
Submitted:
30 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
Over the few decades, cancer-associated mortalities and morbidities were continuously increased worldwide despite sophisticated technological advancements. Pharmaceutical interventions associated with drugs exhibit a high degree of side effects and toxicities in addition to very high costs. Subsequently, to reduce or to vanish the side effects and high costs, researchers are now exploring natural bioactive compounds such as quercetin in its nanoformulations along with biologics as cargo delivery vehicles. Quercetin along with mesenchymal stromal lineage-derived extracellular vesicles (EVs) possesses an anti-cancer potential that can be explored to treat hepatocellular carcinoma (HCC). Exerting enhanced effect, nano quercetin synergistically with EVs triggers the anti-cancer mechanisms by regulating and dysregulating several signalling mechanisms including NF-κB, p53, JAK/STAT, MAPK, Wnt/β-catenin and PI3K/AKT, in addition to PBX3/ERK1/2/CDK2, and miRNAs modulation. In addition, findings regarding the potential checkpoints of anti-cancer signalling pathways were investigated that offer opportunities to develop engineered EVs incorporated with nano quercetin for the development of novel therapeutics to treat HCC in future. In this present mechanistic review, we abridged the regulation of such signalling mechanisms synergetic approach of nano quercetin and EVs. The regulatory role of EVs in the manifestation of innumerable miRNAs has also been tailed with special context to HCC.
Keywords:
nanoquercetin
; extracellular vesicles
; hepatocellular carcinoma
; therapeutics
; anticancer
; drug delivery
1. Background
Liver cancer or primary hepatic malignancy accounts for the sixth most common form of human cancer worldwide and among this 90% of the liver cancer cases exhibit hepatocellular carcinoma (HCC) [1,2]. The major risk factors for HCC include hepatitis B and C infections, fatty liver disease and excess alcohol intake [3]. Among these risk factors, hepatitis B virus infection is among the prominent risk factor for the development of HCC, which alone accounts for 50% of cases [4]. Unrelenting virological response (UVR) with the usage of antiviral drugs has significantly diminished the risk of HCC attributed to hepatitis C virus infection [5]. However, in the West, non-alcoholic steatohepatitis (NASH) attributed to metabolic disorders including diabetes mellitus and obesity is increasing at an alarming rate contributing to the aetiology of HCC [6,7]. Age is also considered to be the contributory risk factor in the progression of non-alcoholic fatty liver disease (NAFLD) related HCC. In one of the previously published studies, it was observed that patients with NAFLD-attributed HCC were more aged compared to virus-associated HCC [8]. Age-associated gut microbiota modulation in patients presenting NAFLD is also considered to be at high risk of developing HCC [9].
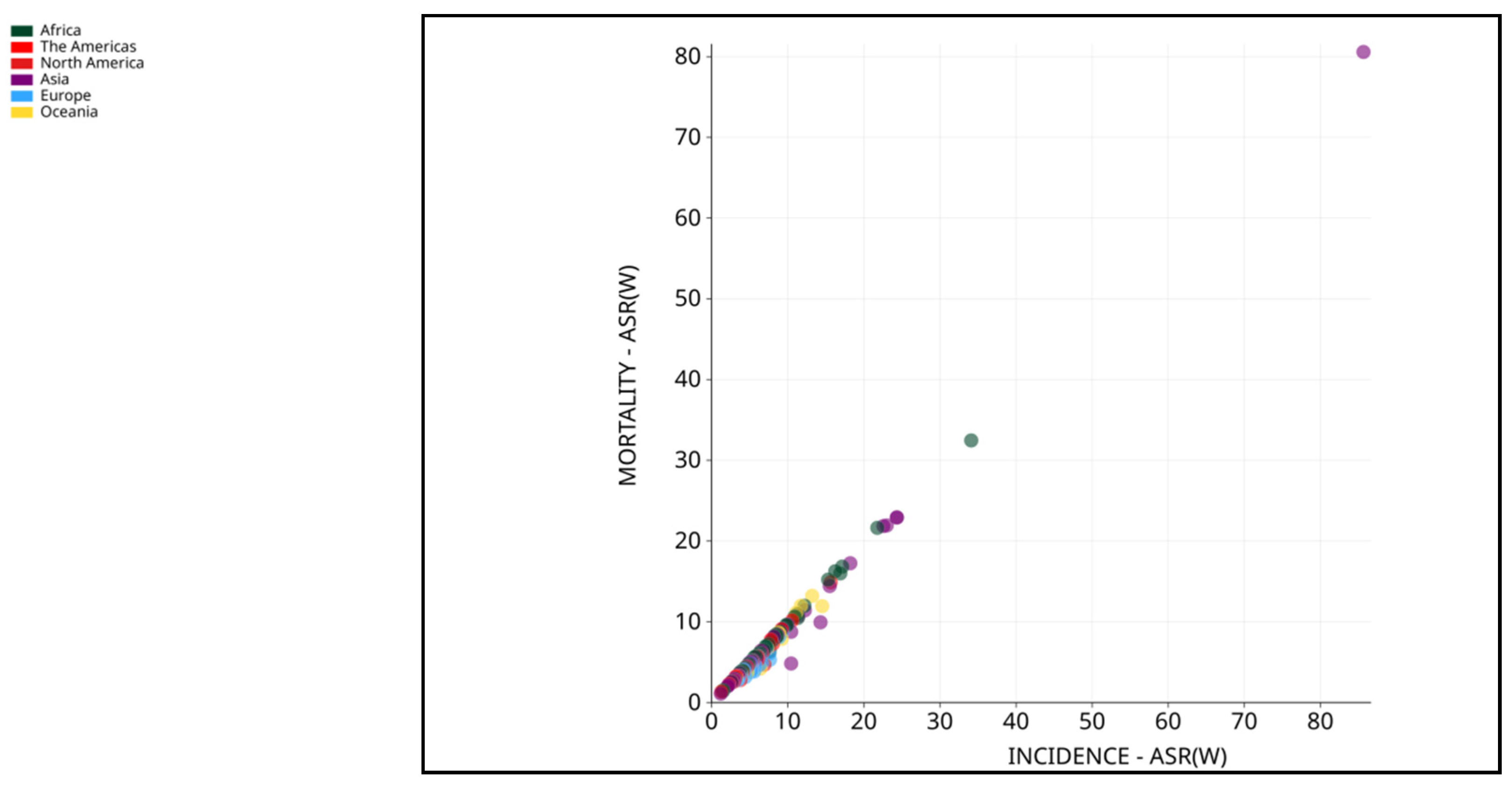
World Health Organization (WHO) considered liver cancer the prime cause of cancer-related mortalities worldwide. It is also estimated that in the year 2020, about 0.83 million people died due to it [10]. In Asia, HCC is among the most common form of liver cancer that accounts for 0.5 million deaths with 0.6 million new cases in 2020 [11,12]. Asian men demonstrated higher incidence and mortality compared to Asian women making it fourth highest incidence and second highest mortality. Moreover, among Asian women, liver cancer accounted seventh-highest incidence and sixth-highest mortality in the year 2020 [10]. A sharp decline in the average annual percent change (AAPC) in incidence rates was observed in cases of liver cancer in South Korea (-2.2%), Japan, China (-1.6%) and the Philippines (-1.7%) as documented by previously published study [12]. On the contrary, South-Western Asian countries including Israel showed a hike in AAPC [12]. In another report published by GLOBOCAN 2020, countries including Iran, Afghanistan, Qatar, Azerbaijan, Iraq, and Nepal also showed a worrying trend [13]. Studies demonstrated that eastern Asia, northern Africa and Micronesia are with highest incidence rates while the highest mortality rates were shown by eastern Asia, northern Africa and south-eastern Asia [14,15] (Figure 1, 2, and 3). With the increased incidence and mortality rates of HCC in several parts of the world, it is necessary to focus on such issues with newer technologies and interventions.
Figure 1.
Estimated age-standardized incidence rates (ASR) (worldwide) for liver cancer, both sexes and all ages, in 2020. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-map?v=2020&mode=population&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=11&type=0&statistic=5&prevalence=0&population_group=earth&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=10&group_cancer=1&include_nmsc=0&include_nmsc_other=0&projection=natural-earth&color_palette=default&map_scale=quantile&map_nb_colors=5&continent=0&show_ranking=0&rotate=%255B10%252C0%255D. (Ref [3]).
Figure 1.
Estimated age-standardized incidence rates (ASR) (worldwide) for liver cancer, both sexes and all ages, in 2020. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-map?v=2020&mode=population&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=11&type=0&statistic=5&prevalence=0&population_group=earth&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=10&group_cancer=1&include_nmsc=0&include_nmsc_other=0&projection=natural-earth&color_palette=default&map_scale=quantile&map_nb_colors=5&continent=0&show_ranking=0&rotate=%255B10%252C0%255D. (Ref [3]).
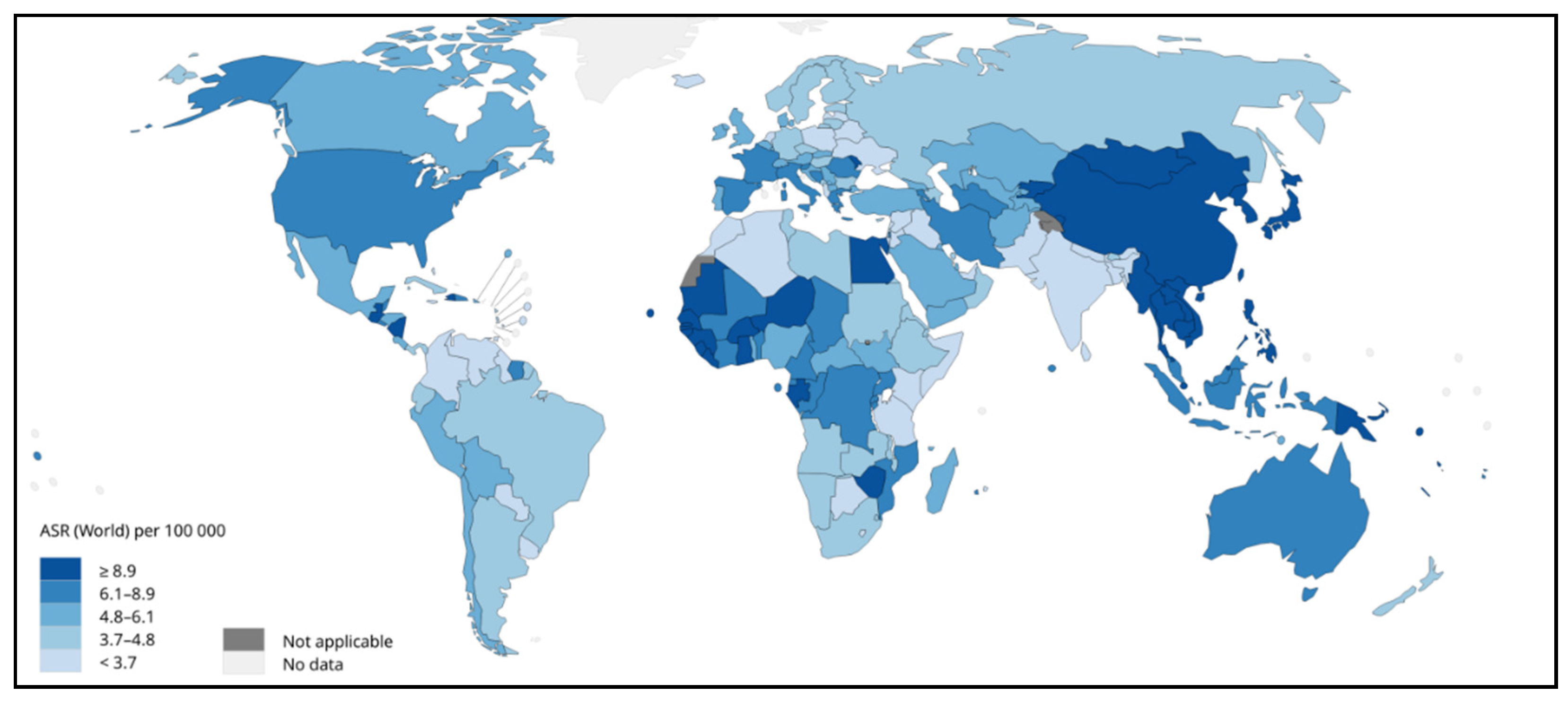
Figure 2.
Estimated age-standardized mortality rates (worldwide) in 2020 for liver cancer, both sexes and all ages. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-map?v=2020&mode=population&mode_population=regions&population=250&populations=250&key=asr&sex=0&cancer=39&type=1&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=10&group_cancer=0&include_nmsc=0&include_nmsc_other=1&projection=globe&color_palette=default&map_scale=quantile&map_nb_colors=3&continent=0&show_ranking=0&rotate=%255B10%252C0%255D (Ref 3).
Figure 2.
Estimated age-standardized mortality rates (worldwide) in 2020 for liver cancer, both sexes and all ages. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-map?v=2020&mode=population&mode_population=regions&population=250&populations=250&key=asr&sex=0&cancer=39&type=1&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=10&group_cancer=0&include_nmsc=0&include_nmsc_other=1&projection=globe&color_palette=default&map_scale=quantile&map_nb_colors=3&continent=0&show_ranking=0&rotate=%255B10%252C0%255D (Ref 3).
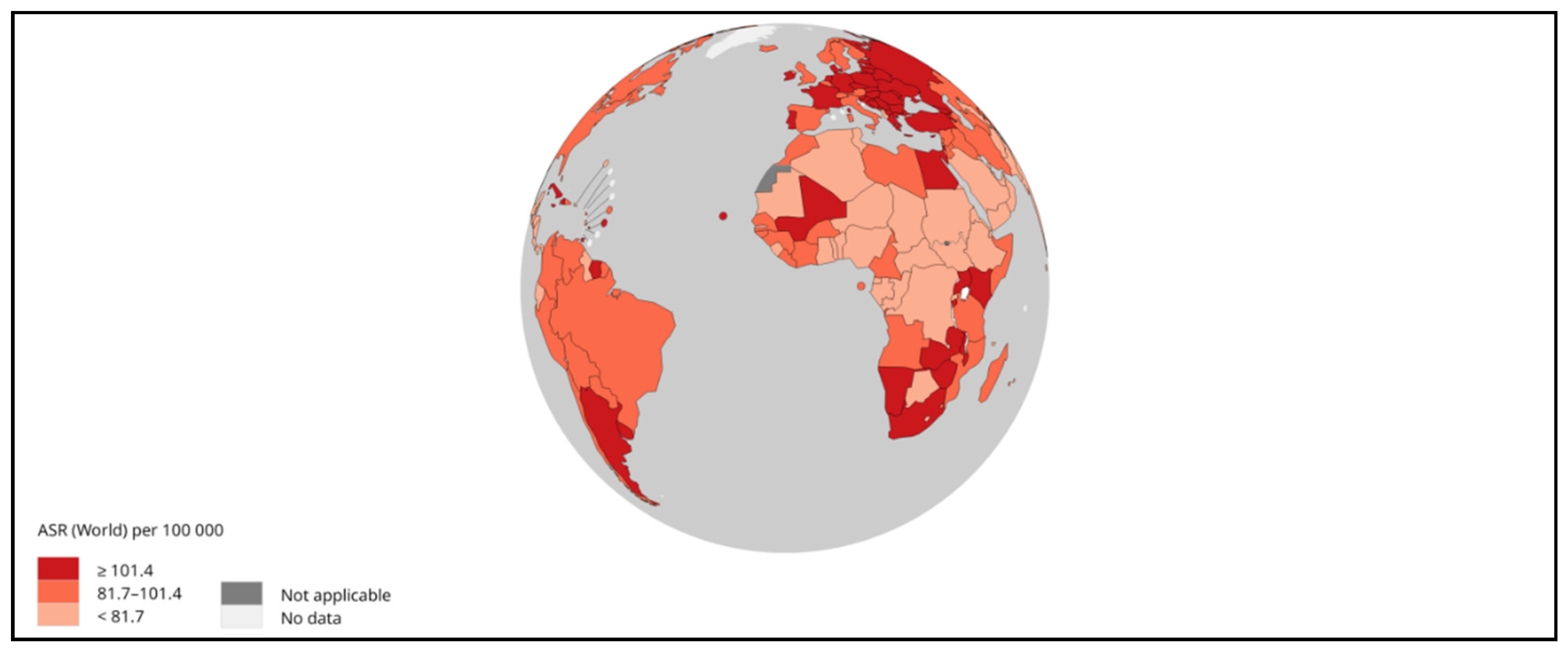
Figure 3.
Mortality-ASR (worldwide) vs. Incidence-ASR (worldwide) in 2020, for both sexes and all ages. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-scatter-plot?v=2020&mode=cancer&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=11_11&type=0&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=5&group_cancer=0&include_nmsc=0&include_nmsc_other=1&fit_to_screen=1&same_scale=1&axis_indicators=%257B%2522x%2522%253A%2522inc%2522%252C%2522y%2522%253A%2522mort%2522%257D&axis_keys=%257B%2522x%2522%253A%2522asr%2522%252C%2522y%2522%253A%2522asr%2522%252C%2522log_scale_x%2522%253Afalse%252C%2522log_scale_y%2522%253Afalse%257D (Ref 3).
Figure 3.
Mortality-ASR (worldwide) vs. Incidence-ASR (worldwide) in 2020, for both sexes and all ages. Data source: GLOBOCAN 2020. Map production: IARC (http://gco.iarc.fr/today) World Health Organization. (Copy Rights 2020). https://gco.iarc.fr/today/online-analysis-scatter-plot?v=2020&mode=cancer&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=11_11&type=0&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&nb_items=5&group_cancer=0&include_nmsc=0&include_nmsc_other=1&fit_to_screen=1&same_scale=1&axis_indicators=%257B%2522x%2522%253A%2522inc%2522%252C%2522y%2522%253A%2522mort%2522%257D&axis_keys=%257B%2522x%2522%253A%2522asr%2522%252C%2522y%2522%253A%2522asr%2522%252C%2522log_scale_x%2522%253Afalse%252C%2522log_scale_y%2522%253Afalse%257D (Ref 3).
Several staging approaches were fabricated to classify HCC including Hong Kong Liver Cancer (HKLC), cancer of the liver Italian program (CLIP), Okuda, Barcelona clinic liver cancer (BCLC) (Figure 4), American Association for the Study of Liver Diseases (AASLD) [16,17]. Among these staging approaches, the latter two were widely used worldwide. Some other staging regimes involve the classification of HCC using molecular genetics, metabolism, immunological properties and chromosomal arrangements [18].
Current interventional approaches for HCC include liver resection, transplantation, transarterial therapy, the implication of tyrosine kinase inhibitors (systemic therapy), and local ablative therapy. Moreover, along with conventional therapies, several drugs including sorafenib, lenvatinib, atezolizumab and bevacizumab along with chemotherapeutic agents such as doxorubicin (DOX) showed restricted effects along with associated extrinsic and intrinsic drug resistance [19].
Despite multiple pharmacological interventions, treatment of advanced-stage HCC does not fulfil the standard health outcomes. Such unsatisfied outcomes may be attributed to several reasons including drugs associated adverse effects, low bioavailability, high toxicity, non-specific delivery of pharmacological agents, high cost at large-scale production, immune complications, and anaphylactic responses [20]. To overcome such limitations, researchers are now looking to explore possibilities of using nanoparticles and extracellular vesicles (EVs) as drug delivery vessels for the treatment of almost every cancer. The present review will focus on exploring the potential of EVs and quercetin nanoparticles (Qnps) for the treatment of HCC. Moreover, the mechanistic pathways inhibiting the pathogenesis of HCC with Qnps and EVs are also explored. We further examined the evidence supporting the use of biological molecules including EVs and quercetin for treatment regimes for HCC. Finally, we performed a critical analysis of current clinically approved drugs/immunotherapies for HCC and ongoing preclinical/clinical trials using EVs and Qnps as anti-cancer agents in patients with HCC.
2. Pathophysiology of HCC
The pathophysiology of HCC exhibits complex multifactor mechanisms. HCC progression and hepatocytes' malignant transformation depend on the interplay between various factors including genetic predisposition, viral and non-viral elements, the severity of liver disease and the cellular microenvironment at its early stage. It was seen that nearly 80% of liver cirrhotic patients develop HCC attributed due to molecular alterations [2]. Viral elements include etiological infections associated with HCV and HBV while the non-viral elements include alcohol consumption, NASH, use of aflatoxin, tobacco and aristolochic acid has been identified as a trigger of cancer mechanisms in the liver [21]. In addition to aforesaid factors, some specific immune and molecular causes were identified as an initiator of HCC [21]. In this respect, studies of such molecular and immunological checkpoints are necessary to understand the onset, progression and treatment using biopharmaceuticals for these targets. Some of the major checkpoints were extensively studied elsewhere [2,22,23].
3. Molecular triggers of hepatocellular carcinoma
In patients with liver cirrhosis, the neoplasm advances through a sequential cascade of histopathological modulations ultimately initiating HCC. Histomorphological characteristics of HCC include highly vascularized tumours with prominent acinar and wide trabeculae along with loss of Kupffer cells and reticulin network [24]. In advanced HCC, tumours were found to be encapsulated with the presence of septae that are positive for CD34 and α-smooth muscle actin (SMA). Studies have found that mature hepatocytes are the primary cells responsible for HCC origin and progression in addition to liver stromal cells [23,25,26]. Studies in past demonstrated that repetitive stress to regenerating hepatocytes triggers genetic lesions that initiate transformation and oncogenesis progression [27]. The study observed that alterations in cyclin-A2 or E1 proteins of the cell cycle favour the progression of HCC, especially in non-cirrhotic patients which is further mediated by activation of E2F and ATR transcriptional pathways along with inactivation of RB1 and PTEN [27].
In patients with NASH, CD8+PD1+ T cells promote hepatocyte death and thereby favour the micro-environment for HCC pathogenesis and progression [28]. On contrary, somatic, genomic and epigenetic modulations also trigger the HCC. A study showed the single nucleotide polymorphisms (SNP) of PNPLA3 (rs738409), TM6SF2 (rs585542926) and HSD17B13 (rs72613567) predispose to liver carcinogenesis that increases the probability of HCC [29]. Genotoxic compounds including aflatoxin B1 and aristolochic acid (promote inversion of T to A) were known to trigger somatic mutations that again increased the risk of HCC progression [30].
4. Checkpoint targets of hepatocellular carcinoma
Hepatocellular carcinoma pathogenesis is triggered by several mechanistic pathways that involved numerous checkpoints and can be explored as targeted therapies in HCC. The following checkpoints were considered to play a pivotal role in HCC.
4.1. Wnt–β-catenin signalling
CTNNB1
A CTNNB1-related active mutation is a major canonical component of the Wnt signalling pathway and is exhibited in nearly 11-41 % of patients with liver cancer [32,33,34]. CTNNB1 is actively involved in the synthesis of actin cytoskeleton responsible for halting cell division [35]. Indeed mutations of CTNNB1 were reported to be significantly correlated with TERT promoter, NFE2L2, MLL2, ARID2, and APOB [36,37]. Studies related to human HCC found that CTNNB1 mutations concurrently occurred with the upregulation of Met, Myc, or Nrf2 [38,39,40]. Drugs including sorafenib and gamma-secretase inhibitors were also studied as effective targets indulging the CTNNB1 mechanism [30,41].
Adenomatous Polyposis Coli (APC)
Human APC mutations originated within the central core region of the open reading frame (ORF) commonly known as MCR (mutation cluster region) that produces truncated proteins [42]. Moreover, this event triggers the loss of several factors including β-catenin binding sites (20R), nuclear localization sequences (NLS), axin binding sites (ABS) and C-terminal basic domain (CTBD), which are responsible for cytoskeletal interfaces. Sporadic APC mutations are considered to be the contributory factor for tumorigenesis. Mutations in APC significantly modulate the Wnt–β-catenin signalling, that in turn initiates the origin and progression of HCC.
AXIN1
AXIN1 mutations were found to be associated with nearly 5-19% of patients with liver cancer [32,33]. AXIN1 negatively regulate the Wnt/β-catenin signalling by modulating the expression of β-catenin [43]. A study found that upregulated expression of wild-type AXIN1 intimidated the cellular proliferation in HCC along with induction of programmed cell death, and thereby can be used as a molecular target to treat HCC [43]. In continuation with this study, another author used adenovirus-mediated gene transfer of AXIN1 and initiated HCC cell apoptosis [44]. AXIN was found to be an inhibitor of tankyrase 1 and 2 through XAV 939 and hence can be used as a novel therapeutic target within Wnt signalling [45].
4.2. Telomere maintenance
TERT
TERT promoter mutations were known to be associated with the pathogenesis of HCC. It was reported by previous studies that the TERT promoter showed mutation at the upstream of ATG translation start site at positions -124 (G>A) and -146 (G>A) [46,47]. Mutations in TERT promoter sequences produce a de novo consensus binding region for the ETS (E-twenty-six) transcription factor that further triggers the increased production of TERT proteins that in turn attenuate the telomerase activity and length [48,49,50]. A recent study reported mutation of the TERT promoter in HCC patients at -297 (C>T) upstream of the ATG translational region generating an AP2 consensus sequence [51]. It has been found that the protein expressed by RB/E2F gene regulates the activity of the TERT promoter and contributed to liver cancer [52]. A past study also showed that TERT gene activation was triggered by the binding of RNA-binding fox-1 homolog 3 (RBFOX3) with AP2β that in turn activates telomerase and promotes HCC [53]. In another study, it was found that SP1 and YAP1 activate the TERT gene expression in the HepG2 cell line [54].
4.3. Cell cycle regulation
TP53
Nearly 13-48% of patients with liver cancer exhibit TP53 mutations [32,33]. TP53 gene suppresses the tumours by arresting the growth and apoptosis of cancerous cells [32]. A previously published study from West demonstrated that mutations in the TP53 gene especially in patients with HCC are associated with poor health outcomes and prognosis [33]. In another study, it was found that non-inflamed tumours exhibit T-cell exclusion mediated either through TP53 gene mutations also known as an intermediate class [55]. Authors of another study concluded that TP53 mutations especially known as hot spot mutations at R249S and V157F were associated with poor outcomes and prognosis of patients with HCC [56].
CDKN2A, CCND1, FGF3, FGF4 or FGF19
It seems that nearly 8% of the HCC cases exhibit CDKN2A deletions mutation [36]. It was known that CDKN2A is also a tumour suppressor gene that triggers the arresting of the cell cycle at the G1 and G2 phases and can act as a potential checkpoint for HCC therapy. Moreover, it also inhibits the expression of CDK4/6 and MDM2, which are responsible for oncogenic action [64]. A previous study reported that loss of CDKN2A in patients with HCC attenuates the rise of CDK4/6 inhibitors in the advanced stage [65].
In liver cancer, it was found that nearly 5-7% and 4-6% of the patients exhibit mutations of CCND1 and FGF3, FGF4, or FGF19 respectively [36,66]. It was studied that augmentation of CCND1, FGF3, FGF4, or FGF19 in patients with resected HCC is associated with poor prognosis and outcome [36]. A plausible study showed suppression of 11q13.3 amplicon by anti-FGF19 antibody along with anti-sense RNA mediated knockdown of FGF19 or CCND1 [67].
4.4. Oxidative stress
Hepatocytes exhibit numerous fatty acids that trigger oxidative stress along with endoplasmic reticulum (ER) stress. Furthermore, these stresses cause cellular damage and inflammation [68]. In one of the animal studies, it was found that ER stress can cause NASH-triggered HCC due to the activation of several pathways including NF- κB and TNF [69]. One of the previously published studies suggested that ER stress is mediated through the activation of hepatosteatosis and secondly due to the promotion of SREBP1 activation and increasing the process of lipogenesis [69]. ER stress in association with steatosis generates ROS in hepatocytes that are the primary cause of oxidative stress and oncogenic mutations. These ROS triggers the lipotoxic death of hepatocytes and thereby activate the macrophages. Further release of TNF-α also triggers the activation of chemokines and growth factors that attenuate the inflammatory microenvironment of hepatocytes [69]. In addition, ROS production induces DNA damage due to mitochondrial dysfunction and hence contributes to the pathophysiology of HCC in humans [70].
In the previously published study, it was found that mTORC2 activations within the hepatocytes trigger the concentration of sphingolipid glucosylceramide and thereby increased ROS generation which in turn leads to HCC [71]. Impaired cholesterol metabolism also triggers the pathophysiology of HCC [72]. A clinical study demonstrated the trend of HCC in patients and found that NASH posed a higher risk for HCC pathogenesis compared to NAFLD [73].
5. Potential anti-cancer mechanism of nano quercetin in HCC
Quercetin belongs to the naturally occurring flavonoid class and is widely known for its therapeutic effect including pro-apoptotic, proliferative and antioxidant [74]. It is a well-known inhibitor of casein kinase-2α that is responsible for HCC pathogenesis [75]. Some studies also decipher the role of casein kinase-2α in the apoptosis mechanism and activation of death receptors pathways [76,77,78]. Moreover, studies showed that nanoformulation of quercetin improves the mechanistic action and therapeutic properties compared to pure quercetin form due to several limitations including less bioavailability, slow absorption and short action. Therefore, nano quercetin showed enhanced anti-cancer activities by significantly modulating the signalling pathways as shown below:-
5.1. Wnt/β-catenin signaling pathway
The wnt/β-catenin signalling pathway regulates several biological processes including cell differentiation, proliferation, migration, and APC/β-catenin/Tcf pathway [79]. In another in-vitro study, it was found that quercetin showed inhibition of SOX2, Nanog, and Oct4 expression along with β-catenin nuclear translocation that in turn resulted in downregulated expression of β-catenin-dependent transcriptional activity [80]. In another study, it was found that 20µM quercetin showed reduced viability through regulating DKK1, 2 and 3 proteins that in turn act as checkpoints of Wnt signalling [81].
5.2. PI3K/AKT pathway
PI3K mediates the translocation process of AKT to the plasma membrane and regulates the mechanism of cell cycle progression, differentiation, cell survival and cell proliferation [82]. PI3K/AKT is also observed to regulate the expression of Bax (Bcl-2 protein family member) which is responsible for the anti-apoptotic mechanism [83]. Authors from the previously published study reported the anti-cancer activity of quercetin against HCC1937 PTEN cancer cell lines through regulation of AKT/PKB phosphorylation [84].
In one of the studies, it was suggested that flavonoids directly or indirectly inhibit the mTOR signalling mechanism [85]. It is known that PDK1 is considered to be a major kinase necessary for the development of the mammalian cell. It is found that in the cancer microenvironment, the degree of phosphorylation of AKT kinase at Thr-308 was significantly increased [86]. Another study found that quercetin triggers the down-regulation of phosphorylation of PDK1 and is hence considered to be the therapeutic target of quercetin and regulatory checkpoints at Ser-473 and Thr-308 [87]. Quercetin is considered to be a broad-spectrum inhibitor of PI3K/AKT1/2 as found by a previous study [88]. Hence, it was considered that quercetin inhibits the AKT1/2 by acting directly by inducing Ser/Thr kinase activity and down-regulating the PI3K.
5.3. JAK/STAT signalling
JAK/STAT signalling mechanism regulates the immune microenvironment, cell death, proliferation, division and tumour growth. It is known that JAK/STAT pathway is controlled by ERK, MAPK and PI3 kinase. It was reported that carcinomas were associated with the deregulation of the JAK/STAT pathway [89]. Qin and their coworkers observed the role of quercetin on the JAK/STAT pathway and observed that MGC-803 cells were arrested at the G2/M stage of the cell cycle mediated through p-STAT3 signalling and also reduces the expression of leptin along with its corresponding receptors [90]. It was also reported that quercetin inhibits the IL-6-triggered glioblastoma cell migration, proliferation and growth by regulating the STAT-3 signalling mechanism mediated through reduced expression of GP130 and JAK1 [91].
Quercetin is known to modulate apoptosis through activation of caspase 3, 8 and PARP cleavage that enable the cell to arrest in the sub-G0/G1 phase of the cell cycle along with reduction of p-JAK1, MMP-9 and p-STAT3 expression [92]. Authors from previous studies claimed that quercetin regulates the apoptosis and autophagy mechanism through the expression of caspase-3 that is further inhibited by JAK2 along with cyclin D1 and mTOR that in turn suppressed STAT3/5 signalling mechanism [93,94]. Moreover, quercetin was found to show reduced proliferation potential of HCC along with an increased rate of cellular apoptosis due to regulation of the cell cycle through the expression of CyclinB1 protein [95]. CyclinB1 is a cell cycle protein which is synthesized in S and G2/M phases. Therefore it is believed that quercetin inhibits the cell cycle at the G2/M phase along with triggering apoptosis.
5.4. The MAPK signalling
Mitogen-Activated Protein Kinase (MAPK) exhibits three primary classes of kinases including ERKs, JNK/SAPK and p38s. It is considered that MAPK 14,7 and 12 regulate cellular proliferation, gene expression, differentiation, growth, mitosis and apoptosis [96]. In addition, one study done on SMMC7221 cells found that quercetin suppresses the proliferation, and reduces the lipopolysaccharide-initiated oxidation along with inhibition of the MAPK signalling pathway [97]. It was also found that quercetin significantly suppresses the activity of p38 MAPK in the fibrotic liver of rats [98]. It was found that isoquercetin activates the caspases 3, 8 and 9 that in turn significantly increases the apoptosis and triggers the JNK phosphorylation through suppression of ERK and p38 MAPK as shown in Figure 5 [99].
5.5. NF-ĸB, p53 and apoptotic signaling
Quercetin triggers the stimulation of 5-fluorouracil-initiated apoptosis in a p53-dependent manner [100]. A similar study also observed that quercetin and p53 work in a synergistic manner [100]. In another study, quercetin along with doxorubicin down regulates the Bcl-xl in a p53-dependent phase [101]. Quercetin was found to promote cell death-associated gene expression including p53 along with downregulation of AKT and Bcl-2 expression [102]. It was also found that quercetin suppresses the mTOR expression simultaneously activation of p53, Sestin-2 via AMPK. In a relevant study performed using nano-quercetin of HepG2 cells it was observed that it activates the p53-ROS crosstalk and triggers apoptosis along with modification at the epigenetic level and cell cycle arrest at the sub-G phase [103]. Another study showed that quercetin activates the p21, p53 and GADD45 signalling mechanism along with simultaneous suppression of JNK mediated through Foxo3a [104].
6. Protective mechanism of EVs in HCC
Extracellular vesicles are known to enhance the anti-cancerous potential by hampering several signalling pathways involved in the metastasis of HCC. In one of the previously published studies, it was found that the Vps4A level is higher in EVs derived from HCC which inhibits the PI3K-Akt signalling pathway that in turn inhibits the HCC progression and metastasis [105]. EVs also served as a mediator in the regulation of intracellular micro-RNA (miRNAs). In one of the studies it was found that Vps4A exhibits two oncogenic miRNAs (miR-27b-3p and miR-92a-3p) and that is found to be upregulated in SMMC-Vps4A [105].
In addition to this SMMC-Vps4A also possesses miR-193a-3p, miR-320a, and miR-132-3p as tumour suppressor miRNAs [105]. Similar study further detected six tumor suppressor miRNAs (miR-122-5p, miR-33a-5p, miR-34a-5p, miR-193a-3p, miR-16-5p, and miR-29b-3p) that showed upregulated trend [105]. The findings of this study were supported by the other authors and they found the role of tumour suppressor miRNAs miR-122-5p, miR-33a-5p, miR-34a-5p, miR-16-5p, and miR-29b-3p in HCC [106]. The authors of a previously published study performed western blotting and found that overexpression of Vps4A leads to the inactivation of the PI3K/Akt signalling pathway that also modulates the miRNAs [105]. So, it was concluded that Vps4A showed a therapeutic target against HCC in miRNAs dependent and independent manner and can be explored as checkpoints for the treatment of patients with HCC.
Another study found that expression of SENP3-EIF4A1 and SENP3-EIF4A1 in secretory EVs suppress the HCC proliferation through miR-9-5p mediated action of ZFP36 [107]. Moreover, IncRNA 85 controls the cancer cell invasion by acting on miR-324-5p by regulating the expression of MMPs, ETS1 and SP1 in HCC [108]. EVs containing miR-320a showed a protective effect against HCC through suppression of the PBX3/ERK1/2/CDK2 signaling pathway [109].
Mesenchymal stromal cells (MSCs) derived EVs are known to exhibit anti-cancer properties and can be explored for the treatment of HCC. A previous study showed that umbilical cord-derived MSCs significantly improved the anti-tumour response of NKT cells in liver cancer by inhibiting oxidative stress [110]. In another study, it was found that miR-122 provides an anti-cancer effect against HCC by suppressing the PI3-K/Akt signaling pathway as shown in Figure 6 [111]. In another study, it was found that fibroblast-derived EVs exhibit less quantity of miR-320a and thus inhibit HCC by suppressing the MAPK signalling pathway [109]. Another study claimed that fibroblast-derived EVs were rich in miR-150-3p and exhibit anti-cancer properties against HCC [112]. Some studies found that miR-195 presence in fibroblast-derived EVs suppresses the activation of VEGF, CDC42, CDK1, CDK4, CDK6, and CDC25 and is considered to be a new therapeutic target for HCC [113,114]. In another study, mi331-3p also inhibits the progression of HCC through the regulation of BAK1 [115].
7. Challenges and perspectives of anti-cancer EVs biopharmaceuticals
In the last decades, the unrelenting progress of biologics encourages to development of a thorough understanding and technological advancement of biopharmaceutical manufacturing procedures. This sharp evolution has deepened the interest of the biopharmaceutical industries in process analytical technology (PAT), which is known as a system for designing, analyzing and controlling the manufacturing of the products along with ensuring the final quality of the product [116]. Nanoparticles are well known to deliver many biologics including proteins, peptides and antibodies. However, such particles face severe challenges including physiological barriers, fast wash-off from targeted sites, poor permeation-retention effect etc. With the spent of time, technological advancements help researchers to overcome several hurdles with the advent of extracellular vesicles. EVs if compared to synthetic drug delivery nanomaterial exhibits natural site targeted features along with improved stability, biocompatibility, and increased bioavailability. Therefore, EVs are considered to be the biggest opportunity for the biopharmaceutical industry to be used as drug/nanoparticle delivery vehicles. Although substantial breakthroughs were fabricated using these engineered EVs as anti-cancer therapy still some challenges may hinder the path to making a bench to bedside products. The complex structure of EVs is associated with a high degree of inconsistency that might affect the therapeutic properties. Moreover, large-scale isolation and purification approaches of EVs still compromised their yield. Researchers nowadays concentrate their research on testing customized EVs in preclinical animal models but data is still lacking in clinical trials. Cargo loading efficiency is still an un-addressable issue and needs serious attention. Summing up the issues of biosafety, bioavailability, biocompatibility, and biostability are some of the peculiar challenges for future clinical translational research.
Conclusions
Quercetin is a polyphenolic flavonoid exhibiting anti-cancerous features that exert its therapeutic mechanism in hepatocellular carcinoma through dysregulation of several signalling mechanisms including PI3K/AKT, NF-κB, P53, Wnt/β-catenin, MAPK, JAK/STAT and Hedgehog pathway. Moreover, quercetin is known to modulate several intracellular signalling biologics including TNF-α, Bax, Bcl-2, caspases, and VEGF. The anticancer potential of quercetin was extensively studied in various cancers including hepatocellular carcinoma. However, the majority of the research was evident in preclinical studies. Studies are lacking in demonstrating clinical trials. EVs derived from mesenchymal lineages were considered to trigger anti-cancerous effects through several miRNAs and IncRNAs. Not a single study was conducted in past that demonstrated the synergistic effect of quercetin and mesenchymal stem cells derived EVs at the clinical trial phase. There are significantly high expectations of such phase III trials focusing on all stages of HCC.
Author Contributions
AR and GBJ: Conceptualization, Methodology, Software AR.: Data curation, Writing—Original draft preparation. AR: Visualization, Investigation. GBJ: Supervision.: AR: Software, Validation.: GBJ: Writing—Reviewing and Editing.
Funding
None.
Ethics approval and consent to participate
Not applicable.
Consent for publication
The consent for publication has been given by all authors.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the Brain Pool Program funded by the Ministry of Science and ICT through the National Research Foundation of Korea (Grant Number 2022H1D3A2A01096346) for supporting this research. We also thank Editage (www.editage.co.kr) for English language editing.
Conflicts of Interest
The authors declare no competing interests.
Abbreviations
| HCC | Hepatocellular carcinoma |
| UVR | Unrelenting virological response |
| NASH | Non-alcoholic steatohepatitis |
| NAFLD | non-alcoholic fatty liver disease |
| EVs | Extracellular vesicles |
| AAPC | Average annual percent change |
| HKLC | Hong Kong Liver Cancer |
| CLIP | Cancer of the liver Italian program |
| BCLC | Barcelona clinic liver cancer |
| AASLD | American Association for the Study of Liver Diseases |
References
- Global Burden of Disease Cancer Collaboration; Fitzmaurice C, Allen C, Barber RM, Barregard L, Bhutta ZA, Brenner H, Dicker DJ, Chimed-Orchir O, Dandona R, Dandona L, Fleming T, Forouzanfar MH, Hancock J, Hay RJ, Hunter-Merrill R, Huynh C, Hosgood HD, Johnson CO, Jonas JB, Khubchandani J, Kumar GA, Kutz M, Lan Q, Larson HJ, Liang X, Lim SS, Lopez AD, MacIntyre MF, Marczak L, Marquez N, Mokdad AH, Pinho C, Pourmalek F, Salomon JA, Sanabria JR, Sandar L, Sartorius B, Schwartz SM, Shackelford KA, Shibuya K, Stanaway J, Steiner C, Sun J, Takahashi K, Vollset SE, Vos T, Wagner JA, Wang H, Westerman R, Zeeb H, Zoeckler L, Abd-Allah F, Ahmed MB, Alabed S, Alam NK, Aldhahri SF, Alem G, Alemayohu MA, Ali R, Al-Raddadi R, Amare A, Amoako Y, Artaman A, Asayesh H, Atnafu N, Awasthi A, Saleem HB, Barac A, Bedi N, Bensenor I, Berhane A, Bernabé E, Betsu B, Binagwaho A, Boneya D, Campos-Nonato I, Castañeda-Orjuela C, Catalá-López F, Chiang P, Chibueze C, Chitheer A, Choi JY, Cowie B, Damtew S, das Neves J, Dey S, Dharmaratne S, Dhillon P, Ding E, Driscoll T, Ekwueme D, Endries AY, Farvid M, Farzadfar F, Fernandes J, Fischer F, G/Hiwot TT, Gebru A, Gopalani S, Hailu A, Horino M, Horita N, Husseini A, Huybrechts I, Inoue M, Islami F, Jakovljevic M, James S, Javanbakht M, Jee SH, Kasaeian A, Kedir MS, Khader YS, Khang YH, Kim D, Leigh J, Linn S, Lunevicius R, El Razek HMA, Malekzadeh R, Malta DC, Marcenes W, Markos D, Melaku YA, Meles KG, Mendoza W, Mengiste DT, Meretoja TJ, Miller TR, Mohammad KA, Mohammadi A, Mohammed S, Moradi-Lakeh M, Nagel G, Nand D, Le Nguyen Q, Nolte S, Ogbo FA, Oladimeji KE, Oren E, Pa M, Park EK, Pereira DM, Plass D, Qorbani M, Radfar A, Rafay A, Rahman M, Rana SM, Søreide K, Satpathy M, Sawhney M, Sepanlou SG, Shaikh MA, She J, Shiue I, Shore HR, Shrime MG, So S, Soneji S, Stathopoulou V, Stroumpoulis K, Sufiyan MB, Sykes BL, Tabarés-Seisdedos R, Tadese F, Tedla BA, Tessema GA, Thakur JS, Tran BX, Ukwaja KN, Uzochukwu BSC, Vlassov VV, Weiderpass E, Wubshet Terefe M, Yebyo HG, Yimam HH, Yonemoto N, Younis MZ, Yu C, Zaidi Z, Zaki MES, Zenebe ZM, Murray CJL, Naghavi M. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2017 Apr 1;3(4):524-548; Erratum in: JAMA Oncol. 2017 Mar 1;3(3):418. [CrossRef]
- Llovet JM, Kelley RK, Villanueva A, Singal AG, Pikarsky E, Roayaie S, Lencioni R, Koike K, Zucman-Rossi J, Finn RS. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021 Jan 21;7(1):6. [CrossRef]
- International Agency for Research on Cancer. GLOBOCAN 2018. IARC https://gco.iarc.fr/today/online-analysis-map?v=2020&mode=population&mode_population=continents&population=900&populations=900&key=asr&sex=0&cancer=11&type=0&statistic=5&prevalence=0&population_groupearth&color_palette=default&map_scale=quantile&map_nb_colors=5&continent=0&rotate=%255B10%252C0%255D (Access on 03/03/2023).
- Global Burden of Disease Liver Cancer Collaboration; Akinyemiju T, Abera S, Ahmed M, Alam N, Alemayohu MA, Allen C, Al-Raddadi R, Alvis-Guzman N, Amoako Y, Artaman A, Ayele TA, Barac A, Bensenor I, Berhane A, Bhutta Z, Castillo-Rivas J, Chitheer A, Choi JY, Cowie B, Dandona L, Dandona R, Dey S, Dicker D, Phuc H, Ekwueme DU, Zaki MS, Fischer F, Fürst T, Hancock J, Hay SI, Hotez P, Jee SH, Kasaeian A, Khader Y, Khang YH, Kumar A, Kutz M, Larson H, Lopez A, Lunevicius R, Malekzadeh R, McAlinden C, Meier T, Mendoza W, Mokdad A, Moradi-Lakeh M, Nagel G, Nguyen Q, Nguyen G, Ogbo F, Patton G, Pereira DM, Pourmalek F, Qorbani M, Radfar A, Roshandel G, Salomon JA, Sanabria J, Sartorius B, Satpathy M, Sawhney M, Sepanlou S, Shackelford K, Shore H, Sun J, Mengistu DT, Topór-Mądry R, Tran B, Ukwaja KN, Vlassov V, Vollset SE, Vos T, Wakayo T, Weiderpass E, Werdecker A, Yonemoto N, Younis M, Yu C, Zaidi Z, Zhu L, Murray CJL, Naghavi M, Fitzmaurice C. The Burden of Primary Liver Cancer and Underlying Etiologies From 1990 to 2015 at the Global, Regional, and National Level: Results From the Global Burden of Disease Study 2015. JAMA Oncol. 2017 Dec 1;3(12):1683-1691. [CrossRef]
- Kanwal F, Kramer J, Asch SM, Chayanupatkul M, Cao Y, El-Serag HB. Risk of Hepatocellular Cancer in HCV Patients Treated With Direct-Acting Antiviral Agents. Gastroenterology. 2017 Oct;153(4):996-1005.e1. [CrossRef]
- Estes C, Razavi H, Loomba R, Younossi Z, Sanyal AJ. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology. 2018 Jan;67(1):123-133. [CrossRef]
- Nishida, N. Metabolic disease as a risk of hepatocellular carcinoma. Clin Mol Hepatol. 2021 Jan;27(1):87-90. [CrossRef]
- Ascha MS, Hanouneh IA, Lopez R, Tamimi TA, Feldstein AF, Zein NN. The incidence and risk factors of hepatocellular carcinoma in patients with nonalcoholic steatohepatitis. Hepatology. 2010 Jun;51(6):1972-8.
- Zhao, L. The gut microbiota and obesity: from correlation to causality. Nat Rev Microbiol. 2013 Sep;11(9):639-47. [CrossRef]
- WHO. Data Visualization Tools for Exploring the Global Cancer Burden in 2020. 2020. Available online: https://gco.iarc.fr/today/ (accessed on 03/03/2022).
- Kulik L, El-Serag HB. Epidemiology and Management of Hepatocellular Carcinoma. Gastroenterology. 2019 Jan;156(2):477-491.e1. [CrossRef]
- Petrick JL, Florio AA, Znaor A, Ruggieri D, Laversanne M, Alvarez CS, Ferlay J, Valery PC, Bray F, McGlynn KA. International trends in hepatocellular carcinoma incidence, 1978-2012. Int J Cancer. 2020 Jul 15;147(2):317-330. [CrossRef]
- Zhang, C.H.; Cheng, Y.; Zhang, S.; Fan, J.; Gao, Q. Changing epidemiology of hepatocellular carcinoma in Asia. Liver Int. 2022, 42, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71:209–49.
- Ferlay J, Colombet M, Soerjomataram I, Parkin DM, Piñeros M, Znaor A, et al. Cancer statistics for the year 2020: An overview. Int J Cancer. 2021;149:778–89. [CrossRef]
- Tellapuri S, Sutphin PD, Beg MS, Singal AG, Kalva SP. Staging systems of hepatocellular carcinoma: A review. Indian J Gastroenterol. 2018 Nov;37(6):481-491. [CrossRef]
- Liu L, Chen H, Wang M, Zhao Y, Cai G, Qi X, Han G. Combination therapy of sorafenib and TACE for unresectable HCC: a systematic review and meta-analysis. PLoS One. 2014 Mar 20;9(3):e91124. [CrossRef]
- Chidambaranathan-Reghupaty S, Fisher PB, Sarkar D. Hepatocellular carcinoma (HCC): Epidemiology, etiology and molecular classification. Adv Cancer Res. 2021;149:1-61. [CrossRef]
- Lin S, Hoffmann K, Schemmer P. Treatment of hepatocellular carcinoma: a systematic review. Liver Cancer. 2012;1:144–58. [CrossRef]
- Li M, Zhang W, Wang B, Gao Y, Song Z, Zheng QC. Ligand-based targeted therapy: a novel strategy for hepatocellular carcinoma. Int J Nanomedicine. 2016 Oct 31;11:5645-5669. [CrossRef]
- Llovet JM, Castet F, Heikenwalder M, Maini MK, Mazzaferro V, Pinato DJ, Pikarsky E, Zhu AX, Finn RS. Immunotherapies for hepatocellular carcinoma. Nat Rev Clin Oncol. 2022 Mar;19(3):151-172. [CrossRef]
- Zucman-Rossi J, Villanueva A, Nault JC, Llovet JM. Genetic Landscape and Biomarkers of Hepatocellular Carcinoma. Gastroenterology. 2015 Oct;149(5):1226-1239.e4. [CrossRef]
- Sia D, Villanueva A, Friedman SL, Llovet JM. Liver Cancer Cell of Origin, Molecular Class, and Effects on Patient Prognosis. Gastroenterology. 2017 Mar;152(4):745-761. [CrossRef]
- Shafizadeh N, Kakar S. Diagnosis of well-differentiated hepatocellular lesions: role of immunohistochemistry and other ancillary techniques. Adv Anat Pathol. 2011 Nov;18(6):438-45. [CrossRef]
- Michalopoulos, G. K. & Bhushan, B. Liver regeneration: biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 18, 40–55 (2021).
- Bruno S, Herrera Sanchez MB, Chiabotto G, Fonsato V, Navarro-Tableros V, Pasquino C, Tapparo M, Camussi G. Human Liver Stem Cells: A Liver-Derived Mesenchymal Stromal Cell-Like Population With Pro-regenerative Properties. Front Cell Dev Biol. 2021 Apr 26;9:644088. [CrossRef]
- Bayard Q, Meunier L, Peneau C, Renault V, Shinde J, Nault JC, Mami I, Couchy G, Amaddeo G, Tubacher E, Bacq D, Meyer V, La Bella T, Debaillon-Vesque A, Bioulac-Sage P, Seror O, Blanc JF, Calderaro J, Deleuze JF, Imbeaud S, Zucman-Rossi J, Letouzé E. Cyclin A2/E1 activation defines a hepatocellular carcinoma subclass with a rearrangement signature of replication stress. Nat Commun. 2018 Dec 7;9(1):5235. [CrossRef]
- Pfister D, Núñez NG, Pinyol R, Govaere O, Pinter M, Szydlowska M, Gupta R, Qiu M, Deczkowska A, Weiner A, Müller F, Sinha A, Friebel E, Engleitner T, Lenggenhager D, Moncsek A, Heide D, Stirm K, Kosla J, Kotsiliti E, Leone V, Dudek M, Yousuf S, Inverso D, Singh I, Teijeiro A, Castet F, Montironi C, Haber PK, Tiniakos D, Bedossa P, Cockell S, Younes R, Vacca M, Marra F, Schattenberg JM, Allison M, Bugianesi E, Ratziu V, Pressiani T, D'Alessio A, Personeni N, Rimassa L, Daly AK, Scheiner B, Pomej K, Kirstein MM, Vogel A, Peck-Radosavljevic M, Hucke F, Finkelmeier F, Waidmann O, Trojan J, Schulze K, Wege H, Koch S, Weinmann A, Bueter M, Rössler F, Siebenhüner A, De Dosso S, Mallm JP, Umansky V, Jugold M, Luedde T, Schietinger A, Schirmacher P, Emu B, Augustin HG, Billeter A, Müller-Stich B, Kikuchi H, Duda DG, Kütting F, Waldschmidt DT, Ebert MP, Rahbari N, Mei HE, Schulz AR, Ringelhan M, Malek N, Spahn S, Bitzer M, Ruiz de Galarreta M, Lujambio A, Dufour JF, Marron TU, Kaseb A, Kudo M, Huang YH, Djouder N, Wolter K, Zender L, Marche PN, Decaens T, Pinato DJ, Rad R, Mertens JC, Weber A, Unger K, Meissner F, Roth S, Jilkova ZM, Claassen M, Anstee QM, Amit I, Knolle P, Becher B, Llovet JM, Heikenwalder M. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature. 2021 Apr;592(7854):450-456. [CrossRef]
- Gellert-Kristensen H, Richardson TG, Davey Smith G, Nordestgaard BG, Tybjaerg-Hansen A, Stender S. Combined Effect of PNPLA3, TM6SF2, and HSD17B13 Variants on Risk of Cirrhosis and Hepatocellular Carcinoma in the General Population. Hepatology. 2020 Sep;72(3):845-856. [CrossRef]
- Yang JD, Hainaut P, Gores GJ, Amadou A, Plymoth A, Roberts LR. A global view of hepatocellular carcinoma: trends, risk, prevention and management. Nat Rev Gastroenterol Hepatol. 2019 Oct;16(10):589-604. [CrossRef]
- Nault JC, Letouzé E. Mutational Processes in Hepatocellular Carcinoma: The Story of Aristolochic Acid. Semin Liver Dis. 2019 Jul;39(3):334-340. [CrossRef]
- Guichard C, Amaddeo G, Imbeaud S, Ladeiro Y, Pelletier L, Maad IB, Calderaro J, Bioulac-Sage P, Letexier M, Degos F, Clément B, Balabaud C, Chevet E, Laurent A, Couchy G, Letouzé E, Calvo F, Zucman-Rossi J. Integrated analysis of somatic mutations and focal copy-number changes identifies key genes and pathways in hepatocellular carcinoma. Nat Genet. 2012 May 6;44(6):694-8. [CrossRef]
- Takai A, Dang HT, Wang XW. Identification of drivers from cancer genome diversity in hepatocellular carcinoma. Int J Mol Sci. 2014 Jun 20;15(6):11142-60. [CrossRef]
- Ahn SM, Jang SJ, Shim JH, Kim D, Hong SM, Sung CO, Baek D, Haq F, Ansari AA, Lee SY, Chun SM, Choi S, Choi HJ, Kim J, Kim S, Hwang S, Lee YJ, Lee JE, Jung WR, Jang HY, Yang E, Sung WK, Lee NP, Mao M, Lee C, Zucman-Rossi J, Yu E, Lee HC, Kong G. Genomic portrait of resectable hepatocellular carcinomas: implications of RB1 and FGF19 aberrations for patient stratification. Hepatology. 2014 Dec;60(6):1972-82. [CrossRef]
- Belinky F, Nativ N, Stelzer G, Zimmerman S, Iny Stein T, Safran M, Lancet D. PathCards: multi-source consolidation of human biological pathways. Database (Oxford). 2015;2015:bav006.
- Schulze K, Imbeaud S, Letouzé E, Alexandrov LB, Calderaro J, Rebouissou S, Couchy G, Meiller C, Shinde J, Soysouvanh F, Calatayud AL, Pinyol R, Pelletier L, Balabaud C, Laurent A, Blanc JF, Mazzaferro V, Calvo F, Villanueva A, Nault JC, Bioulac-Sage P, Stratton MR, Llovet JM, Zucman-Rossi J. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat Genet. 2015 May;47(5):505-511. [CrossRef]
- National Cancer Institute. The Cancer Genome Atlas. Available online: http://cancergenome.nih.gov/ (accessed on 04 March 2023).
- Tao J, Xu E, Zhao Y, Singh S, Li X, Couchy G, Chen X, Zucman-Rossi J, Chikina M, Monga SP. Modeling a human hepatocellular carcinoma subset in mice through coexpression of met and point-mutant β-catenin. Hepatology. 2016 Nov;64(5):1587-1605. [CrossRef]
- Ruiz de Galarreta M, Bresnahan E, Molina-Sánchez P, Lindblad KE, Maier B, Sia D, Puigvehi M, Miguela V, Casanova-Acebes M, Dhainaut M, Villacorta-Martin C, Singhi AD, Moghe A, von Felden J, Tal Grinspan L, Wang S, Kamphorst AO, Monga SP, Brown BD, Villanueva A, Llovet JM, Merad M, Lujambio A. β-Catenin Activation Promotes Immune Escape and Resistance to Anti-PD-1 Therapy in Hepatocellular Carcinoma. Cancer Discov. 2019 Aug;9(8):1124-1141. [CrossRef]
- Tao J, Krutsenko Y, Moghe A, Singh S, Poddar M, Bell A, Oertel M, Singhi AD, Geller D, Chen X, Lujambio A,Tan Liu S, Monga SP. Nuclear factor erythroid 2-related factor 2 and β-Catenin Coactivation in Hepatocellular Cancer: Biological and Therapeutic Implications. Hepatology. 2021 Aug;74(2):741-759. [CrossRef]
- Tan X, Behari J, Cieply B, Michalopoulos GK, Monga SP. Conditional deletion of beta-catenin reveals its role in liver growth and regeneration. Gastroenterology. 2006 Nov;131(5):1561-72. [CrossRef]
- Miyoshi Y, Nagase H, Ando H, Horii A, Ichii S, Nakatsuru S, Aoki T, Miki Y, Mori T, Nakamura Y. Somatic mutations of the APC gene in colorectal tumors: mutation cluster region in the APC gene. Hum Mol Genet. 1992 Jul;1(4):229-33. [CrossRef]
- Li J, Quan H, Liu Q, Si Z, He Z, Qi H. Alterations of axis inhibition protein 1 (AXIN1) in hepatitis B virus-related hepatocellular carcinoma and overexpression of AXIN1 induces apoptosis in hepatocellular cancer cells. Oncol Res. 2013;20(7):281-8. [CrossRef]
- Satoh S, Daigo Y, Furukawa Y, Kato T, Miwa N, Nishiwaki T, Kawasoe T, Ishiguro H, Fujita M, Tokino T, Sasaki Y, Imaoka S, Murata M, Shimano T, Yamaoka Y, Nakamura Y. AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat Genet. 2000 Mar;24(3):245-50. [CrossRef] [PubMed]
- Huang SM, Mishina YM, Liu S, Cheung A, Stegmeier F, Michaud GA, Charlat O, Wiellette E, Zhang Y, Wiessner S, Hild M, Shi X, Wilson CJ, Mickanin C, Myer V, Fazal A, Tomlinson R, Serluca F, Shao W, Cheng H, Shultz M, Rau C, Schirle M, Schlegl J, Ghidelli S, Fawell S, Lu C, Curtis D, Kirschner MW, Lengauer C, Finan PM, Tallarico JA, Bouwmeester T, Porter JA, Bauer A, Cong F. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature. 2009 Oct 1;461(7264):614-20. [CrossRef]
- Totoki Y, Tatsuno K, Covington KR, Ueda H, Creighton CJ, Kato M, Tsuji S, Donehower LA, Slagle BL, Nakamura H, Yamamoto S, Shinbrot E, Hama N, Lehmkuhl M, Hosoda F, Arai Y, Walker K, Dahdouli M, Gotoh K, Nagae G, Gingras MC, Muzny DM, Ojima H, Shimada K, Midorikawa Y, Goss JA, Cotton R, Hayashi A, Shibahara J, Ishikawa S, Guiteau J, Tanaka M, Urushidate T, Ohashi S, Okada N, Doddapaneni H, Wang M, Zhu Y, Dinh H, Okusaka T, Kokudo N, Kosuge T, Takayama T, Fukayama M, Gibbs RA, Wheeler DA, Aburatani H, Shibata T. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat Genet. 2014 Dec;46(12):1267-73. [CrossRef]
- Pezzuto F, Izzo F, Buonaguro L, Annunziata C, Tatangelo F, Botti G, Buonaguro FM, Tornesello ML. Tumor specific mutations in TERT promoter and CTNNB1 gene in hepatitis B and hepatitis C related hepatocellular carcinoma. Oncotarget. 2016 Aug 23;7(34):54253-54262. [CrossRef]
- Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L, Garraway LA. Highly recurrent TERT promoter mutations in human melanoma. Science. 2013 Feb 22;339(6122):957-9. [CrossRef]
- Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, Kadel S, Moll I, Nagore E, Hemminki K, Schadendorf D, Kumar R. TERT promoter mutations in familial and sporadic melanoma. Science. 2013 Feb 22;339(6122):959-61. [CrossRef]
- Borah S, Xi L, Zaug AJ, Powell NM, Dancik GM, Cohen SB, Costello JC, Theodorescu D, Cech TR. Cancer. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science. 2015 Feb 27;347(6225):1006-10. [CrossRef]
- Lombardo D, Saitta C, Giosa D, Di Tocco FC, Musolino C, Caminiti G, Chines V, Franzè MS, Alibrandi A, Navarra G, Raimondo G, Pollicino T. Frequency of somatic mutations in TERT promoter, TP53 and CTNNB1 genes in patients with hepatocellular carcinoma from Southern Italy. Oncol Lett. 2020 Mar;19(3):2368-2374. [CrossRef]
- Sirma H, Kumar M, Meena JK, Witt B, Weise JM, Lechel A, Ande S, Sakk V, Guguen-Guillouzo C, Zender L, Rudolph KL, Günes C. The promoter of human telomerase reverse transcriptase is activated during liver regeneration and hepatocyte proliferation. Gastroenterology. 2011 Jul;141(1):326-37, 337.e1-3. [CrossRef]
- Liu T, Li W, Lu W, Chen M, Luo M, Zhang C, Li Y, Qin G, Shi D, Xiao B, Qiu H, Yu W, Kang L, Kang T, Huang W, Yu X, Wu X, Deng W. RBFOX3 Promotes Tumor Growth and Progression via hTERT Signaling and Predicts a Poor Prognosis in Hepatocellular Carcinoma. Theranostics. 2017 Jul 22;7(12):3138-3154. [CrossRef]
- Esopi D, Graham MK, Brosnan-Cashman JA, Meyers J, Vaghasia A, Gupta A, Kumar B, Haffner MC, Heaphy CM, De Marzo AM, Meeker AK, Nelson WG, Wheelan SJ, Yegnasubramanian S. Pervasive promoter hypermethylation of silenced TERT alleles in human cancers. Cell Oncol (Dordr). 2020 Oct;43(5):847-861. [CrossRef]
- Montironi C, Castet F, Haber PK, Pinyol R, Torres-Martin M, Torrens L, Mesropian A, Wang H, Puigvehi M, Maeda M, Leow WQ, Harrod E, Taik P, Chinburen J, Taivanbaatar E, Chinbold E, Solé Arqués M, Donovan M, Thung S, Neely J, Mazzaferro V, Anderson J, Roayaie S, Schwartz M, Villanueva A, Friedman SL, Uzilov A, Sia D, Llovet JM. Inflamed and non-inflamed classes of HCC: a revised immunogenomic classification. Gut. 2023 Jan;72(1):129-140. [CrossRef]
- Villanueva A, Hoshida Y. Depicting the role of TP53 in hepatocellular carcinoma progression. J Hepatol. 2011 Sep;55(3):724-725. [CrossRef]
- Jeng YM, Hsu HC. Mutation of the DR5/TRAIL receptor 2 gene is infrequent in hepatocellular carcinoma. Cancer Lett. 2002 Jul 26;181(2):205-8. [CrossRef]
- Liu, S. , Qiu, J., He, G. et al. TRAIL promotes hepatocellular carcinoma apoptosis and inhibits proliferation and migration via interacting with IER3. Cancer Cell Int 21, 63 (2021). [CrossRef]
- Salmena L, Lemmers B, Hakem A, Matysiak-Zablocki E, Murakami K, Au PY, Berry DM, Tamblyn L, Shehabeldin A, Migon E, Wakeham A, Bouchard D, Yeh WC, McGlade JC, Ohashi PS, Hakem R. Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity. Genes Dev. 2003 Apr 1;17(7):883-95. [CrossRef]
- Kim IK, Chung CW, Woo HN, Hong GS, Nagata S, Jung YK. Reconstitution of caspase-8 sensitizes JB6 cells to TRAIL. Biochem Biophys Res Commun. 2000 Oct 22;277(2):311-6. [CrossRef]
- Kischkel FC, Lawrence DA, Chuntharapai A, Schow P, Kim KJ, Ashkenazi A. Apo2L/TRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity. 2000 Jun;12(6):611-20. [CrossRef]
- Lin Y, Devin A, Cook A, Keane MM, Kelliher M, Lipkowitz S, Liu ZG. The death domain kinase RIP is essential for TRAIL (Apo2L)-induced activation of IkappaB kinase and c-Jun N-terminal kinase. Mol Cell Biol. 2000 Sep;20(18):6638-45. [CrossRef]
- Schneider P, Thome M, Burns K, Bodmer JL, Hofmann K, Kataoka T, Holler N, Tschopp J. TRAIL receptors 1 (DR4) and 2 (DR5) signal FADD-dependent apoptosis and activate NF-kappaB. Immunity. 1997 Dec;7(6):831-6. [CrossRef]
- Belinky F, Nativ N, Stelzer G, Zimmerman S, Iny Stein T, Safran M, Lancet D. PathCards: multi-source consolidation of human biological pathways. Database (Oxford). 2015;2015:bav006.
- Knudsen ES, Gopal P, Singal AG. The changing landscape of hepatocellular carcinoma: etiology, genetics, and therapy. Am J Pathol. 2014 Mar;184(3):574-83. [CrossRef]
- Llovet JM, Villanueva A, Lachenmayer A, Finn RS. Advances in targeted therapies for hepatocellular carcinoma in the genomic era. Nat Rev Clin Oncol. 2015 Aug;12(8):436. Epub 2015 Jun 23. Erratum for: Nat Rev Clin Oncol. 2015 Jul;12(7):408-24. [CrossRef]
- Sawey ET, Chanrion M, Cai C, Wu G, Zhang J, Zender L, Zhao A, Busuttil RW, Yee H, Stein L, French DM, Finn RS, Lowe SW, Powers S. Identification of a therapeutic strategy targeting amplified FGF19 in liver cancer by Oncogenomic screening. Cancer Cell. 2011 Mar 8;19(3):347-58. [CrossRef]
- Anstee QM, Reeves HL, Kotsiliti E, Govaere O, Heikenwalder M. From NASH to HCC: current concepts and future challenges. Nat Rev Gastroenterol Hepatol. 2019 Jul;16(7):411-428. [CrossRef]
- Nakagawa H, Umemura A, Taniguchi K, Font-Burgada J, Dhar D, Ogata H, Zhong Z, Valasek MA, Seki E, Hidalgo J, Koike K, Kaufman RJ, Karin M. ER stress cooperates with hypernutrition to trigger TNF-dependent spontaneous HCC development. Cancer Cell. 2014 Sep 8;26(3):331-343. [CrossRef]
- Nishida N, Yada N, Hagiwara S, Sakurai T, Kitano M, Kudo M. Unique features associated with hepatic oxidative DNA damage and DNA methylation in non-alcoholic fatty liver disease. J Gastroenterol Hepatol. 2016 Sep;31(9):1646-53. [CrossRef]
- Guri Y, Colombi M, Dazert E, Hindupur SK, Roszik J, Moes S, Jenoe P, Heim MH, Riezman I, Riezman H, Hall MN. mTORC2 Promotes Tumorigenesis via Lipid Synthesis. Cancer Cell. 2017 Dec 11;32(6):807-823.e12. [CrossRef]
- Liu D, Wong CC, Fu L, Chen H, Zhao L, Li C, Zhou Y, Zhang Y, Xu W, Yang Y, Wu B, Cheng G, Lai PB, Wong N, Sung JJY, Yu J. Squalene epoxidase drives NAFLD-induced hepatocellular carcinoma and is a pharmaceutical target. Sci Transl Med. 2018 Apr 18;10(437):eaap9840. Erratum in: Sci Transl Med. 2018 Sep 26;10(460). [CrossRef]
- Estes C, Razavi H, Loomba R, Younossi Z, Sanyal AJ. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology. 2018 Jan;67(1):123-133. [CrossRef]
- Hsu CL, Yen GC. Phenolic compounds: evidence for inhibitory effects against obesity and their underlying molecular signaling mechanisms. Mol Nutr Food Res. 2008 Jan;52(1):53-61. [CrossRef]
- Zhang HX, Jiang SS, Zhang XF, Zhou ZQ, Pan QZ, Chen CL, Zhao JJ, Tang Y, Xia JC, Weng DS. Protein kinase CK2α catalytic subunit is overexpressed and serves as an unfavorable prognostic marker in primary hepatocellular carcinoma. Oncotarget. 2015 Oct 27;6(33):34800-17. [CrossRef]
- Tawfic S, Yu S, Wang H, Faust R, Davis A, Ahmed K. Protein kinase CK2 signal in neoplasia. Histol Histopathol. 2001 Apr;16(2):573-82. [CrossRef]
- Yenice S, Davis AT, Goueli SA, Akdas A, Limas C, Ahmed K. Nuclear casein kinase 2 (CK-2) activity in human normal, benign hyperplastic, and cancerous prostate. Prostate. 1994;24(1):11-6. [CrossRef]
- Croquelois A, Blindenbacher A, Terracciano L, Wang X, Langer I, Radtke F, Heim MH. Inducible inactivation of Notch1 causes nodular regenerative hyperplasia in mice. Hepatology. 2005 Mar;41(3):487-96. [CrossRef]
- Zhang Y, Wang X. Targeting the Wnt/β-catenin signaling pathway in cancer. J Hematol Oncol. 2020 Dec 4;13(1):165. [CrossRef]
- Mojsin M, Vicentic JM, Schwirtlich M, Topalovic V, Stevanovic M. Quercetin reduces pluripotency, migration and adhesion of human teratocarcinoma cell line NT2/D1 by inhibiting Wnt/β-catenin signaling. Food Funct. 2014 Oct;5(10):2564-73. [CrossRef]
- Kim H, Seo EM, Sharma AR, Ganbold B, Park J, Sharma G, Kang YH, Song DK, Lee SS, Nam JS. Regulation of Wnt signaling activity for growth suppression induced by quercetin in 4T1 murine mammary cancer cells. Int J Oncol. 2013 Oct;43(4):1319-25. [CrossRef]
- Osaki M, Oshimura M, Ito H. PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis. 2004 Nov;9(6):667-76. [CrossRef]
- Xiang T, Fang Y, Wang SX. Quercetin suppresses HeLa cells by blocking PI3K/Akt pathway. J Huazhong Univ Sci Technolog Med Sci. 2014 Oct;34(5):740-744. [CrossRef]
- Gulati N, Laudet B, Zohrabian VM, Murali R, Jhanwar-Uniyal M. The antiproliferative effect of Quercetin in cancer cells is mediated via inhibition of the PI3K-Akt/PKB pathway. Anticancer Res. 2006 Mar-Apr;26(2A):1177-81.
- Tan HK, Moad AI, Tan ML. The mTOR signalling pathway in cancer and the potential mTOR inhibitory activities of natural phytochemicals. Asian Pac J Cancer Prev. 2014;15(16):6463-75. [CrossRef]
- Fyffe C, Falasca M. 3-Phosphoinositide-dependent protein kinase-1 as an emerging target in the management of breast cancer. Cancer Manag Res. 2013 Aug 23;5:271-80. [CrossRef]
- Maurya AK, Vinayak M. PI-103 and Quercetin Attenuate PI3K-AKT Signaling Pathway in T- Cell Lymphoma Exposed to Hydrogen Peroxide. PLoS One. 2016 Aug 5;11(8):e0160686. [CrossRef]
- Navarro-Núñez L, Lozano ML, Martínez C, Vicente V, Rivera J. Effect of quercetin on platelet spreading on collagen and fibrinogen and on multiple platelet kinases. Fitoterapia. 2010 Mar;81(2):75-80. [CrossRef]
- O'Shea JJ, Schwartz DM, Villarino AV, Gadina M, McInnes IB, Laurence A. The JAK-STAT pathway: impact on human disease and therapeutic intervention. Annu Rev Med. 2015;66:311-28. [CrossRef]
- Qin Y, He LY, Chen Y, Wang WY, Zhao XH, Wu MY. Quercetin affects leptin and its receptor in human gastric cancer MGC-803 cells and JAK-STAT pathway. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2012 Jan;28(1):12-6.
- Michaud-Levesque J, Bousquet-Gagnon N, Béliveau R. Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp Cell Res. 2012 May 1;318(8):925-35. [CrossRef]
- Seo HS, Ku JM, Choi HS, Choi YK, Woo JK, Kim M, Kim I, Na CH, Hur H, Jang BH, Shin YC, Ko SG. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol Rep. 2016 Jul;36(1):31-42. [CrossRef]
- Luo CL, Liu YQ, Wang P, Song CH, Wang KJ, Dai LP, Zhang JY, Ye H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomed Pharmacother. 2016 Aug;82:595-605. [CrossRef]
- Wu L, Li J, Liu T, Li S, Feng J, Yu Q, Zhang J, Chen J, Zhou Y, Ji J, Chen K, Mao Y, Wang F, Dai W, Fan X, Wu J, Guo C. Quercetin shows anti-tumor effect in hepatocellular carcinoma LM3 cells by abrogating JAK2/STAT3 signaling pathway. Cancer Med. 2019 Aug;8(10):4806-4820. [CrossRef]
- Panagiotakopoulou M, Lendenmann T, Pramotton FM, Giampietro C, Stefopoulos G, Poulikakos D, Ferrari A. Cell cycle-dependent force transmission in cancer cells. Mol Biol Cell. 2018 Oct 15;29(21):2528-2539. [CrossRef]
- Widmann C, Gibson S, Jarpe MB, Johnson GL. Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol Rev. 1999 Jan;79(1):143-80. [CrossRef]
- Xu D, Hu MJ, Wang YQ, Cui YL. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules. 2019 Mar 21;24(6):1123. [CrossRef]
- Wang R, Zhang H, Wang Y, Song F, Yuan Y. Inhibitory effects of quercetin on the progression of liver fibrosis through the regulation of NF-кB/IкBα, p38 MAPK, and Bcl-2/Bax signaling. Int Immunopharmacol. 2017 Jun;47:126-133. [CrossRef]
- Asgharian P, Tazekand AP, Hosseini K, Forouhandeh H, Ghasemnejad T, Ranjbar M, Hasan M, Kumar M, Beirami SM, Tarhriz V, Soofiyani SR, Kozhamzharova L, Sharifi-Rad J, Calina D, Cho WC. Potential mechanisms of quercetin in cancer prevention: focus on cellular and molecular targets. Cancer Cell Int. 2022 Aug 15;22(1):257. [CrossRef]
- Xavier CP, Lima CF, Rohde M, Pereira-Wilson C. Quercetin enhances 5-fluorouracil-induced apoptosis in MSI colorectal cancer cells through p53 modulation. Cancer Chemother Pharmacol. 2011 Dec;68(6):1449-57. [CrossRef]
- Wang G, Zhang J, Liu L, Sharma S, Dong Q. Quercetin potentiates doxorubicin mediated antitumor effects against liver cancer through p53/Bcl-xl. PLoS One. 2012;7(12):e51764. [CrossRef]
- Bishayee K, Ghosh S, Mukherjee A, Sadhukhan R, Mondal J, Khuda-Bukhsh AR. Quercetin induces cytochrome-c release and ROS accumulation to promote apoptosis and arrest the cell cycle in G2/M, in cervical carcinoma: signal cascade and drug-DNA interaction. Cell Prolif. 2013 Apr;46(2):153-63. [CrossRef]
- Bishayee K, Khuda-Bukhsh AR, Huh SO. PLGA-Loaded Gold-Nanoparticles Precipitated with Quercetin Downregulate HDAC-Akt Activities Controlling Proliferation and Activate p53-ROS Crosstalk to Induce Apoptosis in Hepatocarcinoma Cells. Mol Cells. 2015 Jun;38(6):518-27. [CrossRef]
- Nguyen LT, Lee YH, Sharma AR, Park JB, Jagga S, Sharma G, Lee SS, Nam JS. Quercetin induces apoptosis and cell cycle arrest in triple-negative breast cancer cells through modulation of Foxo3a activity. Korean J Physiol Pharmacol. 2017 Mar;21(2):205-213. [CrossRef]
- Wei JX, Lv LH, Wan YL, Cao Y, Li GL, Lin HM, Zhou R, Shang CZ, Cao J, He H, Han QF, Liu PQ, Zhou G, Min J. Vps4A functions as a tumor suppressor by regulating the secretion and uptake of exosomal microRNAs in human hepatoma cells. Hepatology. 2015 Apr;61(4):1284-94. [CrossRef]
- Giordano S, Columbano A. MicroRNAs: new tools for diagnosis, prognosis, and therapy in hepatocellular carcinoma? Hepatology. 2013 Feb;57(2):840-7. [CrossRef]
- Wang J, Pu J, Zhang Y, Yao T, Luo Z, Li W, Xu G, Liu J, Wei W, Deng Y. Exosome-transmitted long non-coding RNA SENP3-EIF4A1 suppresses the progression of hepatocellular carcinoma. Aging (Albany NY). 2020 Jun 27;12(12):11550-11567. [CrossRef]
- Cao L, Xie B, Yang X, Liang H, Jiang X, Zhang D, Xue P, Chen D, Shao Z. MiR-324-5p Suppresses Hepatocellular Carcinoma Cell Invasion by Counteracting ECM Degradation through Post-Transcriptionally Downregulating ETS1 and SP1. PLoS One. 2015 Jul 15;10(7):e0133074. [CrossRef]
- Zhang Z, Li X, Sun W, Yue S, Yang J, Li J, Ma B, Wang J, Yang X, Pu M, Ruan B, Zhao G, Huang Q, Wang L, Tao K, Dou K. Loss of exosomal miR-320a from cancer-associated fibroblasts contributes to HCC proliferation and metastasis. Cancer Lett. 2017 Jul 1;397:33-42. [CrossRef]
- Jiang W, Tan Y, Cai M, Zhao T, Mao F, Zhang X, Xu W, Yan Z, Qian H, Yan Y. Human Umbilical Cord MSC-Derived Exosomes Suppress the Development of CCl4-Induced Liver Injury through Antioxidant Effect. Stem Cells Int. 2018 Mar 4;2018:6079642. [CrossRef]
- Wang J, Wang X, Zhang X, Shao T, Luo Y, Wang W, Han Y. Extracellular Vesicles and Hepatocellular Carcinoma: Opportunities and Challenges. Front Oncol. 2022 May 25;12:884369. [CrossRef]
- Yugawa K, Yoshizumi T, Mano Y, Itoh S, Harada N, Ikegami T, Kohashi K, Oda Y, Mori M. Cancer-associated fibroblasts promote hepatocellular carcinoma progression through downregulation of exosomal miR-150-3p. Eur J Surg Oncol. 2021 Feb;47(2):384-393. [CrossRef]
- Xu T, Zhu Y, Xiong Y, Ge YY, Yun JP, Zhuang SM. MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cells. Hepatology. 2009 Jul;50(1):113-21. [CrossRef]
- Wang R, Zhao N, Li S, Fang JH, Chen MX, Yang J, Jia WH, Yuan Y, Zhuang SM. MicroRNA-195 suppresses angiogenesis and metastasis of hepatocellular carcinoma by inhibiting the expression of VEGF, VAV2, and CDC42. Hepatology. 2013 Aug;58(2):642-53. [CrossRef]
- Chen W, Quan Y, Fan S, Wang H, Liang J, Huang L, Chen L, Liu Q, He P, Ye Y. Exosome-transmitted circular RNA hsa_circ_0051443 suppresses hepatocellular carcinoma progression. Cancer Lett. 2020 Apr 10;475:119-128. [CrossRef]
- Food and Drug Administration. Guidance for industry, PAT-A framework for innovative pharmaceutical development, manufacturing and quality assurance. http://www. fda. gov/cder/guidance/published.html. 2004. (Accessed on 08/03/2023).
Figure 4.
Barcelona Clinic Liver Cancer (BCLC) staging system.
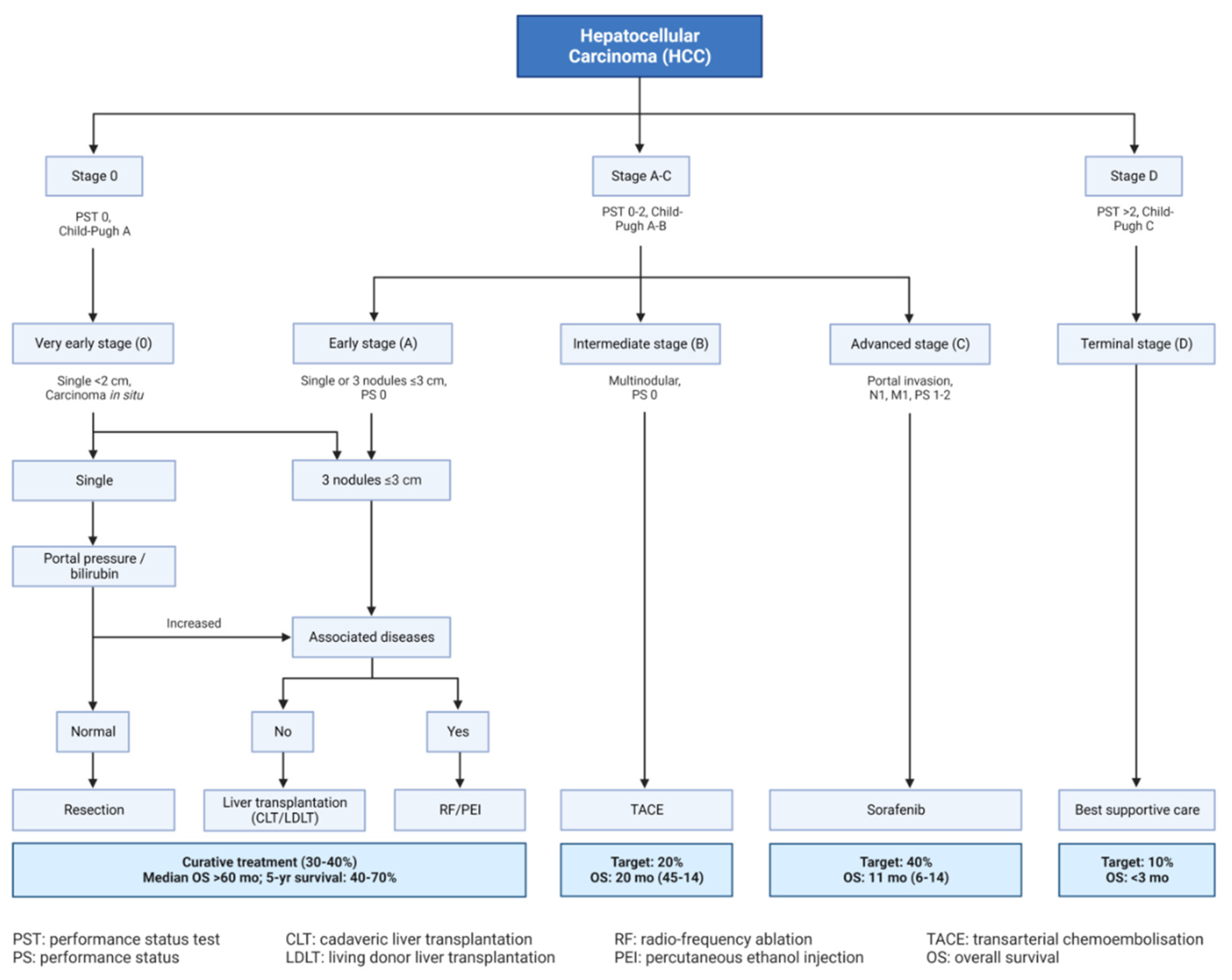
Figure 5.
The most important signalling pathways affected by quercetin during cancer prevention. A) Wnt/β catenin pathway; quercetin inhibits β-catenin translocation to the nucleus, B) PI3K/Akt pathway; inhibition of phosphorylation of PI3k, Akt, and S6K, C) JAK/STAT pathway; inhibition of p-STAT formation; D) MAPK pathway; induced phosphorylation of p38, JNK, and ERK, E) p53 pathway; induced phosphorylation of p53 and induction of apoptosis. (Adopted from Ref. 99 under the terms of the Creative Commons Attribution License (CC BY)).
Figure 5.
The most important signalling pathways affected by quercetin during cancer prevention. A) Wnt/β catenin pathway; quercetin inhibits β-catenin translocation to the nucleus, B) PI3K/Akt pathway; inhibition of phosphorylation of PI3k, Akt, and S6K, C) JAK/STAT pathway; inhibition of p-STAT formation; D) MAPK pathway; induced phosphorylation of p38, JNK, and ERK, E) p53 pathway; induced phosphorylation of p53 and induction of apoptosis. (Adopted from Ref. 99 under the terms of the Creative Commons Attribution License (CC BY)).
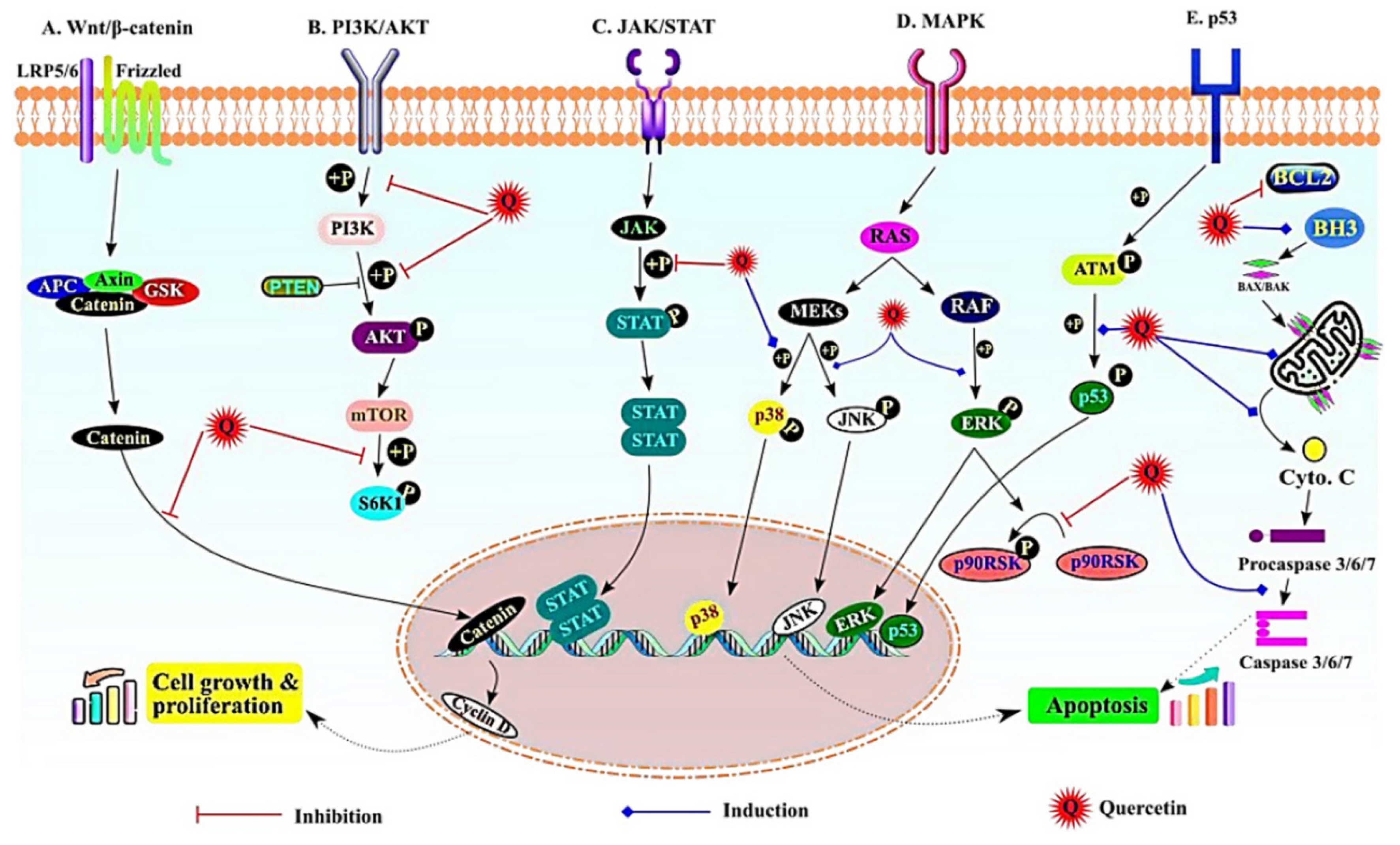
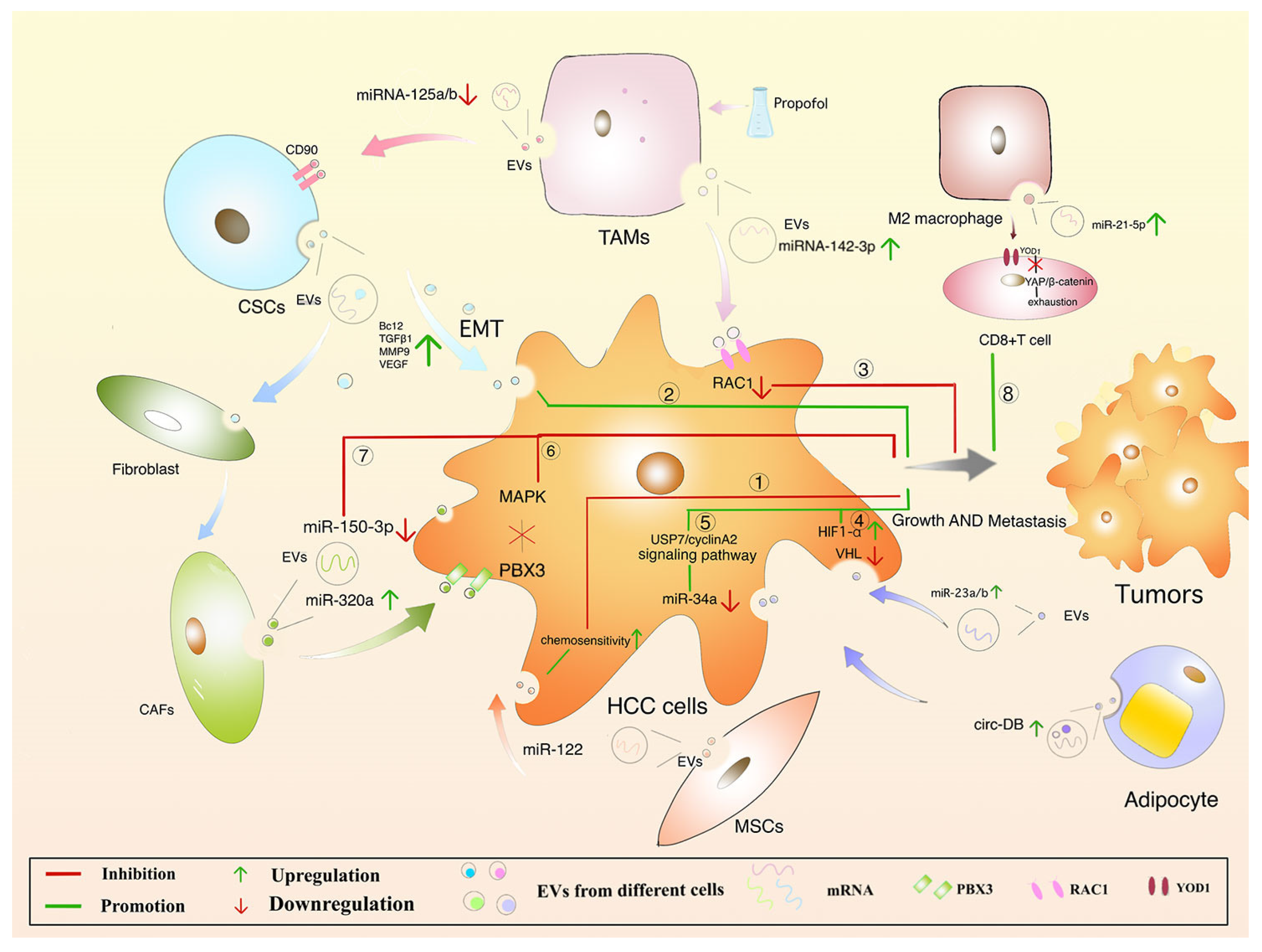
Figure 6.
Regulation of hepatocellular carcinoma by different cell-derived EVs. (Adopted from Ref. 111 under the terms of the Creative Commons Attribution License (CC BY)). TAMs, tumor-associated macrophages; CSCs, cancer stem cells; CAFs, cancer-associated fibroblasts; EMT, epithelial-mesenchymal transition.
Figure 6.
Regulation of hepatocellular carcinoma by different cell-derived EVs. (Adopted from Ref. 111 under the terms of the Creative Commons Attribution License (CC BY)). TAMs, tumor-associated macrophages; CSCs, cancer stem cells; CAFs, cancer-associated fibroblasts; EMT, epithelial-mesenchymal transition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



