
Submitted:
25 November 2023
Posted:
28 November 2023
Read the latest preprint version here
Abstract
From grape cultivation to ripening and harvest timing to processing, each step can be a critical point when it comes to wine quality and phenolic composition. In this study, the influence of winemaking technology on resveratrol and quercetin content, as well as other polyphenolic compounds, was investigated. Resveratrol is a non-flavonoid polyphenolic stilbene synthesized by grape skin when damaged by infectious diseases or ionizing radiation. Quercetin is a phenol found in grape skins and stems and is produced to protect grapes from UV light damage. Trans-resveratrol and quercetin are known to act as antioxidants, reduce the risk of atherosclerosis and type 2 diabetes, inhibit the growth of cancer cells, and prevent the release of allergic and inflammatory molecules. However, the question was whether red wine could be enriched with these phenols using an innovative winemaking technology. The innovative idea was to completely replace the cold maceration process with maceration with the addition of wild yeast (Torulaspora delbrueckii, Td). Maceration with the addition of wild yeast (Td) offers the following advantages over traditional cold maceration: (1) higher concentrations of trans-resveratrol (> 35-40%) and quercetin (> 35-40%) in the final wine, (2) the new wine has a higher potential for human health, (3) the wine has a better aroma and stability due to the higher mannoprotein content, and (4) the better energy efficiency in the production process. The study of stability during storage and aging also included derivatives of benzoic acid and hydroxycinnamic acid, piceid, catechin, naringenin, rutin, kaempherol, hesperetin, and anthocyanins. The study found that younger wines had higher phenolic content, while storage of the wine resulted in a decrease in total phenolic content, especially monomeric stilbenes and quercetin.
Keywords:
innovative technology
; Torulaspora delbrueckii
; non-Saccharomyces yeast
; phenolic compounds
; trans-resveratrol
; quercetin
1. Introduction
Resveratrol (3,5,4’-trihydroxystilbene) is a phenolic phytoalexin produced by grapevines in response to fungal infection [1]. It exists in trans- and cis-isomeric forms, with the former being much more abundant in wine. In red wine, resveratrol also can be found in its glycoside form (resveratrol-3-O-β-mono-D-glucoside; piceid). Trans-resveratrol and piceid are the major active constituents of red wine. In vitro research indicates that resveratrol has chemopreventive effects against cardiovascular disease, aging, and cancer. Trans-piceid is present in red wine to a greater extent than its aglycone, but hydrolysis of this glycosylated derivative may occur in the small intestine and liver, which would increase the amount of biologically active trans-resveratrol. The average content of piceid was ten times higher than that of resveratrol in red wine. In addition, piceid was the most abundant form of resveratrol in nature. Other glycosylated derivatives of resveratrol could possibly be present in wine, but the lack of literature data on their contribution to the total amount of resveratrol suggests their minor importance [2]. A number of studies suggest that piceid may have similar bioactivity to resveratrol, e.g., anticarcinogenic effects, inhibition of platelet aggregation, and antioxidant activity. For this reason, it was of interest to analyze piceid in our innovative wine. It acts as a pro-drug and is stable during transport from the mouth to the small intestine, where it undergoes metabolic conversion to active trans-resveratrol and is reabsorbed into the blood plasma. Piceid preserves resveratrol from degradation in the gastrointestinal tract [2]. In Bordeaux varieties, trans-resveratrol was not found in measurable amounts, but only trans-piceid, in amounts between 0.26 mg/l and 1.25 mg/l [3]. The influence of winemaking techniques and grape varieties on resveratrol content, total phenolic content and antioxidant potential of red wines was previously studied on 10 commercial Serbian red wines. It was clearly found that resveratrol content was very low (0.18-1.31 mg/L) in all studied wines [4]. It was also demonstrated that winemaking techniques influence the amount of phenolic compounds. The highest average resveratrol and total phenolic contents were found in Merlot (4.85 mg/L; 1208 mg/L GAE) and Cabernet Sauvignon (3.78 mg/L; 1410 mg/L GAE) wines [5]. Quercetin has unique biological properties that may improve mental/physical performance and reduce the risk of infection. These properties form the basis for potential benefits to overall health and disease resistance, including anticarcinogenic, anti-inflammatory, antiviral, antioxidant, and psychostimulant activities, as well as the ability to inhibit lipid peroxidation, platelet aggregation, and capillary permeability, and to stimulate mitochondrial biogenesis. The first study on the pharmacokinetics of quercetin in humans showed very low oral bioavailability after a single oral dose (~2%) [6]. In red winemaking, maceration with skin and seeds during fermentation results in a higher concentration of resveratrol and quercetin in red wines compared to white wines. Resveratrol and quercetin content in wine depends on many different factors, including grape variety, harvest year, climatic conditions, UV light, winemaking technique, selected yeast strain, and aging [5,7]. The techniques of skin extraction and enzymatic hydrolysis of the glucoside forms also play an important role in the resulting resveratrol and quercetin concentrations. One way to control phenolic content in wine is through the choice of yeast strain. Not only do yeasts play a role in alcoholic fermentation, but they are also responsible for biochemical, enzymatic and physical reactions during the process, and thus exert a significant influence on wine phenolic composition of the wine. Thus, resveratrol, which is present in glycoside form (piceid), can be hydrolyzed by β-glucosidases, resulting in an increased concentration of free resveratrol [8]. Fermentation of must with yeast strains of Saccharomyces cerevisiae (SC) and fermentation with mixed cultures of Saccharomyces/non-Saccharomyces and lactic acid bacteria are widely used in modern winemaking. Together with the selected microbial cultures, the timing of bacterial inoculation plays an important role in determining the chemical composition of the wine. In traditional winemaking, inoculation with lactic acid bacteria is performed at the end of alcoholic fermentation. This leads to undesirable conditions for malolactic fermentation due to high etanol concentration, low pH, possible degradation of sugars by heterofermentative bacteria, limited antibacterial effect of SO2, etc. [9]. Thus, co-inoculation could overcome these problems, but the interactions between yeasts (Saccharomyces or non-Saccharomyces) and lactic acid bacteria have been little studied [10]. On the other hand, many studies have highlighted the positive influence of non-Saccharomyces yeast strains on the chemical composition of wine [8,10,11]. Recent studies with non-Saccharomyces species described the intense effect of some strains on anthocyanin color and subsequent stability by greatly lowering wine pH of wine during fermentation [12], as well as co-inoculation with lactic acid bacteria [10].
The main focus of this study was to find out the contribution of the innovative technology (co-inoculation with mixed cultures) to increase the content of stilbenes and quercetin in wine. In addition to these compounds, which are of great benefit to human health, the enrichment of wine with other phenolics by co-inoculation with mixed cultures was also investigated. A new winemaking process could improve wine bioactivity, provide guidance to future wine producers, and identify knowledge gaps for future research.
2. Materials and Methods
2.1. Grape
The grapes used in this study came from vineyards in the village of Vinča in Topola (Republic of Serbia), which are characterised by particular microclimatic conditions and soil types (44°13'35.4 "N 20°39'11.1 "E). Cabernet Sauvignon grape variety was harvested in the state of technological maturity. The phytosanitary condition was 100% healthy. The sugar content of the must was 23.4°Bx and the acidity was 6.7 g/l, expressed as tartaric acid.
2.2. Standards and chemicals
Resveratrol > 99%, piceid> 99%, rutin hydrate 95%, caffeic acid, quercetin dihydrate 98%, chlorogenic acid 95%, kaempferol ≥97%, p-coumaric acid, vanillic acid, syringic acid ≥95%, naringenin 95%, hesperetin ≥95%, (+)-catechin ≥90%, trans-cinnamic acid 99+%, and 4-hydroxybenzoic acid were purchased from Sigma-Aldrich (Steinheim, Germany), gallic acid from Alfa Aesar (Heysham Lancaster, UK), and benzoic acid from Lach-Ner (Neratovice, Czech Republic). Anthocyanins, malvidin-3-glucoside (oenin chloride), cyanin-3-glucoside chloride (kuromanin chloride), and delphinidin-3-glucoside chloride (delphinin, myrtillin) were purchased from AppliChem, and petunidin-3-glucoside chloride and peonidin-3-glucoside chloride from Phytolab. Methanol and acetonitrile in HPLC grade were from Promochem LGC (Wesel, Germany), formic and acetic acids from Lach-Ner (Neratovice, Czech Republic), hydrochloric acid 35% from Roth (Karlsruhe, Germany).
2.3. Winemaking
2.3.1. Traditional processing
The first part of the current study was based on traditional winemaking technology (Figure 1a). The sample harvested in small beans (10 kg) after grape harvest was stored at 5℃. The next step was grape destemming and gentle crashing with sulfation (50 mg/kg K2S2O5). Cold maceration lasted 5 days at 8-10°C with daily pumping over (1 volume of juice per day) covered with CO2. After 5 days, the temperature was increased to 15℃ and then inoculated with 25 g/100 L yeast Saccharomyces cerevisiae ICV D 254 (Lallemand, Canada). After one-third of the fermentation time, the nutrient Fermaid E (Lallemand, Canada) was added at a rate of 30 g/100 L. During fermentation, the temperature was not allowed to exceed 24°C. Two delastage per day were performed, one in the morning with oxygenation and the second at night (closed without spraying). After 10 days, when alcoholic fermentation was completed, the temperature was lowered to 18℃, and delastage was performed every second day for the next 6 days (closed without spraying). The total maceration time was 21 days, and then the free run wine was separated from pomace and stored in a stainless steel tank. This pomace was pressed with a pneumatic press until a pressure of 0.8 bar was reached, and the liquid fraction thus obtained was mixed with the free-running wine. After 24 hours, the wine was decanted from the yeast less and the same procedure was repeated after 72 hours and inoculated with the selection bacteria Oenococcus oeni VP 41. Induced malolactic fermentation was carried out at 18℃ and stirring was done twice a week. After completion of malolactic fermentation, SO2 was added (50 mg/L) and the wine was decanted from the lees to large barrels (3800 L). Battonage was performed twice a week. After one month (end of December 2018), the wine was racked off the lees and transferred to barrels. During the next three months, the lees were mixed (battonage) twice a month. In March 2019, the wine was again racked off the lees and during the next 6 months, Nobless (Lallemand, Canada) inactive yeast was added every second month (3 additions) (10 g/100 L). During this period, yeast and Nobless were mixed once a month. In June 2019, the wine was racked off the lees and Noblesse and put back into barrels for the next 12 months. The temperature and the free SO2 content were controlled every month during the wine storage in the barrels (temperature 12±2℃, relative humidity 75%). In addition, the analyzes were performed.
2.3.2. Innovative processing
For the second experiment, the harvested grapes were stored at 16℃ (Figure 1b). In this case, grapes were sulfurized with 15 mg/kg K2S2O5 during destemming and gentle crashing. The enzyme preparation EXV (Lallemand, Canada) was added at a rate of 3 g/100 kg. The pH was 3.58 and was adjusted from 3.58 to 3.4 by adding tartaric acid. Crushed grapes were inoculated with Bio diva (Torulaspora delbrueckii) at a rate of 25 g/100 L. Temperature control (maintained at 18°C) and two delasteges per day, one in the morning and the second during the night (closed delastage without spraying), allowed a very slow fermentation. After 4 days, when the sugar content had decreased by 3°Bx, 20 g/100 L of Fermaid O (organic nutrition – Lallemand) was added and inoculated with the selection yeast Saccharomyces cerevisiae ICV D 254, 25 g/100 L according to the manufacturer's protocol rehydration. After 12 hours the addition of ICV D 254, the selection bacterium VP 41 (Oenococcus oeni – Lallemand, Canada) was inoculated. Two delastage were performed every day, one in the morning with spraying (with oxygenation) and one at night without spraying (closed), until the end of alcoholic fermentation. At 1/3 of the fermentation time, 30 g/100 L of the nutrient Fermaid E was added. The maximum temperature reached during alcoholic fermentation was 24°C. After the alcoholic fermentation was completed (after 14 days), the temperature was lowered to 18℃ and the process continued until the malolactic fermentation was completed on pomace. On every second day, delastage was performed without spraying and malolactic fermentation was monitored. After 8 days, malolactic fermentation was completed (total time on pomace 22 days). Free run wine was separated from the pomace and stored in a stainless steel tank with sulfurization (50 mg/L). Then, the pomace was pressed on a pneumatic press until a pressure of 0.8 bar was reached, and the liquid fraction obtained was mixed with the free run wine. After 24 hours the wine was drawn off the lees and after 72 hours the same procedure was applied, the SO2 content was adjusted and the wine was stored in large barrels (3800 L). To study the influence of wine aging in bottles on polyphenolic composition of the wine, part of the wine was bottled and analyzed after one year. The bottles were stored in horizontal position and closed with a cork. Aging was carried out under cellar conditions (temperature 12±2℃, relative humidity 75%).
2.4. HPLC anaysis
For all analyses performed, the wine samples were prepared by filtration through a membrane philtre (0.45 μm; Sartorius, USA). HPLC analyses were performed using an Agilent 1100 series liquid chromatograph (USA) consisting of a quaternary gradient pump, an autosampler with injection system (10–200 μL), a column heater, a UV–VIS detector and a software package. A reversed-phase Poroshell 120 EC-C18 column (4.6 × 100 mm, 2.7 μm; Agilent, USA) was used for all analyses. Quantification of focal compounds was performed using external calibrations [13].
2.4.1. Phenols
HPLC separation of the phenolic compounds in 5 µL of the sample was performed by gradient elution (A- 0.1% CH3COOH, B- 0.1% CH3COOH in acetonitrile) at a flow rate of 1.0 mL/min on a column heated to 25℃ (Figure 2). Detection was performed with a UV-VIS detector at the following wavelengths: 225 nm (vanillic acid, benzoic acid), 280 nm (gallic acid, 4-hydroxybenzoic acid, catechin, syringic acid, trans-cinnamic acid, hesperetin, naringenin), 305 nm (p-coumaric acid, resveratrol, piceid), 330 nm (chlorogenic acid, caffeic acid), 360 nm (rutin, quercetin, kaempferol) [13].
The stock solutions of the individual compounds were prepared by dissolving them in dimethyl sulfoxide (DMSO). Calibration solutions were prepared by mixing the individual components and appropriate dilution with 0.1% acetic acid in the concentration range 0.5-20/25 mg/L. The calibration curves showed excellent linearity with correlation coefficients over 0.998 for all compounds. The sensitivity of the determination was achieved by increasing the injected sample volume by a factor of five compared to the standard mixture, so that quantification limits of 0.1 mg/L were achieved, which is particularly important for less abundant compounds. The detection limit was tested and confirmed at 0.05 mg/L with a required signal-to-noise ratio (S/N ≥3).
2.4.2. Anthocyanins
Chromatographic separation of the five major anthocyanins in 100 mL of sample was achieved by gradient elution with water/formic acid/acetonitrile mixtures (A – 87:10:3, B - 40:10:50; flow rate 0.8 mL/min; run time 14 min) on a column heated at 40°C (Figure 3). Anthocyanins were detected at 518 nm (method based on Compendium of International Methods of Analysis – OIV, 2013, modified by Majkic et al. [13].
Mixtures of all five anthocyanins, prepared from stock solutions of each anthocyanin in methanol acidified with HCl to 1%, were diluted with the original mobile phase and used to generate calibration curves in the concentration range of 0.1 to 100 mg/L. The linearity of all five corresponding curves was demonstrated by correlation coefficients of more than 0.998. The lowest calibration value of 0.1 mg/L was set as the practical limit of quantification based on a confirmed S/N ≥10, while the detection limit (S/N ≥ 3) was verified at 0.05 mg/L.
2.5. Determination of titratable acidity
A volume of 25 mL was titrated with 0.25 M NaOH to determine the titratable acidity of the wine [14]. The end point of the titration (pH 7.0 ± 0.5) was indicated with a pH meter (Farnell, Denmark).
2.6. Determination of the sugar content
The sugar content (expressed in °Bx) was measured in the grape juice using a PAL-87S refractometer (Atago, Tokyo, Japan).
2.7. Statistical analysis
The statistical analyses were carried out using SPSS Statistic V22.0 software (IBM, Chicago, IL, USA; 2014).
3. Results
3.1. Stilbenes
Stilbenes are mainly found in the grape skins, so yeasts with increased enzymatic activity could increase the extraction of stilbenes from the grape into the wine. By applying the new, innovative technology, the concentration of resveratrol and piceid in Cabernet Sauvignon wine could be significantly increased (by a factor of two and four respectively) compared to the control wine produced using conventional technology (Table 1).
The concentration of piceid and trans-resveratrol in our Cabernet Sauvignon wine was higher in the wine produced with innovative winemaking technology (9.30±0.20 mg/L; 3.10±0.10 mg/L) than in the wine produced with traditional winemaking technology, which is consistent with a study by Escribano-Viana et al. (2019) [15], which showed higher levels of total stilbenes, cis-piceid, trans-piceid and trans-resveratrol compared to fermentation without non-Saccharomyces yeasts. The same study confirms the higher content of piceid than of free resveratrol. The higher piceid content could be attributed to incomplete enzymatic cleavage of trans-piceid during vinification [16]. This suggests that the β-glucosidase activity of yeasts could be inhibited under winemaking conditions due to the low pH, high initial glucose concentration and low aeration at the beginning of fermentation [17]. According to the literature, little is known about the effects of this specific culture mixture on stilbene content, but the study of individual factors (yeasts, enzyme preparation and malolactic bacteria) can be considered [9,16,17,18]. In contrast to traditional fermentation, in innovative fermentation, malolactic bacteria were inoculated at the beginning of fermentation, resulting in a lower pH that could inhibit β-glucosidase activity, which could describe a higher piceid content. However, it is the bacterial enzyme activity that increases the stilbene content [19], probably in synergy with the β-glucosidase activity of the yeasts. Obviously, the innovative technology has increased both the free and glucosidic form of resveratrol (piceid) content, which is characterized by a higher value, probably due to the lower adsorption by yeast and yeast cells [20]. According to Sato et al. (1997) [21], the content of trans-piceid in Cabernet Sauvignon wine varies between 0.37-2.6 mg/L. In general, the average content of trans-piceid was found to be three times higher than that of trans-resveratrol. Kostadinovic et al. (2012) [16] reported that Merlot wine produced with a 6-day maceration using French yeast contained a high concentration of trans-piceid (4.10±0.86 mg/L), which was similar to our results obtained with traditional winemaking technology. The content of resveratrol was determined in red wines from Greece (0.550–2.534 mg/L), which was higher than in our traditionally produced wine [22], but the innovative technology was characterised by a higher content (3.10 mg/L). Higher levels of trans-resveratrol were also found compared to Karaoglan wine from Turkey, whose maximum concentration was 2.68±0.16 mg/L after 15 days of maceration [23]. In the Croatian wine Crljenak kaštelanski, the resveratrol concentration ranged from 0.51 mg/L in the wine produced by the addition of Vinozym Vintage to 1.07 mg/L in the wine produced by the addition of SihazymExtro [24], which was less abundant compared to the values measured in our innovative wine (Table 1). This suggests that the enzyme preparation EXV used in our study in combination with mixed co-inoculation contributed to a higher resveratrol content. According to Alencar et al. [25], the maximum value for trans-resveratrol was reached after 10 days of maceration (2.2 mg/L). In some other studies, trans-resveratrol concentrations of 0–2.0 mg/L were found in red wine from Queensland [26], 2.1–2.5 mg/L in Italian wines [27] and 0.56–2.86 mg/L in Italian red wines [28]. Experiments with the Tempranillo grape variety confirmed that the composition of stilbenes in wine can be modulated by the use of specific fermentation starters such as Torulaspora delbrueckii and other non-Saccharomyces strains compared to SC [15]. It is hypothesised that the enzymatic activity of yeasts improves their extraction as well as the enzymatic activities of early inoculated lactic acid bacteria. Yeasts with β-glucosidase activity not only release aroma precursors during alcoholic fermentation of grape must, but can also hydrolyze resveratrol glucosides from grapes [17]. The use of only SC and EXV enzyme preparations on the same grape variety resulted in a lower content of trans-resveratrol [29]. Torulaspora delbrueckii produces glycerol or pyruvic acid and lower levels of acetic acid than most non-Saccharomyces yeasts, so it contributes to better organoleptic wine characteristics [30]. Its use as a starter for controlled slow fermentation and inoculation with SC after 4 days resulted in wines with higher stilbene content.
3.2. Quercetin
The quercetin content in our innovative wine was higher than that of other Cabernet Sauvignon wines produced in Serbia reported by Radovanović et al. (2010) [31], as well as the study by Lisov et al. (2020) [29], whose results did not exceed 1.73 mg/L using different combinations of yeasts and enzymatic preparations. Similar observations were made for the Croatian autochthonous grape variety Crljenak kaštelanski obtained with different enzymes and a five-day maceration [24], but the quercetin content was lower than in our innovative Cabernet Sauvignon. According to Hernández et al. (2007) [18], the amount of free flavonols in red wines could depend on the Lactobacillus strain carrying out malolactic fermentation. Regarding the use of mixed cultures, a recent study reported that the timing of bacterial inoculation did not result in significant changes in flavonol content [32]. The higher quercetin content in the innovative wine could therefore be due to the enzymatic activity of the inoculated yeast Torulaspora delbrueckii. The use of EXV as a highly concentrated pectinase preparation with a secondary hemicellulase activity could result in a higher quercetin content [24,33]. As reported by Artem et al. (2021) [34], the quercetin content in wines from 2016 was up to 5.94±1.7 mg/L, while the wine from 2017 was not as rich in this flavonol.
3.3. Biologically active phenols
The concentrations of other biologically active phenols from red wine (catechin, rutin, kaempherol, Table 2) were higher in our innovative wine than in the wine produced by traditional winemaking.
Among the flavonoids, catechin was the most abundant phenolic compound, which is consistent with other reports [24,35]. The innovative technology resulted in better extraction of catechin compared to the traditional method (Table 2), leading to higher catechin levels compared to Cabernet Sauvignon wine analysed in a previous Serbian study [31] and Monasterll wine from Spain, which ranged from 7.8 to 14.0 mg/L [33]. Prolonged contact of solids and must resulted in a higher catechin content in wine [29,34]. Thus, a 21-day maceration promoted a high catechin content of up to 29.2 mg/L in innovative wine. The combination of Saccharomyces cerevisiae and Torulaspora delbrueckii wild yeast used resulted in a higher catechin content, which was consistent with the results of Ngqumba et al. [11], who experimented with the same mixture of strains for the Chenin blanc grape variety. Syrah wines produced with mixed co-inoculations of Saccharomyces cerevisiae/Lactobacillus thermotolerans/Torulaspora delbrueckii had lower concentrations of total flavan-3-ols compared to SC-inoculated wines [10], which is consistent with the results reported in this study for catechin.
3.4. Phenolic acids
The concentrations of phenolic acids in Cabernet Sauvignon wines produced using traditional and innovative technology are shown in Table 3.
The gallic acid content was in the range of its average content in red wines (41.8± 24.0 mg/L) [36] and higher than reported by Radovanovic et al. [31] for Cabernet Sauvignon wine from Serbia (22.82±0.98 mg/L). According to the literature, different winemaking techniques such as pre-fermentative maceration, addition of enzymes, tannins, oak chips or Ganimede autowinemaker [37] did not result in higher gallic acid content than that obtained in our wine by both traditional and innovative winemaking techniques. As reported by Artem et al. [34], gallic acid was the predominant phenolic acid in wines, with values ranging from 9.77 to 57.44 mg/L. According to Ngqumbo et al [11], Torulaspora delbrueckii strains had a positive effect on gallic acid concentration, but no significant difference was found in our study with respect to the use of different yeast strains. The concentrations of other analysed phenolic acids detected in our study were lower compared to the results reported by Soto-Vazques et al. [37].
The content of p-coumaric acid in innovative and traditional wine were almost identical (up to 3.0 mg/L). It was reported that the p-coumaric acid concentration in Syrah wine was quite low at 0.2 mg/L [25]. However, Karaoglan wine was richer in this phenolic acid compared to our wines [23]. In contrast to our results, wines produced with a combination of Saccharomyces cerevisiae and Torulaspora delbrueckii yeast had higher levels of p-coumaric acid [11].
3.5. Anthocyanins
The anthocyanin profiles of the traditional and innovative wines are shown in Table 4. A statistically significant difference was found between the anthocyanin content of innovative and traditional wines (p˂0.05).
Yeasts are known to affect wine colour in several ways, including the adhsion/adsorption of pigment compounds to yeast cells. The final amount of anthocyanins in wine depends on environmental/agronomic factors and the conditions of fermentation (time, temperature), but the anthocyanin profile of different wine varieties is considered to be relatively stable [38].
As expected, malvidin-3-O-glucoside, the most abundant anthocyanin in grapes, was the dominant anthocyanin compound in both wines produced with traditional and innovative technology (Table 4), similar to the findings of Generalić Mekinić et al. [24]. The concentration of anthocyanins analysed in the different winemaking techniques was comparable to a study published for red wines from Spanish grape varieties [37]. According to Soto-Vázquez et al [37], the wines produced with the innovative technology had higher anthocyanin contents than those produced with conventional vinification, pre-fermentative maceration, the addition of enzymes and tannins, oak chips, the delestage technique and the Ganimede autowinemaker. Moreover, our results were consistent with those obtained for wines produced with a combination of Saccharomyces, non-Saccharomyces and Oenococcus oeni in successive inoculations compared to Saccharomyces cerevisiae reference wines [10]. Recently, Chen et al. [39] found an increase in total anthocyanins during the fermentation of Torulaspora delbrueckii/Saccharomyces cerevisiae in comparison with fermentation conducted by Saccharomyces cerevisiae alone. Mixed fermentation cultures can change the chemical profiles of the wine. The absorption of anthocyanins could be different between species and strains [30]. In general, Torulaspora delbrueckii has been reported to increase anthocyanin content during alcoholic fermentation [15] and probably competes with Saccharomyces yeast, which adsorbs these compounds less through the cell walls. As for O. oeni, with a cell size several orders of magnitude smaller than that of S. cerevisae, its cells have a much smaller surface area [40]. In this study, malvidin-3-O-glucoside was the most abundant compound among the anthocyanins, possibly due to its low polarity and lower probability of being adsorbed by the yeast cells [40]. Significantly higher levels of malvidin-3-O-glucoside have been reported compared to our wine [41,42]. According to Minnaar et al. (2019) [10], the content of malvidin-3-O-glucoside in Saccharomyces wines after malolactic fermentation (MLF) was significantly lower than in wines with mixed culture inoculation (SC/non-Saccharomyces and O. Oeni), which is consistent with our results. According to Burns and Osborne (2014) [40], significantly higher levels of all individual anthocyanins were found in wines inoculated with Oenococcus oeni compared to the control (without MLF). It is known that malolactic bacteria produce enzymes such as glycosidases which act on the acylated anthocyanins and could therefore influence the anthocyanin profile of the wines [32]. When comparing the co-inoculation of yeasts and malolactic bacteria with sequential traditional MLF, a higher content of monomeric anthocyanins was found in co-inoculated wines. This suggests the involvement of bacterial enzymatic activities such as glycosidases during early inoculation [32], which is consistent with our experiment. Co-inoculation of Saccharomyces and non-Saccharomyces yeasts as well as the addition of malolactic bacteria during alcoholic fermentation may contribute to higher anthocyanin content in wine. In addition to Saccharomyces yeasts, Torulaspora yeasts have been reported to contribute to the intensity and stability of wine colour through the production of pyrucic acid [30].
3.6. Wine after storage
The storage and maturation of the wine in the bottle has an influence on the polyphenol content due to the interactions between the phenolic compounds. As shown in Figure 4a, no decrease in anthocyanins was observed during storage, which is in contrast to the reports of Ivanova et al. [43]. A smaller decrease in anthocyanin content was observed in wines which were macerated longer [43], as they are stabilized by participation in complexes or polymers with flavan-3-ols. In contrast to other studies [44,45], our wines did not undergo degradation of the monomeric anthocyanins. During wine storage, there is a chemical equilibrium between monomeric and copigmented anthocyanins. A decrease in the concentration of monomeric anthocyanins due to polymerization reactions alters this chemical balance. Another possibility would be the dissociation of copigmented anthocyanins, which increases the concentration of monomeric anthocyanins [46], which can be related to our results. Similarly, in a study by Carew et al. [47], no significant decrease in average anthocyanin content was found for any yeast treatment between bottling and 6 months of bottle age. According to Castillo-Sánchez et al. [44], the anthocyanin contents in young red wines depend strongly on the type of vinification, but these differences are significantly smaller in aged red wines. During storage, the anthocyanins are polymerized with other compounds and cannot be determined by HPLC. The differences between the anthocyanin contents in aged red wines determined by HPLC and spectrophotometric methods were greater than the differences observed between red wines at the time of bottling and aged wines [44].
A decrease in the content of catechin, rutin, quercetin and kaempherol (Figure 4b) was observed, but these changes were not significant (p˃0.05). According to other studies, a decrease in flavan-3-ols was observed, partly due to polymerization with anthocyanins and partly due to the formation of procyanidins [45,48]. The decrease in flavonols is related to the fact that they can be oxidized by coupled reactions or act as co-pigments with anthocyanins in co-pigmentation processes. A decrease in flavonol glycosides and an increase in the corresponding aglycones was observed, which is attributed to enzymatic hydrolysis [49]. Quercetin was under the influence of oxygenation, indicating its high reactivity with oxygen [50], which could be a reason for the degradation in our wine during the two-year maturation in the barrel before storage in the bottle.
Our results also showed a decrease in the content of phenolic acids (Figure 5a) and flavonols during wine storage, which is in agreement with Suprun et al. [51]. Bautista Ortin et al. [33] reported that the content of p-coumaric acid increased by one third after 8 months of bottle storage. In our study, the increase in p-coumaric acid could be due to the enzymatic hydrolysis of trans-coumaric acid and especially trans-caftaric acid in favour of caffeic acid and coumaric acid, which is in agreement with García-Falcón et al. [52]. The increased coumaric acid content can also be attributed to the degradation of cumarylanthocyanins during storage of the wines [52].
As shown in Figure 5b, aged wine contained a lower concentration of piceid (a bound form of resveratrol), which is due to its hydrolysis. As expected [53], the concentration of free trans-resveratrol in our wine increased with storage, which is due to the natural acidic hydrolysis of sugar–resveratrol complexes at storage temperature. Six months of storage resulted in a 1.2-fold decrease in total stilbene content [51]. In contrast to our results, Suprun et al. [51] reported that after storage the content of trans-resveratrol decreased from 14.1 % to 12.4 % of the total stilbenes detected.
3.6.1. Principal component analysis
Principal component analysis (PCA) was used to determine which winemaking process was most suitable in terms of the highest phenolic content or the potential aging of the wine. This statistical technique allows the reduction of the multidimensionality of the data, thus facilitating the search for relationships between the objects, the determination of the correlation structure of the variables and the study of how many components (a linear combination of the original characteristics) are required to explain most of the variance containing the characteristics of the reduced elements [29,54].
The grouping of wines was based on the content of anthocyanins, phenolic acids, stilbenes, flavonols and flavanols analyzed for three different winemaking processes (two winemaking processes – innovative and traditional and maturation of the innovative wine). After performing a PCA analysis of the wines, a component with a total diversity of 98.46% was extracted. The parameters of the statistical PCA analysis were the Kaiser-Meyer-Olkin criterion 0.763 and the Bartlett test of sphericity with statistical significance (p<0.05). In this case, the analysis without rotation was applied, as it allowed a maximum description of the variances by new components. Comparing the most significant factor loadings, the innovative wine shows a significantly different phenolic composition after bottling and after maturation and compared to the traditionally produced wine.
4. Conclusions
The proposed co-inoculation protocol is a good choice for the innovative vinification of Cabernet Sauvignon wines. Interestingly, the concentrations of resveratrol, piceid and quercetin were higher in these wines made with the innovative technology than in the wines made with the traditional technology - cold maceration. It is therefore clear that our innovative red wine production technology has achieved its objective: it has produced a wine rich in biologically active phenols and with numerous health benefits. Mixed culture co-inoculations with Saccharomyces cerevisiae cultures together with non-Saccharomyces and O.oeni represent a practical way to improve the phenolic composition of wine. Overall, Torulaspora delbueckii appears to be a good non-Saccharomyces yeast strain capable of increasing the content of anthocyanins and stilbenes as well as other bioactive compounds. The use of non-Saccharomyces yeast strains in co-inoculation with malolactic bacteria and the use of pectolytic enzyme preparations could replace cold maceration and thus improve the quality and health properties of the wine.
Author Contributions
conceptualization, A.P. and V.N.; methodology, J.C. and L.T.; formal analysis, A.P. and L.T.; investigation, N.Ž. and A.B.; resources, A.P. and L.G.B.; writing —creation of the original draft, N.Ž. and A.P.; writing — review and editing, L.T. and A.P.; visualization, A.B.; supervision, A.P.; project administration, A.P.; and fundraising, A.P. and L.G.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the Ministry of Science, Technological Development and Innovation of the Republic of Serbia and the University of Belgrade - Faculty of Agriculture (No. 451-03-47/2023-01/200116 and 451-03-1202/2021-09) and by The Innovation Fund Serbia (Cabernet Sauvignon wine with Td-enriched resveratrol and quercetin concentrations, 50138).
Conflicts of Interest
The authors declare that there are no conflicts of interest. The funders had no influence on the design of the study, the collection, analysis or interpretation of the data, the writing of the manuscript or the decision to publish the results.
References
- Langcake, P.; Pryce, R. J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Physiol. Plant Pathol. 1976, 9(1), 77-86. [CrossRef]
- Téguo, P.; Hawthorne, M. E.; Cuendet, M.; Mérillon, J. M.; Kinghorn, A. D.; Pezzuto, J. M.; Mehta, R. G. Potential cancer-chemopreventive activities of wine stilbenoids and flavans extracted from grape (Vitis vinifera) cell cultures. Nutr. Cancer. 2001, 40(2), 173-179. [CrossRef]
- Balga, I.; Lesko, A.; Ladanyi, M.; Kallay, M. Influence of ageing on changes in polyphenolic compounds in red wines. Czech J. Food Sci. 2015, 32, 563-569. http://dx.doi.org/10.17221/138/2014-CJFS.
- Cvejic, J. M.; Djekic, S. V.; Petrovic, A. V.; Atanackovic, M. T.; Jovic, S. M.; Brceski, I. D.; Gojkovic-Bukarica, L. C. Determination of trans-and cis-resveratrol in Serbian commercial wines. J. Chromatogr. Sci. 2010, 48(3), 229-234. [CrossRef]
- Atanacković, M.; Petrović, A.; Jović, S.; Gojković-Bukarica, L.; Bursać, M.; Cvejić, J. Influence of winemaking techniques on the resveratrol content, total phenolic content and antioxidant potential of red wines. Food Chem. 2012, 131(2), 513-518. [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M. T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8(3), 167. [CrossRef]
- Cantos, E.; Espín, J. C.; Fernández, M. J.; Oliva, J.; Tomás-Barberán, F. A. Postharvest UV-C-irradiated grapes as a potential source for producing stilbene-enriched red wines. J. Agric. Food Chem. 2003, 51(5), 1208-1214. [CrossRef]
- Zhang, P.; Ma, W.; Meng, Y.; Zhang, Y.; Jin, G.; Fang, Z. Wine phenolic profile altered by yeast: Mechanisms and influences. Compr. Rev. Food Sci. Food Saf., 2021, 20(4), 3579-3619. [CrossRef]
- Tristezza, M.; di Feo, L.; Tufariello, M.; Grieco, F.; Capozzi, V.; Spano, G.; Mita, G. Simultaneous inoculation of yeasts and lactic acid bacteria: Effects on fermentation dynamics and chemical composition of Negroamaro wine. LWT-Food Sci. Technol. 2016, 66, 406-412. [CrossRef]
- Minnaar, P. P.; Du Plessis, H. W.; Jolly, N. P.; Van Der Rijst, M.; Du Toit, M. Non-Saccharomyces yeast and lactic acid bacteria in Co-inoculated fermentations with two Saccharomyces cerevisiae yeast strains: A strategy to improve the phenolic content of Syrah wine. Food Chem. 2019, 4, 100070. [CrossRef]
- Ngqumba, Z.; Ntushelo, N.; Jolly, N. P.; Ximba, B. J.; Minnaar, P. P. Effect of Torulaspora delbrueckii yeast treatment on flavanols and phenolic acids of Chenin blanc wines. S. Afr. J. Enol. 2017, 38(2), 192-200. http://dx.doi.org/10.21548/38-2-1391.
- Morata, A.; Escott, C.; Loira, I.; Del Fresno, J. M.; González, C.; Suárez-Lepe, J. A. Influence of Saccharomyces and non-Saccharomyces yeasts in the formation of pyranoanthocyanins and polymeric pigments during red wine making. Molecules 2019, 24(24), 4490. http://dx.doi.org/10.3390/molecules24244490.
- Majkić, T. M.; Torović, L. D.; Lesjak, M. M.; Četojević-Simin, D. D.; Beara, I. N. Activity profiling of Serbian and some other European Merlot wines in inflammation and oxidation processes. Food Res. Int. 2019, 121, 151-160. [CrossRef]
- Tanner, H.; Brunner, H.R. Gentranke-Analytik; Verlag Heller-Chemie und Verwaltunsgesellschaft mbH: Darmstadt, Germany, 1979.
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A. R.; González-Arenzana, L. Effect of the sequential inoculation of non-Saccharomyces/Saccharomyces on the anthocyans and stilbenes composition of tempranillo wines. Front. Microbiol. 2019, 10, 773. https://doi.10.3389/fmicb.2019.00773.
- Kostadinović, S.; Wilkens, A.; Stefova, M.; Ivanova, V.; Vojnoski, B.; Mirhosseini, H.; Winterhalter, P. Stilbene levels and antioxidant activity of Vranec and Merlot wines from Macedonia: Effect of variety and enological practices. Food Chem. 2012, 135(4), 3003-3009. [CrossRef]
- Gaensly, F.; Agustini, B. C.; da Silva, G. A.; Picheth, G.; Bonfim, T. M. B. Autochthonous yeasts with β-glucosidase activity increase resveratrol concentration during the alcoholic fermentation of Vitis labrusca grape must. J. Funct. Foods 2015, 19, 288-295. http://dx.doi.org/10.1016/j.jff.2015.09.041.
- Hernández, T.; Estrella, I.; Pérez-Gordo, M.; Alegría, E. G.; Tenorio, C.; Ruiz-Larrrea, F.; Moreno-Arribas, M. V. Contribution of malolactic fermentation by Oenococcus oeni and Lactobacillus plantarum to the changes in the nonanthocyanin polyphenolic composition of red wine. J. Agric. Food Chem. 2007, 55, 5260-5266. [CrossRef]
- Poussier M.; Guilloux-Benatier M.; Torres M.; Heras E.; Adrian M. Influence of different maceration techniques and microbial enzymatic activities on wine stilbene content. Am. J. Enol. Vitic. 2003, 54, 261-266. http://dx.doi.org/10.5344/ajev.2003.54.4.261.
- Tıraş,Z. S. E.; Okur, H. H.; Günay, Z.; Yıldırım, H. K. Different approaches to enhance resveratrol content in wine. Ciência Téc. Vitiv. 2022, 37, 13-28. [CrossRef]
- Sato, M.; Suzuki, Y.; Okuda, T.; Yokotsuka, K. Contents of resveratrol, piceid, and their isomers in commercially available wines made from grapes cultivated in Japan. Biosci. Biotechnol. Biochem. 1997, 61(11), 1800-1805. [CrossRef]
- Gerogiannaki-Christopoulou, M.; Athanasopoulos, P.; Kyriakidis, N.; Gerogiannaki, I. A.; Spanos, M. trans-Resveratrol in wines from the major Greek red and white grape varieties. Food Control 2006, 17(9), 700-706. [CrossRef]
- Kocabey, N.; Yilmaztekin, M.; Hayaloglu, A. A. Effect of maceration duration on physicochemical characteristics, organic acid, phenolic compounds and antioxidant activity of red wine from Vitis vinifera L. Karaoglan. J. Food Sci. Technol. 2016, 53(9), 3557-3565. http://dx.doi.org/10.1007/s13197-016-2335-4.
- Generalić Mekinić, I.; Skračić, Ž.; Kokeza, A.; Soldo, B.; Ljubenkov, I.; Banović, M.; Skroza, D. Effect of winemaking on phenolic profile, colour components and antioxidants in Crljenak kaštelanski (sin. Zinfandel, Primitivo, Tribidrag) wine. J. Food Sci. Technol. 2019, 56(4), 1841–1853. [CrossRef]
- Alencar, N. M. M.; Cazarin, C. B. B.; Corrêa, L. C.; Maróstica Junior, M. R.; Biasoto, A. C. T.; Behrens, J. H. Influence of maceration time on phenolic compounds and antioxidant activity of the Syrah must and wine. J. Food Biochem. 2017, 42, e12471. http://dx.doi.org/10.1111/jfbc.12471.
- Ginjom, I.; D’Arcy, B.; Caffin, N.; Gidley, M. Phenolic compound profiles in selected Queensland red wines at all stages of the wine-making process. Food Chem. 2011, 125(3), 823-834. [CrossRef]
- Gambuti, A.; Strollo, D.; Ugliano, M.; Lecce, L.; Moio, L. trans-Resveratrol, quercetin,(+)-catechin, and (−)-epicatechin content in south Italian monovarietal wines: relationship with maceration time and marc pressing during winemaking. J. Agric. Food Chem. 2004, 52(18), 5747-5751. [CrossRef]
- Careri, M.; Corradini, C.; Elviri, L.; Nicoletti, I.; Zagnoni, I. Direct HPLC analysis of quercetin and trans-resveratrol in red wine, grape, and winemaking by products. J. Agric. Food Chem. 2003, 51(18), 5226-5231. [CrossRef]
- Lisov, N.; Petrovic, A.; Čakar, U.; Jadranin, M.; Tešević, V.; Bukarica-Gojković, L. Extraction kinetic of some phenolic compounds during Cabernet Sauvignon alcoholic fermentation and antioxidant properties of derived wines. Maced. J. Chem. Chem. 2020, 39(2), 185-196. [CrossRef]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911-1922. [CrossRef]
- Radovanović, B. C.; Radovanović, A. N.; Souquet, J. M. Phenolic profile and free radical-scavenging activity of Cabernet Sauvignon wines of different geographical origins from the Balkan region. J. Agric. Food Chem. 2010, 90(14), 2455-2461. [CrossRef]
- Devi, A.; Anu-Appaiah, K. A.; Lin, T. F. Timing of inoculation of Oenococcus oeni and Lactobacillus plantarum in mixed malo-lactic culture along with compatible native yeast influences the polyphenolic, volatile and sensory profile of the Shiraz wines. LWT 2022, 158, 113130. [CrossRef]
- Bautista-Ortín, A. B.; Martínez-Cutillas, A.; Ros-García, J. M.; López-Roca, J. M.; Gómez-Plaza, E. Improving colour extraction and stability in red wines: the use of maceration enzymes and enological tannins. Int. J. Food Sci. Technol. 2005, 40(8), 867-878. [CrossRef]
- Artem, V.; Antoce, A. O.; Geana, E. I.; Ranca, A. Effect of grape yield and maceration time on phenolic composition of ‘Fetească neagră’organic wine. Not. Bot. Horti Agrobot. 2021, 49, 12345-12345. [CrossRef]
- Xia, E. Q.; Deng, G. F.; Guo, Y. J.; Li, H. B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11(2), 622-646. [CrossRef]
- Heras-Roger, J.; Díaz-Romero, C.; Darias-Martín, J. A comprehensive study of red wine properties according to variety. Food Chem. 2016, 196, 1224-1231. [CrossRef]
- Soto Vázquez, E.; Río Segade, S.; Orriols Fernández, I. Effect of the winemaking technique on phenolic composition and chromatic characteristics in young red wines. Eur. Food Res. Technol. 2010, 231(5), 789-802. http://dx.doi.org/10.1007/s00217-010-1332-5.
- [38]Garrido, J.; Borges, F. Wine and grape polyphenols—A chemical perspective. Food Res. Int. 2013, 54(2), 1844-1858. [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; Del Fresno, J. M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J. A.; Han. S.; Benito, S.. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51-63. [CrossRef]
- Burns, T. R.; Osborne, J. P. Loss of Pinot noir wine color and polymeric pigment after malolactic fermentation and potential causes. Am. J. Enol. Vitic. 2015, 66(2), 130-137. http://dx.doi.org/10.5344/ajev.2014.14061.
- Álvarez, I.; Aleixandre, J. L.; García, M. J.; Lizama, V. Impact of prefermentative maceration on the phenolic and volatile compounds in Monastrell red wines. Anal. Chim. Acta. 2006, 563(1-2), 109-115. [CrossRef]
- Pérez-Lamela, C.; García-Falcón, M. S.; Simal-Gándara, J.; Orriols-Fernández, I. Influence of grape variety, vine system and enological treatments on the colour stability of young red wines. Food Chem. 2007, 101(2), 601-606. [CrossRef]
- Ivanova, V.; Vojnoski, B.; Stefova, M. Effect of winemaking treatment and wine aging on phenolic content in Vranec wines. J. Food Sci. Technol. 2012, 49(2), 161-172. [CrossRef]
- Castillo-Sánchez, J. X.; García-Falcón, M. S.; Garrido, J.; Martínez-Carballo, E.; Martins-Dias, L. R.; Mejuto, X. C. Phenolic compounds and colour stability of Vinhao wines: Influence of wine-making protocol and fining agents. Food Chem. 2008, 106(1), 18-26. [CrossRef]
- Gómez-Plaza, E.; Gil-Muñoz, R.; López-Roca, J. M.; Martı́nez-Cutillas, A.; Fernández-Fernández, J. I. Maintenance of colour composition of a red wine during storage. Influence of prefermentative practices, maceration time and storage. LWT-Food Sci Technol. 2002, 35(1), 46-53. [CrossRef]
- Bimpilas, A.; Panagopoulou, M.; Tsimogiannis, D.; Oreopoulou, V. Anthocyanin copigmentation and color of wine: The effect of naturally obtained hydroxycinnamic acids as cofactors. Food Chem. 2016, 197, 39-46. [CrossRef]
- Carew, A. L.; Smith, P.; Close, D. C.; Curtin, C.; Dambergs, R. G. Yeast effects on Pinot noir wine phenolics, color, and tannin composition. J. Agric. Food Chem. 2013, 61(41), 9892-9898. [CrossRef]
- Marquez, A.; Serratosa, M. P.; Merida, J. Influence of bottle storage time on colour, phenolic composition and sensory properties of sweet red wines. Food Chem. 2014, 146, 507-514. [CrossRef]
- Bimpilas, A.; Tsimogiannis, D.; Balta-Brouma, K.; Lymperopoulou, T.; Oreopoulou, V. Evolution of phenolic compounds and metal content of wine during alcoholic fermentation and storage. Food Chem. 2015, 178, 164-171. [CrossRef]
- Castellari, M.; Matricardi, L.; Arfelli, G.; Galassi, S.; Amati, A. Level of single bioactive phenolics in red wine as a function of the oxygen supplied during storage. Food Chem. 2000, 69(1), 61-67. [CrossRef]
- Suprun, A. R.; Dubrovina, A. S.; Tyunin, A. P.; Kiselev, K. V. Profile of stilbenes and other phenolics in Fanagoria white and red Russian wines. Metabolites 2021, 11(4), 231. [CrossRef]
- García-Falcón, M. S.; Pérez-Lamela, C.; Martínez-Carballo, E.; Simal-Gándara, J. Determination of phenolic compounds in wines: Influence of bottle storage of young red wines on their evolution. Food Chem. 2007, 105(1), 248-259. [CrossRef]
- Naiker, M.; Anderson, S.; Johnson, J. B.; Mani, J. S.; Wakeling, L.; Bowry, V. Loss of trans-resveratrol during storage and ageing of red wines. Aust. J. Grape Wine Res. 2020, 26(4), 385-387. [CrossRef]
- Milovanovic, M.; Žeravík, J.; Obořil, M.; Pelcová, M.; Lacina, K.; Cakar, U.; Petrovic, A.; Glatz, Z.; Skládal, P. A novel method for classification of wine based on organic acids. Food Chem. 2019, 284, 296-302. [CrossRef]
Figure 1.
(a) Traditional processing; (b) Innovative processing.

Figure 2.
Chromatographic separation of piceid, resveratrol and quercetin (2 mg/L).
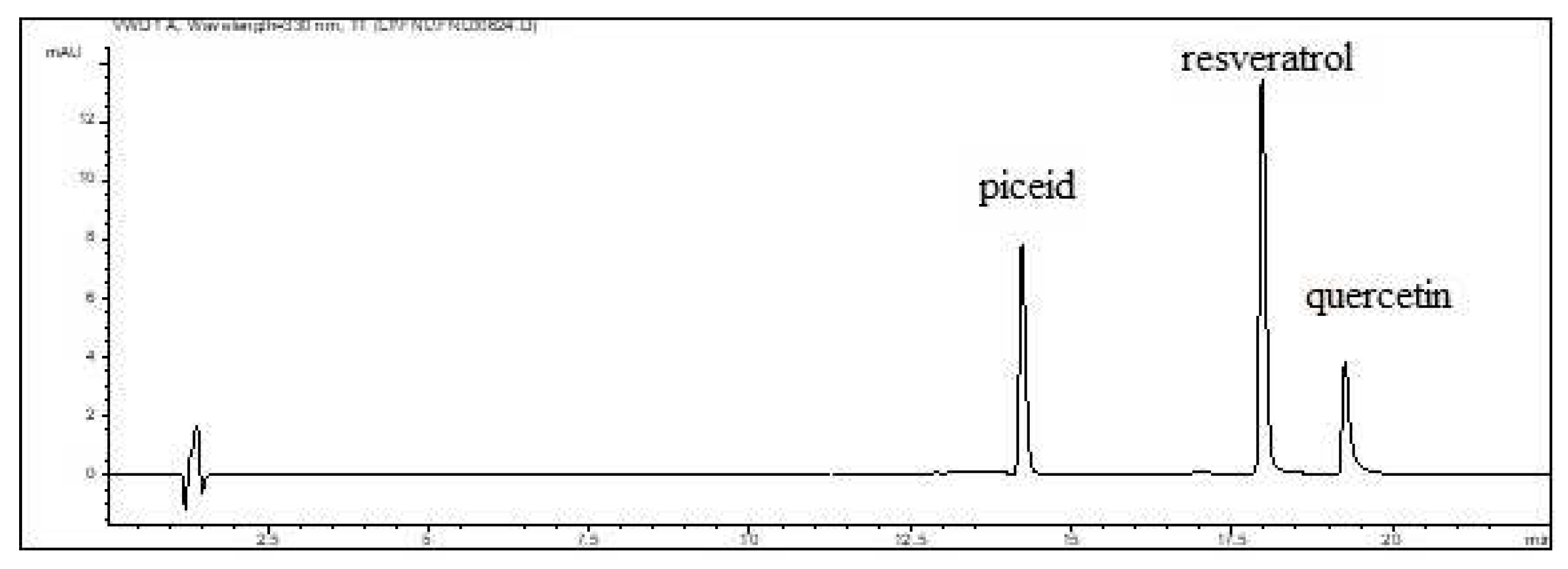
Figure 3.
Chromatographic separation of five anthocyanins (delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin 3-O-glucoside, peonidin 3-O-glucoside, malvidin 3-O-glucoside, respectively; 10 mg/L).
Figure 3.
Chromatographic separation of five anthocyanins (delphinidin 3-O-glucoside, cyanidin 3-O-glucoside, petunidin 3-O-glucoside, peonidin 3-O-glucoside, malvidin 3-O-glucoside, respectively; 10 mg/L).
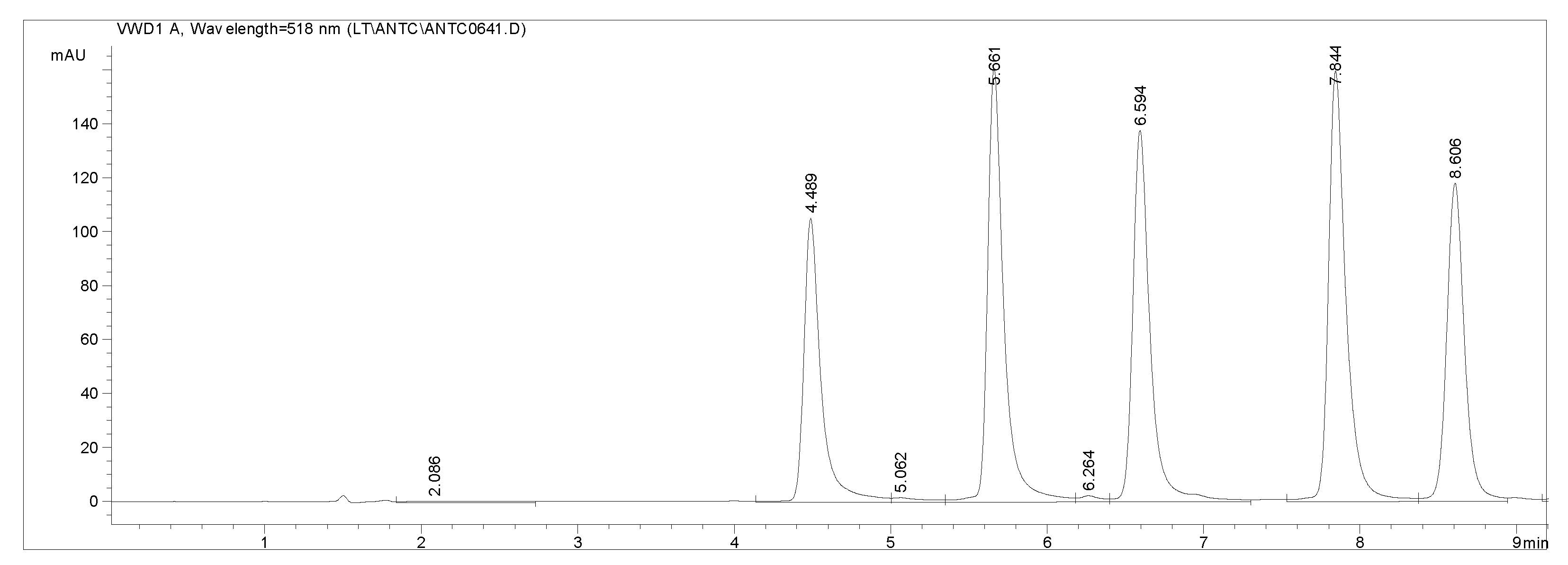
Figure 4.
(a) Content of anthocyanins in wine after storage in the bottle; (b) content of catechin, rutin, quercetin and kaempherol in wine after storage in the bottle.
Figure 4.
(a) Content of anthocyanins in wine after storage in the bottle; (b) content of catechin, rutin, quercetin and kaempherol in wine after storage in the bottle.
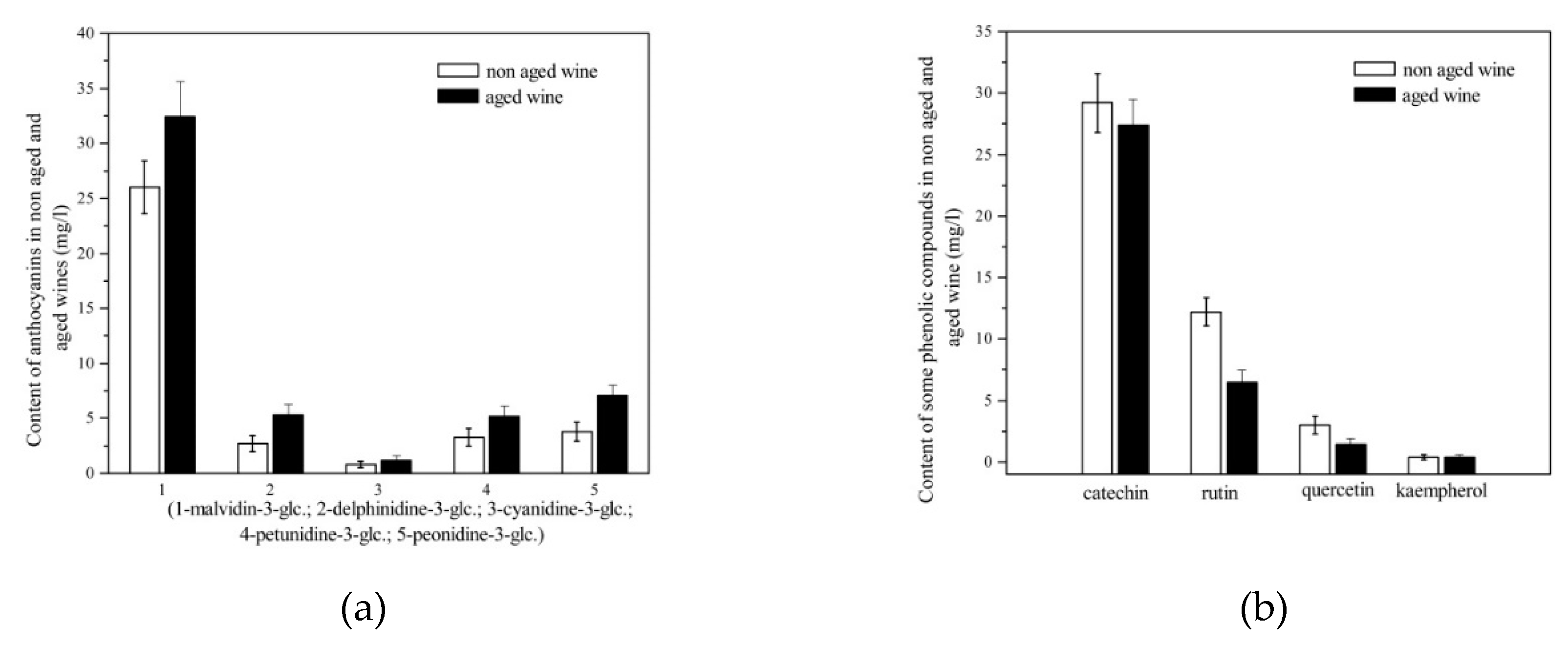
Figure 5.
(a) Content of phenolic acids in wine after storage in the bottle; (b) content of resveratrol and piceid in wine after storage in the bottle.
Figure 5.
(a) Content of phenolic acids in wine after storage in the bottle; (b) content of resveratrol and piceid in wine after storage in the bottle.
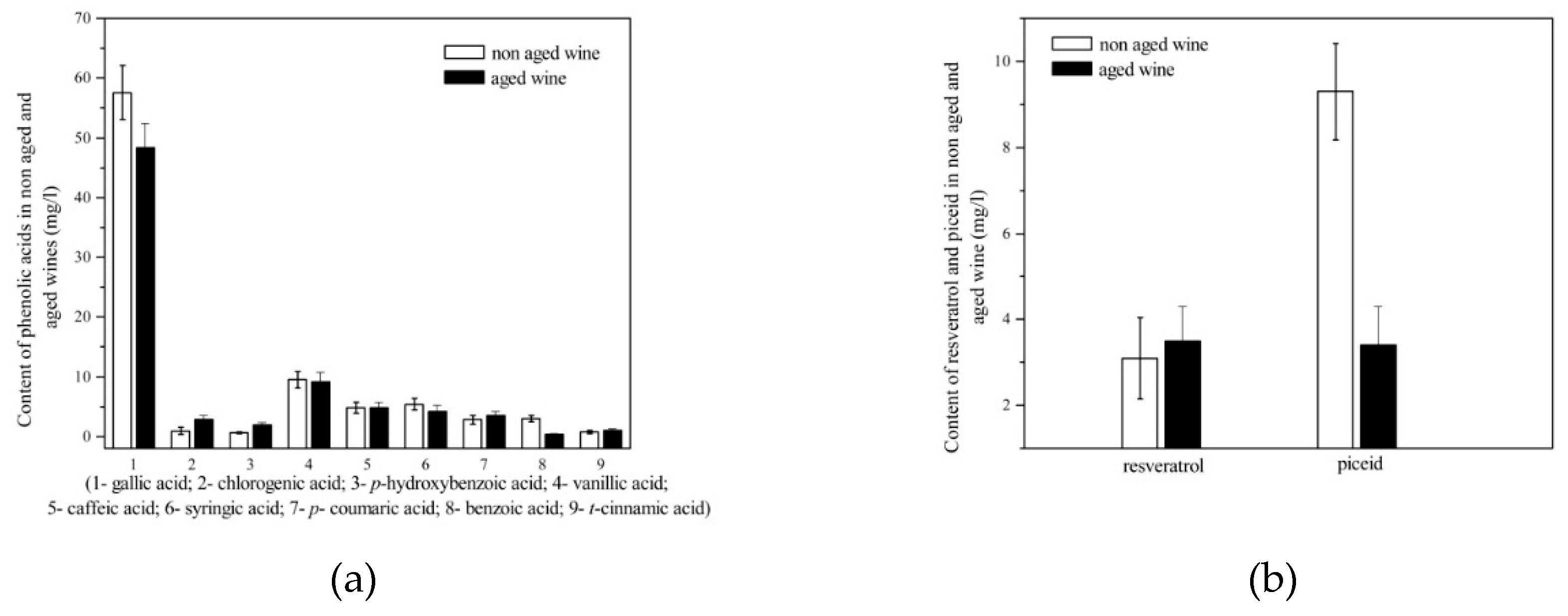

Table 1.
Concentration of stilbenes and quercetin in Cabernet Sauvignon wines produced by traditional and innovative technology.
Table 1.
Concentration of stilbenes and quercetin in Cabernet Sauvignon wines produced by traditional and innovative technology.
| Wine | Traditional winemaking technology |
Innovative winemaking technology |
% increase in content |
| trans-resveratrol (mg/L) | 1.30±0.09 | 3.10±0.10 | 138.5 |
| Piceid (mg/L) | 4.80±0.18 | 9.30±0.20 | 93.8 |
| Quercetin (mg/L) | 0.70±0.04 | 3.0±0.08 | 328.6 |
Table 2.
Concentration of other flavonoids in Cabernet Sauvignon wines produced by traditional and innovative technology.
Table 2.
Concentration of other flavonoids in Cabernet Sauvignon wines produced by traditional and innovative technology.
| Wine |
Traditional winemaking technology |
Innovative winemaking technology |
% increase in content |
| Catechin (mg/L) | 25.3±0.90 | 29.2±1.10 | 15.4 |
| Rutin (mg/L) | 7.3±0.20 | 12.2±0.20 | 67.1 |
| Kaempherol (mg/L) | n.d.* | 0.40±0.04 | 100.0 |
*nd- not detected; analysed, but not detected: naringenin and hesperetin.
Table 3.
Concentration of phenolic acids in Cabernet Sauvignon wines produced by traditional and innovative technology.
Table 3.
Concentration of phenolic acids in Cabernet Sauvignon wines produced by traditional and innovative technology.
| Phenolic acids (mg/l) |
Traditional winemaking technology |
Innovative winemaking technology |
| Gallic acid | 58.90±1.60 | 57.60±1.20 |
| Chlorogenic acid | 0.80±0.05 | 0.95±0.09 |
| p-hidroxybenzoic acid | 0.70±0.04 | 0.60±0.02 |
| Vanillic acid | 10.40±0.25 | 9.50±0.40 |
| Caffeic acid | 3.70±0.20 | 4.80±0.25 |
| Syringic acid | 6.90±0.30 | 5.40±0.20 |
| p-coumaric acid | 3.0±0.12 | 2.80±0.15 |
| Benzoic acid | 2.50±0.10 | 3.0±0.10 |
| t-cinnamic acid | 0.10±0.02 | 0.75±0.04 |
Table 4.
Concentration of anthocyanins in Cabernet Sauvignon wines produced by traditional and innovative technology.
Table 4.
Concentration of anthocyanins in Cabernet Sauvignon wines produced by traditional and innovative technology.
| Anthocyanin (mg/l) | Traditional winemaking technology |
Innovative winemaking technology |
% increase in content |
| delphinidin 3-O-glucoside |
1.60±0.15 | 2.70±0.25 | 68.80 |
| cyanidin 3-O-glucoside |
0.40±0.04 | 0.80±0.10 | 100.0 |
| petunidin 3-O-glucoside |
2.0±0.05 | 3.30±0.15 | 65.0 |
| peonidin 3-O-glucoside |
2.0±0.05 | 3.80±0.25 | 90.0 |
| malvidin 3-O-glucoside |
24.20±0.80 | 26.0±0.95 | 7.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



