
Submitted:
29 November 2023
Posted:
29 November 2023
You are already at the latest version
Abstract
In small animal models of severe acute respiratory syndrome coronaviruses (SARS-CoV and SARS-CoV-2) infection, ferrets (Mustela putorius furo) have been used to investigate the pathogenesis. Podoplanin (PDPN) is an essential marker in lung type I alveolar epithelial cells, kidney podocytes, and lymphatic endothelial cells. Monoclonal antibodies (mAbs) against ferret PDPN (ferPDPN) are useful for the pathological analyses of those tissues. We previously established an anti-ferPDPN mAb, PMab-292 using the Cell-Based Immunization and Screening (CBIS) method. In this study, we determined the critical epitope of PMab-292 using flow cytometry. The N-terminal ferPDPN deletion mutants analysis revealed that the Val34 is located at the N-terminus of the PMab-292 epitope. Furthermore, the PA tag-substituted analysis (PA scanning) showed that Asp39 is located at the C-terminus of PMab-292 epitope. The epitope sequence (34-VRPEDD-39) also exists between Val26 and Asp31 of ferPDPN, indicating that PMab-292recognizes the tandem repeat of the VRPEDD sequence of ferPDPN.
Keywords:
ferret podoplanin
; monoclonal antibody
; epitope
; PA scanning
1. Introduction
Ferrets (Mustela putorius furo) have been used as an animal model for investigating the transmission and pathogenicity of human severe acute respiratory syndrome coronaviruses (SARS-CoV [1] and SARS-CoV-2 [2]). In SARS-CoV-2-infected ferrets, inflammation within alveolar spaces and perivascular mononuclear parts were observed. Furthermore, SARS-CoV-2-infected ferrets exhibited mild alveolar or broncho-alveolar inflammation [2]. Due to the lack of antibodies to distinguish the specific type of cells in the lung, the pathological analysis has been limited.
Podoplanin (PDPN) is an important marker protein in lung type I alveolar epithelial cells [3,4], podocytes in kidney [5], and lymphatic endothelial cells [6,7]. PDPN possesses a heavily glycosylated N-terminal extracellular domain, a single transmembrane domain, and a short intracellular domain [8,9]. A repeat sequence of EDxxVTPG, named PLAG domains are present in the N-terminal domain. “PLAG” is derived from the platelet aggregation-stimulating function of PDPN [10]. Furthermore, several PLAG-like domains (PLDs) have been identified [11].
To investigate the pathogenesis of lung type I alveolar epithelial cells in SARS-CoVs infected ferret, anti-ferret PDPN (ferPDPN) monoclonal antibodies (mAbs) have been thought as an essential tool. We previously established an anti-ferPDPN mAb, PMab-292 using the Cell-Based Immunization and Screening method. PMab-292 can be used for flow cytometry, western blotting, and immunohistochemistry [12]. To clarify further characteristics of PMab-292, we performed epitope mapping using flow cytometry.
2. Materials and Methods
2.1. Antibodies
2.2. Plasmid Construction and Transfection
Synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding ferPDPN plus an N-terminal MAP tag (GDGMVPPGIEDK) [16,17] were subcloned into a pCAG-Ble vector. The ferPDPN deletion mutants (dN33 to dN43) were produced using a KAPA HiFi HotStart ReadyMix PCR Kit (Kapa Biosystems, Wilmington, MA), and subcloned into the pCAG-Ble vector.
For PA scanning, the substitution of PA tag (GVAMPGAEDDVV) in ferPDPN dN34 was performed with oligonucleotides containing PA tag sequence at the desired position. For example, for the substitution of the PA tag from Val34 to Asn45 of ferPDPN, we constructed Thr33-GVAMPGAEDDVV-Asn46 (34-PA-45) in ferPDPN dN34.
2.3. Flow Cytometry
CHO-K1 cells and transfectants were treated with PMab-292 (1 μg/mL), PMab-241 (1 μg/mL), PMab-1 (1 μg/mL), or NZ-1 (1 μg/mL) for 30 min at 4°C. The cells were further treated with Alexa Fluor 488-conjugated anti-mouse IgG or anti-rat IgG (1:1000). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
3. Results
3.1. Epitope Mapping of PMab-292 Using the N-Terminal Deletion Mutants of ferPDPN
We previously established an anti-ferPDPN mAb (PMab-292) by the CBIS method. To determine the PMab-292 epitope, we examined the reactivity to the N-terminal ferPDPN deletion mutants (from dN33 to dN43)-overexpressed CHO-K1 cells by flow cytometry (Figure 1A). As shown in Figure 1B, PMab-292 reacted with dN33 and dN34. In contrast, the reactivity completely disappeared in dN35, dN36, dN37, dN38, dN39, dN40, dN41, dN42, and dN43. We confirmed the cell surface expression of all deletion mutants using PMab-241, originally developed as an anti-bear PDPN mAb, which crossreacts with ferPDPN (epitope amino acids: Thr86, Asp87, and Arg89) [18]. These results indicated that the Val34 is located at the N-terminus of PMab-292 epitope.
To identify the binding epitope of PMab-292, we generated 20 alanine (or glycine)-substituted ferPDPN (Supplemental Figure S1A) and investigated the reactivity of PMab-292 against CHO-K1 cells, which overexpressed the ferPDPN mutants transiently. PMab-292 reacted with all alanine (or glycine)-substituted mutants and wild-type (WT) (Supplemental Figure S1B). We also examined the reactivity of PMab-292 against 2 × alanine (or glycine)-substituted ferPDPN (Supplemental Figure S2A); however, PMab-292 reacted with all mutants (Supplemental Figure S2B). Therefore, we could not determine the epitope of PMab-292 using 1×Ala or 2×Ala scanning methods.
3.2. Flow Cytometry Using PMab-292 with PA Tag-Substituted ferPDPN
As shown in Figure 1B, PMab-292 reacted with the ferPDPN dN34 mutant. We next generated PA tag (GVAMPGAEDDVV)-substituted ferPDPN dN34 mutants as shown in Figure 2A to determine the C-terminus of the PMab-292 epitope. As shown in Figure 2B, the reactivity of PMab-292 almost completely disappeared in 34-PA-45, 35-PA-46, 36-PA-47, 37-PA-48, 38-PA-49, and 39-PA-50 mutants of ferPDPN dN34. In contrast, the reactivity of PMab-292 was observed in the substituted mutants from 40th to 59th amino acids. These results indicated that the epitope of PMab-292 contains the 34-VRPEDD-39 sequence of ferPDPN.
4. Discussion
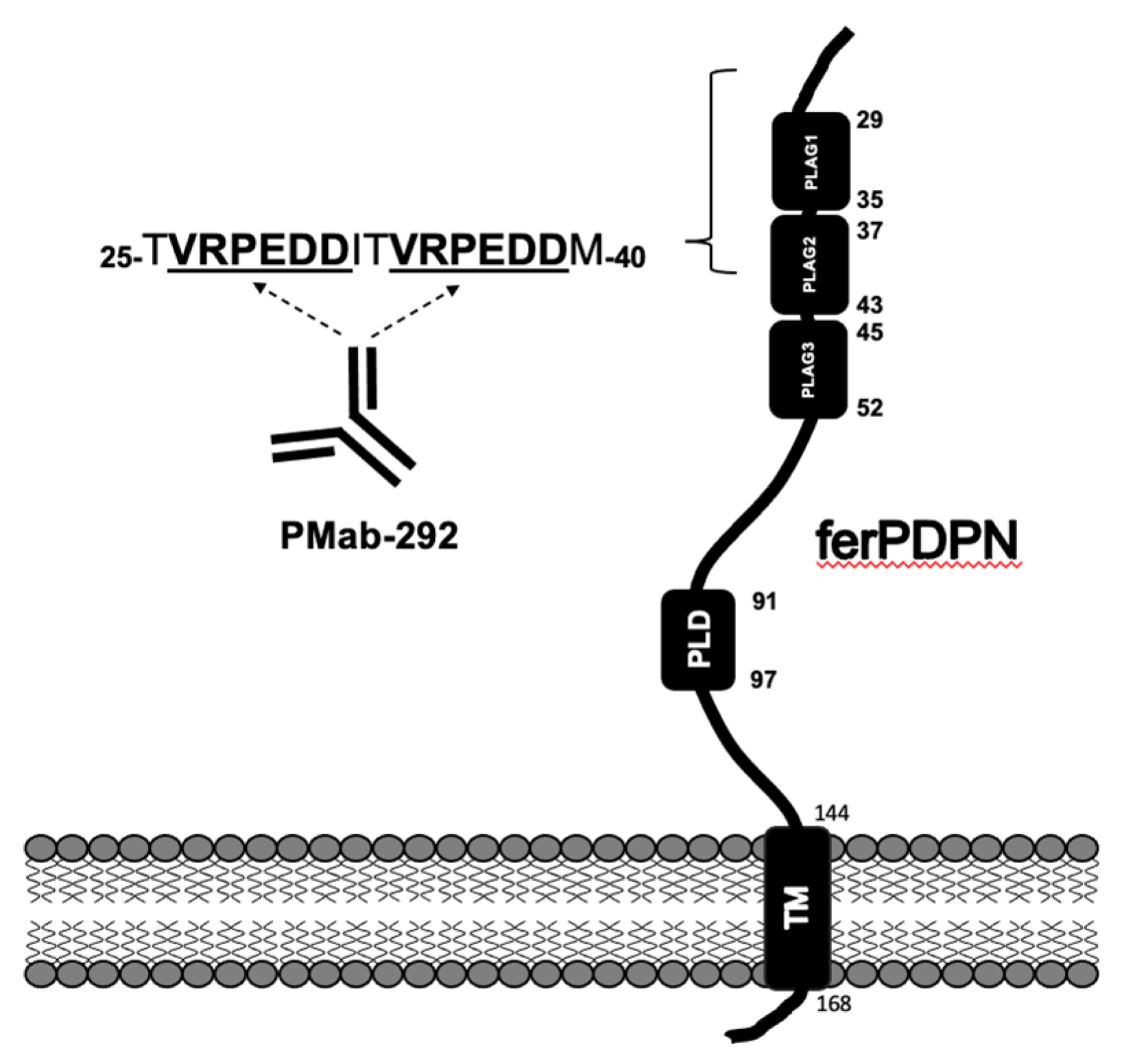
In this study, we performed the flow cytometry-mediated epitope mapping of PMab-292 using ferPDPN deletion mutants (Figure 1) and PA scanning of ferPDPN (Figure 2). We found that 34-VRPEDD-39 sequence of ferPDPN is an important sequence of PMab-292 epitope. In contrast, we could not identify the critical amino acids using 1×Ala scanning (Supplemental Figure S1) or 2×Ala scanning (Supplemental Figure S2) using flow cytometry. Figure 3 shows the surrounding sequence of 34-VRPEDD-39, and found that the same VRPEDD sequence is present between the 26th and 31st amino acids of ferPDPN. This might be the reason why we could not determine the critical amino acids of the ferPDPN epitope using 1×Ala scanning and 2×Ala scanning in the 34-VRPEDD-39 sequence. In other words, PMab-292 probably recognizes the tandem sequences from PLAG1 to PLAG2 domains (Figure 3). Therefore, the alanine-substitution of two same amino acids in both 26-VRPEDD-31 and 34-VRPEDD-39 sequence is essential to determine the critical amino acids of PMab-292 epitope.
PLAG domains play a critical in platelet aggregation. In humans, the O-glycosylation in PLAG3 or PLD domain is crucial for PDPN-induced platelet aggregation [19,20]. A platelet receptor, CLEC-2 recognizes the sialylated PLAG3 or PLD domains of PDPN, which is essential for PDPN-induced platelet aggregation. We have not examined whether ferPDPN can induce platelet aggregation, and have not determined the essential O-glycosylated amino acids for platelet aggregation in PLAG domains. However, PMab-292 could contribute to the study of platelet aggregation by ferPDPN.
We have determined the epitopes of anti-PDPN mAbs against various species [11]. It is the first example that the tandem sequence is recognized by an anti-PDPN mAb. We performed immunohistochemistry using PMab-292 and could detect kidney podocytes and lung alveolar epithelial cells with high sensitivity [12]. The property of PMab-292 may contribute to the high sensitivity of immunohistochemical analysis, which is important for the pathological analysis of lung injury by SARS-CoVs.
References
- van den Brand, J.M.; Haagmans, B.L.; van Riel, D.; Osterhaus, A.D.; Kuiken, T. The pathology and pathogenesis of experimental severe acute respiratory syndrome and influenza in animal models. J. Comp. Pathol. 2014, 151, 83–112. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Dobbs, L.G.; Williams, M.C.; Gonzalez, R. Monoclonal antibodies specific to apical surfaces of rat alveolar type I cells bind to surfaces of cultured, but not freshly isolated, type II cells. Biochim. Biophys. Acta 1988, 970, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Rishi, A.K.; Joyce-Brady, M.; Fisher, J.; Dobbs, L.G.; Floros, J.; VanderSpek, J.; Brody, J.S.; Williams, M.C. Cloning, characterization, and development expression of a rat lung alveolar type I cell gene in embryonic endodermal and neural derivatives. Dev. Biol. 1995, 167, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder-Geleff, S.; Matsui, K.; Soleiman, A.; Meraner, P.; Poczewski, H.; Kalt, R.; Schaffner, G.; Kerjaschki, D. Podoplanin, novel 43-kd membrane protein of glomerular epithelial cells, is down-regulated in puromycin nephrosis. Am. J. Pathol. 1997, 151, 1141–1152. [Google Scholar] [PubMed]
- Hirakawa, S.; Hong, Y.K.; Harvey, N.; Schacht, V.; Matsuda, K.; Libermann, T.; Detmar, M. Identification of vascular lineage-specific genes by transcriptional profiling of isolated blood vascular and lymphatic endothelial cells. Am. J. Pathol. 2003, 162, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Petrova, T.V.; Mäkinen, T.; Mäkelä, T.P.; Saarela, J.; Virtanen, I.; Ferrell, R.E.; Finegold, D.N.; Kerjaschki, D.; Ylä-Herttuala, S.; Alitalo, K. Lymphatic endothelial reprogramming of vascular endothelial cells by the Prox-1 homeobox transcription factor. Embo J. 2002, 21, 4593–4599. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.; Rayes, J.; Miyashita, T.; Ishii, G.; Retzbach, E.P.; Sheehan, S.A.; Takemoto, A.; Chang, Y.W.; Yoneda, K.; Asai, J.; et al. Podoplanin: An emerging cancer biomarker and therapeutic target. Cancer Sci. 2018, 109, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, M.; Montero-Montero, L.; Renart, J.; Martín-Villar, E. Podoplanin in Inflammation and Cancer. Int. J. Mol. Sci. 2019, 20, 707. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Fujita, N.; Kunita, A.; Sato, S.; Kaneko, M.; Osawa, M.; Tsuruo, T. Molecular identification of Aggrus/T1alpha as a platelet aggregation-inducing factor expressed in colorectal tumors. J. Biol. Chem. 2003, 278, 51599–51605. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Kaneko, M.K.; Kato, Y. Roles of Podoplanin in Malignant Progression of Tumor. Cells 2022, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Monoclonal Antibody PMab-292 Against Ferret Podoplanin. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Yamada, S.; Konnai, S.; Ishinazaka, T.; Shimozuru, M.; Kaneko, M.K.; Kato, Y. PMab-241 Specifically Detects Bear Podoplanin of Lymphatic Endothelial Cells in the Lung of Brown Bear. Monoclon. Antib. Immunodiagn. Immunother. 2019, 38, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Kaji, C.; Tsujimoto, Y.; Kato Kaneko, M.; Kato, Y.; Sawa, Y. Immunohistochemical Examination of Novel Rat Monoclonal Antibodies against Mouse and Human Podoplanin. Acta Histochem. Cytochem. 2012, 45, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kaneko, M.K.; Kuno, A.; Uchiyama, N.; Amano, K.; Chiba, Y.; Hasegawa, Y.; Hirabayashi, J.; Narimatsu, H.; Mishima, K.; et al. Inhibition of tumor cell-induced platelet aggregation using a novel anti-podoplanin antibody reacting with its platelet-aggregation-stimulating domain. Biochem. Biophys. Res. Commun. 2006, 349, 1301–1307. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon. Antib. Immunodiagn. Immunother. 2016, 35, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, A.; Kaneko, M.K.; Kato, Y.; Takagi, J.; Arimori, T. Site-specific epitope insertion into recombinant proteins using the MAP tag system. J. Biochem. 2020, 168, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Sayama, Y.; Sano, M.; Asano, T.; Furusawa, Y.; Takei, J.; Nakamura, T.; Yanaka, M.; Okamoto, S.; Handa, S.; Komatsu, Y.; et al. Epitope Mapping of PMab-241, a Lymphatic Endothelial Cell-Specific Anti-Bear Podoplanin Monoclonal Antibody. Monoclon. Antib. Immunodiagn. Immunother. 2020, 39, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Kato, Y.; Kameyama, A.; Ito, H.; Kuno, A.; Hirabayashi, J.; Kubota, T.; Amano, K.; Chiba, Y.; Hasegawa, Y.; et al. Functional glycosylation of human podoplanin: Glycan structure of platelet aggregation-inducing factor. FEBS Lett. 2007, 581, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, T.; Takemoto, A.; Takagi, S.; Takatori, K.; Sato, S.; Takami, M.; Fujita, N. Targeting a novel domain in podoplanin for inhibiting platelet-mediated tumor metastasis. Oncotarget 2016, 7, 3934–3946. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Epitope determination of PMab-292 using deletion mutants of ferPDPN. (A) The ferPDPN deletion mutants were transiently expressed in CHO-K1 cells. (B) The ferPDPN mutants-expressed CHO-K1 cells were incubated with 1 μg/mL of PMab-292 (B, red line) or 1 μg/mL of PMab-241 (C, red line), or control blocking buffer (black line), followed by secondary antibodies treatment. The data were analyzed using the SA3800 Cell Analyzer.
Figure 1.
Epitope determination of PMab-292 using deletion mutants of ferPDPN. (A) The ferPDPN deletion mutants were transiently expressed in CHO-K1 cells. (B) The ferPDPN mutants-expressed CHO-K1 cells were incubated with 1 μg/mL of PMab-292 (B, red line) or 1 μg/mL of PMab-241 (C, red line), or control blocking buffer (black line), followed by secondary antibodies treatment. The data were analyzed using the SA3800 Cell Analyzer.
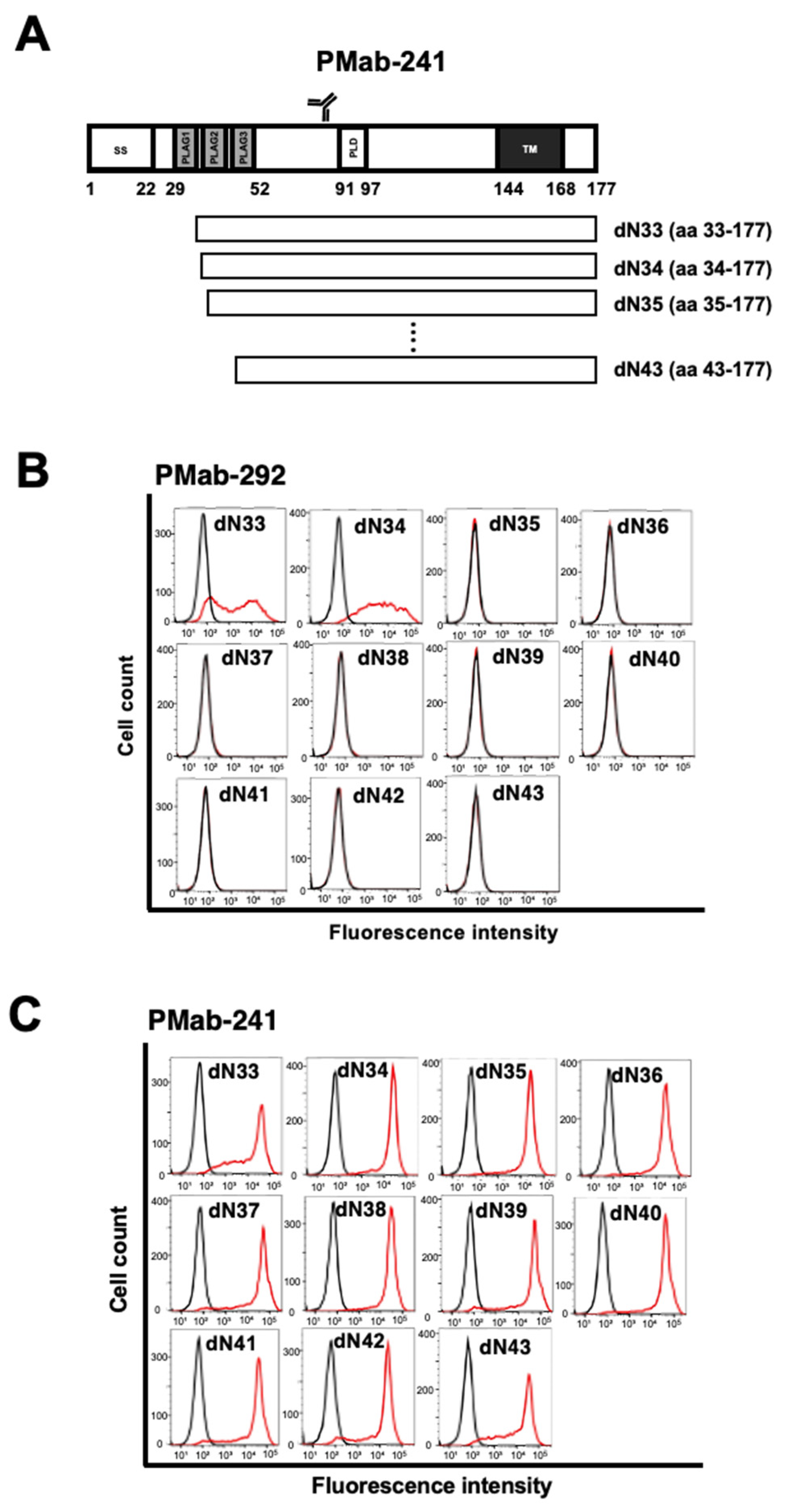
Figure 2.
Epitope determination of PMab-292 using PA tag-substituted mutants of ferPDPN. (A) The ferPDPN dN34 PA tag-substituted mutants were transiently expressed in CHO-K1 cells. (B) The mutants-expressed CHO-K1 cells were incubated with 1 μg/mL of PMab-292 (B, red line), 1 μg/mL of PMab-241 (C, red line), 1 μg/mL of NZ-1 (D, red line), or control blocking buffer (black line), followed by secondary antibodies treatment. The data were analyzed using the SA3800 Cell Analyzer.
Figure 2.
Epitope determination of PMab-292 using PA tag-substituted mutants of ferPDPN. (A) The ferPDPN dN34 PA tag-substituted mutants were transiently expressed in CHO-K1 cells. (B) The mutants-expressed CHO-K1 cells were incubated with 1 μg/mL of PMab-292 (B, red line), 1 μg/mL of PMab-241 (C, red line), 1 μg/mL of NZ-1 (D, red line), or control blocking buffer (black line), followed by secondary antibodies treatment. The data were analyzed using the SA3800 Cell Analyzer.
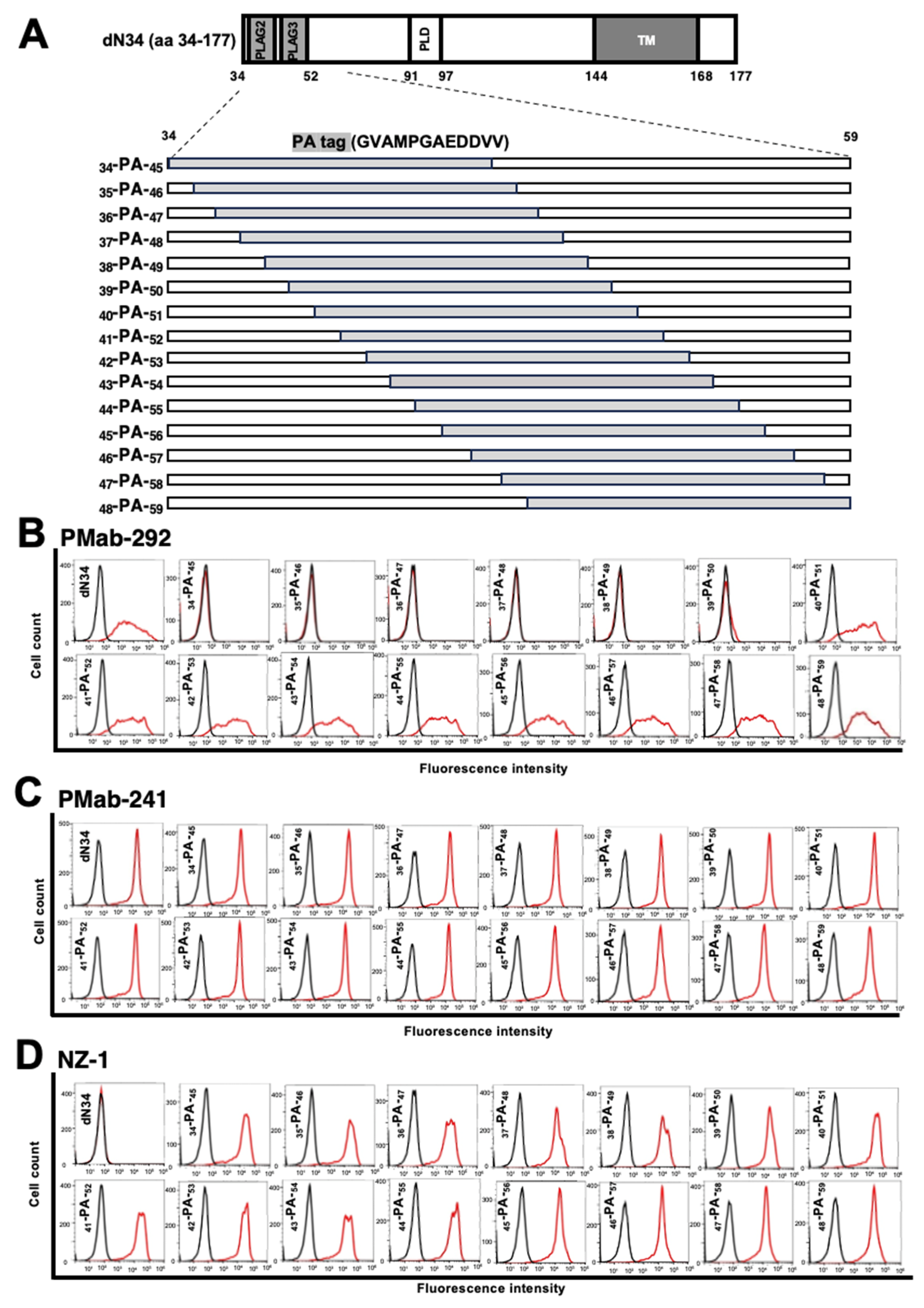
Figure 3.
Schematic illustration of ferPDPN and the PMab-292 epitope. The PMab-292 epitope involves tandem sequence (VRPEDD) of ferPDPN. PLAG, platelet aggregation-stimulating; PLD, PLAG-like domain; TM, transmembrane domain.
Figure 3.
Schematic illustration of ferPDPN and the PMab-292 epitope. The PMab-292 epitope involves tandem sequence (VRPEDD) of ferPDPN. PLAG, platelet aggregation-stimulating; PLD, PLAG-like domain; TM, transmembrane domain.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



