
Submitted:
24 November 2023
Posted:
26 November 2023
You are already at the latest version
Abstract
Protein-drug interactions are crucial for understanding drug delivery and cell functions. Jacalin is a suitable molecule for such targeting, as it specifically recognizes the tu-mor-associated Thomsen-Friedenreich antigen that is expressed on the glycosylated proteins in cancer cells.
The present paper describes the interaction of curcumin and jacalin, a possible carrier molecule for the delivery of antitumor drugs due to its ability to recognize tumor cells. Our results have shown that both steady state fluorescence and fluorescent label-ling of jacalin are two reliable methods to determine jacalin-curcumin interactions. The affinity of jacalin for curcumin is consistently within the micromolar range (using flu-orescence and microscale thermophoresis) showing high-affinity binding of the com-plex. In vitro experiments on the triple negative breast cancer MDA-MB-231 cells indi-cated inhibition of cell growth after treating with the jacalin-curcumin complex for 48 h. The cell survival fraction was significantly reduced to 50% after combined treatment. In this paper, we report for the first time about the jacalin-curcumin interaction. We quantified this unique biomolecular interaction and gathered additional information on the binding event.
We observed that the jacalin-curcumin complex inhibits the proliferation of the triple negative breast cancer MDA-MB-231 cells.
Keywords:
Protein-ligand interaction
; Curcumin
; Jacalin
; Lectin
; Fluorescence
; Cytotoxicity
; Breast cancer cells
; MDA-MB-231
1. Introduction
Cancer is still one of the leading diseases that cause death in the world. According to the World Health Organization, it is the second leading cause of death globally, accounting for an estimated 10 million deaths, or one in six deaths in the last years. Breast cancer is one of the five most common cancers among women. It is known that cancer treatments include surgical intervention, radiation, and chemotherapeutic drugs, which kill not only cancer cells but also healthy cells, and cause severe side effects in the patients. Therefore, researchers are looking for new alternatives, including natural compounds, some of which have shown proven anticancer activity.
Recently, natural treatments have become very promising of curing for which they attract scientific attention [1] compared to conventional treatments for breast cancer which are costly and have serious side effects. Fruits and vegetables have been shown to contain a diverse source of phytochemicals such as carotenoids, tocopherols and polyphenols which possess chemo-preventive properties at multiple stages of carcinogenesis [2,3].
Jacalin is a galactose-binding lectin isolated from Artocarpus integrifolia (jackfruit, used as a fruit in Asia)[4]. It is a tetrameric protein with a molecular weight of 66 kDa that has various biological activities. The lectin that is extracted from jackfruit (Artocarpus heterophyllus) [5], specifically recognizes the Thomsen-Friedenreich antigen (TF-Gal𝛽1-3GalNAc) [6] and is noncytotoxic for normal cells. This TF antigen is expressed in more than 85% of human carcinomas. Expression of CD176 (TF antigen) is established in the MDA-MB-231 cell line of triple negative breast cancer in 5 - 30% of the cells [7]. Due to its ability to recognize TF antigens, jacalin is considered as a promising carrier molecule in cancer treatment [8,9,10,11,12,13].
Recently, jacalin-PEG phthalocyanine gold nanoparticles were investigated for application in photodynamic therapy. The observed strong phototoxicity in HT-29 cancer cells (95–98%) was mainly due to the specific interactions between jacalin and the TF antigen expressed onto the tumor cell membrane [8].
Curcumin is an active substance of turmeric that inhibits cell proliferation and survival. It induces apoptotic and non-apoptotic cell death, and reduces invasion and migration in different types of malignant cancer cells [14,15,16,17,18,19,20]. In a recent study, it was found that curcumin induces apoptosis in MCF-7 cells in a dose-dependent manner and the cell survival rate actually depended on the curcumin concentration [21].
All these data motivated us to study the jacalin–curcumin interaction in relation to the potential application of jacalin as a drug delivery molecule in cancer treatment. Drug–protein interactions are critical in the delivery of small molecules in biological systems [22,23,24,25,26,27,28]. Understanding the molecular basis of the drug–protein interactions is important in designing novel therapeutic systems or to improve selective drug delivery. Furthermore we also studied the effect of the jacalin-curcumin complex for breast cancer cells. This paper is the first report about the novel jacalin-curcumin binding. We have quantified this unique biomolecular interaction, gathering additional information on the binding event. The antiproliferative effect of the complex for MDA-MB-231 cells was also investigated.
2. Results
It is reported that many plant lectins have hydrophobic sites where they can accommodate different compounds, drugs and physiological ligands. It has been shown that some plant lectins have multiple binding sites allowing the interaction with carbohydrate structures but also the binding with noncarbohydrate ligands [29]. Of special interest to us was to study the interaction between jacalin and curcumin (Figure 1) using steady state fluorescence and microscale thermophoresis.
2.1. Interaction of jacalin with curcumin, registered by steady state fluorescence
We studied the interaction between curcumin and jacalin using a fluorescence titration method. For this purpose, we performed titration of curcumin with increasing concentrations of jacalin (0.28-2.42 µM) and calculated the apparent dissociation constant KD of 1.1±0.22 µM, suggesting high affinity binding of jacalin to curcumin, similar to jacalin-porphyrin complexes [29,30].
The fluorescence titration curve (Figure 2a) is indicative for the binding of nonfluorescent protein to the ligand. In Figure 2b the fluorescence emission spectra of curcumin without jacalin and with increasing concentrations of the protein are presented. The binding of jacalin to curcumin caused a significant increase in the fluorescence intensity (Figure 2b). In addition, a fluorescence emission shift of 18 nm is observed, which is indicative for the accommodation of curcumin within the jacalin molecule.
2.2. Interaction of curcumin with fluorescently labeled jacalin
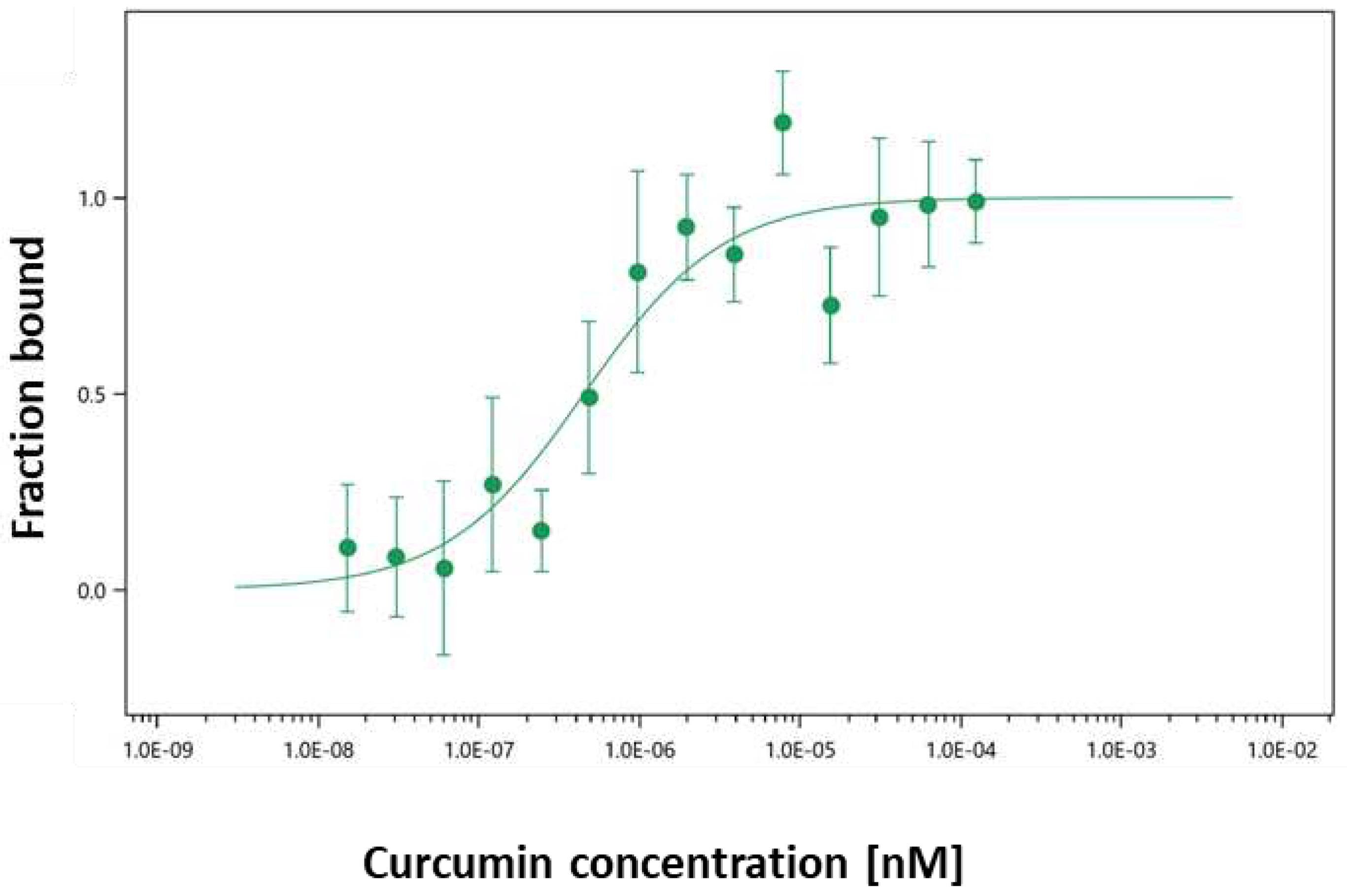
MST is a biophysical method useful to quantify the binding affinities of protein-ligand interactions, respectively jacalin-curcumin interactions. The method uses the ability of jacalin labelled with a fluorescent tag to move along a thermal gradient. Figure 3 shows the differences in thermophoresis as a function of the curcumin concentrations and allows to measure the interaction between curcumin and jacalin. The obtained results have shown that the target protein jacalin interacts with the ligand curcumin with a KD of 0,45 µM ± 0, 2 (Figure 3) in the same range as for steady state fluorescence.
2.3. Cell viability results
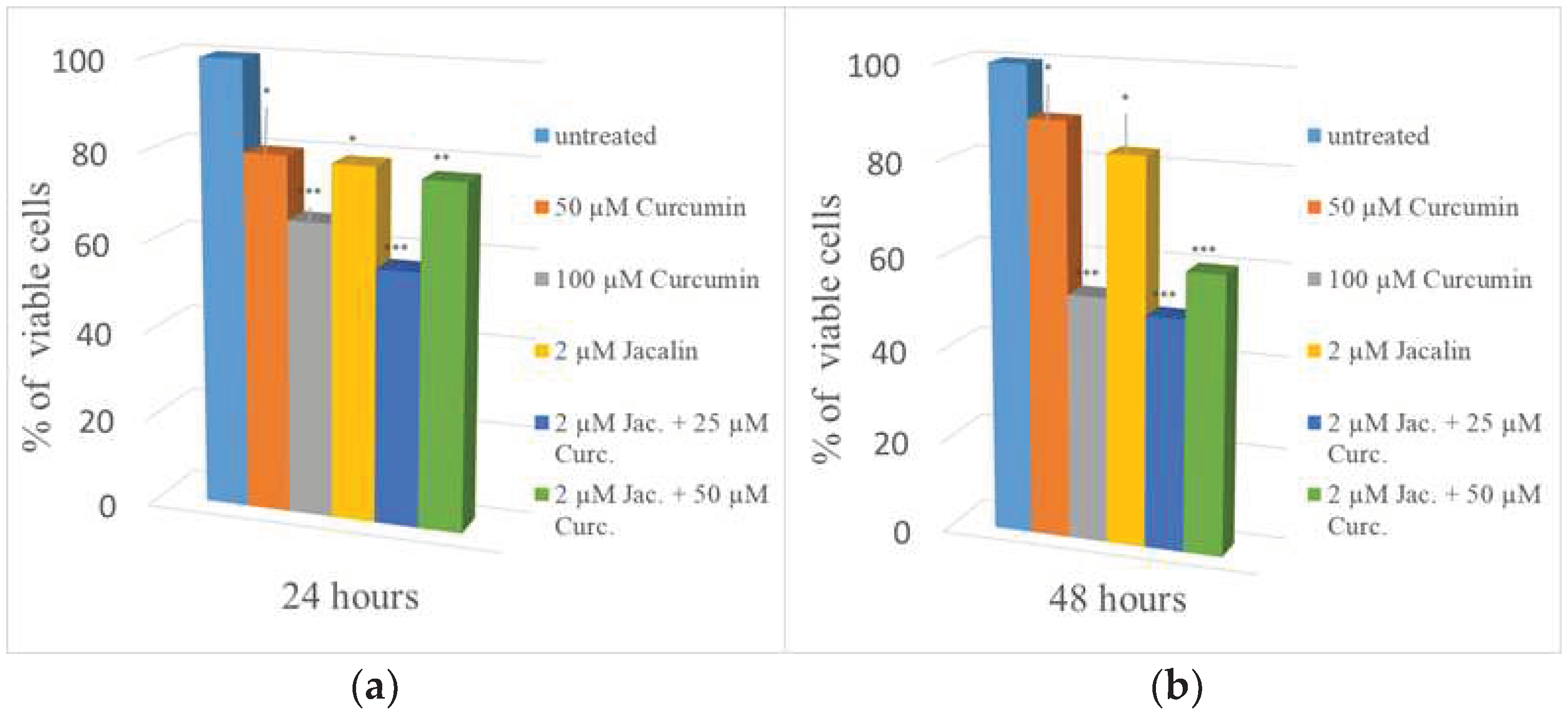
The cytotoxic effect of jacalin (alone) and curcumin (alone), and the jacalin-curcumin complexes (with different combinations: 2 µM:25 µM, 2 µM:50 µM) were studied on the triple negative breast cancer cell line MDA-MB-231, after incubation for 24 h and 48 h (Figure 4). First, cell viability was determined after treatment with jacalin (2 µM). As shown in Figure 4. a, comparatively weak cytotoxicity with a survival of about 79%-83% was induced by jacalin after 24h and 48h incubation, respectfully.
Figure 4 clearly displays the effect of the treatment using two different concentrations of curcumin (50 µM; 100 µM). The results showed that curcumin decreased cell proliferation of MDA-MB-231 cells. The experimental data showed that 24 h incubation with 2 µM jacalin or 50 µM curcumin induced the same percentage (about 20 %) of cell toxicity (Figure 4a). Although the curcumin concentration is 25 orders of magnitude higher than the lectin concentration, both compounds showed similar effects upon MDA-MB-231 cells after 24 h or 48 h incubation. Finally, the effects of jacalin-curcumin complexes (with different combinations: 2 µM:25 µM, 2 µM:50 µM) on the viability of MDA-MB-231 cancer cells were also investigated using the MTT assay. Interestingly, the highest cytotoxicity with cell viability of 50%±1 was induced by the jacalin-curcumin complexes 2 µM:25 µM (Figure 4b). The treatment combination of 2µM jacalin and 50 µM curcumin is less effective reaching a decrease of the cell survival about 60%±2.
Our results confirmed the ability of the jacalin-curcumin complex to inhibit the proliferation of MDA-MB-231 cancer cells. According to the results, relatively high doses of curcumin are necessary to cause a marked decrease in cellular viability.
Figure 4.
Cell viability of breast cancer cell line MDA-MB-231 after treatment with 2 µM jacalin, 50 µM and 100 µM curcumin, and combinations of them (2 µM jacalin/25 µM curcumin; 2 µM jacalin/50 µM curcumin; for 24 h and 48 h. Untreated samples are used as controls. The standard deviations in percentage are presented on the histogram. *- the means are statistically significant at p ≤0.05; **- the means are statistically significant at p≤0.01; ***- the means are statistically significant at p≤0.001.
Figure 4.
Cell viability of breast cancer cell line MDA-MB-231 after treatment with 2 µM jacalin, 50 µM and 100 µM curcumin, and combinations of them (2 µM jacalin/25 µM curcumin; 2 µM jacalin/50 µM curcumin; for 24 h and 48 h. Untreated samples are used as controls. The standard deviations in percentage are presented on the histogram. *- the means are statistically significant at p ≤0.05; **- the means are statistically significant at p≤0.01; ***- the means are statistically significant at p≤0.001.
3. Discussion and conclusions
A universal feature of cancer cells is the altered glycosylation and particular glycan structures are considered as glyco-markers of tumor progression. It is established that the malignant transformation is accompanied by significant alterations in N- and O-glycosylation processes in healthy cells. In cancer cells, the aberrant O-glycans expressed at the cancer cell surface occur as carbohydrate components of membrane-bound N-acetyl galactosamine (O-GalNAc), glycoproteins (T and Tn antigen) and glycolipids (Sialyl-Lewis A and Sialyl-Lewis X). The aberrant O-glycans are considered as potential glyco-markers of cancer diseases [31,32]. It is reported that plant lectins can target O-glycans at the cell surface as tools for cancer diagnosis, prognosis and therapy [33].
The plant lectin jacalin specifically recognizes the tumor-associated Thomsen-Friedenreich antigen and has antiproliferative effects on human colon cancer cells, highlighting its potential antitumor activity [10].
Our results have shown the formation of jacalin-curcumin complexes using fluo-rescence spectroscopy and MST. Binding of jacalin to curcumin enhanced the emission intensity, because of the interaction (Figure 2). The titration curve indicates the interaction of jacalin with curcumin molecules as a function of the protein concentration (Figure 2a). In our investigations, the affinity of jacalin for curcumin is consistently within the micromolar range (KD = 1.1 µM using steady state fluorescence, KD = 0,45 µM using MST experiments) and it reveals that the binding of the lectin for the compound is several orders of magnitude stronger than that of jacalin for porphyrins and carbohyrates [30]. Interestingly, the binding involves hydrophobic interactions [30,34], also for jacalin-curcumin interactions in the present study.
Carbohydrate binding activity has been considered as the major activity of plant lectins for many years. In addition, several plant lectins also showed specific interactions with hydrophobic ligands. These interactions occur at binding sites distinct from the carbohydrate binding site [25].
Jacalin binds AgNPs [12] and the interaction is similar to that of jacalin and other lectins with hydrophobic ligands and plant growth hormones, such as: porphyrins, 2,6-toluidinylnapthalene-sulphonic acid, adenine, auxins and cytokinins (Ka ≈ 1.0 × 103 to 1.0 × 106 M−1) [29,30,35,36].
Jacalin is a D-galactose specific binding lectin with a subunit molecular mass of ~16.5 kDa and is composed of 133 amino acids. The X-ray-crystallographic structure of a jacalin–α-methyl-mannose complex has also been determined. Structural analyses demonstrated the presence of a large carbohydrate-binding site within the jacalin β-prism structure which is a sufficiently flexible structural scaffold that can accommodate carbohydrate structures [37].
Jacalin, being a tumor specific lectin can bind T/Tn antigens at these carbohydrate-binding sites [38].
The jacalin structure demonstrated that native jacalin consists of the tight association of four identical protomers and the protein can be considered as a tetravalent lectin [37]. Interaction of jacalin with curcumin could also be explained with the flexible protein structure that can accommodate curcumin molecules.
Our results provide the first spectroscopic data regarding the interaction of jacalin with curcumin molecules. Curcumin is a fluorescent compound and its fluorescence spectrum is affected by its microenvironment. We registered changes in the fluorescence emission position (registered at 525 nm) indicating the microenvironmental changes due to the lectin binding, resulting in blue shifting (18 nm) and an increase of the specific curcumin fluorescence. Similar to our results, the emission maximum of curcumin in hydrophobically modified starch (HMS) solution is shifted to 531 nm when encapsulated in HMS solution. Interestingly, the anticancer activity of HMS-encapsulated curcumin is higher than that of DMSO dissolved curcumin, suggesting that HMS encapsulation is more effective to deliver curcumin to cancer cells [39].
Curcumin is an active constituent of turmeric; it has been shown to possess an inhibitory effect on cell proliferation and induces apoptosis in a broad range of tumors. Cell inhibition activities of curcumin are different in various cell types. Curcumin induces apoptosis in MCF-7 cells in a dose-dependent manner and its anti-cancer effect on MCF-7 cells under 3D culture conditions could increase the effectiveness of treatment. The cell survival rate depends on the curcumin concentration [21].
In our study we evaluated the curcumin inhibition on breast cancer cells. As jacalin selectively recognizes T-antigens Galb-1–3-GalNAc [6], it can be utilized for targeted cancer therapy. This motivated us to study the effect of the jacalin-curcumin complex upon breast cancer cell line. We chose MDA-MB-231 cells, an invasive and poorly differentiated triple-negative breast cancer (TNBC) cell line which lacks estrogen receptors (ER), progesterone receptors (PR), and the human epidermal growth factor receptors (HER2). MDA-MB-231 does not respond to anticancer compounds which target these receptors [40,41,42,43] but also the hormonal replacement therapy is ineffective. Hence MDA-MB-231 is a good model system for the evaluation of potential new therapeutic drugs and complexes.
Natural compounds are good alternatives to established anticancer drugs and we chose to study their activity. The effects of two natural compounds of plant origin have been evaluated in our work.
First, we studied the cytotoxic effect of jacalin upon cancer cells.
In a recent study, researchers found that jacalin was able to induce macrophage mediated proinflammatory cytokines antitumor activity. Also, it is reported that cytokines were released via the NF-κB signalling pathway. This led to apoptosis in the human colon HT-29 and breast MCF-7 cell lines, respectively. [13].
In mouse and rat bladder cancer models, jacalin showed strong discrimination between normal urothelium and neoplastic urothelium [44]. Recent reports announced that this lectin induced cell cycle arrest in human breast cancer cell line MCF7, in comparison to crude extract, purified jacalin and jacalin standard. IC50 for MCF7 was achieved at concentration of 125 μg/mL [45].
However, in the present study jacalin (alone) was found to inhibit slightly the cell growth of MDA-MB-231 cells. In literature the effect of jacalin (alone) was reversible as the cells recovered after the removal of the lectin. Similar to this finding and in contrary to our expectations, the cytotoxic effect of jacalin (2 µM) alone was lower after 48 h (reaching 17 %), different from the higher effect of jacalin after 24 hours (reaching 21 %). Then, we evaluated the effect of curcumin treatment on the MDA-MB-231 cell line and registered cell death in a dose-dependent manner (Figure 4). These results are in accordance with the dose dependent effects of curcumin on the MCF7 cell line [21].
Finally, we have studied that a combination treatment of low jacalin concentration (2 µM), and 13 orders of magnitude higher concentration of curcumin (25 µM) significantly reduces cell viability. Treating with the unique jacalin-curcumin complex is very promising as both compounds are non-toxic for healthy cells in contrast to chemotherapeutics [46]. Usually, standard chemotherapeutic drugs are effective at high concentrations for triple negative breast cancers. Various combinations including an anticancer drug and a natural compound are the subject of studies on triple negative cancer cell lines. They are applied to minimize the acquired and multi-drug resistance [47]. For example, the treatment combination of 11,65 µM curcumin and 93.95 µM melphalan reduced cell viability to 27% after 24h and to 72,5% after 48h in MDA-MB-231cells [48]. Although there are serious side effects of melphalan treatment (such as iris and chorioretinal atrophy, retinal detachment, etc.), it is still applied for retinoblastoma treatment, and in 2021, it received its first approval for the treatment of adults in the USA [49].
The combined treatment of jacalin (40µg/ml) with the established anti-cancer drug taxol (50 µM) reduced cell viability of the triple negative MDA-MB-468 cell line to 50% after 48h. To reduce the high dose treatment and potential side effects, taxol is combined with the natural compound jacalin [50]. Similar to this, our study shows that jacalin- curcumin treatment causes a significant inhibition of cell growth of 50% after 48h, using 2 times lower concentration of curcumin (25 µM) than taxol (50 µM).
The obtained results showed synergistic effect of both compounds on the triple negative breast cancer cell line MDA-MB-231. Our results confirmed the ability of curcumin-jacalin combination to inhibit the proliferation of MBA-MD-231 cells similar to curcumin induced apoptosis in MCF-7 cells [21].
According to Liu et. al., the antitumor activity of curcumin in mediating the breast cancer cell proliferative rate and invasion is by down-regulating the NF-kB inducing genes. [51].
In conclusion, our results have demonstrated that both steady state fluorescence and fluorescent labelling of jacalin are two reliable methods to determine jacalin-curcumin interactions. The affinity of jacalin for curcumin is consistently within the micromolar range (using fluorescence and microscale thermophoresis) showing high-affinity binding of the complex.
Jacalin potentiates and facilitates the cytotoxic effect of curcumin. The obtained results have shown a synergistic effect of the jacalin-curcumin complex on the triple negative breast cancer cell line MDA-MB-231.
The present study characterizes the capacity of jacalin to bind curcumin, in particular the complex could be an appropriate alternative to the classical anti-cancer drugs, and reveals new perspectives in the drug delivery of anti-cancer agents. Jacalin-curcumin complexes could be used also for cancer prevention or as additives to relieve the side effects of standard chemotherapeutic drugs. The analysis of the protein binding to curcumin not only gives fundamental insights into biomolecular binding but may also direct the way towards improved disease treatment. This study reveals new perspectives for biomedical application of jacalin-curcumin complexes.
4. Materials and Methods
4.1. Materials
All reagents for cell cultures treatments and cell culture assay tests were purchased from Sigma-Aldrich (St. Louis, MO, USA). Curcumin was purchased from Sigma Aldrich. The protein jacalin was dissolved in phosphate buffered saline (PBS) (20mM phosphate buffer containing 0.15M NaCl, pH 7.4) and the concentration of the protein was determined spectrophotometrically using the extinction coefficient. The concentration of curcumin was calculated by the absorbance at 422 nm (e422=3.7x105M-1 cm-1). DMEM medium, L-glutamine, fetal bovine serum (FBS), sodium pyruvate, nonessential amino acids, trypsin and PBS were obtained from Sigma-Aldrich (St. Louis, MO, USA). Antibiotic/antimycotic solution (from Sigma-Aldrich) and absolute ethanol from (Sigma Aldrich), the MTT solution; 3-(4, 5 dimethylthiazol-2-yl)-2, 5-diphenyltetrazoliumbromide) from Sigma-Aldrich (St. Louis, MO). Other chemicals were of the highest quality commercially available.
Measurements were performed on a Monolith NT.115. Proteins were covalently labelled with Red-NHS 2nd generation labeling kit (Alexa Fluor 647), columns and normal/ premium capillaries were purchased from NanoTemper GmbH (Munich, Germany).
4.2. Cell Culture
Breast cancer cell line MDA-MB-231 (ATCC cell culture collection N N HTB-26™) was used in the experiments. The cell line was grown in DMEM medium with high glucose (4.5 g/L) supplemented with 10% FBS and antibiotic/antimycotic solution. Amino acid mix solution (from Sigma-Aldrich) was added to the medium. The cells were cultivated to 80% confluence in CO2 incubator at 37°C in an atmosphere of 5% CO2 – 95% air, subcultured twice a week. Cells were trypsinized with trypsin/EDTA solution. The cells were centrifuged, suspended in FBS with 5% DMSO.
4.3. Purification of Jacalin
Jacalin was isolated from jackfruit seeds by affinity chromatography on immobilized galactose. Briefly, dry seeds (20 g) were soaked overnight in distilled water at 2 °C and homogenized in a Waring blender in 200 ml of 0.1 M Tris/HCl (pH 7.5) containing 0.2 M NaCl. The homogenate was centrifuged at 9000 g for 15 min and the resulting supernatant filtered through filter paper (Whatman 3MM). The cleared extract was applied to a column (2.5 cm x 10 cm; approx. 50 ml bed volume) of galactose–Sepharose 4B equilibrated with 0.1 M PBS (pH 7.5). After loading the extract, the column was washed with the same buffer until the A280 value fell below 0.01. The bound jacalin was eluted with a solution of 0.1 M galactose in PBS and dialyzed against PBS. To remove any possible trace of KM+, the lectin preparation was rechromatographed on galactose–Sepharose 4B, re-eluted with 0.1 M galactose in PBS, dialyzed against the appropriate buffer and stored at ˗ 20 °C until use, as previously described [37].
4.4. Steady state fluorescence
Fluorescence spectroscopy (FS) was performed with a Shimadzu spectrofluorometer RF-5000U (Japan). The emission spectra of the curcumin was recorded in a 1-cm quartz cell (Hellma Analytics, Germany). The curcumin sample was excited at 445 nm with an excitation band pass of 10 nm and an emission band pass of 10 nm. Total fluorescence was calculated after correction for dilution. Absorbance was measured using a spectrophotometer (Beckman, Brea, CA, USA). All measurements were performed at 25 °C.
Curcumin-protein interaction was studied using a fluorescence titration method ideal due to its intrinsic sensitivity and simplicity. For this aim, curcumin was titrated with increasing concentrations of jacalin. The binding of jacalin (0,28-2,42 μM) to curcumin was estimated by a variation of the fluorescence emission of curcumin (excitation at 440 nm). Experimental data (from three independent experiments) were processed by a non-linear regression analysis computed with the GraphPad Prism4 program. Data were statistically analyzed by a binding model. The maximal increase of the curcumin fluorescence due to the saturation of the binding sites by the ligand (Fmax) was estimated from the titration data.
4.5. Jacalin-Curcumin Interaction Studies Using Microscale Thermophoresis
MST experiments were performed by the Monolith NT.115 (NanoTemper GmbH, Munich, Germany) instrument to characterize the protein-ligand interaction. The instrument is equipped with a blue/red filter set, and only the “red” channel was used. An infrared (IR) laser is coupled into the light path of fluorescence excitation and emission and red excitation (605–645 nm, emission 680–685 nm) was used. Experiments were carried out in MST Buffer. The protein jacalin contained lysine residues. Monolith protein labelling kit Red-NHS 2nd generation (Alexa Fluor 647) was used for fluorescent labeling of the protein jacalin following the manufacturer’s recommendations. After elution from the column, the level of the labeled jacalin was checked spectrophotometrically. After that the titration series with non-labeled ligand curcumin was performed. MST measures equilibrium-binding events. The 16 ligand concentrations were prepared with a serial dilution and mixed with an equal volume of the fluorescent labeled jacalin whose concentration was fixed. At this point, all buffers were supplemented to 5% (w/v) DMSO. The final concentration of the protein was 10 nM. The mixture with increasing concentrations of curcumin and the complex was loaded into coated capillaries (NanoTemper). A typical MST experiment featured LED illumination at 100% power and IR illumination between 20% to 60% power, at 24 °C. The instrument was set following the NanoTemper instructions. All data were analyzed using M.O. Affinity Analysis version 2.3, NanoTemper.
4.6. Mitochondrial dehydrogenase assay (MTT assay)
Cells were seeded at 104 cells per well to confluence of 80% into DMEM medium. After 24 hours, the cells were treated with jacalin or curcumin and the combination of both compounds. The following concentrations were applied: curcumin (50 µM and 100 µM); jacalin (2 µM) and two combinations (jacalin 2 µM /curcumin50 µM) and jacalin (2 µM /curcumin 100 µM). The viability was determined at two time points: 24 hours, 48 hours.
MTT test followed the standard procedure (Ref. 1). 20 µl of MTT stock solution (5 mg/ml) was added to the well and the plates were incubated for 4 hours in CO2 incubator. The formazan crystals were solubilized in 150 µl dimethyl sulphoxide. The colour intensity was measured spectrophotometrically, using the wavelength of 600 nm (by ELISA Reader Fluostar Optima/ BMG Tech). The obtained data were processed and compared to the untreated cells. The untreated samples were used as a control with 100 % viability. Assays were performed at least in triplicate. The standard deviation bars were presented in percentage in the graph.
4.7. Statistical Analysis
In all experiments of cell survival (MTT assay) were run at least in triplicate or several repeats. Data were presented as a mean value (+SD or ±SD) from three independent experiments for the MTT assay.
Statistical analyses were performed with GraphPad Prism4 software (GraphPad Software Inc., La Jolla, CA, USA). Data are presented as the mean ± SEM. Differences in the mean of samples were analyzed by one-way ANOVA with selected comparisons using Tukey’s test or by two-way ANOVA with selected comparisons using Bonferroni post hoc test, and the differences were considered significant at p < 0.05 *, p < 0.01 **, or p < 0.001 ***. The results are presented with standard deviations bars [LINK 2]. The absolute values of MTT assay of breast cancer cell line data for different treatment conditions are processed in Student’s t-test for two different conditions (control versus experiment).
Author Contributions
Conceptualization, V.B.; methodology, V.B., D.Z., M.L.; software, V.B., L.P. D.Z. M.L.; validation, N.G., M.S., S.Z. ; formal analysis, L.P, M.L.D.Z.; investigation, N.G., M.S. S. Z.; resources, V.B. , M. L, N.L. D. Z.; data curation, V.B., M. L, D.Z. ; writing—original draft preparation, V. B and L. P.; writing—review and editing, V.B., E.D., N.L.,M. L., D.Z.; visualization, L.P., N.G., M.S., M.L., ; supervision, V.B. ; project administration, V.B. ; funding acquisition, V.B. and L. P.
Funding
This research was 100% funded by the support of Bulgarian National Science Fund, project KП 06-41/09 to V.B.
Acknowledgments
This manuscript was 100% funded by the support of Bulgarian National Fund, project KП 06-41/09 to V.B. The authors highly appreciate the help of Milena Mourdjeva (Institute of Biology and Immunology of Reproduction, Bulgarian Academy of Sciences) for kindly providing cell line MDA-MB-231.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, H.; Khor, T.O.; Shu, L.; Su, Z.-Y.; Fuentes, F.; Lee, J.-H.; Kong, A.-N.T. Plants vs. Cancer: A Review on Natural Phytochemicals in Preventing and Treating Cancers and Their Druggability. Anticancer. Agents Med. Chem. 2012, 12, 1281–1305. [Google Scholar] [CrossRef] [PubMed]
- Naczk, M.; Shahidi, F. Phenolics in Cereals, Fruits and Vegetables: Occurrence, Extraction and Analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabuddhe, A.A.; Ahmed, N.; Krishnasastry, M.V. Stress-Induced Phosphorylation of Caveolin-1 and P38, and down-Regulation of EGFr and ERK by the Dietary Lectin Jacalin in Two Human Carcinoma Cell Lines. Cell Stress Chaperones 2006, 11, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Chatterjee, B.P. Further Characterization and Immunochemical Studies on the Carbohydrate Specificity of Jackfruit (Artocarpus integrifolia) Lectin. J. Biol. Chem. 1989, 264, 9365–9372. [Google Scholar] [CrossRef] [PubMed]
- Kabir, S. The Isolation and Characterisation of Jacalin [Artocarpus Heterophyllus (Jackfruit) Lectin] Based on Its Charge Properties. Int. J. Biochem. Cell Biol. 1995, 27, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Sastry, M.V.; Banarjee, P.; Patanjali, S.R.; Swamy, M.J.; Swarnalatha, G.V.; Surolia, A. Analysis of Saccharide Binding to Artocarpus Integrifolia Lectin Reveals Specific Recognition of T-Antigen (Beta-D-Gal(1----3)D-GalNAc). J. Biol. Chem. 1986, 261, 11726–11733. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-M.; Karsten, U.; Goletz, S.; Cheng, R.-C.; Cao, Y. Expression of CD176 (Thomsen-Friedenreich Antigen) on Lung, Breast and Liver Cancer-Initiating Cells. Int. J. Exp. Pathol. 2011, 92, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Obaid, G.; Chambrier, I.; Cook, M.J.; Russell, D.A. Targeting the Oncofetal Thomsen-Friedenreich Disaccharide Using Jacalin-PEG Phthalocyanine Gold Nanoparticles for Photodynamic Cancer Therapy. Angew. Chem. Int. Ed. 2012, 51, 6158–6162. [Google Scholar] [CrossRef] [PubMed]
- Obaid, G.; Chambrier, I.; Cook, M.J.; Russell, D.A. Cancer Targeting with Biomolecules: A Comparative Study of Photodynamic Therapy Efficacy Using Antibody or Lectin Conjugated Phthalocyanine-PEG Gold Nanoparticles. Photochem. Photobiol. Sci. 2015, 14, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Geraldino, T.H.; Modiano, P.; Veronez, L.C.; Flória-Santos, M.; Garcia, S.B.; Pereira-da-Silva, G. Jacalin Has Chemopreventive Effects on Colon Cancer Development. BioMed. Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Fukase, Y.; Katagiri, T.; Ishikawa, N.; Irie, S.; Sato, T.-A.; Ito, H.; Nakayama, H.; Miyagi, Y.; Tsuchiya, E.; et al. Targeted Serum Glycoproteomics for the Discovery of Lung Cancer-Associated Glycosylation Disorders Using Lectin-Coupled ProteinChip Arrays. Proteomics 2009, 9, 2182–2192. [Google Scholar] [CrossRef] [PubMed]
- Ayaz Ahmed, K.B.; Mohammed, A.S.; Veerappan, A. Interaction of Sugar Stabilized Silver Nanoparticles with the T-Antigen Specific Lectin, Jacalin from Artocarpus Integrifolia. Spectrochim. Acta. A Mol. Biomol. Spectrosc. 2015, 145, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Danella Polli, C.; Pereira Ruas, L.; Chain Veronez, L.; Herrero Geraldino, T.; Rossetto De Morais, F.; Roque-Barreira, M.C.; Pereira-da-Silva, G. Jacalin-Activated Macrophages Exhibit an Antitumor Phenotype. BioMed. Res. Int. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Basnet, P.; Skalko-Basnet, N. Curcumin: An Anti-Inflammatory Molecule from a Curry Spice on the Path to Cancer Treatment. Molecules 2011, 16, 4567–4598. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shu, W.; Chen, W.; Wu, Q.; Liu, H.; Cui, G. Curcumin, Both Histone Deacetylase and P300/CBP-Specific Inhibitor, Represses the Activity of Nuclear Factor Kappa B and Notch 1 in Raji Cells. Basic Clin. Pharmacol. Toxicol. 2007, 101, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From Kitchen to Clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xu, Y.; Meng, L.; Huang, L.; Sun, H. Curcumin Inhibits Proliferation and Promotes Apoptosis of Breast Cancer Cells. Exp. Ther. Med. 2018, 16, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer Potential of Curcumin: Preclinical and Clinical Studies. Anticancer Res. 2003, 23 (1A), 363–398. [Google Scholar]
- Sinha, D.; Biswas, J.; Sung, B.; B. Aggarwal, B.; Bishayee, A. Chemopreventive and Chemotherapeutic Potential of Curcumin in Breast Cancer. Curr. Drug Targets 2012, 13, 1799–1819. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.-D.; Liu, X.-P.; Zhao, W.-J.; Dong, Q.; Li, F.-N.; Wang, H.-B.; Kong, B. Curcumin Induces Apoptosis in Breast Cancer Cells and Inhibits Tumor Growth in Vitro and in Vivo. Int. J. Clin. Exp. Pathol. 2014, 7, 2818–2824. [Google Scholar] [PubMed]
- Zargan, S.; Salehi Barough, M.; Zargan, J.; Shayesteh, M.; Haji Noor Mohammadi, A.; Mousavi, M.; Keshavarz Alikhani, H. Evaluation of the Anti-Cancer Effect of Curcumin on MCF-7 Cells in 3D Culture Conditions to Increase the Efficacy of Breast Cancer Treatment. J. Appl. Biotechnol. Rep. 2022, 9. [Google Scholar] [CrossRef]
- D’Auria, S.; Petrova, L.; John, C.; Russev, G.; Varriale, A.; Bogoeva, V. Tumor-Specific Protein Human Galectin-1 Interacts with Anticancer Agents. Mol. Biosyst. 2009, 5, 1331. [Google Scholar] [CrossRef] [PubMed]
- Bogoeva, V.P.; Petrova, L.P.; Ivanov, I.B.; Kulina, H.N.; Buchvarov, I. Ch. Characterization of Metalloanticancer Capacity of an Agglutinin from Wheat. Mol. Biosyst. 2012, 8, 2633. [Google Scholar] [CrossRef]
- Bogoeva, V.; Petrova, L.; Kubát, P. Binding of Palladium (II) 5, 10, 15, 20-Tetrakis (4-Sulfonatophenyl) Porphyrin to a Lectin for Photosensitizer Targeted Delivery. J. Photochem. Photobiol. B 2015, 153, 276–280. [Google Scholar] [CrossRef]
- Bogoeva, V.; Petrova, L.; Bouckaert, J.; Yordanova, A.; Ivanov, I.; Vanderesse, R.; Frochot, C. Dual Function of Lectins — New Perspectives in Targeted Photodynamic Therapy. J. Porphyr. Phthalocyanines 2019, 23 (11n12), 1241–1250. [Google Scholar] [CrossRef]
- Kunwar, A.; Barik, A.; Pandey, R.; Priyadarsini, K.I. Transport of Liposomal and Albumin Loaded Curcumin to Living Cells: An Absorption and Fluorescence Spectroscopic Study. Biochim. Biophys. Acta BBA - Gen. Subj. 2006, 1760, 1513–1520. [Google Scholar] [CrossRef]
- Ayaz Ahmed, K.B.; Raja, M.R. C.; Mahapatra, S.K.; Anbazhagan, V. Interaction of Cadmium Sulfide Quantum Dots with Jacalin for Specific Recognition of Cancer Cells. J. Lumin. 2017, 182, 283–288. [Google Scholar] [CrossRef]
- Sahu, A.; Kasoju, N.; Bora, U. Fluorescence Study of the Curcumin−Casein Micelle Complexation and Its Application as a Drug Nanocarrier to Cancer Cells. Biomacromolecules 2008, 9, 2905–2912. [Google Scholar] [CrossRef]
- Komath, S.S.; Kavitha, M.; Swamy, M.J. Beyond Carbohydrate Binding: New Directions in Plant Lectin Research. Org. Biomol. Chem. 2006, 4, 973. [Google Scholar] [CrossRef]
- Komath, S.S.; Bhanu, K.; Maiya, B.G.; Swamy, M.J. Binding of Porphyrins by the Tumor-Specific Lectin, Jacalin [Jack Fruit (Artocarpus Integrifolia) Agglutinin]. Biosci. Rep. 2000, 20, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Fuster, M.M.; Esko, J.D. The Sweet and Sour of Cancer: Glycans as Novel Therapeutic Targets. Nat. Rev. Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Glavey, S.V.; Huynh, D.; Reagan, M.R.; Manier, S.; Moschetta, M.; Kawano, Y.; Roccaro, A.M.; Ghobrial, I.M.; Joshi, L.; O’Dwyer, M.E. The Cancer Glycome: Carbohydrates as Mediators of Metastasis. Blood Rev. 2015, 29, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Poiroux, G.; Barre, A.; Van Damme, E.; Benoist, H.; Rougé, P. Plant Lectins Targeting O-Glycans at the Cell Surface as Tools for Cancer Diagnosis, Prognosis and Therapy. Int. J. Mol. Sci. 2017, 18, 1232. [Google Scholar] [CrossRef]
- Pandey, G.; Fatma, T.; Cowsik, S.M.; Komath, S.S. Specific Interaction of Jacalin with Phycocyanin, a Fluorescent Phycobiliprotein. J. Photochem. Photobiol. B 2009, 97, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.D.; Goldstein, I.J. Binding of Hydrophobic Ligands to Plant Lectins: Titration with Arylaminonaphthalenesulfonates. Arch. Biochem. Biophys. 1983, 224, 479–484. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Lectins: Carbohydrate-Specific Proteins That Mediate Cellular Recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Astoul, C.H.; Zamboni, V.; Peumans, W.J.; Menu-Bouaouiche, L.; Van Damme, E.J.M.; Barre, A.; Rougé, P. Structural Basis for the Unusual Carbohydrate-Binding Specificity of Jacalin towards Galactose and Mannose. Biochem. J. 2002, 364 (Pt 1) Pt 1, 173–180. [Google Scholar] [CrossRef]
- Arockia Jeyaprakash, A.; Jayashree, G.; Mahanta, S.K.; Swaminathan, C.P.; Sekar, K.; Surolia, A.; Vijayan, M. Structural Basis for the Energetics of Jacalin–Sugar Interactions: Promiscuity Versus Specificity. J. Mol. Biol. 2005, 347, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, Q. Enhanced in vitro Anti-Cancer Activity of Curcumin Encapsulated in Hydrophobically Modified Starch. Food Chem. 2010, 119, 669–674. [Google Scholar] [CrossRef]
- Zhang, L.; Mu, C.; Zhang, T.; Yang, D.; Wang, C.; Chen, Q.; Tang, L.; Fan, L.; Liu, C.; Shen, J.; et al. Development of Targeted Therapy Therapeutics to Sensitize Triple-Negative Breast Cancer Chemosensitivity Utilizing Bacteriophage Phi29 Derived Packaging RNA. J. Nanobiotechnology 2021, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahmood, S.; Sapiezynski, J.; Garbuzenko, O.B.; Minko, T. Metastatic and Triple-Negative Breast Cancer: Challenges and Treatment Options. Drug Deliv. Transl. Res. 2018, 8, 1483–1507. [Google Scholar] [CrossRef] [PubMed]
- Latif, F.; Bint Abdul Jabbar, H.; Malik, H.; Sadaf, H.; Sarfraz, A.; Sarfraz, Z.; Cherrez-Ojeda, I. Atezolizumab and Pembrolizumab in Triple-Negative Breast Cancer: A Meta-Analysis. Expert Rev. Anticancer Ther. 2022, 22, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Shao, S.; Zhang, Y.; Luo, A.; Zhao, L.; Zhang, L.; Gu, S.; Zhao, X. BRD4 Inhibition Suppresses PD-L1 Expression in Triple-Negative Breast Cancer. Exp. Cell Res. 2020, 392, 112034. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, D.; Kreft, M.E.; Romih, R. Selective Binding of Lectins to Normal and Neoplastic Urothelium in Rat and Mouse Bladder Carcinogenesis Models. Protoplasma 2014, 251, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Zuraidah, M.A.; John, B.A.; Kamaruzzaman, Y. Cytotoxicity on mcf7 cell lines exposed to an extract of the jacalin from jackfruit seed. Sci. Herit. J. 2017, 1, 16–18. [Google Scholar] [CrossRef]
- Shrihastini, V.; Muthuramalingam, P.; Adarshan, S.; Sujitha, M.; Chen, J.-T.; Shin, H.; Ramesh, M. Plant Derived Bioactive Compounds, Their Anti-Cancer Effects and In Silico Approaches as an Alternative Target Treatment Strategy for Breast Cancer: An Updated Overview. Cancers 2021, 13, 6222. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug Resistance in Cancer: Role of ATP–Dependent Transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Passos, C.L.A.; Polinati, R.M.; Ferreira, C.; Dos Santos, N.A.N.; Lima, D.G.V.; Da Silva, J.L.; Fialho, E. Curcumin and Melphalan Cotreatment Induces Cell Cycle Arrest and Apoptosis in MDA-MB-231 Breast Cancer Cells. Sci. Rep. 2023, 13, 13446. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Smith, B.D.; Mohney, B.G. Ocular Side Effects Following Intravitreal Injection Therapy for Retinoblastoma: A Systematic Review. Br. J. Ophthalmol. 2014, 98, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.A.; Waseem, M.; Jamal, S.; Ahmed, N. Effects of Jacalin- a Galactose Binding Lectin on MDA-MB-468, a Triple-Negative Breast Cancer Cell Line, and Its Combinatorial Effect with Taxol; preprint, in Review. 2022. [Google Scholar] [CrossRef]
- Liu, J.-L.; Pan, Y.-Y.; Chen, O.; Luan, Y.; Xue, X.; Zhao, J.-J.; Liu, L.; Jia, H.-Y. Curcumin Inhibits MCF-7 Cells by Modulating the NF-κB Signaling Pathway. Oncol. Lett. 2017, 14, 5581–5584. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of curcumin.
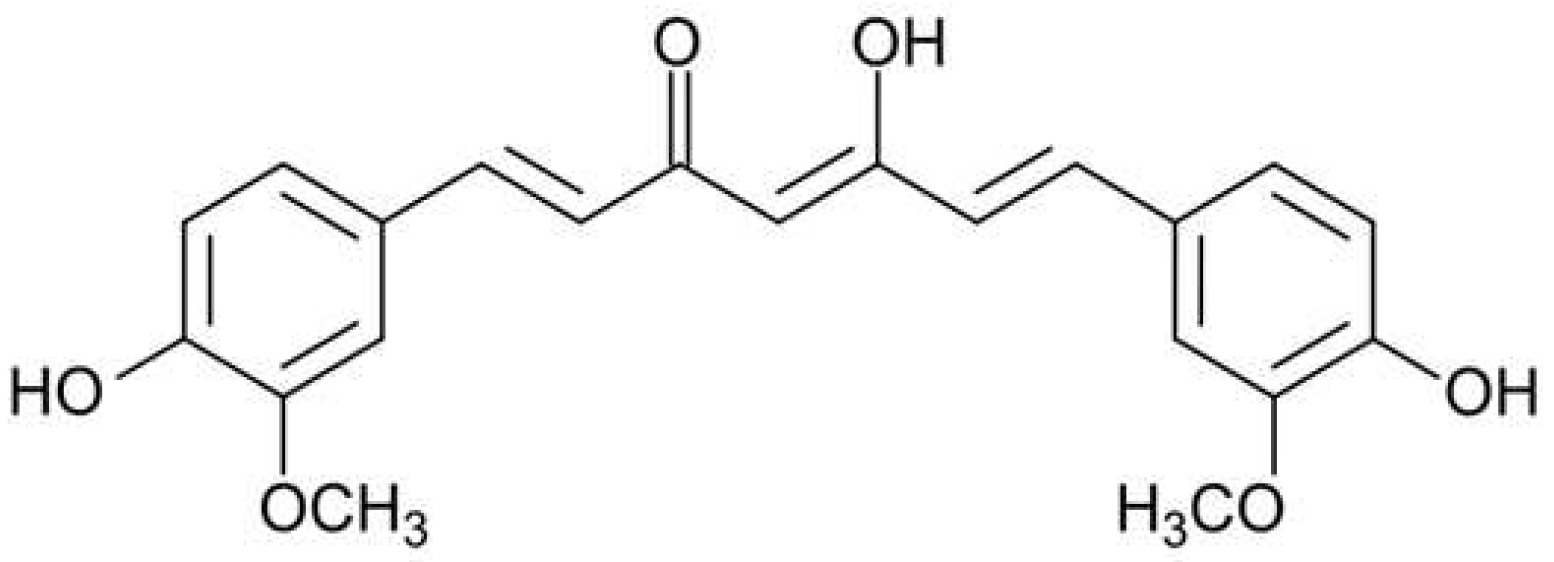
Figure 2.
(а) Fluorescence titration curve, showing the interaction of jacalin with curcumin. The curve is representative of 3 experimental sets. (b) Fluorescence emission spectra of 1 - curcumin (without jacalin), 2-9 curcumin with increasing concentrations of jacalin (0.28 µM - 2.42 µM).
Figure 2.
(а) Fluorescence titration curve, showing the interaction of jacalin with curcumin. The curve is representative of 3 experimental sets. (b) Fluorescence emission spectra of 1 - curcumin (without jacalin), 2-9 curcumin with increasing concentrations of jacalin (0.28 µM - 2.42 µM).
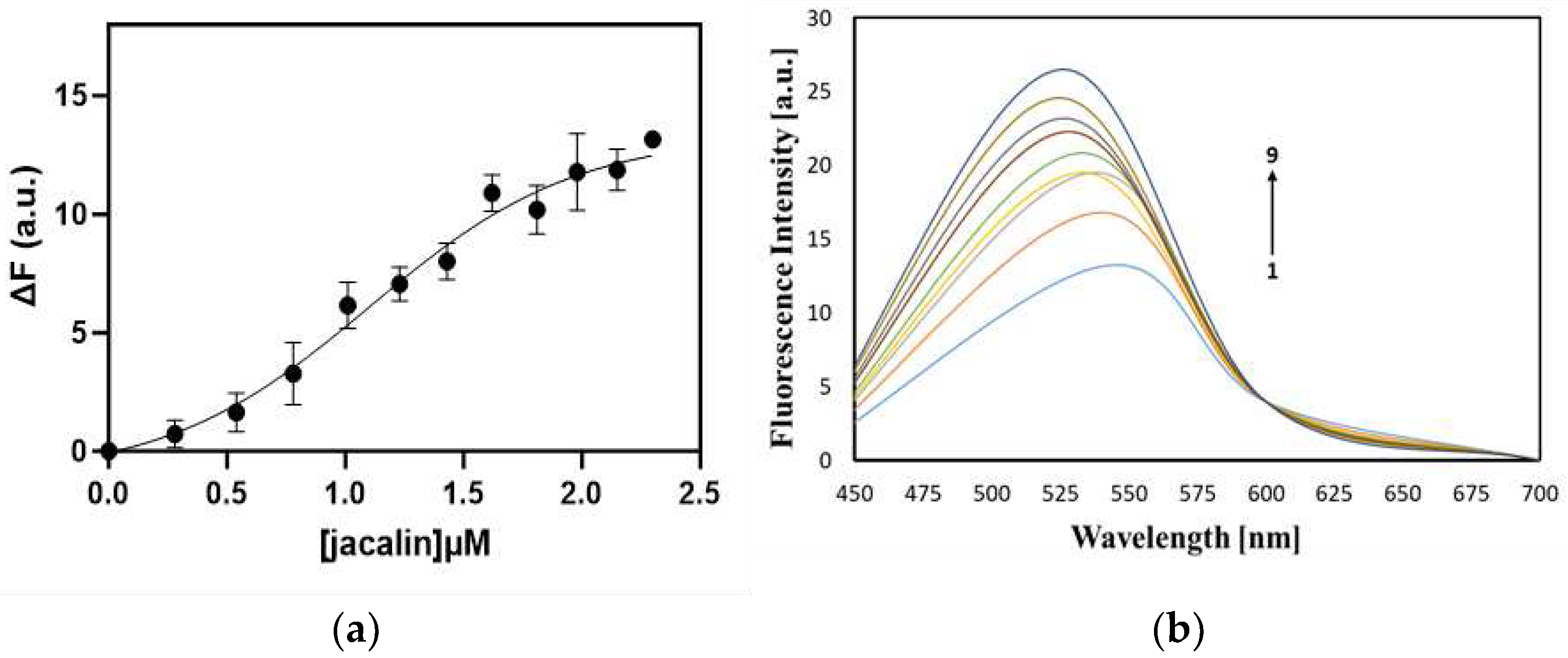
Figure 3.
Binding of jacalin with curcumin by Microscale thermophoresis (MST).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



