
Submitted:
01 December 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
Furunculosis, caused by Aeromonas salmonicida, poses a significant threat to both salmonid and non-salmonid fish in diverse aquatic environments. This study explores the genomic intricacies of re-emergent A. salmonicida outbreaks in Atlantic salmon (Salmo salar). Previous clinical cases have exhibited pathological characteristics, such as periorbital hemorrhages and gastrointestinal abnormalities. Genomic sequencing of three Chilean isolates (ASA04, ASA05, and CIBA_5017) and 25 previously described genomes determined the pan-genome, phylogenomics, insertion sequences, and restriction-modification systems. Unique gene families have contributed to an improved understanding of the psychrophilic and mesophilic clades, while phylogenomic analysis has been used to identify mesophilic and psychrophilic strains, thereby further differentiating between typical and atypical psychrophilic isolates. Diverse insertion sequences and restriction-modification patterns have highlighted genomic structural differences and virulence factor predictions can emphasize exotoxin disparities, especially between psychrophilic and mesophilic strains. Thus, a novel plasmid was characterized, which emphasized the role of plasmids in virulence and antibiotic resistance. The analysis of antibiotic resistance factors revealed resistance against various drug classes in Chilean strains. Overall, this study elucidates the genomic dynamics of re-emergent A. salmonicida and provides novel insights into their virulence, antibiotic resistance, and population structure. However, rigorous monitoring and control in aquaculture remain crucial to mitigate the spread of antibiotic resistance.
Keywords:
furunculosis
; Aeromonas salmonicida
; Salmo salar
; genomic diversity
1. Introduction
Furunculosis is a bacterial disease that is caused by Aeromonas salmonicida, which affects various species of salmonid and non-salmonid fish, in both freshwater and marine environments and is characterized by high mortality and morbidity. A. salmonicida is a Gram-negative, facultatively anaerobic, and nonmotile bacterium, which is classified within the genus Aeromonas and belongs to the class Gammaproteobacteria and the family Aeromonadaceae [1]. The bacterium exhibits a rod-shaped morphology, measures approximately 1.3–2.0 by 0.8–1.3 μm, and displays optimal growth at temperatures ranging from 22 to 25 °C. Moreover, it possesses the ability to both ferment and oxidize glucose and exhibits positive results in catalase and cytochrome oxidase tests. It particularly affects salmonids and trout, thereby making it a significant concern in aquaculture. A. salmonicida can be divided into five subspecies: salmonicida, smithia, achromogenes, masoucida, and pectinolytica [2,3,4,5,6,7,8,9]. The A. salmonicida subspecies salmonicida represents the classical causative agent of furunculosis in salmonids, although atypical strains of A. salmonicida such as masoucida, achromogenes, and smithia can cause a variant in non-salmonids and warm-water fish known as atypical furunculosis [10,11,12]. These strains show optimal growth at temperatures ranging from 22 to 25 °C. Notably, among the known subspecies of A. salmonicida, only A. salmonicida pectinolytica has been observed growing at temperatures exceeding 35 °C.
The first report of A. salmonicida atypica isolates in Atlantic salmon (S. salar) cultured in Chilean seawater was [13]. Subsequently, atypical furunculosis spread rapidly in estuarine, marine, and freshwater fish farms in 2000, with the freshwater stage of the disease only being reported in the localities of Osorno and Punta Arenas in the fall of 2009 [14]. In 2001, an injectable intraperitoneal vaccination containing inactivated A. salmonicida atypica was initiated in Atlantic salmon (S. salar), which reduced disease prevalence. However, despite vaccination becoming an established strategy in salmon farms, sporadic outbreaks continue to occur in Atlantic salmon (S. salar) in lakes associated with vaccination management, estuary fattening, and endemic fish farms. Moreover, between 2021 and 2023, the prevalence of clinical cases in fish farms using recirculating aquaculture technology (RAS) increased among unvaccinated fish groups.
Recently, various aspects contributing to the genomic diversity of A. salmonicida have been described. These distinctive patterns appear to be primarily associated with the accessory genome, thereby enabling differentiation between psychrophilic and mesophilic clades. The detection of a novel genomic entity, denoted as AsaGEI1a, within the 01-B526 strain has been a pivotal revelation since it encompasses a 51 kb chromosomal island, which contains prophage genes. This entity exhibits distinct variants, AsaGEI1b and AsaGEI2a, which are characterized by specific geographical distributions. Thus, 139 isolates were screened via PCR, and revealing a noteworthy geographical association, whereby AsaGEI1a and AsaGEI2a were predominantly identified in North American isolates. In contrast, AsaGEI1b was prevalent, while there was a conspicuous absence of other common genomic elements (GEIs) in European isolates. The identification of an additional prophage (prophage 3) intricately linked to AsaGEI2a further enriched the understanding of genomic dynamics by demonstrating an exclusive geographic prevalence in North America, with a single exception also found in a European isolate [15]. Moreover, the establishment and implementation of a vapA-centric typing system facilitated the subtyping of 675 isolates of A. salmonicida. This innovative approach led to the identification of nine novel subtypes (15–23), with certain subtypes exhibiting a discernible affinity for specific fish species, notably, Cyprinidae and Pleuronectidae [16]. Furthermore, the sequencing and comparative analyses of entire genomes from A. salmonicida strains provided further insights into the genomic distinctions between typical and atypical strains. The genomic disparities were scrutinized by evaluating the insertion sequences (ISs), along with identifying variations in the presence and absence of genes associated with virulence factors, transcriptional regulators, and non-coding RNAs. Additionally, a discernible role of the plasmidome in both the virulence and genomic adaptability of A. salmonicida has been highlighted [17]. Similarly, a comparative examination of 25 complete A. salmonicida genomes revealed gene clusters exclusive to the psychrophilic clades, which are associated with the lateral flagella, outer membrane proteins (including A-layer and T2SS proteins), and insertion sequences (specifically ISAs4, ISAs7, and ISAs29). In contrast, the mesophilic clade exhibited the exclusive presence of MSH type IV pili, which suggests that discernible genetic factors are linked to the specific lifestyle of A. salmonicida within this particular clade [18]. Furthermore, the comprehensive analysis of extensive genomic data compiled from hundreds of Aeromonas strains in aquaculture has enabled the detailed profiling of resistome patterns and can offer insights into associations by isolation year, country of origin, and species characteristics. Indeed, over 100 antibiotic resistance genes (ARGs) have been identified within a dataset encompassing approximately 400 A. salmonicida, A. hydrophila, A. veronii, A. media, and A. sobria genomes, which display either species-specific or non-species-specific patterns. Notably, species such as A. salmonicida and A. media exhibit higher proportions of species-specific ARGs, indicating distinctive patterns in the acquisition of antibiotic resistance. The prevalence of genes such as sul1, tet(A), and tet(D) has also been determined in specific regions of Asia and North America. Recently, there is a noteworthy surge in ARGs these aquatic environmental strains. This underscores the importance of regulating and controlling the excessive use of antibiotics in aquaculture to mitigate the escalation of antibiotic resistance [19].
Prokaryotic defense mechanisms against foreign genomes involve crucial components known as restriction-modification (R-M) systems, which are present in a diverse range of unicellular organisms, encompassing both eubacteria and archaea [20,21]. These systems include two distinct enzymatic functions: restriction endonuclease (REase) and methyltransferase (MTase) functions. The REase targets and cleaves foreign DNA sequences at specific locations, while MTase activity maintains a distinction between the DNA of the host and any external DNA by adding methyl groups to the same specified DNA sequences. R-M systems exhibit diversity in their classifications, with distinctions based on cleavage position, co-factor requirements, sequence recognition, subunit composition, and substrate specificity, thus resulting in four primary types [22]. However, R-M systems seem to play multifaceted roles in bacterial biology, and their functions extend beyond merely protecting against foreign genetic material, and include modulating horizontal gene transfer (HGT) and differentiating among bacterial strains. Due to the rampant impact of HGT on prokaryotic species boundaries, bacterial species are sustained through genetic isolation. REases operate as regulators by restricting foreign DNA with non-native methylation patterns to control the genetic influx. This creates a barrier, which could also serve to preserve the bacterial species [23]. In this study, we describe the histopathological and genomic profile of re-emergent A. salmonicida outbreaks in Chile. We corroborate recent findings regarding genomic variation patterns among typical psychrophilic, atypical psychrophilic, and mesophilic strains of A. salmonicida by comparing the genomes described in this study with 25 closed genomes available on NCBI [24]. Finally, we describe the R-M landscape among A. salmonicida strains.
2. Materials and Methods
2.1. Sample Collection and Gross Pathology
The investigated fish were obtained from 4 different hatcheries located in the regions of Los Lagos and Magallanes at the pre-smolt and smolt stages. All hatcheries followed a vaccination protocol to combat A. salmonicida during the early stages of cultivation as part of their health strategy and implemented RAS. Fish samples were collected at each hatchery by selecting the tanks with high mortality rates and clinical or laboratory diagnoses in the week prior to the visit. Fish exhibiting at least one of the following symptoms were sampled: erratic or surface swimming, exophthalmia, mycosis, fin base hemorrhages, or skin lesions. Necropsies were performed to confirm the diagnosis and establish pathological characteristics in each case, following the description provided by [25]. Additionally, samples were taken from internal organs (kidney and spleen) for molecular biology analysis (PCR for atypical A. salmonicida), internal organs fixed in formalin were selected for histopathological analysis, and fresh fish samples were sent to the laboratory for culturing and to isolate atypical A. salmonicida onto solid media (Tryptic Soy Agar). For cases where successful bacterial isolation was achieved, genomic DNA (gDNA) was directly extracted from colonies, and whole bacterial genome sequencing (WGS) was performed.
2.2. Histopathology
Tissue samples intended for histological analysis were gathered and preserved in 10% buffered formalin. Following collection, they underwent standard processing procedures. The resulting sections, measuring 3–4 µm in thickness, were subsequently stained with hematoxylin and eosin (H & E) as per the standard protocol [26], to identify the significant microscopic morphological changes.
2.3. DNA Extraction and PCR
Here, automated tissue homogenization was conducted on the samples using the MagNA Lyser instrument (Roche, Basel, Switzerland). Subsequently, a robot was employed to perform total DNA extraction (Roche MagNA Pure LC instrument, Basel, Switzerland) alongside the MagNA Pure LC RNA isolation kit III—Tissue, following the manufacturer’s instructions. The extracted DNA was eluted in 50 µL of nuclease-free water, and DNA yields were quantified, while the DNA purity was analyzed by using a NanoPhotometer® P 300 (Implen, Westlake Village, CA, USA), using the OD260/280 ratio. Following quantitation, the eluted DNA was promptly tested for the detection of A. salmonicida, using thermocycling conditions and specific primers, as previously described [27]. Piscine orthoreovirus (PRV), infectious salmon anemia virus (ISAV), and infectious pancreatic necrosis virus (IPNV) detection was also performed as described previously [28,29,30].
2.4. Bacterial Isolation
The main organs selected for bacterial isolation were the spleen, kidney, liver, and skin. The selected organs were aseptically streaked onto tryptic soy agar (TSA) plates and incubated at 16 °C for 5 days. Genomic material extraction was performed from colonies showing suggestive coloration and morphology, which were subsequently confirmed through PCR for atypical A. salmonicida. gDNA was extracted using the Total DNA kit (Omega-Biotek, Norcross, Georgia), following the manufacturer’s instructions. The purified gDNA concentration was determined using a NanoDrop spectrophotometer. In total, 100 ng of gDNA from each strain was utilized for subsequent library preparations.
2.5. Whole Genome Sequencing (WGS) and Bioinformatic Analysis
The ASA04 and ASA05 strains were sequenced using NanoPore technology. The extracted gDNA was prepared for sequencing using the Rapid Sequencing Kit V14, following the manufacturer’s guidelines: This kit is specifically optimized for the MinION Mk1C sequencer (Oxford Nanopore Technologies, Oxford, UK), thereby ensuring efficient library preparation and optimal sequencing performance. The prepared libraries were loaded onto a FlowCell R10.4.1 and sequenced by using the MinION Mk1C sequencer. The device was calibrated for dual-read sequencing to enhance accuracy by reading both DNA strands. After sequencing, the raw electric signal data from the MinION Mk1C were converted into nucleotide sequences through the Guppy basecalling software. Default parameters were employed, as recommended by the manufacturer. Then, the basecalled reads obtained from Guppy were utilized for genome assembly. Two separate assemblers, Canu [31] and Flye [32], were engaged to ensure that the assembly was accurate and robust. The achieved coverage values for the assembled genomes were 99.73× and 99.04×, respectively.
The CIBA_5017 strain was sequenced by PacBio Technology. The extracted gDNA was prepared for sequencing using the SMRTbell prep kit 3.0. The prepared library was sequenced using the Sequel II sequencer. Using subreads as input, circular consensus sequences (CCS) were generated with a minimum of 3 passes and a minimum predicted accuracy of 0.99 by using the CCS (v.4.2.0) tool available in SMRT Link v9.0. This allows for the resulting CCS reads to have a high accuracy of > 99% and Q > 20. The resulting CCS reads were used as input for downstream analysis. The FASTQ file reads were checked for quality using fastp [33]. The PacBio reads were assembled with Unicycler [34], and the quality of the assemblies was checked using the QUAST tool [35].
Genome annotation and pan-genome analysis were performed using Prokka [36] and PIRATE [37], respectively. A core genome assembly encompassing SNPs was built with Snippy [38], and from it, a maximum likelihood phylogenic tree was built using IQ-TREE [39,40,41]. This tree was visualized using iTOL [42]. Virulence factors, antibiotic resistance factors, insertion sequences, and R-M system enzymes were predicted using the VFDB [43], CARD [44], ISFinder [45], and REBASE [46] databases, respectively, through local alignments with BLAST [47]. Plasmid composition was visualized with Proksee [48].
3. Results
3.1. Laboratory Analysis of Clinical Cases
Overall, 27 clinical cases of atypical furunculosis were reported in Atlantic salmon (S. salar) during 2022 and analyzed in this study. The epidemiological background and laboratory results are summarized in Table 1. These showed an average mortality rate of 4.66% and an outbreak time of 8 weeks. Three bacterial isolates obtained from clinical cases were selected for genome sequencing. All the isolates were from clinically diseased fish, specifically those exhibiting the highest frequency in the clinical findings.
3.2. Gross Pathology
Some of the clinical signs illustrated were decreased appetites, increased lethargy, and exophthalmia (protrusion of the eyeball). Externally, the affected fish may exhibit periorbital hemorrhages (around the eyes), hemorrhages at the base of the fins, and hemorrhages in the abdomen, anus, and the peduncle. Sometimes, the presence of bullae, which are fluctuating areas upon palpation called furuncles, which correspond to muscular necrosis, can also be observed. Further, desquamation, erosion, and/or skin ulcers, whether singular or multiple, individual or coalescent, resulted in exposure of the musculature and edges displaying a white or red coloration. Internally, the affected fish exhibit petechial hemorrhages in the liver, swim bladder, heart, visceral fat, and perirenal hemorrhages; variable findings may include nephromegaly (enlarged kidneys) and splenomegaly (enlarged spleen). Finally, hemorrhagic enteritis and hyperemic gastric mucosa were observed in the gastrointestinal system (Figure 1 and Figure 2).
3.3. Histopathology
Variable colonies of bacteria were found to be replacing the tissue in the affected fish, with limited inflammatory responses. All organs were affected by the bacterial colonies, with the musculature, heart, gills, spleen, kidney, and liver being the most frequently affected tissues. In the skin, necrosis of the epidermis and dermis was observed, which deeply affected the adjacent musculature, with necrosis of the muscle fibers and associated hemorrhages, along with an abundance of bacterial colonies. There was hemorrhagic necrosis in the affected muscles, which led to the formation of cavities containing bloody fluids, sometimes with a viscous consistency (Figure 3). Additionally, the presence of hyperplasia and nephropathy was detected in a variable manner, which was possibly associated with suboptimal water quality parameters. Myocarditis, myositis, and hepatic necrosis were also observed, although these findings can be explained by infections from PRV, the causal agent of skeletal and cardiac muscle disease (HSMI) in Atlantic salmon (S. salar). Furthermore, peritonitis findings were identified, which are potentially linked to secondary reactions resulting from the intraperitoneal vaccination.
3.4. Pan-Genome Characterization
The three genomes analyzed in this study were included alongside the previous pan-genome characterization [18]; the combined analysis predicted 7296 gene families in the 28 genomes analyzed. From these, the core genome was composed of 3129 gene families. The genomes described in this study, ASA04, ASA05, and CIBA_5017, contained 17, 2, and 2 unique gene families, respectively. The analysis of the present/absent gene families showed remarkably distinct patterns between the mesophilic (grow well at 35–37 °C) and psychrophilic (optimum growth at 22–25 °C, with no growth at 37 °C) strains, as well as within the typical and atypical psychrophilic strains. Moreover, the landscape of the present gene families seems to be heterogeneous among the atypical psychrophilic strains (Figure 4).
3.5. Phylogenomics
The obtained maximum likelihood dendrogram shows the grouping of the 28 analyzed A. salmonicida genomes into two main clusters (Figure 5). One is composed of mesophilic strains (red in Figure 5) and is distantly located toward the second cluster—composed of psychrophilic strains. Interestingly, the psychrophilic group is further divided into two subclusters, one composed of typical psychrophilic strains (green in Figure 5), while the second is composed of atypical psychrophilic strains (yellow in Figure 5). The three strains described in this study cluster together within the atypical psychrophilic group.
3.6. Insertion Sequences and Restriction-Modification System Abundance
To further characterize the genomic structure of the genomes analyzed in this study, their insertion sequence (IS) and R-M system repertoires were described. Three main patterns of IS composition were found in the 28 genomes included in the analysis, as described elsewhere [18]. This pattern shows a trend indicating that the mesophilic, typical psychrophilic, and atypical psychrophilic strains include clear differences in their IS composition. The three genomes described in this study fall within the expected pattern of atypical psychrophilic strains, whereby the IS5 family is the most abundant. Notably, the ASA04 and ASA05 genomes show a higher number of insertion sequences compared to other atypical psychrophilic strains (Supplementary Figure S1). Furthermore, distinctive patterns of R-M coding sequences were found among the mesophilic, typical psychrophilic, and atypical psychrophilic strains (Figure 6). Remarkably, contrasting patterns appeared in the ISs and R-M coding sequences observed in the atypical psychrophilic strains, with respect to other strains. The IS number is more abundant in the atypical psychrophilic strains compared to the mesophilic strains. In contrast, the number of R-M coding sequences identified in the atypical psychrophilic strains is less abundant than in the mesophilic strains. Moreover, two distinctive patterns of R-M coding sequences are evident within the mesophilic strains, as well as among the atypical psychrophilic strains.
3.7. Virulence Factors
Virulence factors (VFs) were predicted in the genomes included in the present study and compared with the complete A. salmonicida genomes available from NCBI [24]. The presence of virulence factor classes was mostly homogeneous between the compared strains (Supplementary Figure S2). However, regarding exotoxin factors, the mesophilic strains showed the presence of actin cross-linking domain (ACD)-containing toxin factors, which were absent in the psychrophilic strains (Figure 7). Additionally, minor differences were observed regarding the pattern of exotoxin factors between the typical and atypical psychrophilic isolates. Specifically, the DNAse/genotoxin factor clbF was present in all typical psychrophilic but not in the atypical psychrophilic isolates, except for in samples J410, J411, and J409. Moreover, the pore-forming toxin factor aerA/act is present in most typical psychrophilic strains and in all mesophilic isolates, yet was not in the atypical psychrophilic samples. Furthermore, the RNA N-glycosidase factor stx2eA highlights the differences within the atypical psychrophilic strains since it is present only in the S44, S121, S68, BR1900YR, RFAS1, AS1, AS2, and RZ6S-1 isolates.
3.8. Characterization of Plasmids
All three genomes described in this study contained plasmids with distinct factor compositions (Figure 8). Genomes ASA04 and ASA05 contained two plasmids each, while genome CIBA_5017 contained one plasmid. However, the total number of unique plasmids was three since these were subsequently found to be redundant. Plasmid p01CIBA_5017 was found in the ASA04, ASA05, and CIBA_5017 strains. This plasmid was identified as the A. salmonicida strain S44 plasmid pS44-3. In contrast, plasmid p02ASA04 was only found in ASA04, and after performing a BLASTn search against the NCBI database, this plasmid produced hits illustrating a maximum query coverage of 53%, which suggests that it has not been described previously. Finally, plasmid p01ASA05 did not include virulence or antibiotic resistance factors and was only found in the ASA05 strain (not shown).
3.9. Antibiotic Resistance Factors
The three genomes described in this study contain factors associated with antibiotic resistance factors (ARs) effective against several drug classes, including peptides, fluoroquinolones, diaminopyrimidines, cephalosporines, and carbapenems (Figure 9). Notably, analyzing the ARs present in plasmids from Chilean strains shows that the ASA04p2 plasmid also includes the tet(A) and floR factors with high similarity percentages (Figure 8). These factors are associated with resistance against the tetracycline and phenicol drug classes, respectively, which are commonly used in the Chilean aquaculture industry.
4. Discussion
Atypical furunculosis represents an endemic bacterial disease with significant morbidity and mortality, which has traditionally been controlled through vaccination strategies. Notably, a substantial increase in the incidence of this disease was observed between 2022 and 2023, with a widespread geographical distribution in various freshwater production systems. This clinical and histopathological manifestation of cases is consistent with patterns described for furunculosis observed in salmonid fish. The histopathological findings are characterized by the presence of basophilic coccobacillary bacterial colonies in tissues, with inflammatory reactions being scant or absent. Other specific histological findings can be identified, such as cardiac inflammation, and hepatic and pancreatic necrosis, which are associated with co-infections with PRV and IPNV.
The genomic analysis of re-emergent A. salmonicida outbreaks in Atlantic salmon (S. salar) has provided significant insights into the pathogenesis and genomic diversity of this bacterial pathogen. The identified distinctive genomic patterns provide a comprehensive understanding of the complex dynamics associated with furunculosis, emphasizing its relevance in both salmonid and non-salmonid fish species across diverse aquatic environments. This study comprehensively examined clinical cases of atypical furunculosis in Atlantic salmon (S. salar) aquaculture in Chile, offering valuable insights into the epidemiological and clinical aspects of this disease. The 4.66% observed mortality rate and an outbreak duration of 8 weeks serve as indicators of the disease’s significant impact on the aquaculture industry. The selection of bacterial isolates from clinically diseased fish, characterized by the highest frequency of clinical signs, enabled a detailed genomic investigation, and shed light on the pathogenesis and antibiotic resistance profiles of this bacterium in Chile. The gross pathology and histopathological findings corroborate previous observations [14], which noted a range of clinical signs, including periorbital hemorrhages, abdominal hemorrhages, and the presence of furuncles in affected fish. The histopathological analysis underscores extensive tissue colonization by A. salmonicida, primarily characterized by bacterial colonies that replace tissues with limited inflammatory responses. The prevalence of necrotic lesions in the epidermis and dermis is a key pathological feature, which aligns with the observed clinical manifestations.
A pivotal contribution of this study is the genomic exploration of A. salmonicida, particularly in the distinctions between the mesophilic and psychrophilic strains, to confirm and extend recent findings in the field. The phylogenomic analysis revealed three distinct clusters, typical psychrophilic, atypical psychrophilic, and mesophilic, which demonstrates the genetic diversity within this species and validates previous observations [17,18]. The strains investigated in this study are classified within the atypical psychrophilic cluster. Five subspecies of Aeromonas salmonicida have been described; however, the assignment of different isolates to each of these subspecies has been debated, primarily through a comparative analysis of the complete genomes. Such analyses have revealed that this subspecies classification does not reflect divergence at the core genome level. In contrast to the classification into typical psychrophilic, atypical psychrophilic, and mesophilic groups, the last appears to have a genomic basis that is evident at the phylogenetic level and in the distribution of ISs and coding sequences in the R-M system.
The unique gene families found in these strains highlight the genomic complexity within atypical psychrophilic A. salmonicida and further emphasize the need for tailored genomic characterization. Thus, the analysis of virulence factors is of paramount importance. Specifically, since variations in exotoxin factors, particularly the actin cross-linking domain (ACD)-containing toxins, are present in the mesophilic strains but not in the psychrophilic strains, the potential role of these factors in pathogenicity is underscored. Distinctions in the presence of virulence genes such as clbF and aerA/act further accentuate the genetic diversity within typical and atypical psychrophilic strains. The selective presence of the RNA N-glycosidase factor stx2eA among atypical psychrophilic strains serves as an essential marker for distinguishing between subgroups within this classification. Plasmid characterization reveals differences in plasmid content, emphasizing the contribution of each plasmid to virulence and genome plasticity. The presence of antibiotic resistance factors within these genomes raises concerns regarding the development of antibiotic resistance and necessitates careful management strategies in Chilean aquaculture settings.
Furthermore, the diverse patterns in the insertion sequences and restriction-modification system coding sequences underscore the genomic structural differences among strains. REases function as regulatory agents, limiting the entry of foreign DNA with non-native methylation patterns, to modulate genetic influx. This impediment, in effect, may also play a role in preserving bacterial species. Corroborating this, many E. coli and Salmonella enterica serovar Typhimurium strains possess specific genomic loci, referred to as “immigration control regions”, which are abundant in R-M systems [49]. Therefore, R-M systems could achieve greater genetic isolation by regulating the uptake of environmental DNA. This would allow distinctive R-M patterns to be established among closely related strains, further genetically isolating them. As several genetic variants distinctly accumulate, the related bacterial strains gradually evolve and become more distant, until eventually becoming different species entirely. Additionally, studies on type III-like R-M enzymes have uncovered the function of these REases, which acted as a significant obstacle to the HGT in clinical variants of methicillin-resistant S. aureus [50]. The distinctive R-M coding sequence patterns observed among typical psychrophilic, atypical psychrophilic, and mesophilic A. salmonicida strains highlight their genomic diversity. Further, two distinctive patterns of R-M coding sequences are observed within the atypical psychrophilic and mesophilic strains. However, the impact that the R-M system may have on the genomic diversity of A. salmonicida requires further investigation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Distribution of Insertion sequences (ISs) classified by family. Figure S2: Virulence factors landscape of Aeromonas salmonicida complete genomes.
Author Contributions
Conceptualization, M.G., A.M, R.S, and H.B; formal analysis, M.M.O., D.C, K.K, H.B; writing—original draft preparation, M.G., M.M.O, R.S, J.P.P, D.C, K.K.; writing—review and editing, M.G., M.M.O, A.M, J.P.P, H.B, M.K, F.K, S.B; funding acquisition, M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Centro de Investigaciones Biológicas Aplicadas, grant number: clinical_genomic_Bacteria_100.
Data Availability Statement
The genome assemblies described in this study are openly available on NCBI, reference number PRJNA1045586.
Acknowledgments
FONDECYT DE INICIACION N ◦ 11230401 (to J.P.P).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martin-Carnahan, A.; Joseph, S.W. Aeromonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: New York, NY, USA, 2015; pp. 1–44, ISBN 978-1-118-96060-8.
- Austin, D.A.; McIntosh, D.; Austin, B. Taxonomy of Fish Associated Aeromonas Spp., with the Description of Aeromonas salmonicida Subsp. Smithia Subsp. Nov. Syst. Appl. Microbiol. 1989, 11, 277–290. [CrossRef]
- Bernoth, E.-M. Furunculosis: Multidisciplinary Fish Disease Research; Academic Press: San Diego, CA, USA, 1997; ISBN 978-0-12-093040-1.
- Emmerich, R. Über Eine Durch Bacterien Erzeugte Seuche Unter Den Forellen; 1894.
- KIMURA, T. A New Subspecies of Aeromonas Salmonicida as an Etiological Agent of Furunculosis on “Sakuramasu” (Oncorhynchus masou) and Pink salmon (O. gorbuscha) Rearing for Maturity. Part 1. On the Morphological and Physiological Properties. Fish Pathol. 1969, 3, 34–44.
- KIMURA, T. A New Subspecies of Aeromonas Salmonicida as an Etiological Agent of Furunculosis on “Sakuramasu”(Oncorhynchus masou) and Pink Salmon (O. gorbuscha) Rearing for Maturity Part 2 On the Serological Properties. Fish Pathol. 1969, 3, 45–52.
- Mccarthy, D.H. FISH FURUNCULOSIS CAUSED BY Aeromonas salmonicida VAR. achromogenes. J. Wildl. Dis. 1975, 11, 489–493. [CrossRef]
- Smith, I.W. The Classification of “Bacterium Salmonicida”. J. Gen. Microbiol. 1963, 33, 263–274. [CrossRef]
- Pavan, M.E.; Abbott, S.L.; Zorzópulos, J.; Janda, J.M. Aeromonas salmonicida Subsp. Pectinolytica Subsp. Nov., a New Pectinase-Positive Subspecies Isolated from a Heavily Polluted River. Int. J. Syst. Evol. Microbiol. 2000, 50, 1119–1124. [CrossRef]
- Austin, B.; Austin, D.A. Aeromonadaceae Representative (Aeromonas Salmonicida). In Bacterial Fish Pathogens; Springer: Dordrecht, The Netherlands, 2012; pp. 147–228, ISBN 978-94-007-4883-5.
- Beaz, R.; Jos, M. Molecular Detection and Characterization of Furunculosis and Other Aeromonas Fish Infections. In Health and Environment in Aquaculture; Carvalho, E., Ed.; InTech: London, UK, 2012; ISBN 978-953-51-0497-1.
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas Salmonicida: Updates on an Old Acquaintance. Dis. Aquat. Organ. 2016, 120, 49–68. [CrossRef]
- Bravo, S. Atypical Furunculosis in Atlantic salmon. 1999.
- Godoy, M.; Gherardelli, V.; Heisinger, A.; Fernández, J.; Olmos, P.; Ovalle, L.; Ilardi, P.; Avendaño-Herrera, R. First Description of Atypical Furunculosis in Freshwater Farmed Atlantic salmon, Salmo Salar L., in Chile. J. Fish Dis. 2010, 33, 441–449. [CrossRef]
- Emond-Rheault, J.-G.; Vincent, A.T.; Trudel, M.V.; Brochu, F.; Boyle, B.; Tanaka, K.H.; Attéré, S.A.; Jubinville, É.; Loch, T.P.; Winters, A.D.; et al. Variants of a Genomic Island in Aeromonas salmonicida Subsp. Salmonicida Link Isolates with Their Geographical Origins. Vet. Microbiol. 2015, 175, 68–76. [CrossRef]
- Gulla, S.; Bayliss, S.; Björnsdóttir, B.; Dalsgaard, I.; Haenen, O.; Jansson, E.; McCarthy, U.; Scholz, F.; Vercauteren, M.; Verner-Jeffreys, D.; et al. Biogeography of the Fish Pathogen Aeromonas Salmonicida Inferred by vapA Genotyping. FEMS Microbiol. Lett. 2019, 366, fnz074. [CrossRef]
- Vasquez, I.; Hossain, A.; Gnanagobal, H.; Valderrama, K.; Campbell, B.; Ness, M.; Charette, S.J.; Gamperl, A.K.; Cipriano, R.; Segovia, C.; et al. Comparative Genomics of Typical and Atypical Aeromonas Salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution. Microorganisms 2022, 10, 189. [CrossRef]
- Long, M.; Fan, H.; Gan, Z.; Jiang, Z.; Tang, S.; Xia, H.; Lu, Y. Comparative Genomic Analysis Provides Insights into Taxonomy and Temperature Adaption of Aeromonas salmonicida. J. Fish Dis. 2023, 46, 545–561. [CrossRef]
- Roh, H.; Kannimuthu, D. Comparative Resistome Analysis of Aeromonas Species in Aquaculture Reveals Antibiotic Resistance Patterns and Phylogeographic Distribution. Environ. Res. 2023, 239, 117273. [CrossRef]
- Bickle, T.A.; Krüger, D.H. Biology of DNA Restriction. Microbiol. Rev. 1993, 57, 434–450. [CrossRef]
- Pingoud, A.; Fuxreiter, M.; Pingoud, V.; Wende, W. Type II Restriction Endonucleases: Structure and Mechanism. Cell. Mol. Life Sci. 2005, 62, 685–707. [CrossRef]
- Roberts, R.J. A Nomenclature for Restriction Enzymes, DNA Methyltransferases, Homing Endonucleases and Their Genes. Nucleic Acids Res. 2003, 31, 1805–1812. [CrossRef]
- Murray, N.E. Immigration Control of DNA in Bacteria: Self versus Non-Self. Microbiology 2002, 148, 3–20. [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2010; ISBN 978-0-8138-0697-6.
- Prophet, E.; Armed Forces Institute of Pathology (U.S.). In Laboratory Methods in Histotechnology, 7th ed.; American Registry of Pathology: Arlington, TX, USA, 1992.
- Keeling, S.E.; Brosnahan, C.L.; Johnston, C.; Wallis, R.; Gudkovs, N.; McDonald, W.L. Development and Validation of a Real-time PCR Assay for the Detection of A Eromonas salmonicida. J. Fish Dis. 2013, 36, 495–503. [CrossRef]
- Zhao, J.; Vendramin, N.; Cuenca, A.; Polinski, M.; Hawley, L.M.; Garver, K.A. Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR. Pathogens 2021, 10, 1548. [CrossRef]
- Snow, M.; McKay, P.; McBeath, A.J.A.; Black, J.; Doig, F.; Kerr, R.; Cunningham, C.O.; Nylund, A.; Devold, M. Development, Application and Validation of a Taqman Real-Time RT-PCR Assay for the Detection of Infectious Salmon Anaemia Virus (ISAV) in Atlantic salmon (Salmo salar). Dev. Biol. 2006, 126, 133–145; discussion 325-326.
- Ingerslev, H.-C.; Rønneseth, A.; Pettersen, E.F.; Wergeland, H.I. Differential Expression of Immune Genes in Atlantic salmon (Salmo Salar L.) Challenged Intraperitoneally or by Cohabitation with IPNV. Scand. J. Immunol. 2009, 69, 90–98. [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and Accurate Long-Read Assembly via Adaptive k -Mer Weighting and Repeat Separation. Genome Res. 2017, 27, 722–736. [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [CrossRef]
- Bayliss, S.C.; Thorpe, H.A.; Coyle, N.M.; Sheppard, S.K.; Feil, E.J. PIRATE: A Fast and Scalable Pangenomics Toolbox for Clustering Diverged Orthologues in Bacteria. GigaScience 2019, 8, giz119. [CrossRef]
- Seemann, T. Snippy: Fast Bacterial Variant Calling from NGS Reads 2015. https://github.com/tseemann/snippy accessed on 01 October 2023.
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A Comparative Pathogenomic Platform with an Interactive Web Interface. Nucleic Acids Res. 2019, 47, D687–D692. [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, gkz935. [CrossRef]
- Siguier, P. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, D32–D36. [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE: A Database for DNA Restriction and Modification: Enzymes, Genes and Genomes. Nucleic Acids Res. 2023, 51, D629–D630. [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Res. 2023, 51, W484–W492. [CrossRef]
- Sibley, M.H. Cassette-like Variation of Restriction Enzyme Genes in Escherichia Coli C and Relatives. Nucleic Acids Res. 2004, 32, 522–534. [CrossRef]
- Corvaglia, A.R.; François, P.; Hernandez, D.; Perron, K.; Linder, P.; Schrenzel, J. A Type III-like Restriction Endonuclease Functions as a Major Barrier to Horizontal Gene Transfer in Clinical Staphylococcus aureus Strains. Proc. Natl. Acad. Sci. USA 2010, 107, 11954–11958. [CrossRef]
Figure 1.
Atlantic salmon (Salmo salar) in the freshwater phase are affected by a clinical presentation of atypical furunculosis caused by atypical Aeromonas salmonicida. (A,B) Multiple ulcers are observed, with some surrounded by a white-colored border. (C) Loss of structure and muscle hemorrhages are observed. (D) Petechial hemorrhages are observed in the swim bladder. (E) Atlantic salmon (S. salar) affected by a clinical picture of atypical furunculosis (atypical A. salmonicida). The presence of perirenal muscle hemorrhages is observed. (F) Atlantic salmon (S. salar) affected by a clinical picture of atypical furunculosis (atypical A. salmonicida). Periocular hemorrhage, exophthalmia, and multiple ulcers are noted, some of which have coalesced.
Figure 1.
Atlantic salmon (Salmo salar) in the freshwater phase are affected by a clinical presentation of atypical furunculosis caused by atypical Aeromonas salmonicida. (A,B) Multiple ulcers are observed, with some surrounded by a white-colored border. (C) Loss of structure and muscle hemorrhages are observed. (D) Petechial hemorrhages are observed in the swim bladder. (E) Atlantic salmon (S. salar) affected by a clinical picture of atypical furunculosis (atypical A. salmonicida). The presence of perirenal muscle hemorrhages is observed. (F) Atlantic salmon (S. salar) affected by a clinical picture of atypical furunculosis (atypical A. salmonicida). Periocular hemorrhage, exophthalmia, and multiple ulcers are noted, some of which have coalesced.

Figure 2.
Gross pathology findings were observed in Atlantic salmon (Salmo salar) presenting clinical cases of atypical furunculosis between 2022 and 2023, in a total sample size (n) of 294.
Figure 2.
Gross pathology findings were observed in Atlantic salmon (Salmo salar) presenting clinical cases of atypical furunculosis between 2022 and 2023, in a total sample size (n) of 294.
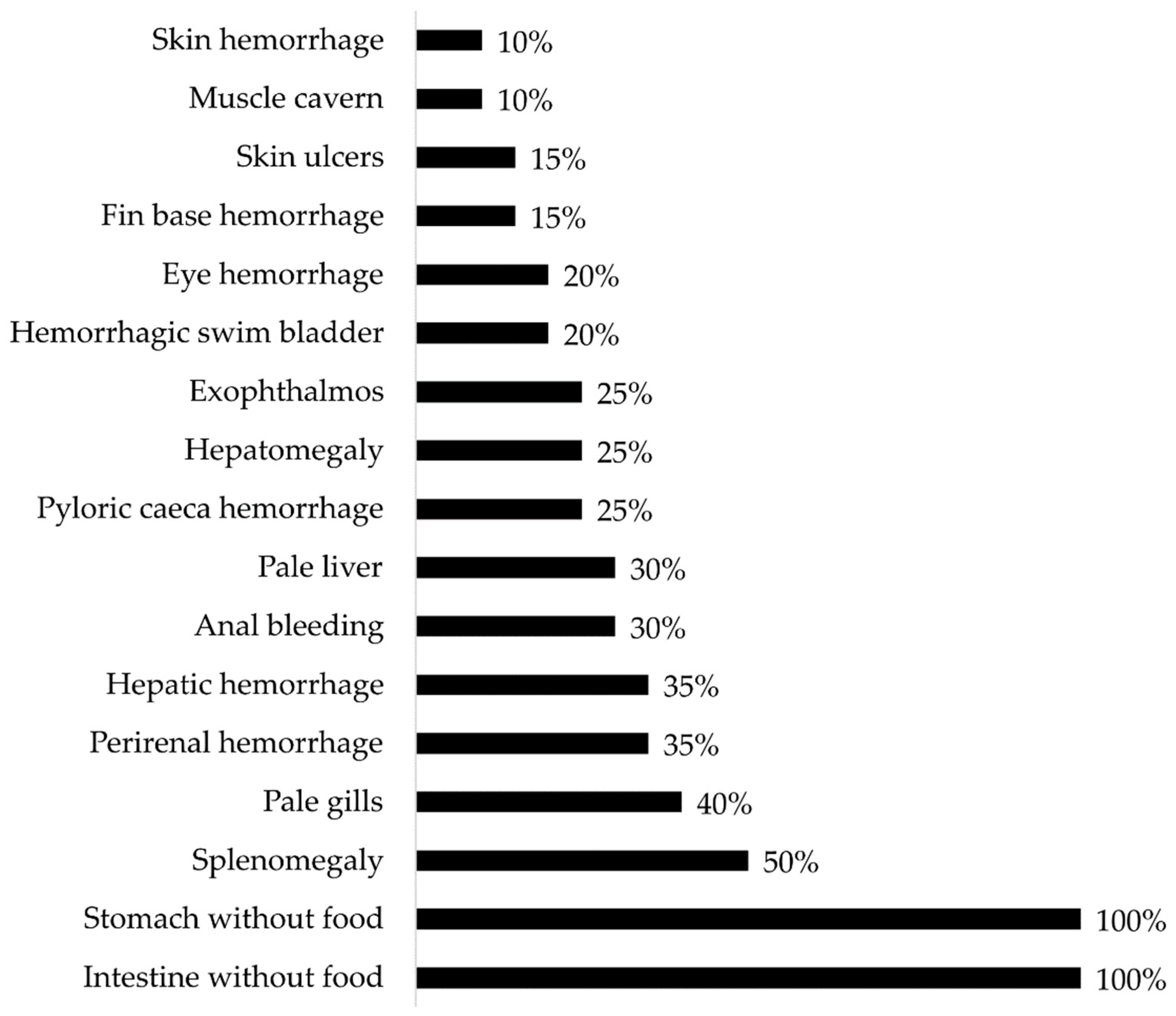
Figure 3.
(A) Atlantic salmon (Salmo salar), skin, and red muscle (H & E). Colonies of bacteria with coccobacillary morphology are observed in red muscle, consistent with atypical Aeromonas salmonicida infection (arrow). (B) Atlantic salmon (S. salar), red muscle (H & E). Observation of diffused infection of bacteria with coccobacillary morphology, consistent with atypical A. salmonicida infection. Additionally, hemorrhage and necrosis of the muscle tissue are observed. (C) Surrounding the muscular tissue, the presence of coccobacillary bacterial colonies consistent with infection by atypical A. salmonicida is observed (arrow). Additionally, muscular necrosis and hemorrhage are observed. (D) Atlantic salmon (S. salar), spleen (H & E). In the splenic parenchyma, multiple colonies of bacteria with coccobacillary morphology are observed, consistent with atypical A. salmonicida infection (arrows). (E) Colonies of bacteria with coccobacillary morphology are observed in the auricle, consistent with atypical A. salmonicida infection (arrow). (F) Atlantic salmon (S. salar), gill (H & E). Bacterial infiltration of the center gill filament with coccobacillary morphology is observed, consistent with atypical A. salmonicida infection (arrow).
Figure 3.
(A) Atlantic salmon (Salmo salar), skin, and red muscle (H & E). Colonies of bacteria with coccobacillary morphology are observed in red muscle, consistent with atypical Aeromonas salmonicida infection (arrow). (B) Atlantic salmon (S. salar), red muscle (H & E). Observation of diffused infection of bacteria with coccobacillary morphology, consistent with atypical A. salmonicida infection. Additionally, hemorrhage and necrosis of the muscle tissue are observed. (C) Surrounding the muscular tissue, the presence of coccobacillary bacterial colonies consistent with infection by atypical A. salmonicida is observed (arrow). Additionally, muscular necrosis and hemorrhage are observed. (D) Atlantic salmon (S. salar), spleen (H & E). In the splenic parenchyma, multiple colonies of bacteria with coccobacillary morphology are observed, consistent with atypical A. salmonicida infection (arrows). (E) Colonies of bacteria with coccobacillary morphology are observed in the auricle, consistent with atypical A. salmonicida infection (arrow). (F) Atlantic salmon (S. salar), gill (H & E). Bacterial infiltration of the center gill filament with coccobacillary morphology is observed, consistent with atypical A. salmonicida infection (arrow).
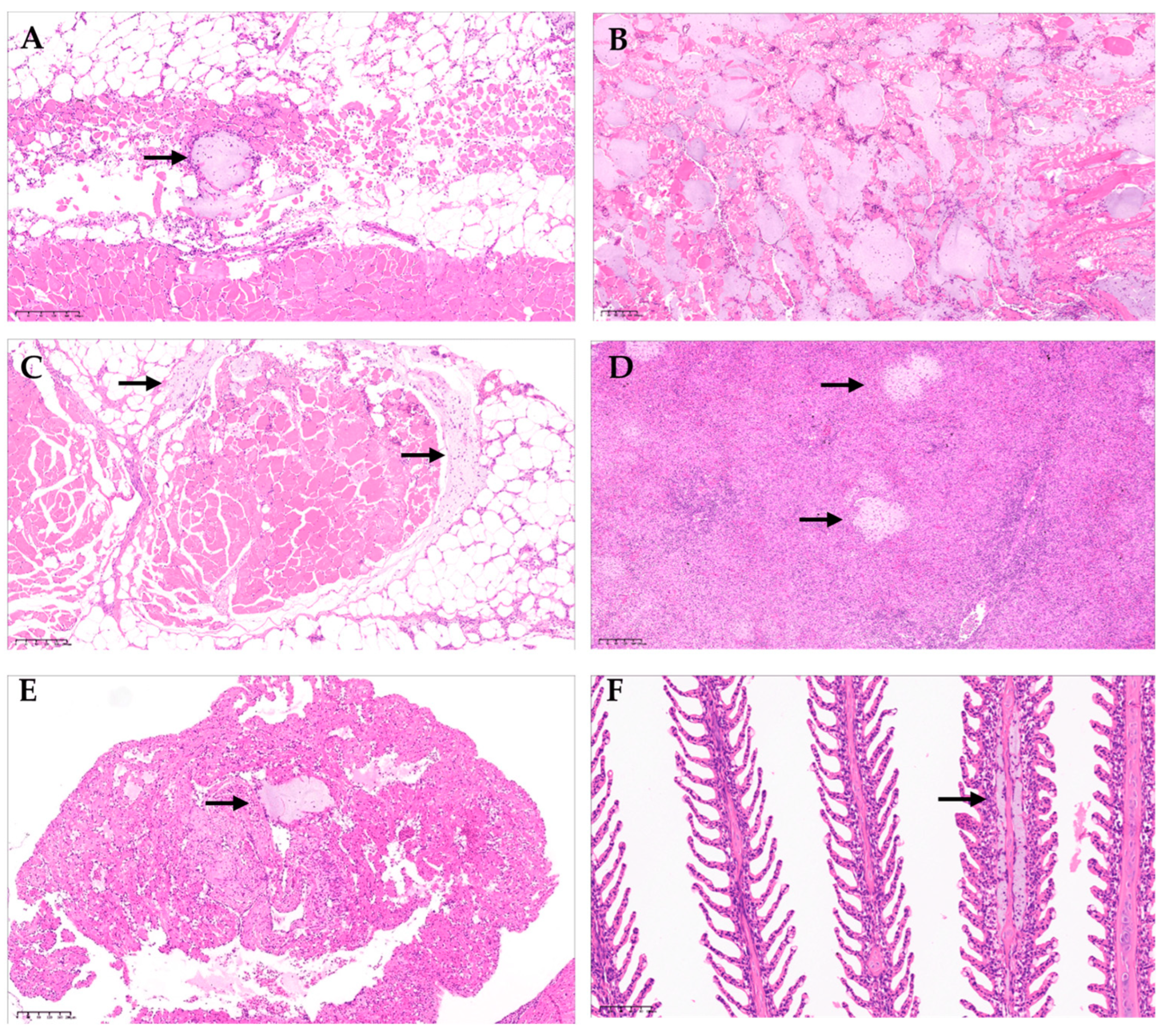
Figure 4.
Pan-genome analysis of Aeromonas salmonicida strains. (A) The proportion of genomes with gene families present. Gene families are classified as stable (in blue) when they contain only a single allele with a 98% similarity in their amino acid sequence, and as divergent (in red) when they have more than one allele. (B) Core and accessory gene/allele estimates. The estimates represent “allelic” variation. Core gene families are characterized as being present in > 95% of genomes. (C) Shared gene presence per isolate, ordered alongside the phylogenetic tree. Gene family presence is indicated by a blue block per column. (D) Core and unique gene families per genome. *: Genomes described in this study.
Figure 4.
Pan-genome analysis of Aeromonas salmonicida strains. (A) The proportion of genomes with gene families present. Gene families are classified as stable (in blue) when they contain only a single allele with a 98% similarity in their amino acid sequence, and as divergent (in red) when they have more than one allele. (B) Core and accessory gene/allele estimates. The estimates represent “allelic” variation. Core gene families are characterized as being present in > 95% of genomes. (C) Shared gene presence per isolate, ordered alongside the phylogenetic tree. Gene family presence is indicated by a blue block per column. (D) Core and unique gene families per genome. *: Genomes described in this study.
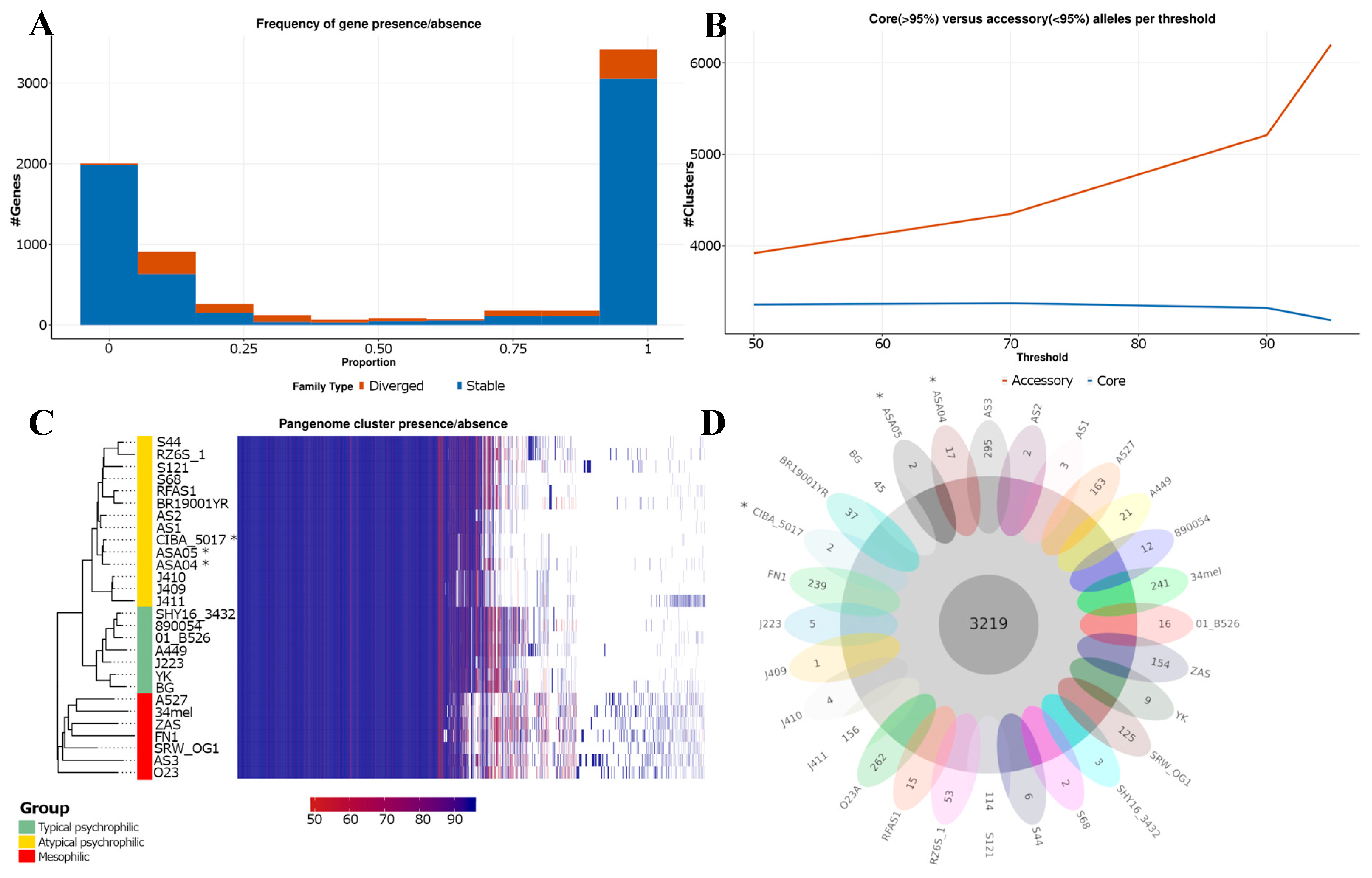
Figure 5.
Phylogenomic analysis of Aeromonas salmonicida strains. Maximum likelihood phylogenic tree constructed from a core genome alignment encompassing 89,915 SNPs. *: Genomes described in this study. Bootstrap frequencies above 70% are represented as red circles. Other representative Aeromonas species are included as outgroup isolates.
Figure 5.
Phylogenomic analysis of Aeromonas salmonicida strains. Maximum likelihood phylogenic tree constructed from a core genome alignment encompassing 89,915 SNPs. *: Genomes described in this study. Bootstrap frequencies above 70% are represented as red circles. Other representative Aeromonas species are included as outgroup isolates.

Figure 6.
Distribution of restriction-modification (R-M) coding sequences.
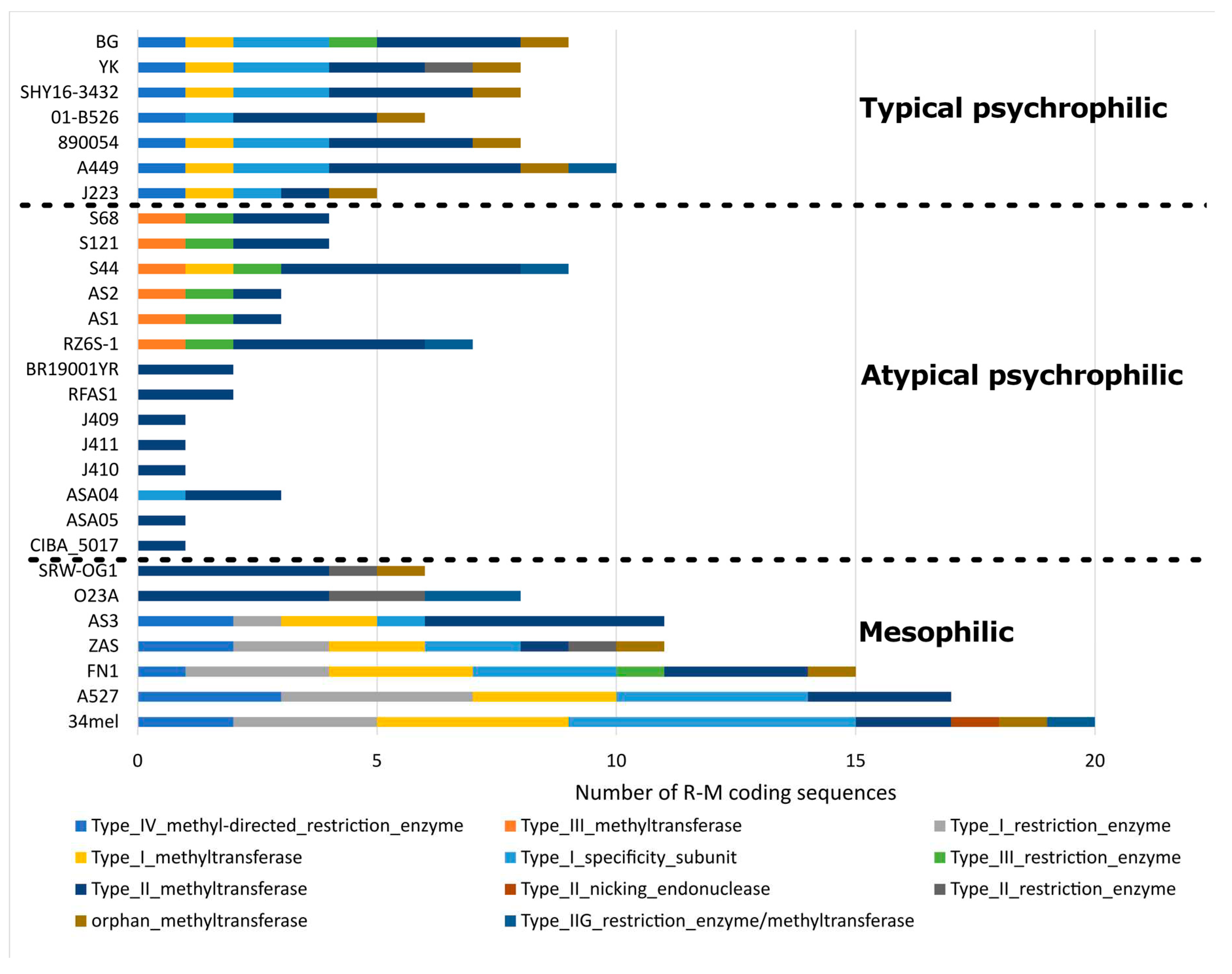
Figure 7.
Profile of exotoxin virulence factors in complete Aeromonas salmonicida genomes. The presence and absence of exotoxin factors were predicted based on local alignments with the VFDB entries. The similarity percentage with the identified factors is shown. *: Genomes reported in this study, with their results highlighted in the green box.
Figure 7.
Profile of exotoxin virulence factors in complete Aeromonas salmonicida genomes. The presence and absence of exotoxin factors were predicted based on local alignments with the VFDB entries. The similarity percentage with the identified factors is shown. *: Genomes reported in this study, with their results highlighted in the green box.
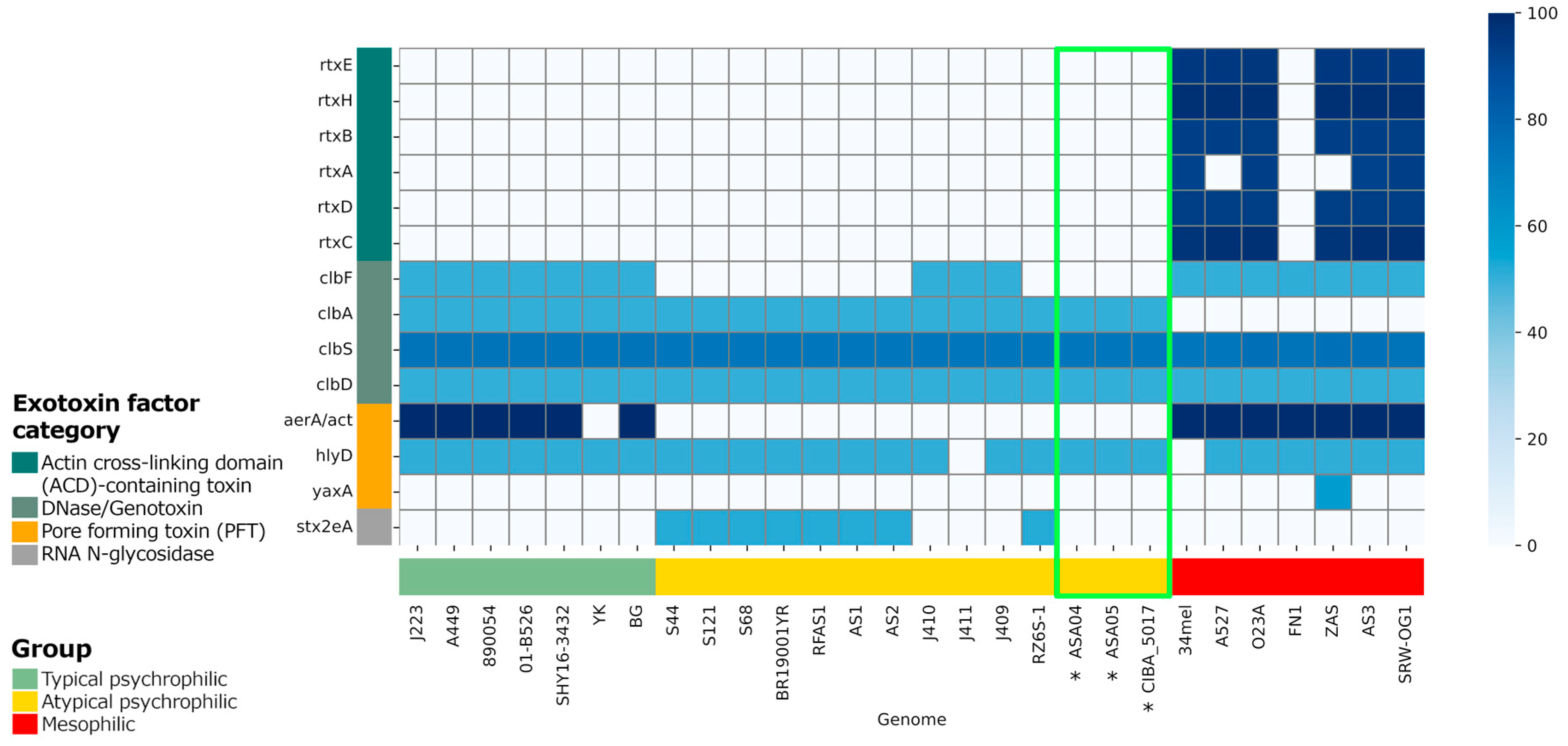
Figure 8.
Virulence and antibiotic resistance factors identified in Aeromonas salmonicida plasmids.
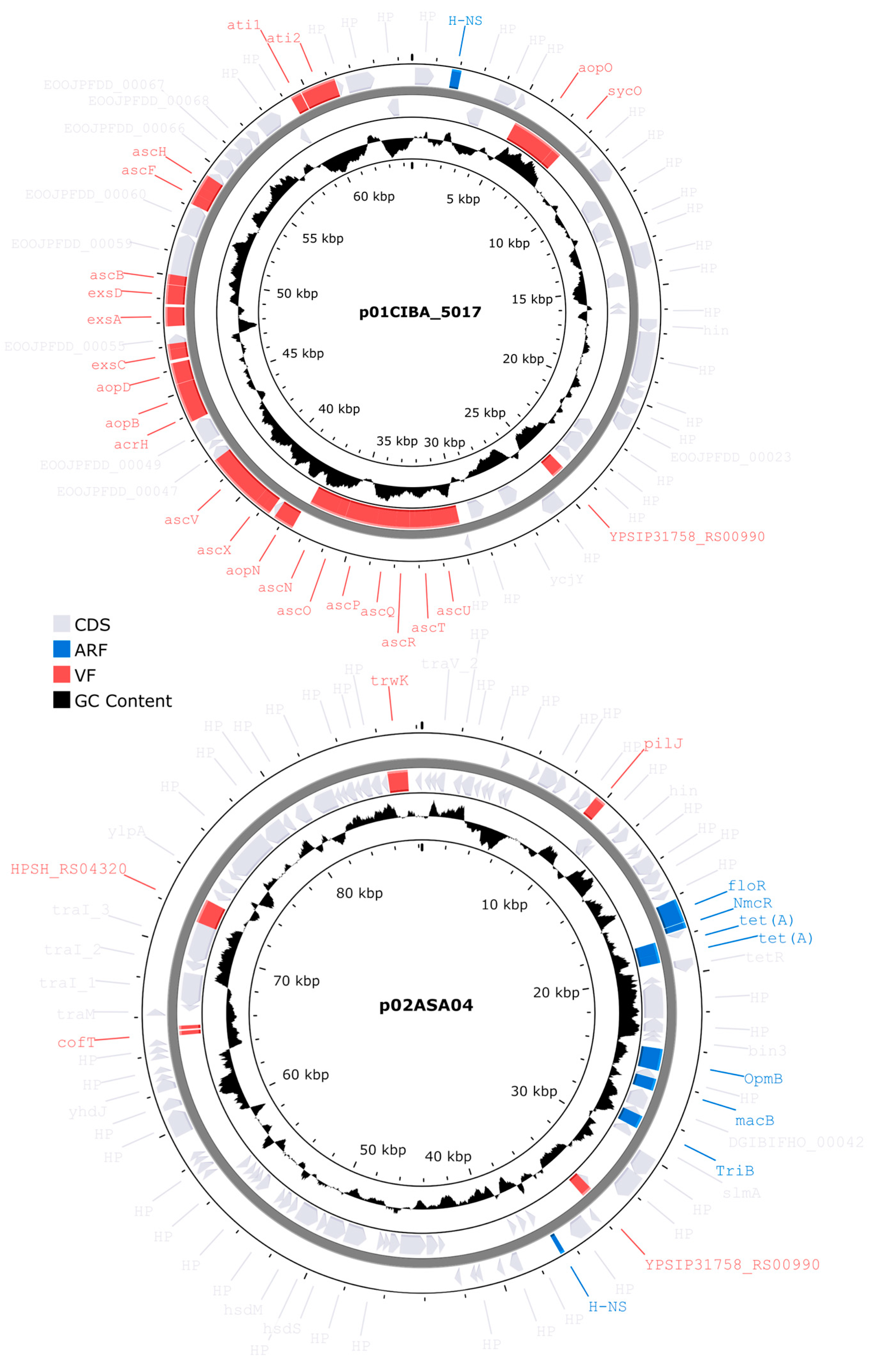
Figure 9.
Antibiotic resistance repertoire in Aeromonas salmonicida complete genomes. The presence and absence of the factor are represented in blue and white, respectively. *: Genomes reported in this study, with their results highlighted in the green box.
Figure 9.
Antibiotic resistance repertoire in Aeromonas salmonicida complete genomes. The presence and absence of the factor are represented in blue and white, respectively. *: Genomes reported in this study, with their results highlighted in the green box.
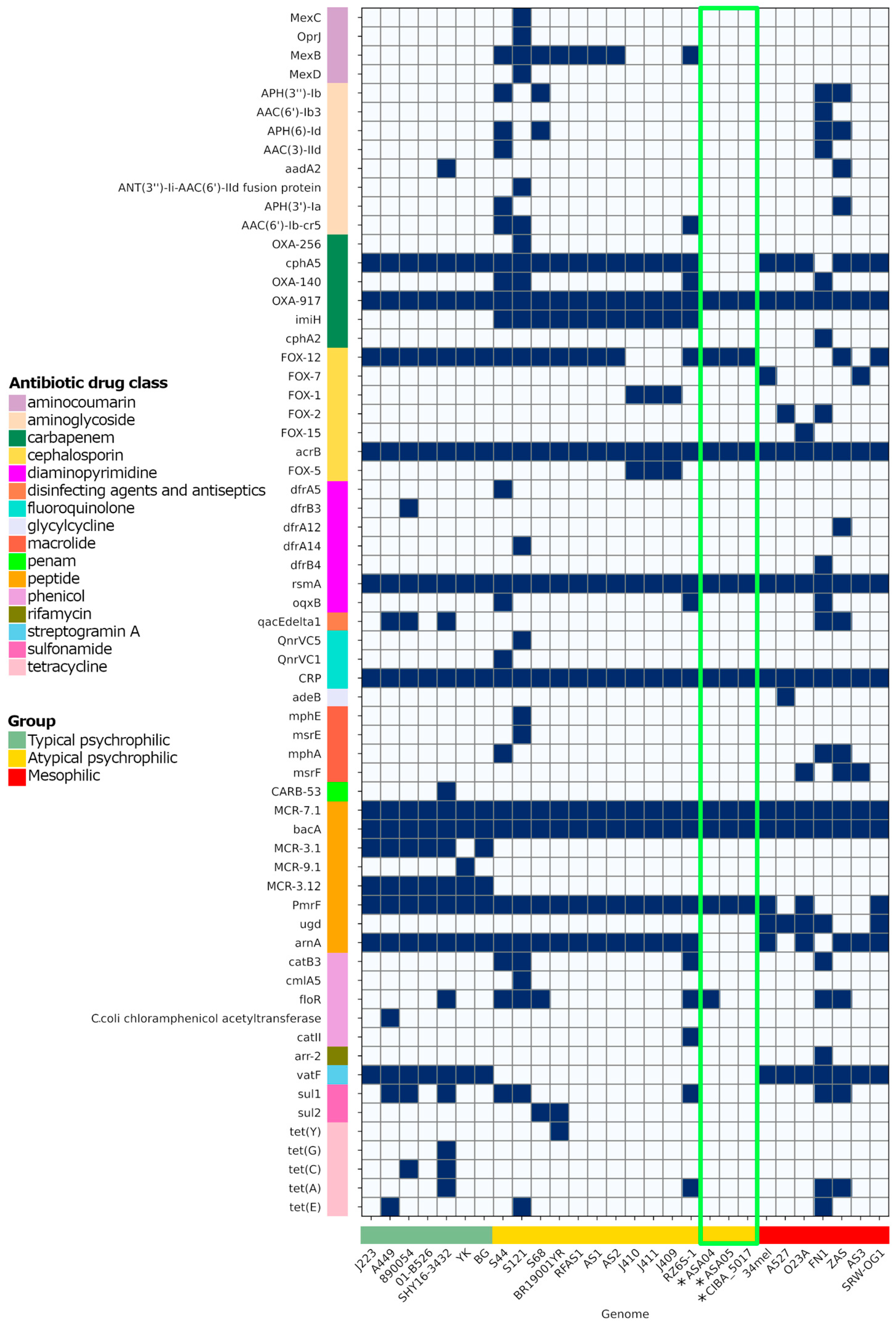

Table 1.
Epidemiological background and laboratory results for the cases analyzed in this study.
| Case ID | 4978 | 4988 | 5131 | 7003 | 7031 | 7081 | 7088 | 7124 | 7167 | 7175 | 7174 | 7215 | 7216 | 7214 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Date | 10-Mar | 14-Mar | 11-May | 23-May | 03-Jun | 23-Jun | 30-Jun | 21-Jul | 08-Aug | 12-Aug | 12-Aug | 18-Aug | 24-Aug | 25-Aug |
| Region | X | X | X | X | X | XII | XII | XII | X | X | X | XII | XII | X |
| Salinity | FW | FW | BW | FW | FW | BW | BW | BW | BW | BW | FW | BW | BW | BW |
| Development stage | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt |
| Fish sampled | 20 | 7 | 10 | 30 | 6 | 7 | 7 | 10 | 10 | 11 | 24 | 1 | 1 | 1 |
| Average ASA Ct | 22.8 | 26.1 | 29.2 | 28.4 | 23.8 | 27.5 | 26.2 | 24.4 | 27.1 | 29.8 | 21.4 | 26.1 | 32.8 | 22.5 |
| Coinfection | PRV | IPNV | IPNV | R. salmoninarum | PRV | IPNV-F.psy | IPNV-PRV | IPNV-PRV | - | - | IPNV-PRV | - | - | - |
| Bacterial isolation | Positive | Positive | Positive | - | - | - | - | - | Positive | - | - | - | - | - |
| GenBank accession | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
Table 1.
(continued) Epidemiological background and laboratory results for the cases analyzed in this study.
Table 1.
(continued) Epidemiological background and laboratory results for the cases analyzed in this study.
| Case ID | 7198 | 7213 | 7217 | 7385 | 7388 | 7387 | 7412 | 7416 | 7435 | CIBA-5017 | ASA04 | ASA05 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Date | 26-Aug | 26-Aug | 06-Sept | 20-Oct | 21-Oct | 21-Oct | 27-Oct | 28-Oct | 04-Nov | 25-Mar | 22-Nov | 23-Nov |
| Region | X | X | X | X | X | X | X | X | X | X | X | X |
| Salinity | FW | FW | FW | FW | BW | FW | FW | FW | BW | FW | FW | FW |
| Development stage | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Pre-smolt | Smolt | Smolt | Smolt |
| Fish sampled | 10 | 1 | 7 | 24 | 24 | 26 | 15 | 10 | 15 | 20 | 25 | 11 |
| Average ASA Ct | 29.6 | 26.2 | 23.2 | 30.6 | 26.6 | 24.3 | 30.8 | 29.1 | 27.0 | 28.8 | 29.7 | 26.1 |
| Coinfection | IPNV | - | IPNV | - | - | - | - | PRV | PRV | - | IPNV-PRV | IPNV-PRV |
| Bacterial isolation | Positive | - | - | Negative | - | Positive | Negative | Negative | - | Positive | Positive | Positive |
| GenBank accession | - | - | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



