioxid Redox Signal 2015, 22(8), 679–85. [Google Scholar] [CrossRef]
- Castro-Marrero, J.; Saez-Francas, N.; Segundo, M.J.; Calvo, N.; Faro, M.; Aliste, L.; Fernandez de Sevilla, T.; Alegre, J. Effect of coenzyme Q10 plus nicotinamide adenine dinucleotide supplementation on maximum heart rate after exercise testing in chronic fatigue syndrome: a randomized, controlled, double-blind trial. Clin Nutr 2016, 35(4), 826–34. [Google Scholar] [CrossRef] [PubMed]
- Castro-Marrero, J.; Segundo, M.J.; Lacasa, M.; Martinez-Martinez, A.; Sentanes, R.S.; Alegre-Martin, J. Effect of Dietary Coenzyme Q10 Plus NADH Supplementation on Fatigue Perception and Health-Related Quality of Life in Individuals with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Prospective, Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2021, 13(8). [Google Scholar] [CrossRef] [PubMed]
- Castro-Marrero, J.; Domingo, J.C.; Cordobilla, B.; Ferrer, R.; Giralt, M.; Sanmartin-Sentanes, R.; Alegre-Martin, J. Does Coenzyme Q10 Plus Selenium Supplementation Ameliorate Clinical Outcomes by Modulating Oxidative Stress and Inflammation in Individuals with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome? Antioxid Redox Signal 2022, 36(10-12), 729–739. [Google Scholar] [CrossRef] [PubMed]
- Banoth, B.; Cassel, S.L. Mitochondria in innate immune signaling. Transl Res 2018, 202, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, C.; Horner, S.M. MAVS Coordination of Antiviral Innate Immunity. J Virol 2015, 89(14), 6974–7. [Google Scholar] [CrossRef] [PubMed]
- Roingeard, P.; Eymieux, S.; Burlaud-Gaillard, J.; Hourioux, C.; Patient, R.; Blanchard, E. The double-membrane vesicle (DMV): a virus-induced organelle dedicated to the replication of SARS-CoV-2 and other positive-sense single-stranded RNA viruses. Cell Mol Life Sci 2022, 79(8), 425. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.; Melia, C.E.; Snijder, E.J.; Barcena, M. Double-Membrane Vesicles as Platforms for Viral Replication. Trends Microbiol 2020, 28(12), 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Amar, S.; Gehlot, P.; Patra, S.K.; Kanwar, N.; Kanwal, A. Mitochondrial Modulations, Autophagy Pathways Shifts in Viral Infections: Consequences of COVID-19. Int J Mol Sci 2021, 22(15). [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hou, P.; Ma, W.; Wang, X.; Wang, H.; Yu, Z.; Chang, H.; Wang, T.; Jin, S.; Wang, X.; Wang, W.; Zhao, Y.; Zhao, Y.; Xu, C.; Ma, X.; Gao, Y.; He, H. SARS-CoV-2 ORF10 suppresses the antiviral innate immune response by degrading MAVS through mitophagy. Cell Mol Immunol 2022, 19(1), 67–78. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Das, N.; Mukherjee, P. Picking up a Fight: Fine Tuning Mitochondrial Innate Immune Defenses Against RNA Viruses. Front Microbiol 2020, 11, 1990. [Google Scholar] [CrossRef] [PubMed]
- Gvozdjakova, A.; Kucharska, J.; Sumbalova, Z.; Rausova, Z.; Chladekova, A.; Komlosi, M.; Szamosova, M.; Mojto, V. The importance of coenzyme Q10 and its ratio to cholesterol in the progress of chronic kidney diseases linked to non-communicable diseases. Bratisl Lek Listy 2020, 121(10), 693–699. [Google Scholar] [CrossRef] [PubMed]
- Barletta, M.A.; Marino, G.; Spagnolo, B.; Bianchi, F.P.; Falappone, P.C.F.; Spagnolo, L.; Gatti, P. Coenzyme Q10 + alpha lipoic acid for chronic COVID syndrome. Clin Exp Med 2023, 23(3), 667–678. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.S.; Mogensen, T.H.; Agergaard, J.; Schiottz-Christensen, B.; Ostergaard, L.; Vibholm, L.K.; Leth, S. High-dose coenzyme Q10 therapy versus placebo in patients with post COVID-19 condition: a randomized, phase 2, crossover trial. Lancet Reg Health Eur 2023, 24, 100539. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.D.; Diaz-Parrado, E.; Carrion, A.M.; Alfonsi, S.; Sanchez-Alcazar, J.A.; Bullon, P.; Battino, M.; de Miguel, M. Is inflammation a mitochondrial dysfunction-dependent event in fibromyalgia? Antioxid Redox Signal 2013, 18(7), 800–7. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, T.; Seki, M.; Naga, T.; Uchino, S.; Asazuma, H.; Yoshida, T.; Iizuka, Y.; Kikuchi, M.; Imagawa, T.; Natsumeda, Y.; Yokota, S.; Yamamoto, Y. Increased oxidative stress and coenzyme Q10 deficiency in juvenile fibromyalgia: amelioration of hypercholesterolemia and fatigue by ubiquinol-10 supplementation. Redox Rep 2013, 18(1), 12–9. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.D.; Alcocer-Gomez, E.; de Miguel, M.; Culic, O.; Carrion, A.M.; Alvarez-Suarez, J.M.; Bullon, P.; Battino, M.; Fernandez-Rodriguez, A.; Sanchez-Alcazar, J.A. Can coenzyme q10 improve clinical and molecular parameters in fibromyalgia? Antioxid Redox Signal 2013, 19(12), 1356–61. [Google Scholar] [CrossRef] [PubMed]
- Alcocer-Gomez, E.; Sanchez-Alcazar, J.A.; Cordero, M.D. Coenzyme q10 regulates serotonin levels and depressive symptoms in fibromyalgia patients: results of a small clinical trial. J Clin Psychopharmacol 2014, 34(2), 277–8. [Google Scholar] [CrossRef]
- Alcocer-Gomez, E.; Culic, O.; Navarro-Pando, J.M.; Sanchez-Alcazar, J.A.; Bullon, P. Effect of Coenzyme Q(10) on Psychopathological Symptoms in Fibromyalgia Patients. CNS Neurosci Ther 2017, 23(2), 188–189. [Google Scholar] [CrossRef] [PubMed]
- Cordero, M.D.; Cano-Garcia, F.J.; Alcocer-Gomez, E.; De Miguel, M.; Sanchez-Alcazar, J.A. Oxidative stress correlates with headache symptoms in fibromyalgia: coenzyme Q10 effect on clinical improvement. PLoS One 2012, 7(4), e35677. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F.; Rossi, A.; Consensi, A.; Giacomelli, C.; Bazzichi, L. Role for a water-soluble form of CoQ10 in female subjects affected by fibromyalgia. A preliminary study. Clin Exp Rheumatol 2017, 35 Suppl 105(3), 20–27. [Google Scholar] [PubMed]
- Sawaddiruk, P.; Apaijai, N.; Paiboonworachat, S.; Kaewchur, T.; Kasitanon, N.; Jaiwongkam, T.; Kerdphoo, S.; Chattipakorn, N.; Chattipakorn, S.C. Coenzyme Q10 supplementation alleviates pain in pregabalin-treated fibromyalgia patients via reducing brain activity and mitochondrial dysfunction. Free Radic Res 2019, 53(8), 901–909. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.E. An open, pilot study to evaluate the potential benefits of coenzyme Q10 combined with Ginkgo biloba extract in fibromyalgia syndrome. J Int Med Res 2002, 30(2), 195–9. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, V.; Secchettin, E.; Castellani, C.; Martini, A.; Mazzocchi, E.; Picelli, A.; Polati, E.; Donadello, K.; Valenti, M.T.; Dalle Carbonare, L. Comparison between Acupuncture and Nutraceutical Treatment with Migratens((R)) in Patients with Fibromyalgia Syndrome: A Prospective Randomized Clinical Trial. Nutrients 2020, 12(3). [Google Scholar] [CrossRef]
- Israel, A.; Schaffer, A.A.; Cicurel, A.; Feldhamer, I.; Tal, A.; Cheng, K.; Sinha, S.; Schiff, E.; Lavie, G.; Ruppin, E. Identification of drugs associated with reduced severity of COVID-19: A case-control study in a large population. medRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Moreno Fernandez-Ayala, D.J.; Navas, P.; Lopez-Lluch, G. Age-related mitochondrial dysfunction as a key factor in COVID-19 disease. Exp Gerontol 2020, 142, 111147. [Google Scholar] [CrossRef] [PubMed]
- Caruso, F.; Rossi, M.; Pedersen, J.Z.; Incerpi, S. Computational studies reveal mechanism by which quinone derivatives can inhibit SARS-CoV-2. Study of embelin and two therapeutic compounds of interest, methyl prednisolone and dexamethasone. J Infect Public Health 2020, 13(12), 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, E.; Bowe, B.; Al-Aly, Z. Long-term cardiovascular outcomes of COVID-19. Nat Med 2022, 28(3), 583–590. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.I.; Saad, J.M.; Han, Y.; Alahdab, F.; Malahfji, M.; Nabi, F.; Mahmarian, J.J.; Cooke, J.P.; Zoghbi, W.A.; Al-Mallah, M.H. Coronary Microvascular Health in Patients With Prior COVID-19 Infection. JACC Cardiovasc Imaging 2022, 15(12), 2153–2155. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.J.; Sykes, R.; McIntosh, A.; Kamdar, A.; Bagot, C.; Bayes, H.K.; Blyth, K.G.; Briscoe, M.; Bulluck, H.; Carrick, D.; Church, C.; Corcoran, D.; Findlay, I.; Gibson, V.B.; Gillespie, L.; Grieve, D.; Hall Barrientos, P.; Ho, A.; Lang, N.N.; Lennie, V.; Lowe, D.J.; Macfarlane, P.W.; Mark, P.B.; Mayne, K.J.; McConnachie, A.; McGeoch, R.; McGinley, C.; McKee, C.; Nordin, S.; Payne, A.; Rankin, A.J.; Robertson, K.E.; Roditi, G.; Ryan, N.; Sattar, N.; Allwood-Spiers, S.; Stobo, D.; Touyz, R.M.; Veldtman, G.; Watkins, S.; Weeden, S.; Weir, R.A.; Welsh, P.; Wereski, R.; Consortium, C.-.; Mangion, K.; Berry, C. A multisystem, cardio-renal investigation of post-COVID-19 illness. Nat Med 2022, 28 (6), 1303-1313. [CrossRef]
- Mantle, D.; Heaton, R.A.; Hargreaves, I.P. Coenzyme Q10 and Immune Function: An Overview. Antioxidants (Basel) 2021, 10(5). [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, I.R.; Mantle, D. COVID-19, Coenzyme Q10 and Selenium. Adv Exp Med Biol 2021, 1327, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Johansson, P.; Bjornstedt, M.; Rosen, A.; Dahlstrom, U. Cardiovascular mortality and N-terminal-proBNP reduced after combined selenium and coenzyme Q10 supplementation: a 5-year prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. Int J Cardiol 2013, 167(5), 1860–6. [Google Scholar] [CrossRef]
- Mortensen, S.A.; Rosenfeldt, F.; Kumar, A.; Dolliner, P.; Filipiak, K.J.; Pella, D.; Alehagen, U.; Steurer, G.; Littarru, G.P.; Investigators, Q.S.S. The effect of coenzyme Q10 on morbidity and mortality in chronic heart failure: results from Q-SYMBIO: a randomized double-blind trial. JACC Heart Fail 2014, 2(6), 641–9. [Google Scholar] [CrossRef] [PubMed]
- Nacul, L.; Authier, F.J.; Scheibenbogen, C.; Lorusso, L.; Helland, I.B.; Martin, J.A.; Sirbu, C.A.; Mengshoel, A.M.; Polo, O.; Behrends, U.; Nielsen, H.; Grabowski, P.; Sekulic, S.; Sepulveda, N.; Estevez-Lopez, F.; Zalewski, P.; Pheby, D.F.H.; Castro-Marrero, J.; Sakkas, G.K.; Capelli, E.; Brundsdlund, I.; Cullinan, J.; Krumina, A.; Bergquist, J.; Murovska, M.; Vermuelen, R.C.W.; Lacerda, E.M. European Network on Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (EUROMENE): Expert Consensus on the Diagnosis, Service Provision, and Care of People with ME/CFS in Europe. Medicina (Kaunas) 2021, 57(5). [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Nojima, J.; Kajimoto, O.; Yamaguti, K.; Nakatomi, Y.; Kuratsune, H.; Watanabe, Y. Ubiquinol-10 supplementation improves autonomic nervous function and cognitive function in chronic fatigue syndrome. Biofactors 2016, 42(4), 431–40. [Google Scholar] [CrossRef] [PubMed]
- Todic, M. Dossier for marketing authorization in the European Union. Bosn J Basic Med Sci 2003, 3(1), 56–60. [Google Scholar] [CrossRef] [PubMed]
- Sardella, M.; Belcher, G.; Lungu, C.; Ignoni, T.; Camisa, M.; Stenver, D.I.; Porcelli, P.; D'Antuono, M.; Castiglione, N.G.; Adams, A.; Furlan, G.; Grisoni, I.; Hall, S.; Boga, L.; Mancini, V.; Ciuca, M.; Chonzi, D.; Edwards, B.; Mangoni, A.A.; Tuccori, M.; Prokofyeva, E.; De Gregorio, F.; Bertazzoli Grabinski Broglio, M.; van Leeuwen, B.; Kruger, P.; Rausch, C.; Le Louet, H. Monitoring the manufacturing and quality of medicines: a fundamental task of pharmacovigilance. Ther Adv Drug Saf 2021, 12, 20420986211038436. [Google Scholar] [CrossRef] [PubMed]
- Kressmann, S.; Muller, W.E.; Blume, H.H. Pharmaceutical quality of different Ginkgo biloba brands. J Pharm Pharmacol 2002, 54(5), 661–9. [Google Scholar] [CrossRef] [PubMed]
- Czigle, S.; Toth, J.; Jedlinszki, N.; Haznagy-Radnai, E.; Csupor, D.; Tekelova, D. Ginkgo biloba Food Supplements on the European Market - Adulteration Patterns Revealed by Quality Control of Selected Samples. Planta Med 2018, 84(6-07), 475–482. [Google Scholar] [CrossRef]
- Walkowiak, A.; Wnuk, K.; Cyrankiewicz, M.; Kupcewicz, B. Discrimination of Adulterated Ginkgo Biloba Products Based on 2T2D Correlation Spectroscopy in UV-Vis Range. Molecules 2022, 27(2). [Google Scholar] [CrossRef] [PubMed]
- Mantle, D.; Millichap, L.; Castro-Marrero, J.; Hargreaves, I.P. Primary Coenzyme Q10 Deficiency: An Update. Antioxidants (Basel) 2023, 12(8). [Google Scholar] [CrossRef] [PubMed]
- Mantle, D.; Dybring, A. Bioavailability of Coenzyme Q10: An Overview of the Absorption Process and Subsequent Metabolism. Antioxidants (Basel) 2020, 9(5). [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lluch, G.; Del Pozo-Cruz, J.; Sanchez-Cuesta, A.; Cortes-Rodriguez, A.B.; Navas, P. Bioavailability of coenzyme Q10 supplements depends on carrier lipids and solubilization. Nutrition 2019, 57, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Mantle, D.; Lopez-Lluch, G.; Hargreaves, I.P. Coenzyme Q10 Metabolism: A Review of Unresolved Issues. Int J Mol Sci 2023, 24(3). [Google Scholar] [CrossRef] [PubMed]
- Mantle, D.; Hargreaves, I.P. Mitochondrial Dysfunction and Neurodegenerative Disorders: Role of Nutritional Supplementation. Int J Mol Sci 2022, 23(20). [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of mitochondrial dysfunction involved in the pathogenesis of post-viral fatigue syndrome. The most prevalent clinical features in these disorders affect brain, heart, skeletal muscle, immune cells and intestine (colon). Dysfunctions are commonly caused in multiple organ systems with a clinical spectrum that varies within and between patients. The resulting fall in ATP production due to reduced mitochondrial membrane potential (ΔΨm) and increased ROS generation has a deleterious effect on a number of major ATP-consuming organs in PVFS causing the cardinal symptoms shown.
Figure 1.
Schematic representation of mitochondrial dysfunction involved in the pathogenesis of post-viral fatigue syndrome. The most prevalent clinical features in these disorders affect brain, heart, skeletal muscle, immune cells and intestine (colon). Dysfunctions are commonly caused in multiple organ systems with a clinical spectrum that varies within and between patients. The resulting fall in ATP production due to reduced mitochondrial membrane potential (ΔΨm) and increased ROS generation has a deleterious effect on a number of major ATP-consuming organs in PVFS causing the cardinal symptoms shown.
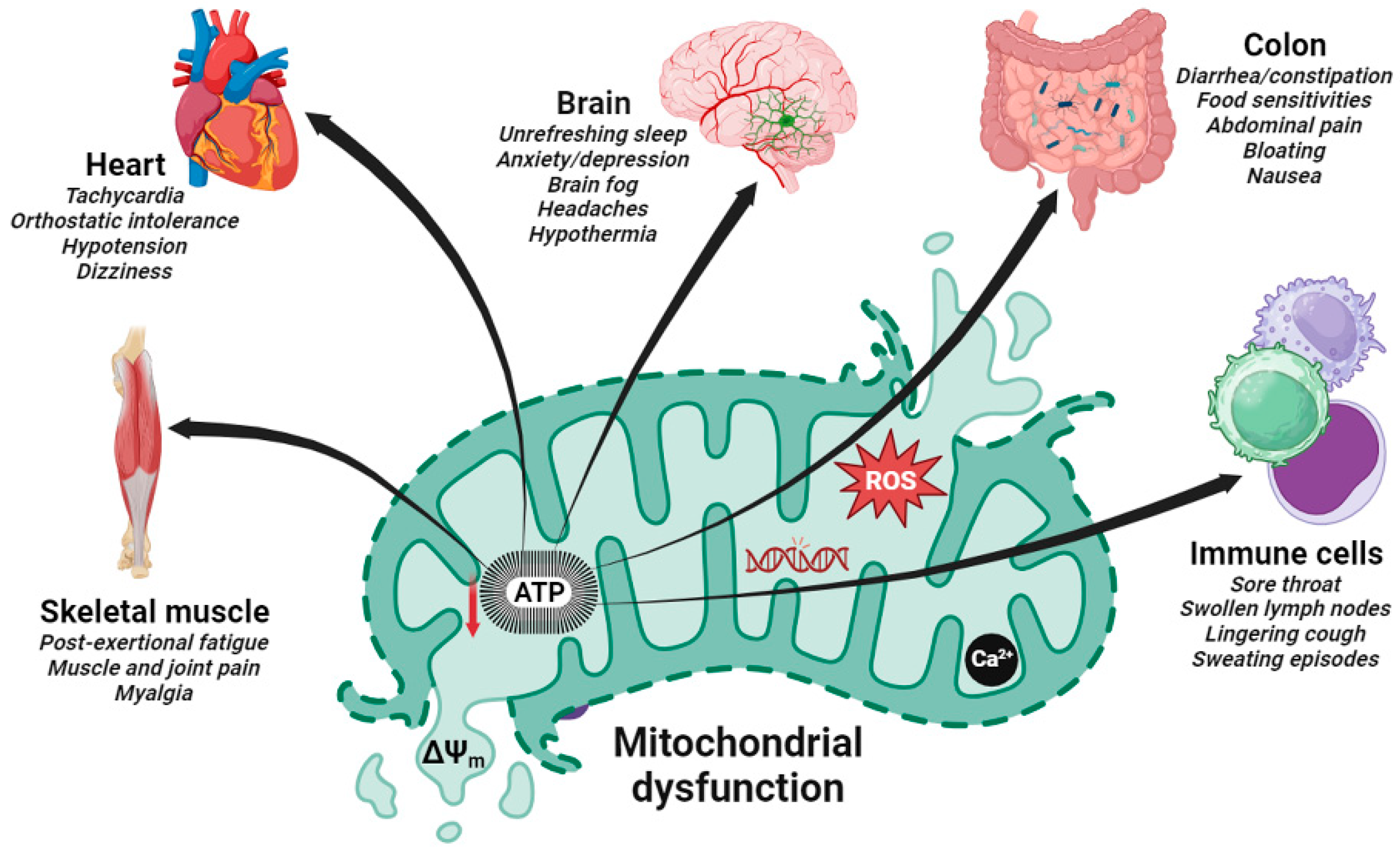

Table 1.
Summary of clinical trials conducted on mitochondria dysfunction-targeted CoQ10 supplementation in post-viral fatigue syndrome.
Table 1.
Summary of clinical trials conducted on mitochondria dysfunction-targeted CoQ10 supplementation in post-viral fatigue syndrome.
| Population | Case criteria |
CoQ10 daily dose and duration | Sample size |
Study design |
Outcomes | Refs. |
|---|---|---|---|---|---|---|
| FM | 1990 ACR | 300 mg for 40 days |
20 | RCT | Reduced chronic pain and fatigue, improved mitochondrial bioenergetic function, reduced oxidative stress and inflammation | Cordero et al. 2013 [77] |
| FM | 2010 ACR | 300 mg for 40 days |
20 | RCT | Regulated serotonin levels in platelets and improved depression symptoms |
Alcocer-Gómez et al. 2014 & 2017 [78,79] |
| FM | 1990 ACR | 300 mg for 40 days |
20 cases 15 controls |
Case-control study | Significant negative correlations between CoQ10 or catalase levels in PBMCs and headache parameters, restored biochemical parameters and improved clinical symptoms | Cordero et al. 2012 [80] |
| FM | 1990 ACR | 2 x 200 mg for 6 months |
22 | Open label crossover study | Significantly improved most pain-related outcomes by 24-37%, including fatigue (by ~22%) and sleep disturbance (by ~33%) |
Di Pierro et al. 2017 [81] |
| FM | 2010 ACR | Pregabalin with CoQ10 or pregabalin with placebo for 40 days | 11 | RCT crossover study | Reduced greater pain, anxiety, inflammation, and mitochondrial oxidative stress and also increased GSH levels and superoxide dismutase levels | Sawaddiruk et al. 2019 [82] |
| FM | Not given | 200 mg CoQ10 and 200 mg Ginkgo biloba extract for 84 days | 25 | Open label pilot study | Improved quality-of-life and improved self-rating with 64% claiming to be better and only 9% claiming to feel worse | Lister et al. 2002 [83] |
| FM | 2016 ACR | 150 mg CoQ10 combined with vitamin D, alpha-lipoic acid, magnesium, and tryptophan or acupuncture treatment, both for 3 months | 55 | RCT | Reduced pain at one month after the start of therapy, strengthened after 3 months with the maintenance of treatment | Schweiger et al. 2020 [84] |
| Long COVID | eHRs from Israel Clalit Health Services provider | Not given |
6953 cases 6530 controls |
Retrospective case-control study | Case studies showing ubiquinone associated with significantly reduced odds for COVID-19 hospitalization (OR = 0.185, 95% CI (0.058 to 0.458), p < 0.001) | Israel et al 2021 [85] |
| Long COVID | 2015 IOM/NIH criteria for ME/CFS | 100 mg CoQ10 and 100 mg alpha-lipoic acid (n= 116) or placebo (n= 58) for 60 days | 174 |
Prospective observational study | Complete Fatigue Severity Scale response was reached more frequently in treatment group (53.5%) than in placebo (3.5%) | Barletta et al. 2023 [73] |
| Long COVID | 2021 WHO clinical case definition | 500 mg or placebo for 6 weeks, with cross-over treatment after a 4-week washout period | 119 | RCT crossover trial | No significant benefit on chronic COVID-19 symptoms | Hansen et al. 2023 [74] |
| ME/CFS | 1994 CDC/Fukuda definition | 200 mg CoQ10 and 20 mg of NADH (n = 104) or placebo (n = 103) for 12 weeks |
207 | RCT | Reduced cognitive fatigue perception and overall FIS-40 score and improved quality of life; improved sleep duration at 4 weeks and improved habitual sleep efficiency at 8 weeks | Castro-Marrero et al. 2021 [63] |
| ME/CFS | 1994 CDC/Fukuda definition | 400 mg CoQ10 and 200 mcg selenium for 8 weeks |
27 | Open label exploratory study | Improved overall fatigue severity and global quality of life but no significant effect on the sleep disturbances; increased total antioxidant capacity, reduced lipoperoxides levels, and decreased circulating cytokine levels | Castro-Marrero et al. 2022 [64] |
| ME/CFS | 1994 CDC/Fukuda | 150 mg ubiquinol or placebo for 3 months | 20 | RCT | Improved autonomic function and cognitive function | Fukuda et al. 2016 [96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



