
Submitted:
24 November 2023
Posted:
24 November 2023
You are already at the latest version
Abstract
Maintaining the high overall health of farmed animals is a central tenant of their well-being and care. Intense animal crowding in aquaculture promotes animal especially in the absence of straightforward methods for monitoring their health. Here, we used bacterial 16S ribosomal RNA gene sequencing to measure population dynamics occurring during P. salmonis outbreaks. Our findings revealed a complex bacterial community consisting of a previously undescribed core pathobiome. Notably, we detected Aliivibrio wodanis and Tenacibaculum dicentrarchi on the skin ulcers of salmon infected with P. salmonis, while Vibrio spp. were enriched on infected gills. The prevalence of these co-occurring networks indicates the influence that coinfection plays in P. salmonis pathogenicity.
Keywords:
Piscirickettsia salmonis
; Salmo salar
; skin
; ulcer
; microbiome
1. Introduction
Salmonid rickettsial septicemia (SRS) is an intracellular infection caused by P. salmonis. This disease is of considerable importance for various salmonid species, Salmo salar) [1], and other fish species globally, including Epinephelus melanostigma [2], Atractoscion nobilis [3], and Dicentrarchus labrax [4]. SRS is a primary cause of both fish mortality and biomass loss within the Chilean salmon industry, prompting increasing antibiotic use. SRS infections present as ulcerative, systemic, and granulomatous, with muscle necrosis symptoms [5,6,7].
Closely monitoring fish skin health is vital in aquaculture for maintaining fish well-being. Skin variations, including ulcers, are crucial indicators of underlying health problems, and skin disorders are a prominent issue in aquaculture, contributing significantly to fish mortality. Approximately 1.1–2.5% of farmed fish succumb to ulceration [8], with an additional 0.7–3.8% down-classified at harvest due to reduced market quality [9].
Fish skin serves as a protective and dynamic tissue layer in constant interaction with the environment. It houses host-derived antimicrobial compounds and immunological components, acting as the primary defense against infectious pathogens [10,11]. Salmon skin is covered by a secreting mucosa. This layer is colonized by a complex microbial community. Evidence indicates that positive associations between mucosa-associated bacteria and the host immune system protects the fish from colonization by pathogenic bacteria [12]. Shifts in the skin microbiota due to environmental changes can lead to the dominance of opportunistic pathogenic bacteria, posing a threat to fish health [13]. The fish skin microbiome, in constant contact with the aquatic environment, facilitates microbial exchange with surrounding water and potentially other hosts [14]. Fish gills have a significant surface area per unit weight and represent the largest organ interfacing with the environment in teleost fish [15,16]. This feature makes them useful for monitoring fish health [17]. Some researchers, such as Koppang et al., 2015; Streit, 1998 [15,16], estimated that fish gills represent around 0.1 to 0.4 m2/kg of body weight. Gill tissues react quickly to unfavorable environmental conditions [17]. Several factors influence the composition of the gill microbiome, such as the fish health status and nutritional intake and the condition of the water [18,19,20]. Information on the gill microbiome is scarce, and comprehensive comparisons across various fish species, focusing on the gill and its external environment, are not widely available [21]. The rapid and acute responsiveness of gills to external alterations positions them as an optimal organ for monitoring fish health [22] and a possible point for identifying microbial biomarkers associated with gill health. The same also applies to fish skin, due to its high sensitivity. Understanding the dynamics of microbial communities within both the skin and gills is instrumental not only for the management of wild fish populations but also for enhancing the performance and growth of captive fish. This can facilitate the swift diagnosis of fish diseases, ultimately contributing to more effective aquaculture practices [18,23].
In contrast to extensively researched microbiomes like the human skin microbiome [24,25,26] and the fish gut microbiome, studies on the skin and, in particular, gill microbiomes in salmonids remain limited [27]. The body of literature focusing on the Salmo salar skin microbiome and its dynamics during infection outbreaks is notably sparse. Some studies have delved into gill microbiomes, exploring aspects such as the influence of Flavobacterium psychrophilum on susceptible and resistant lines [28] and the effects of ploidy and salmonid alphavirus infection [29]. For instance, Casadei et al. (2023) [30] conducted an evaluation of microbiota changes in groups of Atlantic salmon (Salmo salar), including both females and males, following treatment to eliminate infections caused by the ectoparasite Lepeophtheirus salmonis, commonly known as sea lice. In a study conducted in Chile [31], Valenzuela-Miranda et al. (2022) investigated the sea lice Caligus rogercresseyi as a possible vector for pathogenic bacterial groups within S. salar farms. Despite the evident significance of these mucosal microbiomes, there remains a relative dearth of research focusing on the microbiomes of salmonid skin and gills. This knowledge gap underscores the need for further investigations to unravel the complexities of these microbiomes and their responses to various environmental stressors and infectious agents.
For many years, it has been well recognized that microorganisms have a higher chance of survival within communities. However, this fundamental concept has long been overlooked in fish health diagnosis. The prevailing paradigm in aquaculture, influenced by Koch’s postulate [32], has traditionally focused on a one-disease, one-pathogen framework. Consequently, the diagnosis of coinfections involving fungal, bacterial, or viral agents in fish farms has been scarce. There are several factors contributing to this oversight, including the lack of adequate analytical techniques capable of detecting multiple pathogens simultaneously, a heavy reliance on traditional diagnostic methods primarily based on histological and pathological criteria, suboptimal sampling procedures, inadequate sample transportation, and a prevailing focus on analyzing fish farm mortality data. Some experts have emphasized the potential threats posed by coinfections involving opportunistic pathogens to aquatic animals, which can often persist and spread in aquatic environments, exacerbating health problems in wider fish populations [33]. In the context of such coinfections, P. salmonis stands as an illustrative case, with reports of coinfections involving this pathogen being notably scarce to date. Limited investigations have explored the relationship between P. salmonis and the parasite C. rogercresseyi [34,35,36]. Another study delved into the presence of different genogroups of the P. salmonis pathogen in Atlantic salmon samples collected from farms that experienced outbreaks of two distinct P. salmonis genogroups [37].
Continuing research on pathogen interactions and their impact on fish health is crucial for improving fish health management and supporting the aquaculture industry. The primary goal of our study is to investigate the alterations in the microbiome of Atlantic salmon skin and gills during ulcerative outbreaks caused by P. salmonis.
2. Materials and Methods
2.1. Fish Sampling
We conducted our analysis on Atlantic salmon (S. salar) during the grown-out phase, which had an average weight of 2869.34 g (±3160.42 g). Our focus was on ulcerated salmon, selected from seven distinct Chilean salmon farming sites, denoted as C1, C2, C3, C4, C5, C6, and C7, all located in the Los Lagos and Aysén Regions (Table 1). To establish a control group, we randomly chose twelve healthy fish from farm sites C8 and C9.
Our field sampling involved collecting three types of samples for microbiome analysis. Initially, we gathered swabs from S. salar individuals displaying skin ulcerations, swabs from the non-ulcerated skin areas of those fish with ulcers, and swabs from the first-gill arch of each fish. In the control group, we took samples from the midline of the fish’s skin and their first-gill arch. These samples were carefully stored in RNAlater transport medium within sterile Eppendorf tubes.
Samples of the anterior kidney, heart, and spleen were obtained from the salmon at farms (C1 to C7) to confirm the presence of P. salmonis in the fish from the infected centers. This confirmation was achieved through PCR analysis, following the method outlined by Karatas et al., 2008 [38]. Samples from farms C8 and C9 were also collected to validate the negative status of the control groups.
Finally, we collected samples of both skin and ulcerated skin for histological analysis. These tissue samples were preserved in 10% buffered formalin and were processed according to standard procedures. Sections measuring 3–4 mm were prepared and stained with hematoxylin and eosin (H&E), as per the method outlined by Prophet et al., 1992 [39]. This process allowed us to describe significant microscopic morphological changes.
2.2. Gross Pathology and Immunohistochemistry for Detecting Piscirickettsia salmonis in Skin Tissue
Veterinarians from the Centro de Investigaciones Biológicas Aplicadas (CIBA) carried out the gross pathology analysis, documenting external and internal gross lesions. Frequency of occurrence was determined using the methods outlined by Noga., 2010 [40]. Sections of healthy skin and ulcerated skin were preserved in 10% buffered formalin and subjected to immunohistochemistry staining to detect P. salmonis antigens. In our study, the detection of P. salmonis was achieved through immunohistochemical techniques adapted from the guidelines presented by Prophet et al. in 1992 [39]. We initiated the process by utilizing the monoclonal anti-Piscirickettsia salmonis antibody (Clone 7G4/D9) diluted 1:500 to specifically target the pathogen within skin tissue sections. These sections, sliced to a thickness of 3 µm from paraffin-embedded samples, were prepared on slides and left to dry overnight.
The deparaffinization step involved three successive xylene washes, each lasting three minutes. We then transitioned to a series of ethanol solutions, decreasing in concentration from 100% to 50%, to rehydrate the sections. This was followed by rinsing in deionized water. For the antigen retrieval phase, we placed the slides in a steaming container with sodium citrate buffer for 40 min. After a quick rinse in phosphate buffered saline (PBS) provided by Merck, we applied a 3% hydrogen peroxide solution for 10 min within a humid chamber to block endogenous peroxidase activity. After a subsequent rinse through immersion in PBS, we blocked endogenous proteins using either a protein blocker or bovine serum albumin (BSA protein) sourced from Thermo Fisher, also for a duration of 10 min, in a moist environment. The primary antibody incubation was conducted with the Clone 7G4/D9 monoclonal antibody at a 1:500 dilution for 40 min, maintaining the slides in a humid chamber heated to 45 °C to prevent drying. Post-incubation, we utilized the HiDef Detection™ HRP Polymer System (Sigma Merck, Product Number: 954D), following the manufacturer’s guidelines. This step was crucial for amplifying the visualization of the antigen–antibody interactions within the tissue sections.
Subsequent to this, we halted the staining process by washing the slides with distilled water. Counterstaining was performed using Mayer’s hematoxylin for three minutes, followed by rinsing in tap water for five minutes. The final preparation for microscopic examination involved mounting the slides with a glycerol-based, water-soluble mounting solution. Control slides, which included P. salmonis tissues, were processed identically, except for the substitution of the primary antibody with normal rabbit serum.
2.3. DNA Isolation, 16S rRNA Gene Amplification, and Sequencing
DNA was isolated from tissue samples of skin, ulcers, and gills using the Qiagen DNA Microbiome extraction kit (Qiagen, Catalog No. [51704]), following the guidelines provided by the kit manufacturer. The DNA’s concentration was determined with 2 µL of each sample, utilizing the Invitrogen Qubit dsDNA BR Assay kit (Invitrogen, Catalog No. [Q33265]). We amplified the V4 hypervariable region of the bacterial 16S rRNA gene from each DNA sample. This was achieved using a primer set specifically targeting the V4 region: 515F/806R. The PCR process for each sample involved a 35-cycle reaction using the HotStarTaq Plus Master Mix Kit (Qiagen, Catalog No. [203645]), with a specific temperature and time protocol. Following PCR, the amplicons from different samples were combined in equal measures and purified. Sequencing was then performed on the Illumina NovaSeq system, adhering to the provided kit protocols. Finally, all the sequenced data were uploaded to the NCBI Sequence Read Archive under the BioProject code PRJNA1044012.
2.4. Data Filtering, Amplicon Sequence Variants Production, and Taxonomic Assignment
The raw sequencing data were processed and refined using Fastp version 0.20.1 [41], which involved trimming the initial 10 nucleotides and excluding sequences containing any ambiguous nucleotides (Ns). A sliding window approach from right to left, with a window size of 4, was employed to eliminate fragments with a quality score (Q-score) below 30. The resulting high-quality sequences were then subjected to de-noising, removal of chimeric PCR artifacts, and merging. This was accomplished using DADA2 version 2.1.18 [42], a package available in R, resulting in refined 16S amplicon sequence variants (ASVs). Taxonomic classification of these ASVs was carried out using the same R package, DADA2, with the SILVA database version 138.1 [43] as the reference. The classified taxonomic data and ASV read counts were compiled into a phyloseq version 1.4 [44] object to facilitate subsequent analyses. For assessing alpha diversity, we applied rarefaction to the samples and analyzed them using the phyloseq version 1.4 package in R. To visualize the diversity, rarefaction curves were generated using the “rarecurve” function from the vegan package version 2.6.2 [45] in R.
2.5. Beta Diversity: Nonmetric Multidimensional Scaling (NMDS) Analysis
To analyze 16S rRNA amplicon sequencing variants (ASVs), we employed non-metric multidimensional scaling (NMDS) using the Bray–Curtis method. The Bray-Curtis dissimilarity index was computed for each sample using the “vegan” R package, specifically employing the “adonis2” function. Subsequent NMDS plots displayed differentiation between controls and fish with skin ulcers. The skin, gills, and ulcers of infected fish were represented as green dots, while the skin and gill mucosa of control salmon were shown as blue-green dots. Furthermore, permutational multivariate analysis of variance (PERMANOVA) was conducted on distance matrices (p = 0.001), underscoring dissimilarities (beta diversity) between the two groups under investigation: the ‘healthy’ Atlantic salmon control group and the P. salmonis-infected Atlantic salmon.
2.6. Alpha Diversity: Shannon Index
To assess species diversity within the ecological communities present in our samples, we employed the Shannon diversity index, also known as the Shannon–Wiener index. This metric elucidates two principal facets of diversity: species richness and the even distribution of individuals across these species. Calculations were executed using the “vegan” R package across the five investigated tissues. Subsequently, Bray–Curtis analysis was performed at the tissue level, comparing the “healthy” Atlantic salmon control group to the P. salmonis-infected Atlantic salmon group (p = 0.001). Notably, the resulting analysis presented distinct separations among the sample types, even though some overlap was observed between them.
2.7. Linear Discriminant Analysis (LDA)
We utilized the LEfSe (linear discriminant analysis effect size) algorithm [46] to discern significant taxonomic differences among tissues. The LEfSe algorithm involves performing a Wilcoxon test for each taxonomic group, identifying amplicon sequence variants (ASVs) that stand out in their abundance. In line with the criteria set for our study, ASVs with p-values less than 0.05. Employing this approach enabled us to pinpoint ASVs that played a key role in differentiating the microbial profiles of the respective tissues.
2.8. Co-Occurrence Network Analyses
We employed cooccur v1.3 [47] for an in-depth co-occurrence network analysis ensuring adherence to a stringent p-value threshold of 0.05. The derived networks were then rendered visible with visNetwork v2.1 (https://datastorm-open.github.io/visNetwork/) accessed in 2021. Further examination and insights into these networks were achieved through the NetShift v1 [48] tool. A probabilistic model was utilized in our co-occurrence network analysis to further delve into the inter-relationships of operational taxonomic units (ASVs) within the healthy skin and ulcers of Atlantic salmon. Comparative networks for infected gills vs. healthy tissue and infected skin vs. healthy tissue were also studied, with the results presented in Appendix D and E. The aim in this stage of our analysis was to assess the prevalence and patterns of ASV co-occurrences within each tissue type. These co-occurrences were categorized as:
Positive Associations (Green): Signifying co-occurrence frequencies that surpassed expectations, hinting at potential cooperative interactions between ASVs.
Negative Associations (Red): Indicating co-occurrence frequencies below expected levels, suggesting potential competitive or exclusionary relationships.
Random Associations (Blue): Representing co-occurrence frequencies that aligned with expectations.
Within the network structure, the size of each node was determined based on the scaled NESH (Neighbor Shift) score. Notably, nodes transitioning from control to case scenarios and turning red were identified as dominant “drivers” within the entire network, playing a significant role in shaping microbial interactions.
3. Results
3.1. Gross Pathology and Immunohistochemistry, PCR
The fish exhibited scaling, raised scales, single to coalescent ulcers with a distinct white border, exposure of musculature, and variable sizes (Figure 1). Internally, the fish showed an absence of food in the digestive system, splenomegaly, and varying degrees of renomegaly. Visceral fat congestion was observed, and in a reduced percentage of the sampled fish, hepatomegaly and white nodules were present in the liver. Histologically, the analyzed cutaneous tissue showed a disruption of the epidermis and partial involvement of the dermis. Additionally, inflammation, hemorrhages, and necrosis of the epidermis, dermis, and hypodermis were observed. The application of the immunohistochemical (IHC) technique, using monoclonal antibodies specific for P. salmonis on paraffin-embedded tissues, revealed a positive reaction (brown color), indicating the presence of coccoid structures consistent with the presence of P. salmonis in the skin lesions (Figure 2).
3.2. Beta Diversity
Bray–Curtis Method
Figure 3 shows a non-metric multidimensional scaling (NMDS) plot derived from a Bray–Curtis analysis, delineating the microbial communities of Atlantic salmon under two distinct conditions: healthy and P. salmonis-infected. The microbiota profile of healthy salmon, represented by red dots, show predominant occupation in the upper left quadrant of the plot, with a relatively tight clustering suggesting a consistent microbial community composition among the healthy samples. Conversely, the microbiota associated with P. salmonis-infected salmon, depicted as blue-green dots, are largely situated in the lower right quadrant. This distribution might indicate varying degrees of infection among the samples, as suggested by the more dispersed nature of the blue-green cluster. The notable intersection of red and blue-green dots in the central region of the plot underscores the microbial communities common in both conditions, potentially hinting at a transitional microbiota state or the presence of foundational microbes resilient to infection. The axes NMDS1 (X-axis) and NMDS2 (Y-axis), spanning values from −0.50 to 0.50, facilitate the spatial representation of microbial dissimilarity. In the NMDS plot, the spatial arrangement of points carries significance. Points situated closely together signify greater similarity in microbial community composition, while those spaced further apart suggest distinct microbial compositions. Figure 3 offers an in-depth juxtaposition of the microbial landscapes in the salmon under the two conditions.
In Figure 4, the Nonmetric Multidimensional Scaling (NMDS) plot, derived from the Bray–Curtis dissimilarity matrix, visualizes microbial community diversities across various tissue levels in Atlantic salmon. For each data point, its placement on the NMDS plot represents the microbial composition, with the position on the NMDS1 (horizontal) and NMDS2 (vertical) axes indicating the primary axes of microbial variation.
Healthy skin samples, depicted in blue-yellow, predominantly cluster in the right quadrant, indicating positive values on the NMDS1 axis. This placement is emblematic of a unique microbial composition intrinsic to healthy skin tissues. Conversely, the infected skin samples, in blue-green, show more dispersion but still tend toward the positive extremities of the NMDS1 axis. Their proximal positioning to the healthy skin samples on the plot suggests shared microbial taxa, but with evident compositional nuances.
The infected ulcer samples, represented in pink, resided toward the lower values on the NMDS2 axis, often converging near the central to the negative spectrum of the NMDS1 axis. This positioning accentuates a significant departure in their microbial communities, potentially indicative of a unique microbial consortium associated with ulcerated conditions.
There is clear divergence between healthy gills (in red) and infected gill samples (in green) along the NMDS1 dimension. Healthy gill samples are predominantly situated toward the upper-right quadrant, demonstrating positive values on both NMDS axes. In stark contrast, the infected gill samples are more centrally sprawled, with many straying into the negative domain of the NMDS1 axis. This spatial demarcation reflects the microbial perturbations induced by P. salmonis infections in gill tissues.
The arrangement of samples in Figure 4 offers a detailed insight into the microbial dynamics across different tissue types in Atlantic salmon. Their distinct positions along the NMDS axes highlight the variations and commonalities in microbial composition.
3.3. Alpha Diversity
Shannon Index
Following alpha diversity analysis of the microbial profiles comparing between healthy skin (indicated in yellow) and infected ulcer (illustrated in pink), a pronounced differentiation was observed (Figure 5). A Wilcoxon test corroborated this observation, showing a statistically significant divergence with a p-value of 0.00014. This divergence suggests a profound alteration in the microbial ecosystem of the skin post-ulceration, likely indicative of a shift in the dominant microbial species or the emergence of rare taxa.
Contrastingly, when examining the microbial landscape of healthy skin (yellow) against infected skin without ulceration (blue-green), the divergence, though less pronounced, remains significant. A p-value of 0.042 from the Wilcoxon test reaffirmed this distinction. It hints at initial changes in the microbial composition due to infection, even before ulcerative manifestations appear.
The results from comparison of the microbial profiles between infected skin (represented in blue-green) and infected ulcers (depicted in pink) introduce additional intricacy. Although both conditions result from infection, they display distinct microbial diversities, as underscored by a p-value of 0.0016. Such variance indicates the microbiota’s dynamic adaptation or response to the advancing stages of the disease.
Lastly, upon inspection of the healthy gills (depicted in red) and the P. salmonis-infected gills (shown in green), a distinct pattern emerges. Both gill tissues appear to have similar alpha diversity. However, the alpha diversity values revealed that the gills of infected fish possessed a numerically higher level of diversity. The underlying reasons for this increased diversity are not immediately apparent, but it could indicate a microbial defense mechanism in action, or perhaps it is the result of specific microbial species taking advantage of the infected environment.
3.4. Taxonomic Assignment
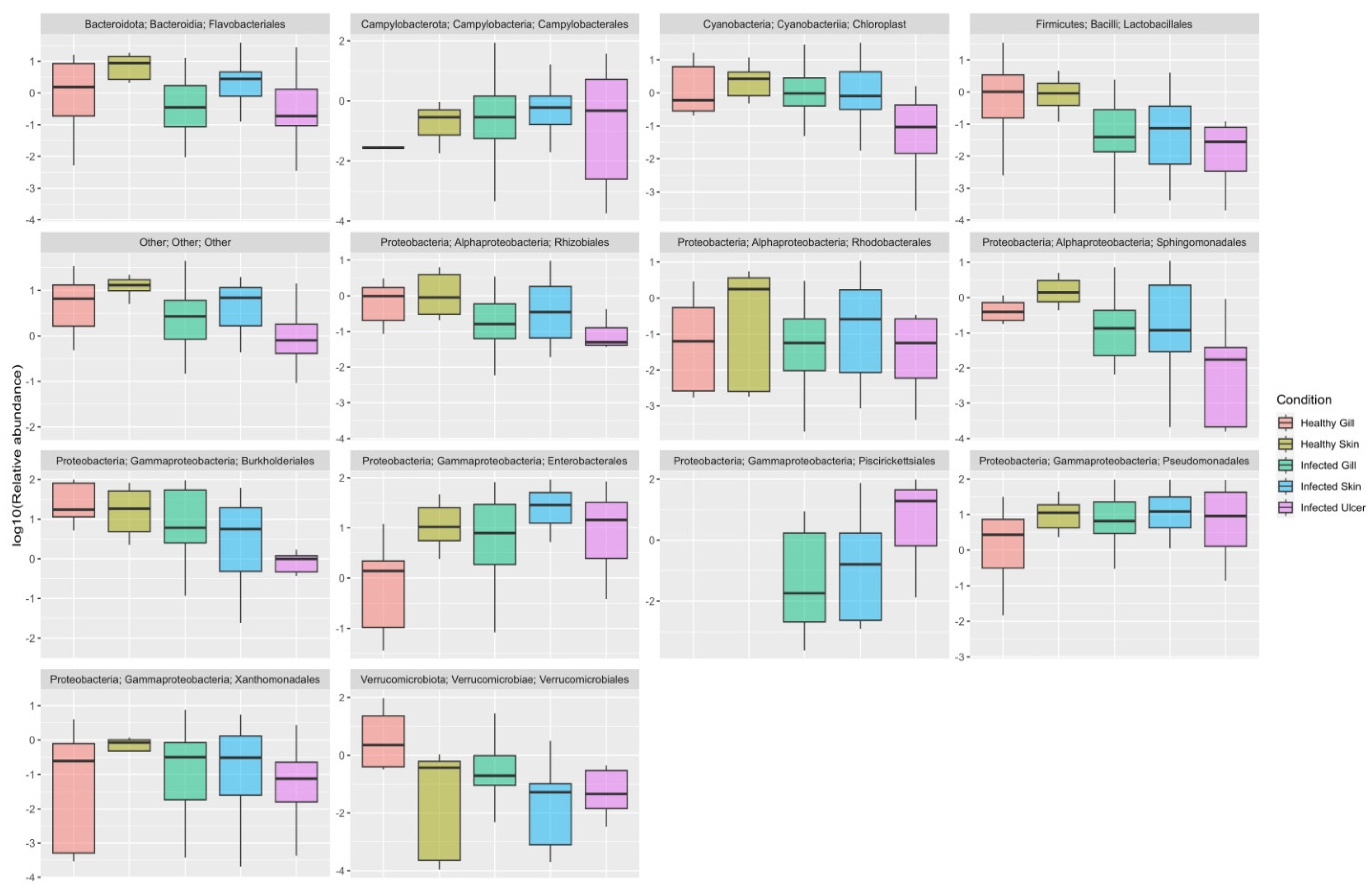
Figure 6 presents the relative abundance of the bacterial microbiome. At the phylum level, each analyzed tissue in Atlantic salmon exhibits a prevalence of the phylum Proteobacteria (74.41%), followed by the phyla Bacteroidota (5.33%), Verrucomicrobiota (3.59%), and Campylobacterota (2.61%). Furthermore, at the order level, a clear differentiation is evident among the samples analyzed. Notably, the presence of the Piscirickettsiales, Flavobacteriales, and Pseudomonales orders was observed in the infected skin and ulcer samples. Furthermore, an overabundance of the genus Pseudomonas spp. was noted in the infected microbiome samples. These particular bacterial groups are commonly associated with freshwater infections and are often indicative of microbial dysbiosis in fish.
At the species level, our primary aim is to detect the presence of the pathogen P. salmonis within each of the analyzed tissues. To achieve this, we analyzed the amplicon sequence variants (ASVs) specific to this microorganism, as illustrated in Figure 7. Our examination revealed the presence of P. salmonis in all three infected tissues, with ASVs counts of 48 in the gills, 54 in the skin, and 53 in the ulcers. Importantly, no ASVs (N.D) corresponding to the pathogen were detected in the tissues of the control groups. This same analysis was extended to the level of bacterial families for all tissues, as presented in Appendix C, further confirming this observation.
3.5. Linear Discriminant Analysis (LDA)
From the LDA analysis, it is evident that the ulcers in the Atlantic salmon were primarily colonized by P. salmonis, Tenacibaculum dicentrarchi, and Aliivibrio wodanis (Figure 8). This particular grouping of bacteria indicates a close correlation between these taxa and the ulcerative state observed in Atlantic salmon. In the infected skin tissue, there is a significant presence of the genus Vibrio spp. The predominance of this genus in the infected skin tissue suggests its potential role in skin infections in salmon. On the other hand, the healthy skin tissue showcases a more diverse microbial profile, characterized by taxa such as Photobacterium, Psychrobacter, and Ralstonia. This assortment of microbes implies a balanced microbial community, possibly contributing to the health and protective functions of the salmon’s skin. Observing this genus in both infected skin and gill tissues might indicate a shared pathogenic route or mechanism affecting both these sites during an infection episode. Contrarily, the microbial profile of the healthy gill, as inferred from the LDA analysis, is markedly distinct from its infected counterpart. The noticeable lack of explicitly pathogenic taxa like Vibrio spp. in the healthy gill may indicate equilibrium in terms of microbial composition.
3.6. Co-Occurrence Network
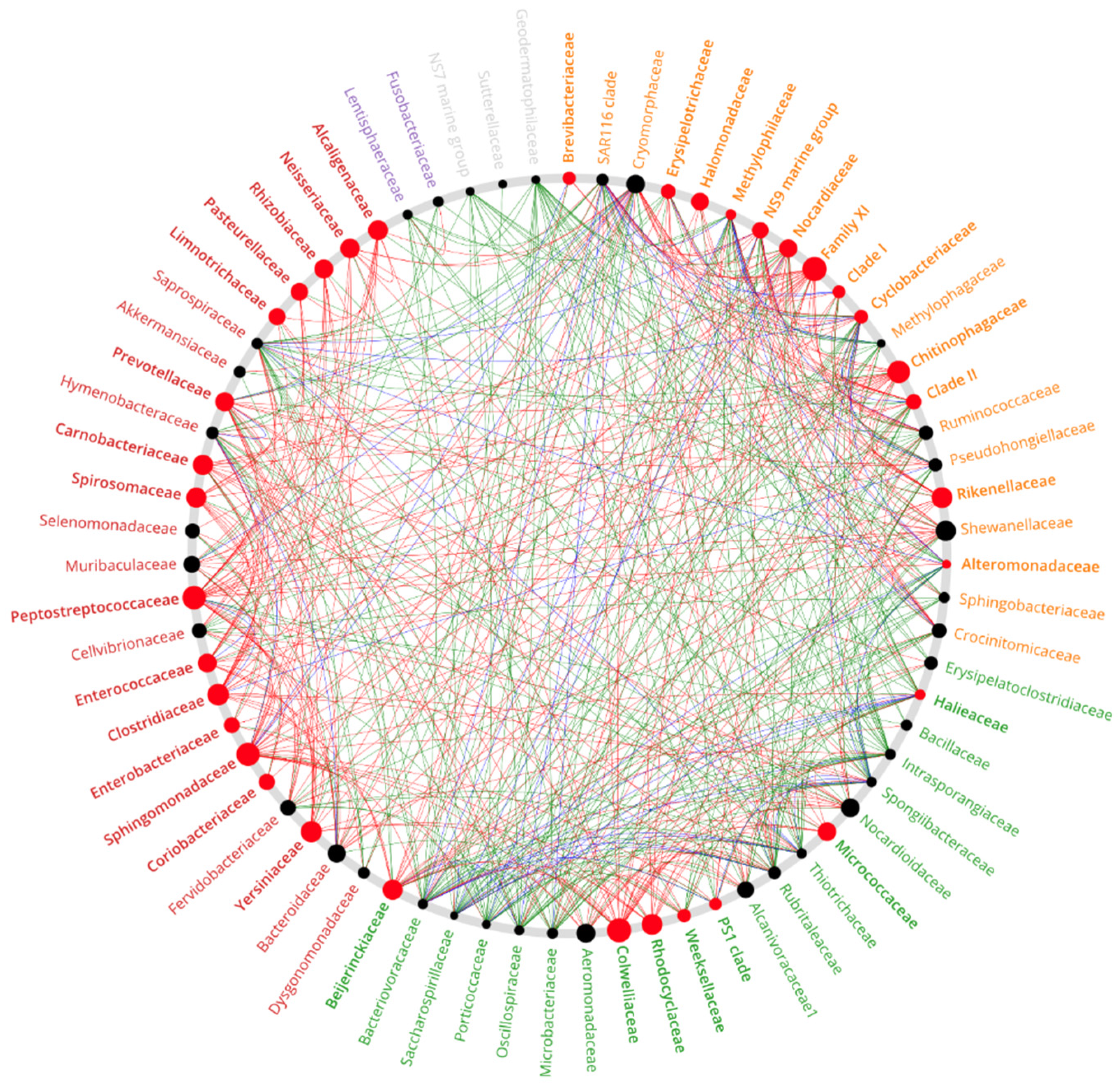
Our analysis of microbial co-occurrence networks in Atlantic salmon revealed intricate patterns. For the healthy skin (Figure 9), we discerned a substantial network comprising 3051 positive and 161 negative interactions, with the dominance of positive correlations hinting at pronounced microbial diversity-dependent effects in maintaining skin health. These findings are consistent with Friedman and Alm (2012) [49], indicating that two perfectly correlated ASVs are operating in synergy. Within this framework, families like Rhodocyclaceae, Micrococcaceae, and Carnobacteriaceae, marked by high NESH scores, stand out as potential beneficial microbes for skin health.
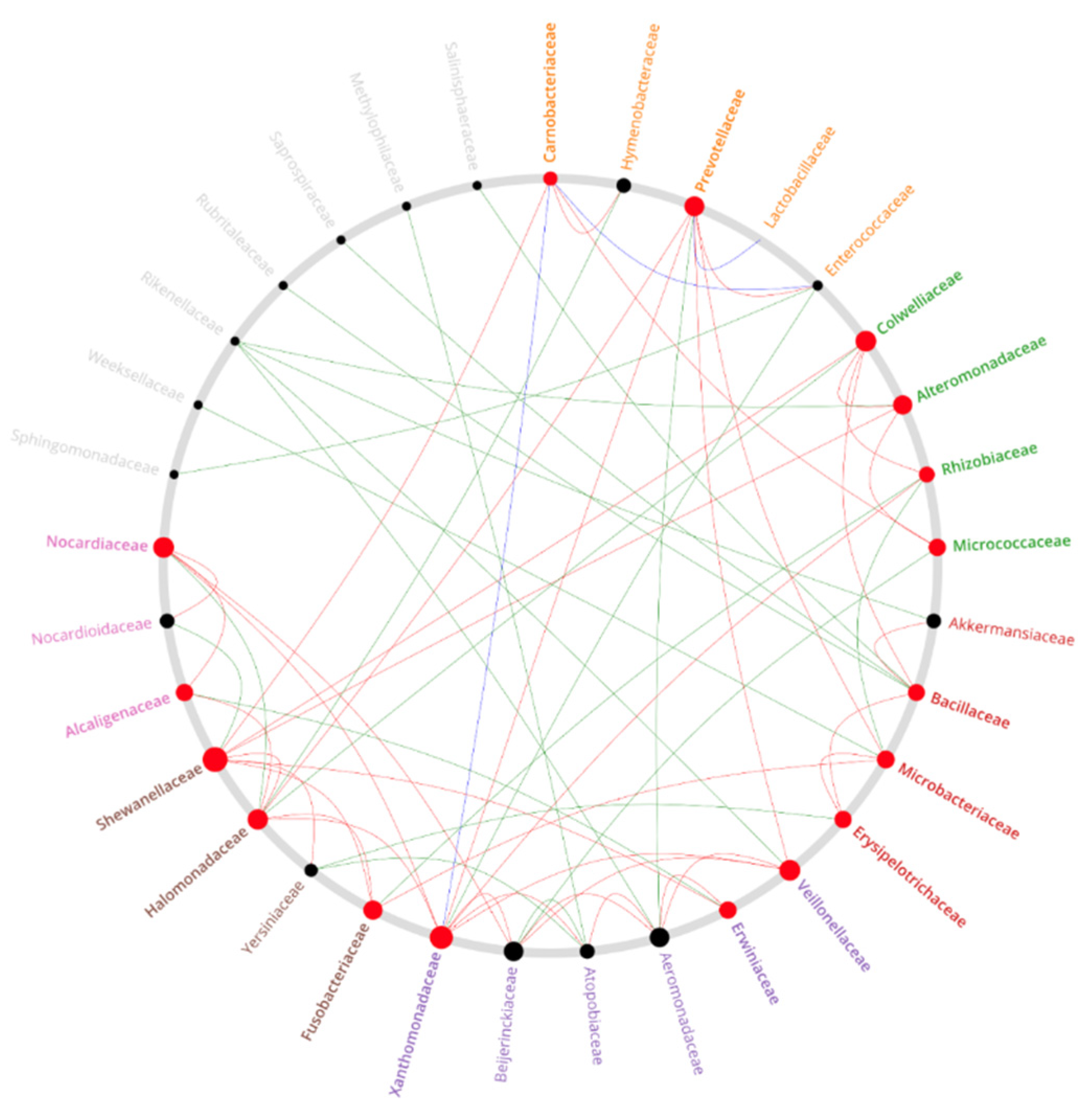
For our analysis of the gills (Appendix D), characterized by a relatively lower bacterial abundance, 161 positive and 87 negative interactions were identified. Taxa with significant NESH scores, specifically Micrococcaceae, Rhizobiaceae, and Akkermansiaceae, manifested predominantly positive interactions, underlining cooperative dynamics. In stark contrast, families like Nocardiaceae and Alcaligenaceae emerged as central to the competitive dynamics.
The infected skin (Appendix E) showcased a remarkable interaction network, with 2282 negative and 1222 positive interactions. Herein, among the bacterial pathogen P. salmonis ASVs detected in infected salmon tissues, 18 positive and 48 negative interactions were observed, highlighting potential dysbiosis. This pattern, marked by the nearly equal negative and positive interactions among ASVs, underscores the intricacies of infection dynamics.
4. Discussion
In our study of the Atlantic salmon skin microbiome during P. salmonis outbreaks, we discerned notable differences between the control and infected fish using beta diversity analysis, specifically PCoA and PERMANOVA analysis (see Figure 3 and Figure 4). Progression of the ulcerative process due to P. salmonis is displayed in Figure 4. There is a stark contrast in the microbiome of healthy skin of salmon in control groups compared with ulcerated skin. Infections without ulcers fall in between these extremes in the representation. Yet, this pattern is not always consistent, as fish species and environmental conditions seem to impact it. For instance, Kashinskaya et al., 2021 [50], while studying Prussian carp and its ectoparasitic crustaceans, Lernaea cyprinacea and Argulus foliaceus, aimed to understand the microbiome alterations during coexisting ulcers and parasitic infestations. Their beta diversity analysis showed there were no significant differences in the microbiota of intact skin mucosa in affected fish, suggesting ulcers and ectoparasites did not alter the microbiome. However, they noted negative correlations between certain bacterial species and the number of ulcers. In our research, we found marked differences between the healthy and ulcerated skin of Atlantic salmon (with p-values of 0.00014 and 0.0016, respectively), indicating a substantial impact of the ulcerative process on skin microbiome diversity.
Our examination of specific taxonomic groups by lesion type revealed surprising bacterial group involvement in the outbreak. Specifically, in advanced ulcerated skin stages, we found an overrepresentation of four bacterial families (refer to Figure 6): Vibrionaceae, Flavobacteriaceae, Piscirickettsiacea, and Pseudomonadaceae, all previously recognized as pathogens or opportunists in aquaculture. One of our most notable discoveries was the detection of the aquatic pathogen A. wodanis from the Vibrionaceae family (see Figure 8) in this Atlantic salmon ulcerative outbreak in Chile. Though A. wodanis is found in the core microbiome of the parasite C. rogercresseyi in Chile [51], it has not been a significant health issue for Chilean salmon farms. It predominantly affects salmon farms in Norway [52] and Canada [53]. A. wodanis is associated with ulcerative skin problems in farmed fish [50] and is considered a pathogen linked to winter ulcer disease in Atlantic salmon in the northern hemisphere.
The specific pathogenic role of A. wodanis in winter ulcer disease is still controversial. While some research points to Moritella viscosa as the primary pathogen behind winter ulcer outbreaks [47,48,49,50,51,52,52,53,54,55,56], the relationship between A. wodanis and M. viscosa is complex. For example, Karlsen et al., 2012 [57] explored the dynamics between these bacteria. They observed that the diminished presence of A. wodanis in healthy fish skin might be due to the propensity of M. viscosa to colonize ulcerated skin. Their findings imply that M. viscosa colonization might require pre-existing skin damage, often associated with secondary infections like “fin rot.” Additionally, the study raised the possibility that A. wodanis might produce toxins that could further compromise colonized surfaces, making them more prone to infections. Hjerde et al., 2015 [58] arrived at a similar conclusion in their research, emphasizing the multifaceted nature of winter ulcer disease, where “winter ulcer” typically refers to infections by M. viscosa. In a more recent study, Maharajan et al., 2021 [59] delved into the quorum sensing (QS) system of A. wodanis. They uncovered that A. wodanis produces virulence factors that might encourage inter- or intraspecies interactions, furthering competition and adaptation during the progression of winter ulcers.
We did not identify A. wodanis alongside M. viscosa, as M. viscosa is not a known issue in Chilean farms. However, we did identify T. dicentrarchi as a prominent microorganism in Atlantic salmon ulcers, as shown in Figure 8. This bacterium was first isolated from skin lesions of the European sea bass in Spain. Subsequent studies, like Olsen et al., 2017 [60], indicated that in Norway, other Tenacibaculum spp. may overshadow T. dicentrarchi. This research also underscored the potentially higher pathogenicity of T. dicentrarchi in non-salmonid fish. Tenacibaculum spp. has been recognized as a genus grouping various opportunistic pathogens, where factors such as dysbiosis, isolate variation, virulence, and host genetics can influence the colonization of affected fish skin [61,62]. Tenacibaculum spp. is often identified as part of various fish coinfection outbreaks. For instance, Apablaza et al., 2017 [63] identified Tenacibaculum maritimum as the causative agent of Tenacibaculosis in Chilean Atlantic salmon (S. salar), and this was associated with a Pseudochattonella spp. algal bloom. Similarly, Avendaño-Herrera., 2006 [64] noted that during Tenacibaculosis outbreaks in Platax orbicularis, the presence of T. maritimum was commonly associated with co-occurrences of other pathogenic genera, specifically Vibrio spp., aligning with our findings. It has been suggested that certain Tenacibaculum species might be an integral part of specific fish species microbiomes, thereby creating a stable pathobiome that harmonizes with the host. Data from Wynne et al., 2020 [61] support this notion. In their study, T. maritimum was observed in the skin microbiome of healthy Atlantic salmon smolt as well as in those recovered from Tenacibaculum infection. This implies the remarkable adaptability of T. maritimum within its ecological niche. Moreover, several researchers have emphasized the presence of multiple species within the Tenacibaculum genus [65] in Atlantic salmon, alongside Vibrio spp. and Aliivibrio [55,66,67]. The diversity of phylogenetic relationships within Tenacibaculum sequences in our research further bolsters the idea of varied Tenacibaculum spp. cultures on Atlantic salmon skin. However, further supporting evidence is required before it can be categorically stated that Tenacibaculum spp. is a constant presence in the healthy microbiome. The fish skin microbiome is dynamic, with variations influenced by species, developmental stage [68], seasonal changes [68], water temperature [69], salinity [70], and geography [71]. As a result, the prevalence and diversity of Tenacibaculum spp. on the skin of healthy Atlantic salmon may exhibit considerable variation across different regions or farming areas, characterized by the unique environmental conditions of each.
Another bacterial genus of note in fish with skin ulcerations is Pseudomonas. Generally viewed as opportunistic pathogens, many species within this genus are recognized as primary agents of diseases in farmed fish, including notable species like Pseudomonas aeruginosa, Pseudomonas anguilliseptica, Pseudomonas chlororaphis, and Pseudomonas fluorescens, among others, as noted by Austin et al., 2016 [72]. In trout farming, P. fluorescens [73] and Pseudomonas putida [74] have posed significant challenges. In studies such as including that by Rao et al., 2019 [75], the significant role of Pseudomonas spp. in fish disease outbreaks has been highlighted. Many of these species are known to produce exopolysaccharides, which are essential for biofilm formation [76]. An illustrative example is P. aeruginosa, known to possess the pyoverdine siderophore biosynthesis gene. This gene is essential for iron acquisition and biofilm development in multi-bacterial environments. This aligns with findings from Levipan et al., 2019 [77], who found that T. dicentrarchi strains can form biofilms in vitro, hinting at a potential synergistic relationship between Pseudomonas spp. and Tenacibaculum spp. Cornelis and Bodilis, 2009 [78] detailed the mechanism by which P. aeruginosa harnesses siderophores from other bacteria to import iron through various receptors. Many pathogens have evolved refined iron acquisition methods to counteract host-imposed iron limitations. In a study by Avendaño-Herrera et al., 2005 [79], for instance, two distinct iron acquisition systems were discovered in T. maritimum. While one involves siderophore synthesis, the other facilitates heme group utilization as an iron source. Calquin et al., 2017 [80] further showed that P. salmonis produces functional siderophores for iron uptake. However, the specific interactions and iron utilization tactics of these pathogens in Atlantic salmon ulcers are still unclear.
P. salmonis was found to be the most prevalent among the ASVs in the infected and ulcerated tissues of Atlantic salmon (see Figure 7). This is consistent with its association with ulceration outbreaks in Chilean fish farms. Figure 9 illustrates the complexity of microbial diversity within ulcers and the intricate interactions among various bacterial families. Yet, the exact mechanisms driving these interactions between bacterial taxa are still unclear. To understand the microbial dynamics during ulcerative outbreaks fully, it is essential to explore how P. salmonis enters Atlantic salmon.
Previous research indicates that infections mainly occur in salmonids through the skin and gills, as suggested by Smith et al., 1999 [81] and Smith et al., 2004 [82]. They showed that P. salmonis can invade Coho salmon (Oncorhynchus kisutch) via the skin and mucous membranes, even without exhibiting visible damage. They hypothesized that P. salmonis may begin its infection process by adhering to microscopic lesions, which are common in cultured salmon [83]. This suggests the skin is a potential primary entry point for this bacterium in salmonid fish.
Notably, there are limited reports of coinfections involving P. salmonis with other pathogens in aquaculture. In previous studies, coinfection interactions between P. salmonis and the parasite C. rogercresseyi have been explored in Chile [28,81,84,85]. However, bacterial coinfections have not been given the same attention, often due to the prevailing belief that outbreaks result from a single pathogenic agent. This oversight becomes even more significant given the diverse range of pathogens we have identified in the affected salmon tissues, suggesting possible bacterial coinfections.
While we have illuminated the microbial dynamics at the advanced stages of ulcerative processes in Atlantic salmon, we may have missed the subtleties of microbial interactions in their early stages. Identifying multiple pathogens accentuates the importance of discerning their collective impact on infections, a dimension we did not deeply explore. We did not consider potential external influencers of microbial communities like water temperature or farming practices. Furthermore, while our genomic methodology was key in identifying specific microbial taxa, it does not guarantee their viability, activity, or exhaustive detection.
5. Conclusions
In conclusion, our examination of the ulcerative outbreak in farmed Atlantic salmon linked to P. salmonis indicates the presence of a multifaceted bacterial community, with revelation of a core pathobiome that was previously unidentified. This underscores the potential of 16S rRNA analysis in discerning intricate bacterial interactions during infections. Comprehending the dynamics of pathogenic evolution in ulcers is pivotal for demystifying the infection mechanism of P. salmonis and tracing its colonization pathway within the host. This discovery raises pertinent questions about the specific roles of the detected infectious and opportunistic taxa, notably Vibrionaceae, Flavobacteriaceae, Piscirickettsiacea, and Pseudomonadaceae, in the ulcerative onset. Thus, further research, incorporating both in vitro and in vivo methodologies centered on the identified species, T. maritimum, P. salmonis, and A. wodanis, is essential.
Author Contributions
Conceptualization, M.G., R.S., Y.C. and C.S.-N.; methodology and study design, M.G., J.P.P. and R.S.; sampling, D.C. and R.S., bioinformatic analysis, V.M.-C. and R.A.-C.; interpretation, Y.C., M.G. and M.M.d.O.; writing—original draft preparation, Y.C. and I.B.; writing—review and editing, M.G., C.S.-N., L.R.-C., J.W.B. and Y.C.; supervision, C.S.-N. and M.G. All authors have provided their consent and agree to the final version of the manuscript as submitted for publication.
Funding
This study was supported by Centro de Investigaciones Biológicas Aplicadas.
Institutional Review Board Statement
The Centro de Investigaciones Biológicas Aplicadas (CIBA) in Puerto Montt, Chile, is certified for aquaculture diagnostic services. All associated experimental protocols received approval from the Chilean National Fish and Aquaculture Service (Servicio Nacional de Pesca y Acuicultura de Chile, http://www.sernapesca.cl/, accessed on 10 November 2022). Our procedures complied with Chile’s Ley 20.380 de Chile sobre Protección de Animales, concerning animal welfare in biomedical research, and closely aligned with the EU Directive 2010/63/EU on animal protection in scientific endeavors. Fish were euthanized humanely with a 20% benzocaine overdose as prescribed by the EU Directive’s Annex IV. No live fish were subjected to procedures for this research. Tissue usage adhered to the directive’s principles: refine, reduce, and replace animal testing.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw data from this study is available at NCBI under the BioProject code PRJNA1044012.
Acknowledgments
We extend our gratitude to our colleagues in the Chilean Salmon Industry for permitting the collection of samples for this study. We deeply appreciate the technician and veterinary teams at the Centro de Investigaciones Biológicas Aplicadas for their invaluable support. Furthermore, we give our thanks to the Agencia Nacional de Investigación y Desarrollo de Chile (ANID) for funding Y.C.R.’s doctoral studies.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
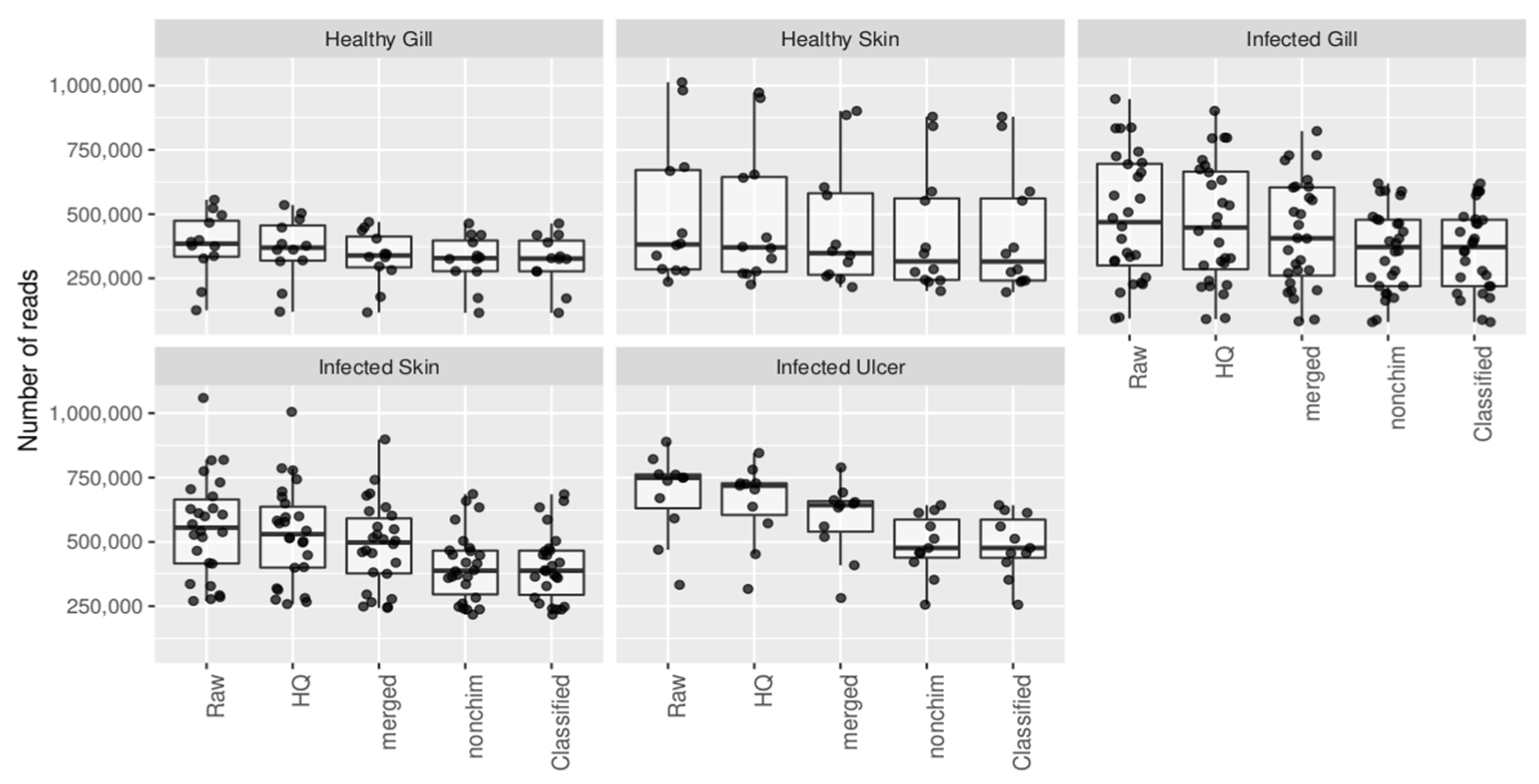
Figure A1.
Numbers of reads for tissue analyzed from healthy Atlantic salmon (healthy gill, healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Figure A1.
Numbers of reads for tissue analyzed from healthy Atlantic salmon (healthy gill, healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Appendix B
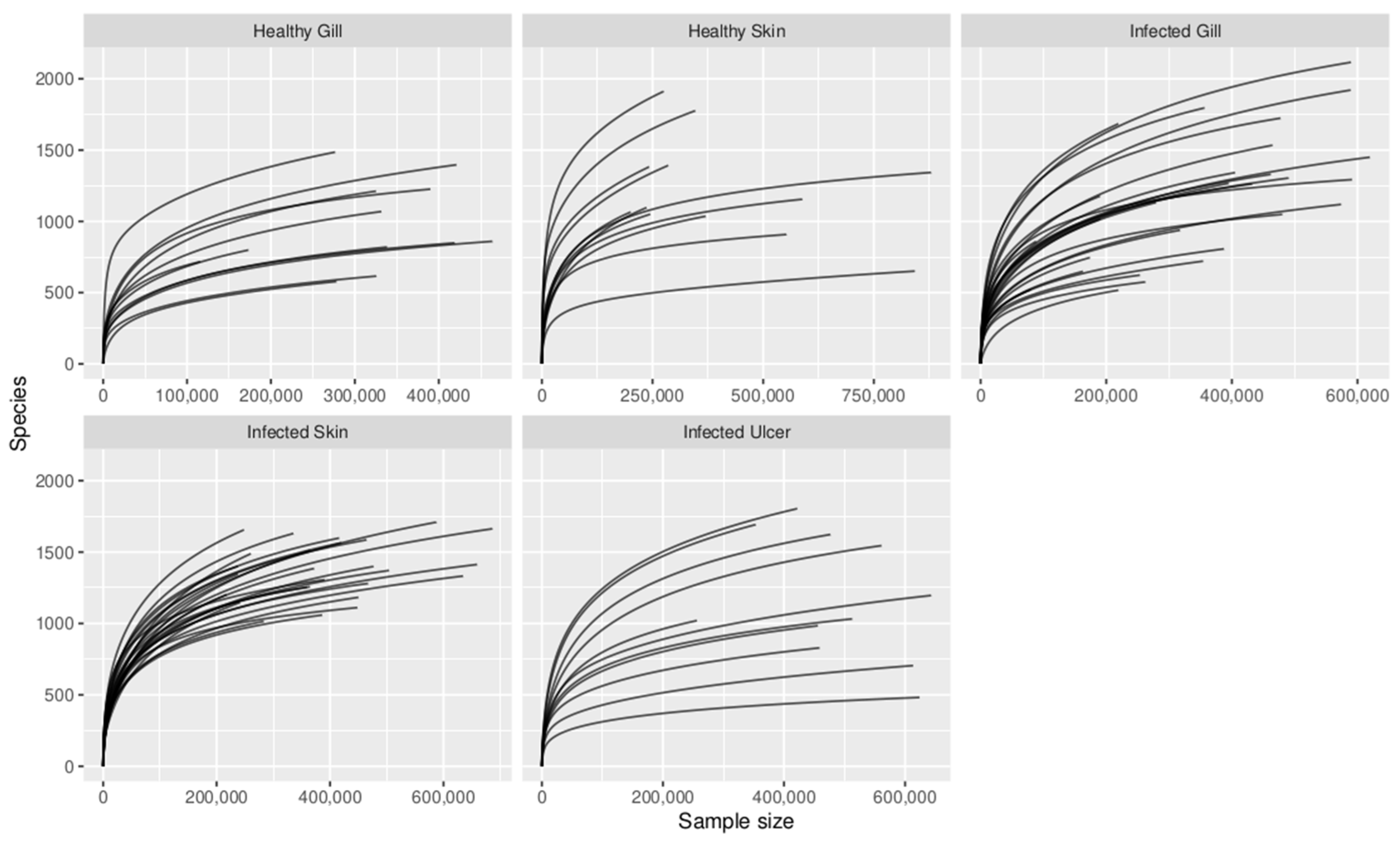
Figure A2.
Rarefaction curves for analyzed tissues of healthy Atlantic salmon (healthy gill, healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Figure A2.
Rarefaction curves for analyzed tissues of healthy Atlantic salmon (healthy gill, healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Appendix C
Figure A3.
Relative abundance (log10) of bacterial families in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer): red (healthy gill, corresponding to the control group), green-yellow (healthy skin, corresponding to the control group), green (infected gill, corresponding to the P. salmonis-infected group), blue-green (infected skin, corresponding to the P. salmonis-infected group), and pink (infected ulcer, corresponding to the P. salmonis-infected group).
Figure A3.
Relative abundance (log10) of bacterial families in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer): red (healthy gill, corresponding to the control group), green-yellow (healthy skin, corresponding to the control group), green (infected gill, corresponding to the P. salmonis-infected group), blue-green (infected skin, corresponding to the P. salmonis-infected group), and pink (infected ulcer, corresponding to the P. salmonis-infected group).
Appendix D
Figure A4.
Co-occurrence network between the principal genera present in the Atlantic salmon infected gill versus the healthy gill of the Atlantic salmon control group.
Figure A4.
Co-occurrence network between the principal genera present in the Atlantic salmon infected gill versus the healthy gill of the Atlantic salmon control group.
Appendix E
Figure A5.
Co-occurrence network between the principal taxa present in the Atlantic salmon infected skin versus the healthy skin of the Atlantic salmon control group.
Figure A5.
Co-occurrence network between the principal taxa present in the Atlantic salmon infected skin versus the healthy skin of the Atlantic salmon control group.
References
- Garcés, L.H.; Larenas, J.J.; Smith, P.A.; Sandino, S.; Lannan, C.N.; Fryer, J.L. Infectivity of a rickettsia isolated from coho salmon Oncorhynchus kisutch. Dis. Aquat. Organ. 1991, 11, 93–97. [Google Scholar] [CrossRef]
- Chen, S.C.; Wang, P.C.; Tung, M.C.; Thompson, K.D.; Adams, A. A Piscirickettsia salmonis-like organism in grouper, Epinephelus melanostigma, in Taiwan. J. Fish Dis. 2000, 23, 415–418. [Google Scholar] [CrossRef]
- Arkush, K.D.; McBride, A.M.; Mendonca, H.L.; Okihiro, M.S.; Andree, K.B.; Marshall, S.; Henriquez, V.; Hedrick, R.P. Genetic characterization and experimental pathogenesis of Piscirickettsia salmonis isolated from white sea bass Atractoscion nobilis. Dis. Aquat. Org. 2005, 63, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Zrnčić, S.; Vendramin, N.; Boutrup, T.S.; Boye, M.; Madsen, L.; Nonneman, B.; Brnić, D.; Oraić, D. First description and diagnostics of disease caused by Piscirickettsia salmonis in farmed European sea bass (Dicentrarchus labrax Linnaeus) from Croatia. J. Fish Dis. 2021, 44, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Cvitanich, J.; Gárate, O.; Smith, C.E. Etiological agent in a Chilean coho disease isolated and confirmed by Koch’s postulates. Am. Fish Soc. Newsl. 1990, 18, 1–2. [Google Scholar]
- Cvitanich, J.; Gárate, O.; Smith, C.E. The isolation of a rickettsia-like organism causing disease and mortality in Chilean salmonids and its confirmation by Koch‘s postulate. J. Fish Dis. 1991, 14, 121–145. [Google Scholar] [CrossRef]
- Cvitanich, J.; Gárate, O.; Smith, C.E. Isolation of a new rickettsia-like organism from Atlantic salmon in Chile. Am. Fish Soc. Newsl. 1995, 23, 1–2. [Google Scholar]
- Karlsen, C.; Ottem, K.; Brevik, Ø.J.; Davey, M.; Sørum, H.; Winther-Larsen, H. The environmental and host-associated bacterial microbiota of Arctic seawater-farmed Atlantic salmon with ulcerative disorders. J. Fish. Dis. 2017, 40, 1645–1663. [Google Scholar] [CrossRef]
- Takle, H. Wounds and Skin Welfare in Atlantic Salmon and Rainbow Trout; Nofima: Tromsø, Norway, 2015. [Google Scholar]
- Salinas, I. The mucosal immune system of teleost fish. Biology 2015, 4, 525–539. [Google Scholar] [CrossRef]
- Salinas, I.; Fernández-Montero, A.; Ding, Y.; Sunyer, J.O. Mucosal immunoglobulins of teleost fish: A decade of advances. Dev. Comp. Immunol. 2021, 121, 104079. [Google Scholar] [CrossRef]
- Kelly, C.; Salinas, I. Under pressure: Interactions between commensal microbiota and the teleost immune system. Front. Immunol. 2017, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Hess, S.; Wenger, A.S.; Ainsworth, T.D.; Rummer, J.L. Exposure of clownfish larvae to suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and microbiome. Sci. Rep. 2015, 5, 10561. [Google Scholar] [CrossRef]
- Gomez, J.A.; Primm, T.P. A Slimy Business: The Future of Fish Skin Microbiome Studies. Microb. Ecol. 2021, 82, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Koppang, E.O.; Kvellestad, A.; Fischer, U. 5—Fish mucosal immunity: Gill. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 93–133. [Google Scholar]
- Streit, B. Bioaccumulation of contaminants in fish. In Fish Ecotoxicology; Braunbeck, T., Hinton, D.E., Streit, B., Eds.; Birkhauser Basel: Basel, Switzerland, 1998; pp. 353–387. [Google Scholar]
- Poleksic, V.; Mitrovic-Tutundzic, V. Fish Gills as a Monitor of Sublethal and Chronic Effects of Pollution. In Fishing News Books Oxford; Fishing News Books Ltd.: Farnham, UK, 1994. [Google Scholar]
- Legrand, T.P.; Catalano, S.R.; Wos-Oxley, M.L.; Stephens, F.; Landos, M.; Bansemer, M.S. The inner workings of the outer surface: Skin and gill microbiota as indicators of changing gut health in yellowtail kingfish. Front. Microbiol. 2018, 8, 26–64. [Google Scholar] [CrossRef] [PubMed]
- Pratte, Z.A.; Besson, M.; Hollman, R.D.; Stewart, F.J. The gills of reef fish support a distinct microbiome influenced by host-specific factors. Appl. Environ. Microbiol. 2018, 84, e00063-18. [Google Scholar] [CrossRef]
- Minich, J.J.; Poore, G.D.; Jantawongsri, K.; Johnston, C.; Bowie, K.; Bowman, J. Microbial ecology of Atlantic salmon (Salmo salar) hatcheries: Impacts of the built environment on fish mucosal microbiota. Appl. Environ. Microbiol. 2020, 86, e00411-20. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I. Microbiome composition and function in aquatic vertebrates: Small organisms making big impacts on aquatic animal health. Front. Microbiol. 2021, 12, 358. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.L.R.M.; Nishanth, T.; Renju, V.B. Bioconcentration of zinc and its effect on the biochemical constituents of the gill tissues of Labeo rohita: An ft-ir study. Infrared Phys. Technol. 2010, 53, 103–111. [Google Scholar] [CrossRef]
- Wu, S.; Wang, G.; Angert, E.R.; Wang, W.; Li, W.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, 30440. [Google Scholar] [CrossRef]
- Schommer, N.N.; Gallo, R.L. Structure and function of the human skin microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef]
- Dorrestein, P.C.; Gallo, R.L.; Knight, R. Microbial skin inhabitants: Friends forever. Cell 2016, 165, 771–772. [Google Scholar] [CrossRef]
- Gallo, R.L. Human skin is the largest epithelial surface for interaction with microbes. J. Investig. Dermatol. 2017, 137, 1213–1214. [Google Scholar] [CrossRef] [PubMed]
- Lorgen-Ritchie, M.; Clarkson, M.; Chalmers, L.; Taylor, J.F.; Migaud, H.; Martin, S.A.M. Temporal changes in skin and gill microbiomes of Atlantic salmon in a recirculating aquaculture system—Why do they matter? Aquaculture 2022, 558, 738352. [Google Scholar] [CrossRef]
- Brown, R.M.; Wiens, G.D.; Salinas, I. Analysis of the gut and gill microbiome of resistant and susceptible lines of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2019, 86, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.; Moore, L.; Mani, A.; Patel, S.; Salinas, I. Effects of ploidy and salmonid alphavirus infection on the skin and gill microbiome of Atlantic Salmon (Salmo Salar). PLoS ONE 2021, 16, e0243684. [Google Scholar] [CrossRef]
- Casadei, E.; Mani, A.; Cisco, M.; Vågnes, Ø.; Salinas, I.; Patel, S. Sex-dependent effects of mechanical delousing on the skin microbiome of broodstock Atlantic salmon (Salmo salar L.). Sci. Rep. 2023, 13, 10824. [Google Scholar] [CrossRef]
- Valenzuela-Miranda, D.; Gonçalves, A.T.; Valenzuela-Muñoz, V.; Nuñez-Acuña, G.; Liachko, I.; Nelson, B.; Gallardo-Escarate, C. Proximity ligation strategy for the genomic reconstruction of microbial communities associated with the ectoparasite Caligus rogercresseyi. Sci. Rep. 2022, 12, 783. [Google Scholar] [CrossRef] [PubMed]
- Koch, R. Xth International Congress of Medicine, Berlin. 1890.
- Brown, S.P.; Cornforth, D.M.; Mideo, N. Evolution of virulence in opportunistic pathogens: Generalism, plasticity, and control. Trends Microbiol. 2012, 20, 336–342. [Google Scholar] [CrossRef]
- Lhorente, J.P.; Gallardo, J.A.; Villanueva, B.; Carabaño, M.J.; Neira, R. Disease resistance in Atlantic salmon (Salmo salar): Coinfection of the intracellular bacterial pathogen Piscirickettsia salmonis and the sea louse Caligus rogercresseyi. PLoS ONE 2014, 9, e95397. [Google Scholar] [CrossRef]
- Arriagada, G.; Hamilton-West, C.; Nekouei, O.; Foerster, C.; Müller, A.; Lara, M.; Gallardo-Escárate, C. Caligus rogercresseyi infestation is associated with Piscirickettsia salmonis-attributed mortalities in farmed salmonids in Chile. Prev. Vet. Med. 2019, 171, 104771. [Google Scholar] [CrossRef]
- Bustos P, Figueroa C, Cádiz B, Santander T, Dixon B, Gallardo JA, Conejeros P. Immune response induced by coinfection of the sea louse Caligus rogercresseyi and the intracellular bacteria Piscirickettsia salmonis in vaccinated Atlantic salmon. J Fish Dis. 2023 Dec;46(12):1337-1342. [CrossRef]
- Rozas-Serri, M.; Peña, A.; Gardner, I.; Peñaloza, E.; Maldonado, L.; Muñoz, A.; Mardones, F.O.; Rodríguez, C.; Ildefonso, R.; Senn, C.; et al. Co-Infection by LF-89-Like and EM-90-Like Genogroups of Piscirickettsia Salmonis in Farmed Atlantic Salmon in Chile: Implications for Surveillance and Control of Piscirickettsiosis. Pathogens 2023, 12, 450. [Google Scholar] [CrossRef] [PubMed]
- Karatas, S.; Mikalsen, J.; Steinum, T.M.; Taksdal, T.; Bordevik, M.; Colquhoun, D.J. Real time PCR detection of Piscirickettsia salmonis from formalin-fixed paraffin-embedded tissues. J. Fish Dis. 2008, 31, 747–753. [Google Scholar] [CrossRef]
- Prophet, E.B.; Mills, B.; Arrington, J.B.; Sobin, L.H. Laboratory methods in histotechnology. Armed Forces Inst. Pathol. 1992, 56, 151–164. [Google Scholar]
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2010; ISBN 978-0-8138-0697-6. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- LEfSe. (Genome Biology, 2011 Jun 24;12(6):R60). Available online: http://huttenhower.sph.harvard.edu/lefse/.
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic species co-occurrence analysis in R. J. Stat. Softw. 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Kuntal, B.K.; Chandrakar, P.; Sadhu, S.; Mande, S.S. ‘NetShift’: A methodology for understanding ‘driver microbes’ from healthy and disease microbiome datasets. ISME J. 2019, 13, 442–454. [Google Scholar] [CrossRef]
- Friedman, J.; Alm, E.J. Inferring Correlation Networks from Genomic Survey Data. PLoS Comput. Biol. 2012, 8, e1002687. [Google Scholar] [CrossRef] [PubMed]
- Kashinskaya, E.N.; Simonov, E.P.; Andree, K.B.; Vlasenko, P.G.; Polenogova, O.V.; Kiriukhin, B.A.; Solovyev, M.M. Microbial community structure in a host-parasite system: The case of Prussian carp and its parasitic crustaceans. J. Appl. Microbiol. 2021, 131, 1722–1741. [Google Scholar] [CrossRef]
- Morales-Rivera, M.F.; Valenzuela-Miranda, D.; Valenzuela-Muñoz, V.; Nuñez-Acuña, G.; Avendaño-Herrera, R.; Gallardo-Escárate, C. Nanopore sequencing evidenced the presence of fish bacterial pathogens in the sea louse (Caligus rogercresseyi) microbiota collected from distant salmon farms in Chile. Aquaculture 2022, 552, 738026. [Google Scholar] [CrossRef]
- Lunder, T.; Evensen, Ø.; Holstad, G.; Hastein, T. Winter ulcer in the Atlantic salmon Salmo Salar—Pathological and bacteriological investigations and transmission experiments. Dis. Aquat. Org. 1995, 23, 39–49. [Google Scholar] [CrossRef]
- Whitman, K.; Backman, S.; Benediktsdottir, E.; Coles, M.; Johnson, G.R. Isolation and characterization of a new Vibrio spp. (Vibrio wodanis) associated with winter ulcer disease in sea water raised Atlantic salmon (Salmo salar L.) in New Brunswick. Aquac. Can. 2001, 2000, 115. [Google Scholar]
- Karlsen, C.; Vanberg, C.; Mikkelsen, H.; Sørum, H. Co-infection of Atlantic salmon (Salmo salar), by Moritella viscosa and Aliivibrio wodanis, development of disease and host colonization. Vet. Microbiol. 2014, 171, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Benediktsdóttir, E.; Helgason, S.; Sigurjónsdóttir, H. Vibrio spp. isolated from salmonids with shallow skin lesions and reared at low temperatures. J. Fish Dis. 1998, 21, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.W.; Griffiths, J.; Petrie, J.; Hastings, T.S. Vibrio viscosus in farmed Atlantic salmon Salmo salar in Scotland: Field and experimental observations. Dis. Aquat. Org. 1998, 34, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, C.; Sørum, H.; Willassen, N.P.; Asbakk, K. Moritella viscosa bypasses Atlantic salmon epidermal keratocyte clearing activity and might use skin surfaces as a port of infection. Vet. Microbiol. 2012, 154, 353–362. [Google Scholar] [CrossRef]
- Hjerde, E.; Karlsen, C.; Sørum, H.; Parkhill, J.; Willassen, N.P.; Thomson, N.R. Co-cultivation and transcriptome sequencing of two co-existing fish pathogens Moritella viscosa and Aliivibrio wodanis. BMC Genom. 2015, 10, 16–447. [Google Scholar] [CrossRef]
- Maharajan, A.D.; Hansen, H.; Khider, M.; Willassen, N.P. Quorum sensing in Aliivibrio wodanis 06/09/139 and its role in controlling various phenotypic traits. PeerJ 2021, 9, e11980. [Google Scholar] [CrossRef]
- Olsen, A.B.; Gulla, S.; Steinum, T.; Colquhoun, D.J.; Nilsen, H.K.; Duchaud, E. Multilocus sequence analysis reveals extensive genetic variety within Tenacibaculum spp. associated with ulcers in sea-farmed fish in Norway. Vet. Microbiol. 2017, 205, 39–45. [Google Scholar] [CrossRef]
- Wynne, J.W.; Thakur, K.K.; Slinger, J.; Samsing, F.; Milligan, B.; Powell, J.F.; Siah, A. Microbiome profiling reveals a microbial dysbiosis during a natural outbreak of tenacibaculosis (Yellow mouth) in Atlantic salmon. Front. Microb. 2020, 11, 586387. [Google Scholar] [CrossRef] [PubMed]
- Slinger, J.; Adams, M.B.; Wynne, J.W. Bacteriomic profiling of branchial lesions induced by Neoparamoeba perurans challenge reveals commensal dysbiosis and an association with Tenacibaculum dicentrarchi in AGD-affected Atlantic salmon (Salmo salar L.). Microorganisms 2020, 8, 1189. [Google Scholar] [CrossRef]
- Apablaza, P.; Frisch, K.; Brevik, Ø.J.; Småge, S.B.; Vallestad, C.; Duesund, H.; Mendoza, J.; Nylund, A. Primary Isolation and Characterization of Tenacibaculum maritimum from Chilean Atlantic Salmon Mortalities Associated with a Pseudochattonella spp. Algal Bloom. J. Aquat. Anim. Health 2017, 29, 143–149. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Toranzo, A.; Magariños, B. Tenacibaculosis infection in marine fish caused by Tenacibaculum maritimum: A review. Dis. Aquat. Org. 2006, 71, 255–266. [Google Scholar] [CrossRef]
- Småge, S.B.; Brevik, Ø.J.; Duesund, H.; Ottem, K.F.; Watanabe, K.; Nylund, A. Tenacibaculum finnmarkense sp. nov., a fish pathogenic bacterium of the family Flavobacteriaceae isolated from Atlantic salmon. Antonie Van Leeuwenhoek 2016, 109, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Benediktsdóttir, E.; Verdonck, L.; Sproer, C.; Helgason, S.; Swings, J. Characterization of Vibrio viscosus and Vibrio wodanis isolated at different geographical locations: A proposal for reclassification of Vibrio viscosus as Moritella viscosa comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Lunder, T.; Sørum, H.; Holstad, G.; Steigerwalt, A.G.; Mowinc-kel, P.; Brenner, D.J. Phenotypic and genotypic characterization of Vibrio viscosus sp. nov. and Vibrio wodanis sp. nov. isolated from Atlantic salmon (Salmo salar) with ‘winter ulcer’. Int. J. Syst. Evol. Microbiol. 2000, 50, 427–450. [Google Scholar] [CrossRef]
- Hovda, M.B.; Fontanillas, R.; McGurk, C.; Obach, A.; Rosnes, J.T. Seasonal variations in the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.). Aquac. Res. 2012, 43, 154–159. [Google Scholar] [CrossRef]
- Hamdan, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2013, 30, 759–795. [Google Scholar] [CrossRef]
- Rudi, K.; Angell, I.L.; Pope, P.B.; Vik, J.O.; Sandve, S.R.; Snipen, L.G. Stable core gut microbiota across the freshwater-to-saltwater transition for farmed Atlantic salmon. Appl. Environ. Microbiol. 2018, 84, e01974-17. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; McGinnity, P.; Dionne, M.; Letourneau, J.; Thonier, F.; Carvalho, G.R.; Derome, N. The biogeography of the Atlantic salmon (Salmo salar) gut microbiome. Multidiscip. J. Microb. Ecol. 2015, 10, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens. Disease of Farmed and Wild Fish; Springer International Publishing: Cham, Switzerland, 2016; pp. 475–498. [Google Scholar] [CrossRef]
- Attia, M.M.; Abdelsalam, M.; Elgendy, M.Y.; Sherif, A.H. Dactylogyrus extensus and Pseudomonas fluorescens dual infection in farmed common carp (Cyprinus carpio). Microb. Pathog. 2022, 173, 105867. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.T.; Kim, J.H.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.G.; Kim, S.W.; Han, S.J.; Kwon, J.; et al. Genetic characterization and pathological analysis of a novel bacterial pathogen, Pseudomonas tructae, in rainbow trout (Oncorhynchus mykiss). Microorganisms 2019, 7, 432. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Liu, Y.; Chen, C.; Lin, Q.; Ren, L.; Huang, M.; Tu, J.; Luo, T. Pseudomonas ovata sp. nov., isolated from the skin of the tail of Farmed Murray cod (Maccullochella peelii peelii) with a profound ulceration. Curr. Microbiol. 2019, 76, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival, and persistence. Front. Cell Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Levipan, H.A.; Irgang, R.; Tapia-Cammas, D.; Avendaño-Herrera, R. A high-throughput analysis of biofilm formation by the fish pathogen Tenacibaculum dicentrarchi. J. Fish Dis. 2019, 42, 617–621. [Google Scholar] [CrossRef]
- Cornelis, P.; Bodilis, J. A survey of TonB-dependent receptors in fluorescent pseudomonads. Environ. Microbiol. Rep. 2009, 1, 256–262. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Toranzo, A.E.; Romalde, J.L.; Lemos, M.L.; Magariños, B. Iron uptake mechanisms in the fish pathogen Tenacibaculum maritimum. Appl. Environ. Microbiol. 2005, 71, 6947–6953. [Google Scholar] [CrossRef]
- Calquín, P.; Ruiz, P.; Oliver, C.; Sánchez, P.; Haro, R.; Oliva, H. Physiological evidence that Piscirickettsia salmonis produces siderophores and uses iron from different sources. J. Fish Dis. 2017, 41, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A.; Pizarro, P.; Ojeda, P.; Contreras, J.; Oyanedel, S.; Larenas, J. Routes of entry of Piscirickettsia salmonis in rainbow trout Oncorhynchus mykiss. Dis. Aquat. Org. 1999, 37, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A.; Rojas, M.E.; Guajardo, A.; Contreras, J.; Morales, M.A.; Larenas, J. Experimental infection of coho salmon Oncorhynchus kisutch by exposure of skin, gills, and intestine with Piscirickettsia salmonis. Dis. Aquat. Org. 2004, 61, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, J.F.; Richards, R.H.; Robertson, D.A. Gross, histological and scanning electron microscopic appearance of dorsal fin rot in farmed Atlantic salmon, Salmo salar L., parr. J. Fish Dis. 1996, 19, 415–427. [Google Scholar] [CrossRef]
- Yañez, J.M.; Lhorente, J.P.; Bassini, L.N.; Oyarzún, M.; Neira, R.; Newman, S. Genetic co-variation between resistance against both Caligus rogercresseyi and Piscirickettsia salmonis, and body weight in Atlantic salmon (Salmo salar). Aquaculture 2014, 433, 295–298. [Google Scholar] [CrossRef]
- González, L.; Robles, C.; San Martín, M.C. Management issues regarding caligidosis treatment on salmon farms in Chile affected by infection salmon anaemia virus (ISAV), Piscirickettsia salmonis and Neoparamoeba perurans. Ocean Coast. Manag. 2016, 123, 74–83. [Google Scholar] [CrossRef]
Figure 1.
Atlantic salmon (S. salar) affected by clinical manifestations of salmonid rickettsial septicemia (SRS), characterized primarily by cutaneous signs (a–c). Affected fish exhibit raised scales, scaling, cutaneous hemorrhages, and the presence of multiple ulcers, some coalescent, varying in size, with distinct white-bordered edges.
Figure 1.
Atlantic salmon (S. salar) affected by clinical manifestations of salmonid rickettsial septicemia (SRS), characterized primarily by cutaneous signs (a–c). Affected fish exhibit raised scales, scaling, cutaneous hemorrhages, and the presence of multiple ulcers, some coalescent, varying in size, with distinct white-bordered edges.

Figure 2.
Atlantic salmon (S. salar), affected by a cutaneous clinical presentation of salmonid rickettsial septicemia (SRS). Immunohistochemistry was performed on paraffin-embedded tissue using a monoclonal antibody specific to P. salmonis. A positive reaction is evident as brown staining (a-b) of coccoid structures (a-b), with invasion of the dermis, hypodermis, and adjacent musculature.
Figure 2.
Atlantic salmon (S. salar), affected by a cutaneous clinical presentation of salmonid rickettsial septicemia (SRS). Immunohistochemistry was performed on paraffin-embedded tissue using a monoclonal antibody specific to P. salmonis. A positive reaction is evident as brown staining (a-b) of coccoid structures (a-b), with invasion of the dermis, hypodermis, and adjacent musculature.
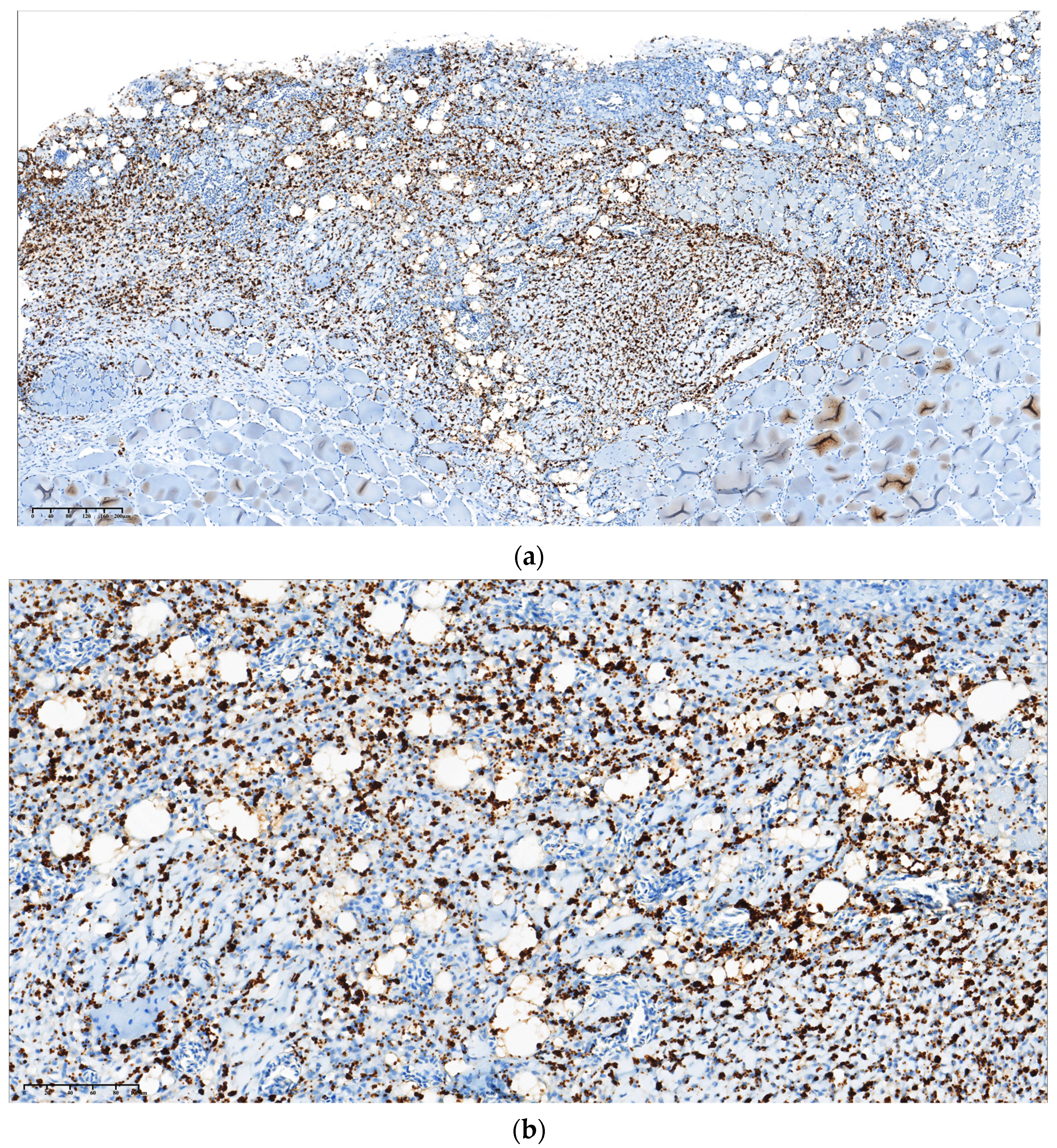
Figure 3.
Non-metric Multidimensional Scaling (NMDS) ordination plots illustrating the distribution of amplicon sequence variants (ASVs) across the sampled salmon population, which was divided into two distinct groups: the “healthy” Atlantic salmon (control group) and P. salmonis-infected Atlantic salmon. The NMDS analysis provides a visual representation of the compositional differences between the microbiota of these two groups.
Figure 3.
Non-metric Multidimensional Scaling (NMDS) ordination plots illustrating the distribution of amplicon sequence variants (ASVs) across the sampled salmon population, which was divided into two distinct groups: the “healthy” Atlantic salmon (control group) and P. salmonis-infected Atlantic salmon. The NMDS analysis provides a visual representation of the compositional differences between the microbiota of these two groups.
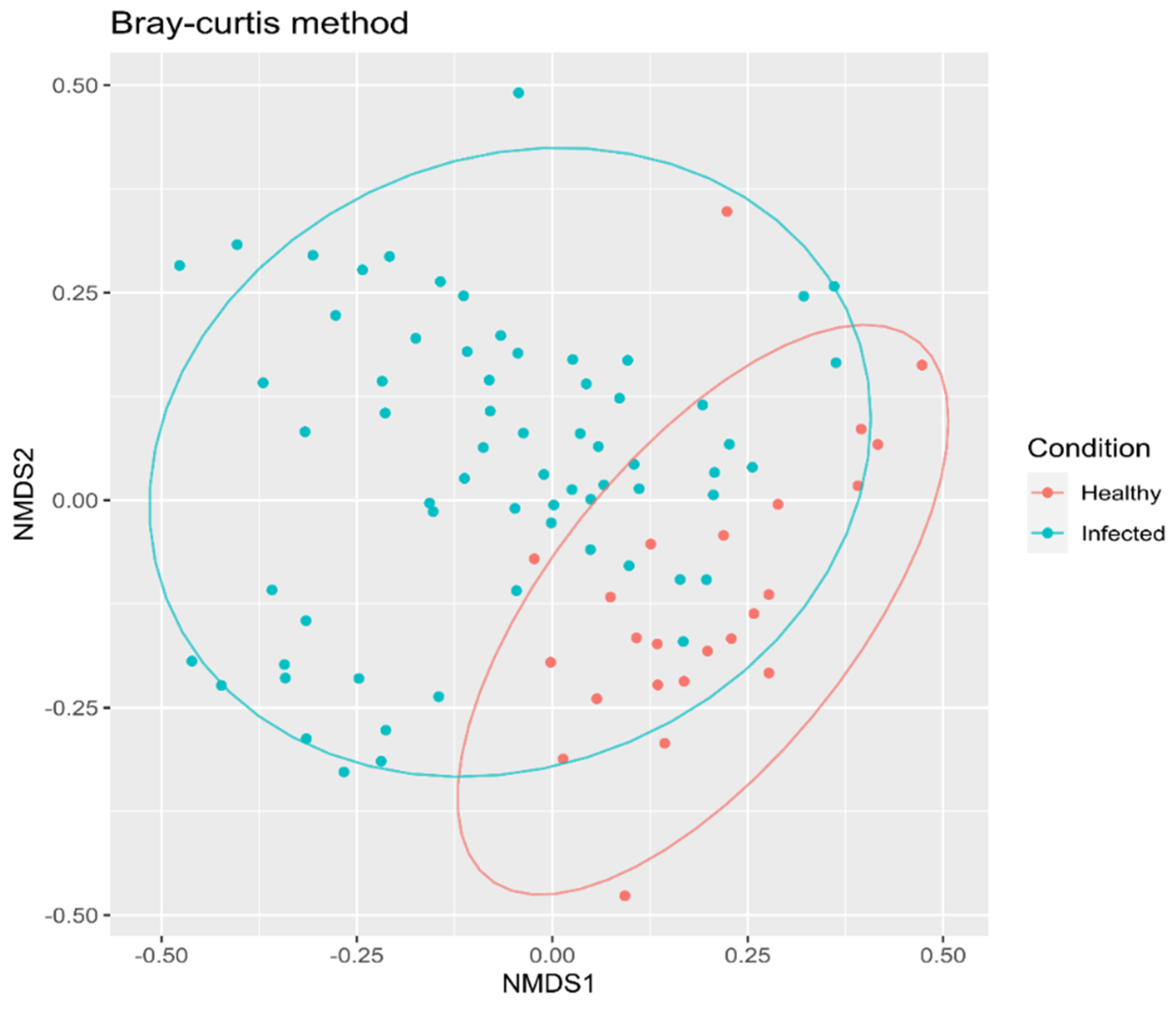
Figure 4.
(a) Non-metric Multidimensional Scaling (NMDS) ordination plots representing the distribution of amplicon sequence variants (ASVs) within tissues of both “healthy” (control group) and P. salmonis-infected Atlantic salmon. Color-coded labels are used to differentiate the tissue types: red (healthy gill, control group), green-yellow (healthy skin, control group), green (infected gill, P. salmonis-infected group), blue-green (infected skin, P. salmonis-infected group), and pink (infected ulcer, P. salmonis-infected group). (b) Schematic representation of the ulcerative progression in Atlantic salmon (Created with BioRender.com).
Figure 4.
(a) Non-metric Multidimensional Scaling (NMDS) ordination plots representing the distribution of amplicon sequence variants (ASVs) within tissues of both “healthy” (control group) and P. salmonis-infected Atlantic salmon. Color-coded labels are used to differentiate the tissue types: red (healthy gill, control group), green-yellow (healthy skin, control group), green (infected gill, P. salmonis-infected group), blue-green (infected skin, P. salmonis-infected group), and pink (infected ulcer, P. salmonis-infected group). (b) Schematic representation of the ulcerative progression in Atlantic salmon (Created with BioRender.com).

Figure 5.
Microbial alpha diversity metrics by tissue (species richness and Shannon diversity), comparing the tissues of healthy Atlantic salmon (control group) and P. salmonis-infected Atlantic salmon: red (healthy gill, corresponding to the control group), green-yellow (healthy skin, corresponding to the control group), green (infected gill, corresponding to the P. salmonis-infected group), blue-green (infected skin, corresponding to the P. salmonis-infected group), and pink (infected ulcer, corresponding to the P. salmonis-infected group).
Figure 5.
Microbial alpha diversity metrics by tissue (species richness and Shannon diversity), comparing the tissues of healthy Atlantic salmon (control group) and P. salmonis-infected Atlantic salmon: red (healthy gill, corresponding to the control group), green-yellow (healthy skin, corresponding to the control group), green (infected gill, corresponding to the P. salmonis-infected group), blue-green (infected skin, corresponding to the P. salmonis-infected group), and pink (infected ulcer, corresponding to the P. salmonis-infected group).
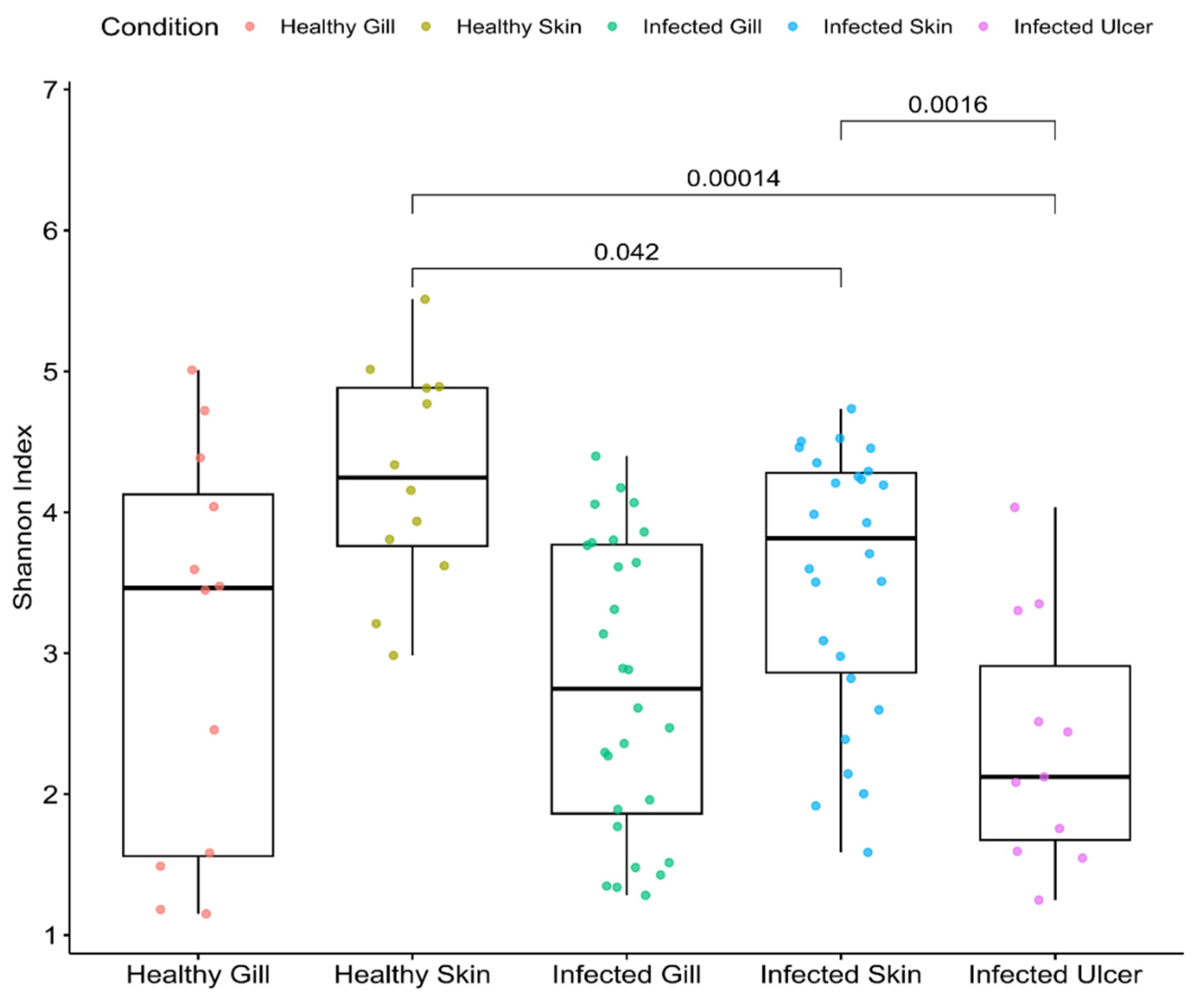
Figure 6.
Mean relative abundances of amplicon sequence variants (ASVs) visualized at the bacterial phylum, class, and order level in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Figure 6.
Mean relative abundances of amplicon sequence variants (ASVs) visualized at the bacterial phylum, class, and order level in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
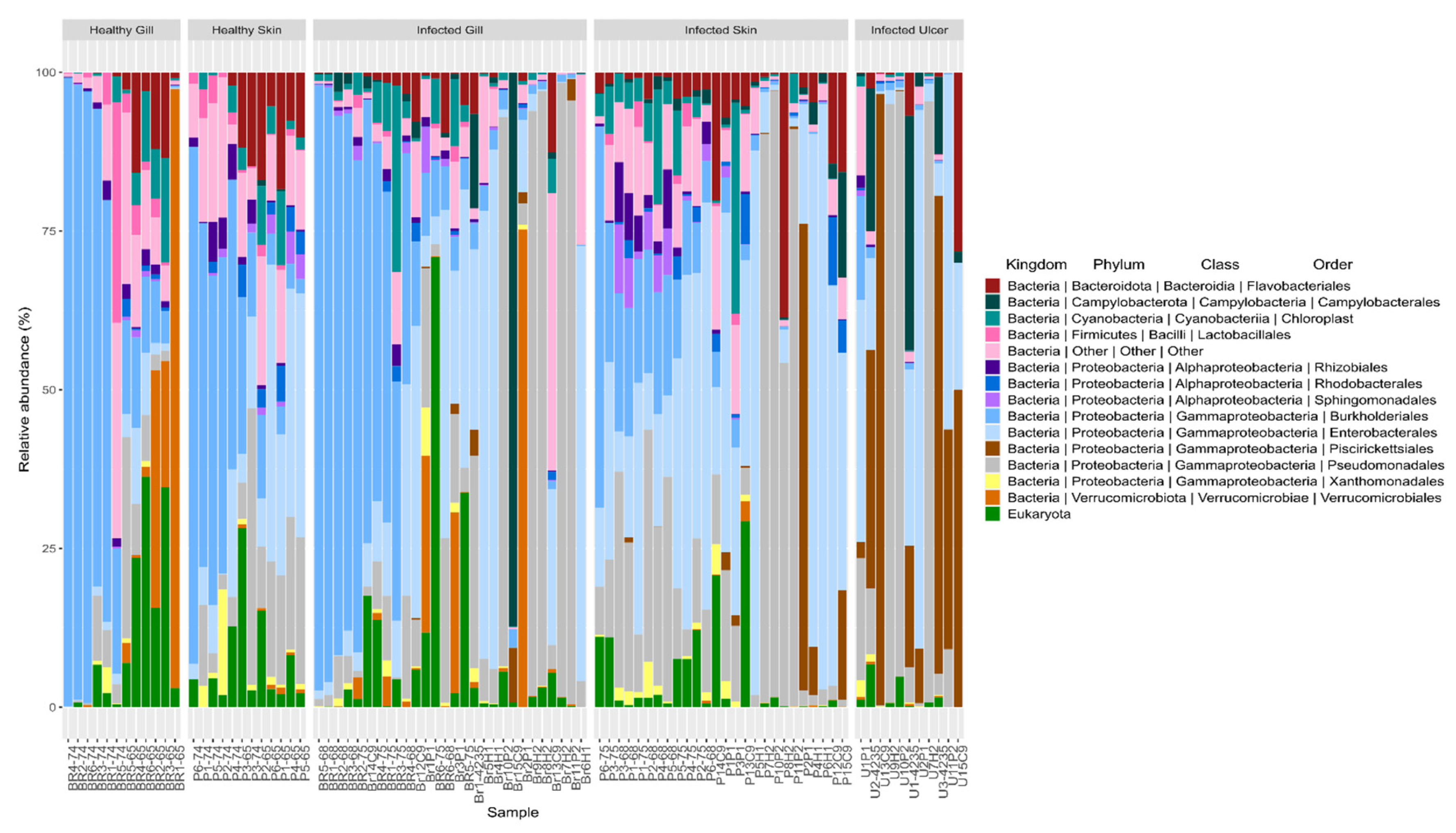
Figure 7.
Mean relative abundances of amplicon sequence variants (ASVs) of P. salmonis in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
Figure 7.
Mean relative abundances of amplicon sequence variants (ASVs) of P. salmonis in the tissues of the healthy Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer).
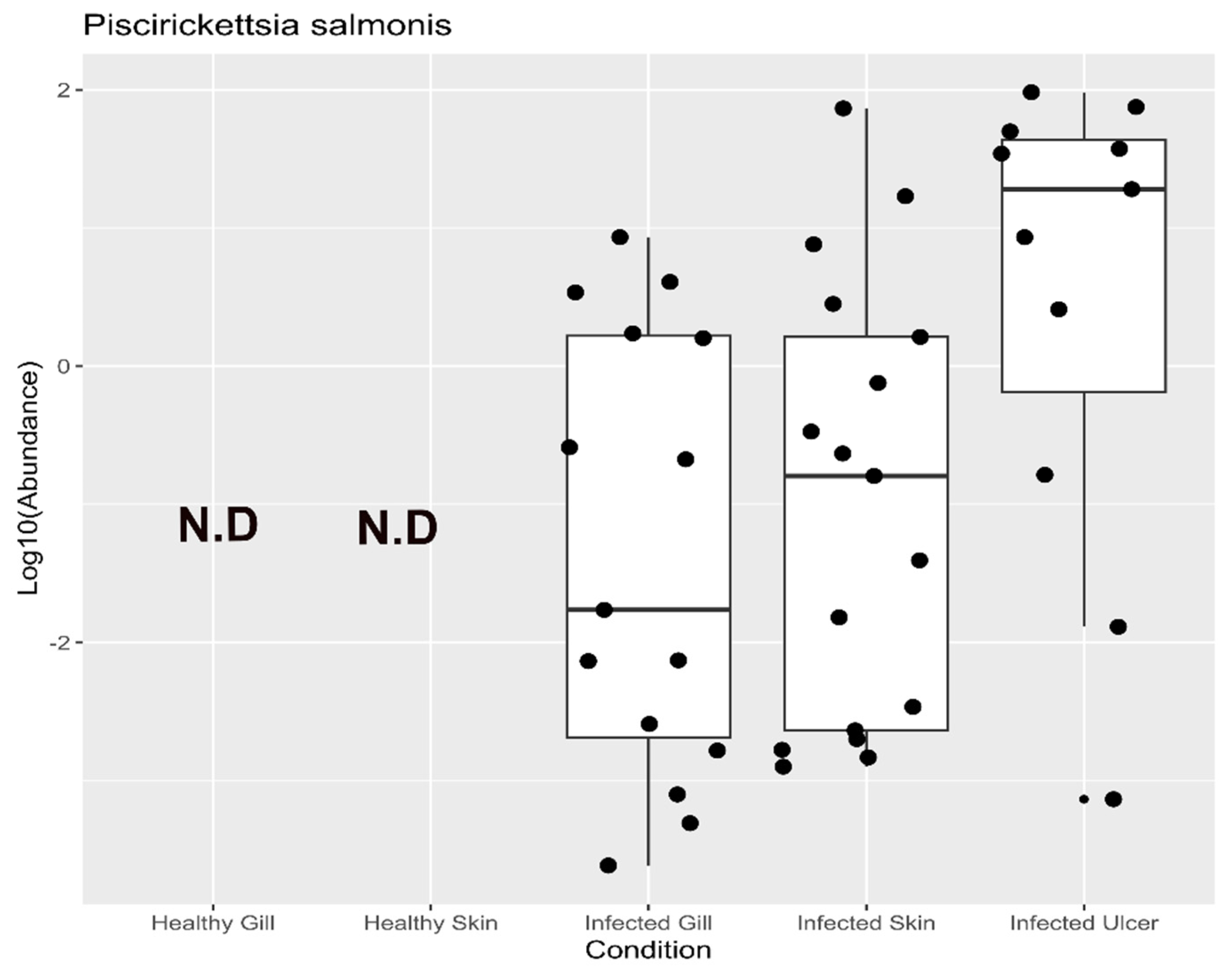
Figure 8.
Linear discriminant analysis effect size (LEfSe) and linear discriminant analysis (LDA) characterization of microbiomes across tissues in the “healthy” Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer). Tissue categories are color-coded: pink (infected ulcer, P. salmonis-infected group), blue-green (infected skin, P. salmonis-infected group), green (infected gill, P. salmonis-infected group), green-yellow (healthy skin, control group), and red (healthy gill, control group).
Figure 8.
Linear discriminant analysis effect size (LEfSe) and linear discriminant analysis (LDA) characterization of microbiomes across tissues in the “healthy” Atlantic salmon group (healthy gill and healthy skin) and P. salmonis-infected Atlantic salmon (infected gill, infected skin, and infected ulcer). Tissue categories are color-coded: pink (infected ulcer, P. salmonis-infected group), blue-green (infected skin, P. salmonis-infected group), green (infected gill, P. salmonis-infected group), green-yellow (healthy skin, control group), and red (healthy gill, control group).
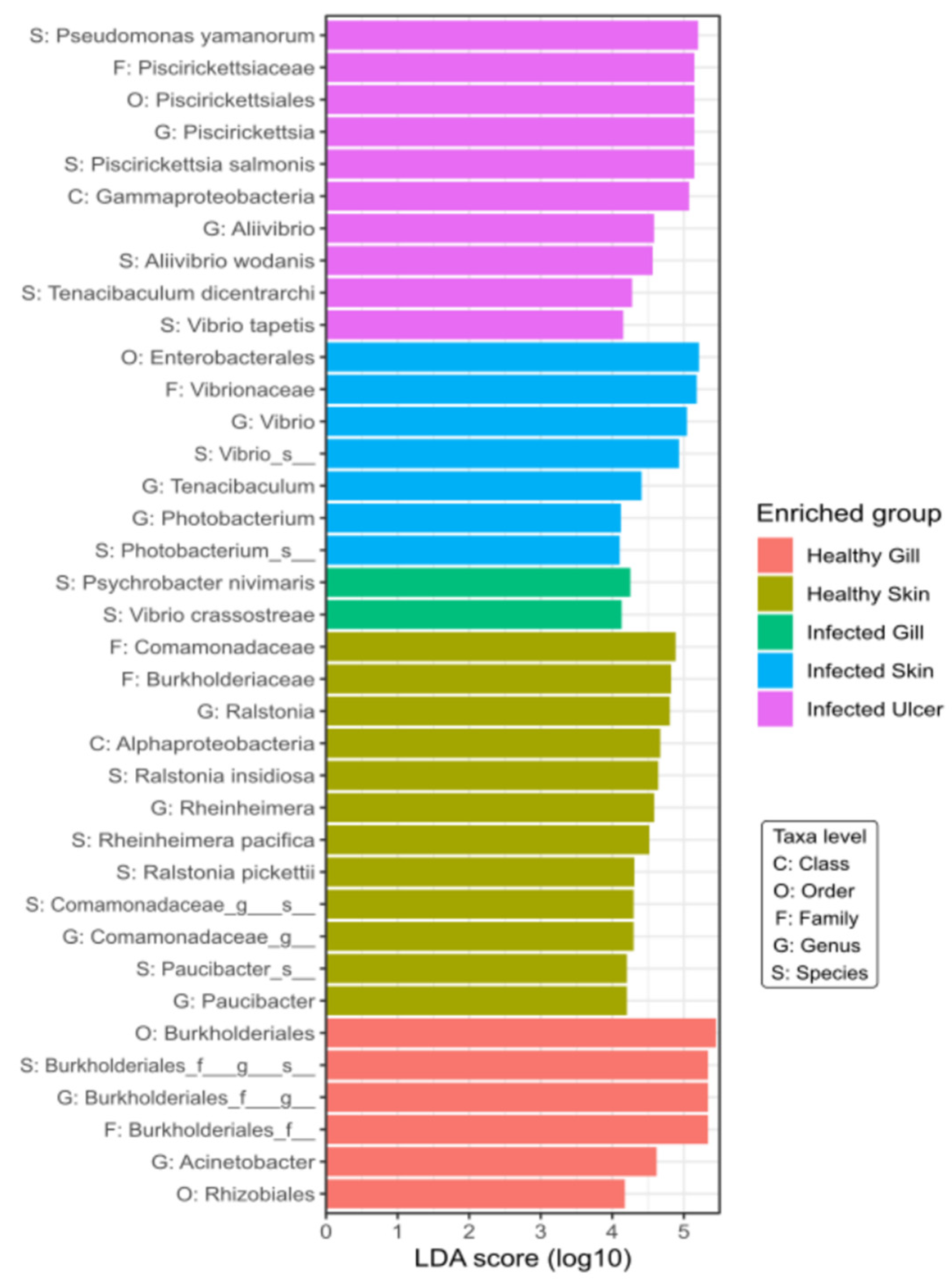
Figure 9.
Co-occurrence network depicting the relationships among the principal genera within the Atlantic salmon ulcer tissue compared to the healthy skin of the Atlantic salmon control group.
Figure 9.
Co-occurrence network depicting the relationships among the principal genera within the Atlantic salmon ulcer tissue compared to the healthy skin of the Atlantic salmon control group.


Table 1.
Atlantic salmon (Salmo salar) farms sampled in Los Lagos and Aysén Regions (Chile) during infectious salmonid rickettsial septicemia (SRS) outbreaks.
Table 1.
Atlantic salmon (Salmo salar) farms sampled in Los Lagos and Aysén Regions (Chile) during infectious salmonid rickettsial septicemia (SRS) outbreaks.
| Farmed Sampled | Health Condition | Sampled Type | Region |
| C1 | Infected with P. salmonis | Skin, gill, and ulcer | Los Lagos |
| C2 | Infected with P. salmonis | Skin, gill, and ulcer | Los Lagos |
| C3 | Infected with P. salmonis | Skin, gill, and ulcer | Aysén |
| C4 | Infected with P. salmonis | Skin, gill, and ulcer | Los Lagos |
| C5 | Infected with P. salmonis | Skin, gill, and ulcer | Los Lagos |
| C6 | Infected with P. salmonis | Skin, gill, and ulcer | Los Lagos |
| C7 | Infected with P. salmonis | Skin, gill, and ulcer | Aysén |
| C8 | Control Farm | Skin and gill | Aysén |
| C9 | Control Farm | Skin and gill | Los Lagos |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



