
Submitted:
30 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
Using qRT-PCR analysis, the LcIL-8 expression level indicated broad expression in most detected tissues, with the most predominant expression in whole blood at 6 to 24 h after infection with S. iniae at concentrations of 105 colony-forming unit (CFU)/fish (p < 0.05). After immersion with F. covae, the LcIL-8 transcript was upregulated in the gills, liver and intestine, with the highest expression level found in the gills. However, LcIL-8 was downregulated in all tested tissues at 48 and 96 h after infection with the two pathogenic strains, indicating that Lc-IL8 has a short half-life during the early immune responses to pathogens. MIC analysis of rLcIL-8 protein against S. iniae was 10.42 ± 3.61 µg/mL. Furthermore, the functional analyses clearly demonstrated that 10 and 100 µg of rLcIL-8 protein efficiently enhanced the phagocytic activity of Asian seabass phagocytes in vitro (p < 0.05). Additionally, in vivo injection of S. iniae following the rLcIL-8 protein indicated that 50 and 100 µg of rLc-IL-8 were highly effective in protecting fish from the tested pathogen (p < 0.001). These results indicate that rLcIL-8 plays a crucial role in the defense mechanisms against bacterial infections in the target fish.
Keywords:
proinflammatory cytokine
; interleukin-8
; asian seabass
; Streptococcus iniae
; Flavobacterium covae
; antimicrobial activity
1. Introduction
Asian seabass (Lates calcarifer Bloch, 1790) is considered an economically significant fish in Thailand and several other countries in the Asian region. The major export markets for Asian seabass include China, Germany, the Netherlands, Indonesia, and Vietnam [1]. Thailand is the 16th largest exporter of Asian seabass in the world. Currently, Asian seabass aquaculture has expanded throughout Thailand, becoming an intensive culture system. However, this system causes poor water quality and rapid changes in the aquatic environment during certain stages of cultivation. These effects directly increase stress in the seabass and induce susceptibility to severe diseases caused by various parasites, viruses, and bacteria. Recently, in Thailand, major pathogenic bacteria, including Flavobacterium covae and Vibrio spp. and Streptococcus spp., particularly S. iniae and F. covae, which are the causative agents of streptococcosis and columnaris, respectively, have been identified in Asian seabass farming. These diseases can lead to fish mortality within 24-72 hours, resulting in substantial losses in the Asian seabass farming industry in Thailand [2,3]. The utilization of antibiotic and chemical treatments for disease management in these fish has been on the decline, a trend primarily attributed to concerns about potential health risks for consumers, compounded by the association of many fish pathogens with the development of antibiotic resistance [4].
To address these problems and sustain Asian seabass aquaculture, an evaluation of the molecular structure and functional analysis of immune-related genes is needed for the effective development of prophylactic and therapeutic strategies. Based on current knowledge, it is known that the immunity of fish, especially teleost fish, can be divided into innate and specific or acquired immunity. The innate immune system is the first line of defense against pathogens that attempt to enter the body of all animals [5]. It is a nonspecific pathway, has limited memory, and responds rapidly to a variety of microbial agents. This system can be divided into three crucial components: physical, cellular and humoral defenses. Both humoral and cellular defenses are crucial components that are typically closely associated with the specific immune system. In particular, the humoral response depends upon the activity of several vital molecules, such as lytic enzymes, complements, transferrin, and cytokines [6]. Cytokines are a group of molecules that certain cells produce to perform important functions in cell signaling during wound healing, inflammation and pathogen infections, especially those caused by bacteria or viruses [7]. In the inflammatory response process, cytokines that promote the initial stages of inflammation are also known as “proinflammatory cytokines”. These molecules are not secreted under ordinary conditions. However, they are secreted when the immune system is triggered by pathogens [8]. Among these molecules, interleukin 8 (IL-8), referred to as “CXCL8 or NAP-1 (neutrophil-activating peptide)”, was the first chemokine to be identified and was initially purified from LPS-stimulated human blood monocytes [9]. Currently, it is widely acknowledged that IL-8 can be produced by various cell types, including nonimmune cells such as fibroblasts and endothelial cells, in response to a wide range of pathogenic stimuli [10]. It is a pivotal chemokine involved in the pro-inflammatory process. This chemokine belongs to the subfamily of CXC chemokines and serves as a chemotactic factor, attracting white blood cells during the early stages of inflammation. Moreover, it possesses the capability to stimulate cells by initiating respiratory bursts, degranulation of storage proteins and production of lipid mediators [11,12,13]. The ability of IL-8 to attract specific cells is attributed to the absence of an ELR motif (Glu-Leu-Arg) adjacent to a CXC motif at the N-terminus [14]. However, in certain fish species, such as Atlantic cod and haddock, the presence of the ELR motif has been discovered to have the capacity to attract PMN cells, especially during inflammation. In fish, the first IL-8 was identified in lamprey (Lampetra fluviatilis) [15]. Subsequently, IL-8 has been characterized in several other fish species, such as flounder [16], trout [17,18], common carp [19], catfish [20], zebrafish [21] and Atlantic cod [22].
Our understanding of the immune function and regulatory processes governing IL-8 in Asian seabass is currently limited. Therefore, this study aims to investigate the structure of the complementary DNA (cDNA) of the IL-8 gene and its transcriptional level expression under pathogenic bacterial infection. Additionally, it aims to elucidate the function of this molecule under various conditions. The insights obtained from the current study are crucial for elucidating the immune mechanisms and functional roles of LcIL-8 to inform the development of prophylactic and therapeutic methods that improve disease resistance against pathogenic bacteria in the Asian seabass aquaculture industry.
2. Materials and Methods
2.1. Experimental Animals
Healthy Asian seabasses (10 g) were obtained from a commercial nursery farm in Chachoengsao Province, Thailand. The fish were acclimatized under laboratory conditions at the Laboratory of Aquatic Animal Health Management (LAAHM), Department of Aquaculture, Faculty of Fisheries, Kasetsart University (Bangkok, Thailand). Subsequently, the experimental fish were maintained in a quarantined 3000-L fiberglass tank containing clean dechlorinated water and adjusted to a salinity of 10 ppt with an aeration system. During acclimatization, the fish were fed twice a day with Seabass feed C-5003 (Uni-President, Vietnam) at 5% body weight for 1 week before the beginning of the experiment.
2.2. Cloning and Characterization of the cDNA Encoding the Mature LcIL-8 Protein of Asian Seabass
Total RNA was isolated from the head kidney of euthanized fish using TRIzol reagent (Invitrogen, Waltham, MA, USA) following the manufacturer’s instructions. First-strand cDNA was synthesized from the total RNA using ReverTra AceTM qPCR RT Master Mix with gDNA Remover (TOYOBO Bio-Technology, CO., LTD., Osaka, Japan) to eliminate contaminating genomic DNA. The open reading frame (ORF) of Asian seabass IL-8 was obtained from the GenBank database (XM_018695863.2) (http://www.ncbi.nlm.nih.gov) and cloned by PCR amplification. Specific primers were designed to amplify the sequence encoding Lc-IL8 starting from the beginning of the leader sequence to the stop codon (LcIL-8 F and LcIL-8 R, Table 1). The cycling conditions were as follows: 95 °C for 5 min; 40 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min; and an elongation step of 72 °C for 10 min. The PCR products were separated by electrophoresis on a 1.5% agarose gel and further purified using a QIAquick® gel extraction kit (Qiagen, Venlo, Netherlands). The obtained PCR DNA fragment was ligated into the pGEM T-easy vector following the provided protocol (Promega, Madison, WI, USA) and then transformed into Escherichia coli (strain JM109) competent cells using the heat-cold shock method. To identify positive clones, a blue‒white colony screening was performed on Luria–Bertani (LB) agar plates supplemented with ampicillin (100 mg/mL), isopropyl-β-d-1-thiogalactopyranoside (IPTG) (1 mM) and X-gal (50 mg/mL). Plasmids were extracted from selected positive clones using a NucleoSpin® Plasmid QuickPure Kit (Macherey-Nagel, Germany). Finally, nucleotide sequencing of the target plasmid at both the 5′ and 3′ directions was conducted using the primers M13F and M13R by Macrogen, Inc. (South Korea). The analysis of the cDNA and the deduced amino acid sequence of Lc-IL8 was conducted using the BLAST program available through the National Center for Biotechnology Information (NCBI; http://www.ncbi.nlm.nih.gov) and the ExPASy Molecular Biology server (http://us.expasy.org).
2.3. Homology and Phylogenetic Analysis of LcIL-8 and Various Interleukin-8 Genes of Other Vertebrates
The amino acid sequence identity and similarity of LcIL-8 cDNA and other reported IL-8 gene sequences were assessed using MatGAT version 2.01 (http://www.angelfire.com/nj2/arabidopsis/MatGAT.html). To investigate evolutionary relationships, a phylogenetic tree of IL-8 genes from various vertebrate species was constructed. This tree was generated by aligning the amino acid sequences of LcIL-8 cDNA with those of known IL-8 genes from diverse vertebrate species using ClustalW (http://ebi.ac.uk/Tools/clustalw/index.html). The phylogenetic tree was constructed using the neighbor-joining method in MEGA 11.0 software (www.megasoftware.net) with 1000 bootstrap analyses.
2.4. Quantitative Reverse-Transcription Real-Time PCR (qRT‒PCR) Analysis of LcIL-8 Gene Expression in Various Tissues of Healthy Fish
2.4.1. Total RNA Isolation and First-Strand cDNA Synthesis
Four fish randomly collected from the previous section were used for tissue extraction, encompassing 12 organs: brain (BR), gills (GL), heart (HR), liver (LV), spleen (SP), head kidney (HK), trunk kidney (TK), intestine (INT), stomach (SM), muscle (MC), skin (SK), and whole blood (WB). Whole blood was obtained by withdrawal from the caudal vein using a 1-mL heparinized syringe equipped with a 23-G needle. Total RNA was isolated from these 12 tissues using TRIzol reagent (Invitrogen, Waltham, MA, USA) according to the manufacturer’s protocol. The RNA pellets from all tissues were air-dried, and the total RNA was dissolved in sterile nuclease-free water. Subsequently, 1 µg of total RNA from each tissue was separately used to synthesize first-strand cDNA using ReverTra AceTM qPCR RT Master Mix with gDNA Remover (TOYOBO Bio-Technology, CO., LTD., Osaka, Japan). The first-strand cDNA was carefully synthesized based on the recommendation of the company’s protocol and stored at -20 °C until use.
2.4.2. qRT‒PCR Analysis
To assess the mRNA levels of the LcIL-8 transcripts in the 12 different tissues of normal fish, qRT‒PCR was conducted with the specific primers LcIL-8 qF and LcIL-8 qR (Table 1). One microgram of first-strand cDNA from every organ was employed using Brilliant III Ultra-Fast SYBR® Green qRT‒PCR Master Mix (Agilent Technologies, Inc., CA, USA), which was performed using an Mx3005P real-time PCR system (Stratagene, USA) equipped with analytical software version 4.0, following the manufacturer’s recommended protocol. Each reaction was performed in a final volume of 25 µL containing 1.0 µL of first-strand cDNA, 12.5 µL of Brilliant III Ultra-Fast SYBR Green qRT‒PCR master mix, 9.5 µL of sterile distilled water and 1.0 µL of each specific primer pair, including LcIL-8 qF and LcIL-8 qR (Table 1). The PCR cycling conditions were set as follows: an initial step of denaturation at 95 °C for 10 min, followed by 40 cycles of denaturation at the same temperature for a precisely timed 30 s, annealing at 56 °C for 30 s, and extension at 72 °C for 1 min. To standardize the results and eliminate variations in mRNA and cDNA quantity and quality, the expression level of the β-actin gene was used as the internal control, with primers Lc-β-actin qF and Lc-β-actin qF (Table 1). Primer specificity was confirmed through DNA melting curve analysis. Triplicate reactions were performed for both the LcIL-8 and β-actin genes in each sample. All primers for gene expression analysis were designed based on the sequences indicated in the Asian seabass genome project (ASM164080v1). A standard plasmid containing the LcIL-8 and β-actin genes was serially diluted in 10-fold increments to generate standard curves to assess PCR efficiency. The relative copy number of the target mRNA was calculated using 2-ΔΔCT analysis, and the expression ratio in the muscle was used as a calibrator according to the protocol by Livak and Schmittgen (2001) [23].
2.5. Response Analysis of LcIL-8 under Stimulation with S. iniae and F. covae Using qRT‒PCR
2.5.1. Bacterial Strains and Preparation
Pathogenic bacterial strains, S. iniae and F. covae were obtained from the LAAHM. A single colony of S. iniae was cultured in 100 mL of trypticase soy broth (TSB) medium and incubated in a shaking incubator at 30 °C for 18–24 hours (hs). Subsequently, the bacterial cells were harvested by centrifugation at 2,300 × g for 15 min and then resuspended in sterile 0.85% NaCl solution until the absorbance at an optical density (OD) of 600 reached 1.00, resulting in a concentration of 1×109 colony-forming unit (CFU)/mL. Then, the same solution was diluted to obtain final concentrations of 1×102, 1×104 and 1×106 CFU/mL. Similarly, a pure colony of F. covae was cultured in 200 mL of Shieh’s medium broth in an incubator shaker at 25-30 °C for 24 h. Then, the obtained bacterial culture was inoculated into 1 L of Shieh’s broth and incubated under the same conditions. Finally, the pellet was resuspended in a sterile 0.85% NaCl solution. The concentrations of F. covae were adjusted to 1×102, 1×104 and 1×106 CFU/mL. This bacterial suspension was used for other experiments below.
2.5.2. Experimental Animals and Design
Two hundred Asian seabass from the previous section were selected and divided into 6 fiberglass tanks, each with a 250-L capacity (30 fish/tank). Tanks 1-3 were filled with salt water at a salinity of 5 ppt, while tanks 4–6 were filled with fresh water and acclimatized as described above for 7 days. After the acclimatization period, all healthy Asian seabass in tanks 1–3 were intraperitoneally injected with 100 µL of S. iniae solution prepared in 0.85% NaCl solution at concentrations of 1×102, 1×104 and 1×106 CFU/mL, respectively. Meanwhile, fish in tanks 4-6, all individuals were removed and placed into 10-L tanks containing clean freshwater, which was prepared with the F. covae bacterial solution and had the same concentration as the S. iniae solution. Immersion was carried out for a duration of 30 min, and then the fish were returned to each tank as before. Sampling was conducted at 0 (Control), 6, 12, 24, 48 and 96 hours after injection; the liver, spleen, head kidney, gills and intestine were collected from 4 fish in each bacterial injected group.
2.5.3. Total RNA Extraction and First-Strand cDNA Synthesis
After infection, tissue samples were collected for total RNA extraction with the methods described above. Total RNA templates were prepared at a concentration of 1,000 ng/mL from each group at different time points. First-strand cDNA was subsequently synthesized using the same method described previously. The resulting cDNA was stored at -20 °C until use.
2.5.4. Quantitative Real-Time PCR Assay
First-strand cDNAs from the liver, spleen, head kidney, gills and intestine of the infected fish at each time point were analyzed using qRT‒PCR as described above with slight modifications, where the relative expression levels of each gene at h 0 were used as calibrators.
2.6. Overexpression, Production and Purification of Recombinant LcIL-8 Protein (rLcIL-8)
2.6.1. Construction of Recombinant LcIL-8 DNA
The plasmid DNA containing the complete nucleotide sequence obtained in the experiment detailed in section 2.2 was subjected to double-restriction enzyme digestion (Nde I and Xho I) following the recommendations of the company’s protocol (Thermo Scientific, Waltham, MA, USA). The resulting products were separated by electrophoresis on a 1.5% agarose gel. Subsequently, the desired digested DNA fragment was purified using a QIAquick® gel extraction kit (Qiagen, Venlo, Netherlands). This purified DNA fragment was ligated into an Xho I/Nde I-cut pET28b (+) expression vector with an N-terminal 6× His-tag and transformed into JM109 competent cells. The transformed bacteria were selected and cultured at 37 °C in LB broth containing 100 µg/mL kanamycin. The positive clones were isolated, preserved, and subjected to plasmid DNA extraction, following the previously described procedure. Subsequently, the rLcIL-8 plasmid was retransformed into E. coli strain BL21 (DE3) using heat-cold shock techniques. The positive cells containing the rLcIL-8 plasmid were sequenced using a similar method as previously mentioned to confirm the correctness of the LcIL-8 sequence.
2.6.2. Overexpression of Recombinant LcIL-8 Protein
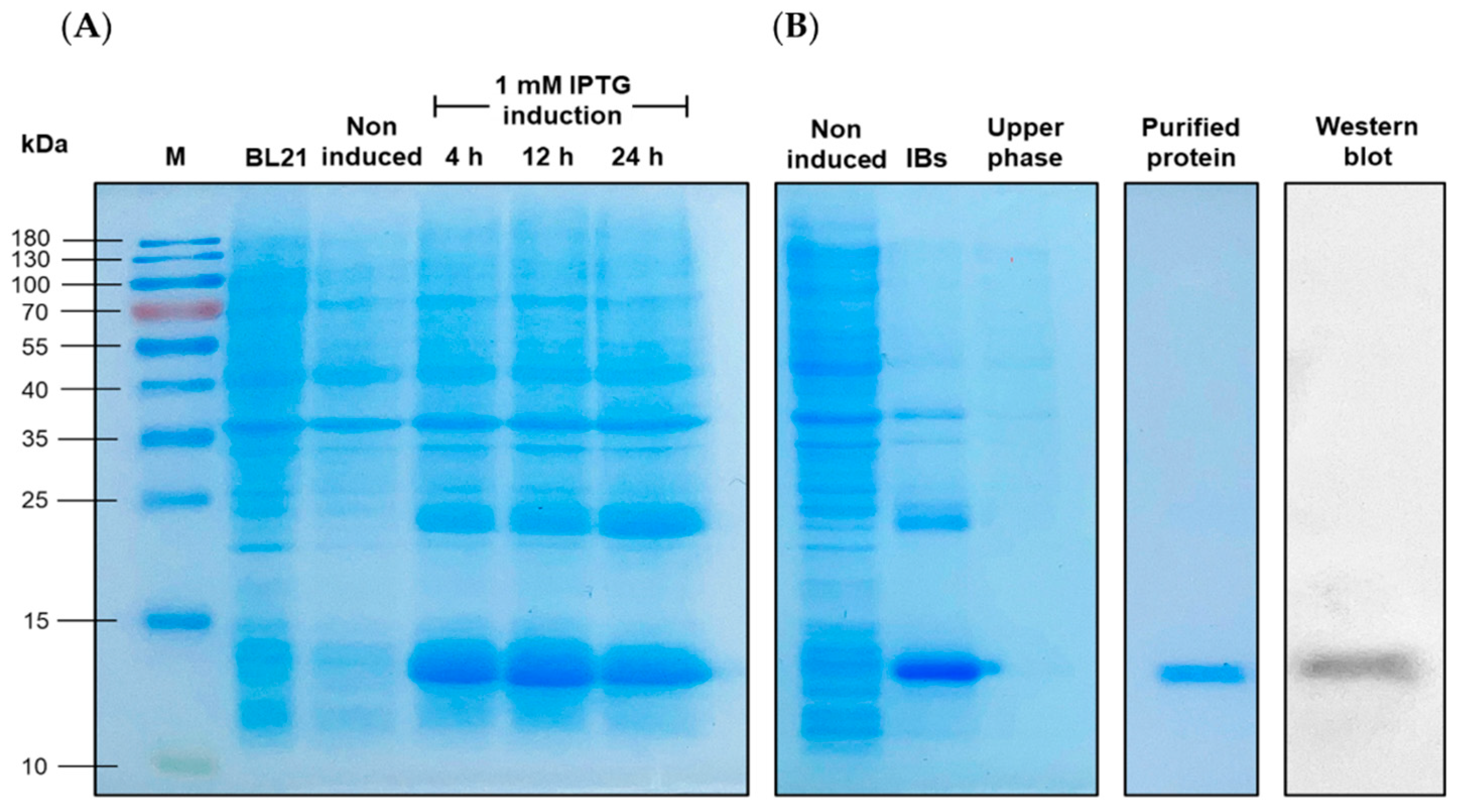
A single positive colony of BL21 cells containing rLcIL-8, as confirmed by nucleotide sequencing, was inoculated in 10 mL of LB broth supplemented with kanamycin. The culture was allowed to incubate overnight in a shaking incubator at 37 °C. Then, 1 mL of prepared bacterial preculture was transferred into 10 mL of LB broth containing kanamycin and incubated in a shaking incubator until the absorbance of the culture reached 0.6 at OD 600 nm. To induce overexpression, IPTG was added to the bacterial culture at a concentration of 1.0 mM. The bacterial culture was then cultivated continuously for a period of 24 hours. During the incubation period, samples were collected at specific time points: 0, 4, 12, and 24 h post-IPTG induction, and each 1 mL aliquot of the bacterial culture was subjected to centrifugation at 7,300 × g for 5 min to facilitate the optimization of protein expression conditions. The expression of recombinant proteins at each time point was analyzed using sodium dodecyl sulfate‒polyacrylamide gel electrophoresis (SDS‒PAGE) (Thermo Fisher Scientific, Waltham, MA, USA).
2.6.3. Purification of Recombinant LcIL-8
The induced recombinant protein (rLcIL-8) was purified with a HiTrap™ Protein A HP column (GE Healthcare, NJ, USA) and an ÄKTA pure protein purification system (ÄKTA pure 25 L, GE Healthcare, NJ, USA) following the manufacturer’s instructions, with a modified protocol designed for inclusion body proteins. Briefly, a 100 mL aliquot of the bacterial culture solution was harvested by centrifugation at 2,300 × g for 5 min. The cell pellets were then resuspended in 10 mL of 20 mM Tris-HCl pH 8.0 and subjected to sonication on ice. Following sonication, the suspension was centrifuged at 12,000 × g for 10 min to pellet the inclusion bodies. These inclusion bodies were subsequently solubilized using 15 mL of solubilization buffer composed of 20 mM Tris-HCl, 0.5 M NaCl, 5 mM imidazole, 6 M urea, and 1 mM 2-mercaptoethanol at pH 8.0. After purification, the recombinant protein was eluted with a linear gradient of 20-500 mM imidazole within an elution buffer at pH 8.0. The resulting protein fractions were collected within fresh Eppendorf tubes and subjected to analysis using SDS‒PAGE techniques, as previously described. The obtained recombinant protein was dialyzed against a solution containing 150 mM NaCl, 20 mM Tris-HCl, pH 8.0 and 20% glycerol, with this process occurring overnight at 4 °C to eliminate small unwanted molecules, such as 2-mercaptoethanol, imidazole and urea. To determine protein concentrations, the Bradford protein assay was employed, and the obtained results were compared to standard serial twofold protein dilutions starting from 2 mg/mL albumin. The quantification of all concentrations was conducted by measuring absorbance at 595 nm utilizing an iMark™ Microplate Absorbance Reader (Bio-Rad, Hercules, CA, USA).
2.6.4. Western Blot Analysis
Western blot analysis was used to separate and determine the molecular weight of the obtained rLcIL-8 protein. The recombinant LcIL-8 protein was separated on a 12% SDS‒PAGE gel and subsequently electrophoretically transferred onto a polyvinylidene difluoride (PVDF) membrane using the Invitrogen™ Power Blotter XL System (Invitrogen, Waltham, MA, USA) with transfer buffer at 100 V for 15 min. Following the transfer, the membrane was blocked with a one-step blocking solution and incubated with a 1:5000 dilution of an anti-6× His tag® antibody (Thermo Fisher Scientific, Waltham, MA, USA) for 2 h at room temperature. Next, the membrane was subjected to three washes with 0.05% Tween-TST. The hybridized protein was visualized using UltraScence Pico Ultra Western Substrate (Bio-Helix, Taiwan) for 1–2 min until color appeared, enabling the identification of specific bands corresponding to the rLcIL-8 protein on the PVDF membrane.
2.7. Effects of rLcIL-8 Protein on Phagocytic Activity (In Vitro)
A phagocytosis assay was employed to investigate the biological activity of the rLcIL-8 protein, which was modified from Puangkaew et al. (2004)[24]. In this experiment, white blood cells were isolated from approximately 100 g of 4 healthy Asian seabasses using a heparinized syringe equipped with a 21-G needle. A volume of 1 mL of whole blood was drawn from the caudal vein and transferred into a 15 mL PE tube containing 2 mL of RPMI 1640 medium (Caissonlabs, North Logan, UT, USA). After gentle mixing, the mixture was then loaded into PE tubes containing 3 mL of LymphoprepTM (Serumwerk Bernburg AG, Oslo, Norway). The subsequent step involved centrifugation in a swing rotor at 600 × g at 25 °C for 30 min. The A band, consisting of lymphocytes and monocytes, was harvested in 1 mL, mixed with 2 mL of Roswell Park Memorial Institute (RPMI) medium, and subjected to centrifugation at 300 × g at 25 °C for 15 min, which was repeated three times. Following dilution, the white blood cell pellets were adjusted to a final concentration of 5 × 106 cells/mL. Subsequently, 200 µL of the prepared white blood cell suspension was carefully loaded onto twelve 22 × 22 mm2 cover glasses, and these cells were allowed to adhere to the glass surface over a 2-hr incubation period. Meanwhile, latex beads (Sigma Aldrich, St Louis, MO, USA) were adjusted to a concentration of 1 × 108 beads/mL using RPMI medium. Four experimental groups were created for each rLcIL-8 protein concentration. In groups 1, 2, and 3, the latex beads were separately incubated with 1, 10 and 100 µg/mL in RPMI medium, respectively. The latex beads in group 4 (control) were solely incubated with RPMI 1640 medium. These experiments were conducted in triplicate.
After 2 h, unattached cells were washed three times with RPMI medium. Subsequently, 200 µL of four latex bead groups, each incubated with different rLcIL-8 protein concentrations, were added to the cover glasses. Phagocytes and latex beads in each group were co-incubated at room temperature for 1.5 h to initiate the phagocytosis process. After 1.5 h, the unattached cells and excess beads were washed three times with RPMI medium. All attached cells in each treatment were stained with Dip-Quick Staining dye (Thermo Fisher Scientific, MA, USA). Subsequently, a minimum of 200 cells were counted under microscopy. The percentage of phagocytosis (PA) and the phagocytic index (PI) were calculated according to the formulas below:
Percent PA = (Number of phagocytic cells with engulfed latex beads
/Number of phagocytes) × 100
PI = Number of engulfed latex beads/phagocytic cells
2.8. Minimum Inhibitory Concentration (MIC) of rLcIL-8 Protein (In Vitro)
In this part, S. iniae was only selected based on its systemic infection properties and for the challenge experiment. It was prepared as detailed in Section 2.5.1. were separately employed to determine the minimal inhibitory concentrations (MICs) using liquid growth inhibition methods. Mueller-Hinton (MH) broth served as a test medium in a 96-well microtiter plate. rLcIL-8 protein was serially diluted twofold from 100 to 0.195 µg/mL (in triplicate). Subsequently, 100 µL of the S. iniae solution with a concentration of 1 × 105 CFU/mL was added (1×104 CFU per well), and the total volume in each well was adjusted to 200 µL. Wells containing only MH broth and bacteria were designated as a negative control, while the positive control group contained only MH broth. Following a 24-hr incubation at 30 °C, the lowest concentration that caused complete inhibition indicated by clear broth was defined as the MIC. Subsequently, the solutions in each well were pipetted and spread onto MH agar. The plates were incubated under the same conditions, and bacterial growth was monitored by colony counting within the range of 30–300 colonies. Bacterial quantity was calculated as colony-forming units per milliliter (CFU/mL) to assess the inhibitory effect on bacterial growth.
2.9. Effects of the rLcIL-8 Protein on Resistance to S. inaie in Asian Seabass (In Vivo)
2.9.1. Experimental Animals
The one hundred and twenty healthy Asian seabass (10 g) detailed in the previous section were used. The fish were randomly selected and placed into twelve separate 250-L fiberglass tanks, each containing 200 L of 5 ppt saline water (10 fish/tank). These tanks maintained the same environmental conditions as previously described. Four groups (3 tanks/group) were set for further experiments below.
2.9.2. Bacterial Challenge
S. iniae strains were cultured and prepared as described above to obtain a bacterial suspension with a final concentration of 1×106 CFU/mL using 0.85% NaCl. The experiment was conducted in triplicate. In each of the four groups, ten fish received intraperitoneal injections with 100 µL of a bacterial suspension. One hour later, three different concentrations of rLcIL-8, including 10, 50 and 100 µg/mL, were injected under the same conditions. In the fourth group, every fish in each tank was similarly injected with 100 µL 0.85% NaCl to serve as controls. Subsequently, the experimental fish in each group were reared for 14 days. During this period, daily mortality was recorded, and any moribund fish found in each tank were examined for bacterial infection in the liver and spleen using the streak plate method on trypticase soy agar (TSA) to confirm S. iniae infection.
2.10. Statistical and Data Analysis
The relative expression ratio of LcIL-8 mRNAs was assessed at different time points by two-way analysis of variance (ANOVA) based on a 3 × 6 factorial design in a completely randomized design (CRD) of 2 factors of 3 bacterial concentrations and 6 detection periods. The percent phagocytosis (PA) and the phagocytic index (PI) were analyzed with one-way ANOVA based on the CRD design. The significance of the means of target parameters was determined by Duncan’s new multiple range test (DMRT) method with SPSS software version 20.0 (IBM SPSS Statistics 24.0). Interactions of the 2 target factors were analyzed throughout. A significance level of p < 0.05 (*) to p < 0.001 (***) was considered statistically significant.
A survival analysis of the Asian seabass in the challenge experiments in each group was performed using the Kaplan‒Meier method with the same software previously mentioned. The level of statistical significance between the control and experimental groups was indicated as *p < 0.05 and ***p < 0.001 using the Student’s t-test.
3. Results
3.1. Characterization of the LcIL-8 cDNA
After obtaining the mature mRNA of the LcIL-8 gene from cloning, a comparison with the GenBank database revealed its 100% identity to the full-length cDNA encoding the interleukin-8 gene of Asian seabass (GenBank accession No. XM_018695863.2). This sequence was named LcIL-8. The LcIL-8 nucleotide sequences contained 237 bp, equal to 79 amino acids (Figure 1A,B). Mature LcIL-8 exhibited a signature arrangement of four cysteines (C30, C32, C56, and C73), two of which were separated by an arginine residue, as observed in the CXC chemokine signature of other vertebrate CXC chemokines. However, the ELR (Glu-Leu-Arg) signature upstream of the CXC motif was not found in LcIL-8 and was replaced by the ILR (Ile-Leu-Arg) motif. A structural analysis of the deduced amino acid sequence of the LcIL-8 protein showed that this protein had a molecular weight of 12.2 kDa and a theoretical isoelectric point (pI) of 8.44.
3.2. Homology and Phylogenetic Analysis of IL-8 Genes in Vertebrates
Homology comparisons of the deduced amino acid sequences revealed that LcIL-8 exhibited 24.8-76.6% sequence identity with IL-8 genes in various fish species and higher vertebrates. Comparisons of the amino acid sequence identity and similarity of LcIL-8 in Asian seabass and other organisms showed that the highest levels of both identity and similarity were shared with yellowfin seabream (Acanthopagrus latus), exhibiting scores of 76.6% and 86.2%, respectively (Supplementary material Table S1).
The phylogenetic tree of the Asian seabass IL-8 protein (LcIL-8), along with other known vertebrate IL-8 proteins, was analyzed using previously reported data. The tree included 37 IL-8 proteins from various vertebrate species derived from 14 higher vertebrates and 23 fish species. The results revealed the presence of three clades of IL-8 proteins. Clade 1 contained five different subgroups of mammals, birds, Western clawed frogs, cartilaginous fish and European seabass IL-8 proteins, while clades 2 and 3 were associated with the teleost fish IL-8 group. LcIL-8 was subclustered within the fish IL-8 first subgroup of clade 2 and exhibited close evolutionary relationships with yellowfin seabream (Acanthopagrus latus) and yellow croaker (Larimichthys crocea) (Figure 2).
3.3. qRT‒PCR Analysis of LcIL-8 Transcripts in Various Tissues of Healthy Asian Seabass
The distribution of the LcIL-8 transcript in healthy Asian seabass tissues was assessed using qRT‒PCR techniques in 12 different tissues from four normal fish. The expression analysis revealed constitutive expression of LcIL-8 among all tested tissues, including muscle, whole blood, heart, liver, spleen, head kidney, trunk kidney, gills, intestine, skin, brain and stomach. Remarkably, the highest expression level was found in the gills, at a level 18.28 ± 0.63-fold higher than that in the muscle (p < 0.05). Additionally, moderate expression levels were observed in the intestine, spleen and stomach at 10.46 ± 0.31, 9.73 ± 0.14 and 6.06 ± 0.53-fold, respectively). In contrast, LcIL-8 mRNA exhibited relatively low expression levels in the muscle, whole blood, heart, liver, head kidney, trunk kidney, skin, and brain (Figure 3).
3.4. Analysis of LcIL-8 Expression in Response to Different Concentrations of S. iniae and F. covae Using qRT‒PCR
3.4.1. Expression Level Analysis in Response to S. iniae Infection
Quantitative real-time PCR was used to quantify LcIL-8 mRNA expression in Asian seabass in response to three concentrations of S. iniae at different time points. In whole blood, a significant upregulation of LcIL-8 mRNA levels was observed in fish injected with 1×105 CFU/fish S. iniae from 6 to 24 h, resulting in relative expression levels of 5.44 ± 0.21, 6.20 ± 0.18, and 9.27 ± 0.09, respectively (p < 0.05). This corresponds to an overall trend in expression levels, indicating a significant upregulation of LcIL-8 from 6 to 24 h for concentrations of 1×105 CFU/fish. However, at 48 and 96 h, all the bacterial concentrations resulted in a significant decrease in the mRNA levels of LcIL-8 (Figure 4A). In the head kidney, the concentrations of 1×105 CFU/fish S. iniae significantly upregulated LcIL-8 mRNA levels at 12 h postinjection, with expression levels of 3.93 ± 0.37-fold, respectively (p < 0.05). Interestingly, at 48 and 96 h postinjection, LcIL-8 exhibited downregulation across all the bacterial concentrations, with extremely low expression levels (Figure 4B).
In the liver (Figure 4C), a similar trend to that observed in the spleen (Figure 4D) at 24 h was noted, with the fish exposed to the highest concentration of bacteria exhibiting a 4.46 ± 0.41-fold upregulation of the target gene, while in the spleen, it was 3.14 ± 0.18-fold (p < 0.05). However, at 48 and 96 h, all the bacterial concentrations resulted in significantly lower expression levels compared to these peak levels (Figure 4C,D). Similarly, in the gills, the highest concentration of S. iniae induced a 3.43 ± 0.05-fold upregulation of LcIL-8 expression at 6 h postinjection (p < 0.05) (Figure 4E). In the intestine, LcIL-8 transcript levels were significantly enhanced by a concentration of 1×103 CFU/fish S. iniae at 12 and 24 h, which resulted in 2.63 ± 0.12 and 3.01 ± 0.37-fold changes, respectively (p < 0.05). An analysis of overall time points and concentrations in the intestine revealed that the highest expression levels of LcIL-8 were observed in fish injected with the moderate dose at 12 and 24 h (Figure 4F).
3.4.2. Expression Level Analysis in Response to F. covae Infection
The qRT‒PCR analysis of LcIL-8 mRNA levels in the six Asian seabass tissues in response to F. covae at different time points revealed that at 6 and 12 h, the highest bacterial concentration significantly induced higher mRNA expression of LcIL-8 in all tested tissues (p < 0.05). However, at 24, 48 and 96 h postinfection, this bacterial concentration resulted in significantly lower expression levels (Figure 5A–F). The mRNA expression showed the highest levels in the liver, gills and intestine Figure 5C, 5E and 5F, respectively). In particular, high levels were observed in the gills at 12 h in fish exposed to 1×106 CFU/mL F. covae, with levels of 11.14 ± 0.05-fold (p < 0.05). Interestingly, by 24 h, the expression levels of LcIL-8 were downregulated and continued to decrease until 48 h postinfection for all concentrations of F. covae. Subsequently, at 96 h, this gene was significantly upregulated again (Figure 5E). In the liver, LcIL-8 mRNA levels gradually increased from 6 to 12 h postinfection at both concentrations of bacteria (1×104 and 1×106 CFU/mL) and were downregulated to normal levels at 24 to 96 h postinfection (Figure 5C). Similarly, in the intestine, the expression of the target gene was significantly upregulated at 6 h in response to concentrations of 1×106 CFU/mL, with relative expression levels of 8.18 ± 0.12-fold (p < 0.05). However, at 12 h postinfection, LcIL-8 gene expression gradually decreased and reached basal levels from 48 to 96 h postinfection at all the bacterial concentrations (Figure 5F).
In the whole blood (Figure 5A), echoing the observed similar trend in the spleen (Figure 5D), LcIL-8 mRNA exhibited a clear and significant increase at 6 and 12 h postinfection. However, when fish were exposed to a bacterial concentration of 1×104 CFU/mL, LcIL-8 mRNA levels were downregulated in the spleen, with a fold-change of 0.97 ± 0.03, while elevated expression persisted exclusively in whole blood at 2.91 ± 0.25-fold (p < 0.05) (Figure 5A,D). In the head kidney, LcIL-8 expression was upregulated from 6 to 12 h postinfection, with the maximal increase occurring at 6 h in response to the highest bacterial concentrations, with fold-changes of 4.69 ± 0.12 (p < 0.05). This trend was consistent with the overall time points and concentrations, with LcIL-8 expression peaking at 6 h in fish injected with the highest dose (Figure 5B).
Figure 6.
SDS-PAGE analysis of the rLcIL-8. Induction analysis of rLcIL-8 at different time points (A). M: Protein marker; BL21; protein form BL21 cells; Non induced; Non-induced BL21 cells containing pET28b (+) rLcIL-8; Lane 4-6; BL21 cells containing pET28b (+) rLcIL-8 induced by 1 mM IPTG at 4, 12 and 24 h, respectively. Characterization inclusion body (IBs) rLcIL-8 and its upper phase in overexpression experiment and purification of the rLcIL-8 and Western blot analysis of refolding rLcIL-8 (B).
Figure 6.
SDS-PAGE analysis of the rLcIL-8. Induction analysis of rLcIL-8 at different time points (A). M: Protein marker; BL21; protein form BL21 cells; Non induced; Non-induced BL21 cells containing pET28b (+) rLcIL-8; Lane 4-6; BL21 cells containing pET28b (+) rLcIL-8 induced by 1 mM IPTG at 4, 12 and 24 h, respectively. Characterization inclusion body (IBs) rLcIL-8 and its upper phase in overexpression experiment and purification of the rLcIL-8 and Western blot analysis of refolding rLcIL-8 (B).
3.6. Effects of rLcIL-8 Protein on Phagocytic Activity and Phagocytic Index
The effects of rLcIL-8 on the phagocytic activity (PA) and phagocytic index (PI) in Asian seabass phagocytes at three different concentrations are illustrated in Figure 7A–C. The results demonstrate that compared to the control group, the rLcIL-8 group exhibited significantly enhanced PA at all concentrations (1, 10 and 100 µg), with PA values of 46.33 ± 3.05, 47.33 ± 1.53, and 58.33 ± 4.16%, respectively (p < 0.05). Notably, the protein concentration of 100 µg exhibited strong PA-inducing ability (Figure 7A). However, the PI showed a significant increase only at protein concentrations of 10 and 100 µg in this group compared to the control, which resulted in indices of 1.56 ± 0.06 and 1.71 ± 0.16, respectively (p < 0.05) (Figure 7B).
3.7. MIC Analysis of the rLcIL-8 Protein against S. iniae
The efficacy of the rLcIL-8 protein against the pathogenic S. iniae strain was assessed via MIC analysis. The results revealed that rLcIL-8 exhibited MIC values ranging from 6.25 to 12.50 μg/mL, with an average value of 10.42 ± 3.61 µg/mL. Furthermore, when examining the bacterial growth rate in CFU/mL units, an inversely proportional relationship was observed between increased protein concentration and decreased number of bacteria. A decline in bacterial growth was evident at a protein concentration of 0.39 µg/mL, with 6.75 ± 0.15 Log CFU/mL (p < 0.05). However, the decrease in the bacterial growth rate tended to remain constant when exposed to protein concentrations ranging from 12.50 to 100.0 µg/mL (Figure 8).
3.8. Anti-S. iniae Effect of the rLcIL-8 Protein against S. iniae
The survival rates of fish treated with rLcIL-8 protein for 14 days after challenge with S. iniae are shown in Figure 9. It is evident that all concentrations of rLcIL-8 resulted in increased survival rates after exposure to the virulent fish pathogen. Specifically, fish that received 50 and 100 µg of rLcIL-8 showed strong resistance to infection, with significantly higher survival rates compared to the control group, which had survival rates of 93.3 ± 0.0% and 83.4 ± 1.7%, respectively (p < 0.001). In addition, fish injected with 10 µg of rLcIL-8 during this period also exhibited significantly higher survival than the control fish at 70.0 ± 2.8% (p < 0.05). However, starting from day 5, the survival rate in each group stabilized and remained consistent until the end of the 14-day observation period.
4. Discussion
In this study, we successfully cloned and characterized cDNA encoding the mature interleukin-8 protein of Asian seabass (LcIL-8). The deduced LcIL-8 sequence exhibited the characteristic arrangement of four cysteine residues (C30, C32, C56, and C73), commonly found in other vertebrate IL-8s. These four conserved cysteine residues played a crucial role in the formation of the tertiary structure and, consequently, influenced the functions of the molecules [25,26]. In addition, another typical feature of the IL-8 gene is its arrangement of amino acids in the CXC pattern, also referred to as the CXC chemokine. In the case of LcIL-8, we observed that two N-terminal cysteines of CXC chemokines were separated by a single nonconserved amino acid residue (Arg31) and connected by disulfide bonds. Basically, CXC chemokines can be classified into two groups based on the presence of the tripeptide motif ELR [27]. All mammalian IL-8 proteins are known to contain the ELR motif; however, the ELR motif is notably absent in teleosts, except for in haddock and Atlantic cod [22,28]. Our sequence analysis revealed an ILR motif upstream of the CXC motif in the deduced LcIL-8 protein, where leucine replaced glutamic acid. Such an amino acid substitution has never been reported in studies of other fish species. Despite the replacement of the ELR motif, which is responsible for recruiting neutrophils and promoting angiogenesis in mammals [14,29], with ILR or other motifs, recombinant proteins of some fish IL-8 variants continue to demonstrate chemotactic activity on neutrophils and macrophages [30,31]. Therefore, LcIL-8 may represent an alternative form of ILR with similar functions. Nevertheless, the exact influence of each motif on the chemoattractant effect in teleost IL-8 still needs further investigation.
The amino acid sequence and phylogenetic analyses of the 37 IL-8 members obtained from fish and higher vertebrates revealed the presence of three distinct clades. Specifically, clades 2 and 3 exhibited associations with the IL-8 group found in teleost fish. Notably, within fish clade 2 of IL-8, a further distinction could be separated into two subgroups. It is known that there are at least two conserved IL-8 lineages in teleosts, in contrast to mammals, which possess only one IL-8 lineage, consistent with previous reports indicating the presence of two IL-8 lineages in fish [32,33]. These two IL-8-like lineages are believed to have diverged through a chromosome duplication event in a teleost ancestor [34]. Among these, LcIL-8 was clustered within the first subgroup of the second clade and exhibited close evolutionary relationships with yellowfin seabream and yellow croaker. Intriguingly, IL-8 from mammals, birds, Western clawed frogs, cartilaginous fish, and European seabass IL-8s were all grouped within the same clade. This observation suggests the possibility that multiple evolutionary events may have occurred in both teleosts and higher vertebrates.
The tissue distribution of LcIL-8 mRNA in healthy Asian seabass was analyzed by qRT‒PCR. The results from qRT‒PCR demonstrated ubiquitous expression in all tested tissues, which was consistent with reports in several fish, such as Atlantic cod [22], Japanese sea perch [35] and blunt snout bream [36]. Additionally, the highest LcIL-8 expression level was observed in the gills, consistent with results from previous studies on IL-8 in other teleost species [37,38], where IL-8 genes were predominantly expressed in the gills, a common target tissue of many pathogens. These results suggest that IL-8 may play a crucial role in the host defense system by promoting the migration of local immune cells, including lymphocytes, monocytes, macrophages and neutrophils, to inflammatory sites and their subsequent adherence to endothelial cells [39,40]. Conversely, the liver, which exhibited relatively weak expression of LcIL-8, was found to have high expression levels in many other teleosts, such as half-smooth tongue sole [41], large yellow croaker [33,42] and South American fish [43]. These results underscore highly inconsistent expression patterns observed in various tissues between different species. Therefore, it could be suggested that IL-8 cytokines have multiple roles in homeostatic functions in both immune and nonimmune tissues due to their species-specific expression in teleost fish.
The route of infection is known to have a significant impact on the host‒pathogen relationship. Therefore, a deeper understanding of the relationship between fish immune responses and different infection routes is necessary. In our current study, we analyzed the expression of LcIL-8 in response to both internal infections by S. iniae and external infection by F. covae using qRT‒PCR in fish immune-related organs. Most of these are the major lymphoid organs in teleost fish and serve as the primary source of macrophages involved in bacterial phagocytosis in innate immunity [44]. The results from this study showed that LcIL-8 was significantly increased in all tested tissues after exposure to both pathogenic bacteria, suggesting that these tissues are the primary sources of immune cells in response to these pathogens. Notably, the highest expression was observed in whole blood after injection with 1×105 CFU/fish S. iniae. This tissue contains numerous immune cells, including lymphocytes, monocytes, and phagocytes, which function as the major sources responsible for producing and releasing cytokines that play a crucial role in the chemotactic activity of various types of immune cells [45]. With the F. covae challenge, a high level of LcIL-8 mRNA was observed in the gills at 12 h in fish infected with 1×106 CFU/mL F. covae. This finding reflects the fact that S. iniae is more systemic in inducing immune defenses than F. covae, which prefers to externally colonize the surface areas of the host, such as the gills and skin, during the early stage of infection. In addition, significant upregulation of LcIL-8 mRNA was also observed in the liver and intestine in response to F. covae. In teleosts, the liver is one of the most important hematopoietic tissues and is responsible for the production of acute phase proteins (APPs), such as transferrin, ceruloplasmin and C-reactive protein (CRP), which are controlled by a variety of different cytokines released during the inflammatory process [46]. Furthermore, epithelial cells in the intestine play a crucial role in producing IL-8 in response to invasive agents [47], and this may be one of the initial signals of acute mucosal inflammation in bacterial infections [48]. Therefore, the strong induction of LcIL-8 after exposure to both pathogenic bacteria in this study indicates its potential proinflammatory role as a chemoattractant, recruiting inflammatory cells to the infected site.
Moreover, in the early stages, at 6–24 h following injection with S. iniae and 6-12 h following immersion with F. covae, respectively, LcIL-8 mRNA displayed varying levels of upregulation in all tested tissues, suggesting that during this critical period, IL-8 mRNAs may be produced vigorously by various cells, including monocytes, macrophages and endothelial cells, in the infected site to eliminate foreign antigens [49]. The results align with those of previous studies in which fish IL-8s were observed to be regulated in response to various stimuli, including bacteria, LPS, and polyinosinic-polycytidylic acid (poly I:C). For example, large yellow croaker exhibited a response to LPS, poly I:C and Vibrio parahaemolyticus [33], Japanese flounder after stimulation with LPS [50] and Nile tilapia challenged with two pathogens of S. agalactiae and Aeromonas hydrophila [51].
However, the expression of the LcIL-8 gene was downregulated and reached basal levels from 48 to 96 h across all the bacterial concentrations of S. iniae. This suggests that IL-8 genes have a brief short half-life in response to this pathogen due to the presence of AU-rich elements (AREs) in the 3′ UTR, which play a critical role in terminating the response [52]. During F. covae challenge, LcIL-8 mRNA expression in most tested tissues, except the gills, gradually decreased and was restored to the basal level from 24 to 96 h postinfection at all the bacterial concentrations. This suggests a suppressive role of IL-8 cytokines on their transcription by feedback inhibition via membrane proteins [53]. Thus, they exhibit a rapid response in the early stages of infection and subsequently degrade later. Furthermore, the regulatory and inhibitory mechanisms of other cytokines are essential since excessive expression of IL-8 genes is directly associated with host tissue damage [54].
Interestingly, in the gills, the expression of LcIL-8 levels in response to all concentrations of F. covae was suppressed at 24 to 48 h, consistent with the mRNA levels of LcIL-8 in the other mentioned tissues. However, at 96 h, this gene was upregulated once again, indicating that the gills are the primary target organs during a natural route infection. These gills not only function as physical barriers against environmental hazards but also serve as gill-associated lymphoid tissue (GIALT) to contain leucocyte populations and produce inflammatory cytokines, including interleukins (ILs) and tumor necrosis factor α (TNFα) [55,56]. It is plausible that the bacteria attempt to breach the gills to enter the bloodstream and initiate a systemic infection [57]. Thus, certain innate immune components within the gills may require longer periods to eliminate F. covae in this experiment.
Based on dose-dependent induction, it was observed that the highest concentration of both bacterial solutions strongly induced LcIL-8 mRNA, followed by the moderate dose, while the lower concentration showed weaker induction. Similar results were observed in Nile tilapia after being induced with the highest dose of 1×109 CFU/mL S. agalactiae, which potentially stimulated CXC chemokine expression compared to lower concentrations of pathogenic bacteria [58]. These findings suggest that a higher bacterial concentration can enhance their ability to effectively spread, colonize, and invade host cells and tissues [59], which are considered one of the primary factors leading to disease in the host.
In the present study, we produced the recombinant LcIL-8 protein to assess its biological functions. The chemotactic activity of various rLcIL-8 proteins has been widely studied in a variety of fish species, including black seabream [60], half-smooth tongue sole [41], Mandarin fish [61], and blunt snout bream [36]. Additionally, in this study, rLcIL-8 was used to perform a phagocytic assay, given that phagocytosis is another important role for most chemokine proteins. It is well known that phagocytosis is fundamental for host defense against invading pathogens and plays a crucial role in immune and inflammatory responses [62]. The results of this study demonstrated a substantial increase in the phagocytic activity (PA) of Asian seabass phagocytes at all concentrations (1, 10, and 100 µg) of rLcIL-8. The highest PA level was observed in the experimental group treated with 100 µg of purified protein, demonstrating the effectiveness of all rLcIL-8 concentrations in enhancing in vitro phagocytic activity. These results are consistent with the findings of a previous report [58], demonstrating that cells treated with 1 and 10 µg/mL rOn-CXC1 and rOn-CXC2 significantly enhance the PA of PBLs in Nile tilapia. Additionally, the experimental group treated with 10 µg/mL of both rOn-CXC1 and rOn-CXC2 exhibited the highest level of PI. In our study, the highest PI level was achieved in the experimental groups treated with 10 and 100 µg of rLcIL-8. This suggests that both concentrations can stimulate phagocytes by enhancing their engulfment capability, while a concentration of 1 µg is more effective at enhancing the number of phagocytic cells rather than improving antigen engulfment.
To determine the efficacy of LcIL-8 in mediating antimicrobial activity against pathogenic S. iniae, an MIC assay was employed. The results revealed that 10.42 ± 3.61 µg/mL rLcIL-8 protein is the minimum concentration needed to inhibit the growth of S. iniae. Furthermore, the inhibitory effect was observed in a dose-dependent manner. These results are similar to those found in other fish, where the fragment derived from the C-terminal end of IL-8 was also determined by antibacterial assay, due to generating IL-8 by cutting at the N-terminus leading to more biologically active forms. Sáenz-Martínez et al. (2021) [63] reported that 10 μM OmIL-8α80-97 synthetic peptide from rainbow trout produced a 50% inhibition of bacterial growth in all tested bacterial strains, including A. salmonicida, Y. ruckeri, Pseudomonas aeruginosa, E. coli and Staphylococcus aureus. Additionally, the IL-8-derived peptide WS12 of snakehead murrel has been demonstrated to possess antibacterial activity against E. coli and Bacillus cereus at concentrations between 3.125 and 50 μM [64]. Previous studies on humans have shown that the IL-8-derived peptide exhibits antimicrobial activity against certain gram-positive and gram-negative bacteria [65]. These findings suggest that IL-8 may play an additional functional role as an antimicrobial effector, either through direct action or by cleaving a peptide derived from its C-terminal end.
Our previous results indicated the additional significant ability of rLcIL-8, which enhances resistance against virulent S. iniae. pathogen. The increased survival rates at all protein concentrations after exposure to S. iniae indicate the protein’s efficacy in protecting Asian seabass against this harmful pathogen. This protective mechanism is supported by its capacity to inhibit S. iniae in vitro and improve antigen engulfment efficiency. Furthermore, another supporting result, as reported by Li et al. (2023)[51], in vitro experiments showed that rOn-IL8 has multiple immune functions in lymphocytes of the head kidney, including downregulation of P38 and P65, upregulation of MyD88 and STAT3, and promotion of the inflammatory response and apoptosis. However, the study by Wang et al. (2016) [66] in channel catfish demonstrated that the cumulative survival of fish intramuscularly injected with PcIL-8 for 4 weeks and then challenged by S. iniae resulted in a low survival rate of 20.0%, which was not significantly different from the control group. These results imply that the protective effects of IL-8 proteins in fish against pathogenic bacteria may depend on (1) fish species, (2) the type, source, isoform, and concentration of IL-8, and (3) the method and duration of protein administration to fish, which may involve different protective mechanisms.
5. Conclusions
In summary, LcIL-8 was successfully characterized for its structure and evolutionary relationships at the molecular level and compared with various IL-8s reported among vertebrates. The qRT‒PCR analysis information obtained in this study demonstrates significant responses of LcIL-8 to two severely pathogenic S. iniae and F. covae. Additionally, the rLcIL-8 protein efficiently enhances the phagocytic activity of Asian seabass phagocytes. Importantly, rLcIL-8 serves as a mediator of antimicrobial activity in vitro and perfectly exhibits its ability to protect fish from streptococcosis, which severely affects the Asian seabass aquaculture industry. Therefore, the insights obtained from this study are crucial for developing effective strategies, including highly potent vaccine adjuvants for preventing diseases caused by harmful pathogenic bacteria in Asian seabass aquaculture.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Homological analyses of LcIL-8 and various IL-8s of other vertebrates.
Author Contributions
Conceptualization, C.M., A.B. and P.Y.; methodology, C.M., A.B. and P.S.; software, C.M. and P.S.; validation, C.M., A.B. and P.S.; formal analysis, C.M and P.S.; investigation, P.S.; resources, C.M.; data curation, C.M.; writing—original draft preparation, C.M. and P.S.; writing—review and editing, C.M.; visualization, A.B. and P.S.; supervision, P.S.; project administration, P.S.; funding acquisition, P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding authors.
Acknowledgments
The authors are grateful to the Faculty of Fisheries, Kasetsart University (KU) for providing generous experimental spaces and equipment support to the student, who is the first author of this article.
Conflicts of Interest
The authors declare that this research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.
References
- The Organisation for Economic Co-operation and Development. Aquaculture production; Species Barramundi (Lates calcarifer). 2022. Available online: https://stats.oecd.org/Index.aspx?DataSetCode=FISH_ AQUA, (accessed on 7 April 2023).
- Suanyuk, N.; Sukkasame, N.; Tanmark, N.; Yoshida, T.; Itami, T.; Thune, R.L.; Tantikitti, C.; Supamattaya, K. Streptococcus iniae infection in cultured Asian seabass (Lates calcarifer) and red tilapia (Oreochromis sp.) in southern Thailand. Warasan Songkhla Nakharin 2010, 32, 341–348. [Google Scholar]
- Chokmangmeepisarn, P.; Thangsunan, P.; Kayansamruaj, P.; Rodkhum, C. Resistome characterization of Flavobacterium columnare isolated from freshwater cultured Asian sea bass (Lates calcarifer) revealed diversity of quinolone resistance associated genes. Aquaculture 2021, 544, 737149. [Google Scholar] [CrossRef]
- Gazal, L.E.D.S.; Brito, K.C.T.D.; Kobayashi, R.K.T.; Nakazato, G.; Cavalli, L.S.; Otutumi, L.K.; Brito, B.G.D. Antimicrobials and resistant bacteria in global fish farming and the possible risk for public health. Arq. Inst. Biol. 2020, 87, e0362019. [Google Scholar] [CrossRef]
- Janeway, C.A. Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Trede, N.S.; Langenau, D.M.; Traver, D.; Thomas, L.A.; Zon, L.I. The use of zebrafish to understand immunity. Immunity 2004, 20, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, J. Expression of cytokines in bacterial and viral infections and their biochemical aspects. J. Biochem. 2000, 127, 525–530. [Google Scholar] [CrossRef]
- Sakai, M.; Hikima, J.I.; Kono, T. Fish cytokines: current research and applications. Fish. Sci. 2021, 87, 1–9. [Google Scholar] [CrossRef]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish Shellfish Immunol. 2013, 35, 1703–1718. [Google Scholar] [CrossRef]
- Larsen, C.G.; Anderson, A.O.; Oppenheim, J.J.; Matsushima, K. Production of interleukin-8 by human dermal fibroblasts and keratinocytes in response to interleukin-1 or tumor necrosis factor. Immunology 1989, 68, 31–36. [Google Scholar]
- Sims, J.E.; Nicklin, M.J.; Bazan, J.F.; Barton, J.L.; Busfield, S.J.; Ford, J.E.; Kastelein, R.A.; Kumar, S.; Lin, H.; Mulero, J.J.; Pan, J.; Pan, Y.; Smith, D.E.; Young, P.R. A new nomenclature for IL-1-family genes. Trends Immunol. 2001, 22, 536–537. [Google Scholar] [CrossRef]
- Bird, S.; Wang, T.; Zou, J.; Cunningham, C.; Secombes, C.J. The first cytokine sequence within cartilaginous fish: IL-1β in the small-spotted catshark (Scyliorhinus canicula). Immunol. 2002, 168, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; Gorman, D.M.; Bazan, J.F.; Kastelein, R.A. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; Van Damme, J.; Walz, A.; Marriott, D. The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar] [CrossRef]
- Najakshin, A.M.; Mechetina, L.V.; Alabyev, B.Y.; Taranin, A.V. Identification of an IL-8 homolog in lamprey (Lampetra fluviatilis): Early evolutionary divergence of chemokines. Eur. J. Immunol. 1999, 29, 375–382. [Google Scholar] [CrossRef]
- Lee, E.Y.; Park, H.H.; Kim, Y.T.; Chung, J.K.; Choi, T.J. Cloning and sequence analysis of the interleukin-8 gene from flounder (Paralichthys olivaceous). Gene 2001, 274, 237–243. [Google Scholar] [CrossRef]
- Laing, K.J.; Zou, J.J.; Wang, T.; Bols, N.; Hirono, I.; Aoki, T.; Secombes, C.J. Identification and analysis of an interleukin 8-like molecule in rainbow trout Oncorhynchus mykiss. Dev. Comp. Immunol. 2002, 26, 433–444. [Google Scholar] [CrossRef]
- Sangrador-Vegas, A.; Lennington, J.B.; Smith, T.J. Molecular cloning of an IL-8-like CXC chemokine and tissue factor in rainbow trout (Oncorhynchus mykiss) by use of suppression subtractive hybridization. Cytokine 2002, 17, 66–70. [Google Scholar] [CrossRef]
- Huising, M.O.; Stet, R.J.; Kruiswijk, C.P.; Savelkoul, H.F.; Verburg-van Kemenade, B.M. Molecular evolution of CXC chemokines: extant CXC chemokines originate from the CNS. Trends Immunol. 2003, 24, 306–312. [Google Scholar] [CrossRef]
- Chen, L.; He, C.; Baoprasertkul, P.; Xu, P.; Li, P.; Serapion, J.; Waldbieser, G.; Wolters, W.; Liu, Z. Analysis of a catfish gene resembling interleukin-8: cDNA cloning, gene structure, and expression after infection with Edwardsiella ictaluri. Dev. Comp. Immunol. 2005, 29, 135–142. [Google Scholar] [CrossRef]
- Nomiyama, H.; Hieshima, K.; Osada, N.; Kato-Unoki, Y.; Otsuka-Ono, K.; Takegawa, S.; Izawa, T.; Yoshizawa, A.; Kikuchi, Y.; Tanase, S.; Miura, R.; Kusuda, J.; Nakao, M.; Yoshie, O. Extensive expansion and diversification of the chemokine gene family in zebrafish: identification of a novel chemokine subfamily CX. BMC Genom. 2008, 9, 222. [Google Scholar] [CrossRef]
- Seppola, M.; Larsen, A.N.; Steiro, K.; Robertsen, B.; Jensen, I. Characterisation and expression analysis of the interleukin genes, IL-1beta, IL-8 and IL-10, in Atlantic cod (Gadus morhua L.). Mol. Immunol. 2008, 45, 887–897. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Puangkaew, J.; Kiron, V.; Somamoto, T.; Okamoto, N.; Satoh, S.; Takeuchi, T.; Watanabe, T. Nonspecific immune response of rainbow trout (Oncorhynchus mykiss Walbaum) in relation to the different status of vitamin E and highly unsaturated fatty acids. Fish Shellfish Immunol. 2004, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, E.T.; Weber, I.T.; St Charles, R.; Xuan, J.C.; Appella, E.; Yamada, M.; Matsushima, K.; Edwards, G.C.; Gronenborn, A.M. Crystal structure of interleukin 8: symbiosis of NMR and crystallography. Proc. Natl. Acad. Sci. 1991, 88, 502–506. [Google Scholar] [CrossRef]
- Rajarathnam, K.; Sykes, B.D.; Dewald, B.; Baggiolini, M.; Clark-Lewis, I. Disulfide bridges in interleukin-8 probed using non-natural disulfide analogues: Dissociation of roles in structure from function. Biochemistry 1999, 38, 7653–7658. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.C.; Mayo, K.H. Chemokines from a structural perspective. Int. J. Mol. Sci. 2017, 18, 2088. [Google Scholar] [CrossRef] [PubMed]
- Corripio-Miyar, Y.; Bird, S.; Tsamopoulos, K.; Secombes, C.J. Cloning and expression analysis of two pro-inflammatory cytokines, IL-1β and IL-8, in haddock (Melanogrammus aeglefinus). Mol. Immunol. 2007, 44, 1361–1373. [Google Scholar] [CrossRef]
- Kaiser, P.; Rothwell, L.; Avery, S.; Balu, S. Evolution of the interleukins. Dev. Comp. Immunol. 2004, 28, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, G.; Zhang, R.; Liu, L.; Zhu, J.; Zhu, H. Molecular characterization and biological functioning of interleukin-8 in Siberian sturgeon (Acipenser baeri). Fish Shellfish Immunol. 2019, 90, 91–101. [Google Scholar] [CrossRef]
- Umasuthan, N.; Bathige, S.D.N.K.; Thulasitha, W.S.; Oh, M.; Lee, J. Comparative characterization of two CXCl8 homologs in Oplegnathus fasciatus: Genomic, transcriptional and functional analyses. Biomolecules 2020, 10, 1382. [Google Scholar] [CrossRef]
- Abdelkhalek, N.K.; Komiya, A.; Kato-Unoki, Y.; Somamoto, T.; Nakao, M. Molecular evidence for the existence of two distinct IL-8 lineages of teleost CXC-chemokines. Fish Shellfish Immunol. 2009, 27, 763–767. [Google Scholar] [CrossRef]
- Li, C.; Yao, C.L. Molecular and expression characterizations of interleukin-8 gene in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2013, 34, 799–809. [Google Scholar] [CrossRef]
- Ravi, V.; Venkatesh, B. Rapidly evolving fish genomes and teleost diversity. Curr. Opin. Genet. Dev. 2008, 18, 544–550. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, H.; Yang, K.; Jiang, S. Molecular cloning and mRNA expression analysis of interleukin-8 gene in Japanese sea perch (Lateolabrax japonicus). Mol. Biol. Rep. 2009, 36, 1099–1105. [Google Scholar] [CrossRef]
- Li, H.; Li, H.; Liu, Y.; Zheng, Y.; Zhang, M.; Wang, X.; Cui, H.; Wang, H.; Zhao, X.; Chen, X.; Cheng, H.; Xu, J.; Ding, Z. Molecular characterization and expression patterns of CXCL8 gene from blunt snout bream (Megalobrama amblycephala) and its chemotactic effects on macrophages and neutrophils. Dev. Comp. Immunol. 2023, 142, 104658. [Google Scholar] [CrossRef]
- Li, H.; Cai, Y.; Xie, P.; Li, G.; Hao, L.; Xiong, Q. Identification and expression profiles of IL-8 in bighead carp (Aristichthys nobilis) in response to microcystin-LR. Arch. Environ. Contam. Toxicol. 2013, 65, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, H.C.; Park, C.J.; Park, J.W.; Lee, Y.M.; Kim, W.J. Interleukin-8 (IL-8) expression in the olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) challenge. Dev. Reprod. 2019, 23, 231. [Google Scholar] [CrossRef]
- Hoffmann, E.; Dittrich-Breiholz, O.; Holtmann, H.; Kracht, M. Multiple control of interleukin-8 gene expression. J. Leukoc. Biol. 2002, 72, 847–855. [Google Scholar] [CrossRef]
- Rességuier, J.; Dalum, A.S.; Du Pasquier, L.; Zhang, Y.; Koppang, E.O.; Boudinot, P.; Wiegertjes, G.F. Lymphoid tissue in teleost gills: Variations on a theme. Biology 2020, 9, 127. [Google Scholar] [CrossRef]
- Sun, J.S.; Zhao, L.; Sun, L. Interleukin-8 of Cynoglossus semilaevis is a chemoattractant with immunoregulatory property. Fish Shellfish Immunol. 2011, 30, 1362–1367. [Google Scholar] [CrossRef]
- Mu, Y.; Wang, K.; Ao, J.; Chen, X. Molecular characterization and biological effects of a CXCL8 homologue in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2015, 44, 462–470. [Google Scholar] [CrossRef]
- Carriero, M.M.; Henrique-Silva, F.; Meira, C.M.; Gato, I.M.Q.; Caetano, A.R.; Lobo, F.P.; Alves, A.L.; Varela, E.S.; A. A.M, Maia. Molecular characterization and gene expression analysis of the pro-inflammatory cytokines IL-1β and IL-8 in the South American fish Piaractus mesopotamicus challenged with Aeromonas dhakensis. Genet. Mol. Biol. 2020, 43, e20200006. [Google Scholar] [CrossRef]
- Bjørgen, H.; Koppang, E.O. Anatomy of teleost fish immune structures and organs. Immunogenetics 2021, 73, 53–63. [Google Scholar] [CrossRef]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef]
- Bode, J.G.; Albrecht, U.; Häussinger, D.; Heinrich, P.C.; Schaper, F. Hepatic acute phase proteins–regulation by IL-6-and IL-1-type cytokines involving STAT3 and its crosstalk with NF-κB-dependent signaling. Eur. J. Cell Biol. 2012, 91, 496–505. [Google Scholar] [CrossRef]
- Eckmann, L.; Smith, J.R.; Housley, M.P.; Dwinell, M.B.; Kagnoff, M.F. Analysis by high-density cDNA arrays of altered gene expression in human intestinal epithelial cells in response to infection with the invasive enteric bacteria Salmonella. J. Biol. Chem. 2000, 275, 14084–14094. [Google Scholar] [CrossRef]
- Eckmann, L.; Kagnoff, M.F.; Fierer, J. Epithelial cells secrete the chemokine interleukin-8 in response to bacterial entry. Infect. Immun. 1993, 61, 4569–4574. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Zhang, F.; Liu, Y.; Zang, L.; Wang, X.; Qiu, X. Expression analysis of three immune genes IL-8, IL-6 and IL-1β in the Japanese flounder (Paralichthys olivaceus). Isr. J. Aquac. 2021, 73, 1–12. [Google Scholar] [CrossRef]
- Li, X.; Jiang, B.; Zhang, Z.; Huang, M.; Feng, J.; Huang, Y.; Amoah, K.; Huang, Y.; Jian, J. Interleukin-8 involved in Nile tilapia (Oreochromis niloticus) against bacterial infection. Fish Shellfish Immunol. 2023, 141, 109004. [Google Scholar] [CrossRef]
- Mahmoud, L.; Al-Enezi, F.; Al-Saif, M.; Warsy, A.; Khabar, K.S.; Hitti, E.G. Sustained stabilization of Interleukin-8 mRNA in human macrophages. RNA Biol. 2014, 11, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Nibbs, R.J.; Graham, G.J. Immune regulation by atypical chemokine receptors. Nat. Rev. Immunol. 2013, 13, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Wang, A.; Martin, B.; Koehler, D.R.; Zeitlin, P.L.; Tanawell, A.K.; Hu, J. Down-regulation of IL-8 expression in human airway epithelial cells through helper-dependent adenoviral-mediated RNA interference. Cell Res. 2005, 15, 111–119. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Lazado, C.C.; Brinchmann, M.F.; Kiron, V. Infection-induced changes in expression of antibacterial and cytokine genes in the gill epithelial cells of Atlantic cod, Gadus morhua during incubation with bacterial pathogens. Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 2010, 156, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Salinas, I.; Zhang, Y.A.; Sunyer, J.O. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef]
- Declercq, A.M.; Haesebrouck, F.; Van den Broeck, W.; Bossier, P.; Decostere, A. Columnaris disease in fish: A review with emphasis on bacterium-host interactions. Vet. Res. 2013, 44, 1–17. [Google Scholar] [CrossRef]
- Nakharuthai, C.; Srisapoome, P. Molecular identification and dual functions of two different CXC chemokines in Nile tilapia (Oreochromis niloticus) against streptococcus agalactiae and Flavobacterium columnare. Microorganisms 2020, 8, 1058. [Google Scholar] [CrossRef]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Zhonghua, C.; Chunpin, G.; Yong, Z.; Kezhi, X.; Yaou, Z. Cloning and bioactivity analysis of a CXC ligand in black seabream Acanthopagrus schlegeli: The evolutionary clues of ELR+ CXC chemokines. BMC Immunol. 2008, 9, 1–12. [Google Scholar] [CrossRef]
- Wang, G.L.; Wang, M.C.; Zhang, X.W.; Chang, M.X.; Xie, H.X.; Nie, P. Molecular cloning, biological effect, and tissue distribution of interleukin-8 protein in mandarin fish (Siniperca chuasti) upon Flavobacterium columnare infection. Fish Shellfish Immunol. 2017, 66, 112–119. [Google Scholar] [CrossRef]
- Flannagan, R.S.; Jaumouillé, V.; Grinstein, S. The cell biology of phagocytosis. Annu. Rev. Pathol. 2012, 7, 61–98. [Google Scholar] [CrossRef]
- Sáenz-Martínez, D.E.; Santana, P.A.; Aróstica, M.; Forero, J.C.; Guzmán, F.; Mercado, L. Immunodetection of rainbow trout IL-8 cleaved-peptide: Tissue bioavailability and potential antibacterial activity in a bacterial infection context. Dev. Comp. Immunol. 2021, 124, 104182. [Google Scholar] [CrossRef]
- Sathyamoorthi, A.; Bhatt, P.; Ravichandran, G.; Kumaresan, V.; Arasu, M.V.; Al-Dhabi, N.A.; Arockiaraj, J. Gene expression and in silico analysis of snakehead murrel interleukin 8 and antimicrobial activity of C-terminal derived peptide WS12. Vet. Immunol. Immunopathol. 2017, 190, 1–9. [Google Scholar] [CrossRef]
- Björstad, A.; Fu, H.; Karlsson, A.; Dahlgren, C.; Bylund, J. Interleukin-8-derived peptide has antibacterial activity. Antimicrob. Agents Chemother. 2005, 49, 3889–3895. [Google Scholar] [CrossRef]
- Wang, E.; Wang, J.; Long, B.; Wang, K.; He, Y.; Yang, Q.; Chen, D.; Geng, Y.; Huang, X.; Ouyang, P.; Lai, W. Molecular cloning, expression and the adjuvant effects of interleukin-8 of channel catfish (Ictalurus punctatus) against Streptococcus iniae. Sci. Rep. 2016, 6, 29310. [Google Scholar] [CrossRef]
Figure 1.
Structure of the mature cDNA encoding LcIL-8 gene used to produce LcIL-8 protein (A) and a schematic structure of recombinant LcIL-8 protein (rLcIL-8). The upper and lower numbers in Figure 1A indicate nucleotide and amino acid sequences, respectively.
Figure 1.
Structure of the mature cDNA encoding LcIL-8 gene used to produce LcIL-8 protein (A) and a schematic structure of recombinant LcIL-8 protein (rLcIL-8). The upper and lower numbers in Figure 1A indicate nucleotide and amino acid sequences, respectively.
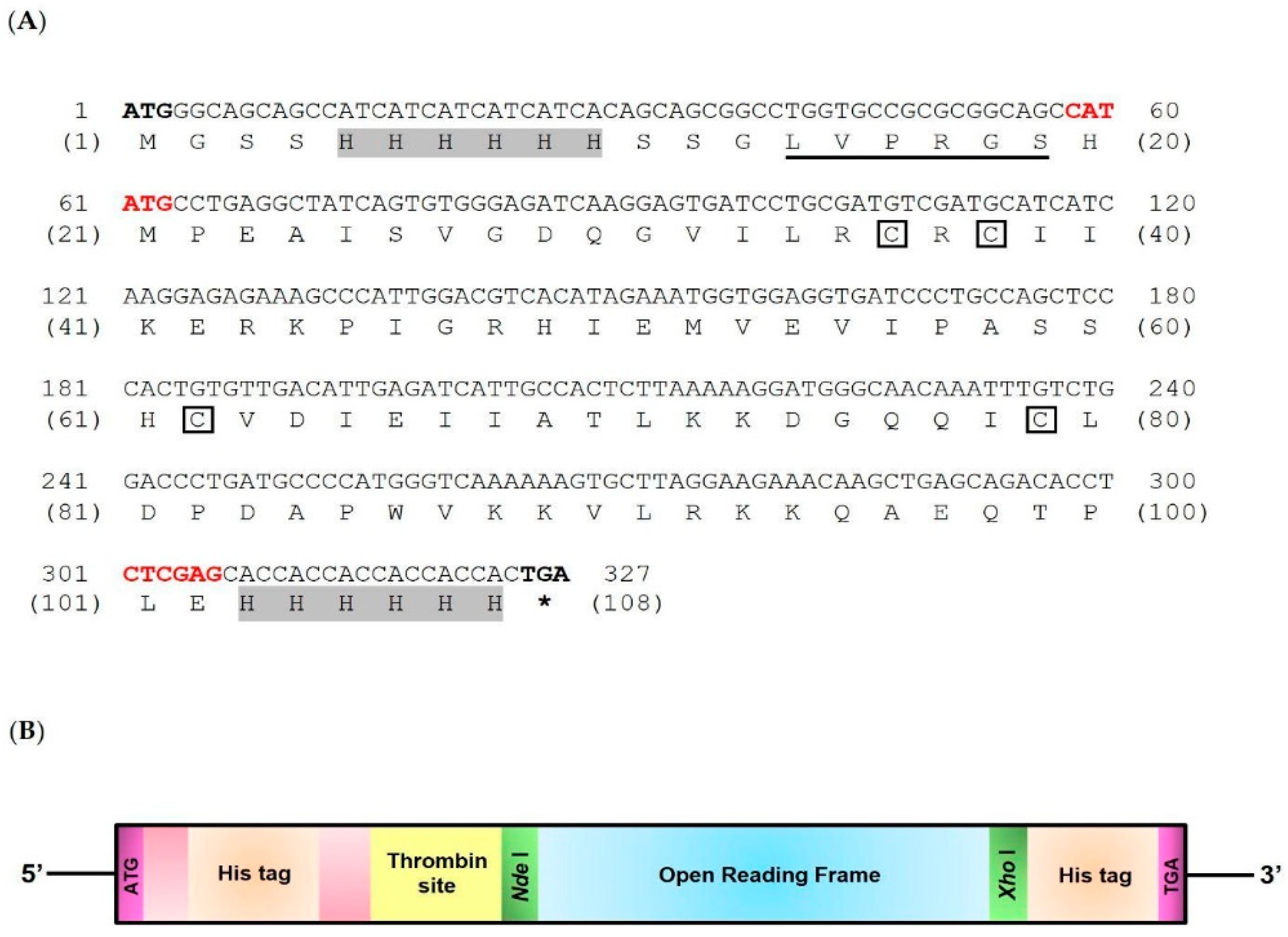
Figure 2.
Phylogenetic analysis of LcIL-8 and other IL-8 proteins of various vertebrates. Accession numbers of each IL-8 are indicated in the parenthesis.
Figure 2.
Phylogenetic analysis of LcIL-8 and other IL-8 proteins of various vertebrates. Accession numbers of each IL-8 are indicated in the parenthesis.
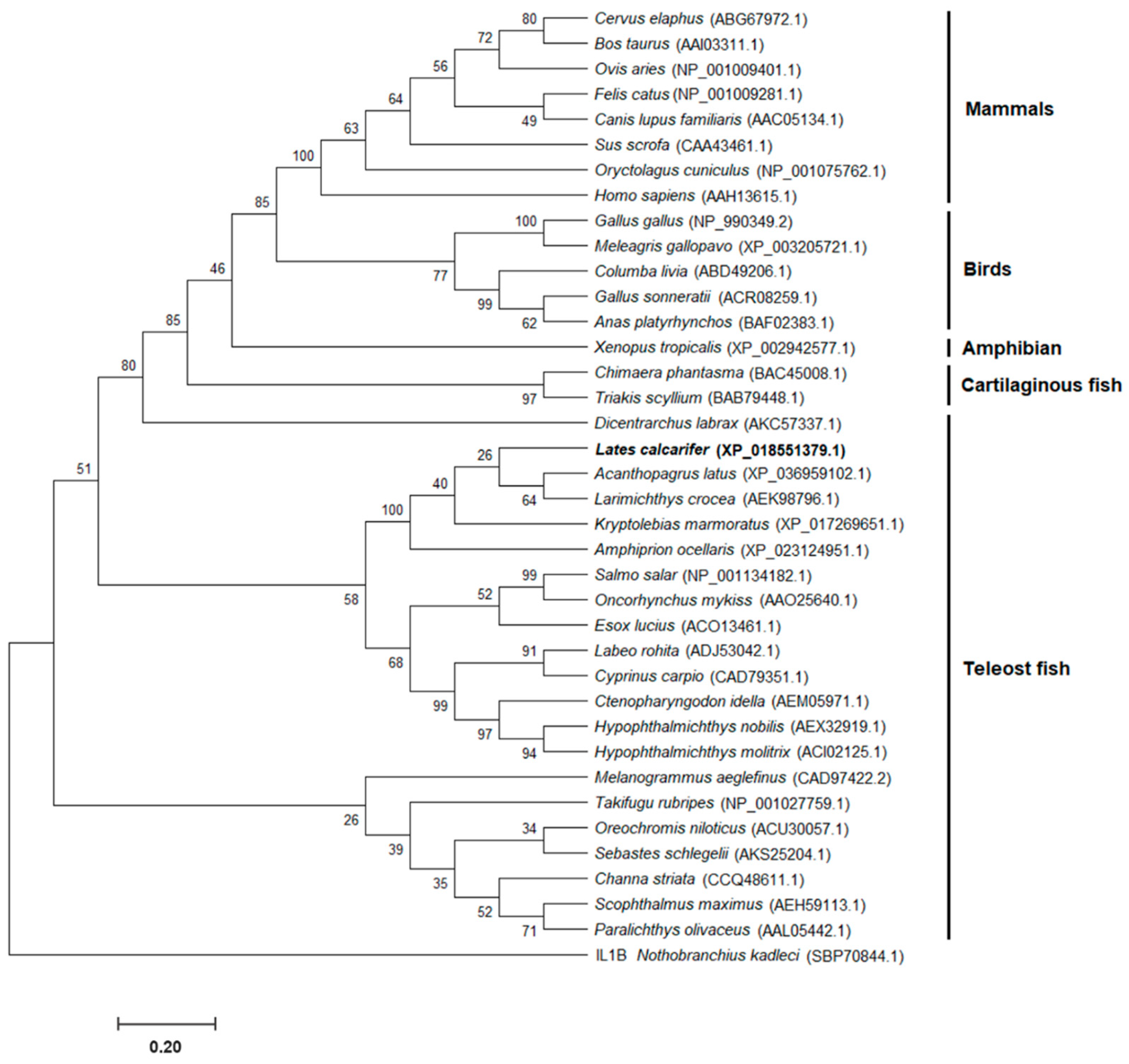
Figure 3.
Quantitative real-time RT-PCR (qRT-PCR) analysis of LcIL-8 transcripts in various tissues. MC; muscle, WB; whole blood, HR; heart, LV; liver, SP; spleen, HK; head kidney, TK; trunk kidney; GL; gills; INT; intestine; SK; skin, BR; brain and SM; stomach. Different letters indicate significant differences (p < 0.05).
Figure 3.
Quantitative real-time RT-PCR (qRT-PCR) analysis of LcIL-8 transcripts in various tissues. MC; muscle, WB; whole blood, HR; heart, LV; liver, SP; spleen, HK; head kidney, TK; trunk kidney; GL; gills; INT; intestine; SK; skin, BR; brain and SM; stomach. Different letters indicate significant differences (p < 0.05).
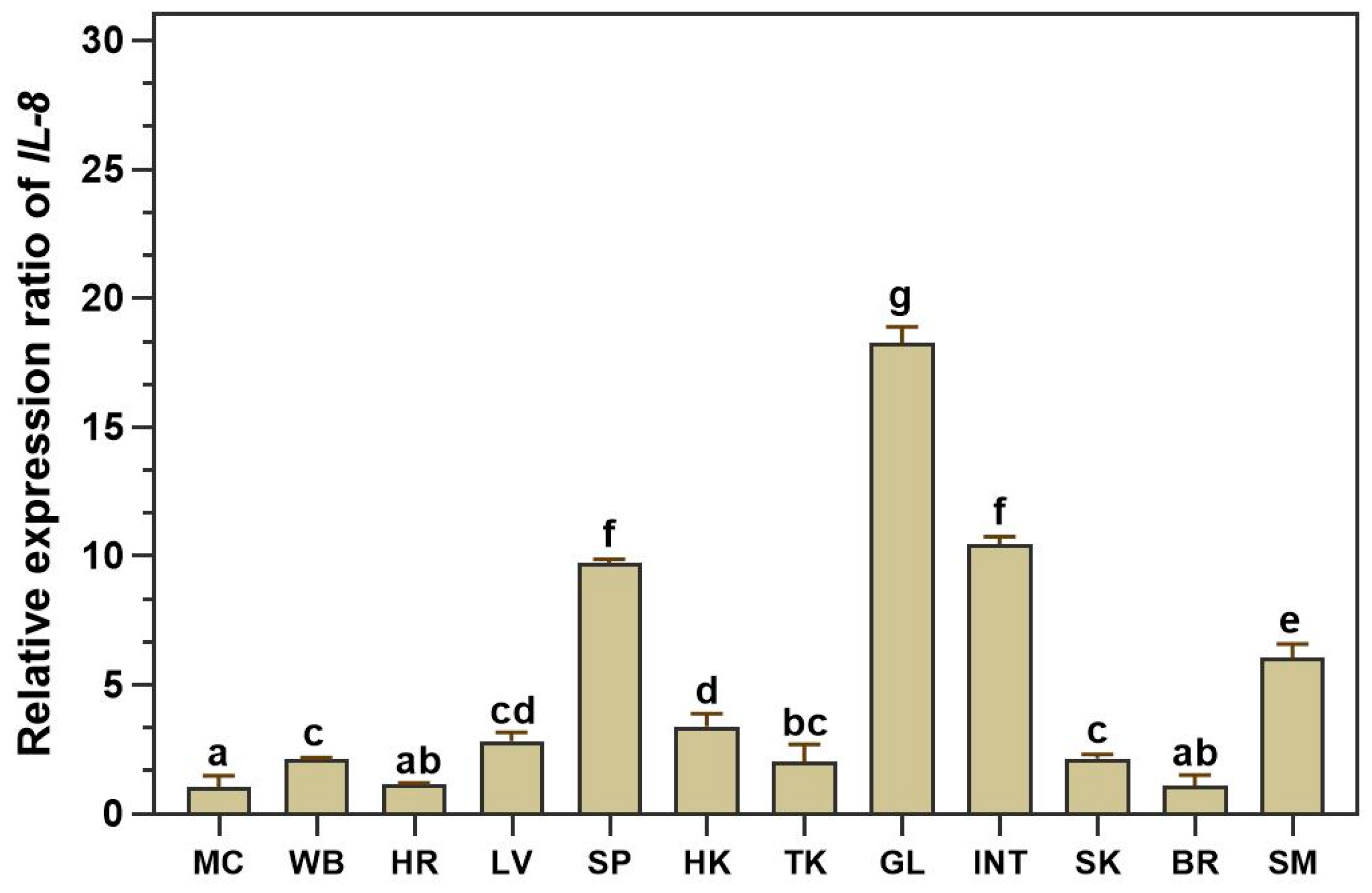
Figure 4.
qRT-PCR analysis of LcIL-8 transcripts in whole blood (A), head kidney (B), liver (C), spleen (D), gills (E) and intestine (F) of Asian seabass injected with different concentrations of S. iniae at different time courses. Different letters indicate significant differences (p < 0.05).
Figure 4.
qRT-PCR analysis of LcIL-8 transcripts in whole blood (A), head kidney (B), liver (C), spleen (D), gills (E) and intestine (F) of Asian seabass injected with different concentrations of S. iniae at different time courses. Different letters indicate significant differences (p < 0.05).
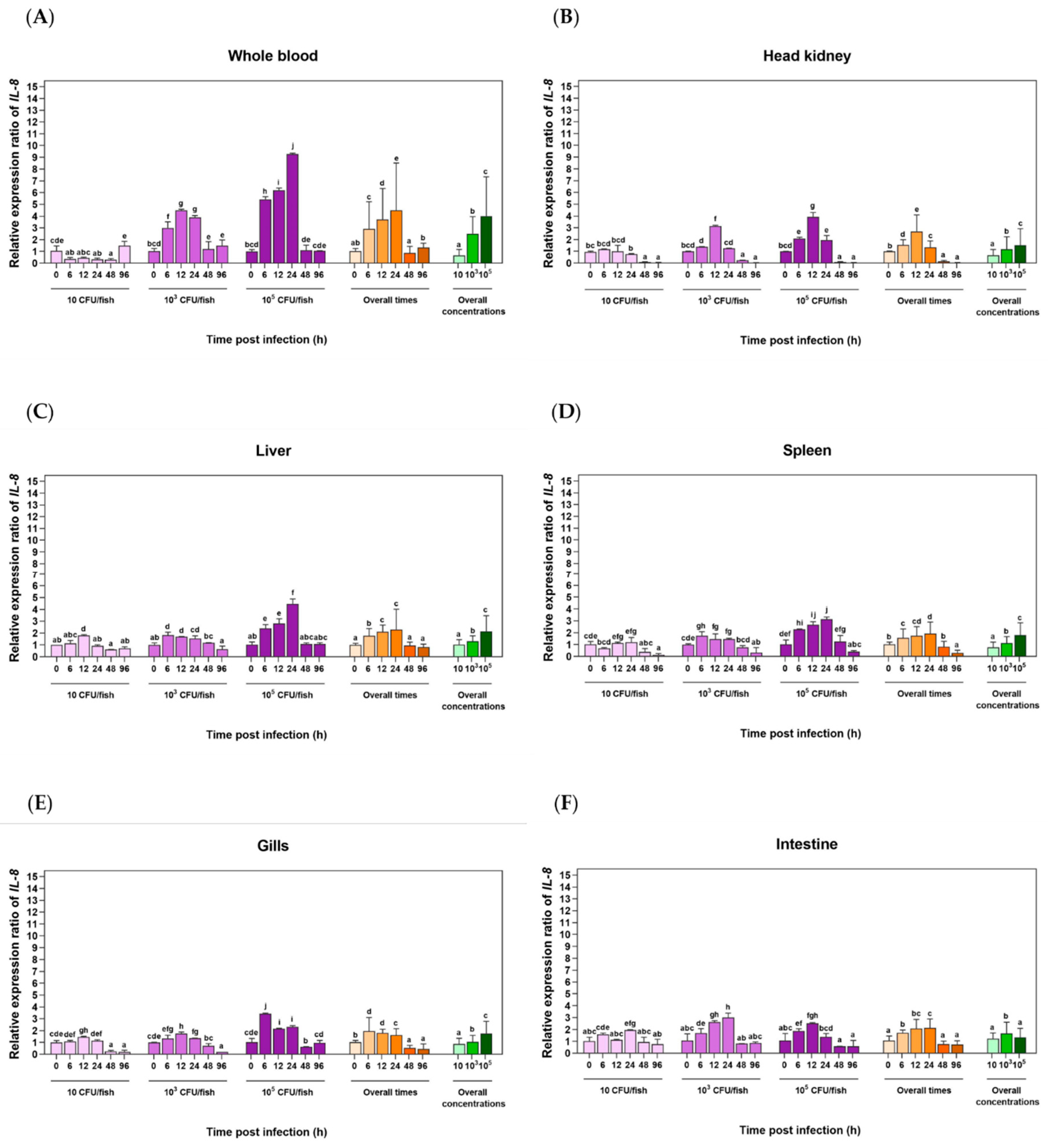
Figure 5.
qRT-PCR analysis of LcIL-8 transcripts in whole blood (A), head kidney (B), liver (C), spleen (D), gills (E) and intestine (F) of Asian seabass immersed with different concentrations of F. covae at different time courses. Different letters indicate significant differences (p < 0.05).
Figure 5.
qRT-PCR analysis of LcIL-8 transcripts in whole blood (A), head kidney (B), liver (C), spleen (D), gills (E) and intestine (F) of Asian seabass immersed with different concentrations of F. covae at different time courses. Different letters indicate significant differences (p < 0.05).
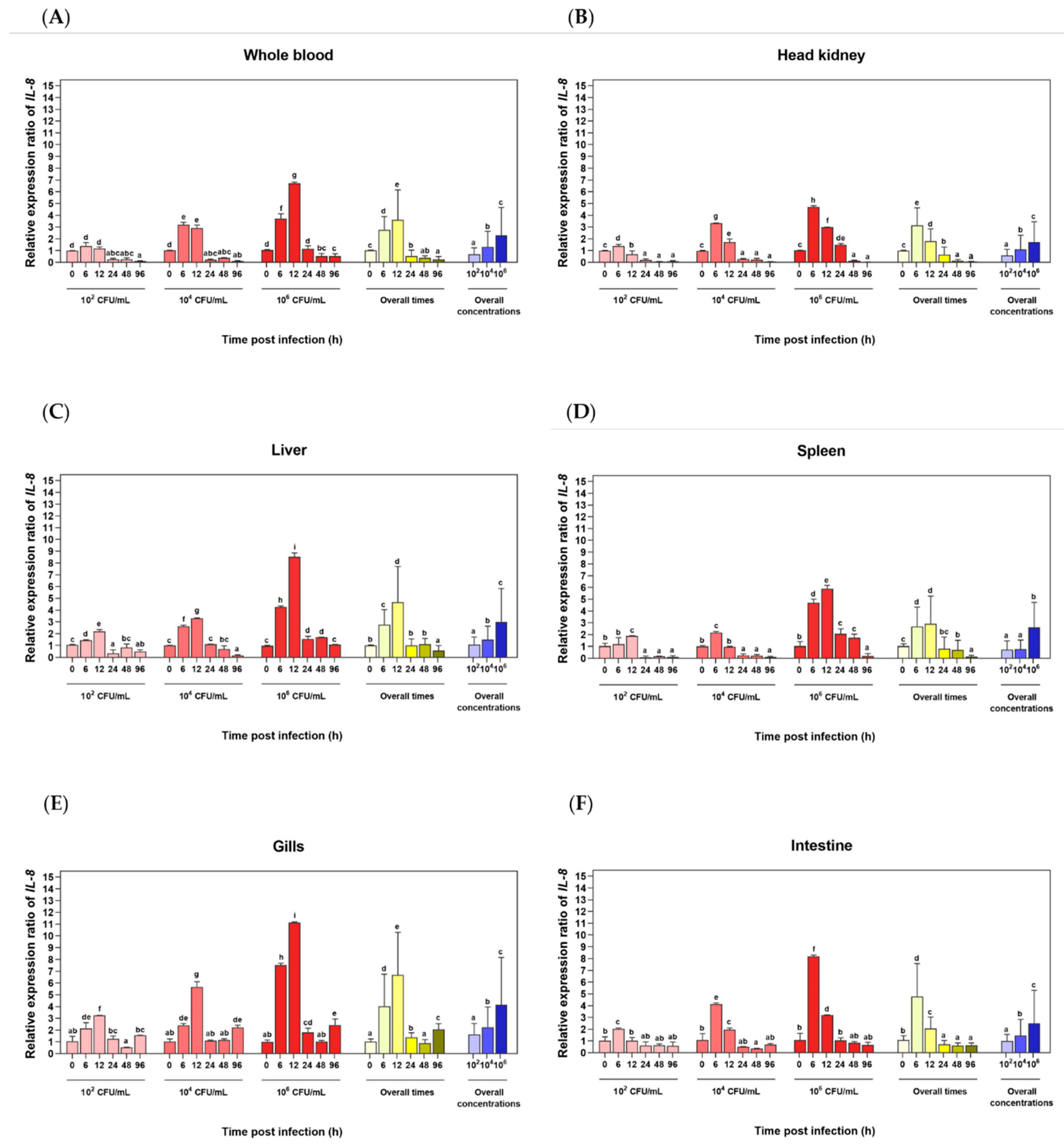
Figure 7.
Phagocytic activity (A), phagocytic index (B) and phagocyting cell pictures of Asian seabass phagocytes induced by various concentrations of rLcIL-8. Different letters indicate significant differences (p < 0.05).
Figure 7.
Phagocytic activity (A), phagocytic index (B) and phagocyting cell pictures of Asian seabass phagocytes induced by various concentrations of rLcIL-8. Different letters indicate significant differences (p < 0.05).
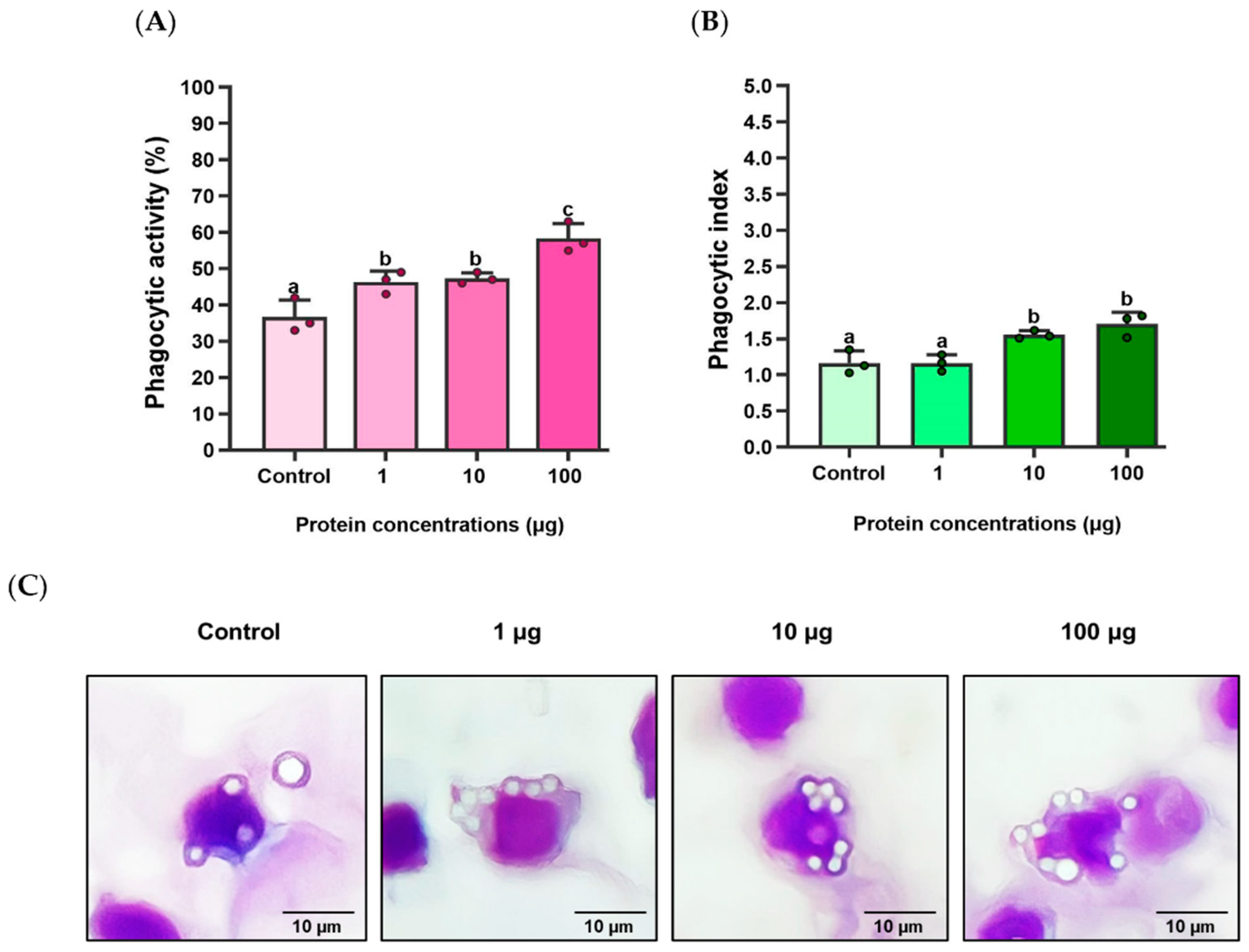
Figure 8.
MIC analysis of S. iniae exposed with various concentrations of rLcIL-8.
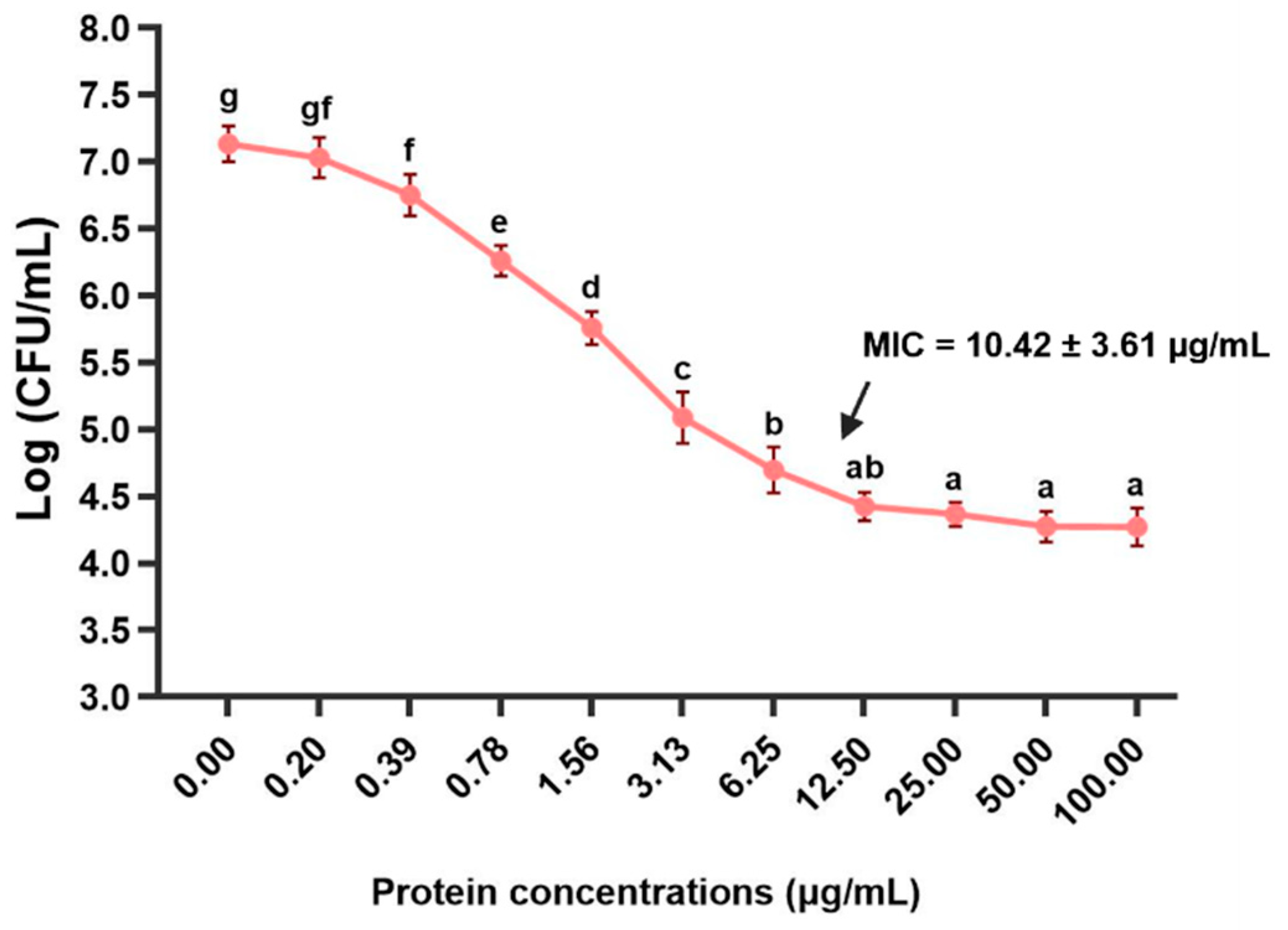
Figure 9.
Survival analysis of Asian seabass was first injected with S. iniae following secondary injection with various concentrations of rLcIL-8. Asterisks “*” and “***” indicate significant differences at p < 0.05 and p < 0.001, respectively.
Figure 9.
Survival analysis of Asian seabass was first injected with S. iniae following secondary injection with various concentrations of rLcIL-8. Asterisks “*” and “***” indicate significant differences at p < 0.05 and p < 0.001, respectively.
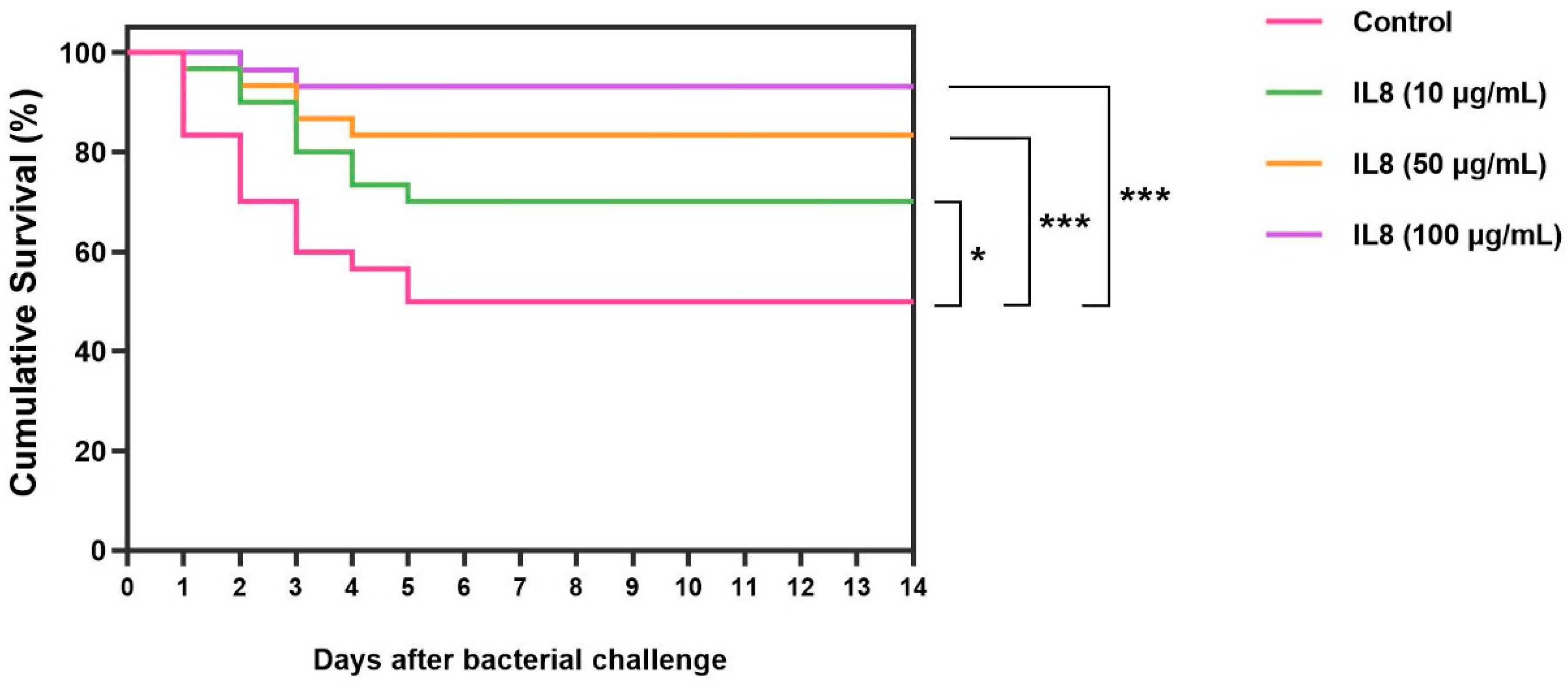

Table 1.
Specific primers used in the current study.
| Primer name | Sequence (5′-3′) | Amplicon size |
|---|---|---|
| LcIL8 F | CATATGCCTGAGGCTATCAGTGTGGGAGAT | 237 bp |
| LcIL8 R | CTCGAGAGGTGTCTGCTCAGCTTGTTTCTT | |
| LcIL-8 qF | TGATCCTGCGATGTCGATGCAT | 206 bp |
| LcIL-8 qR | AGGTGTCTGCTCAGCTTGTTTC | |
| Lc-β-actin qF | TACCCCATTGAGCACGGTATTG | 150 bp |
| Lc-β-actin qF | TCTGGGTCATCTTCTCCCTGTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



