
Submitted:
24 November 2023
Posted:
25 November 2023
You are already at the latest version
Abstract
Flavin-dependent monooxygenases (FMOs) have made substantial influence as catalysts for monooxygenation reactions, impacting diverse field such as drug metabolism, environmental studies, and natural product synthesis. Their application in biocatalysis boasts several advantages over conventional chemical catalysis, such as heightened selectivity, safety, sustainability, and eco-friendliness. In the realm of biomedicine, FMOs are pivotal in antibiotic research, modulating natural products, antimicrobial agents, and pathways involved in drug synthesis. They're also underscored as potential pharmaceutical targets, pivotal in disease progression and viable for therapeutic intervention. Additionally, FMOs play a substantial role in environmental science, especially in pesticide processing and plant vitality. Their involvement in biosynthesizing compounds like polyethers, tropolones, and ω-hydroxy fatty acids, with remarkable regio- and stereoselectivity, renders them indispensable in drug discovery and development. As our comprehension of FMOs' catalytic mechanisms and structures advances, coupled with cutting-edge biotechnologies like computational design and directed evolution, FMOs are poised to occupy an increasingly significant role in both scientific exploration and industrial applications.
Keywords:
flavin-dependent monooxygenases (FMOs)
; natural products biosynthesis
; biocatalysis
; pharmaceutical development
; heteroatom hydroxylation
; baeyer-villiger oxidation
1. Introduction
Flavin-dependent enzymes are a class of enzymes that play a crucial role in biochemical reactions, catalyzing a variety of chemical reactions through their reliance on flavins as cofactors. Flavins, including flavin adenine dinucleotide (FAD) and flavin mononucleotide (FMN), are water-soluble derivatives of vitamin B2, ubiquitously present in organisms. These cofactors act as catalysts in numerous biochemical processes, especially in redox reactions, and are essential for maintaining normal metabolism in living organisms [1].Among the diverse families of flavin-dependent enzymes, flavin-dependent monooxygenases (FMOs) are particularly significant. These enzymes catalyze monooxygenation reactions, where a single oxygen atom is inserted into an organic substrate, utilizing the unique redox properties of flavin cofactors. Biologists and chemists have begun to exploit these incredible catalysts to achieve their catalytic roles in various fields, aiming to provide potential applications for biocatalysis across diverse industries in the future.
Distinct from traditional chemical catalysis, biocatalysts offer unique advantages in terms of high selectivity, safety, sustainability, and eco-friendliness [2]. For flavin-dependent monooxygenases (FMOs), the mediated redox reactions utilize molecular oxygen as a pure chemical oxidant, with one oxygen atom being incorporated into the substrate and the other forming water. This process circumvents the use of challenging and hazardous oxidants. As biocatalysts, FMOs can operate in aqueous environments without the need for extreme temperatures or pressures, simplifying and enhancing the practicality of biocatalytic processes. Despite these advantages, widespread application of biocatalysis across various fields faces significant challenges, such as limited number of characterized enzymes, difficulties in enzyme acquisition, and narrow substrate scope. However, with advances in molecular and structural biology, research on FMOs is deepening, revealing their complex regulatory mechanisms and interactions with biomolecules [3,4]. The ongoing development of biocatalysis has led to an increase in the number of commercial enzymes, and technologies for enzyme modification and evolution have been established to expand substrate scope and enhance enzyme stability [5].
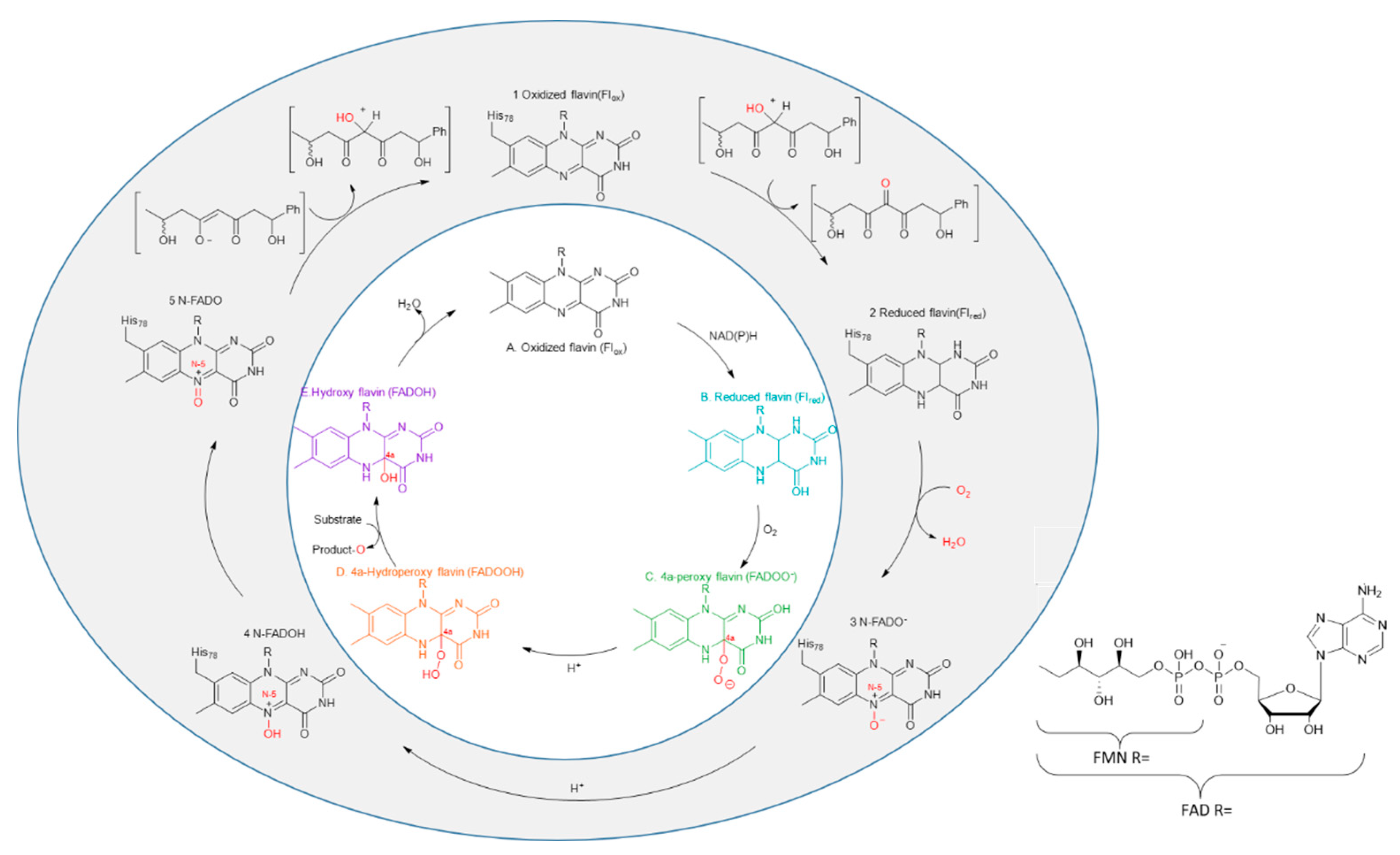
Years of research have elucidated the mechanistic framework of flavin-dependent monooxygenases. As illustrated in Figure 1, flavins undergo a series of oxidative epoxide reactions to transfer oxygen atoms. The catalysis by FMOs is divided into two mechanisms: C4a and N5. The traditional C4a mechanism consists of two half-reactions. In the first half-reaction, the cofactor binds to the enzyme, reducing FAD (or FMN) through two electron transfers. In the second half-reaction, molecular oxygen rapidly reacts with the reduced flavin (Flred), forming a peroxide flavin and oxidizing the substrate [6]. In addition, recently, Robin and colleagues first revealed the N5 redox mechanism in Encm [7]. This mechanism enables the protonation of the N5 hydroxyl group and the hydroxylamine tautomerization in the absence of a cofactor, thereby achieving hydroxylation of the substrate.
Flavin-dependent monooxygenases (FMOs), as key biocatalysts, exhibit tremendous potential in multiple disciplines, including but not limited to drug metabolism [4], degradation of harmful substances [8], and synthesis of bioactive molecules [6]. Importantly, unlike cytochrome P450 enzymes (CYPs), FMOs are self-sufficient, not requiring additional redox partner proteins for activation [9]. Moreover, compared to CYPs, FMOs demonstrate higher catalytic efficiency and lower level of substrate inhibition [10,11]. In recent years, significant progress has been made in the application research of FMOs. This work will discuss the novel applications of flavin-dependent monooxygenases FMOs in various fields such as biomedical science, environmental science, and synthetic biology.
2. Applications of FMOs in the Biomedical Field
2.1. Applications of FMOs in Antibiotic Research
With the advancement of bacterial genomic sequencing and genome mining, the identification of biosynthetic gene clusters for bioactive natural products, such as tumor-inhibiting and antibiotic agents, has accelerated [12]. In the reconstruction of these biological pathways, many new FMOs have been discovered. These FMOs are involved not only in the synthesis of natural products but also in initiating biodegradation metabolism [13,14]. Additionally, this activity enables bacteria to degrade or deactivate antibiotics. MabTetX, an FMO found in Mycobacterium abscessus, belongs to the tetracycline destructase (TetX) family [15]. Recent studies have associated MabTetX with tetracycline resistance [16]. MabTetX catalyzes the modification of tetracycline and doxycycline through oxidation reactions. Studies have also shown that deletion of the MabTetX gene MAD_1496c increases sensitivity to tetracycline and doxycycline. Anhydrotetracycline (ATc), a degradation product of tetracycline, despite its lower antibiotic activity, can effectively inhibit MabTetX, thereby reducing the minimum inhibitory concentrations (MICs) of tetracycline and doxycycline, potentially enhancing their effectiveness against Mycobacterium abscessus [16].
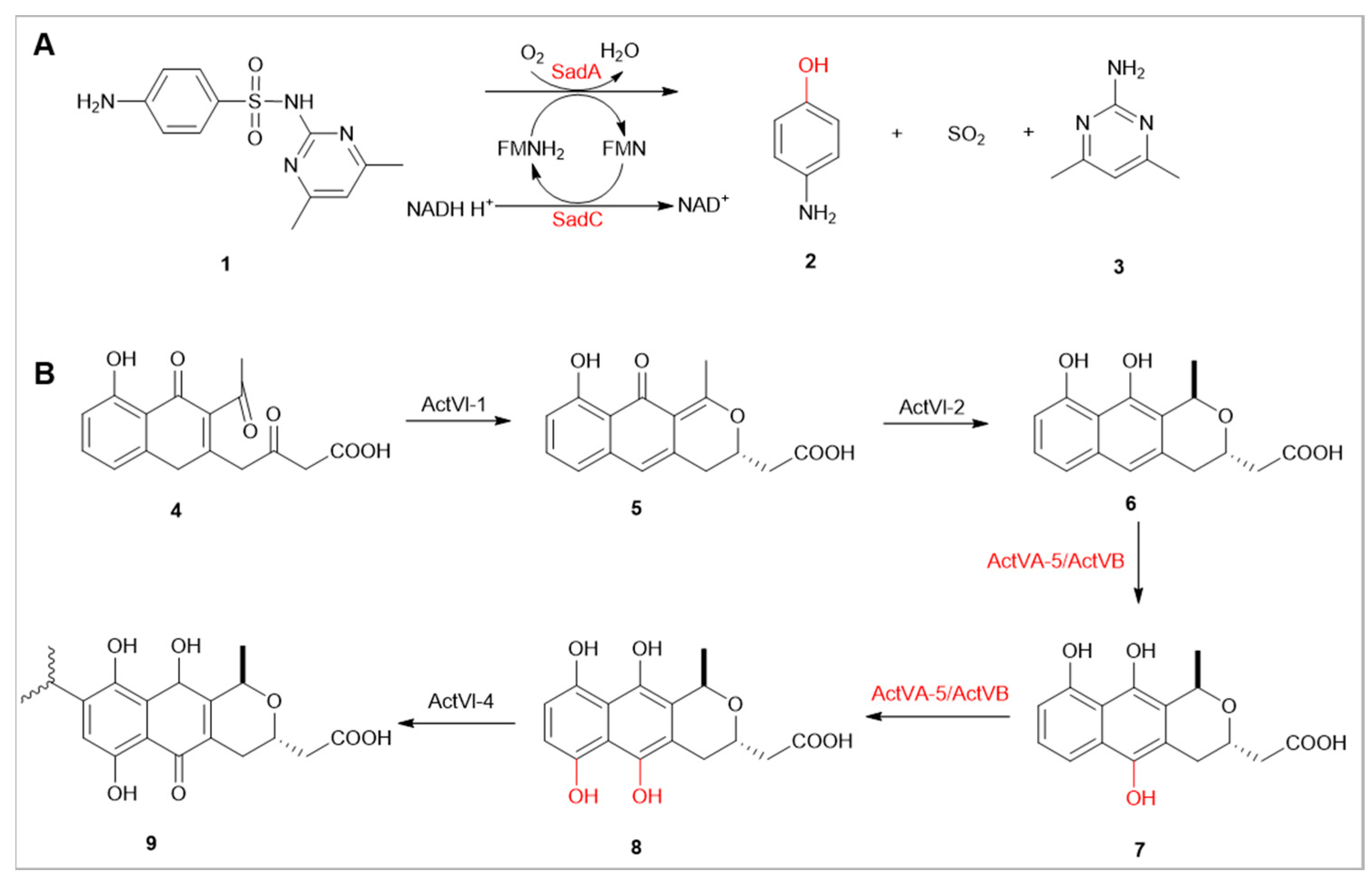
Sulfonamide drugs, synthetic antimicrobial agents, are widely used in human and veterinary medicine [17]. However, sulfonamides are chemically difficult to degrade, and the enzymes involved in their degradation are considered to possess potential resistance mechanisms [18]. In the bacterium Microbacterium sp. CJ77, enzymes SadA and SadC are involved in the degradation of sulfonamides (1) and are associated with resistance. SadA, a FMNH2-dependent monooxygenase, is responsible for the initial ipso-hydroxylation reaction of sulfonamides(2), while SadC, a flavin reductase, participates in the subsequent breakdown of these drugs [18]. Co-expression of this two-component system in Escherichia coli has been shown to decrease sensitivity to sulfamethoxazole.
FMOs are also extensively involved in the degradation of aromatic compounds in the environment and the biosynthesis of secondary metabolites [4]. Recent studies have identified the ActVA-ORF5/ActVB system as being involved in the synthesis of the quinone antibiotic actinorhodin (9). This system specifically participates in two consecutive hydroxylation reactions (7,8), which are crucial steps in the biosynthetic pathway of actinorhodin [19].
Figure 2.
(A) Involvement of SadA/SadC in the Degradation of Sulfonamides. (B) Actinorhodin Biosynthesis Pathway, ActVA-5/ActVB Involved in Consecutive Hydroxylation Twice.
Figure 2.
(A) Involvement of SadA/SadC in the Degradation of Sulfonamides. (B) Actinorhodin Biosynthesis Pathway, ActVA-5/ActVB Involved in Consecutive Hydroxylation Twice.
2.2. Applications of FMOs as Pharmaceutical Targets
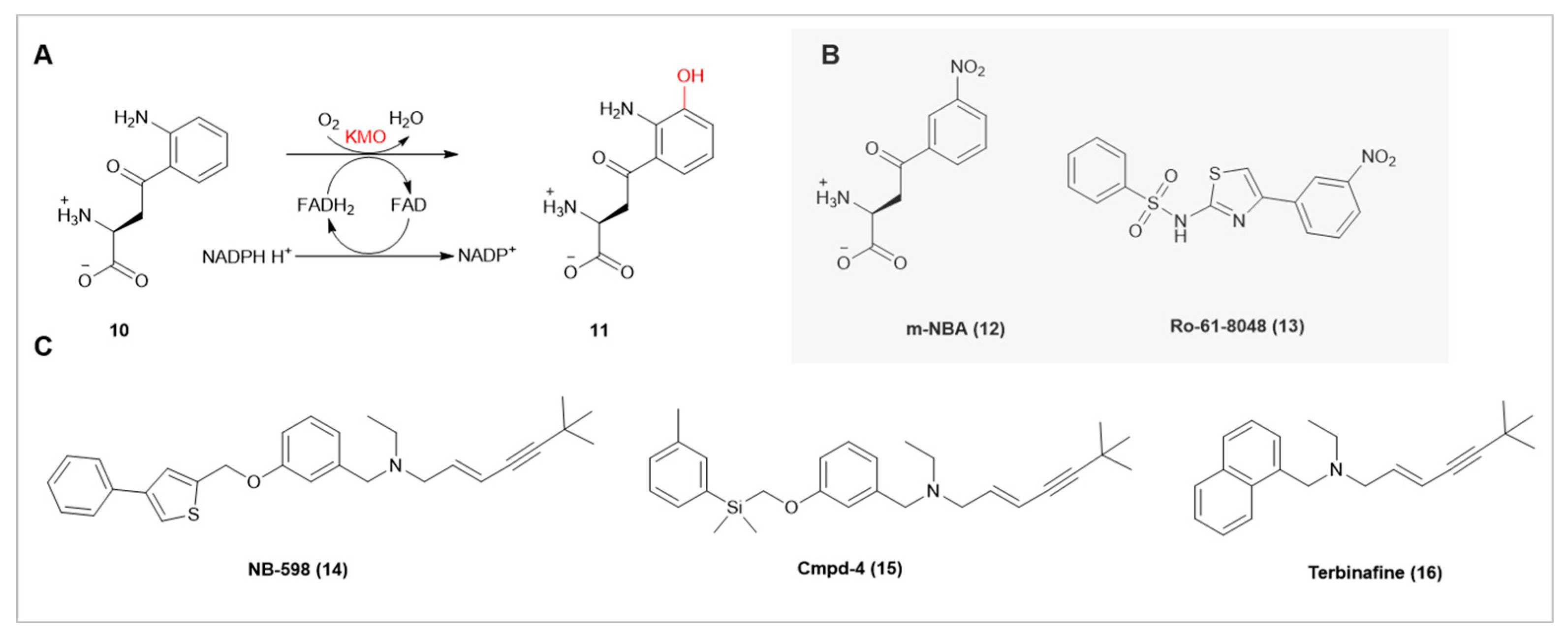
Kynurenine 3-monooxygenase (KMO), a key enzyme in the Class A FMO pathway situated in the mitochondrial outer membrane, is responsible for catalyzing the hydroxylation of L-kynurenine(10), thereby producing the neurotoxin 3-hydroxykynu-renine(11) [20]. Studies indicate that KMO facilitates cancer progression and serves as a prognostic marker for human hepatocellular carcinoma and canine mammary tumors [21]. For instance, in a recent study on triple-negative breast cancer (TNBC), KMO amplification was associated with poorer survival rates. KMO expression is significantly higher in TNBC tumors compared to adjacent normal mammary tissues. In vitro experiments have shown that increased KMO enhances TNBC cell growth, colony and mammosphere formation, as well as migration, invasion, and the expression of mesenchymal markers. KMO also upregulates the expression and promoter activity of pluripotency genes. It regulates pluripotency genes through β-catenin, playing an oncogenic role in the progression of TNBC [22]. Currently, research focuses on developing effective KMO inhibitors(12,13), primarily through high-throughput screening [23] and drug design [24].
Human Squalene Epoxidase (SQLE), a key enzyme in cholesterol biosynthesis, has emerged as a research focus in recent years. NB-598(14) and Cmpd-4(15) have been developed as potent SQLE inhibitors, with IC50 values of 63 nM and 69 nM, respectively. In contrast, the common antifungal drug Terbinafine (16), although sharing a similar tertiary amine structure with these inhibitors, possesses a larger naphthyl group. This structural difference corresponds with its weaker inhibitory effect (IC50 of 7.7 μM) [25]. High-resolution crystallographic analysis has also revealed the binding sites of these inhibitors [26]. The interaction between the Tyr195 residue and the amine group of the inhibitors may be crucial in designing the next generation of SQLE inhibitors.
Figure 3.
(A) KMO Catalyzes the hydroxylation of L-kynurenine. (B) Potential KMO Inhibitors. (C) Three SQLE Inhibitors(NB-598 IC50=63 nM, Cmpd-4 IC50=69 nM, Terbinafine IC50=7.7 μM).
Figure 3.
(A) KMO Catalyzes the hydroxylation of L-kynurenine. (B) Potential KMO Inhibitors. (C) Three SQLE Inhibitors(NB-598 IC50=63 nM, Cmpd-4 IC50=69 nM, Terbinafine IC50=7.7 μM).
2.3. Applications of FMOs in Drug Synthesis
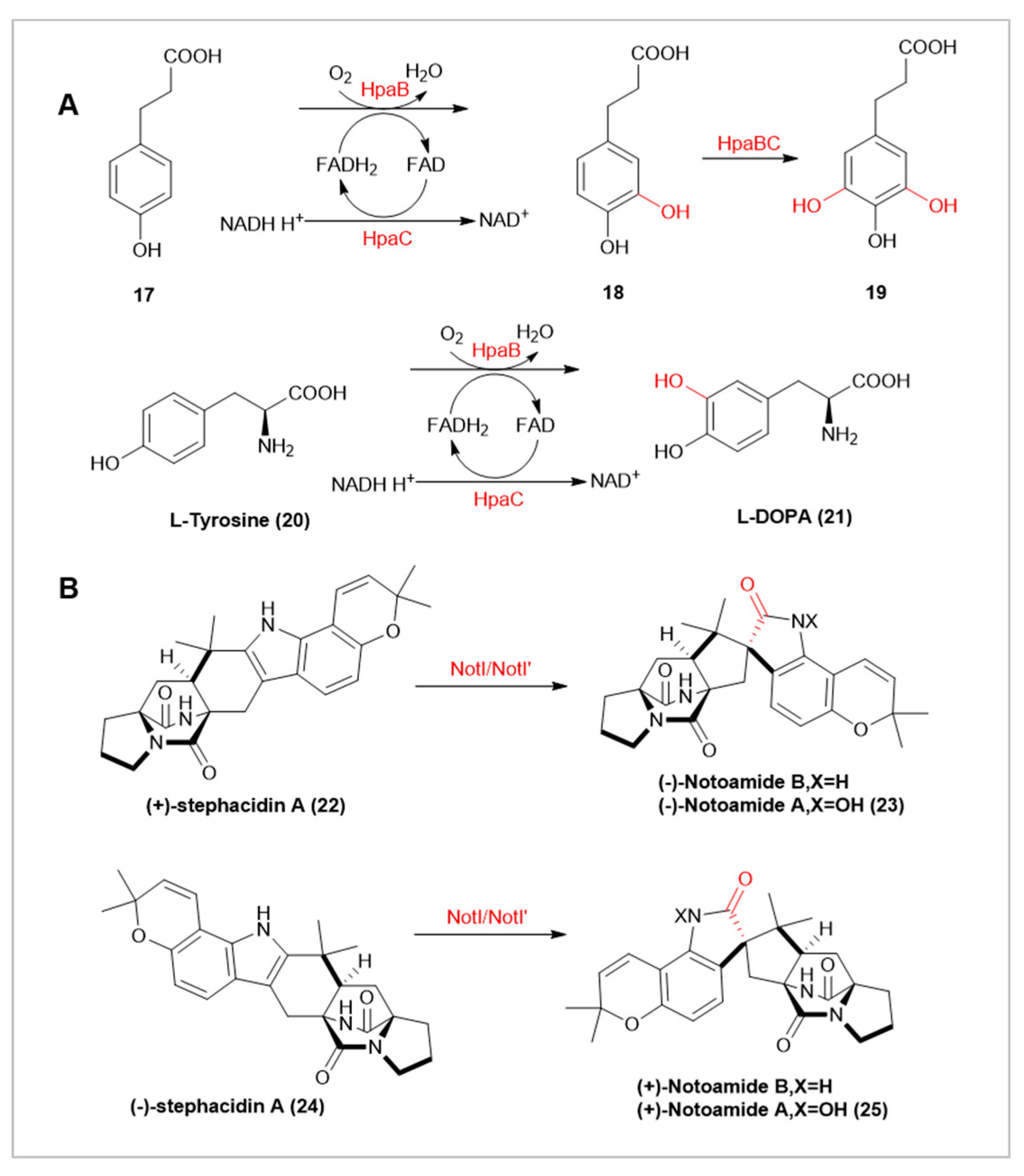
HpaBC, a bifunctional flavin-dependent monooxygenase in Pseudomonas aeruginosa, is involved in catalyzing the hydroxylation of cinnamic acid derivatives [27]. HpaBC demonstrates high activity in the hydroxylation of 3-(4-Hydroxyphenyl)-propanoic acid(17), producing 3-(3,4,5-Trihydroxyphenyl)-propanoic acid(19), a compound with significant anti-proliferative effects on human cancer cells and high medicinal potential [28]. Recent studies have shown that HpaBC can also hydroxylate L-Tyrosine(20) to produce L-DOPA(21), a prescription medication for Parkinson’s disease [29]. Characterization of HpaBC’s function and structure revealed its ability to hydroxylate a range of phenolic substrates, including tyrosol, hydroxyphenyllactic acid, coumaric acid, hydroxybenzoic acid, and phenol [30]. Further enzyme engineering, by substituting selected HpaBC residues with homologous residues from Pseudomonas aeruginosa, revealed that these mutants possess highly region-specific aromatic hydroxylation capabilities [31].
Fungal indole alkaloids are a class of bioactive natural products, with Notoamide exhibiting notable anticancer activity [32]. In the synthesis of these compounds, the flavin-dependent monooxygenases NotI and NotI’ (sharing 85% sequence homology) catalyze the rearrangement of the semi-indole to form the spiro-indole portion. Interestingly, both oxygenases can process any enantiomer of stephacidin A (22,24), leading to the corresponding Notoamide isomers (23,25). Notably, (-)-stephacidin A(24) is more reactive, which might be attributed to the conversion of (+)-stephacidin(22) A being an evolutionary process [32].
Figure 4.
(A) HpaBC-Catalyzed Hydroxylation of 3-(4-Hydroxyphenyl)-propanoic Acid and L-Tyrosine.(B) Stereochemical Selectivity in Notl/Notl’-Catalyzed Hydroxylation.
Figure 4.
(A) HpaBC-Catalyzed Hydroxylation of 3-(4-Hydroxyphenyl)-propanoic Acid and L-Tyrosine.(B) Stereochemical Selectivity in Notl/Notl’-Catalyzed Hydroxylation.
3. Applications of FMOs in the Biomedical Field
3.1. Application of FMOs in the Processing of Pesticides
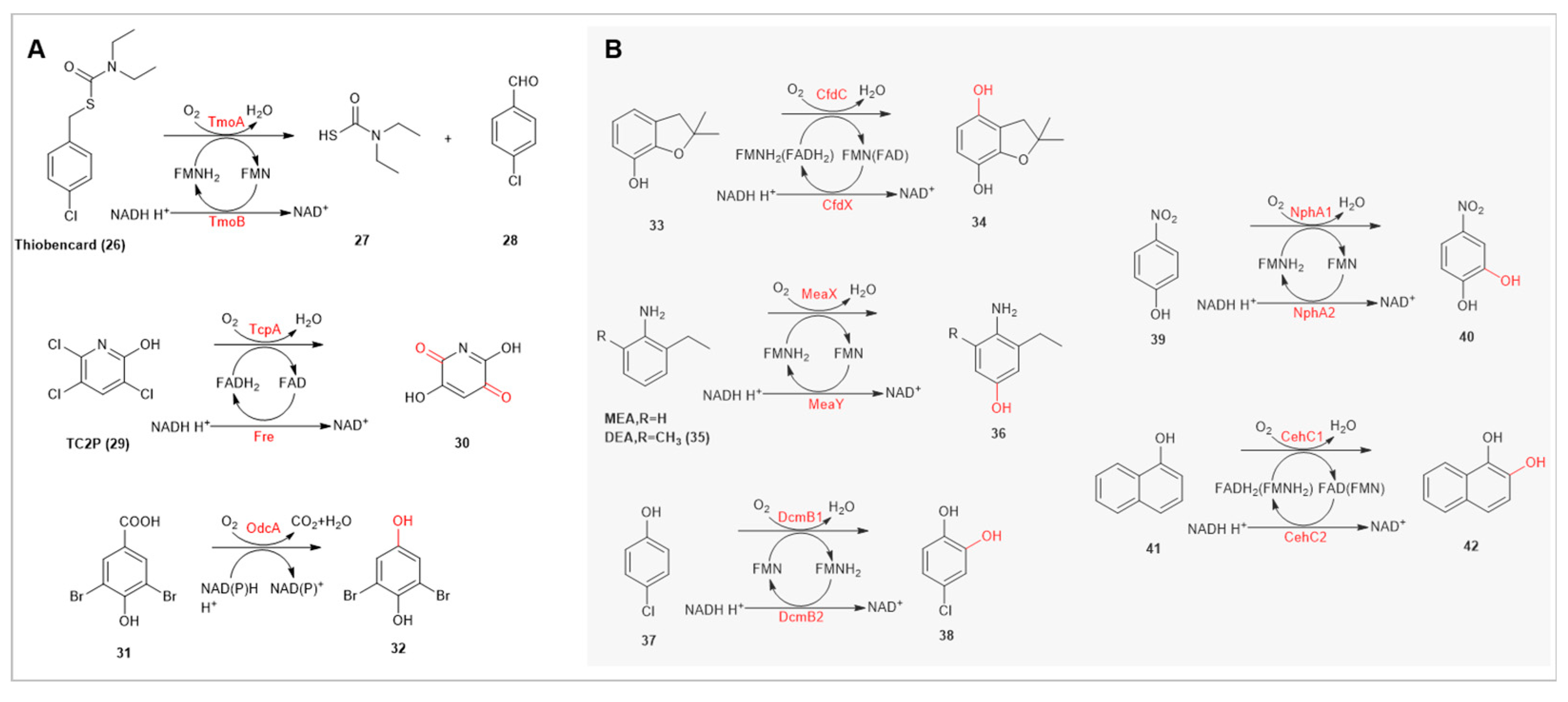
The pesticide degradation process involves several key steps, including the cleavage of functional groups, hydroxylation of aromatic heterocycles, and further breakdown of hydroxylated aromatic heterocycles [33]. In this degradation pathway, the cleavage of functional groups is the initial step, primarily involving the release of heteroatoms, such as dealkylation, decarboxylation, and dehalogenation. For instance, the bifunctional FMO TomAB in Acidovorax sp. is involved in the C-S bond cleavage of the thiocarbamate herbicide thiobencard(26) [34]. Chlorpyrifos, one of the most commonly used organophosphate insecticides, typically degrades in the environment to 3,5,6-trichloro-2-pyridinol (TC2P,29) [35]. TcpA from the X1T strain can progressively dechlorinate TC2P in the presence of the redox partner protein (Fre) [36]. Additionally, the OdcA gene in the Pigmentiphaga sp. strain, identified during the study of the herbicide bromoxynil’s hydrolysis product 3,5-dibromo-4-hydroxybenzoate (DBHB,31), exhibits decarboxylation activity, converting DBHB to 2,6-dibromohydroquinone (32) in the presence of cofactors. However, the activity of OdcA significantly decreases with monobrominated analogs [37].
Aromatic hydroxylation, especially ortho-dihydroxylation, produces dihydroxylated compounds crucial for ring cleavage processes [33]. The bifunctional flavin-dependent monooxygenases CfdCCDS-1 and CdfX from Sphingomonas sp. are capable of hydroxylating furan phenol(33), a hydrolysis product of furan [38]. In Sphingobium baderi DE-13, the bifunctional FMO MeaXY can completely degrade 2-methyl-6-ethyl aniline (MEA,35) and hydroxylate MEA and its metabolic product 2,6-diethyl aniline (EDA,36) to produce para-hydroxylated amino derivatives. MeaXY is also highly conserved in other Sphingomonas strains degrading MEA [39]. Another bifunctional flavin-dependent monooxygenase, DcmB1B2, can ortho-hydroxylate 4-chlorophenol to form 4-chlorocatechol(37) [40]. Similarly, NphA1A2 (56%-79% sequence similarity to DcmB1B2) can convert 4-nitrophenol(39) to 4-nitrocatechol(40) [41]. In Rhizobium sp. X9, the bifunctional FMO CehC1C2 identified in the typical degradation pathway of the carbamate pesticide carbaryl catalyzes the conversion of 1-naphthol(41) to 1,2-dihydroxynaphthalene(42) [42]. CehC1C2 can utilize both FAD and FMN as cofactors, with FAD showing higher catalytic activity(Km and Kcat/Km values of 74.71 ± 16.07 μM and (8.29 ± 2.44) × 10−4 s−1·μM−1).
Figure 5.
(A) Applications of TomAB, TcpA, and OdcA in Functional Group Cleavage. (B) Applications of CfdCX, MeaXY, DcmB1B2, NphA1A2, and CehC1C2 in Aromatic Ring Hydroxylation, with DcmB1B2 and CehC1C2 Catalyzing the Formation of Vicinal Diol Structures.
Figure 5.
(A) Applications of TomAB, TcpA, and OdcA in Functional Group Cleavage. (B) Applications of CfdCX, MeaXY, DcmB1B2, NphA1A2, and CehC1C2 in Aromatic Ring Hydroxylation, with DcmB1B2 and CehC1C2 Catalyzing the Formation of Vicinal Diol Structures.
3.2. Application of FMOs in the Processing of Pesticides
In recent years, studies in mammals, bacteria, and fungi have significantly advanced the research on flavin-dependent monooxygenases (FMOs), particularly in areas closely related to human health and biotechnological applications [43,44,45]. In contrast, although plants possess a greater diversity of FMOs, the endogenous substrates of plant FMOs remain largely unidentified, which considerably hampers the research in this area [46,47].
Plant FMOs are classified as Class B FMOs, characterized by three conserved sequences: the FMO recognition motif (FXGXXXHXXXY/F), the FAD binding motif (GXGXXG), and the NAD(P)H binding motif (GXGXXG) [48]. The FMO recognition motif is located at the binding pocket junction, ensuring proper structural rotation and conformational changes [49]. The primary subclass of plant FMOs is BFMO, but recent studies on BVMOs from mosses have expanded our understanding of the diversity of plant FMOs [50].
Cm-BVMO from Cyanidioschyzon merolae (Cm) and Pp-BVMO from Physcomitrella patens (Pp) are currently the only reported plant-derived BVMOs. Cm-BVMO is known as the most thermostable Type I BVMO, with an apparent melting temperature of 56 °C. Pp-BVMO has been demonstrated to insert oxygen atoms into C-C bonds, catalyzing the conversion of phenylacetone (43) to benzyl acetate (44). Interestingly, mutating the Y in the FxGxxxYxxxWP motif to H in the enzyme sequence resulted in higher activity on most tested substrates. [50] However, the physiological role of Pp-BVMO remains unclear. The recombinant enzyme exhibits relatively low kcat values (in the 0.1–0.2 s−1 range),suggesting a potential role in the secondary metabolism of these photosynthetic organisms.
BFMO is the largest subclass of plant FMOs, typically catalyzing oxidation reactions that lead to the hydroxylation of heteroatoms like nitrogen and sulfur [4,51].YUCCA, the first functionally characterized FMO in plants, is involved in the synthesis of the hormone auxin (IAA,46) [52].In Arabidopsis, a series of YUCCA enzymes (YUC 2-11) have been identified [53,54,55],and other plant species also possess complete YUCCA genes [56,57]. Numerous YUCCAs are involved in catalyzing the oxidative decarboxylation of IPA(45) to IAA(46), as well as the oxidative decarboxylation of phenylpyruvic acid to produce phenylacetic acid [58], and the N-hydroxylation of TAM(47) to produce N-hydroxy TAM(48) [52]. YUC6, the most extensively studied enzyme in this family, is not limited to auxin synthesis but also exhibits reductase partner enzyme activity, enhancing peroxidase activity and the scavenging of reactive oxygen species [59]. Overexpression of YUC6 enhances plant drought tolerance and reduces leaf senescence [60]. Despite significant progress in studying YUCCA functions, the catalytic mechanism of these enzymes remains largely elusive.
The AsFMO1 enzyme from garlic shows high sequence similarity to the S-methyl methanethiosulfinate S-oxygenase (FMOGS-OX) enzyme in Arabidopsis [61,62]. AsFMO1 exhibits high stereoselective S-oxygenation activity towards (+)-alliin [(RCSS)-S-allylcysteine sulfoxide (49), with an apparent Km value of 0.25 mM, resulting in the formation of allicin(50). This compound provides the distinctive flavor of the Allium genus and is beneficial for human health [61].
Plants can induce a broad-spectrum immune response to pathogen infections, known as systemic acquired resistance (SAR). In Arabidopsis, AtFMO1 plays a critical role in inducing and regulating SAR, crucial for disease resistance and plant immunity [63]. As a piperidine N-hydroxylase, AtFMO1 catalyzes the conversion of the lysine catabolite pipecolic acid (Pip,53) into N-hydroxypipecolic acid (NHP,54). The exogenous application of AtFMO1 can overcome the FMO deficiencies caused by a lack of NHP, thereby enhancing immunity against oomycete infections [64].
Figure 6.
The Applications of FMOs in plant life catalytic metabolism.
4. Applications of FMOs in the Natural Product Synthesis
Natural products (NPs) typically do not partake in primary metabolic processes but are encoded and synthesized by secondary metabolic pathways. They possess unique structures and play a crucial role in drug discovery [65,66,67]. By deciphering and reconstructing metabolic pathways in organisms, a variety of naturally occurring compounds with unique bioactivities can be produced in cell factories, such as medicinal terpenes [68,69,70], polyphenols [71,72,73], and alkaloids [72,74,75]. The catalytic reactions of FMOs in synthesizing natural products include hydroxylation, epoxidation, Baeyer-Villiger oxidation, decarboxylation, dehalogenation, and dealkylation [51,76].
4.1. Application of FMOs in the Biosynthesis of Polyether through epoxidations
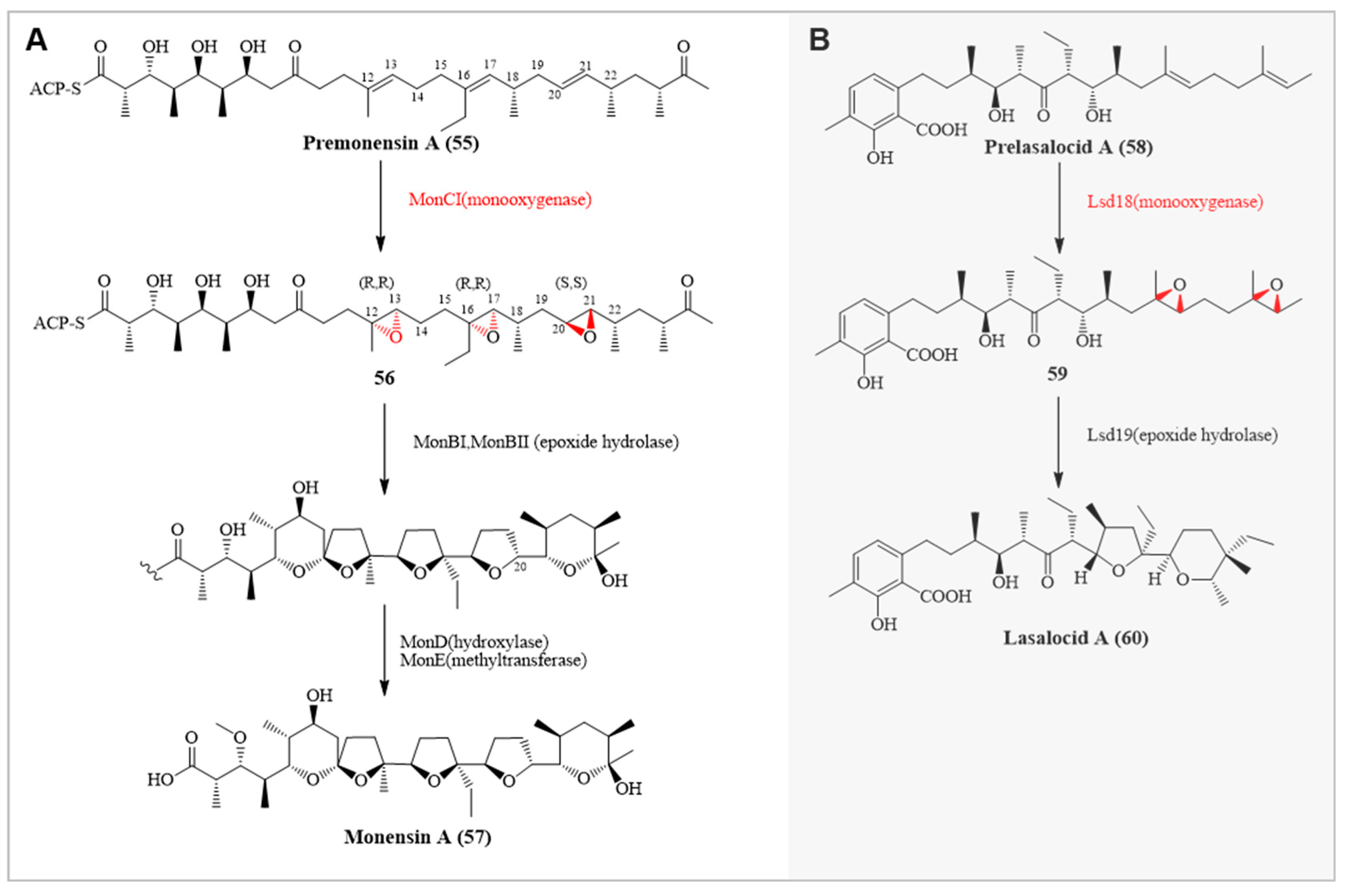
Natural polycyclic polyethers are primarily composed of acetate, propionate, and butyrate units, typically containing multiple chiral centers [77]. Their unique multi-furan ring structure facilitates the transport of metal ions. In polyether carriers, metal ions often chelate with oxygen atoms, enhancing the permeability of ions across biological membranes due to their hydrophobic properties, thus exhibiting significant antibiotic activity [78]. For instance, Monensin A(57), a typical ion-carrier polycyclic polyether antibiotic, disrupts the ion concentration gradient across cell membranes by chelating ions like Na+ and K+, widely used in the treatment of coccidiosis in poultry and cattle [79]. In the biosynthetic pathway of Monensin A, flavin-dependent epoxidase MonCI and epoxide hydrolases MonBI/BII were discovered. MonCI is involved in the stereoselective epoxidation of three double bonds in the precursor substrate [3,80] while MonBI/BII participate in epoxide ring-opening cascade reactions to form new five- and six-membered rings. MonCI catalyzes the triple epoxidation of premonensin A (55) to form (12R, 13R, 16R, 17R, 20S, 21S)-triiepoxypremonenin(56), with each epoxidation occurring in a highly stereospecific manner. This process is facilitated by the unusually large substrate binding cavity of MonCI, accommodating various conformations of premonensin A. This capability of performing multiple stereospecific epoxidations demonstrates the extraordinary functionality and vast potential of FMOs [3].
Lasalocid A(60) is one of the simplest polycyclic polyether ion carriers, similar to Monensin, composed of a tetrahydrofuran ring and a tetrahydropyran ring. A key enzyme in the synthesis of lasalocid A is Lsd18, a flavin-dependent epoxidase [81]. Lsd18 catalyzes the conversion of prelasalocid A(58) to Bisepoxyprelasalocid A(59), an essential step in the formation of the polycyclic polyether compound. Subsequently, lsd19 is involved in the epoxide hydrolysis reaction to form the final polycyclic polyether compound. The entire process is carried out in a stereoselective manner.
Figure 7.
(A) Biosynthesis of Mononemycin A involving MonCI. (B) Biosynthesis of Lasalocid A involving Lsd18.
Figure 7.
(A) Biosynthesis of Mononemycin A involving MonCI. (B) Biosynthesis of Lasalocid A involving Lsd18.
4.2. Application of FMOs in the Biosynthesis of Natural Products through Dearomatization
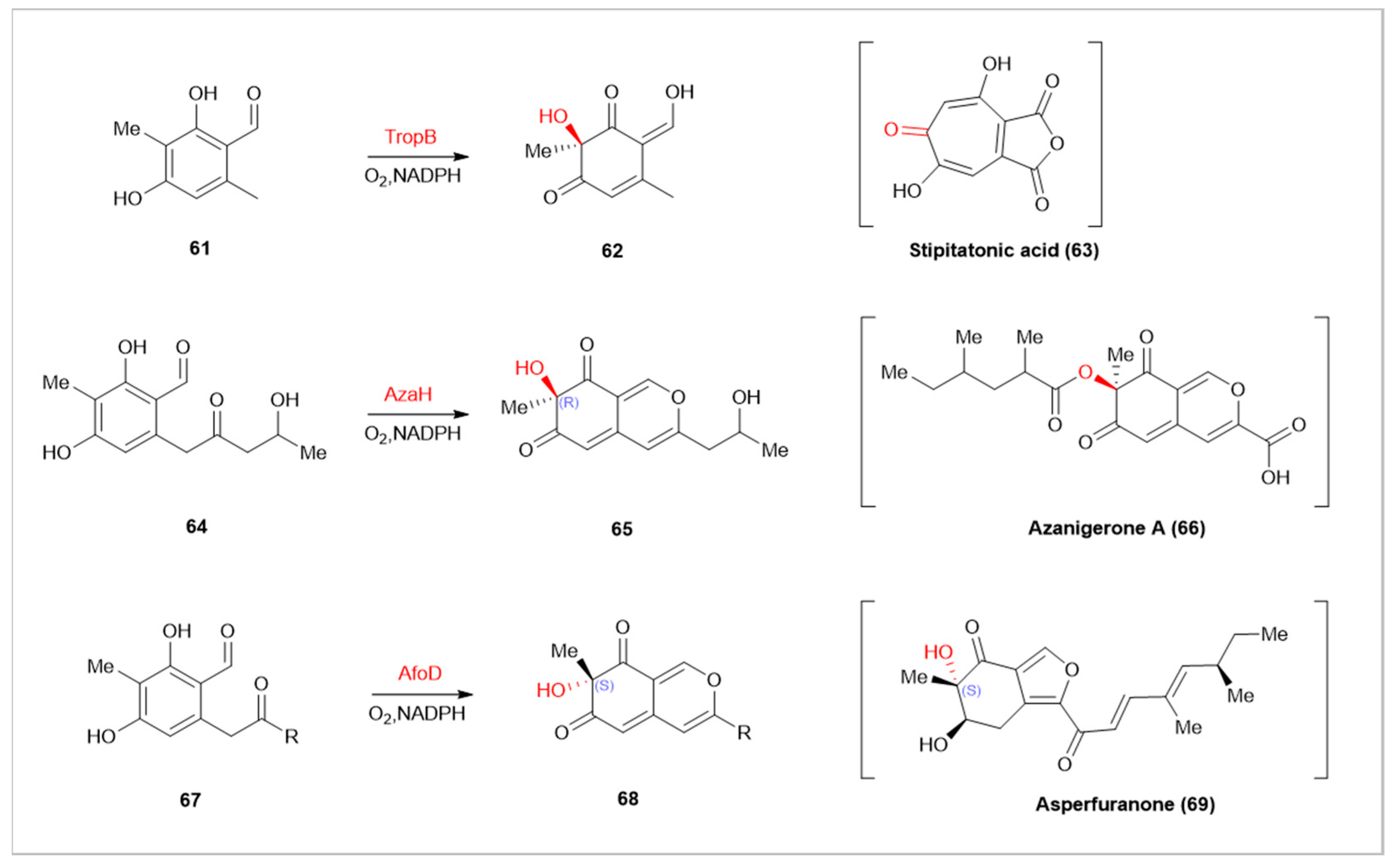
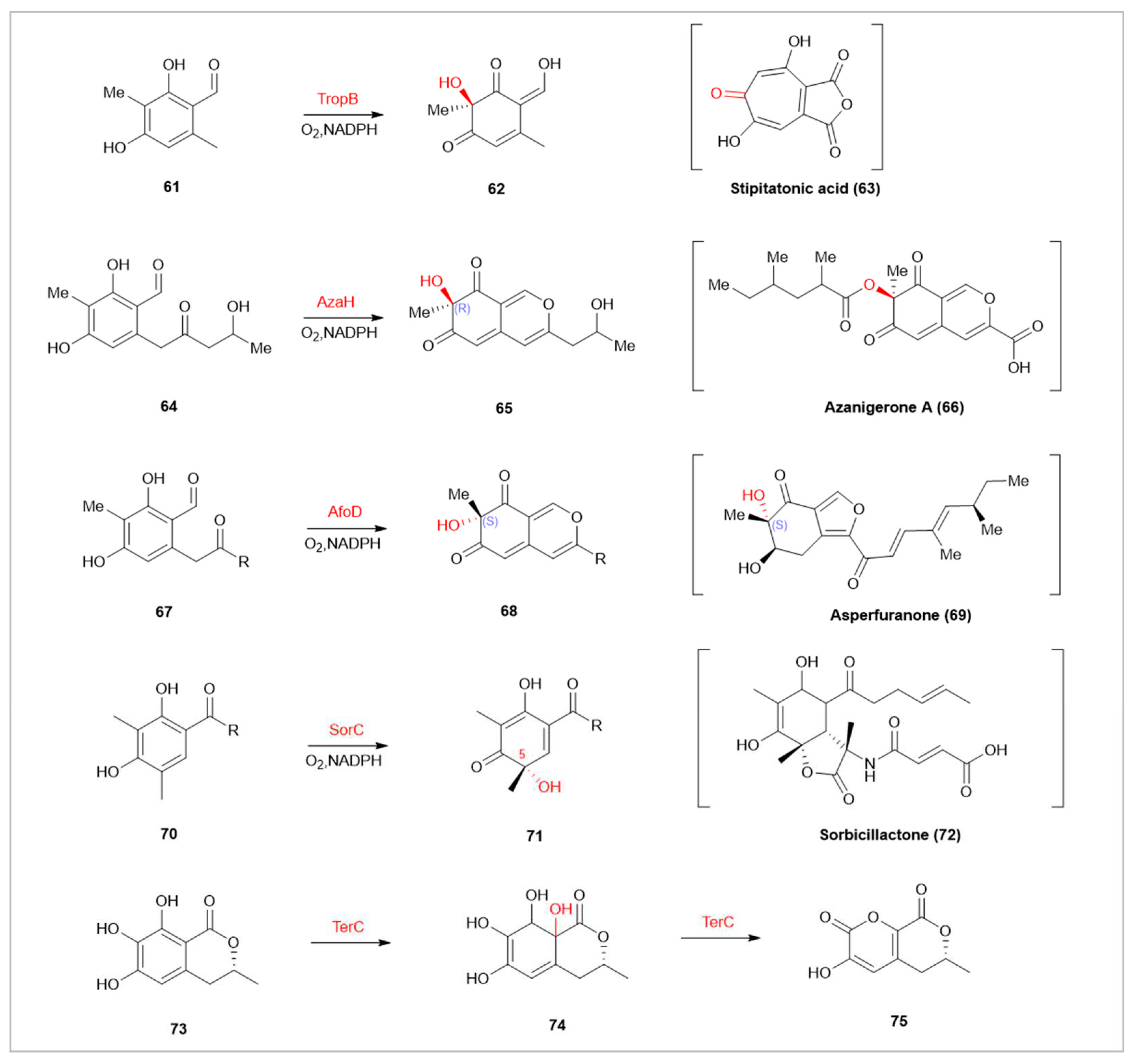
In natural product synthesis, FMOs participate in the oxidative dearomatization of various substances. Compared to traditional metal catalysts like (IIII, IV, PbIV, and CuI), FMOs offer unique stereoselectivity and precise site selectivity, reducing metal byproduct contamination and improving atom economy [82,83]. Tropolone is a class of seven-membered ring natural products, characterized by a core structure containing a ketone and a hydroxyl group [84]. Tropolones and their derivatives display multiple activities in biology and medicine, with puberulic acid (5-hydroxy stipitatic acid) showing potent anti-malarial activity (IC50= 10 ng·mL −1) [85]. TropB, a flavin-dependent monooxygenase encoded in the fungal natural product Stipitatonic acid, selectively hydroxylates 3-methyl-octanal(61) at the C-3 position for dearomatization [84]. Enzymes with similar mechanisms include AzaH and AfoD. AzaH, from a silent Aspergillus niger gene cluster, participates in the synthesis of azanigerone A(66), forming an “R” configuration at the newly formed stereocenter [86]. AfoD is involved in asperfuranone(69) synthesis, producing a complementary “S” configuration [87]. SorbC, involved in sorbicillactone A(72)synthesis, differs in site selectivity from AzaH, AfoD, and TropB [88]. Additionally, the flavin-dependent monooxygenase TerC, encoded by the terCDEF gene, catalyzes the dearomatization of 6-hydroxymellein (6-HM,73) to form 1,4-benzoquinone (75). This reaction, controlled solely by C-7 substitution, triggers a skeleton alteration through a bifurcated reaction cascade, forming benzoquinone or pyrone structures, offering a novel approach to [89].
Figure 8.
Flavin-dependent monooxygenases TropB, AzaH, and AfoD catalyze hydroxylation at the C-3 position.SorbC catalyze hydroxylation at the C-5 position. In parentheses are the final natural products synthesized in the respective pathways.
Figure 8.
Flavin-dependent monooxygenases TropB, AzaH, and AfoD catalyze hydroxylation at the C-3 position.SorbC catalyze hydroxylation at the C-5 position. In parentheses are the final natural products synthesized in the respective pathways.
4.3. Application of Baeyer-Villiger Monooxygenases in Natural Product Synthesis
Baeyer-Villiger monooxygenases (BVMOs) are a class of flavin-dependent monooxygenases that catalyze the Baeyer-Villiger (BV) oxidation of ketones and cyclic ketones into esters or lactones by inserting an oxygen atom near the carbonyl group, in the presence of cofactors. BVMOs can also oxidize heteroatoms, such as N-oxidation or sulfoxidation [90]. Compared to metal catalysts, BVMOs exhibit superior regioselectivity and enantioselectivity, and they can process a wide range of substrates, including cyclic and substituted cyclic, aromatic and linear ketones, aldehydes, bicyclic ketones, and various steroids [91]. BVMOs are categorized into two types based on the flavin cofactor used: Type I BVMOs contain oxidative and reductive domains with two dinucleotide motifs (Rossmann fold) for binding FAD and NAD(P)H [92,93], while Type II BVMOs consist of two different peptide components, one oxidative component binding FMN as a cofactor, and another reductive component utilizing NADH as a cofactor [94].
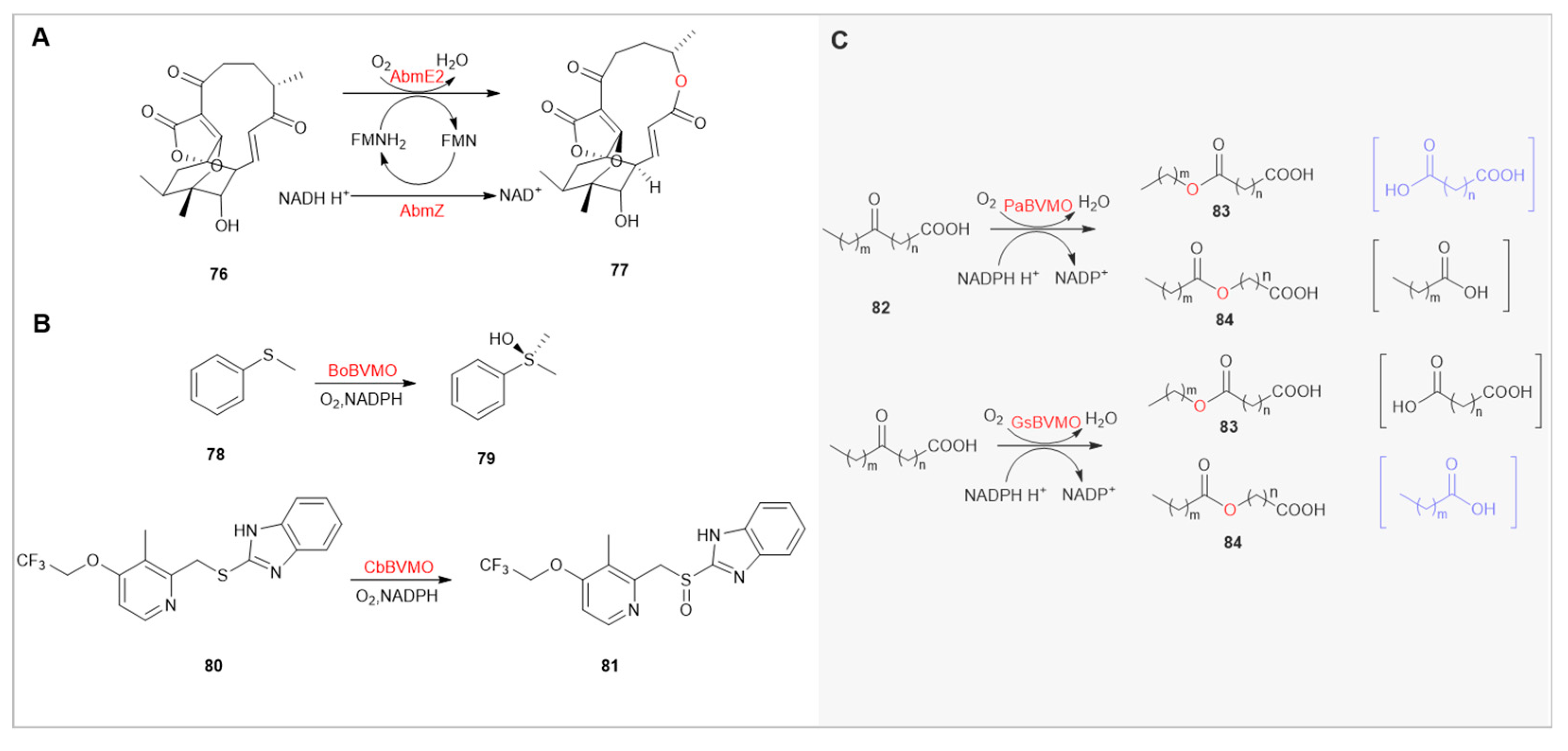
Abyssomicins/neoabyssomicins, isolated from Verrucosispora and Streptomyces species, are a class of anti-infective spirane lactone antibiotics [95,96,97]. AbmE2/AbmZ is a bifunctional BVMO involved in the catalytic conversion of abyssomicin 2(76) to neoabyssomicin B(77), categorized as Type II BVMOs. Interestingly, abyssomicin 2(76) exhibits antibacterial activity against Gram-positive pathogens, including clinically methicillin-resistant Staphylococcus aureus (MRSA), with MIC values of 3-15 μg/mL, while its derivative neoabyssomicin B(77) does not show such activity. This suggests that AbmE2/AbmZ may act as a resistance gene, employing a self-defense strategy through the transformation of toxic substances [98].
Optically active sulfoxide structures are extensively used in triazole drugs, such as the chiral proton pump inhibitors dexlansoprazole (Dexilant™, the R-enantiomer of lansoprazole) [99] dexrabeprazole (Dexpure™, the R-enantiomer of rabeprazole), and esomeprazole (Nexium™, the S-enantiomer of omeprazole) [100], with esomeprazole widely used in clinical treatment of gastrointestinal disorders [101]. BoBVMO is the first natural enzyme capable of catalyzing the asymmetric sulfoxidation of bulky prazole thioethers, showing its highest activity with benzyl methyl sulfide (specific activity of 0.117 U/mg), but only a modest activity of 0.69 mU/mg with lansoprazole sulfide (LPS, precursors for lansoprazole,80) [102]. Through genomic mining of BoBVMO, CbBVMO from Cupriavidus basilensis (62% sequence similarity) exhibited a higher specific activity of 39 mU/mg with LPS(81), completing the full conversion of 10 mM LPS in 35 hours and showing good activity with several other prazole sulfides. Overall, the catalytic efficiency of CbBVMO remains suboptimal. This issue might be addressed adequately through protein engineering and reaction engineering [103].
In biobased chemicals, ω-hydroxy fatty acids (C8-C14) are significant functional compounds, widely used in fragrances, preservatives, adhesives, and pharmaceutical intermediates. These compounds are characterized by having both carboxylic and hydroxyl functional groups at opposite ends of the fatty chain [104,105,106]. Previous studies have shown that PpBVMO from Pseudomonas putida KT2440 catalyzes the insertion of oxygen atoms at high substitution sites in asymmetric linear ketones, followed by hydrolysis to produce corresponding ω-hydroxy fatty acids [107]. Similarly, PfBVMO from Pseudomonas fluorescens DSM 50106 can catalyze the formation of α,ω-dicarboxylic acids [108]. Building on this research, PaBVMO from Pseudomonas aeruginosa demonstrates higher regioselectivity than PfBVMO, especially in long-chain (C16-C19) ketones, producing up to 95% dicarboxylic monoesters [109]. Recent studies have focused on enhancing the selectivity for monocarboxylic esters (PpBVMO 83:84=26:74), as demonstrated by GsBVMO from G. sihwensis, with 54% sequence similarity to PpBVMO, exhibiting high selectivity and activity in catalyzing medium to long-chain ketone acids into monocarboxylic esters. Surprisingly, enzyme engineering studies on GsBVMO revealed that the mutant GsBVMOC308L is an efficient biocatalyst, effectively converting 10-ketostearic acid into 9-nonyloxy nonanoic acid (60.5 gL−1d−1) [110].
Figure 8.
(A) AbmE2/AbmZ catalyzes the formation of Neoabyssomicin B, Devoid of Antibacterial Activity, from Abyssomicin 2.(B) The Application of BoBVMO and CbBVMO in Acetone Synthesis.(C) The Catalytic application of PaBVMO and PpBVMO on linear fatty ketones. Among these, PaBVMO exhibits stereochemical selectivity (83:84=71:29), while GsBVMO demonstrates stereochemical selectivity (83:84=3:97), with the numbers in parentheses indicating the corresponding hydrolysis products.
Figure 8.
(A) AbmE2/AbmZ catalyzes the formation of Neoabyssomicin B, Devoid of Antibacterial Activity, from Abyssomicin 2.(B) The Application of BoBVMO and CbBVMO in Acetone Synthesis.(C) The Catalytic application of PaBVMO and PpBVMO on linear fatty ketones. Among these, PaBVMO exhibits stereochemical selectivity (83:84=71:29), while GsBVMO demonstrates stereochemical selectivity (83:84=3:97), with the numbers in parentheses indicating the corresponding hydrolysis products.
5. Summary and Outlook
Flavin-Dependent Monooxygenases (FMOs) play a significant role in the field of biocatalysis, exhibiting tremendous potential for future biosynthesis and drug development. These enzymes utilize flavin adenine dinucleotide (FAD) and flavin mononucleotide (FMN) as cofactors to catalyze various oxidation reactions, including heteroatom hydroxylation, Baeyer-Villiger oxidation, and epoxidation. FMOs are known for their high regio- and stereoselectivity, crucial in the pharmaceutical industry [6]. The uniqueness of FMOs lies in their diverse reaction types and efficient catalytic capabilities, introducing oxygen atoms through redox processes, thus imparting biological activity to substrates, a key aspect in drug discovery and design. Additionally, the inactivation of FMOs can serve as a target for identifying small molecules, detecting potential diseases, treating microbial infections, and developing specific inhibitors against their catalytic conformations [76]. However, current research on FMOs as drug targets faces significant challenges, as the structures of many key metabolic FMOs remain unresolved, which may become a focal point in future biomedicine research.
In aerobic microorganisms, Flavin-Dependent Monooxygenases (FMOs) play a leading role in the degradation of pesticides. They “attack” chemically stable pesticide substrates through oxygenation, leading to the release of heteroatoms, dearomatization of aromatic rings, and ring cleavage during the pesticide degradation process. Although FMOs have demonstrated their diverse catalytic capabilities, further elucidation of more FMOs is required, along with the combination of multiple enzymes, to develop more convenient, efficient, and safe biodegradation pathways [33].
Research on plant flavin-dependent monooxygenases (FMOs) remains sparse, as highlighted by Schlaich in 2007, stating, “without any known substrates for plant FMOs, the field would stall.” [46]. Indeed, the case of YUC6 suggests that plant FMOs may have additional physiological benefits unrelated to their catalytic oxidation functions. This revelation expands the significance of FMOs in the realm of plant metabolism and opens new directions for future agricultural applications, particularly in crop improvement. Although limited biochemical and structural analyses have hindered the development of plant FMOs research, it is undeniable that plant FMOs, as an underutilized class of enzymes, hold substantial potential [47].
It is well-known that natural products are major sources of various chemical products and pharmaceuticals. Traditional chemical synthesis struggles to replicate complex natural products with unique scaffolds. In this context, Flavin-Dependent Monooxygenases (FMOs) play a crucial role in the synthesis pathways as oxygenating enzymes. Although fewer in number compared to the CYP450 family in catalyzing natural product synthesis [111,112], FMOs offer unique advantages of self-sufficiency, high efficiency, selectivity, and fewer toxic byproducts in catalytic processes [113]. The catalytic mechanism framework of FMOs has been largely established, with the revelation of the N5 mechanism suggesting that some FMOs can complete oxidation in the absence of NAD(P)H [7]. The “IN” state of FADred as demonstrated by the GrhO5 crystal structure reveals that Group A FMOs can complete the catalytic cycle directly with substrates and NAD(P)H via FAD [114]. Despite significant advancements in understanding catalytic mechanisms and reaction types, several challenges remain. For instance, how to engineer and design enzymes rationally based on mechanisms to accommodate a broader range of substrates? How to address the expensive recycling cost of NAD(P)H? There are a few electrochemical and photochemical alternative cycling methods, but their efficiency is still not optimal [115,116].
We firmly believe that, in the current era of rapid research advancements, more catalytic mechanisms and structures related to FMOs will be uncovered. With the aid of advanced biotechnologies like computational design and directed evolution, the development and application of FMOs across various fields are set to expand significantly [117]. This will not only advance progress in biocatalysis and drug development involving FMOs but also open up more possibilities for exploring novel biochemical pathways and innovative therapeutic approaches.
Author Contributions
Y.G. conceptualized the review and wrote the manuscript. X.C. supervised and revised the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China, grant number 2022YFC2106100, the Natural Science Foundation of China, grant number 21807088 and 22377098, Projects of International Cooperation in Shaanxi Province of China, grant number 2023-GHYB-08, Open Project Program of State Key Laboratory of Cancer Biology, grant number CBSKL2022KF13, Scholarship Program for Science and Technology Activities of Returned Overseas Scholars, grant number 2022-004, and Open Funding Project of the State Key Laboratory of Bioreactor Engineering, grant number 2018OPEN05.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Macheroux, P.; Kappes, B.; et al. Flavogenomics—A genomic and structural view of flavin-dependent proteins. Febs Journal 2011, 278, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chemical Reviews 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, N.; et al. Triepoxide formation by a flavin-dependent monooxygenase in monensin biosynthesis. Nature communications 2023, 14, 6273. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, M.M.E.; Montersino, S.; et al. Flavin dependent monooxygenases. Archives of Biochemistry and Biophysics 2014, 544, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Arnold, F.H. Directed Evolution: Bringing New Chemistry to Life. Angewandte Chemie-International Edition 2018, 57, 4143–4148. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.M.; Zhou, Q.; et al. Properties and Mechanisms of Flavin-Dependent Monooxygenases and Their Applications in Natural Product Synthesis. International Journal of Molecular Sciences 2022, 23, 2622. [Google Scholar] [CrossRef] [PubMed]
- Teufel, R.; Miyanaga, A.; et al. Flavin-mediated dual oxidation controls an enzymatic Favorskii-type rearrangement. Nature 2013, 503, 552. [Google Scholar] [CrossRef] [PubMed]
- Eswaramoorthy, S.; Bonanno, J.B.; et al. Mechanism of action of a flavin-containing monooxygenase. Proceedings of the National Academy of Sciences of the United States of America 2006, 103, 9832–9837. [Google Scholar] [CrossRef] [PubMed]
- Cashman, J.R. Some distinctions between flavin-containing and cytochrome P450 monooxygenases. Biochemical and Biophysical Research Communications 2005, 338, 599–604. [Google Scholar] [CrossRef]
- Krueger, S.K.; Williams, D.E. Mammalian flavin-containing monooxygenases: Structure/function, genetic polymorphisms and role in drug metabolism. Pharmacology & Therapeutics 2005, 106, 357–387. [Google Scholar] [CrossRef]
- Ziegler, D.M. An overview of the mechanism, substrate specificities, and structure of FMOs. Drug Metabolism Reviews 2002, 34, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Finefield, J.M.; Frisvad, J.C.; et al. Fungal Origins of the Bicyclo 2.2.2 diazaoctane Ring System of Prenylated Indole Alkaloids. Journal of Natural Products 2012, 75, 812–833. [Google Scholar] [CrossRef]
- Sehlmeyer, S.; Wang, L.Z.; et al. Flavin-Dependent Monooxygenases as a Detoxification Mechanism in Insects: New Insights from the Arctiids (Lepidoptera). PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Kotani, T.; Yurimoto, H.; et al. Novel acetone metabolism in a propane-utilizing bacterium, Gordonia sp strain TY-5. Journal of Bacteriology 2007, 189, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Volkers, G.; Palm, G.J.; et al. Structural basis for a new tetracycline resistance mechanism relying on the TetX monooxygenase. Febs Letters 2011, 585, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Rudra, P.; Hurst-Hess, K.; et al. High Levels of Intrinsic Tetracycline Resistance in Mycobacterium abscessus Are Conferred by a Tetracycline-Modifying Monooxygenase. Antimicrobial Agents and Chemotherapy 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P. Resistance to trimethoprim-sulfamethoxazole. Clinical Infectious Diseases 2001, 32, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.R.; Moore, I.F.; et al. TetX is a flavin-dependent monooxygenase conferring resistance to tetracycline antibiotics. Journal of Biological Chemistry 2004, 279, 52346–52352. [Google Scholar] [CrossRef]
- Hashimoto, M.; Taguchi, T.; et al. Unveiling Two Consecutive Hydroxylations: Mechanisms of Aromatic Hydroxylations Catalyzed by Flavin-Dependent Monooxygenases for the Biosynthesis of Actinorhodin and Related Antibiotics. Chembiochem 2020, 21, 623–627. [Google Scholar] [CrossRef]
- Mole, D.J.; Webster, S.P.; et al. Kynurenine-3-monooxygenase inhibition prevents multiple organ failure in rodent models of acute pancreatitis. Nature Medicine 2016, 22, 202–209. [Google Scholar] [CrossRef]
- Chiang, Y.L.; Lei, H.L.; et al. KMO as a novel diagnostic and prognostic biomarker in canine mammary gland tumors. Cancer Research 2012, 72. [Google Scholar] [CrossRef]
- Huang, T.T.; Tseng, L.M.; et al. Kynurenine 3-monooxygenase upregulates pluripotent genes through β-catenin and promotes triple-negative breast cancer progression. Ebiomedicine 2020, 54. [Google Scholar] [CrossRef]
- Jacobs, K.R.; Guillemin, G.J.; et al. Development of a Rapid Fluorescence-Based High-Throughput Screening Assay to Identify Novel Kynurenine 3-Monooxygenase Inhibitor Scaffolds. Slas Discovery 2018, 23, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.S.; Iradukunda, E.C.; et al. Modulation of Enzyme Activity in the Kynurenine Pathway by Kynurenine Monooxygenase Inhibition. Frontiers in Molecular Biosciences 2019, 6. [Google Scholar] [CrossRef]
- Padyana, A.K.; Gross, S.; et al. Structure and inhibition mechanism of the catalytic domain of human squalene epoxidase. Nature Communications 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Chua, N.K.; et al. The shape of human squalene epoxidase expands the arsenal against cancer. Nature Communications 2019, 10. [Google Scholar] [CrossRef]
- Santos, A.G.; da Rocha, G.O.; et al. Occurrence of the potent mutagens 2-nitrobenzanthrone and 3-nitrobenzanthrone in fine airborne particles. Scientific Reports 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Kino, K. Catalytic activity of the two-component flavin-dependent monooxygenase from Pseudomonas aeruginosa toward cinnamic acid derivatives. Applied Microbiology and Biotechnology 2014, 98, 1145–1154. [Google Scholar] [CrossRef]
- Nakagawa, A.; Nakamura, S.; et al. Selection of the optimal tyrosine hydroxylation enzyme for (S)-reticuline production in Escherichia coli. Applied Microbiology and Biotechnology 2021, 105, 5433–5447. [Google Scholar] [CrossRef]
- Deng, Y.F.; Faivre, B. Structural and Functional Characterization of 4-Hydroxyphenylacetate 3-Hydroxylase from Escherichia coli. Chembiochem 2020, 21, 163–170. [Google Scholar] [CrossRef]
- Herrmann, S.; Dippe, M.; et al. Engineered Bacterial Flavin-Dependent Monooxygenases for the Regiospecific Hydroxylation of Polycyclic Phenols. Chembiochem 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Fraley, A.E.; Tran, H.T.; et al. Flavin-Dependent Monooxygenases NotI and NotI′ Mediate Spiro-Oxindole Formation in Biosynthesis of the Notoamides. Chembiochem 2020, 21, 2449–2454. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.G.; Chen, D.; et al. Oxygenases as Powerful Weapons in the Microbial Degradation of Pesticides. Annual Review of Microbiology 2022, 76, 325–348. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.W.; Liu, B.; et al. A Novel Aerobic Degradation Pathway for Thiobencarb Is Initiated by the TmoAB Two-Component Flavin Mononucleotide-Dependent Monooxygenase System in Acidovorax sp Strain T1. Applied and Environmental Microbiology 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.J.; Zhou, X.Y. Experiment and Simulation on Adsorption of 3,5,6-Trichloro-2-Pyridinol in Typical Farmland of Purple Soil, Southwestern China. Soil & Sediment Contamination 2017, 26, 345–363. [Google Scholar] [CrossRef]
- Fang, L.C.; Shi, T.Z.; et al. Kinetics and Catabolic Pathways of the Insecticide Chlorpyrifos, Annotation of the Degradation Genes, and Characterization of Enzymes TcpA and Fre in Cupriavidus nantongensis X1T. Journal of Agricultural and Food Chemistry 2019, 67, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Mu, Y.; et al. Comparative Transcriptome Analysis Reveals the Mechanism Underlying 3,5-Dibromo-4-Hydroxybenzoate Catabolism via a New Oxidative Decarboxylation Pathway. Applied and Environmental Microbiology 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Jin, W.; et al. Hydrolase CehA and Monooxygenase CfdC Are Responsible for Carbofuran Degradation in Sphingomonas sp Strain CDS-1. Applied and Environmental Microbiology 2018, 84. [Google Scholar] [CrossRef]
- Cheng, M.G.; Meng, Q.; et al. The Two-Component Monooxygenase MeaXY Initiates the Downstream Pathway of Chloroacetanilide Herbicide Catabolism in Sphingomonads. Applied and Environmental Microbiology 2017, 83. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, T.; et al. Two dcm Gene Clusters Essential for the Degradation of Diclofop-methyl in a Microbial Consortium of Rhodococcus sp JT-3 and Brevundimonas sp JT-9. Journal of Agricultural and Food Chemistry 2018, 66, 12217–12226. [Google Scholar] [CrossRef]
- Takeo, M.; Murakami, M.; et al. Mechanism of 4-Nitrophenol Oxidation in Rhodococcus sp Strain PN1: Characterization of the Two-Component 4-Nitrophenol Hydroxylase and Regulation of Its Expression. Journal of Bacteriology 2008, 190, 7367–7374. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Ke, Z.J.; et al. Hydrolase CehA and a Novel Two-Component 1-Naphthol Hydroxylase CehC1C2 are Responsible for the Two Initial Steps of Carbaryl Degradation in Rhizobium sp. X9. Journal of Agricultural and Food Chemistry 2020, 68, 14739–14747. [Google Scholar] [CrossRef]
- Riebel, A.; de Gonzalo, G.; et al. Expanding the biocatalytic toolbox of flavoprotein monooxygenases from Rhodococcus jostii RHA1. Journal of Molecular Catalysis B-Enzymatic 2013, 88, 20–25. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.; et al. Baeyer-Villiger monooxygenases in the biosynthesis of microbial secondary metabolites. Sheng wu gong cheng xue bao = Chinese journal of biotechnology 2019, 35, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Romero, E.; Castellanos, J.R.G.; et al. Same Substrate, Many Reactions: Oxygen Activation in Flavoenzymes. Chemical Reviews 2018, 118, 1742–1769. [Google Scholar] [CrossRef]
- Schlaich, N.L. Flavin-containing monooxygenases in plants: Looking beyond detox. Trends in Plant Science 2007, 12, 412–418. [Google Scholar] [CrossRef]
- Thodberg, S.; Neilson, E.J.H. The “Green” FMOs: Diversity, Functionality and Application of Plant Flavoproteins. Catalysts 2020, 10, 329. [Google Scholar] [CrossRef]
- Fraaije, M.W.; Kamerbeek, N.M. Identification of a Baeyer-Villiger monooxygenase sequence motif. Febs Letters 2002, 518, 43–47. [Google Scholar] [CrossRef]
- Malito, E.; Alfieri, A.; et al. Crystal structure of a Baeyer-Villiger monooxygenase. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 13157–13162. [Google Scholar] [CrossRef] [PubMed]
- Beneventi, E.; Niero, M.; et al. Discovery of Baeyer-Villiger monooxygenases from photosynthetic eukaryotes. Journal of Molecular Catalysis B-Enzymatic 2013, 98, 145–154. [Google Scholar] [CrossRef]
- Mascotti, M.L.; Ayub, M.J.; et al. Chopping and Changing: The Evolution of the Flavin-dependent Monooxygenases. Journal of Molecular Biology 2016, 428, 3131–3146. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D.; Christensen, S.K.; et al. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Jung, J.H.; et al. Activation of a flavin monooxygenase gene YUCCA7 enhances drought resistance in Arabidopsis. Planta 2012, 235, 923–938. [Google Scholar] [CrossRef]
- Won, C.; Shen, X.L.; et al. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 18518–18523. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Tanaka, K.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 18512–18517. [Google Scholar] [CrossRef]
- Cao, X.; Yang, H.L.; et al. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. International Journal of Molecular Sciences 2019, 20, 6343. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.R.; Chen, J.B. Genome-Wide Identification, Expression Analysis, and Potential Roles under Abiotic Stress of the YUCCA Gene Family in Mungbean (Vigna radiata L.). International Journal of Molecular Sciences 2023, 24, 1603. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.H.; Mashiguchi, K.; et al. The Biochemical Mechanism of Auxin Biosynthesis by an Arabidopsis YUCCA Flavin-containing Monooxygenase. Journal of Biological Chemistry 2013, 288, 1448–1457. [Google Scholar] [CrossRef]
- Cha, J.Y.; Kim, W.Y.; et al. A novel thiol-reductase activity of Arabidopsis YUC6 confers drought tolerance independently of auxin biosynthesis. Nature Communications 2015, 6. [Google Scholar] [CrossRef]
- Cha, J.Y.; Kim, M.R.; et al. The Thiol Reductase Activity of YUCCA6 Mediates Delayed Leaf Senescence by Regulating Genes Involved in Auxin Redistribution. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef]
- Yoshimoto, N.; Saito, K. S-Alk(en)ylcysteine sulfoxides in the genus Allium: Proposed biosynthesis, chemical conversion, and bioactivities. Journal of Experimental Botany 2019, 70, 4123–4137. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Onuma, M.; et al. Identification of a flavin-containing S-oxygenating monooxygenase involved in alliin biosynthesis in garlic. Plant Journal 2015, 83, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Mishina, T.E.; Zeier, J. The Arabidopsis flavin-dependent monooxygenase FMO1 is an essential component of biologically induced systemic acquired resistance. Plant Physiology 2006, 141, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Zeier, T.; et al. Flavin Monooxygenase-Generated N-Hydroxypipecolic Acid Is a Critical Element of Plant Systemic Immunity. Cell 2018, 173, 456. [Google Scholar] [CrossRef] [PubMed]
- Pye, C.R.; Bertin, M.J.; et al. Retrospective analysis of natural products provides insights for future discovery trends. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, 5601–5606. [Google Scholar] [CrossRef]
- Tintore, M.; Vidal-Jordana, A.; et al. Treatment of multiple sclerosis—Success from bench to bedside. Nature Reviews Neurology 2019, 15, 53–58. [Google Scholar] [CrossRef]
- Waltenberger, B.; Mocan, A.; et al. Natural Products to Counteract the Epidemic of Cardiovascular and Metabolic Disorders. Molecules 2016, 21, 807. [Google Scholar] [CrossRef]
- Hu, Z.F.; Gu, A.D.; et al. Construction and optimization of microbial cell factories for sustainable production of bioactive dammarenediol-II glucosides. Green Chemistry 2019, 21, 3286–3299. [Google Scholar] [CrossRef]
- Li, B.J.; Wang, H.; et al. Improving 10-deacetylbaccatin III-10-β-O-acetyltransferase catalytic fitness for Taxol production. Nature Communications 2017, 8, 15544. [Google Scholar] [CrossRef]
- Paddon, C.J.; Westfall, P.J.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Vogt, M.; et al. Construction of a Corynebacterium glutamicum platform strain for the production of stilbenes and (2S)-flavanones. Metabolic Engineering 2016, 38, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Araújo, R.G.; et al. Production of curcuminoids from tyrosine by a metabolically engineered Escherichia coli using caffeic acid as an intermediate. Biotechnology Journal 2015, 10, 599–U315. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Q.; Liu, H.; et al. Engineering the Biosynthesis of Caffeic Acid in Saccharomyces cerevisiae with Heterologous Enzyme Combinations. Engineering 2019, 5, 287–295. [Google Scholar] [CrossRef]
- Campbell, A.; Bauchart, P.; et al. Engineering of a Nepetalactol-Producing Platform Strain of Saccharomyces cerevisiae for the Production of Plant Seco-Iridoids. Acs Synthetic Biology 2016, 5, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Galanie, S.; Thodey, K.; et al. SYNTHETIC BIOLOGY Complete biosynthesis of opioids in yeast. Science 2015, 349, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.A.G.; Li, H.; et al. New frontiers in flavin-dependent monooxygenases. Archives of Biochemistry and Biophysics 2021, 699. [Google Scholar] [CrossRef]
- Cane, D.E. Unified Stereochemical Model of Polyether Antibiotic Structure and Biogenesis. J. Am. Chem. SOC. 1983, 105, 3594–3600. [Google Scholar] [CrossRef]
- Leadlay, P.F.; Staunton, J.; et al. Engineering of complex polyketide biosynthesis -: Insights from sequencing of the monensin biosynthetic gene cluster. Journal of Industrial Microbiology & Biotechnology 2001, 27, 360–367. [Google Scholar] [CrossRef]
- Goodrich, R.D.; Garrett, J.E.; et al. Influence of monensin on the performance of cattle. Journal of animal science 1984, 58, 1484–1498. [Google Scholar] [CrossRef]
- Bhatt, A.; Stark, C.B.W.; et al. Accumulation of an E, E, E-triene by the monensin-producing polyketide synthase when oxidative cyclization is blocked. Angewandte Chemie-International Edition 2005, 44, 7075–7078. [Google Scholar] [CrossRef]
- Suzuki, G.; Minami, A.; et al. Analysis of Enantiofacial Selective Epoxidation Catalyzed by Flavin-containing Monooxygenase Lsd18 Involved in Ionophore Polyether Lasalocid Biosynthesis. Chemistry Letters 2014, 43, 1779–1781. [Google Scholar] [CrossRef]
- Pinkerton, D.M.; Banwell, M.G.; et al. Chemoenzymatic Access to Versatile Epoxyquinol Synthons. Organic Letters 2009, 11, 4290–4293. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Romero, E.O.; et al. Chemoenzymatic Total Synthesis of Natural Products. Accounts of Chemical Research 2021, 54, 1374–1384. [Google Scholar] [CrossRef]
- Davison, J.; al Fahad, A.; et al. Genetic, molecular, and biochemical basis of fungal tropolone biosynthesis. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 7642–7647. [Google Scholar] [CrossRef]
- Iwatsuki, M.; Takada, S.; et al. In vitro and in vivo antimalarial activity of puberulic acid and its new analogs, viticolins A-C, produced by Penicillium sp FKI-4410. Journal of Antibiotics 2011, 64, 183–188. [Google Scholar] [CrossRef]
- Zabala, A.O.; Xu, W.; et al. Characterization of a Silent Azaphilone Gene Cluster from Aspergillus niger ATCC 1015 Reveals a Hydroxylation-Mediated Pyran-Ring Formation. Chemistry & Biology 2012, 19, 1049–1059. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Szewczyk, E.; et al. A Gene Cluster Containing Two Fungal Polyketide Synthases Encodes the Biosynthetic Pathway for a Polyketide, Asperfuranone, in Aspergillus nidulans. Journal of the American Chemical Society 2009, 131, 2965–2970. [Google Scholar] [CrossRef]
- al Fahad, A.; Abood, A.; et al. Oxidative dearomatisation: The key step of sorbicillinoid biosynthesis. Chemical Science 2014, 5, 523–527. [Google Scholar] [CrossRef]
- Shu, X.; Wei, G.Z.; et al. TerC Is a Multifunctional and Promiscuous Flavoprotein Monooxygenase That Catalyzes Bimodal Oxidative Transformations. Organic Letters 2021, 23, 8947–8951. [Google Scholar] [CrossRef]
- Leisch, H.; Morley, K.; et al. Baeyer-Villiger Monooxygenases: More Than Just Green Chemistry. Chemical Reviews 2011, 111, 4165–4222. [Google Scholar] [CrossRef]
- Tolmie, C.; Smit, M.S.; et al. Native roles of Baeyer-Villiger monooxygenases in the microbial metabolism of natural compounds. Natural Product Reports 2019, 36, 326–353. [Google Scholar] [CrossRef]
- van Berkel, W.J.H.; Kamerbeek, N.M.; et al. Flavoprotein monooxygenases, a diverse class of oxidative biocatalysts. Journal of Biotechnology 2006, 124, 670–689. [Google Scholar] [CrossRef]
- Yachnin, B.J.; Sprules, T.; et al. The Substrate-Bound Crystal Structure of a Baeyer-Villiger Monooxygenase Exhibits a Criegee-like Conformation. Journal of the American Chemical Society 2012, 134, 7788–7795. [Google Scholar] [CrossRef]
- Isupov, M.N.; Schröder, E.; et al. The oxygenating constituent of 3,6-diketocamphane monooxygenase from the CAM plasmid of Pseudomonas putida: The first crystal structure of a type II Baeyer-Villiger monooxygenase. Acta Crystallographica Section D-Biological Crystallography 2015, 71, 2344–2353. [Google Scholar] [CrossRef]
- Song, Y.X.; Li, Q.L.; et al. Neoabyssomicins A-C, polycyclic macrolactones from the deep-sea derived Streptomyces koyangensis SCSIO 5802. Tetrahedron 2017, 73, 5366–5372. [Google Scholar] [CrossRef]
- Wang, Q.; Song, F.H.; et al. Abyssomicins from the South China Sea Deep-Sea Sediment Verrucosispora sp.: Natural Thioether Michael Addition Adducts as Antitubercular Prodrugs. Angewandte Chemie-International Edition 2013, 52, 1231–1234. [Google Scholar] [CrossRef]
- Tu, J.J.; Li, S.T.; et al. Characterization and heterologous expression of the neoabyssomicin/abyssomicin biosynthetic gene cluster from Streptomyces koyangensis SCSIO 5802. Microbial Cell Factories 2018, 17. [Google Scholar] [CrossRef]
- Ji, X.Q.; Tu, J.J.; et al. A Luciferase-Like Monooxygenase and Flavin Reductase Pair AbmE2/AbmZ Catalyzes Baeyer-Villiger Oxidation in Neoabyssomicin Biosynthesis. Acs Catalysis 2020, 10, 2591–2595. [Google Scholar] [CrossRef]
- Fass, R.; Frazier, R. The role of dexlansoprazole modified-release in the management of gastroesophageal reflux disease. Therapeutic Advances in Gastroenterology 2017, 10, 243–251. [Google Scholar] [CrossRef]
- Mejia, A.; Kraft, W.K. Acid peptic diseases: Pharmacological approach to treatment. Expert review of clinical pharmacology 2009, 2, 295–314. [Google Scholar] [CrossRef]
- Robinson, M. Proton pump inhibitors: Update on their role in acid-related gastrointestinal diseases. International Journal of Clinical Practice 2005, 59, 709–715. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, F.; et al. Discovery of Two Native Baeyer-Villiger Monooxygenases for Asymmetric Synthesis of Bulky Chiral Sulfoxides. Applied and Environmental Microbiology 2018, 84. [Google Scholar] [CrossRef]
- Liu, F.; Shou, C.; et al. A Baeyer-Villiger monooxygenase from Cupriavidus basilensis catalyzes asymmetric synthesis of (R)-lansoprazole and other pharmaco-sulfoxides. Applied Microbiology and Biotechnology 2021, 105, 3169–3180. [Google Scholar] [CrossRef]
- Malca, S.H.; Scheps, D.; et al. Bacterial CYP153A monooxygenases for the synthesis of omega-hydroxylated fatty acids. Chemical Communications 2012, 48, 5115–5117. [Google Scholar] [CrossRef]
- Scheps, D.; Malca, S.H.; et al. Synthesis of ω-hydroxy dodecanoic acid based on an engineered CYP153A fusion construct. Microbial Biotechnology 2013, 6, 694–707. [Google Scholar] [CrossRef]
- Lu, W.H.; Ness, J.E.; et al. Biosynthesis of Monomers for Plastics from Renewable Oils. Journal of the American Chemical Society 2010, 132, 15451–15455. [Google Scholar] [CrossRef]
- Rehdorf, J.; Kirschner, A.; et al. Cloning, expression and characterization of a Baeyer-Villiger monooxygenase from Pseudomonas putida KT2440. Biotechnology Letters 2007, 29, 1393–1398. [Google Scholar] [CrossRef]
- Kirschner, A.; Altenbuchner, J.; et al. Cloning, expression, and characterization of a Baeyer-Villiger monooxygenase from Pseudomonas fluorescens DSM 50106 in E-coli. Applied Microbiology and Biotechnology 2007, 73, 1065–1072. [Google Scholar] [CrossRef]
- Yu, J.M.; Liu, Y.Y.; et al. Direct Access to Medium-Chain α,ω-Dicarboxylic Acids by Using a Baeyer-Villiger Monooxygenase of Abnormal Regioselectivity. Chembiochem 2018, 19, 2049–2054. [Google Scholar] [CrossRef]
- Zhang, G.X.; You, Z.N.; et al. Discovery and Engineering of a Novel Baeyer-Villiger Monooxygenase with High Normal Regioselectivity. Chembiochem 2021, 22, 1190–1195. [Google Scholar]
- Hansen, C.C.; Sorensen, M.; et al. Reconfigured Cyanogenic Glucoside Biosynthesis in Eucalyptus cladocalyx Involves a Cytochrome P450 CYP706C55. Plant Physiology 2018, 178, 1081–1095. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yamamoto, K.; et al. Identification and characterization of CYP79D16 and CYP71AN24 catalyzing the first and second steps in L-phenylalanine-derived cyanogenic glycoside biosynthesis in the Japanese apricot, Prunus mume Sieb. et Zucc. Plant Molecular Biology 2014, 86, 215–223. [Google Scholar] [CrossRef]
- Toplak, M.; Matthews, A.; et al. The devil is in the details: The chemical basis and mechanistic versatility of flavoprotein monooxygenases. Archives of Biochemistry and Biophysics 2021, 698. [Google Scholar] [CrossRef]
- Toplak, M.; Saleem-Batcha, R.; et al. Catalytic Control of Spiroketal Formation in Rubromycin Polyketide Biosynthesis. Angewandte Chemie-International Edition 2021, 60, 26960–26970. [Google Scholar] [CrossRef]
- Hollmann, F.; Taglieber, A.; et al. A light-driven stereoselective biocatalytic oxidation. Angewandte Chemie-International Edition 2007, 46, 2903–2906. [Google Scholar] [CrossRef]
- Hollmann, F.; Hofstetter, K.; et al. Non-enzymatic regeneration of nicotinamide and flavin cofactors for monooxygenase catalysis. Trends in Biotechnology 2006, 24, 163–171. [Google Scholar] [CrossRef]
- Bunzel, H.A.; Anderson, J.L.R.; et al. Designing better enzymes: Insights from directed evolution. Current Opinion in Structural Biology 2021, 67, 212–218. [Google Scholar] [CrossRef]
Figure 1.
The two catalytic mechanisms of FMOs: the C4a mechanism, labeled with alphabetic numeration (in white), and the N5 mechanism, labeled with numeric numeration (in gray). In the C4a catalytic mechanism, the cofactor NAD(P)H acts as the hydrogen donor, whereas, in the N5 mechanism, the substrate itself serves as the hydrogen donor.
Figure 1.
The two catalytic mechanisms of FMOs: the C4a mechanism, labeled with alphabetic numeration (in white), and the N5 mechanism, labeled with numeric numeration (in gray). In the C4a catalytic mechanism, the cofactor NAD(P)H acts as the hydrogen donor, whereas, in the N5 mechanism, the substrate itself serves as the hydrogen donor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



