
Submitted:
27 November 2023
Posted:
28 November 2023
You are already at the latest version
Abstract
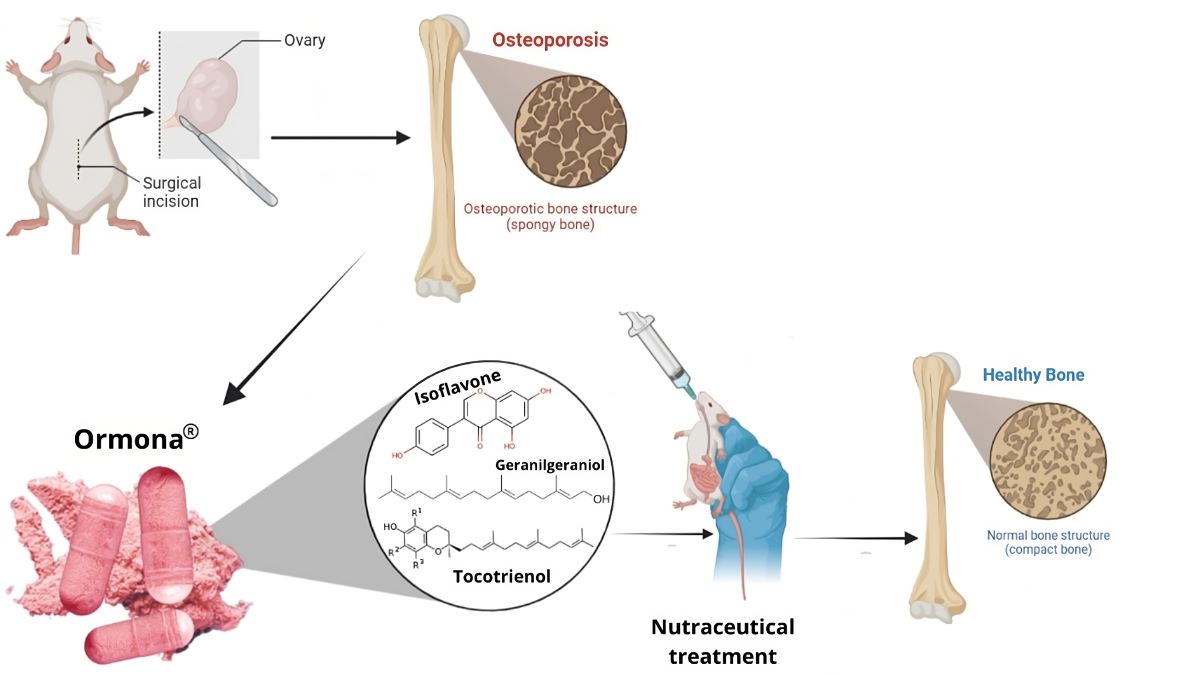
The phytonutraceutical Ormona® is a product composed of: purified oil of Bixa orellana Linné; dry extract of Myrciaria dubia McVaugh; dry extract of Trifolium pratense L. and dry extract of Euterpe oleracea Mart. obtained using Evolve® technology. This study evaluated the effects of Ormona® on Wistar rats affected by ovariectomy-induced osteoporosis. Pre-treatment was conducted for 15 days before surgery and continued for a further 45 days after the surgical procedure. The experimental design consisted of 5 groups (n=5): OVW: treated with distilled water (1 mL/ kg, p.o); ADS: alendronate sodium (4 mg/ kg p.o); EST: conjugated estrogen (2 ug/ kg, p.o); ORM: Ormona® (20 mg/kg, p.o); ORM + EST: Ormona® (20 mg/kg + conjugated estrogen 2 ug/ kg, p.o. Biochemical and hormonal parameters, along with bone histopathology and trabecular and femoral diaphysis size, were evaluated through Scanning Electron Microscopy (SEM) and bone calcium quantification by atomic absorption spectrophotometry. The results show that ovariectomy caused bone alterations such as loss of femoral microarchitecture, decreased bone homeostasis parameters, and changes in the lipid profile. Estrogen supplementation reduced parameters such as cholesterol, LDL, and Ca2+ concentration. However, Ormona® showed higher serum estradiol levels (p< 0.01), effects on the lipid profile, including parameters that estrogen replacement and alendronate sodium did not affect, with an increase in HDL, and positive modulation of bone metabolism, increasing osteocytes and the presence of osteoblasts. Ormona®, therefore, produced better results than the groups treated with estrogen and alendronate sodium.
Keywords:
Isoflavones
; Bone Density
; Osteoporosis
; Climacterium
; Estrogen receptors.
1. Introduction
Osteoporosis, a bone disease characterized by the loss of bone mass and deterioration of bone microarchitecture, is often associated with menopause in women. During menopause, the decrease in hormonal levels, especially estrogen, can trigger a significant increase in bone loss, substantially raising the risk of fractures and other bone complications. The interconnection between menopause and osteoporosis has been extensively documented, highlighting the pressing need for effective therapeutic interventions to mitigate the adverse effects of this debilitating condition. In this condition, the bones become more porous and lose density, becoming more prone to fractures (1–3).
Although hormone replacement therapy (HRT) has been a common approach to combat the effects of menopause on bone health, its potential side effects, such as an increased risk of breast cancer and cardiovascular diseases, have raised significant concerns (4,5). The ongoing search for safe and effective alternatives to HRT has led to a growing interest in exploring phytoestrogen compounds derived from plants, which can partially mimic the effects of endogenous estrogen in the body. Phytoestrogens have been considered promising candidates for managing menopausal symptoms and possibly preventing menopause-related osteoporosis (6). Such compounds have an agonistic effect on estrogen receptors (β-receptors), mimicking the action of physiological hormonal estradiol, thus replacing the estrogen deficit post-menopause (7).
In light of these advances, the research and development of nutraceuticals, defined as food products with health benefits beyond basic nutrition (8), has focused on formulating products that can exert an osteoprotective action, preventing bone loss and strengthening compromised bone structure. In this context, Ormona®, a phytonutraceutical derived from natural sources, has been the subject of interest due to its potential ability to mitigate the negative effects of osteoporosis, offering a promising and potentially safe alternative to conventional therapies.
2. Materials and Methods
2.1. Test Material
The OrmonaⓇ samples were kindly provided by Ages Bioactive Compounds Co. (São Paulo, SP, Brazil). The batch analysis certificate: ORM220702 (Expiry date 07/21/24) specifies the following composition: Purified oil of Bixa orellana Linné; dry extract of Myrciaria dubia McVaugh; dry extract of Trifolium pratense L. and dry extract of Euterpe oleracea Mart. The quantification method and its physicochemical characteristics are detailed in patent application BR102022008408-4.
Two standard products were used as positive controls: Premarin® - conjugated estrogens 0.3 mg (Pfizer Ireland Pharmaceuticals, imported by Wyeth Indústria Farmacêutica Ltda- Itapevi - SP) and Alendronate sodium 70 mg (EMS laboratory, Hortolândia/SP).
2.2. Animals and Ethical Aspects
According to the Brazilian College of Animal Experimentation and the Federal Council of Veterinary Medicine resolutions, this study was based on the Universal Declaration of Animal Rights recommendations. The project was submitted to the Ethics Committee for the Use of Animals in Research (CEUA) of the Federal University of Amapá and was approved under protocol number 017/2022.
The animals were acquired from the Multidisciplinary Center for Biological Research in Laboratory Animal Science (CEMIB) of the State University of Campinas (UNICAMP). The experiment was conducted at the Drug Research Laboratory, which belongs to the Department of Biological and Health Sciences located at the Federal University of Amapá (UNIFAP). Six-month-old female Rattus norvegicus Wistar weighing 200-250 g were used. They were acclimatized throughout the treatment and kept in a polyethylene cage at a controlled temperature of (~21ºC), with food and water ad libitum and a 12h light/dark cycle.
2.3. Experimental design and ovariectomy
The chosen dose of Ormona® was adapted from Tsuang et al. (11). Animals in the Ormona® groups were treated with a dose of 20 mg/kg concerning total isoflavones. The animals were divided into 5 experimental groups (n = 5):
- Negative control group (OVW): ovariectomized rats treated with distilled water (1 mL/kg, p.o);
- Sodium alendronate group (ADS): ovariectomized rats treated with Sodium Alendronate (4 mg/kg, p.o).
- Estrogen group (EST): ovariectomized rats treated with estrogen (2 µg/kg, p.o);
- Ormona® group (ORM): ovariectomized rats treated with Ormona® (20 mg/kg, p.o);
- Experimental group (ORM + EST): ovariectomized rats treated with Ormona® (20 mg/kg + estrogen 2 µg/kg, p.o);
The animals received pre-treatment for 15 days, and after this period, bilateral ovariectomy was performed, following the technique described by Yousefzadeh et al. (12). Once the animals had recovered from the surgery, treatment continued for another 45 days, totaling 60 days of treatment. The animals’ food and water intake were monitored throughout this period to assess their weight development.
2.4. Hormonal and Biochemical Analysis
On the 61st day of the experiment, 1.5 mL of blood was collected from the animals through the retro-orbital plexus. The samples underwent centrifugation for 10 minutes at 5000 rpm/min (13). The evaluated parameters included estrogen, progesterone, total cholesterol (TC) and fractions (HDL and LDL), triglycerides (TG), aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase, urea, and creatinine. The samples were processed at AlchemyPet Laboratory, which specializes in veterinary laboratory tests in Pompeia - São Paulo and registered with CRMV/SP under no. 42697-PJ.
2.5. Scanning electron microscopy (SEM) of the femur
The femurs of the euthanized rats were removed, cleaned, dehydrated in an oven at 100 ºC, and weighed using an electronic analytical balance (Bioprecisa Model, FA2104n). The specimens were sectioned longitudinally and transversely every 0.2 cm for Scanning Electron Microscopy (Hitachi TM3030PLUS) analysis. All the photographs were taken of the same area (on a scale of 1 mm to 2 mm) in all the groups, ensuring uniformity (14).
2.6. Quantification of Calcium in Bone Matrix by atomic absorption spectrophotometry
Approximately 200 mg of the bone matrix were digested with a solution of nitric and perchloric acid in a 2:1 ratio (nitric acid, HNO3; 65%, PA, Vetec; Sigma-Aldrich Ltd., São Paulo, SP, Brazil, and perchloric acid, HClO4; 70%, PA, ACS, Vetec; Sigma-Aldrich Co.), heated to 200 ºC for complete digestion of organic matter. After this step, the samples were transferred to 50 mL volumetric flasks. Calcium concentrations were determined by atomic absorption spectrophotometry (Shimadzu, Tokyo, Japan) with hollow cathode lamps (422.7 nm) and air-acetylene flame. A standard solution of Ca2+ 1000 ppm (Merck 1.09943 Titrisol, St. Louis, MA, USA) was used for the analytical curve (15).
2.7. Histopathological analysis of bone tissue
For the histopathological study, the femurs were collected and kept in 10% neutral buffered formalin for 72 hours for fixation and then decalcified in 8% hydrochloric acid solution and 10% formic acid in phosphate-buffered saline for 7 days. They were then dehydrated with alcohol baths in increasing concentrations, clarified in xylene, and embedded in paraffin (16)
The paraffin blocks with the samples were microtomized into 3 μm sections, placed on glass slides, stained with hematoxylin-eosin and analyzed using optical microscopy (Olympus BX41-69 Micronal), and photographed with a digital camera (MDCE-5C USB 2.0).
2.8. Statistical Analysis
For statistical analysis, all quantitative results were expressed as mean ± standard deviation, and the groups were compared using One-Way ANOVA followed by the Tukey test for multiple comparisons. The ORM and ORM+EST groups were compared using the Student’s t-test. Results with p < 0.001, p < 0.01, and p < 0.05 were considered statistically significant. The statistical software used was GraphPad Prism (version 7.0).
3. Results
3.1. Ponderal development
The effects of the treatments on weight development are shown in Figure 1. The OVW group had lower food and water intake and, consequently, lower body weight (p < 0.05). However, the ADS group had a statistically significant increase in average body weight (p < 0.05), water, and food intake. There was no weight loss in the groups treated with EST, ORM, and ORM+EST.
3.2. Hormonal and biochemical analysis
The hormonal profile of the treated groups after 60 days is illustrated in Figure 2. Serum hormone estradiol levels in the ORM group were significantly higher (p < 0.01) compared to all groups. The Student’s t-test revealed that the group treated with ORM+EST had a statistically lower estradiol level (p < 0.01) than the ORM group. The progesterone concentration tended to be higher in the rats in the OVW group after ovariectomy; however, this was not statistically significant.
Significant differences were observed in all parameters regarding the animals’ lipid profiles. Concerning total cholesterol levels, remarkably significant results were noted, with the lowest values for total cholesterol presented in the EST, ORM, and ORM+EST groups, all with p < 0.01 compared to OVW (Figure 3). Regarding triglycerides, the ORM and ORM+EST groups showed statistically significant lower values (p < 0.05) than the ADS group used as a positive control.
Although not significant, the ORM and ADS groups showed elevated levels of HDL. The EST and ORM+EST groups exhibited statistically significantly lower HDL levels, p < 0.01, than the OVW group. Meanwhile, the LDL level in the ADS group was higher compared to the OVW group. Both the group treated with estradiol alone (EST) and the ORM+EST group recorded the lowest LDL values (p < 0.01 and p < 0.05, respectively) compared to the OVW group. The ORM group showed a statistically significant lower level (p < 0.001) than the positive control group treated with ADS.
The levels of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in the animals in the estradiol-treated group (EST) showed a statistically significant higher level of AST (276.60 ± 102) and ALT (33.80 ± 7.22) (p < 0.01 and p < 0.05, respectively) when compared to the OVW group. In the group treated with OrmonaⓇ (ORM) and the animals treated with sodium alendronate (ADS), elevated levels were observed only for ALT (p < 0.01 and p < 0.0001, respectively), as shown in Table 1. There was no statistical difference between the groups in urea, creatinine, and alkaline phosphatase levels.
3.3. Scanning electron microscopy (SEM) of the femur and calcium quantification
The femur weight of the ORM group was statistically higher compared to the OVW group (p < 0.05) and the ORM+EST group (p < 0.01). Bone calcium concentrations were higher in the ORM group compared to the OVW group (p < 0.01) and the ADS group (p < 0.05). These results are consistent with the data obtained in the analysis of trabecular size and diaphysis thickness, where, despite no statistically significant difference, the ORM group tended to have a more minor trabecular extension and greater femoral diaphysis thickness (Figure 4)
Figure 5 illustrates the cross-sections of the femoral metaphysis with the trabeculae of the animals according to each group, in which the photomicrographs were obtained using scanning electron microscopy. The OVW negative control group (Figure 5A) shows deterioration in bone microarchitecture, with extensive and prominent trabeculae, indicating loss of bone density. In contrast, supplementation with EST (Figure 5C), ORM (Figure 5D), and ADS (Figure 5B) alone preserved the size of the trabeculae compared to the OVW group. However, there was no statistical difference in the average sizes.
Figure 5A and 5E show bone loss, mainly of the trabeculae, and the connection of the bone trabeculae was interrupted with significant bone loss in the OVW group. As shown in Figure 5B, 5C, and 5D, the structure of the bone tissue in the distal femur of the other treatment groups did not suffer bone loss, as evidenced by the uniform tissue morphology, indicating that the trabecular microstructure of the animals was not altered after the treatments.
3.4. Histopathological analysis of bone tissue
H&E staining showed through light microscopy that the thickness of the cortical bone and the trabecular bone area was reduced in the OVW negative control group (Figure 6A), with a thin trabecular meshwork, little medullary tissue, and loss of bone microarchitecture. There was a loss of connectivity in both the primary and secondary cancellous portions, where the endosteal surface of the OVW group appeared worn, with an absence of osteogenic lining. Many osteoporotic cavities, resorption cavities, and calcified cartilage were in the tissue analyzed.
However, the groups treated with Ormona® were able to attenuate the signs of bone degeneration in the ovariectomized rats. In particular, the ORM and ADS groups showed denser trabeculae with a significant presence of medullary tissue inside the trabeculae (Figure 6D and 6B, respectively).
The OVW group showed resorption of bone tissue, marked mineral loss, dispersed trabecular tissue with a fragmented shape, and rare osteogenic activity (Figure 8). In addition, there was a reduced number of osteocytes and lamellae with no medullary content, signaling a process of osteopenia. An improvement in morphological architecture was seen in the bone matrix of the groups treated with ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), ORM + EST (20 mg/kg + 2 µg/kg), revealing cell-rich bone tissue with osteocyte lacunae (arrows marked with OT) on the endosteal surface. It is also possible to observe an increase in trabecular bone density and the thickness of the cortical region in the treated groups (Figure 7B–E).
Treatment of the ORM group attenuated the pathological changes in preserving the spongy portion of the femur, evidenced by smaller trabeculae (Figure 6D). Bone mineral retention and trabecular bone density were marked with the ORM and ORM+EST treatments, and active proliferation of osteoblasts (marked with the acronym OB) was observed around the growth plate (Figure 6D and 6E). Meanwhile, treatment with ADS resulted in a smaller volume of trabecular tissue and minerals, demonstrating a retarding effect on bone loss in the tissue sections with EST, which was similar to that of ORM and ORM+EST.
4. Discussion
Ovariectomy is a preclinical model for post-menopausal osteoporosis and is considered the first choice, most commonly used for such studies (17,18), and recommended by the Food and Drug Administration (19). This experimental model has bone loss characteristics similar to osteoporosis in post-menopausal women. From the age of three months, when they are in the reproductive phase, rodents respond appropriately to the deficiency of sex hormones acquired due to ovariectomy. In this experimental model, in addition to bone loss, there is hypercholesterolemia, an increase in bone remodeling markers, a decrease in calcium absorption, and the protective effect of obesity on bone loss resulting from estrogen deficiency (20).
In this study, ovariectomized rats developed bone alterations similar to those observed during osteoporosis due to hypoestrogenism. As indicated by a decrease in femur weight, bone calcium content, an increase in alkaline phosphatase concentration, deformity of the microarchitecture of the epiphyses, a decrease in the number of osteocytes, and the absence of osteoblasts. Treatment with estrogen suppressed the effects of estrogen deficiency in most parameters. These results are in line with several studies that have used ovariectomy as a model for inducing osteoporosis in vivo (21-24).
In contrast to the clear bone-related benefits of HRT, its uncertain popularity has culminated in a clinical need for well-tolerated nutritional options that can be used safely after menopause and that effectively prevent bone loss and the development of osteoporosis. The phytonutraceutical OrmonaⓇ is a phytocomplex composed of 40% oil extract of annatto (Bixa orellana), with the same composition as ChronicⓇ (25,26). Among these 40%, ~10% are mainly δ-tocotrienols and ~28% are geranylgeraniol. In addition, it contains 20.8% red clover extract (Trifolium pratense), standardized with isoflavones such as daidzein, genistein, Biochanin A, and formononetin. 10% of the formulation is composed of a standardized extract of açaí (Euterpe oleracea) standardized with anthocyanins (mainly cyanidin 3-O-glucoside) (10).
Estrogens are directly involved in regulating energy homeostasis and are crucial in modulating food intake and energy expenditure. The literature reports that reduced levels of estradiol (E2, the main ovarian estrogen) after menopause in women and ovariectomy (OVX) in rodents lead to hyperphagia and reduced energy expenditure (EE), inducing overall weight gain (27-29). There is evidence that the effects of estradiol on food intake are regulated by estrogen receptors in the hypothalamus (for example, the arcuate nucleus - ARC and the hypothalamic paraventricular nucleus - PVN) and also in the nucleus of the solitary tract in the brainstem. However, which estrogen receptor is involved has yet to be made clear (30).
Contrary to what is observed in the literature (28,29,31), in this study, no increase in weight or food intake was observed in the OVX group. However, the "classical view" of estrogen action has been widely regarded as canonical dogma, as it integrates well not only the actions of E2 on the energy balance but also its effects on the hypothalamic-pituitary-gonadal (HPG) axis. A recent study (32) shows that the effect of E2 on eating depends on nutritional status.
However, there was a significant increase in weight in the ADS group. These animals’ higher food consumption could partly explain this. As weight gain occurred in parallel with food consumption in this study, demonstrating that the increase in weight induced by ovariectomy, which has been reported previously (33-35), can be attributed to food efficiency or possible changes in energy metabolism and deprivation of estrogenic activity since this was the only group that did not have estrogen hormone replacement.
The results of the hormonal profile showed that ovariectomy caused a significant decrease in the concentration of serum estradiol and progesterone in the OVX group. In contrast, the group treated only with the phytonutraceutical Ormona® showed a higher serum estradiol level (p < 0.005). Similar results were obtained in previous studies (36,37), which determined the dose-dependent pro-estrogenic profile in ovariectomized rats treated with increasing concentrations of T. pratense extract. Supplementation with T. pratense for 6 weeks increased estrogen and progesterone levels in mice. This result of estrogenic activity can be attributed to isoflavones (38).
Among the isoflavones found in the T. pratense species, four bind competitively to ERα and ERβ receptors (genistein > daidzein > biochanin A > formononetin) with affinities approximately one-thousandth of 17β-estradiol (39,40). In the phytonutraceutical Ormona®, 5.4% daidzein, 2.3% glycitein, and 3.6% genistein were identified (10), justifying the potent estrogenic activity performed by Ormona®.
In 2000, the North American Menopause Society (NAMS) published an evidence-based consensus on the role of isoflavones in menopausal health. Although the health effects observed in humans cannot be attributed to isoflavones alone, it has become clear that foods or products containing these substances have some physiological effects, mainly cardiovascular benefits. Since Ormona® phytonutraceutical is a product that combines various bioactives, the composition of this product favors its use for improving the health of premenopausal and menopausal women (41).
It was also observed that the group treated with the combination of Ormona® + estradiol showed a reduced osteogenic profile (p < 0.005). This result can be explained in part by the potential drug interaction between T. pratense and other anti-osteoporotic drugs, such as tamoxifen since studies report that preparations containing T. pratense can interfere with the steady-state mRNA levels of the main drug-metabolizing enzymes. This interference can be either by induction, as in the case of CYP3A2, or inhibition/suppression, as in the case of CYP 1A2, 2B1/2, 2C11, 2E1. Both situations can lead to changes in the metabolism of coadministered drugs (42).
Furthermore, it can be inferred that the ovariectomized rats model lacks an endogenous estrogen environment, as the higher endogenous estrogen environment can cause competition between isoflavones and estradiol for receptor binding sites, and therefore, the weaker estrogenic potency of isoflavones, as well as the different binding preferences to the α and β Estrogen Receptors (ERs) can lead to an anti-estrogenic effect. In a higher estrogen environment, isoflavones act in vitro as antiestrogens, while in a low estrogen environment, they act as estrogens, which is characteristic of a post-menopausal situation (40,43,44).
Another aspect to consider is the low serum estrogen concentration in the ORM+EST group. The competition for the same binding site between estradiol (an endogenous estrogen) and phytoestrogens (" plant-origin estrogens") is a topic discussed in the scientific literature. Some studies suggest phytoestrogens may have weaker estrogenic effects due to competition for estrogen receptors, leading to decreased serum estrogen levels. However, it is important to note that the extent of this competition and its effects on serum estrogen levels may vary according to several factors, including the amount and type of phytoestrogens consumed, the duration of exposure, and individual characteristics. In addition, studies on this topic can present conflicting results, and there needs to be a clear consensus on the effects of phytoestrogens on regulating serum estrogen levels (38,39).
In terms of biochemical parameters (Figure 3), higher levels of cholesterol (TC), LDL, and triglycerides (TG) were observed in the OVW group, while HDL was not affected. These higher levels were explained by the increase in LDL cholesterol in the OVW group and corroborate other studies that have shown hypercholesterolemia in ovariectomized rats, reporting an association between estrogen deficiency and early onset changes in hepatic cholesterol and triacylglycerol synthesis (45-47).
Women in hypoestrogenic conditions are susceptible to various lipid metabolism disorders after menopause since epidemiological and prospective studies associate estrogen with a relevant role against metabolic syndrome precisely because it has effects on many organ systems that contribute to cardiovascular protection, including the regulation of hepatic lipid metabolism and serum lipoprotein levels (48,49).
The group treated with estrogen (EST) showed lower levels of TC, LDL, and higher HDL (p < 0.01), but it was unable to prevent an increase in TG compared to the negative control group. This can be explained by the hepatic action of estradiol (E2), which regulates the rate of synthesis of structural apolipoproteins for VLDL and HDL, reduces the rate of synthesis of apoB-100 while stimulating the synthesis of apoA-I and apoA-II. The HDL fraction containing apoA-I and apoA-II is necessary for the degradation of chylomicrons and VLDL and for the direct and indirect transportation of cholesterol to the liver. Furthermore, in hepatocytes, estrogens stimulate the synthesis of apoC-III while decreasing the synthesis of hepatic lipase (LH) (49-51).
Meanwhile, both OrmonaⓇ (ORM) and Ormona in combination with estrogen (ORM+EST) reduced TC and TG levels and increased HDL levels. These effects can be explained by the composition of the phytonutraceutical OrmonaⓇ, which has phytochemical markers such as geranylgeraniol and tocotrienols from B. orellana, which can reduce the biosynthesis of cholesterol and triglycerides by negatively regulating HMG-CoA reductase (HGMR) (14,52-54). In addition, reports show that supplementation with T. pratense (the main component of OrmonaⓇ) reduced LDL and TG levels in mice. Phytoestrogens, by inhibiting 7a-hydroxylase and estrogenic effects on the LDL receptor, modulate the PCSK9 enzyme to decrease LDL (37,55).
The significant increase in aspartate aminotransferase (AST) and alanine aminotransferase (ALT) following the administration of estradiol (EST) may be associated with various factors, including direct and indirect effects of estradiol on the liver or hyperandrogenemia. During hepatic metabolization, estradiol can increase blood flow to the liver. This hyperactivation of liver metabolism can cause damage to hepatocytes, releasing AST and ALT into the bloodstream. In addition, the liver can produce toxic by-products that can damage liver cells and release AST and ALT (56).
Although the OVW group showed a higher serum alkaline phosphatase (ALP) serum, the differences were not statistically significant (Table 1). Despite this, the treated groups showed lower levels of ALP, demonstrating a modulating action on bone metabolism. Measuring biochemical markers of bone metabolism leads to understanding bone transition in advanced age. ALP is a component of the cell membrane of many body tissues, with the highest concentrations of this enzyme found in bone cells (osteoblasts) and the liver. ALP is increased in skeletal system diseases associated with increased osteoblastic activity and bone remodeling (57,58). Studies have reported increased ALP in ovariectomized rats (59-63).
The deposition of calcium salts in the process of osteogenesis is a process dependent on the nutritional state of the bone and immunological and biochemical factors (64). When assessing the total weight of the femur, the ORM group had a statistically higher bone mass (p < 0.05) compared to the OVW (Figure 5). Similarly, the ORM group also had higher Ca2+ levels than the OVW and ADS groups (p < 0.05). These results were confirmed in the SEM images, where observation of the microstructure of the femur showed the epiphysis of the cancellous bone preserved (Figure 6), indicating the osteoprotective action of OrmonaⓇ.
In the current study, histopathological evaluation revealed that treatment of ovariectomized rats with OrmonaⓇ resulted in an increase in the thickness of the femoral shaft compared to the OVW group and a decrease in the trabeculae. There was also an increase in the number of osteocytes, the presence of osteoblasts, and the absence of osteoclasts. These results align with the observations of other researchers, who have reported the protective action of T. pratense against bone alterations characteristic of osteoporosis due to hypoestrogenism (21,40,65-68).
Also present in OrmonaⓇ, geranylgeraniol (GGOH) may play an osteoprotective role, as observed in a previous study by our research group (14). Exogenous GGOH may inhibit osteoclast formation by suppressing the receptor activating NF-κB ligand expression (69). However, the mechanism of action proposed for the osteoprotective, and anabolic action of the phytoestrogens present in OrmonaⓇ is based on specific effects on osteoblasts and osteoclasts in vitro and in vivo.
The most abundant isoflavones in T. pratense, biochanin A and formononetin, are metabolized into genistein and daidzein, respectively (70,71). In vitro studies, it has been shown that the phytoestrogens daidzein (72,73), genistein (74,75), biochanin A, and formononetin (73,76,77) promote the differentiation and growth of osteoblasts and stimulate the formation and prevention of bone resorption by regulating calcium absorption, the intestinal microbiota, and various metabolic pathways. By mobilizing bone calcium into the circulation, isoflavones enhance calcium homeostasis during the calcium uptake (65,68,78).
Phytoestrogens are selective modulators of estrogen receptors (ER). Their main affinity is for the beta type (β-RE), which is more present in bone cells, especially in osteoblasts responsible for bone formation. Genistein, daidzein, biochanin A, and formononetin have been implicated in the estrogenic activity of T. pratense and have been shown to competitively bind to ERα and ERβ with similar affinities to 17-β estradiol (72,79,80).
Clinical data on the action of isoflavones shows that aglycones (most commonly found in fermented products) have beneficial effects against bone loss due to estrogen deficiency in women compared to the glycoside form of isoflavones. With this, the EFSA ratified the safety of isoflavones for women in a more comprehensive report (81).
Therefore, ascorbic acid from Myrciaria dubia extract tends to act as a catalyst, favoring the acid hydrolysis necessary to absorb these phytoestrogens. This is due to the pharmacokinetic aspect, since the absorption rates of isoflavones are favored at acidic pH, as it is necessary to break the bonds with the sugars present in the molecule, enabling the release of the aglycones present in glycosylated isoflavones, such as glycitein and daidzein (82).
Zhu et al. (82) showed that ascorbic acid prevented bone density loss in ovariectomized mice, suggesting that the nutrient may help prevent osteoporosis. The combined action of ascorbic acid and polyphenols present in M. dubia extract (approximately 1797.0 mg EAG.100g-1) provides antioxidant effects and benefits against menopausal symptoms (82).
Osteoprotegerin (OPG) and receptor activator ligand NF-j B (RANKL) are crucial in bone remodeling. The modulation of these factors has profound effects on bone formation and resorption. Interleukin 6 (IL-6) can inhibit apoptosis and directly increase the activity of osteoclasts, thus prolonging their lifespan and promoting bone loss by activating OPG/RANKL/RANK (osteoprotegerin/receptor activator of nuclear factor kappa B ligand/receptor activator of the nuclear factor kappa B) (83,84). In contrast, genistein and daidzein could suppress IL-6 release and increase OPG production dose-dependent (85).
In osteoblasts, genistein stimulates their proliferation (osteoblast-like cells MC3T3-E1) and protects against oxidative cell damage from free radicals (86), increases collagen synthesis, and prevents apoptosis in MC3T3-E1 (87). However, in vitro studies show that genistein can inhibit bone resorption by osteoclasts by inhibiting the formation and differentiation of osteoclastic cells from bone marrow cells and induces apoptosis of osteoclasts in their mature form through the mechanism of Ca 2+ channels. This suppressive effect of genistein on these bone cells is due to partial inhibition of protein tyrosine kinase and activation of protein tyrosine phosphatase (88-90).
In vivo tests with subcutaneous administration of genistein in ovariectomized rats showed an increase in the number of osteoblasts (91). Similarly, treatment with daidzein and 17β-estradiol has been shown to affect the maturation and activity of osteoclasts in vitro, providing evidence of inhibition of osteoclast differentiation by the mechanism of apoptosis of their specific precursors (92).
The formation of osteoclasts is regulated in response to the secretion of osteoclast differentiation factor (RANKL) and osteoprotegerin (OPG). RANKL is produced by osteoblasts or osteogenic stromal cells, and it is involved in the formation of osteoclast-like cells, the fusion of osteoclast precursor cells, and bone resorption. OPG inhibits osteoclastogenesis by masking RANKL in osteoblasts or osteogenic stromal cells. Phytoestrogens decreased RANKL mRNA levels and increased OPG mRNA levels through specific inhibition of topoisomerase II (topo II) and protein tyrosine kinase (93).
Therefore, this research shows that the results with the phytonutraceutical OrmonaⓇ were favorable for regulating the activities of bone cells, providing a possible way to support the regulation of menopause-related osteoporosis symptoms in women. However, simultaneous administration with synthetic estrogens requires greater attention, as possible anti-estrogenic activity has been observed. The effects of OrmonaⓇ on the skeletal system of estrogen-deficient rats differed from those exerted by estradiol, indicating a more complex mechanism of action.
5. Conclusions
Based on the biochemical, hormonal, and histopathology results, it can be suggested that the phytonutraceutical OrmonaⓇ promoted the highest serum estradiol concentration compared to the other experimental groups. The positive estrogenic effect was associated with increased bone calcium concentration and preservation of bone microarchitecture, which was also observed in the group treated with estrogen and sodium alendronate. However, only treatment with OrmonaⓇ improved the lipid profile (cholesterol, triglycerides), especially by increasing HDL.
6. Patents
A patent from this work has been submitted to the National Institute of Industrial Property (INPI) under the number BR102022008408-4.
Author Contributions
Concept, Methodology J.C.T.C., A.Ld.N.; G.d.C.F.; H.d.O.C.; H.R.d.S.; F.S.B.; K.C.d.M.; A.O.N.B and R.M.B. Validation, A.L.d.N.; P.F.S. Formal analysis, A,L.d.N and H.d.O.C. Writing - preparation of original manuscript and discussion of results A.L.d.N.; G.d.C. F. and H.R.d.S. Writing - review and editing, J.C.T.C. Supervision, J.C.T.C. Project management, J.C.T.C. Funding acquisition, J.C.T.C. Translation, V.MV. All authors have read and agreed with the published version of the manuscript.
Funding
This work was supported in part by National Council for Scientific and Technological Development – CNPq No. 12/2020 and CNPq No. 068/2022 – Proc.: 403587/2020-4 and 421808/2022-5, Master’s and Doctoral Program for Innovation - MAI/DAI.
Institutional Review Board Statement
The authors followed the guidelines for animal experimentation established by the National Council for Animal Experimentation of Brazil (CONCEA). The Ethics Committee on the Use of Animals (CEUA) of the Federal University of Amapá-UNIFAP approved the project under protocol number 017/2022.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Silva, A.C.V.; Rosa, M.I.; Fernandes, B.; Lumertz, S.; Diniz, R.M.; Damiani ME dos, R. Fatores associados à osteopenia e osteoporose em mulheres submetidas à densitometria óssea. Rev Bras Reumatol, 2015, 55:223–8. [CrossRef]
- Radominski, S.C.; Bernardo, W.; Patrícia De Paula, A.; Albergaria, B.H.; Moreira, C.; Fernandes, C.E.; et al. Brazilian guidelines for the diagnosis and treatment of post-menopausal osteoporosis. Revista brasileira de reumatologia, 2017. [CrossRef]
- Costa, J.R.G da, Tavares A.L de F, Bertolini G.R.F, Costa R.M, Ribeiro L. de F.C. Efeito da reposição fitoterápica com isoflavonas na histomorfometria do tecido ósseo de ratas Wistar ooforectomizadas. Biosaúde. (2021).
- Deli, T.; Orosz, M.; Jakab, A. Hormone Replacement Therapy in Cancer Survivors – Review of the Literature. Pathology & Oncology Research 2019, 26, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y., Ma L., Yang X., Bie J., Li D., Sun C., et al. Menopausal Hormone Replacement Therapy and the Risk of Ovarian Cancer: A Meta-Analysis. Front Endocrinol (Lausanne). 3 de dezembro de 2019, 10:486180. [CrossRef]
- Frigo, M., Barros E. de, Santos P.C. de B. dos, Koehnlein E.A. Isoflavonas como tratamento alternativo na sintomatologia climatérica: uma revisão sistemática. Rev Inst Adolfo Lutz, 2021. [CrossRef]
- Viggiani, M.T.; Polimeno, L.; Di Leo, A.; Barone, M. Phytoestrogens: Dietary Intake, Bioavailability, and Protective Mechanisms against Colorectal Neoproliferative Lesions. Nutrients. 24 de julho de 2019, 11, 1709. [Google Scholar] [CrossRef] [PubMed]
- Aronson, J.K. Defining ’nutraceuticals’: neither nutritious nor pharmaceutical. British journal of clinical pharmacology. 2017, 83, 8–19. [Google Scholar] [CrossRef]
- Rocha, C.F, Flexa C de N.N, de Souza G.C, Pereira A.C.M, Carvalho H. de O., Nascimento A.L, et al. Acute and Reproductive Toxicity Evaluation of Ormona® SI and Ormona® RC-Two New Nutraceuticals with Geranylgeraniol, Tocotrienols, Anthocyanins, and Isoflavones-In Adult Zebrafish. Pharmaceuticals (Basel), 2022 15(11). [CrossRef]
- Rodrigues A.P.S, da Silva B.R, Pereira A.C.M, Batista M.A, Sales P.F, Ferreira A.M, et al. Ormona® SI and Ormona® RC—New Nutraceuticals with Geranylgeraniol, Tocotrienols, Anthocyanins, and Isoflavones—Decrease High-Fat Diet-Induced Dyslipidemia in Wistar Rats. Nutraceuticals, 2022, 2, 311–22. [CrossRef]
- Tsuang Y.H, Chen L.T, Chiang C.J, Wu L.C, Chiang Y.F, Chen P.Y, et al. Isoflavones prevent bone loss following ovariectomy in young adult rats. J Orthop Surg Res, 2008, 3, 1–9. [CrossRef]
- Yousefzadeh, N.; Kashfi, K.; Jeddi, S.; Ghasemi, A. Ovariectomized rat model of osteoporosis: a practical guide. EXCLI J. 10 de janeiro de 2020, 19:89. [CrossRef]
- Oliveira Carvalho H., Farias e Souza B.S, dos Santos I.V.F, Resque R.L, Keita H, Fernandes C.P, et al. Hypoglycemic effect of formulation containing hydroethanolic extract of Calophyllum brasiliense in diabetic rats induced by streptozotocin. Revista Brasileira de Farmacognosia. setembro de 2016, 26, 634–9. [CrossRef]
- Pereira A.C.M, de Oliveira Carvalho H., Gonçalves D.E.S., Picanço K.R.T, de Lima Teixeira dos Santos A.V.T, da Silva H.R, et al. Co-Treatment of Purified Annatto Oil (Bixa orellana L.) and Its Granules (Chronic®) Improves the Blood Lipid Profile and Bone Protective Effects of Testosterone in the Orchiectomy-Induced Osteoporosis in Wistar Rats. Molecules. 4 de agosto de 2021, 26, 4720. [CrossRef]
- Palma, M.N.N.; Rocha, G.C.; Filho, S.C.V.; Detmann, E. Evaluation of Acid Digestion Procedures to Estimate Mineral Contents in Materials from Animal Trials. Asian-Australas J Anim Sci. 4 de agosto de 2015, 28, 1624–8. [Google Scholar] [PubMed]
- Hou, T.; Zhang, L.; Yang, X. Ferulic acid, a natural polyphenol, protects against osteoporosis by activating SIRT1 and NF-κB in neonatal rats with glucocorticoid-induced osteoporosis. Biomedicine & Pharmacotherapy. 1o de dezembro de 2019, 120:109205. [CrossRef]
- Turner, A.S. Animal models of osteoporosis - necessity and limitations. European Cells & Materials. 22 de junho de 2001, 1:66–81. [CrossRef]
- Yamagishi T., Otsuka E., Hagiwara H. Reciprocal Control of Expression of mRNAs for Osteoclast Differentiation Factor and OPG in Osteogenic Stromal Cells by Genistein: Evidence for the Involvement of Topoisomerase II in Osteoclastogenesis. Endocrinology. 1o de agosto de 2001, 142, 3632–7. [CrossRef]
- Thompson D.D., Simmons H.A., Pirie C.M., Ke H.Z. FDA guidelines and animal models for osteoporosis. Bone. Outubro de 1995, 17, S125–33. [CrossRef]
- Silveira, V.A.S. Efeitos iniciais da ovariectomia e do tratamento com estrógeno e isoflavonas da soja, isolados e associados, na reparação óssea alveolar e no útero de ratas [tese]. Faculdade de Odontologia de São José dos Campos, UNESP. 10 de julho de 2007.
- Sharaf, H.A.; Shaffie, N.M.; Morsy, F.A.; Badawi, M.A.; Abbas, N.F. Role of some phytoestrogens in recovering bone loss: histological results from experimental ovariectomized rat models. Journal of The Arab Society for Medical Research. 2015, 10, 65. [Google Scholar] [CrossRef]
- Kalleny, N.K. Histological and morphometric studies on the effect of alpha-lipoic acid on post ovariectomy osteoporosis induced in adult female albino rats. The Egyptian Journal of Histology. Março de 2011, 34, 139–55. [Google Scholar] [CrossRef]
- Park, J.A., Ha S.K., Kang T.H., Oh M.S., Cho M.H., Lee S.Y., et al. Protective effect of apigenin on ovariectomy-induced bone loss in rats. Life Sci. 20 de junho de 2008, 82(25–26):1217–23. [CrossRef]
- Weber K., Kaschig C., Erben R.G. 1α-Hydroxyvitamin D2 and 1α-hydroxyvitamin D3 have anabolic effects on cortical bone, but induce intracortical remodeling at toxic doses in ovariectomized rats. Bone. Setembro de 2004, 35, 704–10. [CrossRef]
- Barlet J.P., Picherit C., Coxam V., Bennetau-Pelissero C., Lebecque P., Kati-Coulibaly S., et al. Daidzein Is More Efficient than Genistein in Preventing Ovariectomy-Induced Bone Loss in Rats. J Nutr. Julho de 2000, 130, 1675–81. [CrossRef]
- Fanti P., Monier-Faugere M.C., Geng Z., Schmidt J., Morris P.E., Cohen D., et al. The phytoestrogen genistein reduces bone loss in short-term ovariectomized rats. Osteoporosis International. 1998, 8, 274-81. [CrossRef]
- López, M.; Tena-Sempere, M. Estrogens and the control of energy homeostasis: a brain perspective. Trends in Endocrinology & Metabolism. 1o de agosto de 2015, 26, 411–21. [Google Scholar] [CrossRef]
- Martínez de Morentin, P.B.; González-García, I.; Martins, L.; Lage, R.; Fernández-Mallo, D.; Martínez-Sánchez, N.; et al. Estradiol Regulates Brown Adipose Tissue Thermogenesis via Hypothalamic, A. M.P.K. Cell Metab. 1º de julho de 2014, 20, 41–53. [Google Scholar] [CrossRef]
- Xu, Y.; López, M. Central regulation of energy metabolism by estrogens. Mol Metab. 23 de maio de 2018, 15:104–15. [CrossRef]
- Hirschberg, A.L. Sex hormones, appetite and eating behaviour in women. Maturitas. Março de 2012, 71, 248–56. [Google Scholar] [CrossRef]
- Liu Z.P., Li W.X., Yu B., Huang .J, Sun J., Huo J.S., et al. Effects of trans-Resveratrol from Polygonum cuspidatum on Bone Loss Using the Ovariectomized Rat Model. J Med Food. 25 de março de 2005, 8, 14–9. [CrossRef]
- Yu, K., He Y., Hyseni I., Pei Z., Yang Y., Xu P., et al. 17β-estradiol promotes acute refeeding in hungry mice via membrane-initiated ERα signaling. Mol Metab. Dezembro de 2020, 42:101053. [CrossRef]
- Li L., Yang Y., Ma C., Li X., Bian X., Fu Y., et al. Effects of soybean isoflavone aglycone on osteoporosis in ovariectomized rats. Front Nutr. 5 de junho de 2023, 10. [CrossRef]
- Xu, H., Liu T., Hu L., Li J., Gan C., Xu J., et al. Effect of caffeine on ovariectomy-induced osteoporosis in rats. Biomedicine & Pharmacotherapy. Abril de 2019, 112:108650. [CrossRef]
- Zhang, Q., Song X., Chen X., Jiang R., Peng K., Tang X., et al. Antiosteoporotic effect of hesperidin against ovariectomy-induced osteoporosis in rats via reduction of oxidative stress and inflammation. J Biochem Mol Toxicol. 24 de agosto de 2021, 35(8). [CrossRef]
- Khattab, H.A.H.; Ardawi, M.S.; Ateeq, R.A.M. Effect of Phytoestrogens Derived from Red Clover (Trifolium Pratense L.) in Ovariectomized Rats. Life Science Journal. Janeiro de 2013, 10, 1485–1497. [Google Scholar]
- Chen, Y.M.; Wang, I.L.; Zhu, X.Y.; Chiu, W.C.; Chiu, Y.S. Red Clover Isoflavones Influence Estradiol Concentration, Exercise Performance, and Gut Microbiota in Female Mice. Front Nutr. 14 de abril de 2021, 8: 623698. [CrossRef]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: estrogenic activity, biological effect and bioavailability. Eur J Drug Metab Pharmacokinet. 17 de março de 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Liu J., Burdette J.E., Xu H., Gu C., van Breemen R.B., Bhat K.P.L., et al. Evaluation of Estrogenic Activity of Plant Extracts for the Potential Treatment of Menopausal Symptoms. J Agric Food Chem. 1o de maio de 2001, 49, 2472–9. [CrossRef]
- Burdette J.E., Liu J., Lantvit D., Lim E., Booth N., Bhat K.P.L., et al. Trifolium pratense (Red Clover) Exhibits Estrogenic Effects In Vivo in Ovariectomized Sprague-Dawley Rats. J Nutr. Janeiro de 2002, 132, 27–30. [CrossRef]
- Greenwood S., Barnes S., Clarkson T.B., Eden J., Helferich W.G., Hughes C., et al. The role of isoflavones in menopausal health: Consensus opinion of the North American Menopause Society. Menopause: The Journal of the North American Menopause Society. 27 de março de 2000, 7, 215–29. [CrossRef]
- Tripathi A., Singh S., Raju K.S.R., Wahajuddin, Gayen J. Effect of Red Clover on CYP Expression: An Investigation of Herb-Drug Interaction at Molecular Level. Indian Journal of Pharmaceutical Sciences. 2014, 76, 261. [CrossRef]
- Hwang, C.S., Kwak H.S., Lim H.J., Lee S.H., Kang Y.S., Choe T.B., et al. Isoflavone metabolites and their in vitro dual functions: They can act as an estrogenic agonist or antagonist depending on the estrogen concentration. J Steroid Biochem Mol Biol. Novembro de 2006, 101(4–5): 246–53. [CrossRef]
- Stubert, J.; Gerber, B. Isoflavones – Mechanism of Action and Impact on Breast Cancer Risk. Breast Care. 20 de fevereiro de 2009, 4, 22–9. [Google Scholar] [CrossRef]
- Medina-Contreras, J.; Villalobos-Molina, R.; Zarain-Herzberg, A.; Balderas-Villalobos, J. Ovariectomized rodents as a menopausal metabolic syndrome model. A minireview. Mol Cell Biochem. 27 de dezembro de 2020, 475(1–2): 261–76. [CrossRef]
- Nigro M., Santos A.T., Barthem C.S., Louzada R.A.N., Fortunato R.S., Ketzer L.A., et al. A Change in Liver Metabolism but Not in Brown Adipose Tissue Thermogenesis Is an Early Event in Ovariectomy-Induced Obesity in Rats. Endocrinology. 1o de agosto de 2014, 155, 2881–91. [CrossRef]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased adipose tissue in male and female estrogen receptor-α knockout mice. Proceedings of the National Academy of Sciences. 7 de novembro de 2000, 97, 12729–34. [Google Scholar] [CrossRef] [PubMed]
- Iorga, A.; Cunningham, C.M.; Moazeni, S.; Ruffenach, G.; Umar, S.; Eghbali, M. The protective role of estrogen and estrogen receptors in cardiovascular disease and the controversial use of estrogen therapy. Biol Sex Differ. 24 de dezembro de 2017, 8, 33. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. Adv Exp Med Biol. 10 de dezembro de 2017, 1043: 227–56. [CrossRef]
- Berman, D.M.; Nicklas, B.J.; Ryan, A.S.; Rogus, E.M.; Dennis, K.E.; Goldberg, A.P. Regulation of Lipolysis and Lipoprotein Lipase after Weight Loss in Obese, Post-menopausal Women. Obes Res. Janeiro de 2004, 12, 32–9. [Google Scholar] [CrossRef]
- Faulds M.H., Zhao C., Dahlman-Wright K/, Gustafsson J.Å. The diversity of sex steroid action: regulation of metabolism by estrogen signaling. Journal of Endocrinology. Janeiro de 2012, 212, 3–12. [CrossRef]
- Pearce, B.C.; Parker, R.A.; Deason, M.E.; Qureshi, A.A.; Wright, J.J.K. Hypocholesterolemic activity of synthetic and natural tocotrienols. J Med Chem. 1o de outubro de 1992, 35, 3595–606. [Google Scholar] [CrossRef]
- Sever, N.; Song, B.L.; Yabe, D.; Goldstein, J.L.; Brown, M.S.; DeBose-Boyd, R.A. Insig-dependent Ubiquitination and Degradation of Mammalian 3-Hydroxy-3-methylglutaryl-CoA Reductase Stimulated by Sterols and Geranylgeraniol. Journal of Biological Chemistry. Dezembro de 2003, 278, 52479–90. [Google Scholar] [CrossRef]
- Qureshi, A.; Khan, D.; Mahjabeen, W.; Qureshi, N. Dose-dependent Modulation of Lipid Parameters, Cytokines and RNA by δ-tocotrienol in Hypercholesterolemic Subjects Restricted to AHA Step-1 Diet. Br J Med Med Res. 10 de janeiro de 2015, 6, 351–66. [Google Scholar] [CrossRef]
- Luís, Â.; Domingues, F.; Pereira, L. Effects of red clover on perimenopausal and post-menopausal women’s blood lipid profile: A meta-analysis. Climacteric. 3 de setembro de 2018, 21, 446–53. [Google Scholar] [CrossRef]
- Chen, M.J.; Chiu, H.M.; Chen, C.L.; Yang, W.S.; Yang, Y.S.; Ho, H.N. Hyperandrogenemia Is Independently Associated with Elevated Alanine Aminotransferase Activity in Young Women with Polycystic Ovary Syndrome. J Clin Endocrinol Metab. 1o de julho de 2010, 95, 3332–41. [Google Scholar] [CrossRef]
- Grigoryan A.V., Dimitrova A.A., Kostov K.G., Russeva A.L., Atanasova M.A., Blagev A.B., et al. Changes of Serum Concentrations of Alkaline Phosphatase and Metalloproteinase-9 in an Ovariectomized Wistar Rat Model of Osteoporosis. Journal of Biomedical and Clinical Research. 1o de novembro de 2017, 10, 32–6. [CrossRef]
- Kim, S.K.; Lee, M.H.; Rhee, M.H. Studies on the Effects of Biomedicinal Agents on Serum Concentration of Ca2+, P and ALP Activity in Osteoporosis-Induced Rats. J Vet Sci. Agosto de 2003, 4, 151–4. [Google Scholar] [CrossRef]
- Devareddy, L.; Hooshmand, S.; Collins, J.K.; Lucas, E.A.; Chai, S.C.; Arjmandi, B.H. Blueberry prevents bone loss in ovariectomized rat model of post-menopausal osteoporosis. J Nutr Biochem. Outubro de 2008, 19, 694–9. [Google Scholar] [CrossRef]
- Zhang, R.; Hu, S.J.; Li, C.; Zhang, F.; Gan, H.Q.; Mei, Q.B. Achyranthes bidentata root extract prevent OVX-induced osteoporosis in rats. J Ethnopharmacol. Janeiro de 2012, 139, 12–8. [Google Scholar] [CrossRef]
- Chang K.L., Hu Y.C., Hsieh B.S., Cheng H.L., Hsu H.W., Huang L.W., et al. Combined effect of soy isoflavones and vitamin D3 on bone loss in ovariectomized rats. Nutrition. Janeiro de 2013, 29, 250–7. [CrossRef]
- Hooshmand, S.; Juma, S.; Arjmandi, B.H. Combination of Genistin and Fructooligosaccharides Prevents Bone Loss in Ovarian Hormone Deficiency. J Med Food. Abril de 2010, 13, 320–5. [Google Scholar] [CrossRef]
- Gao, Y.H.; Yamaguchi, M. Suppressive Effect of Genistein on Rat Bone Osteoclasts: Apoptosis Is Induced through Ca2+ Signaling. Biol Pharm Bull. 15 de agosto de 1999, 22, 805–9. [Google Scholar] [CrossRef]
- Orsatti, F.L.; Nahas, E.P.; Nahas-Neto, J.; Orsatti, C.L.; Teixeira, A.S. Efeito do treinamento contrarresistência e isoflavona na densidade mineral óssea em mulheres na pós-menopausa. Revista Brasileira de Cineantropometria e Desempenho Humano. 27 de agosto de 2013, 15(6). [CrossRef]
- Cegieła, U.; Folwarczna, J.; Pytlik, M.; Zgórka, G. Effects of Extracts from Trifolium medium L. and Trifolium pratense L. on Development of Estrogen Deficiency-Induced Osteoporosis in Rats. Evidence-Based Complementary and Alternative Medicine. 29 de novembro de 2012, 2012: 1–11. [CrossRef]
- AL-Ghaban, N.; Jasem, G. Histomorphometric evaluation of the effects of local application of red cloveroil (trifolium pratense) on bone healing in rats. Journal of Baghdad College of Dentistry. 15 de junho de 2020, 32, 26–31. [Google Scholar] [CrossRef]
- Occhiuto F., De Pasquale R., Guglielmo G., Palumbo D.R., Zangla G., Samperi S., et al. Effects of phytoestrogenic isoflavones from red clover (Trifolium pratense L.) on experimental osteoporosis. Phytotherapy Research. Fevereiro de 2007, 21, 130–4. [CrossRef]
- Gao, Y.H.; Yamaguchi, M. Suppressive effect of genistein on rat bone osteoclasts: involvement of protein kinase inhibition and protein tyrosine phosphatase activation. Int J Mol Med. 1o de março de 2000, 5, 261–7. [Google Scholar] [CrossRef]
- Hiruma, Y.; Nakahama, K.; Fujita, H.; Morita, I. Vitamin K2 and geranylgeraniol, its side chain component, inhibited osteoclast formation in a different manner. Biochem Biophys Res Commun. Janeiro de 2004, 314, 24–30. [Google Scholar] [CrossRef]
- Andres, S.; Hansen, U.; Niemann, B.; Palavinskas, R.; Lampen, A. Determination of the isoflavone composition and estrogenic activity of commercial dietary supplements based on soy or red clover. Food Funct. Junho de 2015, 6, 2017–25. [Google Scholar] [CrossRef]
- Pfitscher, A.; Reiter, E.; Jungbauer, A. Receptor binding and transactivation activities of red clover isoflavones and their metabolites. J Steroid Biochem Mol Biol. Novembro de 2008, 112(1–3): 87–94. [CrossRef]
- Mathey J., Mardon J., Fokialakis N., Puel C., Kati-Coulibaly S., Mitakou S., et al. Modulation of soy isoflavones bioavailability and subsequent effects on bone health in ovariectomized rats: the case for equol. Osteoporosis International. 29 de março de 2007, 18, 671–9. [CrossRef]
- Somjen, D.; Katzburg, S.; Kohen, F.; Gayer, B.; Livne, E. Daidzein but not other phytoestrogens preserves bone architecture in ovariectomized female rats in vivo. J Cell Biochem. 15 de abril de 2008, 103, 1826–32. [Google Scholar] [CrossRef]
- Sehmisch S., Uffenorde J., Maehlmeyer S., Tezval M., Jarry H., Stuermer K.M., et al. Evaluation of bone quality and quantity in osteoporotic mice – The effects of genistein and equol. Phytomedicine. Maio de 2010, 17, 424–30. [CrossRef]
- Anderson, J.J.B.; Ambrose, W.W.; Garner, S.C. Biphasic Effects of Genistein on Bone Tissue in the Ovariectomized, Lactating Rat Model. Proc Soc Exp Biol Med. 1o de março de 1998, 217, 345–50. [Google Scholar] [CrossRef]
- Gautam A.K., Bhargavan B., Tyagi A.M., Srivastava K., Yadav D.K., Kumar M., et al. Differential effects of formononetin and cladrin on osteoblast function, peak bone mass achievement and bioavailability in rats. J Nutr Biochem. Abril de 2011, 22, 318–27. [CrossRef]
- Ha H., Lee H.Y., Lee J.H., Jung D., Choi J., Song K.Y., et al. Formononetin prevents ovariectomy-induced bone loss in rats. Arch Pharm Res. 27 de abril de 2010, 33, 625–32. [CrossRef]
- Harahap, I.A.; Suliburska, J. An overview of dietary isoflavones on bone health: The association between calcium bioavailability and gut microbiota modulation. Materials Today Proceedings. Abril de 2022, 63: S368–72. [CrossRef]
- De Souza, A.P., Gonsalves I.F., Schneider M.J.f., Giannini N.M., Kuroiwa V.Y., Quinones E.M., et al. Uso de Isoflavonas em Casos de Osteoporose nas Mulheres Uma Revisão Bibliográfica. Revista Higei@ - Revista Científica de Saúde. 19 de julho de 2022, 4(7).
- De Franciscis P., Colacurci N., Riemma G., Conte A., Pittana E., Guida M., et al. A Nutraceutical Approach to Menopausal Complaints. Medicina (Kaunas). 28 de agosto de 2019, 55, 544. [CrossRef]
- Lambert, M.N.T.; Jeppesen, P.B. Isoflavones and bone health in perimenopausal and post-menopausal women. Curr Opin Clin Nutr Metab Care. Novembro de 2018, 21, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Zhu L.L., Cao J., Sun M., Yuen T., Zhou R., Li J., et al. Vitamin C Prevents Hypogonadal Bone Loss. PLoS One. 8 de outubro de 2012, 7, e47058. [CrossRef]
- Shi, X.; Jiang, J.; Hong, R.; Xu, F.; Dai, S. Circulating IGFBP-3 and Interleukin 6 as Predictors of Osteoporosis in Post-menopausal Women: A Cross-Sectional Study. Mediators Inflamm. 31 de março de 2023, 2023: 1–6. [CrossRef]
- Kong Y.Y., Yoshida H., Sarosi I., Tan H.L., Timms E., Capparelli C., et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature. Janeiro de 1999, 397, 315–23. [CrossRef]
- Chen, X.W.; Garner, S.C.; Anderson, J.J.B. Isoflavones regulate interleukin-6 and osteoprotegerin synthesis during osteoblast cell differentiation via an estrogen-receptor-dependent pathway. Biochem Biophys Res Commun. 12 de julho de 2002, 295, 417–22. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Chen, X.W.; Anderson, J.J.B. Physiological concentrations of genistein stimulate the proliferation and protect against free radical-induced oxidative damage of MC3T3-E1 osteoblast-like cells. Nutrition Research. Setembro de 2001, 21, 1287–98. [Google Scholar] [CrossRef]
- Choi, E.M.; Suh, K.S.; Kim, Y.S.; Choue, R.W.; Koo, S.J. Soybean ethanol extract increases the function of osteoblastic MC3T3-E1 cells. Phytochemistry. Abril de 2001, 56, 733–9. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Gao, Y.H. Anabolic effect of genistein on bone metabolism in the femoral-metaphyseal tissues of elderly rats is inhibited by the antiestrogen tamoxifen. Research in Experimental Medicine. 24 de março de 1997, 197, 101–7. [Google Scholar] [CrossRef]
Figure 1.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on the clinical parameters of ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference between groups was a p < 0.001, b p < 0.01, and c p < 0.05. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test.
Figure 1.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on the clinical parameters of ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference between groups was a p < 0.001, b p < 0.01, and c p < 0.05. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test.
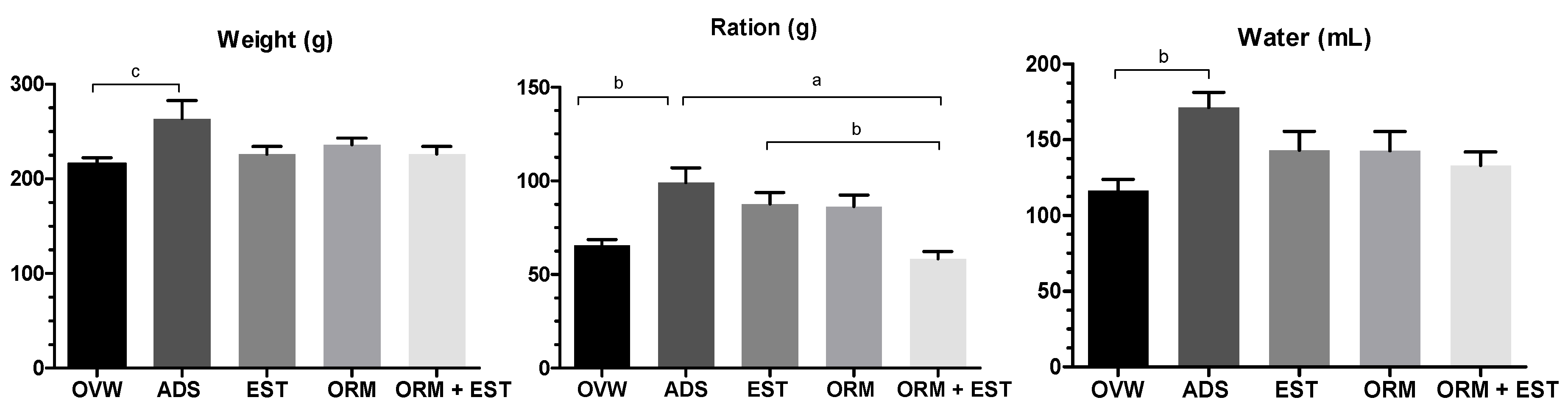
Figure 2.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on hormonal parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference b p < 0.01 and c p < 0.05 between groups. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test. The ORM and ORM+ EST groups were compared using the Student’s t-test.
Figure 2.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on hormonal parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference b p < 0.01 and c p < 0.05 between groups. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test. The ORM and ORM+ EST groups were compared using the Student’s t-test.

Figure 3.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on lipid profile parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference between groups was a p < 0.001, b p < 0.01, and c p < 0.05. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test. The ORM and ORM+ EST groups were compared using the Student’s t-test.
Figure 3.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on lipid profile parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference between groups was a p < 0.001, b p < 0.01, and c p < 0.05. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test. The ORM and ORM+ EST groups were compared using the Student’s t-test.
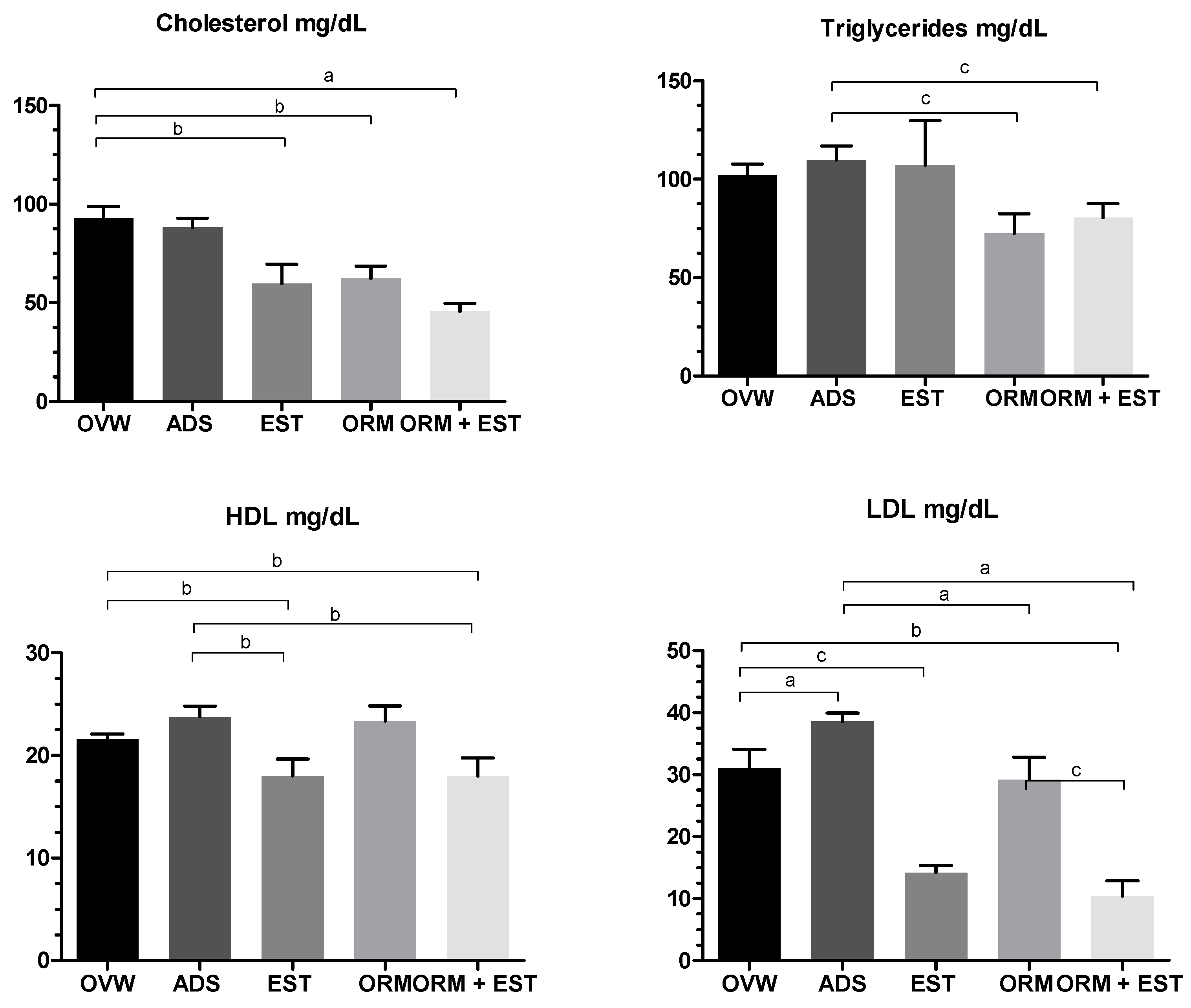
Figure 4.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on bone parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference b p < 0.01 and c p < 0.05 between groups. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test.
Figure 4.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on bone parameters in ovariectomized rats. The columns represent the mean ± standard deviation. Statistical difference b p < 0.01 and c p < 0.05 between groups. One-Way ANOVA test followed by Tukey Multiple Comparisons post-test.
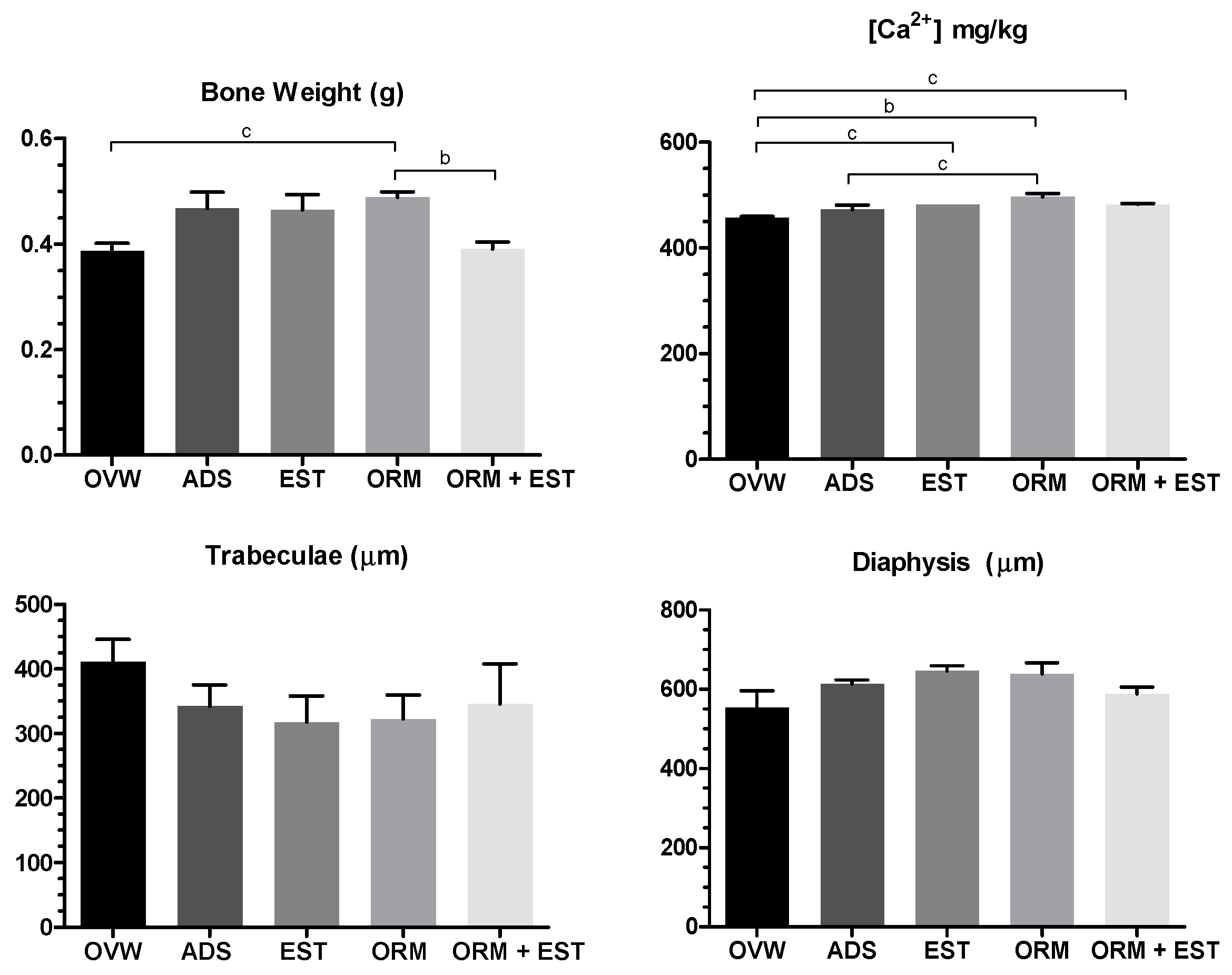
Figure 5.
Representative scanning electron microscopy (SEM) images of cross-sections of the femoral epiphysis with the trabeculae after 60 days of treatment with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D. ORM (20 mg/kg); E: ORM + EST (20 mg/kg + 2 µg/kg): ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg). Photographs were taken of the same area (on a scale of 1 mm to 2 mm) between the groups.
Figure 5.
Representative scanning electron microscopy (SEM) images of cross-sections of the femoral epiphysis with the trabeculae after 60 days of treatment with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D. ORM (20 mg/kg); E: ORM + EST (20 mg/kg + 2 µg/kg): ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg). Photographs were taken of the same area (on a scale of 1 mm to 2 mm) between the groups.
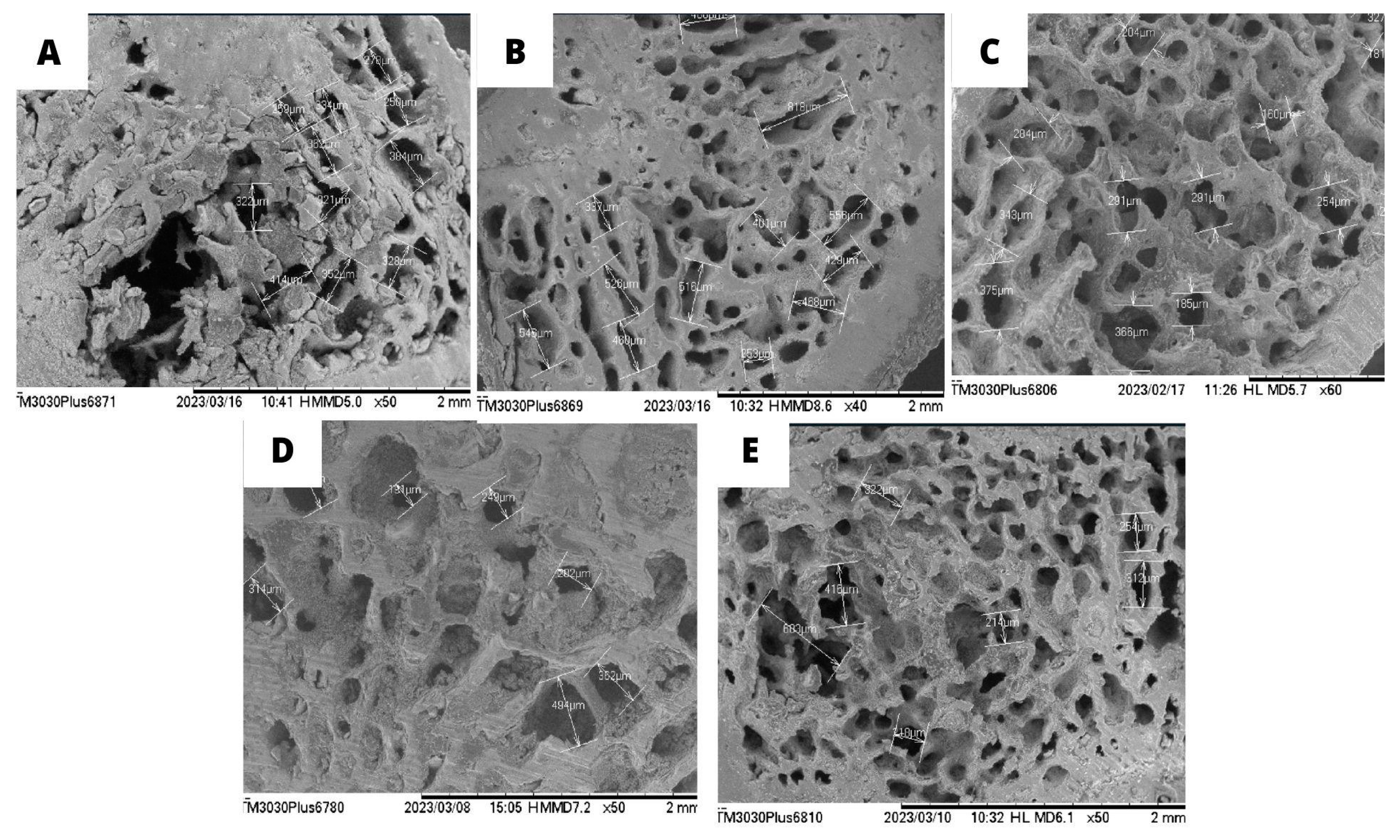
Figure 6.
Effect of oral treatments with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D: ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg) on the optical microscopy parameters of bone tissues from ovariectomized rats. Cross-section of the distal femoral epiphysis. (A) OVW, (B) ADS, (C) EST, (D) ORM, (E) ORM+EST. H.E. 10x magnification.
Figure 6.
Effect of oral treatments with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D: ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg) on the optical microscopy parameters of bone tissues from ovariectomized rats. Cross-section of the distal femoral epiphysis. (A) OVW, (B) ADS, (C) EST, (D) ORM, (E) ORM+EST. H.E. 10x magnification.
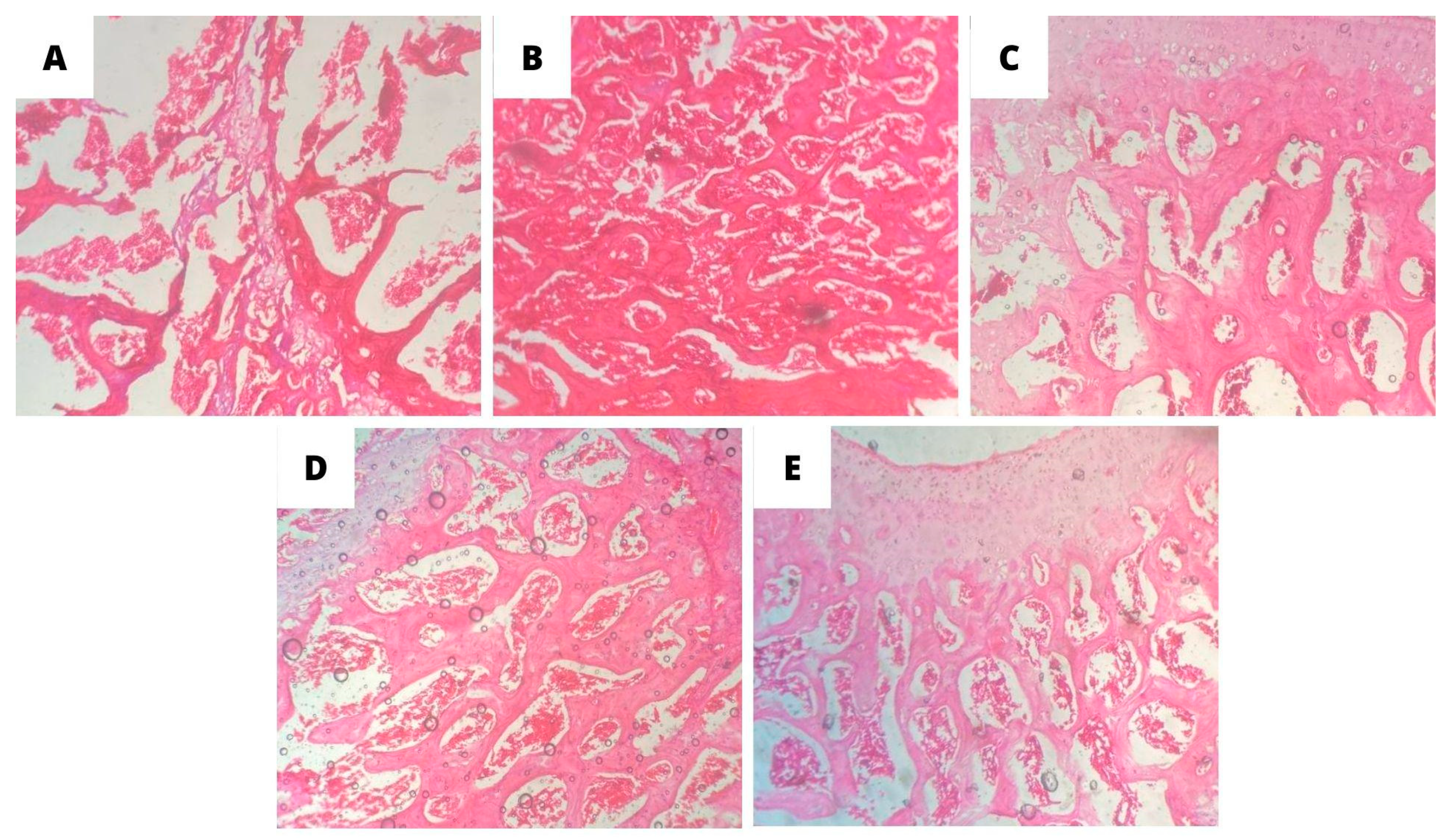
Figure 7.
Effect of oral treatments with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D: ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg) on the optical microscopy parameters of bone tissues from ovariectomized rats. Cross-section of the distal femoral epiphysis groups (A) OVW, (B) ADS, (C) EST, (D) ORM, (E) ORM+EST. H.E. 40x magnification. ot: osteoclasts; ob: osteoblasts.
Figure 7.
Effect of oral treatments with A: OVW (1 mL/kg); B: ADS (4 mg/kg); C: EST (2µg/kg), D: ORM (20 mg/kg): E: ORM + EST (20 mg/kg + 2 µg/kg) on the optical microscopy parameters of bone tissues from ovariectomized rats. Cross-section of the distal femoral epiphysis groups (A) OVW, (B) ADS, (C) EST, (D) ORM, (E) ORM+EST. H.E. 40x magnification. ot: osteoclasts; ob: osteoblasts.
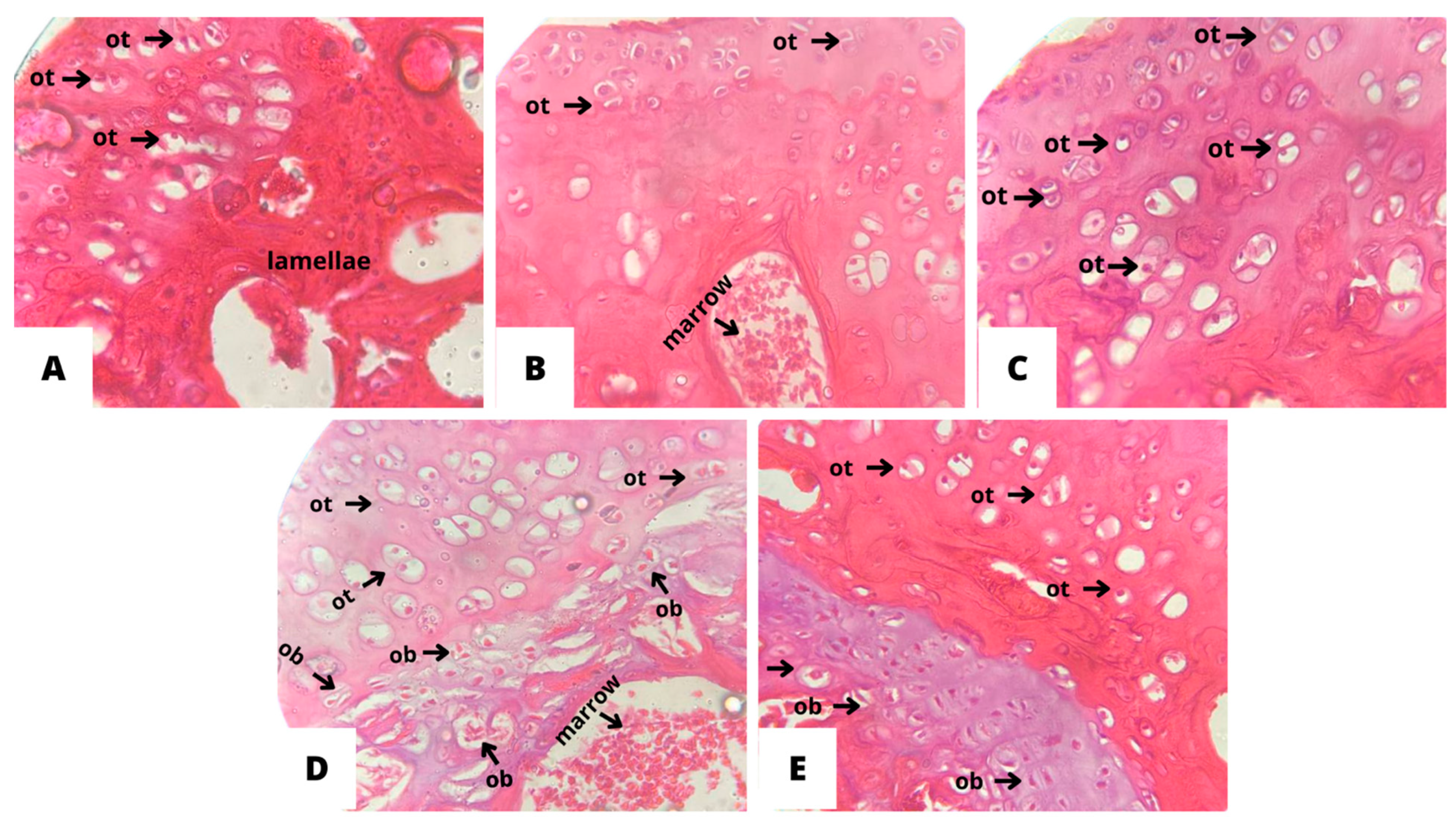

Table 1.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on biochemical parameters. Results are presented as mean ± standard deviation. *** p < 0.0001, **p < 0.01, and * p < 0.05 represent statistically significant results compared to the OVW group. #p < 0.05 represents statistically significant results compared to the ADS group one-way ANOVA test followed by Tukey Multiple Comparisons post-test.
Table 1.
Effect of oral treatments with OVW (1 mL/kg), ADS (4 mg/kg), EST (2µg/kg), ORM (20 mg/kg), and ORM + EST (20 mg/kg + 2 µg/kg) on biochemical parameters. Results are presented as mean ± standard deviation. *** p < 0.0001, **p < 0.01, and * p < 0.05 represent statistically significant results compared to the OVW group. #p < 0.05 represents statistically significant results compared to the ADS group one-way ANOVA test followed by Tukey Multiple Comparisons post-test.
| Parameters | Urea (mg/dL) | Creatinine | AST (U/dL) | ALT (U/dL) | Alkaline Phosphatase (U/L) |
|---|---|---|---|---|---|
| OVW | 29,40 ± 4,10 | 0,42 ± 0,16 | 144,40 ± 24,48 | 20,40 ± 2,07 | 55,00 ± 12,12 |
| ADS | 22,20 ± 6,42 | 0,38 ± 0,08 | 156,60 ± 33,95 | 37,60 ± 2,61*** | 50,80 ± 15,47 |
| EST | 31,40 ± 2,88 | 0,46 ± 0,15 | 276,60 ± 102,10*# | 33,80 ± 7,22** | 44,60 ± 8,53 |
| ORM | 25,40 ± 5,40 | 0,38 ± 0,04 | 201,30 ± 49,27 | 33,00 ± 3,74** | 51,40 ± 19,26 |
| ORM + EST | 26,40 ± 7,27 | 0,48± 0,05 | 156,80 ± 48,02 | 28,80 ± 7,56 | 51,60 ± 4,77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



