
Submitted:
29 November 2023
Posted:
30 November 2023
You are already at the latest version
Abstract
Temperature increases mediated through climate change threaten the survival of species. It is of foremost importance to engage citizens and future generations in understanding the mechanisms through which temperatures imposes its affect. For educators though this is not straight forward as tools for examined the impact of temperature over the lifetime of an animal are prohibitively expensive. At the same time environmental educators need guidance on the appropriate study systems to use with a balance between the species having an obvious response and ensuring the outcomes are ethical and sustainable. In our study, we created and tested a cost-effective experiment meant to be used for environmental education purposes. More specifically, we tested the sensitivity of the Painted lady butterfly Vanessa cardui to temperature variations using a homemade incubator. We describe the design of this experiment and report findings on survival rate, morphological variations, pupation time, and wingspan of adults across a range of biologically relevant temperatures. The information provided give educators options for testing a variety of hypotheses with regards to the impacts of temperature using an affordable and flexible set-up. Furthermore, the findings can be used by students to develop an understanding of the ramifications of the butterflies responses in an ecological context.
Keywords:
active learning
; development
; phenology
; pollinator
; climate change
; incubator
; survival rate
; pupation time
; butterfly
1. Introduction
The active learning paradigm of constructing individual meaning and building internal and personal representations of knowledge require personal experience [1]. Active or experimental learning, in which an individual’s own experiences with participating in practical activities that affirm their learning, strongly influence how individuals’ frame information and develop attitudes [2]. Like that of the kinaesthetic learning (i.e., associating physical movement with learning goals), experimental learning is effective at promoting investment in actioning solutions to problems than simply visual and auditory learning styles among young students [3]. Active learning teaching styles produce students with higher levels of emotional intelligence, i.e., self-awareness, self-regulation, motivation, empathy and social skills [4]. Highlighting the likelihood of an individual choosing to invest in future pro-environmental behaviours as an adult is framed by their own personal experience [5].
A challenging but crucial climate change concept for environmental educators, is to develop active learning approaches, that acutely illustrate the insidious impacts that rising temperatures can have on the biology of organisms [6]. Science recognises that rapid threshold increases in temperature can lead to species extinctions [7,8]. For the wider populace though the link between what seems little increases in mean temperature and the widescale decline and collapse of species is largely intangible. Much of this is possibly because the influence of temperature and the mechanism by which it acts is nuanced, subtle and often realised across subsequent generations rather than having instantaneous consequences [9]. Temperature increases can act directing in changing physical and sensory features that can adversely alter mobility and sensitivity to cues. These modifications though can also be adaptive (i.e., phenotypically plastic), with expressed traits acting to buffer against, avoid or reduce temperature related stress [10,11,12]. Temperature increases of the magnitude we are currently experiencing though is leading to large scale shifts in phenology, i.e., seasonal timings [9,13]. Such shifts have been shown to result in a mismatch between an organism’s readiness to use a resource and the resources availability. Indeed, examples of key relationships have been shown to be out of sync such as pollinators and floral resources [14,15], migratory birds and their prey [16], and herbivores and their host plants [17,18]. The importance of understanding the impact of phenology is evidenced by global initiatives that have been set-up to track these shifts in nature many of which have relied on citizens to collect data [19,20,21]. Such initiatives even suggest that establishing phenology networks has been key to elevating public awareness of the impacts of climate change. The part of the story missing for participants of such initiatives though is the mechanism through which temperature imposes these phenological shifts.
There are significant barriers for educators wishing to explore the mechanisms by which temperature influences species survival and trait development. The first is that controlled experiments typically require expensive infrastructure such as temperature-controlled incubators. Incubators designed for rearing insects used by scientists rather from £500-well over £2000 for equivalent small units. The second barrier is knowledge of a study system that provides reliable outcomes when exposed to a range of temperatures. Phenology shifts are well documented in butterflies with phenology traits promoted as indicators of species response to the changing environment [22]. As a poikilothermic animal, temperature highly influences their life cycle [23]. Over the past two decades, first appearance, mean flight date and flight periods have increased, parallel with an increase of 1-1.5 ̊C of central England spring and summer temperatures [23]. Migratory species can be acutely vulnerable to environmental change since they depend on a series of habitats throughout their migration [24]. Thus, it is likely that they may also have plastic responses to temperature rather than just providing survival data.
For this study we explored the use of Vanessa cardui (Painted Lady) as a candidate for system to study thermal tolerance and plasticity as it appeared to meet our pre-determined eligibility criteria. Our criteria for selection is the species have:
- a conservation status of Least Concern: the British Isles is wholly dependent on immigrants from the continent and they arrive in substantial numbers,
- A wide distribution range to increase the prospect the species may require adaptations to deal with different environmental gradients and to increase the chance the study can be replicated globally: V. cardui is a long-distance migrant with a wide distribution range, inhabiting all continents except for Australia and Antarctica [25].
- Larvae are easily attainable in high numbers and ethically sourced, breeding requires minimal demands for teachers and students with the animal being relatively robust: there are retailers that breed butterflies to sell as pets and they are found almost anywhere, from coastal to urban area, with it being one of the few species that can breed intensively in a variety of habitats;
- There is some indication that the species show gradual and measurable changes across the thermal range for which our study was to be conducted: adults are first seen in late March and numbers continue to rise through May and June as further migrants arrive from the continent [26]. A few publications have shown V. cardui vary in developmental timing and wing morphology across different temperatures.
- Cosmopolitan so the final release of the butterflies is unlikely to cause any environmental damage itself. This species is not a conservation concern.
This study had two main aims the first being to develop and test a cost-effective and experimental flexible set-up that educators can use to give students first-hand experience of the mechanisms by which shifts in temperature can alter the fitness of organisms. The second was to demonstrate the effectiveness of the set-up under proper experimental conditions to address the question of how does temperature influence the biology of V. cardui? It was hypothesised that if the set-up was effective subjecting the larvae to increased temperatures should lead to predictable phenological adaptations. If true, it was predicted: 1. Larvae of V. cardui incubated in warm temperatures will experience early pupation duration and early emergence, 2. Larvae incubated in high temperatures will have a higher mortality rate, 3. With warming temperatures there would be an increased likelihood of phenotypic shifts and trait aberrations. It was also hoped that outcomes of the study could help educators design their own experiments with the knowledge of the sensitivity of V. cardui exposed to a range of temperatures and an understanding of the optimal temperatures for studying survival estimates versus phenotypic responses where optimal is defined as comprise between survivability (i.e., low mortality rate) and seeing clear phenological responses in a timely manner.
2. Materials and Methods
2.1. Source and Husbandry of V. cardui
For this study, three Butterfly Garden Kits were purchased from a conservation education company Insect lore (£64.99 each). Each kit contains: pop-up, reusable 63cm tall clear mesh habitat; 35 live V. cardui larvae; specially formulated, ready to use, larvae food; 35 vials with lids; 70 sticking pads; feeding pipette; plastic spoon; food levelling tool; small transfer brush. This provided 3rd instar V. cardui larvae. Each larva was kept in its own housing which were plastic containers of 5cm x 5cm x 6cm, airholes in the lid and an allocation of larvae food. The larval food was provided by the company and included a classified formular specialised for V. cardui larvae. Each container received one teaspoon of this food which was enough to sustain the larvae until pupation i.e., no food replacement was required (Figure 1).
2.2. Incubator Design
To ensure the study was replicated by schools it was necessary to design a cost-effective and robust incubator set-up. Considerations of the design needed to consider affordability of the incubator design and its capacity to ensure temperatures were reliably maintained. Here we outline the design and some of the design considerations that were made, (Table 1).
The incubator was a modular design made with off the shelf items and easily constructed and included the following design elements; 1. Basking clip-on spot lamp: was used for ease of assembly. The basking spotlight selected was due to the shape of the bulb, categorised as a reflector bulb (code R) this bulb generates diffused heat in all directions creating an evenly distributed heat source throughout the incubator. The wattage of the bulbs was selected due to the desired temperature output for each treatment 25-35 ̊C = 50w and 40 ̊C = 100w. Ideally using the same wattage would be simpler but it was not possible to create 40 ̊C environment using the 50w. 2. The Habistat dimming thermostat was selected for its low-high monitoring range and accuracy. The thermostat was responsible for keeping temperatures consistent between 0.9 of a ̊C for all temperature treatments, for example 25.0 – 25.9 ̊C. The thermostat automatically turns off the light briefly when temperatures are exceeded. 3. The incubator body was created from polystyrene foam boards, this was selected due to its high thermal resistance, ease of assembly and accessibility. Each incubator was 42cm x 30cm x 44cm in size with the door cut to half size. 4. The 30cm butterfly habitat nets were selected to fit within the incubators, allowing for heat, airflow and moisture to be exchangeable. 5. The digital thermometers were selected for accuracy of interpretating a variety of temperature ranges. Connected to the inside of the mesh butterfly net for accuracy, displayed on the outer body for ease of interpreting. 6. A half sheet of polystyrene foam was used to cover half of the incubators front. This aspect of the design experimented with a full-length polystyrene cover and no cover at all. The full cover decreased airflow and increased the temperature past the desired outcome. In contrast, no cover was unable to achieve the desired temperature. The half door design provided adequate airflow whilst maintaining the desired temperature for each treatment. Incubators were set up three days prior to receiving live specimens, this enabled necessary adjustments to heat source placement and thermostat settings to ensure a consistent temperature was present and evenly distributed throughout each incubator. Temperatures continued to be monitored daily throughout the entirety of the experiment. An additional temperature gun was used to ensure accuracy. Finally, all electrical elements were checked, and PAT tested by an electrical and maintenance company Vinchi for compliance to ensure the set-up was safe.
2.2.1. Experimental Design
Twenty 4th instar larvae were exposed to one of 5 temperature ranges. The larvae were received as 3rd instars. Previous studies indicate a higher mortality rate for V. cardui larvae occur at temperatures exceeding 28 ̊C prior to 4th instar stage (Huang et al., 2022). All 3rd instar V. cardui larvae were kept at room temperature (18.5 ̊C) until reaching 4th instar. All individuals were re-measured three days post arrival, with all individuals measuring between 13-16mm, confirming 4th instar stage (Table A1). Temperature gradients included room temperature (18.5 ̊C), 25 ̊C, 30 ̊C, 35 ̊C and 40 ̊C. The room temperature enclosure consisted of one 63cm mesh habitat and a White Python Digital Thermometer Hygrometer without any incubator housing. V. cardui larvae were randomly assigned to temperature treatments using a random number generator. Individuals were placed into the centre of their allocated incubators. Room temperature was selected as a treatment with the idea that it might provide a treatment that the schools do not need an incubator set-up for. There is of course a limitation here as it assumes the room temperature is similar wherever the study is done.
2.2.2. Measurements
Physiological changes of larvae were recorded three days post incubation (Table A2). A colour code key was referenced when noting changes of V. cardui larvae (Figure 2). Survival rate (individuals alive per day) and phenological changes such as pupation date, pupation duration and emergence rate of V. cardui larvae were recorded (Table A3). Post emergence morphological attributes of V. cardui adults were recorded including wingspan (mm) and any visible abnormalities (i.e., deformed wing, missing limb) (Table A4).
2.2.3. Data Analysis for Baseline Butterfly Experiment
Analyses were conducted in R Studios, version 4.1 (RStudio Team, 2020). Calculations of pupation duration (days) were also identified, and wingspan (mm) were recorded. Individuals were coded to represent colour (i.e., B=black, B&W=black and white, W=white). A survival analysis was used to plot the probability of survival of V. cardui larvae to emergence using Survminer [27] and ggplot2 [28]. The analysis was parametrised with survival and event being the number of days since 4th instar. Generalised Linear Mixed Models were used for condition of V.cardui (condition classified as deformed or formed), pupation duration and wingspan size. Package instalments included, Generalized Linear Mixed Model using Template Model Builder (glmmTMB) [29], Diagnostics for Hierarchical Regression Models (DHARMA) [30] and Estimated Marginal Means (emmeans) [31]. To ensure the best fit a variety of family models were tested for each response variable. Selected models for each response variable were, Generalized Poisson distribution (genpois) = pupation duration, genpois = wingspan [29]. Colour variation of V. cardui larvae in relation to temperature treatments was displayed in a bar chart.
3. Results
3.1. Survival Rate of V. cardui to Temperature Variation.
Probability of survival of V. cardui larvae to emergence and the rate of development varied among the different temperature treatments (Figure 3). The highest temperature treatment at 40 ̊C displayed 100% mortality rate, with 98% of those mortalities occurring before pupation. Both the 35 ̊C and 30 ̊C completed their full life cycle in the same amount of time, however the 30 ̊C temperature treatment displayed far less attrition. Whereas room temperature displayed a 100% survival rate but took twice as long to complete their cycles relative to the 30-35 ̊C temperature treatments. Interestingly, the 25 ̊C led to relatively minor attrition but took around 10 days less to complete the cycle relative to the room temperature treatment.
3.2. Morphological Variations of V. cardui Larvae in Response to Temperature Variation
Morphological adaptations were present in V. cardui larvae between 4th – 5th instar under different temperature treatments. Prior to 4th instar stage and exposure to any temperature treatments all 100 individuals were black in colour. Changes in colour varied among the treatments, RT predominantly stayed the same prior to treatment exposure with 20% of larvae changing to black and white variations. As temperature increased the number of larvae to develop white colour variations increased as well as the distribution of white variation patterns until larvae were completely white in colour (Figure 4).
3.3. Morphological and Phenological Variations of V. cardui in Relation to Temperature
With room temperature as the exception, deformity was present in all temperature treatments in surviving V. cardui. The rate of expression of deformity though increased with temperature increase, (the value n represents the total number of individuals that made it to adulthood, the percentage represents the deformity rate out of the total number) RT (n=20) 0%, 25 ̊C (n=18)11.1%, 30 ̊C (n=17) 29.4%, 35 ̊C (n=7) 57.1%. Wingspan of fully formed V. cardui also varied amongst temperature treatments, except for one individual in the RT treatment which displayed a wingspan of 60cm. RT treatment displayed the largest collective of individuals with the smallest wingspan of 62 mm (n=20, 50%), in contrast to 25 ̊C and 30 ̊C (n=16, 12.5%; n=12, 0%). Wingspan displays greater then 62mm are (RT, n=20, 50%; 25 ̊C, n=16, 87.5%; 30 ̊C, n=12, 100%; 35 ̊C, n=4, 75%). Temperature treatment 30 ̊C displayed the largest wingspan of 70mm 16.6%. Phenological variations in the form of pupation duration (days) also varied in relation to temperature treatments. Room temperature displayed the longest pupation period with a mean pupation duration of 12.1 days. In contrast to 25 ̊C = 7.3 days, 30 ̊C = 5.7 days, 35 ̊C = 5.5 days. The rate of pupation duration decreased with temperature increase (Figure 5, Table 2).
4. Discussion
4.1. Temperatures Effect on Phenological and Morphological Attributes of V. cardui
Here we showed how that a basic ecological experiment manipulating temperature variations can alter the phenological traits of V. cardui as seen in other butterfly species. The optimal temperature (optimal defined as comprise between survivability, i.e., low mortality rate and seeing clear phenological responses in a timely manner) was consistent with studies using more sophisticated and expensive equipment at around 28 ̊C [32,33,34]. As anticipated larvae that were exposed to higher temperatures had an increase in development rate, and lower pupation period in line with other similar studies [35]. Likewise, a temperature threshold was reached in which the ability of the larvae to complete their maturation was clearly compromised while wing deformities were more apparent with an increase temperature.
Temperature variation also influenced phenotypic traits. Interestingly, our wingspan results were contrary to the ‘temperature-size rule’ [36] which suggest adults of ecotherms raised at higher temperatures are smaller than their counterparts raised at cooler temperatures [35,36]. This rule is far from absolute with several studies showing contradictory findings [37,38]. Indeed, the likelihood of temperature having a positive, negative or neutral impact on wingspan can be species specific [37] or dependent on the developmental stage at which the temperature increase occurs [38]. Wilson et al [38], for instance, found adults, from different families of butterflies, that were exposed to higher temperatures at late larval stages, as per our experiment, had substantial larger wingspan that those adults subjected at earlier or pupa stages. In fact, it seems different evolutionary outcomes maybe expected among populations or species with different migratory tendencies. In some ways increased wingspan for migratory species at higher temperatures makes sense given they are likely to invest in traits that facilitate migration [39,40]. Wingspan measurements though are rudimentary representatives of mobility potential. Indeed, mobility can be affected by temperature in other ways such as altering wing shape [41] or flight endurance [42].
Larvae also varied with the degree of melanisation decreasing with an increase temperature. Changes in colouration has been documented in the adults with lighter wing colouration occurring in individuals in warmer conditions [43]. As far as we are aware this is the first documentation of colour shifts in the larvae. Although not previously described for V. cardui this has been observed in other species such as Monarchs within both laboratory [44,45] and field conditions [12] and is suggested to be a direct consequence of melanisation being linked to thermoregulation, with greater melanisation needed in colder conditions.
4.2. Flexibility of experimental set-up
Here we demonstrated that the combination of the homemade set-up with the chosen study system is a powerful and scalable tool for testing the impacts of temperature on developmental timing and morphology. The cost effectiveness of the set-up means educators can potentially afford more than one incubator to examine impacts across gradients of temperature. The experiment also provides a range of data types making it scalable from early to late school students and even into early university. For example, the data collection can be as simple as calculating the time it takes from larvae to adult emergence and in this case, there is only a need to compare two temperatures with one possibly being just room temperature. This experiment is ideal for young students who are coming to terms with counting and the concept of days/time. For more advanced students into high school students can do measurements of traits such as wingspan and colour and they can consider the ramifications of when changes in phenology and morphology coexist. While at university students can use this set-up for their own experiments, test across a range of temperatures and look at more advanced investigations of survival rates and colour change, use geometric morphometrics to investigate shape characteristics to value-add to size measurements and finally develop behavioural assays for larvae or adults to test behavioural consequences of the range of changes observed.
4.3. Active learning in environmental education
Relative to this study, experiencing first-hand the effects of a physical environmental process (i.e., temperature and phenotypic shifts) in an active learning environment rather than using visual and auditory learning styles, will produce a more inherent understanding of the biological principle and thus gain a sense of urgency as it has been personally experienced. For example, a mixed method study examined children aged ten-to-twelve over a fifteen-week program. The program combined digital photography and hands on educational activities focused on individual and collaborative change [46]. Children knew significantly more about the social and scientific dimensions of environmental change post program and were motivated by their growing environmental impact awareness to take action to minimize environmental harm [46]. Similarly, related studies also identified environmental engagement activities resulted in changes in skill, attitude and knowledge related to enhancing ecological, social and economic justice [47,48,49,50]. We feel this study provides the means for educators to give a real insight into the impacts of temperature on species survival.
5. Conclusions
We showed the steps to design a homemade and cost-effective experiment for environmental education to show the effect of temperature on V. cardui. We tested the approach under experimental conditions and found the set-up to provide reliable and interesting findings. We suggest to get the most out of the experiment findings, that scenario-based activities proceed in which students need to workshop the ramifications of the findings in an ecological framework. The set-up is used for V. cardui but is adaptable for other study systems and could potential open avenues for environmental educators to test their own hypotheses.
Author Contributions
Conceptualization, C.G., M.C. and M.B.; methodology, C.G., M.C. and M.B.; software, C.G., M.C. and M.B.; validation, C.G., M.C. and M.B.; formal analysis, C.G., M.C. and M.B.; investigation, C.G.; resources, C.G., M.C. and M.B.; data curation, C.G.; writing—original draft preparation, C.G.; writing—review and editing, M.C and M.B.; visualization, C.G., M.C. and M.B.; supervision, M.C. and M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw data can be found in the appendix.
Acknowledgments
We thank the Oakhurst Community Primary School for hosting the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Table A1.
Showing size (mm) and instar stage of painted lady larvae on the date of arrival (10.05.23) and the date they went into incubation (13.05.23).
Table A1.
Showing size (mm) and instar stage of painted lady larvae on the date of arrival (10.05.23) and the date they went into incubation (13.05.23).
| Physiological observation on 10.05.23 | Physiological observations on 13.05.23 | ||||||
|---|---|---|---|---|---|---|---|
| Group 1: Room Temperature (18.5 ̊C) | |||||||
| No. | Length (mm) | Colour | Instar Stage | No. | Length (mm) | Colour | Instar Stage |
| 1 | 10 | B | 3rd | 1 | 14 | B | 4th |
| 2 | 10 | B | 3rd | 2 | 13 | B | 4th |
| 3 | 10 | B | 3rd | 3 | 14 | B | 4th |
| 4 | 9 | B | 3rd | 4 | 13 | B | 4th |
| 5 | 11 | B | 3rd | 5 | 16 | B | 4th |
| 6 | 10 | B | 3rd | 6 | 15 | B | 4th |
| 7 | 11 | B | 3rd | 7 | 16 | B | 4th |
| 8 | 11 | B | 3rd | 8 | 16 | B | 4th |
| 9 | 9 | B | 3rd | 9 | 13 | B | 4th |
| 10 | 9 | B | 3rd | 10 | 13 | B | 4th |
| 11 | 10 | B | 3rd | 11 | 14 | B | 4th |
| 12 | 9 | B | 3rd | 12 | 13 | B | 4th |
| 13 | 11 | B | 3rd | 13 | 15 | B | 4th |
| 14 | 12 | B | 3rd | 14 | 16 | B | 4th |
| 15 | 10 | B | 3rd | 15 | 14 | B | 4th |
| 16 | 10 | B | 3rd | 16 | 14 | B | 4th |
| 17 | 9 | B | 3rd | 17 | 13 | B | 4th |
| 18 | 11 | B | 3rd | 18 | 15 | B | 4th |
| 19 | 10 | B | 3rd | 19 | 15 | B | 4th |
| 20 | 10 | B | 3rd | 20 | 14 | B | 4th |
| Group 2: 25 ̊C | |||||||
| 21 | 10 | B | 3rd | 21 | 15 | B | 4th |
| 22 | 9 | B | 3rd | 22 | 13 | B | 4th |
| 23 | 11 | B | 3rd | 23 | 16 | B | 4th |
| 24 | 11 | B | 3rd | 24 | 16 | B | 4th |
| 25 | 9 | B | 3rd | 25 | 13 | B | 4th |
| 26 | 11 | B | 3rd | 26 | 14 | B | 4th |
| 27 | 10 | B | 3rd | 27 | 13 | B | 4th |
| 28 | 9 | B | 3rd | 28 | 13 | B | 4th |
| 29 | 10 | B | 3rd | 29 | 14 | B | 4th |
| 30 | 10 | B | 3rd | 30 | 14 | B | 4th |
| 31 | 10 | B | 3rd | 31 | 14 | B | 4th |
| 32 | 10 | B | 3rd | 32 | 13 | B | 4th |
| 33 | 11 | B | 3rd | 33 | 16 | B | 4th |
| 34 | 9 | B | 3rd | 34 | 13 | B | 4th |
| 35 | 10 | B | 3rd | 35 | 14 | B | 4th |
| 36 | 11 | B | 3rd | 36 | 15 | B | 4th |
| 37 | 11 | B | 3rd | 37 | 16 | B | 4th |
| 38 | 11 | B | 3rd | 38 | 15 | B | 4th |
| 39 | 9 | B | 3rd | 39 | 13 | B | 4th |
| 40 | 9 | B | 3rd | 40 | 13 | B | 4th |
| Group 3: 30 ̊C | |||||||
| 41 | 10 | B | 3rd | 41 | 15 | B | 4th |
| 42 | 9 | B | 3rd | 42 | 13 | B | 4th |
| 43 | 9 | B | 3rd | 43 | 13 | B | 4th |
| 44 | 11 | B | 3rd | 44 | 15 | B | 4th |
| 45 | 9 | B | 3rd | 45 | 13 | B | 4th |
| 46 | 12 | B | 3rd | 46 | 16 | B | 4th |
| 47 | 10 | B | 3rd | 47 | 14 | B | 4th |
| 48 | 10 | B | 3rd | 48 | 14 | B | 4th |
| 49 | 11 | B | 3rd | 49 | 15 | B | 4th |
| 50 | 10 | B | 3rd | 50 | 14 | B | 4th |
| 51 | 10 | B | 3rd | 51 | 14 | B | 4th |
| 52 | 11 | B | 3rd | 52 | 15 | B | 4th |
| 53 | 12 | B | 3rd | 53 | 16 | B | 4th |
| 54 | 9 | B | 3rd | 54 | 13 | B | 4th |
| 55 | 10 | B | 3rd | 55 | 14 | B | 4th |
| 56 | 12 | B | 3rd | 56 | 16 | B | 4th |
| 57 | 11 | B | 3rd | 57 | 14 | B | 4th |
| 58 | 11 | B | 3rd | 58 | 15 | B | 4th |
| 59 | 10 | B | 3rd | 59 | 14 | B | 4th |
| 60 | 9 | B | 3rd | 60 | 13 | B | 4th |
| Group 4: 35 ̊C | |||||||
| 61 | 10 | B | 3rd | 61 | 14 | B | 4th |
| 62 | 10 | B | 3rd | 62 | 14 | B | 4th |
| 63 | 11 | B | 3rd | 63 | 15 | B | 4th |
| 64 | 9 | B | 3rd | 64 | 13 | B | 4th |
| 65 | 12 | B | 3rd | 65 | 16 | B | 4th |
| 66 | 10 | B | 3rd | 66 | 14 | B | 4th |
| 67 | 10 | B | 3rd | 67 | 14 | B | 4th |
| 68 | 11 | B | 3rd | 68 | 15 | B | 4th |
| 69 | 11 | B | 3rd | 69 | 15 | B | 4th |
| 70 | 11 | B | 3rd | 70 | 16 | B | 4th |
| 71 | 9 | B | 3rd | 71 | 13 | B | 4th |
| 72 | 10 | B | 3rd | 72 | 14 | B | 4th |
| 73 | 9 | B | 3rd | 73 | 13 | B | 4th |
| 74 | 9 | B | 3rd | 74 | 14 | B | 4th |
| 75 | 11 | B | 3rd | 75 | 15 | B | 4th |
| 76 | 9 | B | 3rd | 76 | 13 | B | 4th |
| 77 | 11 | B | 3rd | 77 | 16 | B | 4th |
| 78 | 10 | B | 3rd | 78 | 13 | B | 4th |
| 79 | 10 | B | 3rd | 79 | 14 | B | 4th |
| 80 | 10 | B | 3rd | 80 | 14 | B | 4th |
| Group 5: 40 ̊C | |||||||
| 81 | 10 | B | 3rd | 81 | 15 | B | 4th |
| 82 | 11 | B | 3rd | 82 | 16 | B | 4th |
| 83 | 12 | B | 3rd | 83 | 16 | B | 4th |
| 84 | 9 | B | 3rd | 84 | 13 | B | 4th |
| 85 | 10 | B | 3rd | 85 | 14 | B | 4th |
| 86 | 11 | B | 3rd | 86 | 15 | B | 4th |
| 87 | 11 | B | 3rd | 87 | 15 | B | 4th |
| 88 | 12 | B | 3rd | 88 | 16 | B | 4th |
| 89 | 11 | B | 3rd | 89 | 15 | B | 4th |
| 90 | 10 | B | 3rd | 90 | 13 | B | 4th |
| 91 | 9 | B | 3rd | 91 | 13 | B | 4th |
| 92 | 9 | B | 3rd | 92 | 13 | B | 4th |
| 93 | 9 | B | 3rd | 93 | 13 | B | 4th |
| 94 | 10 | B | 3rd | 94 | 14 | B | 4th |
| 95 | 11 | B | 3rd | 95 | 15 | B | 4th |
| 96 | 10 | B | 3rd | 96 | 14 | B | 4th |
| 97 | 11 | B | 3rd | 97 | 16 | B | 4th |
| 98 | 9 | B | 3rd | 98 | 13 | B | 4th |
| 99 | 9 | B | 3rd | 99 | 13 | B | 4th |
| 100 | 10 | B | 3rd | 100 | 15 | B | 4th |
Table A2.
Penological changes of V. cardui larvae three days post incubation.
| Physiological Changes three days post incubation: Date - 16.06.23 | |||||
|---|---|---|---|---|---|
| Group 1: Room Temp (18.5̊C) | |||||
| Number | Length (mm) | Colour | Number | Length (mm) | Colour |
| 1 | 15 | B | 11 | 20 | B |
| 2 | 17 | B/W | 12 | 16 | B |
| 3 | 17 | B | 13 | 22 | B |
| 4 | 20 | B | 14 | 16 | B/W |
| 5 | 18 | B | 15 | 19 | B |
| 6 | 18 | B | 16 | 17 | B |
| 7 | 20 | B | 17 | 18 | B |
| 8 | 15 | B | 18 | 19 | B |
| 9 | 15 | B | 19 | 17 | B |
| 10 | 20 | B | 20 | 26 | B/W |
| Group 2 : 25̊C | |||||
| 21 | 35 | B/W | 31 | 35 | B/W |
| 22 | 28 | B/W | 32 | 40 | B/W |
| 23 | 40 | B/W | 33 | 37 | B/W |
| 24 | 27 | B/W | 34 | 39 | B/W |
| 25 | 37 | W | 35 | 40 | B/W |
| 26 | 37 | B/W | 36 | 35 | B/W |
| 27 | 33 | B/W | 37 | 37 | B/W |
| 28 | 35 | B/W | 38 | 34 | B/W |
| 29 | 36 | W | 39 | 34 | B/W |
| 30 | 35 | B/W | 40 | 28 | B/W |
| Group 3: 30̊C | |||||
| 41 | 37 | B/W | 51 | 38 | B/W |
| 42 | 36 | B/W | 52 | 40 | B/W |
| 43 | 40 | W | 53 | 37 | W |
| 44 | 40 | W | 54 | 33 | B/W |
| 45 | 37 | B/W | 55 | 40 | W |
| 46 | 33 | W | 56 | 36 | B/W |
| 47 | 33 | W | 57 | 35 | B/W |
| 48 | 35 | B/W | 58 | 37 | B/W |
| 49 | 31 | B/W | 59 | 39 | B/W |
| 50 | 37 | B/W | 60 | 36 | B/W |
| Group 4: 35̊C | |||||
| 61 | - | W | 71 | 41 | W |
| 62 | 35 | W | 72 | 40 | W |
| 63 | 39 | W | 73 | 43 | W |
| 64 | 40 | W | 74 | 42 | B/W |
| 65 | 41 | W | 75 | 37 | W |
| 66 | 32 | B/W | 76 | 37 | W |
| 67 | 38 | W | 77 | 38 | W |
| 68 | 42 | W | 78 | 37 | B/W |
| 69 | 36 | W | 79 | 40 | W |
| 70 | 35 | W | 80 | 41 | W |
| Group 5: 40̊C | |||||
| 81 | - | W | 91 | - | W |
| 82 | - | W | 92 | 30 | W |
| 83 | - | W | 93 | - | W |
| 84 | 30 | W | 94 | 35 | W |
| 85 | - | W | 95 | - | W |
| 86 | - | W | 96 | - | W |
| 87 | - | W | 97 | 34 | W |
| 88 | 39 | W | 98 | - | W |
| 89 | - | W | 99 | - | W |
| 90 | - | W | 100 | 30 | W |
Table A3.
Phenological records of V. cardui emergence.
| Phenological Records | |||||||
|---|---|---|---|---|---|---|---|
| Group 1: Room Temperature (18.5̊C) | |||||||
| No. | Pupation Date | Pupation Emergence | Pupation Duration (days) | No. | PupationDate | Pupation Emergence | Pupation Duration (days) |
| 1 | 23.05.23 | 05.06.23 | 13 | 11 | 23.05.23 | 04.06.23 | 12 |
| 2 | 23.05.23 | 05.06.23 | 13 | 12 | 23.05.23 | 05.06.23 | 13 |
| 3 | 23.05.23 | 04.06.23 | 12 | 13 | 22.05.23 | 03.06.23 | 12 |
| 4 | 24.05.23 | 06.06.23 | 13 | 14 | 23.05.23 | 04.06.23 | 12 |
| 5 | 23.05.23 | 04.06.23 | 12 | 15 | 23.05.23 | 04.06.23 | 12 |
| 6 | 23.05.23 | 05.06.23 | 13 | 16 | 23.05.23 | 04.06.23 | 12 |
| 7 | 22.05.23 | 02.06.23 | 11 | 17 | 24.05.23 | 05.06.23 | 12 |
| 8 | 23.05.23 | 04.06.23 | 12 | 18 | 23.05.23 | 03.06.23 | 11 |
| 9 | 24.05.23 | 05.06.23 | 12 | 19 | 23.05.23 | 04.06.23 | 12 |
| 10 | 23.05.23 | 04.06.23 | 12 | 20 | 22.05.23 | 03.06.23 | 12 |
| Group 2: 25̊C | |||||||
| 21 | 18.05.23 | 25.05.23 | 7 | 31 | 17.05.23 | 24.05.23 | 7 |
| 22 | 18.05.23 | 25.05.23 | 7 | 32 | 17.05.23 | - | - |
| 23 | 18.05.23 | 25.05.23 | 7 | 33 | 18.05.23 | 25.05.23 | 7 |
| 24 | 18.05.23 | 25.05.23 | 7 | 34 | 18.05.23 | 25.05.23 | 7 |
| 25 | 18.05.23 | 25.05.23 | 7 | 35 | 19.05.23 | 27.05.23 | 8 |
| 26 | 18.05.23 | - | - | 36 | 18.05.23 | 25.05.23 | 7 |
| 27 | 17.05.23 | 25.05.23 | 8 | 37 | 17.05.23 | 25.05.23 | 8 |
| 28 | 17.05.23 | 25.05.23 | 8 | 38 | 18.05.23 | 25.05.23 | 7 |
| 29 | 18.05.23 | 25.05.23 | 7 | 39 | 18.05.23 | 25.05.23 | 7 |
| 30 | 18.05.23 | 26.05.23 | 8 | 40 | 18.05.23 | 26.05.23 | 8 |
| Group 3: 30̊C | |||||||
| 41 | 17.05.23 | 24.05.23 | 7 | 51 | 17.05.23 | 21.05.23 | 4 |
| 42 | 17.05.23 | 23.05.23 | 6 | 52 | 17.05.23 | 23.05.23 | 6 |
| 43 | 17.05.23 | 23.05.23 | 6 | 53 | 17.05.23 | 23.05.23 | 6 |
| 44 | 17.05.23 | 23.05.23 | 6 | 54 | - | - | - |
| 45 | 17.05.23 | 22.05.23 | 5 | 55 | 17.05.23 | 23.05.23 | 6 |
| 46 | 17.05.23 | 23.05.23 | 6 | 56 | 17.05.23 | 22.05.23 | 5 |
| 47 | 18.05.23 | 23.05.23 | 5 | 57 | 17.05.23 | 23.05.23 | 6 |
| 48 | 18.05.23 | Failed | - | 58 | 18.05.23 | 24.05.23 | 6 |
| 49 | 18.05.23 | 24.05.23 | 6 | 59 | 17.05.23 | 23.05.23 | 6 |
| 50 | 18.05.23 | Failed | - | 60 | 17.05.23 | 23.05.23 | 6 |
| Group 4: 35̊C | |||||||
| 61 | - | - | - | 71 | 18.05.23 | Failed | - |
| 62 | - | - | - | 72 | - | - | - |
| 63 | 17.05.23 | Failed | - | 73 | 18.05.23 | 23.05.23 | 5 |
| 64 | - | - | - | 74 | 18.05.23 | Failed | - |
| 65 | 17.05.23 | Failed | - | 75 | 18.05.23 | 23.05.23 | 5 |
| 66 | 19.05.23 | Failed | - | 76 | 17.05.23 | Failed | - |
| 67 | 17.05.23 | Failed | - | 77 | 18.05.23 | 24.05.23 | 6 |
| 68 | 18.05.23 | Failed | - | 78 | 17.05.23 | 23.05.23 | 6 |
| 69 | 18.05.23 | Failed | - | 79 | 17.05.23 | 23.05.23 | 6 |
| 70 | 17.05.23 | 23.05.23 | 6 | 80 | 18.05.23 | 23.05.23 | 5 |
| Group 5: 40̊C | |||||||
| 81 | - | - | - | 91 | - | - | - |
| 82 | - | - | - | 92 | 18.05.23 | Failed | - |
| 83 | - | - | - | 93 | - | - | - |
| 84 | 17.05.23 | Failed | - | 94 | 18.05.23 | Failed | - |
| 85 | - | - | - | 95 | - | - | - |
| 86 | - | - | - | 96 | - | - | - |
| 87 | - | - | - | 97 | 17.05.23 | Failed | - |
| 88 | - | - | - | 98 | - | - | - |
| 89 | - | - | - | 99 | - | - | - |
| 90 | - | - | - | 100 | - | - | - |
Table A4.
Morphological attributes of V. cardui upon emergence.
| Morphological Attributes Upon Emergence | |||||||
|---|---|---|---|---|---|---|---|
| Group 1: Room Temperature (18.5̊C) | |||||||
| No. | Pupa Position | Note | Wingspan (mm) | No. | Pupa Position | Note | Wingspan (mm) |
| 1 | Hanging | Perfectly formed | 64 | 11 | Hanging | Perfectly formed | 66 |
| 2 | Hanging | Perfectly formed | 62 | 12 | Hanging | Perfectly formed | 66 |
| 3 | Ground | Perfectly formed | 62 | 13 | Hanging | Perfectly formed | 64 |
| 4 | Hanging | Perfectly formed | 64 | 14 | Hanging | Perfectly formed | 62 |
| 5 | Hanging | Perfectly formed | 60 | 15 | Hanging | Perfectly formed | 62 |
| 6 | Hanging | Perfectly formed | 64 | 16 | Hanging | Perfectly formed | 68 |
| 7 | Hanging | Perfectly formed | 62 | 17 | Hanging | Perfectly formed | 62 |
| 8 | Hanging | Perfectly formed | 64 | 18 | Hanging | Perfectly formed | 62 |
| 9 | Hanging | Perfectly formed | 62 | 19 | Ground | Perfectly formed | 64 |
| 10 | Hanging | Perfectly formed | 66 | 20 | Hanging | Perfectly formed | 62 |
| Group 2: 25̊C | |||||||
| 21 | Hanging | Perfectly formed | 66 | 31 | Hanging | Perfectly formed | 64 |
| 22 | Hanging | Perfectly formed | 64 | 32 | Ground | Failed to emerge | - |
| 23 | Hanging | Perfectly formed | 68 | 33 | Hanging | Perfectly formed | 62 |
| 24 | Ground | Perfectly formed | 64 | 34 | Hanging | Perfectly formed | 68 |
| 25 | Hanging | Perfectly formed | 66 | 35 | Hanging | Perfectly formed | 64 |
| 26 | Ground | Failed to emerge | - | 36 | Hanging | Perfectly formed | 66 |
| 27 | Hanging | Right wings larger than the left | 60 | 37 | Hanging | Perfectly formed | 68 |
| 28 | Ground | Perfectly formed | 68 | 38 | Hanging | Perfectly formed | 64 |
| 29 | Hanging | Perfectly formed | 68 | 39 | Hanging | Perfectly formed | 66 |
| 30 | Hanging | Severely deformed on all wings, stuck in chrysalis | - | 40 | Hanging | Perfectly formed | 62 |
| Group 3: 30̊C | |||||||
| 41 | Hanging | Perfectly formed | 66 | 51 | Ground | Deformity in left forewing | 64 |
| 42 | Hanging | Perfectly formed | 70 | 52 | Hanging | Perfectly formed | 64 |
| 43 | Hanging | Perfectly formed | 70 | 53 | Hanging | Perfectly formed | 66 |
| 44 | Hanging | Perfectly formed | 68 | 54 | - | - | - |
| 45 | Ground | Deformity of right hind wing | 66 | 55 | Hanging | Slight uplift in outer left forewing | 64 |
| 46 | Hanging | Perfectly formed | 64 | 56 | Ground | Perfectly formed | 66 |
| 47 | Hanging | Perfectly formed | 68 | 57 | Hanging | Perfectly formed | 68 |
| 48 | Hanging | Failed to emerge | - | 58 | Hanging | Perfectly formed | 66 |
| 49 | Ground | Small, deformity of both the right hind & forewing | 44 | 59 | Ground | Severely deformed on all wings. Got stuck in chrysalis. | - |
| 50 | Ground | Failed to emerge | - | 60 | Ground | Perfectly formed | 66 |
| Group 4: 35̊C | |||||||
| 61 | - | - | - | 71 | Ground | Failed to emerge | - |
| 62 | - | - | - | 72 | - | - | - |
| 63 | Hanging | Failed to emerge | - | 73 | Ground | Perfectly formed | 66 |
| 64 | - | - | - | 74 | Ground | Failed to emerge | - |
| 65 | Ground | Failed to emerge | - | 75 | Ground | Perfectly formed | 58 |
| 66 | Ground | Failed to emerge | - | 76 | Ground | Failed to emerge | - |
| 67 | Ground | Failed to emerge | - | 77 | Ground | Severely deformed on all wings, got stuck in chrysalis | - |
| 68 | Hanging | Failed to emerge | - | 78 | Hanging | Perfectly formed | 66 |
| 69 | Ground | Failed to emerge | - | 79 | Hanging | Perfectly formed | 64 |
| 70 | Hanging | Severely deformed on all wings, got stuck in chrysalis | - | 80 | Ground | Abdomen stuck in chrysalis | 66 |
| Group 5: 40̊C | |||||||
| 81 | - | - | - | 91 | - | - | - |
| 82 | - | - | - | 92 | Ground | Failed to emerge | - |
| 83 | - | - | - | 93 | - | - | - |
| 84 | Hanging | Failed to emerge | - | 94 | Ground | Failed to emerge | - |
| 85 | - | - | - | 95 | - | - | - |
| 86 | - | - | - | 96 | - | - | - |
| 87 | - | - | - | 97 | Hanging | Failed to emerge | - |
| 88 | - | - | - | 98 | - | - | - |
| 89 | - | - | - | 99 | - | - | - |
| 90 | - | - | - | 100 | - | - | - |
References
- Jones, R; Bursens, P. The effects of active learning environments: how simulations trigger affective learning. Eur. Political Sci. 2015, 14, 254–265. [Google Scholar] [CrossRef]
- Gano-Phillips, S. Affective learning in general education. Special topic: Assessment in University General Education Program 2009, 6, 1–44. [Google Scholar]
- Vaishnav, S.R. Learning Style and Academic Achievement of Secondary School Students. Learning Style and Academic Achievements 2013, 1, 2277–7733. [Google Scholar]
- Marleny, L.; Aloysius, C.; Hadi, S. Emotional Intelligence among Auditory, Reading, and Kinaesthetic Learning Styles of Elementary School Students in Ambon-Indonesia. International Electronic Journal of Elementary Education 2017, 10, 83–91. [Google Scholar]
- Gifford, R.; Nilsson, A. Personal and social factors that influence pro-environmental concern and behaviour: A review. Int. J. Psychol. 2014, 49, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.A.; Heinen, R.; Gols, R.; Thakur, M.P. Climate change-mediated temperature extremes and insects: From outbreaks to breakdowns. Glob. Change Biol. 2020, 26, 6685–6701. [Google Scholar] [CrossRef]
- Song, H.; Kemp, D.B.; Tian, L.; Chu, D.; Song, H.; Dai, X. Thresholds of temperature change for mass extinctions. Nat. Comm. 2021, 12, 4694. [Google Scholar] [CrossRef] [PubMed]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef]
- Chmura, H.E.; Kharouba, H.M.; Ashander, J.; Ehlman, S.M.; Rivest, E.B.; Yang, L.H. The mechanisms of phenology: the patterns and processes of phenological shifts. Ecol. Monogr. 2019, 89, e01337. [Google Scholar] [CrossRef]
- Gérard, M.; Vanderplanck, M.; Wood, T.; Michez, D. Global warming and plant–pollinator mismatches. Emerg. Top. Life Sci. 2020, 4, 77–86. [Google Scholar]
- Buckley, L.B.; Kingsolver, J.G. Environmental variability shapes evolution, plasticity and biogeographic responses to climate change. Glob. Ecol. Biogeogr. 2019, 28, 1456–1468. [Google Scholar] [CrossRef]
- Tseng, M.; Bevanda, C.; Bhatti, S.S.; Black, E.N.; Chang, E.; Chiang, J.; Dhaliwal, H.; Dimitriou, A.; Gong, S.Y.; Halbe, E.; Harris, N. Effects of temperature on monarch caterpillar pigment variation in nature. Insect Conserv. Divers. 2023, 16, 164–171. [Google Scholar] [CrossRef]
- Cohen, J.M.; Lajeunesse, M.J.; Rohr, J.R. A global synthesis of animal phenological responses to climate change. Nat. Clim. Change 2018, 8, 224–228. [Google Scholar] [CrossRef]
- Miller-Struttmann, N.E.; Geib, J.C.; Franklin, J.D.; Kevan, P.G.; Holdo, R.M.; Ebert-May, D.; Lynn, A.M.; Kettenbach, J.A.; Hedrick, E.; Galen, C. Functional mismatch in a bumble bee pollination mutualism under climate change. Science 2015, 349, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Kudo, G.; Cooper, E.J. When spring ephemerals fail to meet pollinators: mechanism of phenological mismatch and its impact on plant reproduction. Proc. Natl. Acad. Sci. USA 2019, 286, 20190573. [Google Scholar] [CrossRef] [PubMed]
- Both, C.; Van Turnhout, C.A.; Bijlsma, R.G.; Siepel, H.; Van Strien, A.J.; Foppen, R.P. Avian population consequences of climate change are most severe for long-distance migrants in seasonal habitats. Proc. R. Soc. Lond., B 2010, 277, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.E.; Holleman, L.J. Warmer springs disrupt the synchrony of oak and winter moth phenology. Proc. R. Soc. Lond., B 2001, 268, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Posledovich, D.; Toftegaard, T.; Wiklund, C.; Ehrlén, J.; Gotthard, K. The developmental race between maturing host plants and their butterfly herbivore–the influence of phenological matching and temperature. J. Anim. Ecol. 2015, 84, 1690–1699. [Google Scholar] [CrossRef]
- Richardson, A.D. PhenoCam: An evolving, open-source tool to study the temporal and spatial variability of ecosystem-scale phenology. Agric. For. Meteorol. 2023, 342, 109751. [Google Scholar] [CrossRef]
- Lawrence, A. The first cuckoo in winter: phenology, recording, credibility and meaning in Britain. Glob. Environ. Change 2019, 19, 173–179. [Google Scholar] [CrossRef]
- van der Kolk, H.J.; WallisDeVries, M.F.; Van Vliet, A.J. Using a phenological network to assess weather influences on first appearance of butterflies in the Netherlands. Ecol. Indic. 2016, 69, 205–212. [Google Scholar] [CrossRef]
- Roy, B.D.; Sparks, H.T. Phenology of British butterflies and climate change. Glob. Change Biol. 2001, 6, 407–416. [Google Scholar] [CrossRef]
- Stefanescu, C.; Penuelas, J.; Filella, I. Effects of climatic change on the phenology of butterflies in the northwest Mediterranean Basin. Glob. Change Biol. 2003, 9, 1494–1506. [Google Scholar] [CrossRef]
- Chowdhury, S.; Fuller, A.R.; Dingle, H.; Chapman, W.J.; Zalucki, M. Migration in butterflies: a global overview. Biol. Rev. 2021, 96, 1462–1483. [Google Scholar] [CrossRef] [PubMed]
- Stefanescu, C.; Paramo, F.; Akesson, S.; Alarcon, M.; Avila, A.; Brereton, T.; Carnicer, J.; Cassar, L.; Fox, R.; Heliola, J.; Hill, J.; Hirneisen, N.; Kjellen, N.; Kuhn, E.; Kuussaari, M.; Leskinen, M.; Liechti, F.; Musche, M.; Regan, E.; Reynolds, D.; Roy, D.; Ryrholm, N.; Schmaljohann, H.; Settele, J.; Thomas, C.; Swaay, C.; Chapman, J. Multi-generational long-distance migration of insects: studying the painted lady butterfly in Western Palaearctic. Echography 2012, 36, 474–486. [Google Scholar] [CrossRef]
- Ubach, A.; Stefanescu, C.; Wiklund, C. Timing of mating, reproductive status and resource availability in relation to migration in the painted lady butterfly. An. Behav. 2021, 172, 145–153. [Google Scholar]
- Kassambara, A.; Kosinski, M.; Biecek, P.; Fabian, S. Survminer: Drawing Survival Curves using ‘ggplot2’. https://cran.rstudio.com/web/packages/survminer/index.html (accessed on 20th November 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. Springer: New York 2016. [CrossRef]
- Brookes, M, E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. “glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modelling”. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F.; Lohse, L. Residual Diagnostics for Hierarchical (Multi-Level/ Mixed Regression Models). https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 20th November 2023).
- Lenth, V.R.; Bolker, B.; Buerkner, P.; Gine-Vazquez, I.; Herve, M.; Jung, M.; Love, J.; Miguez, F.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, aka Least-Squares Means. https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 20th November 2023).
- Babosova, M.; Pereckova, M.; Porhajasova, J. Fixation of the Pupae of Selected Butterfly Species and Factors Affecting their Emerging. Pol. J. Environ. Stud. 2021, 30, 1521–1529. [Google Scholar] [CrossRef]
- Huang, Y.; McPherson, J.; Jiggins, C.; Montejo-Kovacevich, G. Effects of temperature on the development of Heliconius erato butterflies. University of Cambridge: United Kingdom 2022. [CrossRef]
- Koda, K.; Nakamura, H. Effects of temperature on the development and survival of an endangered butterfly, Lycaeides argyrognomon (Lepidoptera: Lycaenidae) with estimation of optimal and threshold temperatures using linear and nonlinear models. Entomol. Sci. 2021, 15, 162–170. [Google Scholar] [CrossRef]
- Stevens, D. Pupal development temperature alters adult phenotype in the speckled wood butterfly, Pararge aegeria. J. Therm. Biol. 2004, 29, 205–210. [Google Scholar] [CrossRef]
- Atkinson, D.; Sibly, R. Why are organisms usually bigger in colder environments? Making sense of a life history puzzle. Trends Ecol. Evol. 1997, 12, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Na, S.; Lee, E.; Kim, H.; Choi, S.; Yi, H. The relationship of mean temperature and 9 collected butterfly species’ wingspan as the response of global warming. J. Ecol. Environ. 2021, 45, 1–8. [Google Scholar] [CrossRef]
- Wilson, R.J.; de Siqueira, A.F.; Brooks, S.J.; Price, B.W.; Simon, L.M.; van der Walt, S.J.; Fenberg, P.B. Applying computer vision to digitised natural history collections for climate change research: Temperature-size responses in British butterflies. Methods Ecol. Evol. 2023, 14, 372–384. [Google Scholar] [CrossRef]
- Sekar, S. A meta-analysis of the traits affecting dispersal ability in butterflies: can wingspan be used as a proxy? J. Anim. Ecol. 2021, 81, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.; Davis, A.K. Populations of monarch butterflies with different migratory behaviors show divergence in wing morphology. Evolution 2010, 64, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Soule, A.; Decker, L.; Hunter, M. Effects of diet and temperature on monarch butterfly wing morphology and flight ability. J. Ins. Conserv. 2020, 24, 961–975. [Google Scholar] [CrossRef]
- Kautz, M.; Imron, A.; Dworschak, K.; Schopf, R. Dispersal variability and associated population-level consequences in tree-killing bark beetles. Mov. Ecol. 2016, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. Stress-induced color-pattern modifications and evolution of the Painted Lady butterflies Vanessa cardui and Vanessa kershawi. Zool. Sci. 2007, 24, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Solensky, M.J.; Larkin, E. Temperature-induced variation in larval coloration in Danaus plexippus (Lepidoptera: Nymphalidae). Ann. Entomol. Soc. Am. 2003, 96, 211–216. [Google Scholar] [CrossRef]
- Yamanaka, A.; Tanaka, A.; Kitazawa, C. Pupal color polyphenism regulated by temperature and photoperiod in the Asian comma butterfly, Polygonia caureum (Lepidoptera: Nymphalidae). Zool. Stud. 2012, 51, 1432–1437. [Google Scholar]
- Trott, C. Reshaping our world: Collaborating with children for community-based climate change action. Action Res. 2021, 17, 42–62. [Google Scholar] [CrossRef]
- Baker, C.; Clayton, S.; Bragg, E. Educating for resilience: parent and teacher perceptions of children’s emotional needs in response to climate change. Environ. Edu. Res. 2021, 27, 687–705. [Google Scholar] [CrossRef]
- Balestri, M.; Campera, M.; Budiadi, B.; Imron, M.A.; Nekaris, K.A.I. Active learning increases knowledge and understanding of wildlife friendly farming in middle school students in Java, Indonesia. Knowledge 2023, 3, 401–413. [Google Scholar] [CrossRef]
- Cordero, E.; Todd, A.; Abellera, D. Climate Change Education and the Ecological Footprint. Bull. Am. Meteorol. Soc. 2008, 89, 865–872. [Google Scholar] [CrossRef]
- Trott, C. Childrens constructive climate change engagement: Empowering awareness, agency, and action. Environ. Edu. Res. 2019, 26, 532–554. [Google Scholar] [CrossRef]
Figure 1.
Image displaying all 100 V. cardui larvae in individual vials.

Figure 2.
Colour code key used for phenological observations. Panel A: Black (B); panel B: Black/White (B/W); panel C: White (W).
Figure 2.
Colour code key used for phenological observations. Panel A: Black (B); panel B: Black/White (B/W); panel C: White (W).

Figure 3.
Displays the survival probability of V. cardui from 4th instar larvae to emergence under differing temperature treatments (RT-18 ̊C, 25 ̊C, 30 ̊C, 35 ̊C, 40 ̊C).
Figure 3.
Displays the survival probability of V. cardui from 4th instar larvae to emergence under differing temperature treatments (RT-18 ̊C, 25 ̊C, 30 ̊C, 35 ̊C, 40 ̊C).
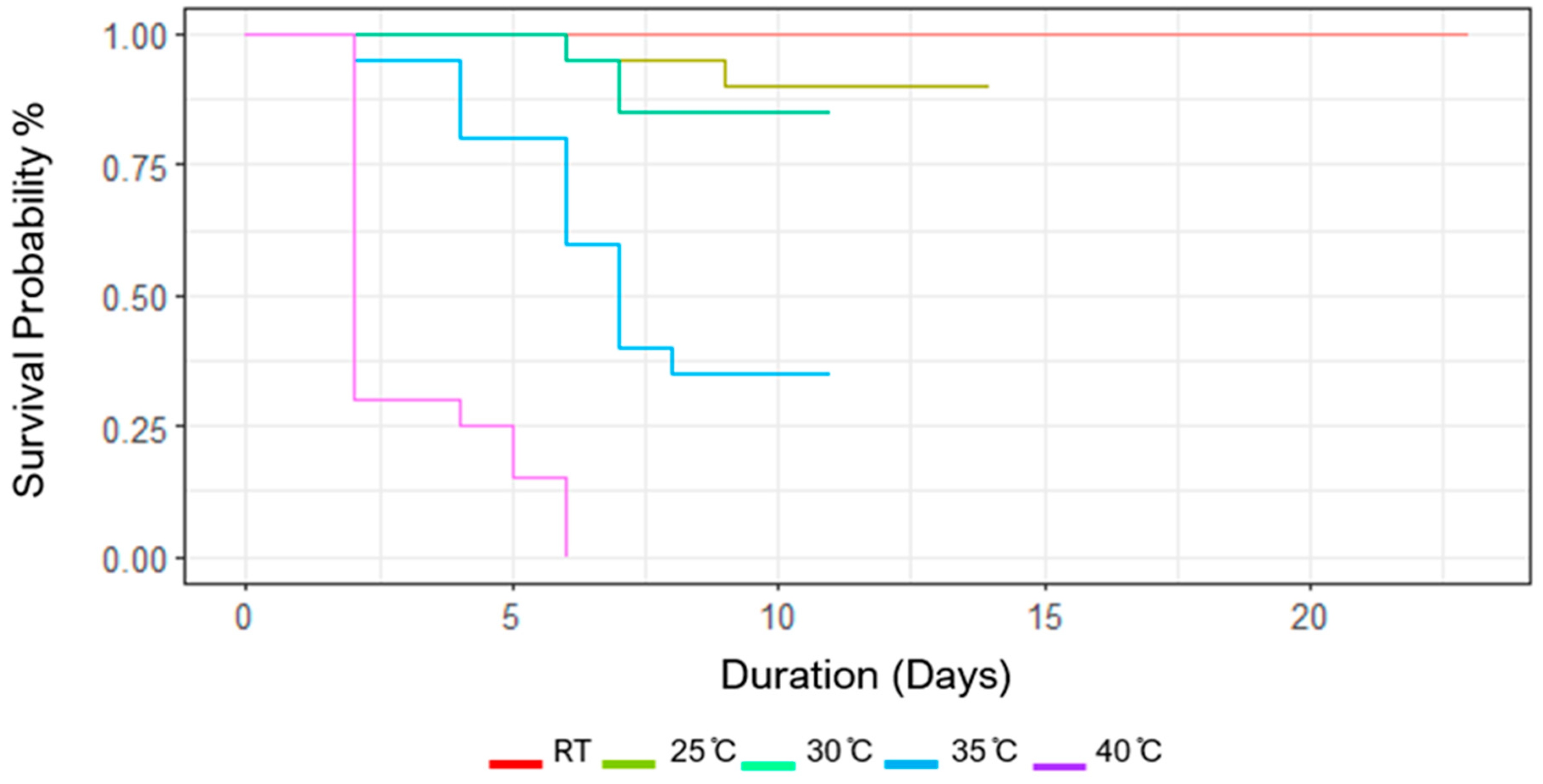
Figure 4.
Displaying colour variation of larvae in relation to temperature treatment.
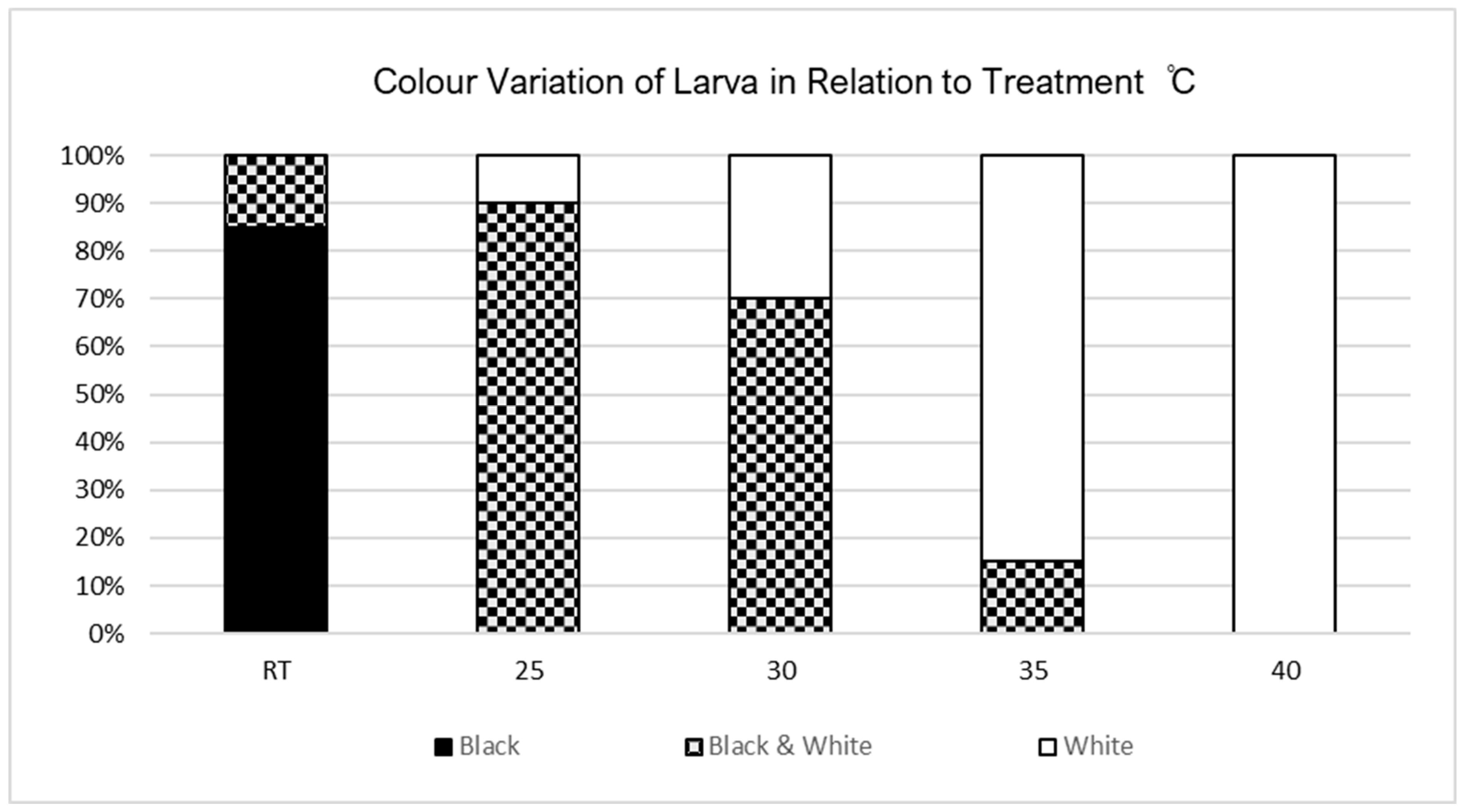
Figure 5.
Shows the mean score and standard error for each temperature gradient for all response variables, pupation duration and wingspan (mm).
Figure 5.
Shows the mean score and standard error for each temperature gradient for all response variables, pupation duration and wingspan (mm).
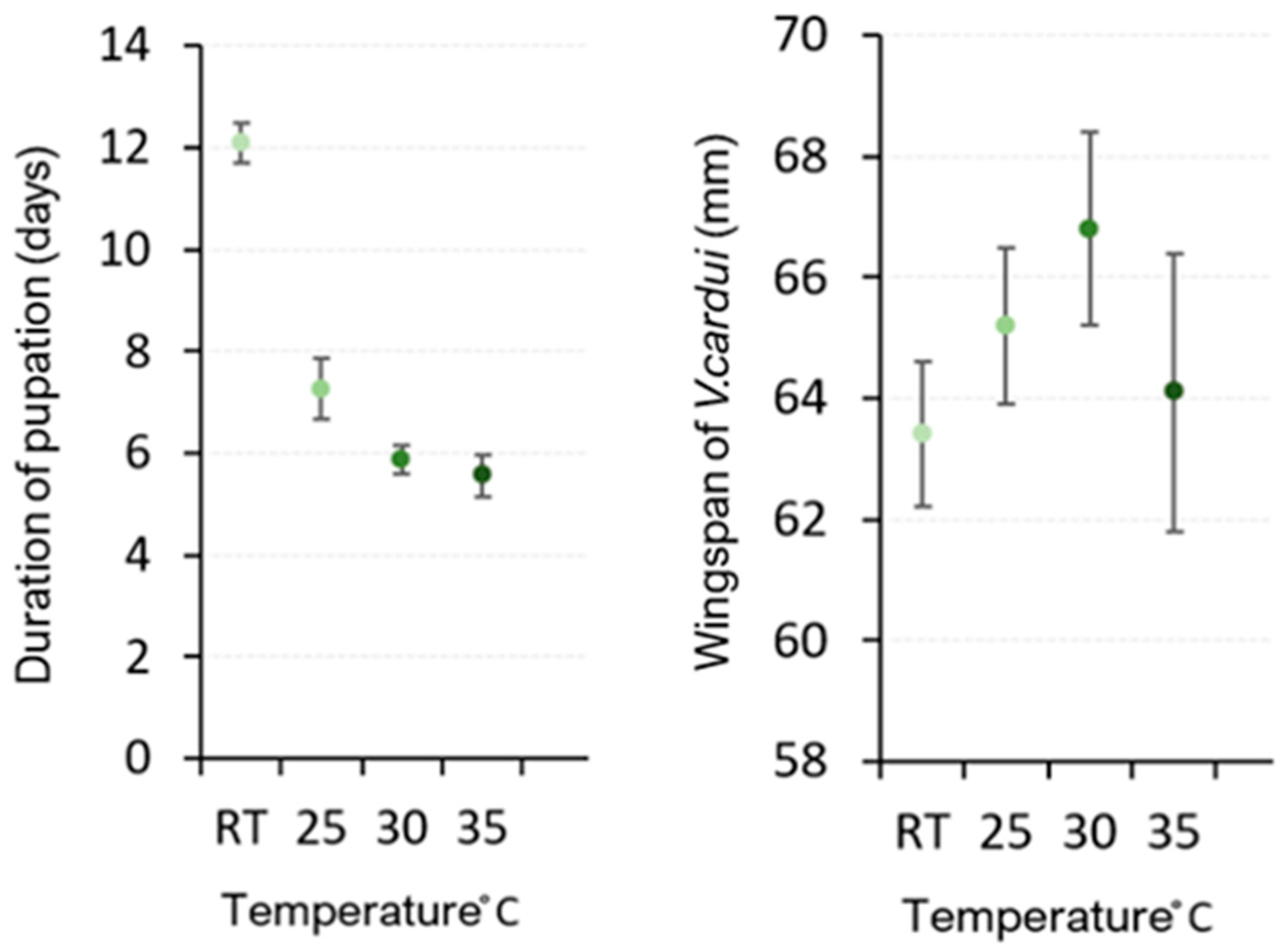
Table 1.
Per unit costings and source of materials for experimental incubator design, price relative to the time of purchase.
Table 1.
Per unit costings and source of materials for experimental incubator design, price relative to the time of purchase.
 |
Equipment | Site | Price |
| Basking clip-on spot lamp | Amazon | £19.99 | |
| Basking spotlight 50w* | Reptilush | £4.80 | |
| Habistat Dimming Thermostat | Swell UK | £55.99 | |
| 20 pack A3 white Polystyrene foam boards | Amazon | £18.99 | |
| 30cm mesh habitat net** | Insect Lore | £15.00 | |
| Total | £130.76 | ||
*for the 40C treatment a 100W spotlight was required which costed £5.10. **generally comes with insect kit when purchasing the butterflies.
Table 2.
Results of the Generalized Linear Mixed Models with the response to pupation duration and wingspan (mm) of 80 V. cardui.
Table 2.
Results of the Generalized Linear Mixed Models with the response to pupation duration and wingspan (mm) of 80 V. cardui.
| Response variable | Predictor | Estimate | Std. Error | z-value | p-value |
|---|---|---|---|---|---|
| Duration of pupation | Intercept | 2.492 | 0.013 | 190.88 | <0.0001*** |
| 25 ̊C | -0.509 | 0.021 | -23.56 | <0.0001*** | |
| 30 ̊C | -0.722 | 0.022 | -32.68 | <0.0001*** | |
| 35 ̊C | -0.778 | 0.033 | -23.53 | <0.0001*** | |
| Wingspan of adults | Intercept | 4.148 | 0.007 | 548.4 | <0.0001*** |
| 25 ̊C | 0.028 | 0.011 | 2.6 | 0.0095** | |
| 30 ̊C | 0.525 | 0.121 | 4.3 | 0.0001*** | |
| 35 ̊C | 0.012 | 0.016 | 0.7 | 0.465 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



