
Submitted:
01 December 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
Vector-borne disease (VBD) accounts for more than 17% of the global infectious disease burden, disproportionately affecting developing nations within tropical and subtropical areas. As Aedes aegypti mosquitoes are the principal vector for the transmission of many Public Health pathogens, our study examined their population dynamics within the seasonal context of tropical dry biomes in El Salvador as incidence of VBD in this region has increased in recent years. Species that rely on sexual reproduction must maintain a balanced sex ratio (1:1) to support their population dynamics, however, our phenological analysis of field-collected A. aegypti specimens revealed significant sex ratio variations during dry and wet seasons. Interestingly, we detected a seasonal shift that favored female mosquitoes during the dry season, highlighting the dynamic nature of mosquito populations in highly urbanized environments like El Salvador. By linking sex ratio distortions to anthropogenic factors, our study underscores the importance of comprehensively characterizing the urban ecology of vectors for the development of effective biocontrol strategies. As such, we advocate for more nuanced vector control approaches that consider ecological factors as well as the human behaviors that influence mosquito breeding habitats to effectively combat the rise of VBDs amid global climate changes.
Keywords:
Neotropics
; Northern Triangle of Central America
; phenology
; tropical dry forests
; urban biology
; vector ecology
; vector-borne disease.
1. Introduction
Vector-borne diseases (VBDs), like malaria and dengue, are caused by microbial pathogens that are primarily or exclusively transmitted through invertebrate vectors (mainly arthropods) when they take a blood meal from an infected host (human or animal). VBDs have a tremendous impact on human mortality and morbidity worldwide, resulting in more than 700,000 deaths annually though disease burden disproportionally affects underdeveloped nations within tropical and subtropical regions [1]. The worldwide distribution of VBDs is dependent on the complex interplay of demographic, ecological, environmental and social factors, with more than half the world’s population at risk for infection with at least one vector-borne pathogen [1]. Human activity (e.g. expanding worldwide trade and travel, urbanization, changes in land use and population growth) coupled with global climate changes (e.g. increased temperatures, altered weather patterns, and changes in seasonal rainfall) have increased the worldwide distribution of many vectors, which is predicted to accelerate the spread of VBDs in the future [1,2,3]. While many arthropod species can act like vectors for disease transmission, mosquitoes and ticks account for the majority of VBD incidence worldwide.
Mosquito vectors acquire vector-borne pathogens because female mosquitoes are haematophagous, taking blood meals during the mating season to sustain the additional nutritional requirements associated with ovulation. Even if the blood meal is taken from an infected host, the mosquito vector must acquire a sufficient pathogen load to establish an infection within the epithelial cells lining the mesenteron (midgut). During the extrinsic incubation period (lasting 1-3 weeks depending on the pathogen), the pathogen must then escape from the midgut epithelium into the hemolymph (body cavity) where it disseminates throughout the mosquito body. However, it is the invasion of the salivary glands that facilitates VBD transmission to a susceptible host during a blood meal [4,5]. Interestingly, mosquitoes remain infectious for life and can transmit the vector-borne pathogen to susceptible hosts during subsequent blood-feedings [2,4]. Vertical transmission can occur when infected female mosquitoes transmit vector-borne pathogens to their offspring through their eggs though the epidemiological significance of this mode of transmission remains unclear [5]. There are over 2,500 different species of mosquitoes throughout the world, however, the mosquito genera that are most associated with the spread of VBD are Anopheles (subfamily Anophelinae), Culex (subfamily Culicinae) and Aedes (subfamily Culicinae). A. aegypti mosquitoes in particular are responsible for the transmission of many important Public Health pathogens, including Yellow Fever, Zika, Chikungunya and Dengue. A. aegypti mosquitoes were first introduced to the American continent during the slave trade in the 1600s and spread worldwide when the shipping industry expanded [2]. This species of mosquito lives in intimate association with humans, preferring to live within human dwellings as they primarily feed on humans and generally breed in dark-colored, man-made water containers [2]. While the average female mosquito lives for approximately 4-6 weeks depending on the environmental conditions, male mosquitoes have significantly shorter lifespans (8-10 days).
The maintenance of a relatively balanced ratio between females and males (1:1) is a fundamental aspect of the population dynamics in sexually reproducing species [6] and several mechanisms contribute to their maintenance at or near parity [7]. Fisher’s principle suggests that, in a sexually reproducing species, the natural selection process favors a balanced sex ratio [8]. When one sex becomes rarer, it has a higher rate of mating success leading to a selective advantage for producing offspring of the rarer sex [9] and a re-establishment of a 1:1 sex ratio within the population [10,11]. Additionally, certain environmental factors can differentially affect the development of males and females [9] by favoring the survival or reproduction of one sex over the other as well as influencing the mortality or fertility of one sex over the other [12]. Density-dependent processes, like the prevalence of predators or the introduction of disease-causing pathogens, can also preferentially select for the prevalence of one sex over the other [9], ultimately leading to distorted sex ratios within the population [13].
The successful implementation of biocontrol strategies to combat the spread of VBD requires a deeper understanding of the pathogen [14,15], the natural behaviors of their vectors [16], as well as the local environmental factors [17] that can be manipulated to break the disease transmission cycle with minimal unintended consequences on non-target organisms or local ecosystems [18]. El Salvador is characterized by tropical dry forests [19,20], consisting of deciduous forests where most tree species shed their leaves during the drought season, and floristically distinct gallery forests where most tree species retain their leaves throughout the year [21]. Seasonally dry tropical forests, such as those in El Salvador, are particularly sensitive to changes in climate, including variations in temperature and precipitation patterns. Studying these ecosystems allows us to observe and understand how climate change influences vegetation composition, arthropod species distributions, and ecosystem dynamics [22]. Dry forests are often more vulnerable to ecological disturbances, such as droughts and wildfires, compared to other tropical ecosystems, providing valuable insights into their resilience and adaptability in the face of environmental change [23,24].
Given that the context-dependent response of interspecific interactions to global climate change remains poorly understood [25,26], we examined the urban ecology of an important mosquito vector in a seasonally dry tropical environment. Our research objective was to analyze the metadata collected by the Salvadoran Ministry of Health and curated by the Center for Ecohealth Research (CENSALUD) at the University of El Salvador to explore the ecological factors that influence the population dynamics of A. aegypti mosquitoes. Specifically, we hypothesized that seasonality would help explain sex ratio biases in A. aegypti populations and provide valuable insight for the identification of context-dependent mitigation strategies to help combat the recent rise in VBD incidence within El Salvador. Our study underscores the importance of understanding vector ecology in the context of a dynamic urban environment as part of continued efforts to implement effective biocontrol strategies in the Northern Triangle of Central America.
2. Materials and Methods
2.1. Mosquito Collection
A total of 150 A. aegypti mosquitoes were collected from the Department of Morazan. Sampling of 50 mosquitoes from the municipality of San Carlos (collected June 28th, 2022) in the hamlet of Barrio El Centro (13°38’48.5"N 88°05’47.4"W), 48 mosquitoes from the municipality of Jocoro (collected July 2nd, 2022) in the hamlet of Barrio San Sebastian (13°36’56.3"N 88°01’21.3"W), and an additional 52 mosquitoes from the municipality of Jocoro (collected July 16th, 2022) in the hamlet of Las Marias Centro (13°37’08.1"N 87°58’56.1"W). Moreover, a total of 150 A. aegypti mosquitoes were collected from the Department of La Union. Sampling of 50 mosquitoes from the municipality of Santa Rosa de Lima (collected July 28th, 2022) in the hamlet of El Limón (13°36’25.8"N 87°55’47.3"W), 50 mosquitoes from the municipality of Pasaquina (collected July 9th, 2022) in the hamlet of Santa Clara (13°35’12.4"N 87°46’47.2"W), and an additional 50 mosquitoes from the municipality of San José la Fuente (collected September 20th, 2022) in the hamlet of Barrio El Calvario (13°33’33.2"N 87°54’16.4"W). An additional 187 A. aegypti mosquitoes were collected from the Department of Usulutan. All these mosquitoes were sampled from the municipality of Ereguayquin (collected March 14th, 2021) in an unnamed hamlet (13°20.5880´N 88°23.3880´W). Furthermore, a total of 89 A. aegypti mosquitoes were obtained from the Department of San Miguel. These mosquitoes were all sampled from the municipality of San Rafael Oriente (collected July 6th, 2021) in the hamlet of Barrio San Benito (13°22.8290’N 88°21.2050’W).
2.2. Sex Ratio Determination
The sex of A. aegypti mosquitoes was determined by examination of specific morphological features, including the proboscis, palps, abdomen, and antennae [27,28]. Female mosquitoes were identified by their distinct elongated proboscis, which is uniquely adapted to pierce the skin of animals to facilitate blood-feeding. In contrast, male mosquitoes have a weaker, less prominent proboscis that is more suited for drinking nectar. The palps, sensory appendages situated immediately in front of the antennae, offered another distinguishing morphological sex characteristic. In females, these palps are notably shorter and are often positions near the proboscis. Conversely, males are equipped with longer, larger, feathery palps. Male mosquitoes also have bushier and more intricate antennae, which are essential sensory organs for detecting the wingbeat frequencies of females during the mating process as well as changes in carbon dioxide levels. Additional morphological distinctions are evident in the abdomen. Female mosquitoes typically have a larger and more rounded abdomen, particularly when gravid. This abdominal enlargement could take on a brownish hue if the female had recently taken a blood meal. In males, the abdomen is thinner and has a streamlined appearance devoid of the engorgement seen in females resulting from recent blood-feeding or egg development.
2.3. Climate Diagram
We generated a climate diagram for San Salvador, El Salvador by utilizing temperature and precipitation data collected between 1981-2010 from CHELSA V2.1 [29]. This climate data was also used to determine the Köppen-Geiger Classification [20] and the Holdridge Life Zone [19]. Changes in the observed sex ratios of male to female (M:F) A. aegypti mosquitos were evaluated relative to an expected homogenized sex ratio (1:1) using a 1-Factor Chi-Square analysis. Additionally, the percent sampling abundance for both male and female mosquitoes relative to all A. aegypti collected across all samples was evaluated relative to an expected homogenized percent sampling distribution (12.5%).
3. Results
The 1981-2010 climate diagram of San Salvador, El Salvador revealed a distinct seasonal pattern with a dry season lasting approximately 6 months (November - April), and a wet season spanning approximately 6 months (May - October) whereby water-saturation of soils persists for at least 5 months (Figure 1). Climate data confirmed that the Köppen-Geiger Classification for this geographic region corresponds to a tropical wet and dry climate (As/Aw), characterized by dry winters and summers. The climate data also established that the Holdridge Life Zone classification was that of a Tropical Dry Forest.
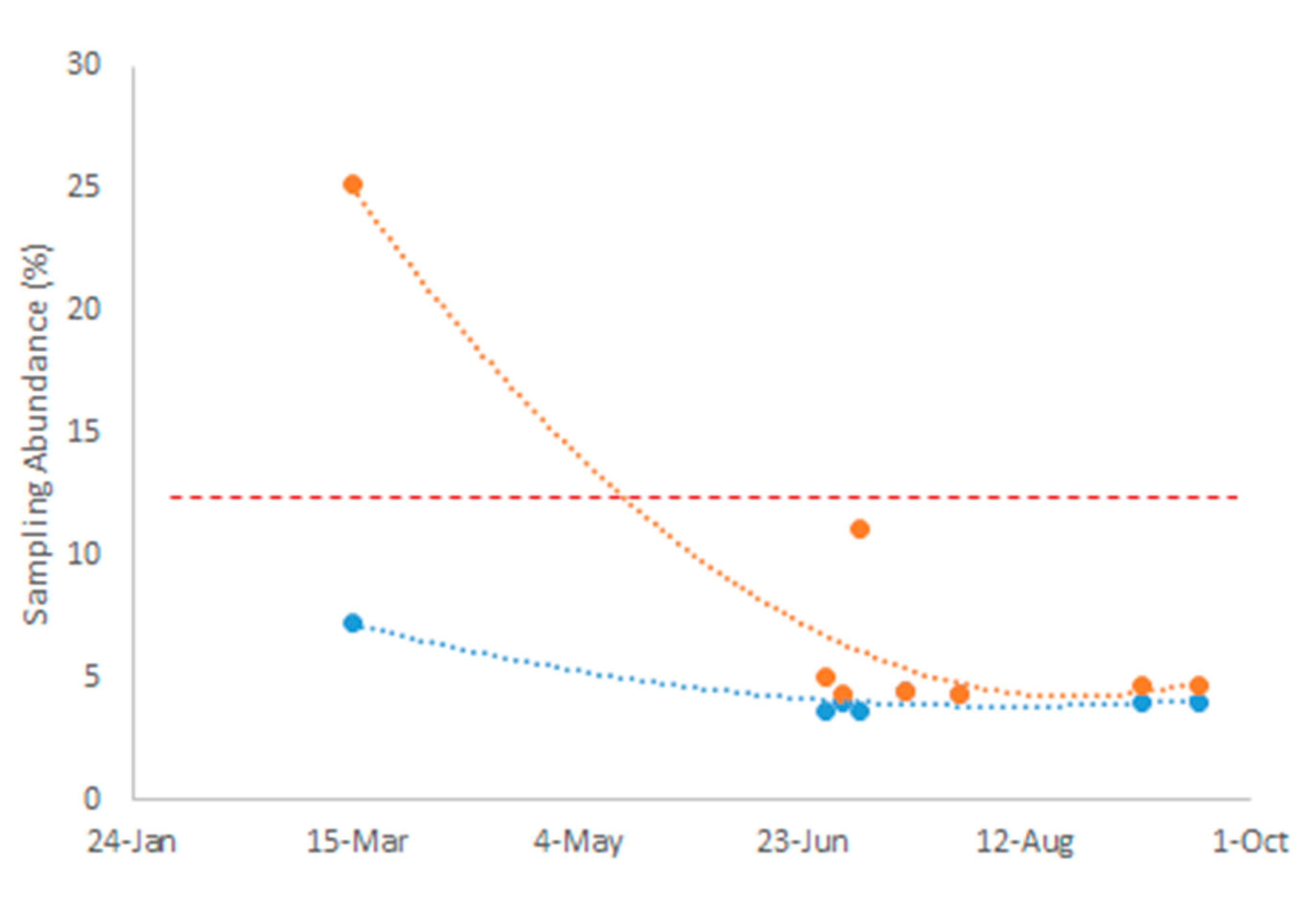
The sex ratio of A. aegypti mosquitoes observed during the sampling period (March 2021 - September 2022) significantly deviated from parity (χ² = 108.2, p value << 0.001), favoring female mosquitoes during the dry season (March) with fewer males. However, the sex ratio shifted during the wet season (June - October), approaching parity between male and female mosquitoes (Figure 2). Additionally, the proportion of male A. aegypti mosquitoes collected during the sampling period relative to the total mosquitoes sampled differed significantly from the expected proportional abundance (χ² = 42.5, p value << 0.001). We found that the observed proportional abundance of males remained below the expected value throughout the dry and wet season sampling periods (Figure 3). Interestingly, the proportion of female A. aegypti mosquitoes collected relative to the total mosquitoes sampled also differed significantly from the expected proportional abundance (χ² = 43.0, p value << 0.001). Although our analysis showed that there were more female mosquitoes than expected during the dry season (March), while female representation fell below expected proportional abundance during the wet season (June - September) (Figure 3). Collectively, our metanalysis showed evidence of a seasonal shift in A. aegypti mosquito sex ratios that favored the abundance of female mosquitoes during the dry season of a highly urbanized environment within El Salvador.
4. Discussion
Understanding of the natural selection mechanisms that influence sex ratios in mosquito populations has been leveraged before as an effective biocontrol strategy [30]. This is an important consideration as disease prevention and vector control mechanisms are the principal methods for managing the spread of VBDs given that there are limited curative therapies and vaccines available [2,31]. Generally, biocontrol mechanisms limit the transmission of vector-borne pathogens by reducing or eliminating human contact with their vectors and there are a wide range of chemical- and non-chemical-based tools available [2,3,30,31]. For example, immature vectors stages can be targeted using chemical or biological larvicides, removing suitable aquatic habitats (either through habitat modifications or manipulations) or through the introduction of predator species [3,32,33,34]. Adult vectors are generally eliminated using chemical sprays within and around human dwellings as well as by reducing human contact with vectors using topical insect repellents, screen doors and insecticide-treated bed nets [3]. However, recent biocontrol mechanisms have employed alternative strategies that require a molecular understanding of VBD pathogenesis and the dynamics of vector ecology, including the introduction of genetically engineered mosquitoes or the use of Wolbachia-infected mosquitoes that cannot support the growth of vector-borne pathogens [2,3,5,35].
Sterile insect techniques are actively being used as a method of biocontrol that exploits the mating behavior and relatively stable sex ratios within natural A. aegypti mosquito populations [2,36,37]. Briefly, male mosquitoes are sterilized through radiation or genetic modification (e.g. introduction of a dominant lethal gene) before being released into the natural mosquito population [2,35,38]. When these sterile males mate with wild A. aegypti females, no viable offspring are produced, leading to a decrease in the mosquito population over time [2]. The advantage of this form of biocontrol is that it facilitates the rapid reduction of mosquito populations though the shortened lifespan of male mosquitoes means this form of vector control is unsustainable and limited to a single breeding cycle. Long-term, sustainable examples of biocontrol include the purposeful infection of A. aegypti mosquitoes with Wolbachia, a natural bacterial pathogen that commonly parasitizes insects [2,39]. Wolbachia bacteria can induce a natural mechanism known as cytoplasmic incompatility (CI) within infected A. aegypti mosquitoes [40], reducing the lifespan of female mosquitoes and increasing their resistance to infection with certain vector-borne pathogens, including the Plasmodium parasites that cause malaria [41,42] and arboviruses like Dengue or Chikungunya [41]. When released into a natural mosquito population, Wolbachia infected mosquitoes can spread the bacterial infection to their mating partners [5] leading to a reduction in reproductive success over time as well as introducing skewed sex ratios that ultimately reduce the mosquito population within that geographic region [39,40,43]. However, the timing for and geographic distribution of these biocontrol mechanisms should consider the ecological and anthropogenic factors that influence mosquito mating behaviors and sex ratios to maximize efforts [2,44].
Our analysis of climatic patterns in El Salvador from 1981 to 2010 revealed a distinct six-month cycle between wet and dry seasons (Figure 1). The dry season persisted from November through April, followed by a wet season spanning May through October, during which conditions of water-saturated soils persisted for at least five months. This marked seasonality helped explain significant changes in sex ratios of A. aegypti mosquito populations in El Salvador. We observed that sex ratios shifted in favor of female A. aegypti mosquitoes during the dry season and then approached parity throughout the wet season (Figure 2). Interestingly, male A. aegypti mosquitoes persisted proportionally below expected abundance year-round while female A. aegypti mosquitoes significantly rose above expected abundance, specifically during the dry season (Figure 3). Our findings underscore the dynamic nature of A. aegypti mosquito populations in seasonally dry neotropical biomes, emphasizing the need for a nuanced approach to vector control that accounts for these seasonal shifts in mosquito sex ratios.
Previous studies have emphasized the low abundance of mosquitoes in seasonally dry tropical biomes during the dry season [45,46], with mosquitoes actively seeking natural and man-made refuges during this period [47,48]. In El Salvador, the intricate interplay between anthropogenic activities and high population density assumes critical importance, especially regarding the abundance of man-made water storage structures that serve as mosquito habitats in urban environments. Boasting the highest population density in Central America, reaching approximately 304 inhabitants/km2 in 2021 [49], the demand for water in this region intensifies during the dry season [50]. While 95.8% of the urban population in El Salvador has access to drinking water, 58% of the overall population lacks water services, relying on natural water sources like rivers, lakes, and rainwater [51]. Consequently, a deeply ingrained practice among Salvadorans involves the domiciliary storage of water in plastic barrels or concrete structures, inadvertently creating potential habitats for A. aegypti mosquitoes [52]. Notably, domiciliary water storage volumes increase during the dry season when water scarcity peaks in natural water sources [51,52], providing the environmental conditions necessary to sustain mosquito breeding cycles near or within human dwellings when female A. aegypti mosquito abundance is high (Figure 2 and Figure 3).
It is important to note that while female mosquitoes engage in blood-feeding, male mosquitoes primarily feed on nectar sources. The scarcity of floral resources during the dry season may further influence the observed ecological dynamics of male mosquitoes, resulting in their low abundance during the dry season in El Salvador (Figure 2 and Figure 3). Given that our study revealed a sex ratio distortion with linkages to anthropogenic factors during a season typically characterized by lower mosquito abundance and dependence on refugia, we propose that developing effective biocontrol strategies for disease transmission mitigation in these seasonally dry tropical biomes would significantly benefit from a comprehensive ecological understanding of the domestic habits of anthropophilic vector species [53]. Specifically, considerations of seasonality, climate variability, and human behaviors influencing mosquito habitats would collectively contribute to more resilient and sustainable approaches to combat VBDs in the face of ongoing global climate changes [54,55]. Ongoing efforts to utlize sterile insect techniques to reduce the spread of VBD should consider how the seasonality of A. aegypti mosquito sex ratios will affect the implementation of proposed biocontrol programs within this region of Central America as the release of Wolbachia-infected or sterile male mosquitoes might be more effective at reducing the incidence of VBD during the dry season (Figure 2 and Figure 3).
Future investigations should further characterize the epidemiological implications of the observed sex ratio bias toward female A. aegypti mosquitoes during the dry season in El Salvador. This involves assessing the prevalence of disease-causing pathogens in female mosquitoes during this period and exploring their potential impact on disease transmission dynamics within local human populations. Such research will inform more nuanced biocontrol approaches as well as enhance our understanding of the intricate relationship between mosquito vector ecology in highly urbanized environments and the transmission of VBDs in seasonally dry neotropical biomes. Ultimately, effective control of VBD throughout the Northern Triangle of Central America will require the implementation of several biocontrol mechanisms as well as active surveillance of vector-borne pathogens within local A. aegypti populations. Given the recent rise in VBD incidence worldwide, a comprehensive understanding of the ecology of A. aegypti vectors and the spacial distribution of the pathogens they transmit is vital for the development of sustained biocontrol mechanisms within El Salvador.
Author Contributions
Conceptualization, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; methodology, A.M.G.P.; software, V.D.C.-G.; validation, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; formal analysis, V.D.C.-G.; investigation, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; resources, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; data curation, A.M.G.P. and V.D.C.-G.; writing—original draft preparation, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; writing—review and editing, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; visualization, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; supervision, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; project administration, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.; funding acquisition, A.M.G.-C., M.A.V.C., A.M.G.P., and V.D.C.-G.. All authors have read and agreed to the published version of the manuscript.
Funding
A.M.G.-C. was funded through the Fulbright U.S. Scholar program at the United States Department of State.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author [A.M.G.-C.] upon reasonable request.
Acknowledgments
We would like to express our sincere gratitude to the Salvadoran Ministry of Health for their invaluable support of scientific endeavors that contribute to a better understanding of the population dynamics of disease vectors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Organization, W.H. Vector-borne diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 28 November 2023).
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat Rev Dis Primers 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl Trop Dis 2020, 14, e0007831. [Google Scholar] [CrossRef]
- Pimenta, P.F.; Touray, M.; Miller, L. The journey of malaria sporozoites in the mosquito salivary gland. J Eukaryot Microbiol 1994, 41, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Ooi, E.E.; Vasudevan, S.; Farrar, J. Dengue and Dengue Hemorrhagic Fever, 2nd Edition; CABI: 2014.
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press: 1982.
- Hamilton, W.D. Extraordinary sex ratios. A sex-ratio theory for sex linkage and inbreeding has new implications in cytogenetics and entomology. Science 1967, 156, 477–488. [Google Scholar] [CrossRef]
- Fisher, R.A. The genetical theory of natural selection: a complete variorum edition; Oxford University Press: 1999.
- A. W. F. Edwards. Natural Selection and the Sex Ratio: Fisher’s Sources. The American Naturalist 1998, 151, 564–569. [CrossRef]
- Andrew Olaf Shelton. The Ecological and Evolutionary Drivers of Female-Biased Sex Ratios: Two-Sex Models of Perennial Seagrasses. The American Naturalist 2010, 175, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Pérez, I.d.L.G.; Carretero, M.A.; Font, E. Intensity of male-male competition predicts morph diversity in a color polymorphic lizard. Evolution 2017, 71, 1832–1840. [Google Scholar] [CrossRef]
- Barrett, S.C.; Yakimowski, S.B.; Field, D.L.; Pickup, M. Ecological genetics of sex ratios in plant populations. Philos Trans R Soc Lond B Biol Sci 2010, 365, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Vergés, A.; Paul, N.A.; Steinberg, P.D. Sex and life-history stage alter herbivore responses to a chemically defended red alga. Ecology 2008, 89, 1334–1343. [Google Scholar] [CrossRef]
- Groat-Carmona, A.M.; Orozco, S.; Friebe, P.; Payne, A.; Kramer, L.; Harris, E. A novel coding-region RNA element modulates infectious dengue virus particle production in both mammalian and mosquito cells and regulates viral replication in Aedes aegypti mosquitoes. Virology 2012, 432, 511–526. [Google Scholar] [CrossRef]
- Groat-Carmona, A.M.; Kain, H.; Brownell, J.; Douglass, A.N.; Aly, A.S.I.; Kappe, S.H.I. A Plasmodium α/β-hydrolase modulates the development of invasive stages. Cellular Microbiology 2015, 17, 1848–1867. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Callahan, A.G.; Yang, Q.; Jasper, M.; Arif, M.A.K.; Afizah, A.N.; Nazni, W.A.; Hoffmann, A.A. An elusive endosymbiont: Does Wolbachia occur naturally in Aedes aegypti? Ecology and Evolution 2020, 10, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Xiong, J.; Lu, D.; Wang, G.; Ai, H. Transcriptome Sequencing Analysis and Functional Identification of Sex Differentiation Genes from the Mosquito Parasitic Nematode, Romanomermis wuchangensis. PLOS ONE 2016, 11, e0163127. [Google Scholar] [CrossRef]
- Simberloff, D. Risks of biological control for conservation purposes. BioControl 2012, 57, 263–276. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of World Plant Formations From Simple Climatic Data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Bullock, S.H.; Mooney, H.A.; Medina, E. Seasonally Dry Tropical Forests; Cambridge University Press: Cambridge, 1995. [Google Scholar]
- Dexter, K.G.; Pennington, R.T.; Oliveira-Filho, A.T.; Bueno, M.L.; Silva de Miranda, P.L.; Neves, D.M. Inserting Tropical Dry Forests Into the Discussion on Biome Transitions in the Tropics. Frontiers in Ecology and Evolution 2018, 6. [Google Scholar] [CrossRef]
- Pablo-Cea, J.D.; Velado-Cano, M.A.; Noriega, J.A. A first step to evaluate the impact of ecotourism on biodiversity in El Salvador: a case study using dung beetles in a National Park. Journal of Ecotourism 2021, 20, 51–69. [Google Scholar] [CrossRef]
- Szabó, B.; Korányi, D.; Gallé, R.; Lövei, G.L.; Bakonyi, G.; Batáry, P. Urbanization decreases species richness, and increases abundance in dry climates whereas decreases in wet climates: A global meta-analysis. Science of The Total Environment 2023, 859, 160145. [Google Scholar] [CrossRef]
- Slominski, A.H.; Burkle, L.A. Solitary bee life history traits and sex mediate responses to manipulated seasonal temperatures and season length. Frontiers in Ecology and Evolution 2019, 7, 314. [Google Scholar] [CrossRef]
- Vides-Hernández, G.L.; Velado-Cano, M.A.; Pablo-Cea, J.D.; Carmona-Galindo, V.D. Patrones de riqueza y diversidad de aves en áreas verdes del centro urbano de San Salvador, El Salvador. Huitzil 2017, 18, 272–280. [Google Scholar] [CrossRef]
- de Araújo, H.R.C.; Kojin, B.B.; Capurro, M.L. Sex determination and Aedes population control. Parasites & Vectors 2018, 11, 644. [Google Scholar] [CrossRef] [PubMed]
- Shettima, A.; Joseph, S.; Ishak, I.H.; Abdul Raiz, S.H.; Abu Hasan, H.; Othman, N. Evaluation of Total Female and Male Aedes aegypti Proteomes Reveals Significant Predictive Protein–Protein Interactions, Functional Ontologies, and Differentially Abundant Proteins. Insects 2021, 12, 752. [Google Scholar] [CrossRef] [PubMed]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Scientific Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Robinson, A.S. Sex-ratio manipulation in relation to insect pest control. Annu Rev Genet 1983, 17, 191–214. [Google Scholar] [CrossRef]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological Control of Mosquito Vectors: Past, Present, and Future. Insects 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Lu, L.; Fu, G.; Zhong, S.; Ding, G.; Xu, R.; Zhu, G.; Shi, N.; Fan, F.; Liu, Q. Epidemiology and vector efficiency during a dengue fever outbreak in Cixi, Zhejiang Province, China. Journal of vector ecology 2009, 34, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E. The sex structure of the larval populations of the urban mosquito Culex pipiens pipiens f. molestus Forskal (Diptera, Culicidae) in St. Petersburg. Entomological review 2011, 91, 729–734. [Google Scholar] [CrossRef]
- Mwingira, V.; Mboera, L.E.; Dicke, M.; Takken, W. Exploiting the chemical ecology of mosquito oviposition behavior in mosquito surveillance and control: a review. Journal of Vector Ecology 2020, 45, 155–179. [Google Scholar] [CrossRef]
- Gubler, D.J. The partnership for dengue control - a new global alliance for the prevention and control of dengue. Vaccine 2015, 33, 1233. [Google Scholar] [CrossRef]
- Lance, D.; McInnis, D. Biological basis of the sterile insect technique. Sterile Insect Technique 2021, 113–142. [Google Scholar]
- Hendrichs, J.; Robinson, A. Sterile insect technique. In Encyclopedia of insects; Elsevier: 2009; pp. 953–957.
- Benedict, M.Q.; Robinson, A.S. The first releases of transgenic mosquitoes: an argument for the sterile insect technique. Trends in parasitology 2003, 19, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Jayachandran, S.; Prabagaran, S.R. Evidence for the natural occurrence of Wolbachia in Aedes aegypti mosquitoes. FEMS microbiology letters 2019, 366, fnz055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zheng, X.; Xi, Z.; Bourtzis, K.; Gilles, J.R. Combining the sterile insect technique with the incompatible insect technique: I-impact of Wolbachia infection on the fitness of triple-and double-infected strains of Aedes albopictus. PloS one 2015, 10, e0121126. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Moreira, L.A. Can Wolbachia be used to control malaria? Memórias do Instituto Oswaldo Cruz 2011, 106, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Zhang, D.; Lees, R.S.; Xi, Z.; Bourtzis, K.; Gilles, J.R. Combining the sterile insect technique with the incompatible insect technique: III-robust mating competitiveness of irradiated triple Wolbachia-infected Aedes albopictus males under semi-field conditions. PloS one 2016, 11, e0151864. [Google Scholar] [CrossRef] [PubMed]
- Jawara, M.; Pinder, M.; Drakeley, C.J.; Nwakanma, D.C.; Jallow, E.; Bogh, C.; Lindsay, S.W.; Conway, D.J. Dry season ecology of Anopheles gambiae complex mosquitoes in The Gambia. Malaria Journal 2008, 7, 1–9. [Google Scholar] [CrossRef]
- Minakawa, N.; Githure, J.I.; Beier, J.C.; Yan, G. Anopheline mosquito survival strategies during the dry period in western Kenya. Journal of medical entomology 2001, 38, 388–392. [Google Scholar] [CrossRef]
- Charlwood, J.; Vij, R.; Billingsley, P. Dry season refugia of malaria-transmitting mosquitoes in a dry savannah zone of east Africa. The American journal of tropical medicine and hygiene 2000, 62, 726–732. [Google Scholar] [CrossRef]
- Animut, A.; Negash, Y. Dry season occurrence of Anopheles mosquitoes and implications in Jabi Tehnan District, west Gojjam Zone, Ethiopia. Malaria Journal 2018, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bank, W. Population density in El Salvador from 2011 to 2021 (in number of inhabitants per square kilometer). Available online: https://www-statista-com.laverne.idm.oclc.org/statistics/882993/population-density-el-salvador/ (accessed on 25 November 2023).
- Nazari, B.; Keshavarz, M. Water population density: Global and regional analysis. Theoretical and Applied Climatology 2023, 1–15. [Google Scholar] [CrossRef]
- Fernández, D.M., Alfredo; Sarmanto, Natalia. Diagnóstico de la prestación de los servicios de agua potable y alcantarillado en El Salvador.; 2023; p. 25 p.
- Joyce, A.L.; Torres, M.M.; Torres, R.; Moreno, M. Genetic variability of the Aedes aegypti (Diptera: Culicidae) mosquito in El Salvador, vector of dengue, yellow fever, chikungunya and Zika. Parasites & vectors 2018, 11, 1–14. [Google Scholar]
- Choi, J.; Carmona-Galindo, V.; Paredes, G.R.; Marín Recinos, M.F.; de Abrego, V.C. An exploration of land use and poverty as an integrative model for mitigating Chagas disease in El Salvador. BIOS 2020, 91, 125–132, 128. [Google Scholar] [CrossRef]
- Benelli, G. Managing mosquitoes and ticks in a rapidly changing world–Facts and trends. Saudi Journal of Biological Sciences 2019, 26, 921–929. [Google Scholar] [CrossRef]
- Ferraguti, M.; Martínez-de La Puente, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effects of landscape anthropization on mosquito community composition and abundance. Scientific reports 2016, 6, 29002. [Google Scholar] [CrossRef]
Figure 1.
Climate diagram of San Salvador, El Salvador (1981-2010).
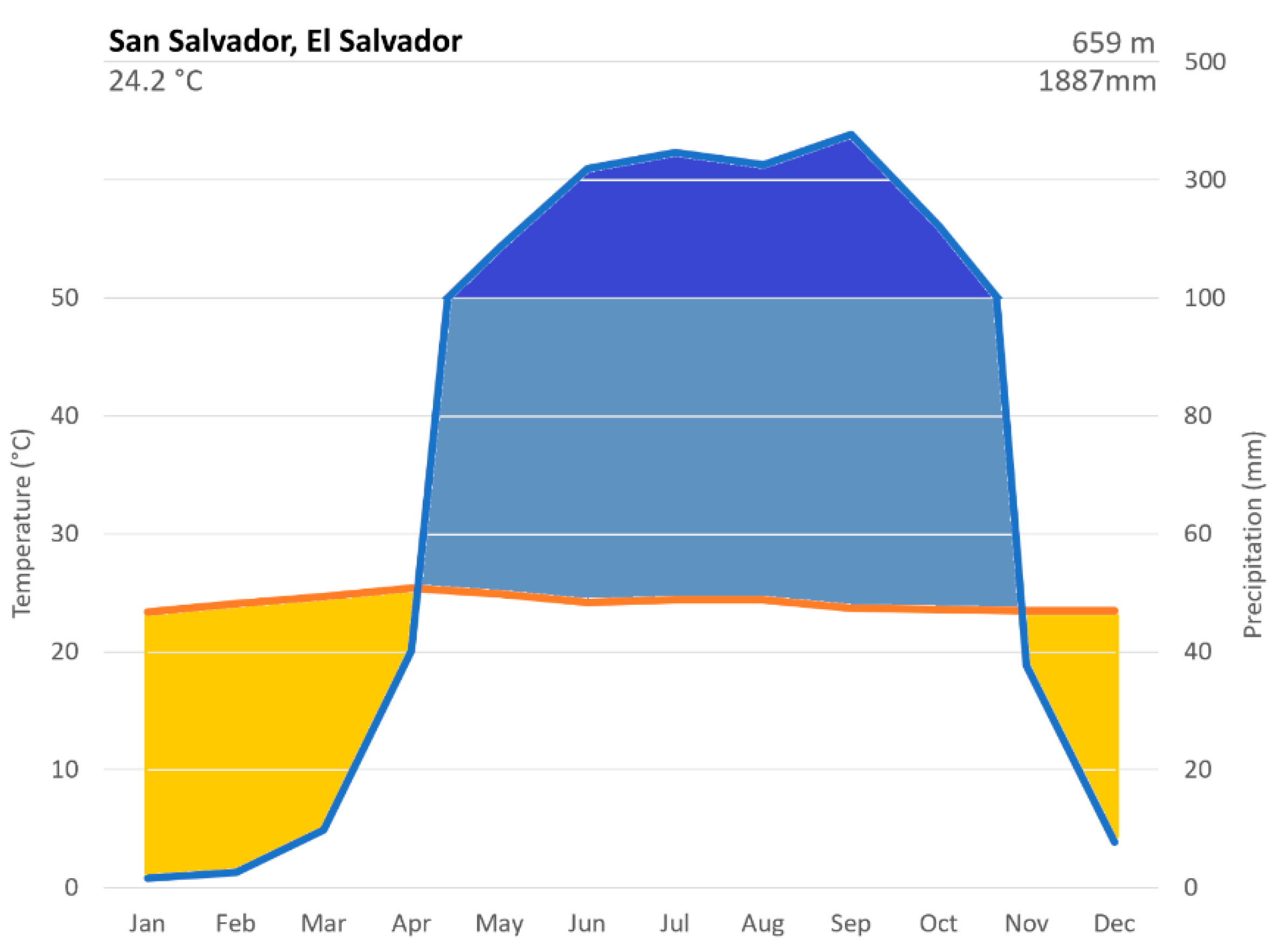
Figure 2.
Observed temporal changes to sex ratio in mosquitoes (blue) relative to an expected parity (red).
Figure 2.
Observed temporal changes to sex ratio in mosquitoes (blue) relative to an expected parity (red).
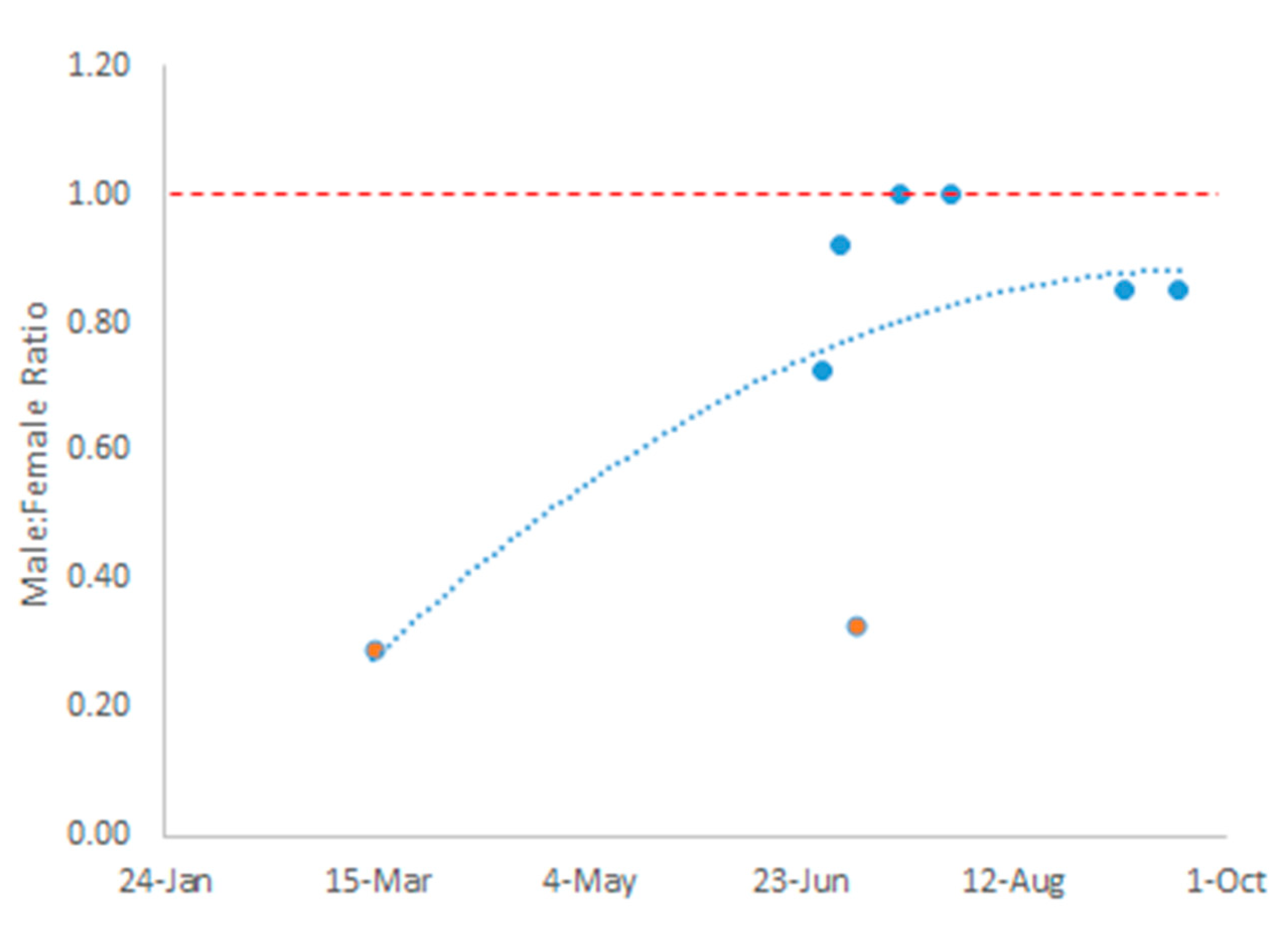
Figure 3.
Observed temporal changes to the proportional abundance of male (blue) and female (orange) mosquitoes relative to the expected sampling abundance (red).
Figure 3.
Observed temporal changes to the proportional abundance of male (blue) and female (orange) mosquitoes relative to the expected sampling abundance (red).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



