
Submitted:
30 November 2023
Posted:
01 December 2023
You are already at the latest version
Abstract
RNA pol II assembly occurs in the cytoplasm before the translocation of the enzyme to the nucleus. Affecting assembly influences mRNA transcription in the nucleus and mRNA decay in the cytoplasm. However, very little is known about the consequences on ncRNA synthesis. In this work, we show that impairment of RNA pol II assembly leads to a decrease in cryptic non-coding RNAs (preferentially CUTs and SUTs). This alteration is partially restored upon overcoming the assembly defect. Notably, this drop in ncRNAs is only partially dependent on the nuclear exosome, which suggests a major specific effect of enzyme assembly. Our data also point out a defect in transcription termination and lead as to proposes that CTD phosphatase Rtr1 could be involved in this process.
Keywords:
transcription
; RNA polymerases
; ncRNAs
; RNA polymerases assembly
; Rtr1 CTD phosphatase
; NNS termination
; exosome
; CUTs
; SUTs
; Saccharomyces cerevisiae
1. Introduction
Transcription is a well-studied step in gene expression. In eukaryotes, transcription is performed by three different RNA polymerases (five in plants) that act in coordination with complex transcriptional machinery [1,2,3]. Specifically, transcription mediated by RNA pol II has been widely studied because it is responsible for transcribing the majority of the genome, including all the mRNAs and some non-coding RNAs (ncRNAs), such as: small nuclear RNAs (snRNAs); small nucleolar RNAs (snoRNAs); small Cajal body-specific RNAs (scaRNAs); microRNAs (miRNAs); long non-coding RNAs (lncRNAs); piwi-interacting RNAs (piRNAs); circular RNAs (circRNAs) and small interfering RNAs (siRNAs) [1,4,5,6]. In addition, the fragmentation of some of these non-coding RNAs produces other types of small regulatory RNAs, such as sno-RNA-derived fragments (sdRNAs) [7].
Pervasive transcription, which refers to the idea that transcripts are not limited to clearly defined functional characteristics like genes [8], can be mediated by RNA pol II and has been reported in several organisms [9,10,11,12,13,14,15]. In budding yeast Saccharomyces cerevisiae, many non-coding transcripts that result from RNA pol II transcription have been identified using high-resolution omics techniques.
The ncRNAs that derive from the pervasive transcription of RNA pol II have been further classified according to their sensitivity to different decay pathways, transcription termination pathways or the specific conditions for their expression (reviewed in [16]). Of these ncRNAs, different types have been described: “Cryptic Unstable Transcripts” (CUTs), degraded by the nuclear exosome, “Stable Unannotated Transcripts” (SUTs) that exist stably in cells, and “Xrn1-sensitive unstable transcripts” (XUTs) that are degraded by cytoplasmic exonuclease Xrn1 [9,10,17,18,19]. While SUTs are detected in wild-type (WT) cells, CUTs and XUTs are detected mainly upon the inactivation of Rrp6 subunit of the nuclear exosome or Xrn1, respectively. In addition, “Nrd1-unterminated transcripts” (NUTs) have been described, which appear upon the depletion of the Nrd1 termination factor [20]. “Set2-repressed antisense transcripts” (SRATs) [21] and “meiotic unannotated transcripts” (MUTs) have also been reported [22]. Most non-coding transcripts have overlapping sequences [23,24], and many NUTs overlap CUTs [20], while XUTs significantly overlap SUTs and CUTs [18,23,24]
As the synthesis of non-functional ncRNAs could interfere with the correct production of protein-coding mRNAs through different mechanisms [25], cells exhibit strategies to control the production of pervasive transcripts. In yeast, the most prominent transcriptome surveillance mechanism coordinates both transcription termination and nuclear RNA degradation thorough the action of the Nrd1-Nab3-Sen1(NNS) transcription termination complex, the nuclear exosome and the TRAMP complex [25,26]. Briefly in this mechanism, CUTs and other snRNAs and snoRNAs (sn(o)RNAs) terminate by the Nrd1-Nab3-Sen1 (NNS) pathway [25,27]). Nrd1, which recognises the RNA pol II phosphorylated at Ser5 residues of the carboxi-terminal domain (CTD) of the Rpb1 subunit, also recruits TRAMP4/5 (Trf4/5-Air2/1-Mtr4-polyadenylation) that add a short poly(A) tail to its targets RNAs [26,27]. This poly(A) tail targets the nuclear exosome to ncRNAs and promotes exosome function to completely degrade CUTs, while it trims sn(o)RNAs precursors to their stable mature forms [19,28]. Conversely, XUTs are poly-adenylated by the canonical poly(A) polymerase, Pap1 [18,24], decapped by Dcp2, and finally degraded by Xrn1 in the cytoplasm [24], mainly by the Nonsense-Mediated mRNA Decay (NMD) pathway [24,29]. SUTs are partially sensitive to the nuclear exosome and are degraded mainly in the cytoplasm by Xrn1 after being decapped by Dcp2 [24,25,30,31]. However, some SUTs are also degraded in the cytoplasm by the NMD pathway [25,32].
Not only the NNS pathway requires Ser5P-CTD phosphorylation for Nrd1 recruitment [27], but also other posttranslational modifications of the CTD of Rpb1. Tyr1-CTD phosphorylation seems to mediate a pausing event that is critical for termination [33], probably in concert with Rtr1 Ser5P-CTD phosphatase [34,35,36], which could act also as Tyr1 phosphatase, as proposed by [35,37]. Notably, Ser7P-CTD co-localises closely with Nrd1 on CUTs, SUTs and sn(o)RNAs genes [38]. Interestingly in human cells, Ser7-CTD phosphorylation influences snRNA production in concert with the Rtr1 orthologue RPAP2 and the integrator complex [39,40,41].
Rtr1 and RPAP2 participate in RNA pol II assembly, which occurs in the cytoplasm before its nuclear import and its nuclear transport [42,43,44,45]. Furthermore, and specifically, Rtr1 is involved in the correct assembly of the Rpb4/7 dimer into complete RNA pol II in the cytoplasm, and its inactivation affects RNA pol II assembly and mRNA decay in Saccharomyces cerevisiae [42].
In this study, we investigated the effect of altering RNA pol II assembly on ncRNA transcription by using an rpb1 mutant of S. cerevisiae carrying a mutation in the conserved foot domain of RNA pol II. We provide evidence that defects in RNA pol II assembly bring about a reduction in cryptic transcription, mainly of CUTs, a feature that is not influenced by the levels of its bidirectional open reading frame (ORF). We demonstrate that low levels of CUTs are not completely suppressed upon exosome inactivation, which suggests a defect in ncRNA transcription as a consequence of the assembly defect of RNA pol II. Finally, we propose that a defect in transcription termination could account for the reduction in ncRNAs in the rpb1 mutant.
2. Results
2.1. Altering assembly of RNA pol II affects ncRNA accumulation
In order to explore how the assembly of RNA pol II influences the transcription of ncRNAs, we used an S. cerevisiae strain with a mutation corresponding to the conserved foot domain of this complex called rpb1-84. This mutation affects the correct association of the Rpb4/7 dimer with the rest of the complex, which leads to changes in mRNA synthesis and stability [47,56].
To do so, a genome-wide RNA-Seq analysis was performed with two independent biological replicates in the rpb1-84 mutant and its isogenic WT strain, grown at 30 ºC in YPD-rich medium. 3′QuantSeq and an approach similar to the 3′T-Fill methodology were employed [57,58]. Each set of two replicates, which showed good similarity (see supplemental Figure S1), were used to analyse the differentially expressed transcripts with the DESeq2 package of Bioconductor [52]. The defined annotations were used to distinguish among ORFs, CUTs, SUTs and sn(o)RNAs [10]. A specific cut-off was applied to define those genes with expression differences higher than 2-fold in the mutant compared to the WT strain (consisting of the Log2 Fold-Change of rpb1-84 vs. a WT higher than ±1 and padj<0.1) (Figure 1A). Regarding ORFs, 76 up-regulated and 249 down-regulated genes were identified. The functional analysis of Gene Ontology (GO) categories related to the biological processes for all differentially expressed mRNAs, using the Funspec software [59], showed functional categories that corresponded to the stress response for the up-regulated mRNAs. The functional categories related to transcription, transcription regulation and termination were identified among the down-regulated mRNAs (Table 1). Notably, these data agreed with the previous transcriptomic analysis for rpb1-84 and a second mutant of the foot domain of RNA pol II [56].
By applying the same cut-off for ncRNA analysis, global down-regulation was identified (Figure 1). Interestingly, while only 4.6% of ORFs were down-regulated in the rpb1-84 mutant vs. the WT strain, 32% of the annotated CUTs and 13% of SUTs were down-regulated (Figure 1B). In addition, about 12% of sn(o)RNAs also decreased. However, a similar increase of about 8% was identified for sn(o)RNAs, while a limited increase of about 1% was observed for other ncRNAs and ORFs.
As S. cerevisiae ncRNAs are classified according to their sensitivity to specific decay pathways [10,17,19], we investigated the accumulation of other ncRNAs. We analysed the accumulation of XUTs, described as ncRNAs detected upon the inactivation of Xrn1 exonuclease [18,23,24] and NUTs, which are Nrd1-unterminated transcripts [20] (Figure 1). As shown, the rpb1-84 mutant displayed a 10.68% reduction in the accumulation of the annotated XUTs, while 22.9% of the total annotated NUTs decreased in relation to the WT strain (Figure 1A and B).
Furthermore, the analyses evidenced CUTs as being the most altered class of the different types of ncRNAs in the rpb1-84 mutant in relation to the WT strain (Figure 1C). As NUTs significantly overlapped CUTs [20], and as XUTs were 3′-extended isoforms of SUTs, and some XUTs overlapped CUTs/NUTs [23,24], we cannot rule out that part of the decrease in XUTs and NUTs could be explained by the reduction in CUTs. Accordingly, we focused on CUTs and SUTs ncRNAs.
These data suggest that affecting RNA pol II assembly decreases relative ncRNA accumulation
2.2. The decrease in CUTs seems to be independent of bidirectional transcription
It has been described that promoters exhibit bidirectionality as an inherent property, and the frequent usage of bidirectional promoters is observed for divergent transcript pairs, which involves both ncRNAs and protein-coding genes transcription [10,20].
We wondered if the promoter bidirectionality could influence the reduction in CUTs accumulation. So we analysed if the decrease in CUTs could correlate with the decrease in the expression of the adjacent sense transcripts. To investigate this, we plotted the data of the gene expression between the WT and the rpb1-84 mutant for CUTs and their corresponding adjacent ORFs (Figure 2A). Lack of correlation suggested that the drop in CUTs accumulation in the rpb1-84 mutant must not be related to an effect of bidirectional transcription.
Previous data have shown that defects in RNA pol II assembly in foot mutants lower the levels of RNA pol II occupancy in protein-coding genes [47]. Accordingly, we wondered whether the decrease in ncRNAs in the rpb1-84 mutant could result from a differential reduction in RNA pol II occupancy in ncRNA transcription units vs. ORFs. Then we analysed the occupancy of RNA pol II in non-coding regions, and also in their corresponding sense ORFs, by chromatin immunoprecipitation using a Rpb1 antibody in the WT and rpb1-84 mutant cells. As shown in Figure 2B, similar lower levels of RNA pol II occupancy were observed for each pair: CUT474 -LYS20 (sense ORF) and SUT228 -PIR1 (sense ORF) for the rpb1-84 mutant. These data suggest that the decrease in ncRNA accumulation in relation to the mRNAs in the foot mutant did not result from the differential RNA pol II occupancy to these type of transcription units. In addition, we analysed Rpb1 occupancy in some sn(o)RNAs transcription units (snR5, snR13 and snR82), which displayed similar behaviour.
Based on the ChIP experiments, which revealed a general reduction in Rpb1 occupancy in the rpb1-84 mutant compared to the WT strain, we speculated about the existence of additional mechanisms that govern the reduction in ncRNA accumulation in the rpb1-84 mutant, which could be independent of RNA pol II occupancy to transcription units.
2.3. The decrease in ncRNA transcription observed in the rpb1-84 mutant does not appear to result from a malfunction of nuclear exosome
As cryptic transcripts are eliminated from the cell by the nuclear exosome [60], we wondered whether the decrease in ncRNAs, provoked by affecting the RNA pol II assembly, could depend on nuclear exosome function.
Therefore, we set out to analyse the effect of inactivating this pathway on the accumulation of ncRNAs in the rpb1-84 mutant by deleting the RRP6 gene coding for the Rrp6 subunit of the nuclear exosome [19]. For this purpose, the RNA-seq for the rpb1-84 rrp6∆ double mutant and the rrp6∆ single mutant, and their isogenic WT strain, was also performed using the same workflow as that employed for the rpb1-84 mutant (the similarity of replicates is shown in Supplemental Figure S1). As Figure 3A (and Supplemental Figures S2 and S3) depicts, and as expected, the deletion of the RRP6 gene led to a general increase in CUTs and SUTs accumulation compared to its isogenic WT strain [19]. Similar results were obtained for sn(o)RNA (not shown). In addition, the rpb1-84 rrp6∆ double mutant also displayed a general increase in ncRNA accumulation vs. the WT strain as a consequence of the rrp6∆ mutation with a slight drop in the median levels compared to the rrp6∆/WT analysis (2.992 vs. 3.1411 for CUTs and 1.930 vs. 2.026 for SUTs). However, and notably, a significant drop in ncRNA accumulation in the rpb1-84 rrp6∆ double mutant strain compared to the rrp6∆ single mutant was observed. This decrease corresponded to 11.5% of the annotated CUTs and 9% of the annotated SUTs. We also analysed the accumulation of XUTs and NUTs, and found a similar profile to that shown for CUTs and SUTs (Supplementary Figures S2 and S3).
Furthermore, the RNA-seq results were corroborated by analysing some selected transcripts by RT-qPCR (Figure 3B). Note that cryptic transcripts are often rapidly degraded, which leads to very low transcript levels under normal growth conditions [19]. Consequently, the analysis of ncRNAs in the rpb1-84 single mutant and the WT cells led to less consistent results, which are not shown.
Taken together, these results suggest that the decrease in ncRNA accumulation in the rpb1-84 mutant did not mainly depend on a defect in the nuclear exosome function.
2.4. Correcting RNA pol II assembly overcomes the decrease in pervasive transcription
The defect in the correct assembly of RNA pol II in foot mutants is overcome with RPB6 overexpression [47]. Therefore, we wondered whether the increased dose of the Rpb6 subunit was sufficient to restore the levels of the ncRNAs in the rpb1-84 mutant. We firstly analysed the effect of RPB6 overexpression on the growth phenotype of the rpb1-84 rrp6∆ double mutant and the rrp6∆ single mutant at different temperatures. As shown in Figure 4A, the temperature sensitivity phenotype of the rpb1-84 rrp6∆ double mutant was partially suppressed with RPB6 overexpression, as previously described for the rpb1-84 single mutant [47], while no additional positive effect on rrp6∆ single mutant was observed. These data indicated that RPB6 overexpression corrected the growth of the rpb1-84 single mutant, but did not seem to affect the growth of the rrp6∆ mutant.
To investigate the effect of RPB6 overexpression on ncRNA accumulation, we performed RT-qPCR experiments in both the rpb1-84 rrp6∆ double mutant and the rrp6∆ single mutant containing a plasmid that allows RPB6 overexpression or an empty plasmid as a control, and analysed two SUTs and two CUTs transcription units. As shown in Figure 4B, the levels of ncRNAs were restored by RPB6 overexpression in the rpb1-84 rrp6∆ double mutant. These data suggested that overcoming the correct RNA pol II assembly was sufficient to suppress the pervasive transcription alteration.
The assembly defect of the rpb1-84 mutant leads to an incorrect association of the Rpb4/7 dimer with the rest of the complex [47,56]. Therefore, to gain insights, we investigated whether lack of Rpb4 could affect ncRNA transcription. For this purpose, RNA-seq analyses were performed for an rpb4∆ mutant and its isogenic WT strain under the same conditions described above for the other used strains (Supplementary Figure S1 showed the similarity of replicates). The analysis of ncRNAs evidenced no major alteration to the rpb4∆ mutant in relation to the WT isogenic strain, except for SUTs (Figure 4C and Supplementary Figure S4). These data indicate that the absence of Rpb4 does not significantly affect ncRNA accumulation.
2.5. The drop in the ncRNA levels in the rpb1-84 mutant may be linked with the alteration in ncRNA transcription termination
A role for Ser5P-CTD phosphatase Rtr1 in preventing the premature termination of ncRNAs by attenuating Nrd1-dependent transcription termination has been recently reported [37]. In fact the rtr1∆ mutant significantly down-regulates ncRNAs, which suggests increased termination at non-coding genes [37]. We have previously shown a genetic interaction between the rtr1∆ mutant and the mutants of the foot domain of RNA pol II, including rpb1-84 [47]. Accordingly, we hypothesise that the assembly defect of the rpb1-84 mutant could affect ncRNA termination.
As it has been proposed that Rtr1 could dephosphorylate Tyr1-CTD of Rpb1 [35], and as Tyr1-CTD phosphorylation has been linked with the transcription termination of ncRNAs [33,61], we aimed to explore the phosphorylation of the Tyr1-CTD residues in the rpb1-84 mutant in relation to its isogenic WT strain. For this purpose, we used the chromatin-enriched fractions protocol (yChEFs) and analysed the chromatin-bound proteins by western blotting. As shown in Figure 5A, the Tyr1P/Rpb1 ratio in the rpb1-84 mutant significantly increased compared to the WT strain.
The recruitment of the human Rtr1 orthologue, RPAP2, is facilitated by Ser7 phosphorylation [41], and it influences the expression of certain snRNAs [39,40,41]. It has also been proposed that Nrd1 co-localises with Ser7P in ncRNAs in S. cerevisiae [38]. Accordingly, we also analysed the phosphorylation of the Ser7-CTD residues in the rpb1-84 mutant in the same samples used above. The western blot analysis of whole-cell extracts and chromatin-enriched fractions also evidenced a significant increase in Ser7P in the rpb1-84 mutant (Figure 5A). These data collectively suggest that an altered CTD phosphorylation pattern (Tyr1P and Ser7P) could be related to the deregulation of the ncRNA expression in the foot mutant.
Furthermore, the analysis of the biological functional categories of differentially expressed genes in the rpb1-84 mutant cells in relation to WT cells (Table 1) identified a group of mRNAs related to the ncRNA transcription termination associated with the nuclear exosome. Particularly, the mRNA encoding the Nrd1 subunit of the NNS complex decreased in the rpb1-84 mutant cells (about 2-fold). Therefore, we analysed the amount of Nrd1 associated with the chromatin in both the WT and rpb1-84 mutant strains by using a Nrd1-TAP allele. However as observed in Figure 5B, no differences in Nrd1 were found. Furthermore, Nrd1 did not seem to vary in whole-cell crude extracts, which indicates no correlation with the alteration observed during the transcriptomic analysis. In any case, we cannot rule out the possibility of partial alterations in Nrd1 occupancy at specific transcriptional units.
Collectively, these findings indicate that the assembly defect in RNA pol II provoked by the foot mutation can potentially lead to deficiencies in transcription termination during ncRNA transcription.
3. Discussion
This work provides evidence for the effect of altering the correct assembly of RNA pol II in the expression of ncRNAs using the rpb1-84 mutant of Rpb1 of RNA pol II.
The rpb1-84 mutation resides within the conserved foot domain of the major subunit of RNA pol II, Rpb1 [62], and affects the assembly of the enzyme by altering the correct association of subunit Rpb6 and dimer Rpb4/7. Altering the assembly of RNA pol II impacts transcription, which leads to a drop in mRNA levels and a general reduction in RNA pol II occupancy [47]. Similar defects in transcription have also been reported for the mutants lacking Rpb4 [56,63,64] or for the mutants of RPB6, which also affects Rpb4/7 association [65]. In agreement with the defects in mRNA expression previously reported for the foot mutant from macroarray experiments [56], our new analyses by RNA-Seq showed similar functional GO categories being altered.
Notably, our results by RNA-seq evidenced that altering RNA pol II assembly also caused a significant down-regulation of the ncRNAs in the rpb1-84 mutant, which was maximal for the transcripts resulting from pervasive transcription, CUTs and SUTs [10]. These data indicate that affecting RNA pol II assembly leads to many consequences in the transcription mediated by this complex. Our data also revealed a major decrease in XUTs and NUTs. However, NUTs have been suggested to be extended isoforms of CUTs and other non-coding RNAs [20], and some XUTs might overlap CUTs/NUTs [23,24]. Thus, we cannot rule out that the defect in these kinds of ncRNAs could be the result of the drop in CUTs accumulation. Nor can we exclude the notion that the marked decrease in CUTs could overshadow the detection of other non-coding transcripts.
In S. cerevisiae, the ncRNAs that results from pervasive transcription are very low cellular abundance RNAs, particularly CUTs [10,19], and they are usually detected following the depletion of the RNA surveillance machinery that stabilises them [9,10,17,18,19]. The deletion of nuclear exosome subunits, TRAMP complex subunits and cofactors, termination NNS complex subunits and cytoplasmic exonuclease Xrn1 specifically results in an up-regulation of the different types of pervasive transcripts [10,17,18,19,23,24,31,66,67,68]. However, and interestingly, the foot mutation that alters the RNA pol II assembly led to a general decrease in ncRNAs, which was clearly observed by the RNA-Seq analysis, and the inactivation of the Rrp6 subunit of the nuclear exosome in the rpb1-84 mutant did not completely compensate the reduction in ncRNAs. All this suggests that additional mechanisms are involved in the transcription of these types of RNAs in this mutant. It is worth noting that there are a few described conditions that bring about a drop in cryptic transcription [37,67]. Of these, one is interestingly: a decrease in ncRNAs has also been described for the rtr1∆ mutant that lacks Ser5-CTD phosphatase Rtr1 [37]. These features might be connected because a functional relation has been reported for Rtr1 and the assembly of RNA pol II [42,47] (see below).
RNA pol II assembly defects in the rpb1-84 mutant are suppressed by RPB6 overexpression [47]. In line with this, the increased Rpb6 dose restored the accumulation of ncRNAs, which supports the fact that a reduction of ncRNAs in foot mutant rpb1-84 is related to a defect in RNA pol II assembly. Although the defect in RNA pol II assembly in foot mutants alters the correct association of the Rpb4 subunit with the rest of the enzyme [42,47], the deletion of RPB4 alone does not lead to a general reduction in non-coding RNA accumulation. This suggests that the decrease in ncRNAs does not seem to depend mainly on the absence of Rpb4. Accordingly, we cannot speculate that other elements are important for ncRNA transcription, which depends on correct RNA pol II assembly.
Most yeast promoters are bidirectional, which results in divergent transcript pairs that encompass both non-coding transcripts and protein-coding genes [10,20]. Furthermore, around 90% of pervasive transcripts share the 5′ NFR with a protein-coding transcript [10]. However, our data evidenced no correlation between ncRNA and ORFs transcription with promoter bidirectionality. Furthermore, despite the fact that foot mutants exhibit a global reduction in RNA pol II occupancy to transcription units [47], we did not find this to be differential between the CUTs transcription units and their bidirectional protein-coding gene. However, given the close proximity and potential overlap of divergent transcript pairs, the sensitivity of the chromatin immunoprecipitation technique might not be adequate for distinguishing between pervasive transcripts and their bidirectional ORFs. Based on our data and those of others who have demonstrated that either affecting the association of Rpb4 with RNA pol II or deleting RPB4 display, reduces Rpb4-mRNA imprinting, which leads to increased mRNA stability [53,56,69], we cannot rule out differential stability mechanisms operating for ncRNAs and mRNAs when RNA pol II assembly is altered.
Our data suggest a possible defect in the transcription termination of ncRNA transcription that could be related to Ser5-CTD phosphatase Rtr1, which has been shown to prevent premature termination via the NNS pathway [37]. Several pieces of evidences support this hypothesis. Firstly, Rtr1, has been described to influence Rpb4 assembly in complete RNA pol II [42], and foot mutants show an assembly defect of the Rpb4/7 dimer [47]. The location of the Rpb4/7 dimer near the CTD of the Rpb1 subunit is potentially important for its interaction with termination factors, such as Seb1 in S. pombe and its homologue in S. cerevisiae, namely the Nrd1 termination factor [70,71]. Secondly, foot mutants in combination with RTR1 deletion are lethal [47]. Thirdly, Rtr1 has been proposed to act as the Tyr1P phosphatase of the CTD [35], and Tyr1 has been reported as being crucial for efficient ncRNAs termination via the NNS-mediated termination pathway [33]. In agreement with this, the rpb1-84 mutant displays high Tyr1 phosphorylation levels in relation to its WT strain. Fourthly, the Rtr1 human orthologous RPAP2 has been related to the efficient transcription of non-coding RNAs, and the recruitment of RPAP2 to snRNAs is facilitated by the Ser7 phosphorylation of the CTD of RPB1 [40]. Interestingly, foot mutants showed high Ser7 phosphorylation levels.
As previously mentioned, ncRNA transcription termination in yeast is mediated by the Nrd1-Nab3-Sen1 (NNS) pathway [27]. Interestingly, our Gene Ontology analysis of rpb1-84 showed the down-regulation of the functional categories related to this termination pathway. Accordingly, an rtr1∆ mutant displayed low NRD1 mRNA levels [37]. Interestingly, and as indicated above, the deletion of RTR1 resulted in reduced ncRNA accumulation [37], similarly to that observed for the rpb1-84 mutant. Furthermore, the human orthologue of Rtr1, RPAP2, which is knocked down by RNAi, also decreased the expression of snRNAs [40].
Nrd1 recruitment has been linked with Ser5 phosphorylation [27,33,71]. According to the role of a Ser5 phosphatase [34,35,36], it has been proposed that the increased Ser5P RNA pol II CTD levels in rtr1Δ cells may facilitate the marked activity of the NNS-dependent termination pathway by reducing cellular ncRNA abundance [37]. In agreement with this, foot mutants also increase Ser5 phosphorylation [47]. Due to the role of Rtr1 as Tyr1P-CTD phosphatase [35] and the relation between RPAP2 and Ser7P in human cells [40], we are tempted to speculate additional mechanisms for Rtr1 to participate in NNS transcription termination, which could be related to Tyr1 and Ser7 phosphorylation. Accordingly, Nrd1 colocalises with Ser7P on ncRNA transcription units, including CUTs, SUTs and sn/snoRNAs [38], and its human orthologue RPAP2 needs Ser7P for its recruitment to snRNA [40].
Tyr1 is necessary to display a pausing event for efficient NNS termination [33]. Furthermore, the mutations that convert Tyr1 into a non-phosphorylatable residue, such as the Y1F mutant, result in non-coding readthrough [33], which provokes a notable increase in both CUTs and SUTs. This coincides with decreased Nrd1 occupancy at the target promoters [72]. Accordingly, the increased Tyr1P of the rpb1-84 mutant could potentially enhance NNS function and, consequently, reduce the pool of ncRNAs as a result of increased transcription termination. However, although we did not find any differences in the global amount of Nrd1, we cannot rule out that this might be observed for specific transcription units.
In summary, we suggest that influencing RNA pol II assembly, particularly through the foot mutation, may impact ncRNA termination through the NNS pathway, likely by affecting the correct function of Rtr1, CTD phosphorylation, and possibly by interacting with other components. Finally, further studies are necessary to uncover the mechanisms that lie behind the outcomes of RNA pol II alterations to ncRNA transcription and to identify additional elements that play a role in this process.
4. Materials and Methods
4.1. Yeast Strains, Plasmid, Genetic Manipulations and Media
Common yeast media, growth conditions, and genetic techniques were employed as previously described [46]. Yeast strains and plasmids are listed and briefly described in Supplementary Tables S1 and S2.
Strain YFN542 contains the rrp6∆::KanMX4 allele and was obtained by chromosomal integration in strain Gr21-2d of the corresponding PCR product amplified by using the genomic DNA from strain Y01777 (Euroscarf) with oligonucleotides Rrp6-501 and Rrp6-301 (Table S3) as a template. Strains YFN543 and YFN545 were derived from YFN542 through plasmid shuffle exchange employing plasmids pYEB220 and pYEB220-84, as described elsewhere [47]. Strain YFN471, containing an rpb1-Δ187:: TRP1 allele, was obtained by integrating the TRP1 maker from plasmid pHT6 [48] into the rpb1-Δ187:: HIS3 marker of strain YFN166 through chromosomal integration.
4.2. RNA extraction, sequencing and bioinformatic analysis
For global RNA extraction, 50 ml cultures were grown in YPD until an OD600 of approximately 0.8 was reached. Cells were collected and utilised for total RNA extraction following the previously described procedure [49].
cDNA libraries were generated for the global expression analysis as outlined before [50]. Briefly, libraries were prepared using the QuantSeq 3′ mRNA-seq Library Prep Kit from Illumina following the manufacturer’s instructions, and samples were sequenced with an Illumina HiSeq 2000 instrument. After obtaining the Fastq sequencing datasets with FastQC, adapters were removed using Fastx Clipper, and sequences were aligned to the S. cerevisiae S288C genome (SGD R64) with Hisat2. Gene counts were determined by employing featureCounts [51]. Differential RNA abundance was analysed with the DESeq2 software [52].
The WT RNA-seq used in the RNA-seq experiment of the rpb1-84 mutant has been previously reported [53] with GEO accession numbers GSM4603016 and GSM4603018.
The WT RNA-seq applied in the RNA-seq experiment of the rpb4∆ mutant has been previously reported [50] with GEO accession numbers GSM3772984 and GSM3772985.
The other samples employed in this work are provided with GEO accession number: GSE248405.
The differential expression results of the different experiments are listed in Supplementary Table S4.
4.3. Reverse transcription and qRT-PCR
First-strand cDNA was synthesised from 0.5 µg of RNA using the iScript cDNA synthesis kit (Bio-Rad) and following the manufacturer’s protocol.
Real-time PCR analyses were conducted as previously outlined [50], utilising a CFX-384 Real-Time PCR instrument (BioRad) and the EvaGreen detection system ‘SsoFast™ EvaGreen® Supermix’ (BioRad). Three independent biological replicates of each sample were analysed in triplicate to obtain a representative average. mRNA amounts were normalised to the steady-state 18S rRNA levels. The employed oligonucleotides are listed in Supplemental Table S3.
4.4. Chromatin immunoprecipitation
The chromatin immunoprecipitation experiments were conducted using the Dynabeads M-280 sheep anti-rabbit (BioRad) and anti Rpb1 antibody (y80; Santa Cruz Biotecnology) following the previously described procedures [47]. Real-time PCR was carried out in a CFX-384 Real-Time PCR instrument (BioRad) with SYBR premix EX Taq (Takara) and the oligonucleotides listed in Supplemental Table S3. At least three independent biological replicates were analysed. For the quantitative real-time PCR, a 1:100 dilution was applied for the input DNA and a 1:4 dilution for the immunoprecipitated samples DNA.
4.5. Chromatin-enriched fractions and western blot analyses
The yChEFs procedure was followed to prepare chromatin-enriched fractions [54,55] using 50 ml of the exponentially grown YPD cultures (OD600~0.6–0.8). The final chromatin-bound proteins were resuspended in 1X SDS-PAGE sample buffer, boiled and analysed by western blot with different antibodies.
Protein electrophoresis and western blots were carried out as described in [47]. The employed antibodies included the anti-Rpb1 (against the CTD; manufactured in our laboratory) [54], anti-RNA pol II CTD phospho Tyr1 (Active Motif), anti-RNA pol II phospho Ser7 (4E12) (Chromotek), anti-phosphoglycerate kinase, Pgk1 (22C5D8; Invitrogen), anti-H3 (ab1791; Abcam) and PAP (Sigma) antibodies.
The IMAGE STUDIO LITE software was utilised to quantify the intensities of the immunoreactive bands.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: F.N., V.P. and A.I.G.-G.; experimental procedures and analyses: A.I.G.-G., I.G. and F.N.; writing: F.N. and A.I.G.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by grants from the Spanish Ministry of Science and Innovation (MCIN) and ERDF (PID2020-112853GB-C33), the Junta de Andalucía (P20-00792 and BIO258) and the University of Jaén to F.N. VP acknowledges funds from the Swedish Research Council [VR 2019-02335, 2020-01480 and 2021-06112], a Wallenberg Academy Fellowship [KAW 2021.0167], Vinnova [2020-03620] and the Karolinska Institutet (SciLifeLab SFO and KI funds).
Acknowledgments
We thank the “Servicios Centrales de Apoyo a la Investigación (SCAI)” of the University of Jaen for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Werner, F.; Grohmann, D., Evolution of multisubunit RNA polymerases in the three domains of life. Nat Rev Microbiol 2011, 9, (2), 85-98. [CrossRef]
- Barba-Aliaga, M.; Alepuz, P.; Pérez-Ortín, J. E., Eukaryotic RNA Polymerases. 2021.
- Cramer, P., Organization and regulation of gene transcription. Nature 2019, 573, (7772), 45-54. [CrossRef]
- Werner, M.; Thuriaux, P.; Soutourina, J., Structure-function analysis of RNA polymerases I and III. Curr Opin Struct Biol 2009, 19, (6), 740-5. [CrossRef]
- Pawlicki, J. M.; Steitz, J. A., Nuclear networking fashions pre-messenger RNA and primary microRNA transcripts for function. Trends Cell Biol 2010, 20, (1), 52-61. [CrossRef]
- Kornienko, A. E.; Guenzl, P. M.; Barlow, D. P.; Pauler, F. M., Gene regulation by the act of long non-coding RNA transcription. BMC Biol 2013, 11, 59. [CrossRef]
- Tuck, A. C.; Tollervey, D., RNA in pieces. Trends Genet 2011, 27, (10), 422-32. [CrossRef]
- Jacquier, A., The complex eukaryotic transcriptome: unexpected pervasive transcription and novel small RNAs. Nature Reviews Genetics 2009, 10, (12), 833-844. [CrossRef]
- Jensen, T. H.; Jacquier, A.; Libri, D., Dealing with pervasive transcription. Molecular cell 2013, 52, (4), 473-484. [CrossRef]
- Xu, Z.; Wei, W.; Gagneur, J.; Perocchi, F.; Clauder-Münster, S.; Camblong, J.; Guffanti, E.; Stutz, F.; Huber, W.; Steinmetz, L. M., Bidirectional promoters generate pervasive transcription in yeast. Nature 2009, 457, (7232), 1033-1037. [CrossRef]
- Bertone, P.; Stolc, V.; Royce, T. E.; Rozowsky, J. S.; Urban, A. E.; Zhu, X.; Rinn, J. L.; Tongprasit, W.; Samanta, M.; Weissman, S., Global identification of human transcribed sequences with genome tiling arrays. Science 2004, 306, (5705), 2242-2246. [CrossRef]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C., The transcriptional landscape of the mammalian genome. science 2005, 309, (5740), 1559-1563. [CrossRef]
- David, L.; Huber, W.; Granovskaia, M.; Toedling, J.; Palm, C. J.; Bofkin, L.; Jones, T.; Davis, R. W.; Steinmetz, L. M., A high-resolution map of transcription in the yeast genome. Proc Natl Acad Sci U S A 2006, 103, (14), 5320-5. [CrossRef]
- Li, L.; Wang, X.; Stolc, V.; Li, X.; Zhang, D.; Su, N.; Tongprasit, W.; Li, S.; Cheng, Z.; Wang, J., Genome-wide transcription analyses in rice using tiling microarrays. Nature genetics 2006, 38, (1), 124-129. [CrossRef]
- Stolc, V.; Gauhar, Z.; Mason, C.; Halasz, G.; Van Batenburg, M. F.; Rifkin, S. A.; Hua, S.; Herreman, T.; Tongprasit, W.; Barbano, P. E., A gene expression map for the euchromatic genome of Drosophila melanogaster. Science 2004, 306, (5696), 655-660. [CrossRef]
- Novačić, A.; Vučenović, I.; Primig, M.; Stuparević, I., Non-coding RNAs as cell wall regulators in Saccharomyces cerevisiae. Critical reviews in microbiology 2020, 46, (1), 15-25. [CrossRef]
- Neil, H.; Malabat, C.; d’Aubenton-Carafa, Y.; Xu, Z.; Steinmetz, L. M.; Jacquier, A., Widespread bidirectional promoters are the major source of cryptic transcripts in yeast. Nature 2009, 457, (7232), 1038-1042. [CrossRef]
- Van Dijk, E. L.; Chen, C. L.; d’Aubenton-Carafa, Y.; Gourvennec, S.; Kwapisz, M.; Roche, V.; Bertrand, C.; Silvain, M.; Legoix-Ne, P.; Loeillet, S., XUTs are a class of Xrn1-sensitive antisense regulatory non-coding RNA in yeast. Nature 2011, 475, (7354), 114-117. [CrossRef]
- Wyers, F.; Rougemaille, M.; Badis, G.; Rousselle, J.-C.; Dufour, M.-E.; Boulay, J.; Régnault, B.; Devaux, F.; Namane, A.; Séraphin, B., Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly (A) polymerase. Cell 2005, 121, (5), 725-737. [CrossRef]
- Schulz, D.; Schwalb, B.; Kiesel, A.; Baejen, C.; Torkler, P.; Gagneur, J.; Soeding, J.; Cramer, P., Transcriptome surveillance by selective termination of noncoding RNA synthesis. Cell 2013, 155, (5), 1075-1087. [CrossRef]
- Venkatesh, S.; Li, H.; Gogol, M. M.; Workman, J. L., Selective suppression of antisense transcription by Set2-mediated H3K36 methylation. Nature communications 2016, 7, (1), 13610. [CrossRef]
- Lardenois, A.; Liu, Y.; Walther, T.; Chalmel, F.; Evrard, B.; Granovskaia, M.; Chu, A.; Davis, R. W.; Steinmetz, L. M.; Primig, M., Execution of the meiotic noncoding RNA expression program and the onset of gametogenesis in yeast require the conserved exosome subunit Rrp6. Proceedings of the National Academy of Sciences 2011, 108, (3), 1058-1063. [CrossRef]
- Wery, M.; Szachnowski, U.; Andjus, S.; de Andres-Pablo, A.; Morillon, A., The RNA helicases Dbp2 and Mtr4 regulate the expression of Xrn1-sensitive long non-coding RNAs in yeast. Frontiers in RNA research 2023, 1. [CrossRef]
- Wery, M.; Descrimes, M.; Vogt, N.; Dallongeville, A.-S.; Gautheret, D.; Morillon, A., Nonsense-mediated decay restricts LncRNA levels in yeast unless blocked by double-stranded RNA structure. Molecular cell 2016, 61, (3), 379-392. [CrossRef]
- Porrua, O.; Libri, D. Porrua, O.; Libri, D., Transcription termination and the control of the transcriptome: why, where and how to stop. Nature reviews Molecular cell biology 2015, 16, (3), 190-202. [CrossRef]
- Tudek, A.; Porrua, O.; Kabzinski, T.; Lidschreiber, M.; Kubicek, K.; Fortova, A.; Lacroute, F.; Vanacova, S.; Cramer, P.; Stefl, R., Molecular basis for coordinating transcription termination with noncoding RNA degradation. Molecular cell 2014, 55, (3), 467-481. [CrossRef]
- Vasiljeva, L.; Kim, M.; Mutschler, H.; Buratowski, S.; Meinhart, A., The Nrd1-Nab3-Sen1 termination complex interacts with the Ser5-phosphorylated RNA polymerase II C-terminal domain. Nat Struct Mol Biol 2008, 15, (8), 795-804. [CrossRef]
- Kilchert, C.; Wittmann, S.; Vasiljeva, L., The regulation and functions of the nuclear RNA exosome complex. Nature Reviews Molecular Cell Biology 2016, 17, (4), 227-239. [CrossRef]
- Malabat, C.; Feuerbach, F.; Ma, L.; Saveanu, C.; Jacquier, A., Quality control of transcription start site selection by nonsense-mediated-mRNA decay. Elife 2015, 4, e06722. [CrossRef]
- Marquardt, S.; Hazelbaker, D. Z.; Buratowski, S., Distinct RNA degradation pathways and 3′extensions of yeast non-coding RNA species. Transcription 2011, 2, (3), 145-154. [CrossRef]
- Sohrabi-Jahromi, S.; Hofmann, K. B.; Boltendahl, A.; Roth, C.; Gressel, S.; Baejen, C.; Soeding, J.; Cramer, P., Transcriptome maps of general eukaryotic RNA degradation factors. Elife 2019, 8, e47040. [CrossRef]
- Smith, J. E.; Alvarez-Dominguez, J. R.; Kline, N.; Huynh, N. J.; Geisler, S.; Hu, W.; Coller, J.; Baker, K. E., Translation of small open reading frames within unannotated RNA transcripts in Saccharomyces cerevisiae. Cell reports 2014, 7, (6), 1858-1866. [CrossRef]
- Collin, P.; Jeronimo, C.; Poitras, C.; Robert, F., RNA Polymerase II CTD Tyrosine 1 Is Required for Efficient Termination by the Nrd1-Nab3-Sen1 Pathway. Molecular cell 2019, 73, (4), 655-669. e7. [CrossRef]
- Mosley, A. L.; Pattenden, S. G.; Carey, M.; Venkatesh, S.; Gilmore, J. M.; Florens, L.; Workman, J. L.; Washburn, M. P., Rtr1 is a CTD phosphatase that regulates RNA polymerase II during the transition from serine 5 to serine 2 phosphorylation. Mol Cell 2009, 34, (2), 168-78. [CrossRef]
- Hsu, P. L.; Yang, F.; Smith-Kinnaman, W.; Yang, W.; Song, J.-E.; Mosley, A. L.; Varani, G., Rtr1 is a dual specificity phosphatase that dephosphorylates Tyr1 and Ser5 on the RNA polymerase II CTD. Journal of molecular biology 2014, 426, (16), 2970-2981. [CrossRef]
- Hunter, G. O.; Fox, M. J.; Smith-Kinnaman, W. R.; Gogol, M.; Fleharty, B.; Mosley, A. L., Phosphatase Rtr1 regulates global levels of serine 5 RNA polymerase II C-terminal domain phosphorylation and cotranscriptional histone methylation. Molecular and cellular biology 2016, 36, (17), 2236-2245. [CrossRef]
- Victorino, J. F.; Fox, M. J.; Smith-Kinnaman, W. R.; Justice, S. A. P.; Burriss, K. H.; Boyd, A. K.; Zimmerly, M. A.; Chan, R. R.; Hunter, G. O.; Liu, Y., RNA Polymerase II CTD phosphatase Rtr1 fine-tunes transcription termination. PLoS genetics 2020, 16, (3), e1008317. [CrossRef]
- Kim, H.; Erickson, B.; Luo, W.; Seward, D.; Graber, J. H.; Pollock, D. D.; Megee, P. C.; Bentley, D. L., Gene-specific RNA polymerase II phosphorylation and the CTD code. Nat Struct Mol Biol 2010. [CrossRef]
- Egloff, S.; O’Reilly, D.; Chapman, R. D.; Taylor, A.; Tanzhaus, K.; Pitts, L.; Eick, D.; Murphy, S., Serine-7 of the RNA polymerase II CTD is specifically required for snRNA gene expression. Science 2007, 318, (5857), 1777-9. [CrossRef]
- Egloff, S.; Zaborowska, J.; Laitem, C.; Kiss, T.; Murphy, S., Ser7 phosphorylation of the CTD recruits the RPAP2 Ser5 phosphatase to snRNA genes. Mol Cell 2012, 45, (1), 111-22. [CrossRef]
- Egloff, S., Role of Ser7 phosphorylation of the CTD during transcription of snRNA genes. RNA biology 2012, 9, (8), 1033-1038. [CrossRef]
- Garrido-Godino, A. I.; Cuevas-Bermúdez, A.; Gutiérrez-Santiago, F.; Mota-Trujillo, M. d. C.; Navarro, F., The Association of Rpb4 with RNA Polymerase II Depends on CTD Ser5P Phosphatase Rtr1 and Influences mRNA Decay in Saccharomyces cerevisiae. International journal of molecular sciences 2022, 23, (4), 2002. [CrossRef]
- Garrido-Godino, A. I.; Gutiérrez-Santiago, F.; Navarro, F., Biogenesis of RNA Polymerases in Yeast. Frontiers in Molecular Biosciences 2021, 8. [CrossRef]
- Gómez-Navarro, N.; Estruch, F., Different pathways for the nuclear import of yeast RNA polymerase II. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2015, 1849, (11), 1354-1362. [CrossRef]
- Forget, D.; Lacombe, A. A.; Cloutier, P.; Lavallee-Adam, M.; Blanchette, M.; Coulombe, B., Nuclear import of RNA polymerase II is coupled with nucleocytoplasmic shuttling of the RNA polymerase II-associated protein 2. Nucleic Acids Res 2013, 41, (14), 6881-91. [CrossRef]
- Garcia-Lopez, M. C.; Miron-Garcia, M. C.; Garrido-Godino, A. I.; Mingorance, C.; Navarro, F., Overexpression of SNG1 causes 6-azauracil resistance in Saccharomyces cerevisiae. Curr Genet 2010, 56, (3), 251-63. [CrossRef]
- Garrido-Godino, A. I.; Garcia-Lopez, M. C.; Navarro, F., Correct Assembly of RNA Polymerase II Depends on the Foot Domain and Is Required for Multiple Steps of Transcription in Saccharomyces cerevisiae. Mol Cell Biol 2013, 33, (18), 3611-26. [CrossRef]
- Cross, F. R., ‘Marker swap’ plasmids: convenient tools for budding yeast molecular genetics. Yeast 1997, 13, (7), 647-53.
- Nonet, M.; Scafe, C.; Sexton, J.; Young, R., Eucaryotic RNA polymerase conditional mutant that rapidly ceases mRNA synthesis. Mol Cell Biol 1987, 7, (5), 1602-11. [CrossRef]
- Martínez-Fernández, V.; Cuevas-Bermúdez, A.; Gutiérrez-Santiago, F.; Garrido-Godino, A. I.; Rodríguez-Galán, O.; Jordán-Pla, A.; Lois, S.; Triviño, J. C.; de la Cruz, J.; Navarro, F., Prefoldin-like Bud27 influences the transcription of ribosomal components and ribosome biogenesis in Saccharomyces cerevisiae. RNA 2020, rna. 075507.120. [CrossRef]
- Liao, Y.; Smyth, G. K.; Shi, W., featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, (7), 923-930. [CrossRef]
- Love, M. I.; Huber, W.; Anders, S., Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome biology 2014, 15, (12), 1-21. [CrossRef]
- Garrido-Godino, A. I.; Gupta, I.; Gutiérrez-Santiago, F.; Martínez-Padilla, A. B.; Alekseenko, A.; Steinmetz, L. M.; Pérez-Ortín, J. E.; Pelechano, V.; Navarro, F., Rpb4 and Puf3 imprint and post-transcriptionally control the stability of a common set of mRNAs in yeast. RNA biology 2021, 18, (8), 1206-1220. [CrossRef]
- Cuevas-Bermúdez, A.; Garrido-Godino, A. I.; Navarro, F., A novel yeast chromatin-enriched fractions purification approach, yChEFs, for the chromatin-associated protein analysis used for chromatin-associated and RNA-dependent chromatin-associated proteome studies from Saccharomyces cerevisiae. Gene Reports 2019, 16, 100450. [CrossRef]
- Cuevas-Bermúdez, A.; Garrido-Godino, A. I.; Gutiérrez-Santiago, F.; Navarro, F., A Yeast Chromatin-enriched Fractions Purification Approach, yChEFs, from Saccharomyces cerevisiae. Bio-Protocol 2020, 10, (1). [CrossRef]
- Garrido-Godino, A.; García-López, M.; García-Martínez, J.; Pelechano, V.; Medina, D.; Pérez-Ortín, J.; Navarro, F., Rpb1 foot mutations demonstrate a major role of Rpb4 in mRNA stability during stress situations in yeast. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2016, 1859, (5), 731-743. [CrossRef]
- Pelechano, V.; Wilkening, S.; Jarvelin, A. I.; Tekkedil, M. M.; Steinmetz, L. M., Genome-wide polyadenylation site mapping. Methods Enzymol 2012, 513, 271-96. [CrossRef]
- Wilkening, S.; Pelechano, V.; Jarvelin, A. I.; Tekkedil, M. M.; Anders, S.; Benes, V.; Steinmetz, L. M., An efficient method for genome-wide polyadenylation site mapping and RNA quantification. Nucleic Acids Res 2013, 41, (5), e65. [CrossRef]
- Robinson, M. D.; Grigull, J.; Mohammad, N.; Hughes, T. R., FunSpec: a web-based cluster interpreter for yeast. BMC bioinformatics 2002, 3, (1), 35. [CrossRef]
- Lykke-Andersen, S.; Jensen, T. H., Nonsense-mediated mRNA decay: an intricate machinery that shapes transcriptomes. Nature reviews Molecular cell biology 2015, 16, (11), 665-677. [CrossRef]
- Mayer, A.; Heidemann, M.; Lidschreiber, M.; Schreieck, A.; Sun, M.; Hintermair, C.; Kremmer, E.; Eick, D.; Cramer, P., CTD tyrosine phosphorylation impairs termination factor recruitment to RNA polymerase II. Science 2012, 336, (6089), 1723-5. [CrossRef]
- García-López, M. C.; Navarro, F., RNA polymerase II conserved protein domains as platforms for protein-protein interactions. Transcription 2011, 2, (4), 193-7. [CrossRef]
- Schulz, D.; Pirkl, N.; Lehmann, E.; Cramer, P., Rpb4 subunit functions mainly in mRNA synthesis by RNA polymerase II. Journal of Biological Chemistry 2014, 289, (25), 17446-17452. [CrossRef]
- Allepuz-Fuster, P.; Martínez-Fernández, V.; Garrido-Godino, A. I.; Alonso-Aguado, S.; Hanes, S. D.; Navarro, F.; Calvo, O., Rpb4/7 facilitates RNA polymerase II CTD dephosphorylation. Nucleic acids research 2014, 42, (22), 13674-13688. [CrossRef]
- Shalem, O.; Groisman, B.; Choder, M.; Dahan, O.; Pilpel, Y., Transcriptome kinetics is governed by a genome-wide coupling of mRNA production and degradation: a role for RNA Pol II. PLoS Genet 2011, 7, (9), e1002273. [CrossRef]
- Fasken, M. B.; Leung, S. W.; Banerjee, A.; Kodani, M. O.; Chavez, R.; Bowman, E. A.; Purohit, M. K.; Rubinson, M. E.; Rubinson, E. H.; Corbett, A. H., Air1 zinc knuckles 4 and 5 and a conserved IWRXY motif are critical for the function and integrity of the Trf4/5-Air1/2-Mtr4 polyadenylation (TRAMP) RNA quality control complex. Journal of Biological Chemistry 2011, 286, (43), 37429-37445. [CrossRef]
- Ellison, M. A.; Lederer, A. R.; Warner, M. H.; Mavrich, T. N.; Raupach, E. A.; Heisler, L. E.; Nislow, C.; Lee, M. T.; Arndt, K. M., The Paf1 complex broadly impacts the transcriptome of Saccharomyces cerevisiae. Genetics 2019, 212, (3), 711-728. [CrossRef]
- Schneider, C.; Kudla, G.; Wlotzka, W.; Tuck, A.; Tollervey, D., Transcriptome-wide analysis of exosome targets. Molecular cell 2012, 48, (3), 422-433. [CrossRef]
- Goler-Baron, V.; Selitrennik, M.; Barkai, O.; Haimovich, G.; Lotan, R.; Choder, M., Transcription in the nucleus and mRNA decay in the cytoplasm are coupled processes. Genes Dev 2008, 22, (15), 2022-7. [CrossRef]
- Mitsuzawa, H.; Kanda, E.; Ishihama, A., Rpb7 subunit of RNA polymerase II interacts with an RNA-binding protein involved in processing of transcripts. Nucleic Acids Res 2003, 31, (16), 4696-701. [CrossRef]
- Kubicek, K.; Cerna, H.; Holub, P.; Pasulka, J.; Hrossova, D.; Loehr, F.; Hofr, C.; Vanacova, S.; Stefl, R., Serine phosphorylation and proline isomerization in RNAP II CTD control recruitment of Nrd1. Genes & development 2012, 26, (17), 1891-1896. [CrossRef]
- Yurko, N.; Liu, X.; Yamazaki, T.; Hoque, M.; Tian, B.; Manley, J. L., MPK1/SLT2 links multiple stress responses with gene expression in budding yeast by phosphorylating Tyr1 of the RNAP II CTD. Molecular cell 2017, 68, (5), 913-925. e3. [CrossRef]
Figure 1.
ncRNAs globally decrease by altering RNA pol II assembly. (A) RNA-seq for the rpb1-84 mutant and its isogenic wild-type (WT) strain. Volcano plot generated to identify the differentially expressed transcripts in the rpb1-84 mutant vs. the WT strain (n=2, DESeq2) [52]. A specific cut-off was applied to define the altered accumulation transcripts, consisting in Log2 Fold-Change of the rpb1-84 mutant vs. WT higher than ±1 (dashed grey lines) and padj<0.1 (dashed red line). Left upper panel shows the mRNA transcripts, whereas specific classes of ncRNAs are coloured in the other panels: CUTs (orange), SUTs (green), sn(o)RNAs (cyan), XUTs (violet) and NUTs (salmon). (B) Percentage of transcripts from each category that are down-regulated (red) and up-regulated (green) in the rpb1-84 mutant vs. the WT strain according to the parameters indicated in panel A. (C) Box-plot showing the distribution for each type of transcripts in the rpb1-84 mutant vs. the WT strain. The indicated p-values were obtained during a two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group.
Figure 1.
ncRNAs globally decrease by altering RNA pol II assembly. (A) RNA-seq for the rpb1-84 mutant and its isogenic wild-type (WT) strain. Volcano plot generated to identify the differentially expressed transcripts in the rpb1-84 mutant vs. the WT strain (n=2, DESeq2) [52]. A specific cut-off was applied to define the altered accumulation transcripts, consisting in Log2 Fold-Change of the rpb1-84 mutant vs. WT higher than ±1 (dashed grey lines) and padj<0.1 (dashed red line). Left upper panel shows the mRNA transcripts, whereas specific classes of ncRNAs are coloured in the other panels: CUTs (orange), SUTs (green), sn(o)RNAs (cyan), XUTs (violet) and NUTs (salmon). (B) Percentage of transcripts from each category that are down-regulated (red) and up-regulated (green) in the rpb1-84 mutant vs. the WT strain according to the parameters indicated in panel A. (C) Box-plot showing the distribution for each type of transcripts in the rpb1-84 mutant vs. the WT strain. The indicated p-values were obtained during a two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group.
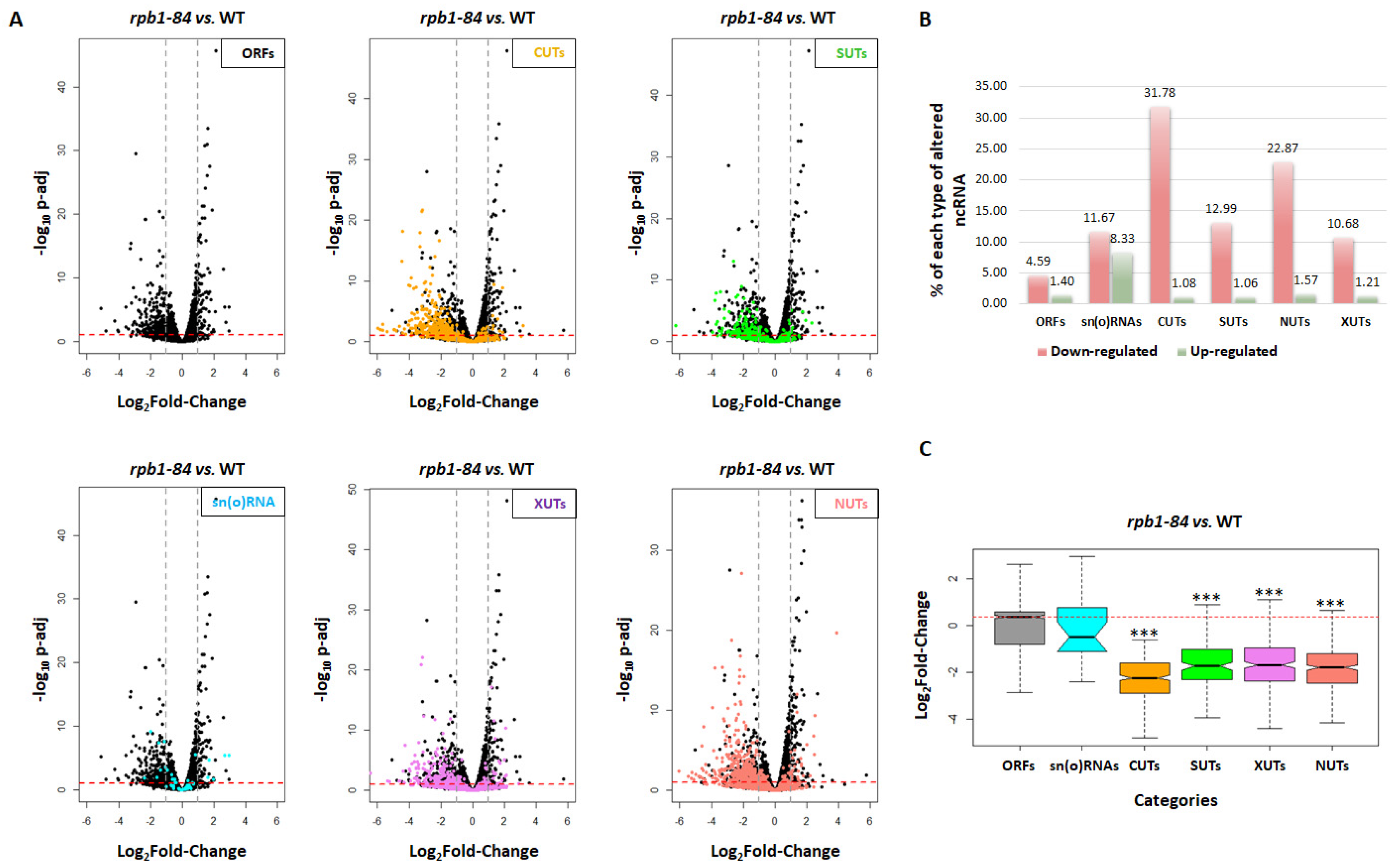
Figure 2.
The down-regulation of ncRNAs is not dependent on bidirectional promoters. (A) Scatterplot showing the correlation between the accumulation of CUTs and their corresponding bidirectional sense mRNAs in the wild-type (WT) strains vs. the rpb1-84 mutant. (B) Chromatin immunoprecipitation (ChIP) analysis for the different transcription units in the WT and rpb1-84 cells, performed with an anti-Rpb1 antibody. A negative control without an antibody is also shown, as well as an intergenic non-transcribed region of chromosome V used as the negative control. The values found for the immunoprecipitated PCR products were compared as % of immunoprecipitated material, IP, vs. INPUT. Bars represent standard deviation. At least three independent experiments were performed.
Figure 2.
The down-regulation of ncRNAs is not dependent on bidirectional promoters. (A) Scatterplot showing the correlation between the accumulation of CUTs and their corresponding bidirectional sense mRNAs in the wild-type (WT) strains vs. the rpb1-84 mutant. (B) Chromatin immunoprecipitation (ChIP) analysis for the different transcription units in the WT and rpb1-84 cells, performed with an anti-Rpb1 antibody. A negative control without an antibody is also shown, as well as an intergenic non-transcribed region of chromosome V used as the negative control. The values found for the immunoprecipitated PCR products were compared as % of immunoprecipitated material, IP, vs. INPUT. Bars represent standard deviation. At least three independent experiments were performed.
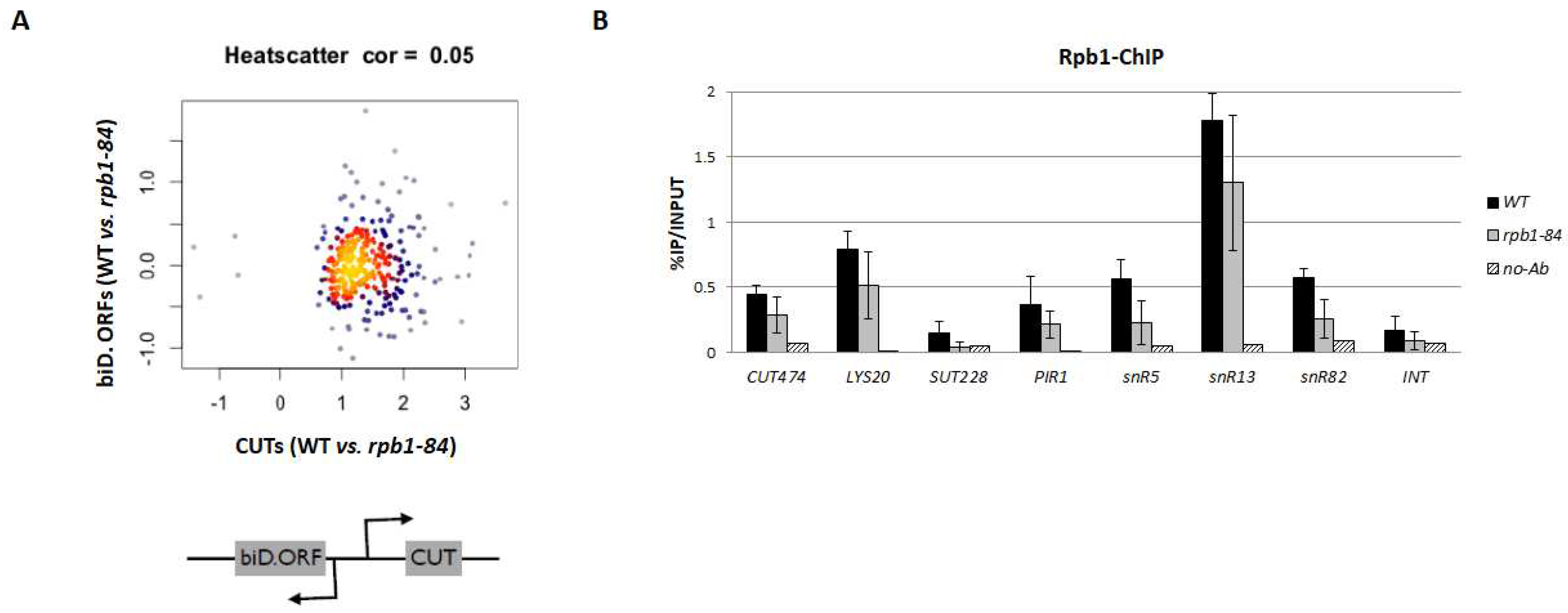
Figure 3.
Exosome inactivation analysis by RNA-seq for the rpb1-84 rrp6∆ double mutant, the rrp6∆ single mutant and their isogenic wild-type strains. (A) Box-plot showing the distribution of ORFs, CUTs and SUTs in the rrp6∆ mutant vs. wild type (WT; left panel), the rpb1-84 rrp6∆ mutant vs. the WT strain (middle panel) and the rpb1-84 rrp6∆ mutant vs. the rrp6∆ mutant strain (right panel). The indicated p-values were obtained during the two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group. (B) The RT-qPCR analysis for the CUTs and SUTs accumulation in the rpb1-84 rrp6∆ vs. rrp6∆ mutant strain. The values obtained for the rrp6∆ single mutant are represented as 1 for all the tested transcripts. Data are shown as the average and standard deviation (SD) of two independent experiments. rRNA 18S was used as a normaliser.
Figure 3.
Exosome inactivation analysis by RNA-seq for the rpb1-84 rrp6∆ double mutant, the rrp6∆ single mutant and their isogenic wild-type strains. (A) Box-plot showing the distribution of ORFs, CUTs and SUTs in the rrp6∆ mutant vs. wild type (WT; left panel), the rpb1-84 rrp6∆ mutant vs. the WT strain (middle panel) and the rpb1-84 rrp6∆ mutant vs. the rrp6∆ mutant strain (right panel). The indicated p-values were obtained during the two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group. (B) The RT-qPCR analysis for the CUTs and SUTs accumulation in the rpb1-84 rrp6∆ vs. rrp6∆ mutant strain. The values obtained for the rrp6∆ single mutant are represented as 1 for all the tested transcripts. Data are shown as the average and standard deviation (SD) of two independent experiments. rRNA 18S was used as a normaliser.
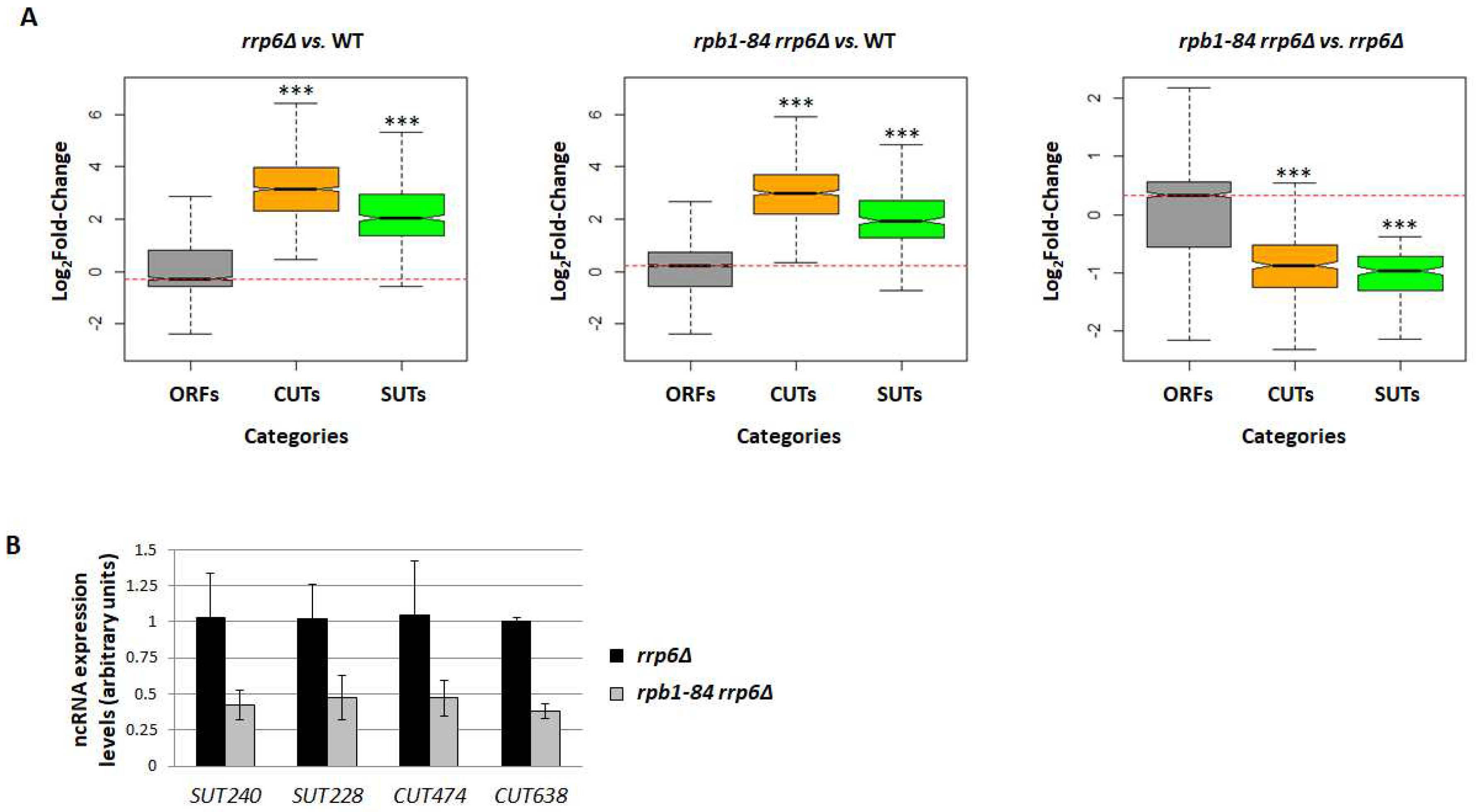
Figure 4.
ncRNA accumulation is dependent on RNA pol II assembly. (A) Growth assay showing the overexpression of RPB6 in the rpb1-84 rrp6∆ double mutant compared to the rrp6∆ single mutant cells grown in SD (-uracil) at different temperatures. (B) The RT-qPCR analysis for the CUTs and SUTs accumulation in the rpb1-84 rrp6∆ vs. the rrp6∆ mutant cells overexpressing RPB6. Data are shown as the average and standard deviation (SD) of two independent experiments. rRNA 18S was used as a normaliser. The values obtained for the rrp6∆ single mutant are represented as 1 for all the tested transcripts. (C) Box-plot showing the distribution of ORFs, CUTs and SUTs in the rpb4∆ mutant vs. the wild type. The indicated p-values were obtained during the two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group.
Figure 4.
ncRNA accumulation is dependent on RNA pol II assembly. (A) Growth assay showing the overexpression of RPB6 in the rpb1-84 rrp6∆ double mutant compared to the rrp6∆ single mutant cells grown in SD (-uracil) at different temperatures. (B) The RT-qPCR analysis for the CUTs and SUTs accumulation in the rpb1-84 rrp6∆ vs. the rrp6∆ mutant cells overexpressing RPB6. Data are shown as the average and standard deviation (SD) of two independent experiments. rRNA 18S was used as a normaliser. The values obtained for the rrp6∆ single mutant are represented as 1 for all the tested transcripts. (C) Box-plot showing the distribution of ORFs, CUTs and SUTs in the rpb4∆ mutant vs. the wild type. The indicated p-values were obtained during the two-sided Wilcoxon rank-sum test. * Wilcoxon-test p-value < 10−3; ** p-value < 10−5; *** p-value < 10−9. Each group of analysed transcripts was compared to the ‘ORFs’ group.
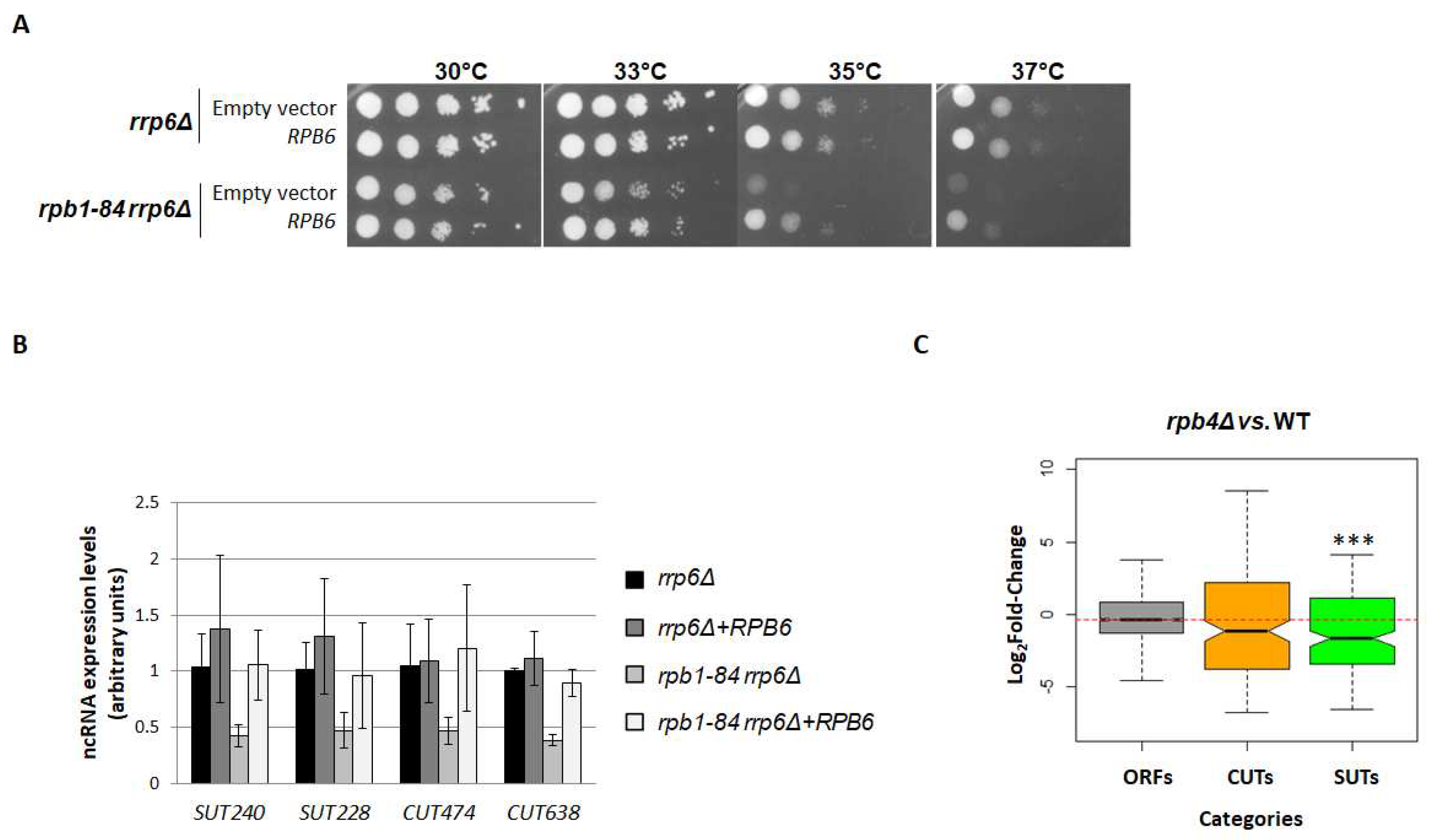
Figure 5.
CTD-phosphorylation and the Nrd1 analysis in chromatin fractions of the foot mutant. (A) Left panel: the whole-cell extract and chromatin-bound proteins isolated by the yChEFs procedure [54,55] from the wild-type (WT) and rpb1-84 strains grown in YPD medium at 30 ºC and analysed by western blotting using specific antibodies against Rpb1, Tyr1P-CTD and Ser7P-CTD. The H3 histone was used as a positive control of chromatin and Pgk1 as a negative control of cytoplasmic contamination. Right panel: quantification of the experiments in the left panel. Levels of the phosphorylated-CTD/Rpb1 ratios for the WT strain are represented as 1. (B) Left panel: whole-cell extract and chromatin-bound proteins isolated by the yChEFs procedure [54,55] from the WT and rpb1-84 strains carrying an Nrd1-TAP allele grown in YPD medium at 30 ºC were analysed by western blotting using specific antibodies against Rpb1 and TAP, and against the H3 histone used as a positive control of chromatin and Pgk1 employed as a negative control of cytoplasmic contamination. Right panel: quantification of the experiments in the left panel. Levels of the Nrd1/Rpb1 ratios and Nrd1/H3 ratios for the WT strain are represented as 1. At least three independent experiments with independent biological replicates were performed.
Figure 5.
CTD-phosphorylation and the Nrd1 analysis in chromatin fractions of the foot mutant. (A) Left panel: the whole-cell extract and chromatin-bound proteins isolated by the yChEFs procedure [54,55] from the wild-type (WT) and rpb1-84 strains grown in YPD medium at 30 ºC and analysed by western blotting using specific antibodies against Rpb1, Tyr1P-CTD and Ser7P-CTD. The H3 histone was used as a positive control of chromatin and Pgk1 as a negative control of cytoplasmic contamination. Right panel: quantification of the experiments in the left panel. Levels of the phosphorylated-CTD/Rpb1 ratios for the WT strain are represented as 1. (B) Left panel: whole-cell extract and chromatin-bound proteins isolated by the yChEFs procedure [54,55] from the WT and rpb1-84 strains carrying an Nrd1-TAP allele grown in YPD medium at 30 ºC were analysed by western blotting using specific antibodies against Rpb1 and TAP, and against the H3 histone used as a positive control of chromatin and Pgk1 employed as a negative control of cytoplasmic contamination. Right panel: quantification of the experiments in the left panel. Levels of the Nrd1/Rpb1 ratios and Nrd1/H3 ratios for the WT strain are represented as 1. At least three independent experiments with independent biological replicates were performed.
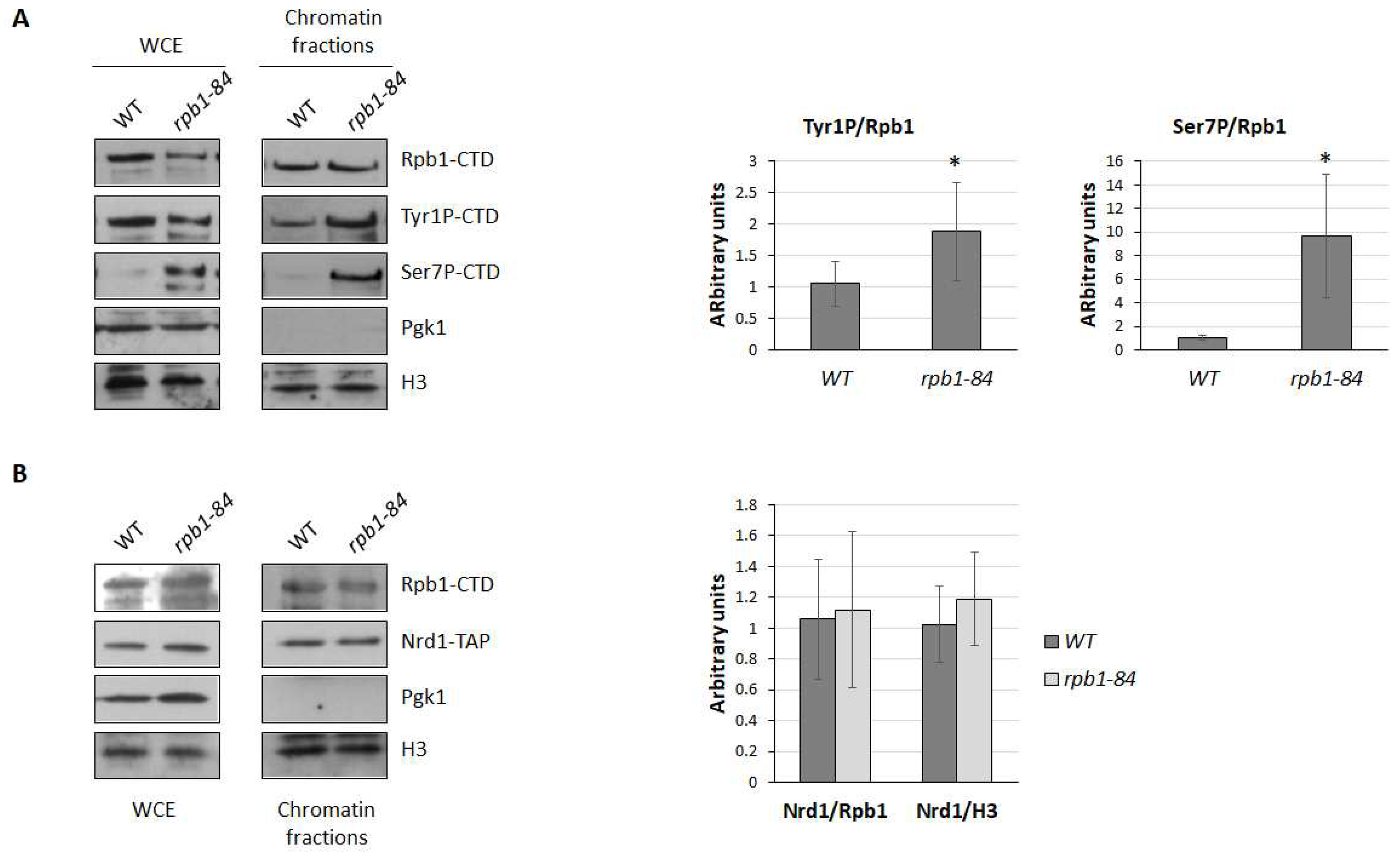

Table 1.
Biological functional categories of differentially expressed genes in rpb1-84 mutant cells with respect to wild-type cells. Analysis was performed using Funspec software [59].
Table 1.
Biological functional categories of differentially expressed genes in rpb1-84 mutant cells with respect to wild-type cells. Analysis was performed using Funspec software [59].
| Functional Category | p-value | |
|---|---|---|
| Up-regulated | purine nucleotide biosynthetic process [GO:0006164] | 7.81559e-05 |
| glycolysis [GO:0006096] | 0.000472358 | |
| response to stress [GO:0006950] | 0.000887445 | |
| metabolic process [GO:0008152] | 0.00134185 | |
| gluconeogenesis [GO:0006094] | 0.001615 | |
| cell adhesion [GO:0007155] | 0.00353101 | |
| manganese ion transport [GO:0006828] | 0.00466732 | |
| ‘de novo’ IMP biosynthetic process [GO:0006189] | 0.00594903 | |
| Down-regulated | transcription, DNA-dependent [GO:0006351] | 0.0009675 |
| sexual reproduction [GO:0019953] | 0.00111532 | |
| mating [GO:0007618] | 0.00111532 | |
| trehalose biosynthetic process [GO:0005992] | 0.00117173 | |
| regulation of transcription, DNA-dependent [GO:0006355] | 0.0014433 | |
| ATP-dependent chromatin remodeling [GO:0043044] | 0.004667 | |
| termination of RNA polymerase II transcription, exosome-dependent [GO:0030847] | 0.00500098 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



